Genetic diversity of the Q fever agent Coxiella burnet Paul Beare Coxiella Pathogenesis Section Rocky Mountain Laboratories LICP/NIAID/NIH

Genetic diversity of the Q fever agent Coxiella burnetii Paul Beare Coxiella Pathogenesis Section Rocky Mountain Laboratories LICP/NIAID/NIH.
Dec 27, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Genetic diversity of the Q fever agent Coxiella burnetii
Paul Beare
Coxiella Pathogenesis SectionRocky Mountain Laboratories
LICP/NIAID/NIH
Coleman et al, J Bact 2004
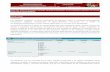
Coxiella burnetii (1937- )
•Obligate intracellular Gram-negative bacterium
•Causative agent of query (Q) fever•Can manifest as acute or chronic disease
•Replicates in a lysosome-like vacoule•Exists as two different developmental forms
•small cell and large cell variant•No systems for genetic manipulation•Category B bioterrorism agent

Ubiquitous 4-legged “bioterrorists”

Ubiquitous 4-legged “bioterrorists”

Ubiquitous 4-legged “bioterrorists”

Ubiquitous 4-legged “bioterrorists”

•Sequence of the Nine Mile phase I RSA493 reference isolate has been determined (Seshadri et al, PNAS, 2003)
•Genome consists of a 1,995,281 bp circular chromosome and 37,393 bp plasmid
•~2,134 coding sequences (~34% hypothetical)
•29 insertion sequence elements
•83 pseudogenes
•Restriction fragment length polymorphism (RFLP) and multispacer sequence typing analyses reveal a range of genetic diversity between different isolates
Coxiella - Genome characteristics

Coxiella – Isolate differentiation
•RFLP analysis
Restriction enzyme fragmentation of genomic DNA followed by an analysis of the associated fragments
•Multispacer sequence typing
Based on the sequencing of multiple intergenic regions
Genomic groupIIIIIIIIIIIIIIIIIIIVIVIVVVVVIUNUNUNUN
Plasmid typeQpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpH1QpRSQpRSQpRS
IPIPIP
QpDGQpDVPTUPTUPTU
IsolateNine Mile RSA493Nine Mile RSA439Nine Mile RSA514 (Crazy)Australia QD RSA425D RSA345Turkey RSA333African RSA 334Giroud RSA431El Tayeb RSA 342Panama RSA335California 33 RSA329California 16 RSA350Ohio 314 RSA370Henzerling RSA331Idaho Goat Q195MSU Goat Q177K Q154P Q173G Q212S Q217Ko Q229Dugway 7E9-12Q321Dugway 5G61-63BDT1Le Bruges
Original source, yrMontana, tick, 1935Montana, tick, 1935Montana, tick, 1935Australia, human blood, ~1939USA, human blood, 1938Turkey, human blood, 1948Central Africa, human blood, 1949Central Africa, human blood, 1949Egypt, tick, 1967Panama, chiggers, 1961California, cow’s milk, 1947California, cow’s milk, 1947Ohio, cow’s milk, 1956Italy, human blood, 1945Idaho, goat, 1981Montana, goat cotyledon, 1980Oregon, human heart valve, 1976California, human heart valve, 1979Nova Scotia, human heart valve, 1981Montana, human livery biopsy, 1981Nova Scotia, human heart valve, 1982Utah, rodent, 1958Russia, cow’s milkUtah, tick, 1958USA, brown dog tick, 1951France, unknown source
Disease---
AcuteAcuteAcuteAcuteAcute
--
PersistantPersistantPersistent
AcuteAbortionAbortion
EndocarditisEndocarditisEndocarditis
HepatitisEndocarditis
-Persistant
---
Isolates used in this study

Study aims
•Examine the genetic diversity of Coxiella isolates
•Phylogenetically group ungrouped isolates
•Examine the potential link between pathotype and genome profile
•Estimate the core Coxiella genome

The Coxiella microarray
•Multipathogen Affymetrix expression array (RMLchip)
•Covers 1,988 chromosomal and 36 plasmid ORFs of the Nine Mile phase I RSA493 reference isolate
•~249,690 Coxiella-specific 25-mer probe pairs
•Each Coxiella ORF is represented by 16 probe pairs
•Maximum of 400 bp coverage for each ORF
•Total of ~30% coverage of the Coxiella genome
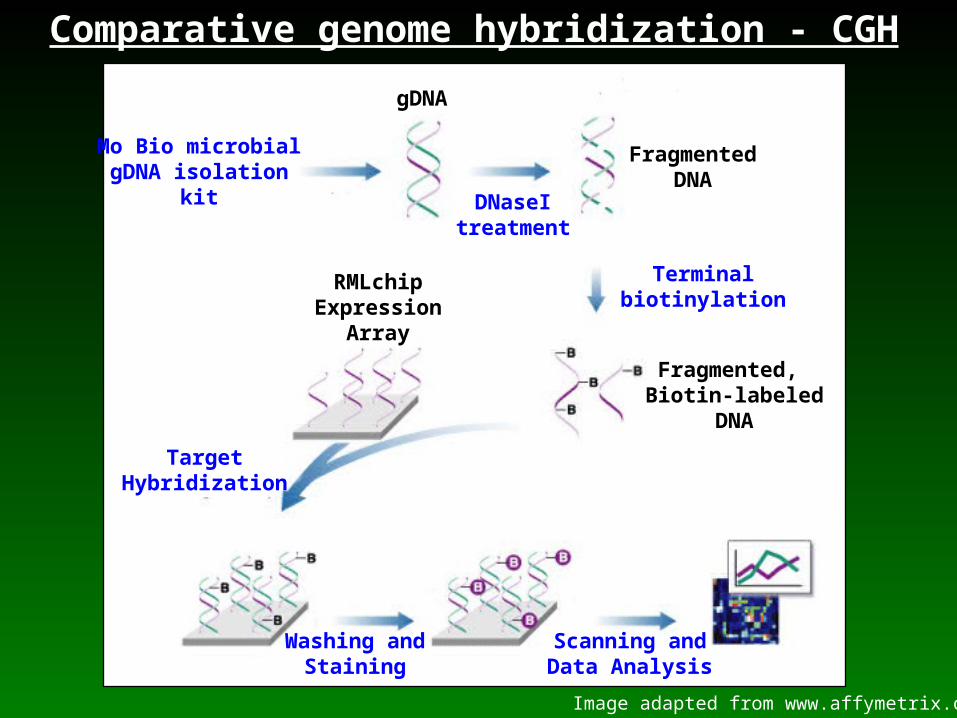
Mo Bio microbialgDNA isolation
kit
gDNA
FragmentedDNA
DNaseItreatment
Terminalbiotinylation
Fragmented, Biotin-labeled
DNA
RMLchipExpression
Array
TargetHybridization
Washing andStaining
Scanning andData Analysis
Image adapted from www.affymetrix.com
Comparative genome hybridization - CGH
Gro
up I
Gro
up II
I
Gro
up IV
Gro
up V
I
Gro
up V
BD
TGro
up II
Key:Present ORFAbsent ORFPartial ORFORF containing a PM or small insertion
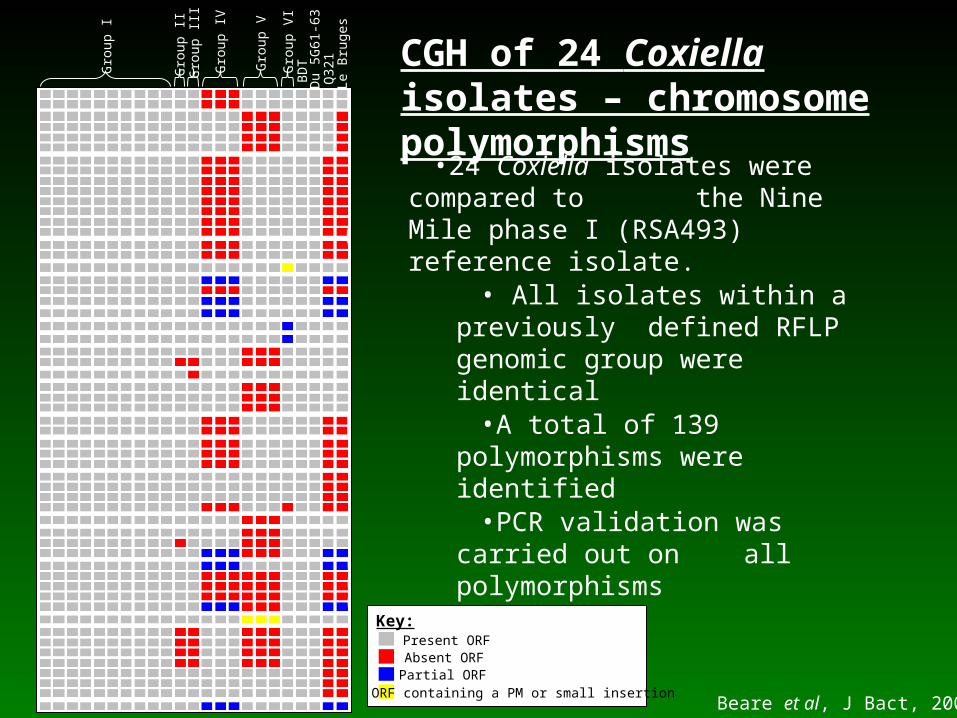
•24 Coxiella isolates were compared to the Nine Mile phase I (RSA493) reference isolate.
• All isolates within a previously defined RFLP genomic group
were identical•A total of 139 polymorphisms
were identified•PCR validation was carried out on all polymorphisms
Beare et al, J Bact, 2006
Du
5G61
-63
Q32
1Le
Bru
ges
CGH of 24 Coxiella isolates – chromosome polymorphisms
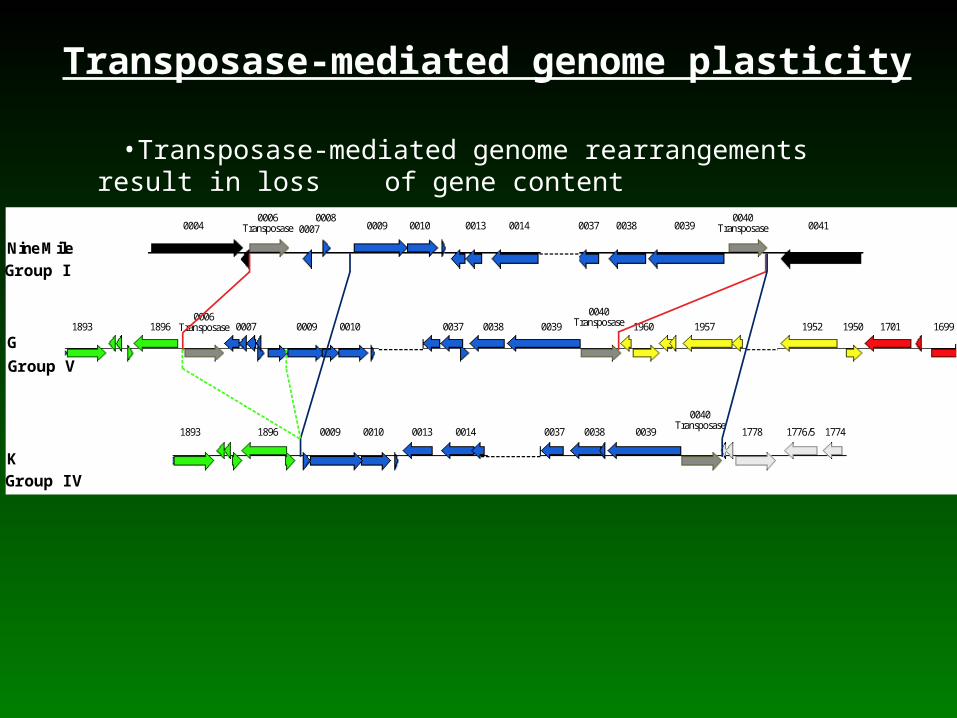
Transposase-mediated genome plasticity
Nine Mile
G
K
0040Transposase
00140013 0039003800370009 001018961893 1778 1776/5 1774
0039003800370009 001018961893 0007 1960 1957 1952 1950 1701 1699
0040Transposase0006
Transposase
0006Transposase
0040Transposase0009 0010
00080007 0039003800370004 004100140013
Group I
Group V
Group IV
•Transposase-mediated genome rearrangements result in loss of gene content
Gro
up I
Gro
up II
I
Gro
up IV
Gro
up V
I
Gro
up V
BD
TGro
up II
Key:Present ORFAbsent ORFPartial ORFORF containing a PM or small insertion
Beare et al, J Bact, 2006
Du
5G61
-63
Q32
1Le
Bru
ges
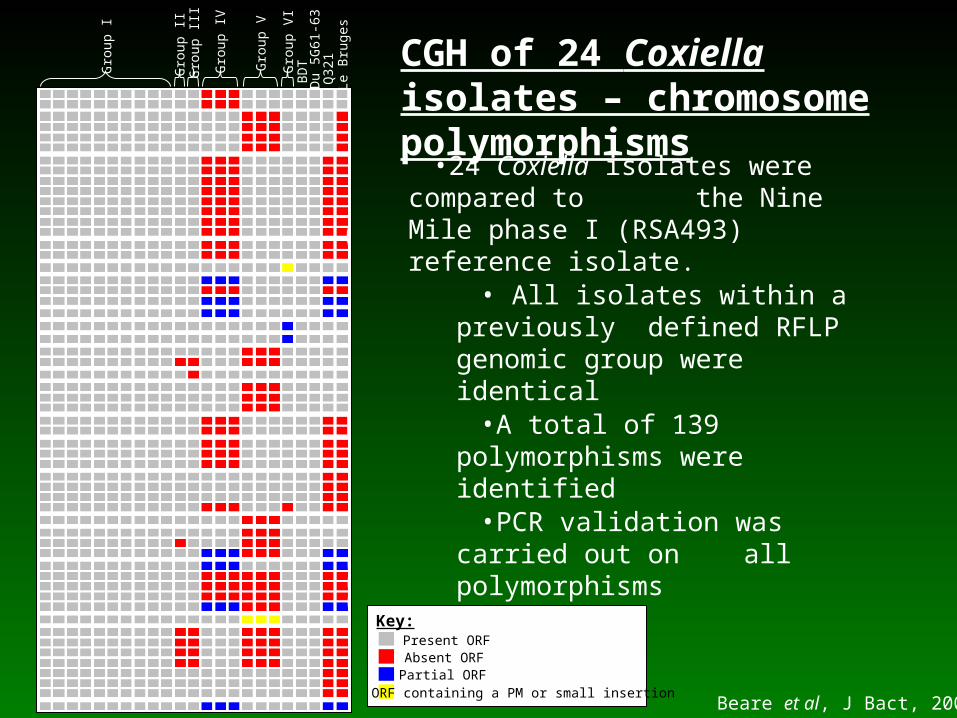
•24 Coxiella isolates were compared to the Nine Mile phase I (RSA493) reference isolate.
• All isolates within a previously defined RFLP genomic group
were identical•A total of 139 polymorphisms
were identified•PCR validation was carried out on all polymorphisms
CGH of 24 Coxiella isolates – chromosome polymorphisms

Gro
up I
Gro
up II
I
Gro
up IV
Gro
up V
I
Gro
up V
Gro
up II
QpH
1
QpH
1
QpH
1
QpH
1
QpD
V
QpD
G
IPQpR
S
•The BDT and Dugway 5G61-63 have the same profile as genomic group I and contain the QpH1
plasmid
•Q321 and Le Bruges both have the same unique profile, suggesting that Le Bruges has the QpDV plasmid
Beare et al, J Bact, 2006
Key:Present ORFAbsent ORFPartial ORFORF containing a PM or small insertion
BD
TD
u 5G
61-6
3Q
321
Le B
ruge
s
CGH of 24 Coxiella isolates – plasmid polymorphisms
Phylogenetic analysis of Coxiella isolates
•The BDT and Du 5G61-63 isolates reside in the genomic group I
•Q321 and Le Bruges represent two new genomic groups, VII and
VIII respectively
Beare et al, J Bact, 2006

Functions of polymorphic genes
• 67% of polymorphic ORFs encode hypothetical proteins
• 12% of polymorphic ORFs reside in pseudogenes
• The remaining ORFs have a range of functions;
• Ankyrin repeat proteins
• Transporters
• Transcriptional regulators
• Proteins that are part of large gene families
CGH analysis of small DNA samples
•Single Coxiella containing vacuoles were isolated by micropipette
•DNA contained in the vacuole suspension was amplified using a phi29 polymerase-based whole genome amplification (WGA) kit
VERO cell monolayer infected with Coxiella
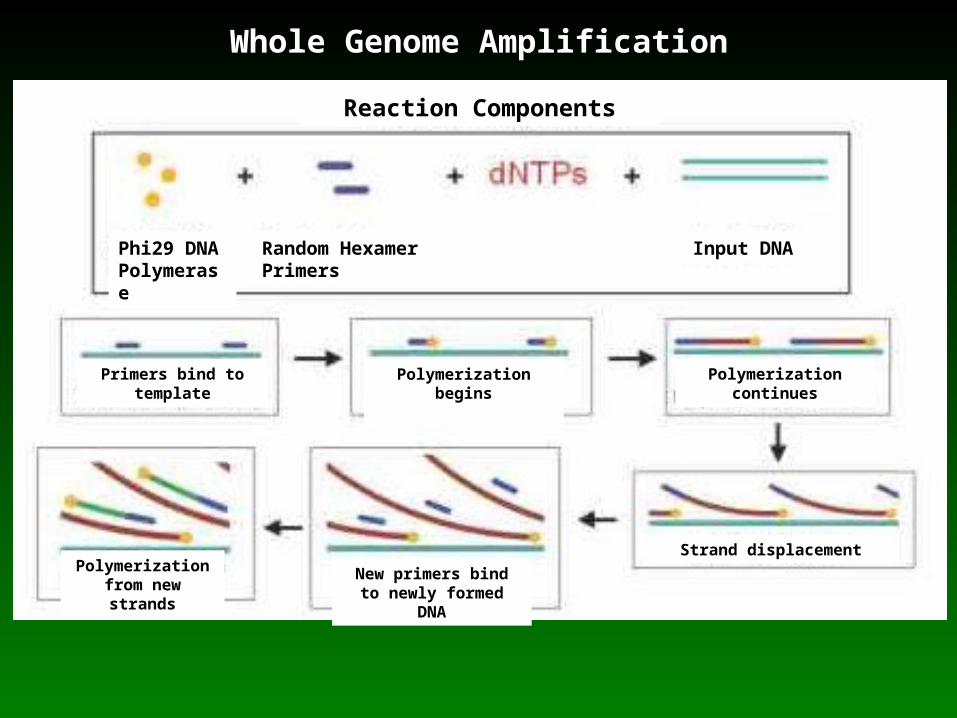
Whole Genome Amplification
Phi29 DNAPolymerase
Random Hexamer Primers
Input DNA
Primers bind to template
Polymerization begins
Polymerization from new strands
Polymerization continues
New primers bind to newly formed DNA
Strand displacement
Reaction Components
CGH analysis of small DNA samples
•Single Coxiella containing vacuoles were isolated by micropipette
•DNA contained in the vacuole suspension was amplified using a phi29 polymerase-based whole genome amplification (WGA) kit
•Following DNA precipitation, 10 g of DNA was hybridized to the Coxiella microarray
•Results obtained using WGA DNA were identical to those of non- amplified genomic DNA
VERO cell monolayer infected with Coxiella

Coxiella lipopolysaccharide (LPS) phase variation
•LPS is the only defined Coxiella virulence factor
•Virulent phase I have full length LPS
•Avirulent phase II have severely truncated LPS
•In vitro serial passing of virulent phase I results in avirulent phase II

LPS phase variation in Coxiella
•Nine Mile phase I (NMI) has a full length LPS and is virulent
•Two cloned variants of NMI have been isolated
•Nine Mile Crazy (NMC) has an intermediate LPS length and intermediate virulence
•Nine Mile phase II (NMII) has a severely truncated LPS and is avirulent
66.2
31.0
45.0
21.5
14.4
NM
I
NM
II
NM
C

66.2
31.0
45.0
21.5
14.4
NM
I
NM
II
NM
C
CGH analysis of Coxiella LPS variants
NM
IN
MII
NM
C
Region containing genes involved in LPS O-antigen biosynthesis
•CGH analysis of the NMII and NMC variants indicated the presence of only a single large genome deletion, which has been previously described (Hoover et al, Infect. Immun., 2002)
•CGH could not explain the why NMII, with a smaller deletion, produces a more truncated LPS
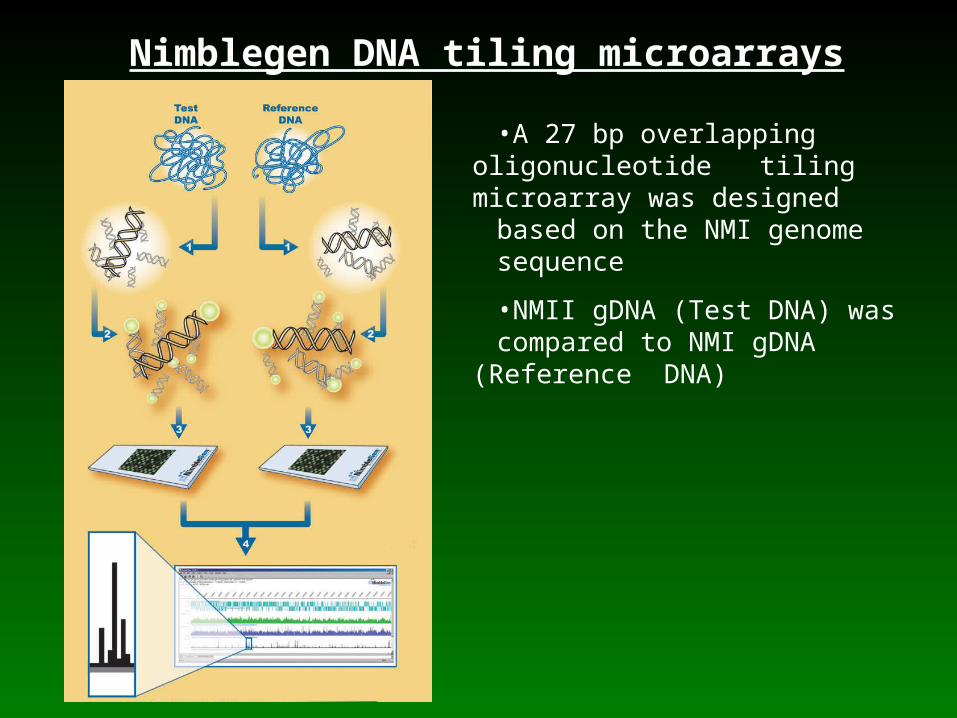
Nimblegen DNA tiling microarrays
•A 27 bp overlapping oligonucleotide tiling microarray was designed based on the NMI genome
sequence
•NMII gDNA (Test DNA) was compared to NMI gDNA (Reference
DNA)
Analysis of LPS variation - Tiling microarray of NMII
NLGNLFFLGDLKIGLWAIPITVIVVLANINAMNMIDGQDGLAGGVALGQALLLLLVSVKL 177
NLGNLFFLGDLKIGLWAIPITVIVVLANINAMNMIDGQDGLAGGVALGQA-LLLLVSVKL 176
NMI
NMII
•NMII has 14 polymorphisms relative to NMI
•One polymorphism resides in an ORF (CBU0533) predicted to be involved in LPS biosynthesis (wecA)
•This polymorphism is a 3 bp deletion that results in an inframe deletion of L168

The WecA protein of Escherichia coli
•A UDP-N-acetylglucosamine (GlcNAc):undecaprenyl-phosphate (Und-P) GlcNAc-1-phosphate transferase
•Participates in the biosynthesis of O antigen LPS in many enteric bacteria
•Transfers GlcNAc 1-phosphate from UDP-GlcNAc onto Und-P to form Und-P-P-GlcNAc
•Product of this reaction serves as an acceptor for the addition of subsequent sugars to complete the biosynthesis of O antigen
•Deletion of wecA in E. coli results in a truncated LPS
Analysis of LPS variation - Tiling microarray of NMII
NLGNLFFLGDLKIGLWAIPITVIVVLANINAMNMIDGQDGLAGGVALGQALLLLLVSVKL 177
NLGNLFFLGDLKIGLWAIPITVIVVLANINAMNMIDGQDGLAGGVALGQA-LLLLVSVKL 176
SLGYIFGSWEMVLGPFGYFLTLFAVWAAINAFNMVDGIDGLLGGLSCVSFAAIGMILWFD 180
.** :* :: :* :. :*::.* * ***:**:** *** **:: . : ::
E. coli
NMI
NMII
•L168 is predicted to be in the transmembrane region (colored green) immediately downstream of the WecA active site
•Preliminary evidence in an E. coli wecA mutant indicates that the NMII WecA is not properly inserted into the inner membrane

Conclusions
•Genetic variation was observed within Coxiella isolates
•Coxiella isolates of unknown genetic backgrounds can be typed and the extent of their genetic variation relative to
NMI analyzed
•Transposases play an important role in Coxiella genetic variation
•Potential pathotype-specific genes have been identified
•Using WGA DNA, very small samples (~1 ng of total DNA) can be analyzed by CGH
•A mutation potentially involved in Coxiella LPS phase variation was identified
•The CBU0533 mutation is also found in a number of isolates producing a severely truncated LPS
Related Documents