CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, 1071-412X/01/$04.0010 DOI: 10.1128/CDLI.8.1.85–92.2001 Jan. 2001, p. 85–92 Vol. 8, No. 1 Copyright © 2001, American Society for Microbiology. All Rights Reserved. Genetic and Biochemical Characterization of Glycerol Uptake in Mycoplasma mycoides subsp. mycoides SC: Its Impact on H 2 O 2 Production and Virulence EDY M. VILEI AND JOACHIM FREY* Institute for Veterinary Bacteriology, University of Berne, CH-3012 Berne, Switzerland Received 14 August 2000/Returned for modification 6 October 2000/Accepted 23 October 2000 Highly virulent strains of Mycoplasma mycoides subsp. mycoides SC belonging to the African cluster contain an operon with the genes gtsA, gtsB, and gtsC, encoding membrane ATP binding cassette transporter proteins GtsA, GtsB, and GtsC, which are involved in glycerol transport. Strain Afade ´ from the African cluster incorporated [U- 14 C]glycerol with a time-dependent increase. The less virulent strain L2 of the European cluster, which lacks gtsB and gtsC, failed to incorporate glycerol. Antibodies against GtsB noncompetitively inhibited glycerol uptake. L-a-Glycerophosphate was not transported by M. mycoides subsp. mycoides SC. It is postulated to be synthesized by phosphorylation of glycerol during transport and subsequently metabolized further to dihydroxyacetone phosphate accompanied by release of H 2 O 2 . Peroxide production in glycerol- containing growth medium was high for the African strain Afade ´ but very low for the European strain L2. Virtually no H 2 O 2 was produced by both strains without glycerol. Hence, the efficient glycerol uptake system found in the virulent strain of the African cluster leads to a strong release of peroxide, a potential virulence factor which is lacking in the less virulent European strains. M. mycoides subsp. mycoides SC might have adopted, as a strategy for virulence, a highly efficient uptake system for glycerol which allows the production of an active metabolic intermediate that damages host cells. Mycoplasma mycoides subsp. mycoides small-colony type (SC) is the etiological agent of contagious bovine pleuropneu- monia (CBPP), a severe, highly contagious disease of cattle that has drastic economical and socioeconomic consequences (16, 39). The disease was eradicated in the middle of the 20th century in the industrialized continents, but it reemerged in Europe in the last two decades in a milder, more insidious form with a low mortality rate (30). Genetic typing methods using insertion elements IS1296 and IS1634 revealed two dis- tinct clonal lineages of M. mycoides subsp. mycoides SC, one containing strains from the reemerging European outbreaks and the other containing isolates from the African and Aus- tralian continents (14, 18, 44). Differences between strains of the European and the African-Australian clusters of M. my- coides subsp. mycoides SC were also evidenced serologically by detection of lipoprotein LppB exclusively in strains of the Af- rican-Australian cluster (43). Controlled differential experi- mental infections of cattle showed that strain L2, a represen- tative strain of the European cluster isolated from the recent reemerging outbreaks, was significantly less virulent than the African strain Afade ´ (1). This confirmed the observations from outbreaks of CBPP in Africa and in Europe by Nicholas et al. (30) and showed that the difference in virulence of CBPP was due to differences of the strains. In spite of the high pathogenicity of M. mycoides subsp. mycoides SC and the enormous losses of livestock production caused by this mycoplasma worldwide, its virulence factors are virtually unknown. This is also true for other pathogenic my- coplasmas and is due to the difficulties encountered in micro- biology and genetics of mycoplasmas. So far, no classical vir- ulence factors such as toxins or invasins have been found in mycoplasmas, as revealed by the full genomic sequences of two species of this organism (17, 21). This might be due to their extremely small genome, leading mycoplasmas to a drastic economization in genetic resources, which are reduced to es- sential functions of life (34, 35). Mycoplasmas seem therefore to adopt endogenous structural and metabolic functions as virulence effectors to cause disease (42). Thus, membrane li- poproteins of several pathogenic mycoplasmas have been sug- gested as possible virulence factors due to their capability to induce blastogenesis and secretion of proinflammatory cyto- kines in vitro (11, 19, 33). Furthermore, the formation of active metabolic intermediates such as hydrogen peroxide (H 2 O 2 ) (15, 23, 28, 29, 38), galactan (13, 25), adhesins (9, 24), and variable surface-located membrane antigens (4, 46) has been suggested as a potential virulence attribute of mycoplasmas. Despite these many proposals, the impact of H 2 O 2 on viru- lence is not clear (27), and direct comparative genetic evidence explaining the basic differences between highly virulent and moderately virulent strains of M. mycoides subsp. mycoides SC is still lacking. Moreover, no tools for efficient gene transfer systems and genetic complementation experiments are avail- able for this pathogen. Because of their parasitic mode of life, mycoplasmas must acquire macromolecular precursors and high-energy com- pounds such as sugars from their environment in order to safeguard their life cycle and to produce active metabolic in- termediates. For this reason, a significant number of mycoplas- mal genes (about 30%) are devoted to adhesins and trans- porter proteins (34). Among the latter, ATP-binding cassette (ABC) transporters, which are membrane proteins ubiqui- * Corresponding author. Mailing address: Institute for Veterinary Bacteriology, University of Berne, La ¨nggass-Strasse 122, CH-3012 Berne, Switzerland. Phone: 41 31 631 2414. Fax: 41 31 631 2634. E-mail: [email protected]. 85

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY,1071-412X/01/$04.0010 DOI: 10.1128/CDLI.8.1.85–92.2001
Jan. 2001, p. 85–92 Vol. 8, No. 1
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Genetic and Biochemical Characterization of Glycerol Uptakein Mycoplasma mycoides subsp. mycoides SC: Its Impact on
H2O2 Production and VirulenceEDY M. VILEI AND JOACHIM FREY*
Institute for Veterinary Bacteriology, University of Berne, CH-3012 Berne, Switzerland
Received 14 August 2000/Returned for modification 6 October 2000/Accepted 23 October 2000
Highly virulent strains of Mycoplasma mycoides subsp. mycoides SC belonging to the African cluster containan operon with the genes gtsA, gtsB, and gtsC, encoding membrane ATP binding cassette transporter proteinsGtsA, GtsB, and GtsC, which are involved in glycerol transport. Strain Afade from the African clusterincorporated [U-14C]glycerol with a time-dependent increase. The less virulent strain L2 of the Europeancluster, which lacks gtsB and gtsC, failed to incorporate glycerol. Antibodies against GtsB noncompetitivelyinhibited glycerol uptake. L-a-Glycerophosphate was not transported by M. mycoides subsp. mycoides SC. It ispostulated to be synthesized by phosphorylation of glycerol during transport and subsequently metabolizedfurther to dihydroxyacetone phosphate accompanied by release of H2O2. Peroxide production in glycerol-containing growth medium was high for the African strain Afade but very low for the European strain L2.Virtually no H2O2 was produced by both strains without glycerol. Hence, the efficient glycerol uptake systemfound in the virulent strain of the African cluster leads to a strong release of peroxide, a potential virulencefactor which is lacking in the less virulent European strains. M. mycoides subsp. mycoides SC might haveadopted, as a strategy for virulence, a highly efficient uptake system for glycerol which allows the productionof an active metabolic intermediate that damages host cells.
Mycoplasma mycoides subsp. mycoides small-colony type(SC) is the etiological agent of contagious bovine pleuropneu-monia (CBPP), a severe, highly contagious disease of cattlethat has drastic economical and socioeconomic consequences(16, 39). The disease was eradicated in the middle of the 20thcentury in the industrialized continents, but it reemerged inEurope in the last two decades in a milder, more insidiousform with a low mortality rate (30). Genetic typing methodsusing insertion elements IS1296 and IS1634 revealed two dis-tinct clonal lineages of M. mycoides subsp. mycoides SC, onecontaining strains from the reemerging European outbreaksand the other containing isolates from the African and Aus-tralian continents (14, 18, 44). Differences between strains ofthe European and the African-Australian clusters of M. my-coides subsp. mycoides SC were also evidenced serologically bydetection of lipoprotein LppB exclusively in strains of the Af-rican-Australian cluster (43). Controlled differential experi-mental infections of cattle showed that strain L2, a represen-tative strain of the European cluster isolated from the recentreemerging outbreaks, was significantly less virulent than theAfrican strain Afade (1). This confirmed the observations fromoutbreaks of CBPP in Africa and in Europe by Nicholas et al.(30) and showed that the difference in virulence of CBPP wasdue to differences of the strains.
In spite of the high pathogenicity of M. mycoides subsp.mycoides SC and the enormous losses of livestock productioncaused by this mycoplasma worldwide, its virulence factors arevirtually unknown. This is also true for other pathogenic my-
coplasmas and is due to the difficulties encountered in micro-biology and genetics of mycoplasmas. So far, no classical vir-ulence factors such as toxins or invasins have been found inmycoplasmas, as revealed by the full genomic sequences of twospecies of this organism (17, 21). This might be due to theirextremely small genome, leading mycoplasmas to a drasticeconomization in genetic resources, which are reduced to es-sential functions of life (34, 35). Mycoplasmas seem thereforeto adopt endogenous structural and metabolic functions asvirulence effectors to cause disease (42). Thus, membrane li-poproteins of several pathogenic mycoplasmas have been sug-gested as possible virulence factors due to their capability toinduce blastogenesis and secretion of proinflammatory cyto-kines in vitro (11, 19, 33). Furthermore, the formation of activemetabolic intermediates such as hydrogen peroxide (H2O2)(15, 23, 28, 29, 38), galactan (13, 25), adhesins (9, 24), andvariable surface-located membrane antigens (4, 46) has beensuggested as a potential virulence attribute of mycoplasmas.Despite these many proposals, the impact of H2O2 on viru-lence is not clear (27), and direct comparative genetic evidenceexplaining the basic differences between highly virulent andmoderately virulent strains of M. mycoides subsp. mycoides SCis still lacking. Moreover, no tools for efficient gene transfersystems and genetic complementation experiments are avail-able for this pathogen.
Because of their parasitic mode of life, mycoplasmas mustacquire macromolecular precursors and high-energy com-pounds such as sugars from their environment in order tosafeguard their life cycle and to produce active metabolic in-termediates. For this reason, a significant number of mycoplas-mal genes (about 30%) are devoted to adhesins and trans-porter proteins (34). Among the latter, ATP-binding cassette(ABC) transporters, which are membrane proteins ubiqui-
* Corresponding author. Mailing address: Institute for VeterinaryBacteriology, University of Berne, Langgass-Strasse 122, CH-3012Berne, Switzerland. Phone: 41 31 631 2414. Fax: 41 31 631 2634.E-mail: [email protected].
85

tously present from bacteria to humans, are involved in theactive ATP-dependent transport of a broad variety of com-pounds, ranging from inorganic ions to large polypeptides (20).These transporters are proteins that are built from combina-tions of conservative domains like the ATP-binding ABC unitsand membrane-bound regions. These membrane proteins nor-mally function as transport ATPases, by hydrolyzing ATP incombination with transporting their substrate moleculesthrough cellular or intracellular membranes. The recently dis-covered major genetic difference between highly virulent Af-rican strains and significantly less virulent European strains ofM. mycoides subsp. mycoides SC, which consists of an 8.84-kbchromosome segment present uniquely in African-Australianstrains (43), revealed several open reading frames (ORFs) forpotential ABC transporters. We present the genetic basis foran efficient glycerol uptake system consisting of three ABCtransporters encoded on the genes gtsA, gtsB, and gtsC andpropose a biochemical pathway involved in the production ofH2O2. Its potential role in virulence of M. mycoides subsp.mycoides SC is discussed.
MATERIALS AND METHODS
Strains, growth conditions, and DNA extraction. The isolates of M. mycoidessubsp. mycoides SC used in this study were strain L2 (isolated recently by ourlaboratory from pathological lung material of a cattle with CBPP, received fromF. G. Santini, Teramo, Italy) as a representative of the European cluster andstrain Afade (isolated in Chad from a bovine from Northern Cameroon in 1969and received from F. Thiaucourt, CIRAD-EMVT, Montpellier, France) as arepresentative of the African-Australian cluster (14). Mycoplasmal cultures weremade in a standard medium (8) to a density of 108 to 109 cells/ml. Crude lysatepreparations with GES buffer (5 M guanidium thiocyanate, 100 mM EDTA,0.5% N-lauroylsarcosine) and DNA extraction were performed as previouslydescribed (14).
Southern blot analysis. Genomic mycoplasmal DNA was digested withHindIII, separated electrophoretically on a 0.7% agarose gel, and transferred toa positively charged nylon membrane (Boehringer Mannheim, Rotkreuz, Swit-zerland). The membrane was briefly rinsed with 13 SSC (150 mM NaCl, 15 mM
sodium citrate [pH 7.7]), and DNA was denatured at 80°C under vacuum for 30min (5). The membrane was preincubated with 10 ml of hybridization buffer,consisting of 53 SSC, 0.02% sodium dodecyl sulfate (SDS), 0.1% N-lauroylsar-cosine, and 1% blocking reagent (Boehringer Mannheim), at 68°C for 2 h andthen hybridized with 5 ml of hybridization buffer, containing 3 ml of digoxigenin-11-dUTP (DIG)-labeled probe, for 15 h at 68°C. The membrane was thenwashed twice for 5 min at room temperature with 23 SSC containing 0.1% SDSand twice for 15 min at room temperature with 0.23 SSC containing 0.1% SDS.The hybridized DIG-labeled probes were detected using phosphatase-labeledanti-DIG antibodies (Boehringer Mannheim) as described in the producer’sprotocol.
Sequencing strategies and gene cloning. DNA sequence analysis of segmentsadjacent to the locus involving the 8.84-kb deletion in European-cluster strains ofM. mycoides subsp. mycoides SC was performed using the Vectorette II System(Genosys Biotechnologies, London, United Kingdom). This system is based on aunidirectional PCR approach. The previously determined DNA sequence of the3,414-bp HindIII fragment (GenBank accession number AF165134) from strainL2 (43) was used to design specific primers. Briefly, genomic DNA from Afadewas digested with BamHI and ligated to Vectorette units. Thereafter, unidirec-tional PCR amplification using one primer specific for the Vectorette unitand primer ORF0-S3 (59-TTTGGCATCAAGTGCTGAAAATGGTTCATC-39), matching bases 3149 to 3178 of AF165134, produced a fragment of about 8kb, which was then sequenced with primer ORF0-S (59-TCTTTGTTTTTGACCACCAG-39), matching bases 3227 to 3246. From this new sequence, anotheroligonucleotide primer, ORF0-rev (59-ATTCAAAATGAGTTAAACAAGC-39), was designed and used with primer ORF0-S2 (59-CCACCAGATAACTGGTTAAC-39), matching bases 3240 to 3259 of AF165134, to produce a DIG-labeled ORF0-specific probe of 323 bp from the genomic DNA of Afade (Fig. 1).
To complete the sequence of ORF0, genomic HindIII fragments of M. my-coides subsp. mycoides SC strain L2 were cloned into vector pBluescriptII SK(2)(Stratagene, La Jolla, Calif.) and reacted by colony screening using the ORF0-specific probe by standard methods (5). Plasmid DNA of the positive colonieswas isolated using the QIAprep spin plasmid kit from Qiagen AG (Basle, Swit-zerland), digested with HindIII, and further verified by Southern blot hybridiza-tion with the ORF0-specific probe. Plasmid pJFFev1.5-L2(MP2.2), containing a1.5-kb HindIII insert with the 59 part of ORF0, was retained and sequenced.Double-stranded nested deletion, using exonuclease III (Pharmacia Biotech,Uppsala, Sweden), was carried out as specified by the manufacturer. The sub-clones obtained were sequenced with a DNA Sequenator AB310 and the TaqDye Deoxy Terminator cycle-sequencing kit (Perkin-Elmer, Norwalk, Conn.),using primers matching the T3 and T7 promoters of the vector and primersprogressively derived from sequenced segments.
FIG. 1. Genetic map of the locus containing the operon for the glycerol transport system in strain Afade. The genetic map of the locus in M.mycoides subsp. mycoides SC which differentiates European strains (top) from African cluster strains (middle) is shown. A magnification of theoperon for the glycerol transport system derived from sequence data (AF251037) is shown at the bottom. Open boxes indicate the copies of thetwo IS elements IS1296 and IS1634. Dotted boxes represent the 478-bp direct repeats created by the IS1634 insertion close to lppB in strain Afade.Boxes with arrowheads indicate ORFs found in the three DNA sequences. HindIII sites are represented by vertical bars marked H. Thin dottedlines align analogous genomic sections between the two different strains. The arrow in the lower part indicates the site of the deletion in strain L2,210 denotes the signal box of the transcription promoter, and the hairpin structure represents the potential transcription terminator signal. Thescales are in kilobase pairs.
86 VILEI AND FREY CLIN. DIAGN. LAB. IMMUNOL.

Production of monospecific rabbit antibodies against the glycerol uptakesystem and immunoblot analysis. To produce polyclonal antibodies against thepeptides encoded on the 39-terminal part of ORF2 or on ORF3, which are absentin the genome of European strains (43), their derived amino acid sequences wereanalyzed to deduce the best antigenic segment(s) for immunization. A peptidesegment of 25 amino acids from the C terminus of the glycerol transport proteinGtsB (ORF2) was calculated to have the best antigenicity and hydrophilicityscore, using the software ProtScale (http://www.expasy.ch/cgi-bin/protscale.pl)(7) and TMpred (http://www.ch.embnet.org/software/TMPRED_form.html)(22). Monospecific, polyclonal antibodies directed against the 25-amino-acidGtsB-derived peptide 1002 (IKSIYKLIKKVILKNRGKYESKNNN; synthesizedby E. Servi, Laboratoire des Peptides, Universite de Lausanne, Lausanne, Swit-zerland) were obtained by immunizing rabbits with 500 mg of GtsB peptide 1002in 500 ml of 0.85% (wt/vol) NaCl mixed with 500 ml of Freund’s completeadjuvant (Difco Laboratories, Detroit, Mich.) followed by a booster immuniza-tion with the same amount of protein in Freund’s incomplete adjuvant (DifcoLaboratories) 3 weeks later. The animals were bled 10 days after the boosterimmunization. Antiserum 89-d32 was prepared from the blood samples andstored at 220°C. The production of monospecific polyclonal rabbit antibodiesdirected against lipoprotein LppB was described previously (43).
Immunoblot analysis with antiserum 89-d32 (at a dilution of 1:1,000) wascarried out by standard methods (5), using SDS-polyacrylamide gel electrophore-sis (10% polyacrylamide) PAGE for separation of the proteins and phosphatase-labeled conjugate goat anti-rabbit immunoglobulin G (heavy plus light chains)(Kirkegaard & Perry Laboratories, Gaithersburg, Md.; catalogue no. 075-1506)at a dilution of 1:5,000.
Time-dependent glycerol- and G3P uptake assays. Measurements of time-dependent glycerol or L-a-glycerophosphate (G3P) incorporation into cellularmaterial of M. mycoides subsp. mycoides SC strains L2 and Afade were carriedout with radioactive substrates as described elsewhere (12, 28, 40). Briefly, 70-mlmycoplasmal cultures of both L2 and Afade, grown to a density of 108 to 109
cells/ml in standard medium (8), were centrifuged at 12,000 3 g for 10 min andthe cell pellets were washed and resuspended in 30 ml of isotonic HEPES buffer(67.6 mM HEPES [pH 7.3], 140 mM NaCl). Cell suspensions of 1.2 ml were thenadjusted to an optical density at 550 nm (OD550) of 1.0. After starvation for 1 hat 37°C, the isotonic buffer was adjusted to 6 mM G3P, 570 nM [U-14C]G3P(from a stock of 147 mCi/mmol; Amersham International plc, Little Chalfont,United Kingdom), 9 mM ATP, and 7 mM MgCl2 for G3P uptake. For glyceroluptake, the isotonic buffer was adjusted to 20 mM glycerol, 280 nM [U-14C]g-lycerol (from a stock of 149 mCi/mmol, Amersham International plc), and 7 mMMgCl2. Aliquots of 200 ml were vacuum filtered at different time intervals (30 s,1 min 45 s, 3 min, 4 min 15 s, and 5 min 30 s), and filters (pore size, 0.22 mm;Millipore, Bedford, Mass.) were washed immediately with HEPES buffer andfinally counted in a scintillation counter. It has to be noted that a minor amountof incorporated glycerol is not measured in this assay since it is catabolized andexcreted by the bacteria.
Kinetic studies for glycerol uptake in Afade. Concentration-dependent glyc-erol incorporation into cellular material of M. mycoides subsp. mycoides SC strainAfade was measured as described above. After starvation of the cells for 1 h at37°C in the isotonic buffer, the final glycerol concentration was adjusted to 0, 2,5, 10, 20, 50, 100, or 200 mM in eight different samples, using a 14C-labeled/unlabeled glycerol molar ratio of 1:357, and 7 mM MgCl2 (final concentration)was added. Aliquots of 200 ml were vacuum filtered after 4 min, and incorpora-tion of 14C-glycerol was measured as above. Inhibition assays with anti-GtsBserum 89-d32 (at a dilution of 1:100) were performed by preincubating cells withthe antibodies for 15 min at 37°C prior to addition of the substrate. As a control,preimmunization serum from the same rabbit was used.
Quantification of H2O2 production. Measurements of time-dependent H2O2
production in M. mycoides subsp. mycoides SC strains L2 and Afade were carriedout using the Merckoquant peroxide test (Merck KgaA, Darmstadt, Germany),which has a detection range of 0.5 to 25 mg of H2O2 per ml of solution. Briefly,30-ml mycoplasmal cultures of both L2 and Afade, grown to a density of ap-proximately 108 cells/ml (8), were centrifuged at 12,000 3 g for 10 min and thecell pellets were washed and resuspended in 10 ml of isotonic HEPES buffercontaining 7 mM MgCl2. From each strain, two cell suspensions of 1.0 ml wereadjusted to an OD550 of 1.0. After starvation for 1 h at 37°C, the isotonic bufferof one sample was adjusted to 100 mM glycerol while the buffer of the othersample was kept without glycerol. At different time intervals, ranging from 5 s to20 min, the test strips were dipped into the suspensions for 1 s and subsequentlyread.
Assessment of hemolytic activity. Mycoplasma strains L2 and Afade weregrown on PPLO agar plates containing 5% washed sheep erythrocytes (8).Cultures were incubated both aerobically and anaerobically at 37°C for 72 h. The
incubations were performed in the absence or in the presence of 1 mM glycerolin the growth medium. Hemolysis was estimated as described below.
Nucleotide sequence accession number. The EMBL/GenBank accession num-ber for the nucleotide sequence of the operon of 3,169 bp for the glyceroltransport system, consisting of gtsA, gtsB, and gtsC, from M. mycoides subsp.mycoides SC strain Afade, is AF251037.
RESULTS
Sequence analysis of the operon for three ABC transportersin M. mycoides subsp. mycoides SC. Cloning and DNA se-quence analysis of a genetic locus, specific to the virulentAfrican-cluster strains of M. mycoides subsp. mycoides SC, re-vealed an operon structure of 3,169 bp containing three ORFs,ORF0, ORF2, and ORF3, which are partially overlapping.These ORFs are preceded by a consensus sequence for a 210signal box of a prokaryotic transcription promoter and arefollowed by a sequence which can form a hairpin structure witha DG of 29.1 kcal/mol, representing a potential rho-indepen-dent transcription termination signal (Fig. 1). ORF0 encodes apeptide of 406 amino acids (aa) with a calculated molecularmass of 47.5 kDa. It is preceded by a canonical ribosomebinding sequence 16 bp upstream of the ATG start codon.ORF0 contains three mycoplasma-specific UGATrp codons. Itis followed by ORF2, which encodes a 342-aa peptide with acalculated molecular mass of 39.8 kDa. ORF2 overlaps ORF0by 23 bp and contains three UGATrp codons. It is followed byORF3, encoding a peptide of 269 aa with a molecular mass of31.6 kDa. ORF3 overlaps ORF2 by 23 bp and contains sevenUGATrp codons. In contrast to the African strain Afade, theEuropean strain L2 has a truncated operon with only an intactORF0 and a partial ORF2. The A1T content of the 3.17-kbsegment is 80.5%, reflecting the A1T-rich genome character-istic for mycoplasmas.
A comparison of the amino acid sequence of ORF0, ORF2,and ORF3 with the complete EMBL/GenBank database forsearch of related sequences was performed using the NationalCenter for Biotechnology Information BLASTP program (3).ORF0 was found to be very similar (28% identical and 50%similar amino acids) to a hypothetical ABC transporter fromM. genitalium, MG187, assumed to be involved in the G3Ptransport system. ORF2 has 25% identical and 44% similaramino acids to the probable ABC transporter MG188 from M.genitalium, and ORF3 has 24% identical and 48% similaramino acids to MG189, another hypothetical ABC transporterof M. genitalium (17).
Using ScanProsite software (http://www.expasy.ch/tools/scnpsit1.html), the protein encoded by ORF0 revealed anATP/GTP binding-site motif A (P-loop) at aa 48 to 55 and anABC transporter family signature at aa 213 to 227. ORF2 andORF3 did not show particular features for ABC transporters.However, TMpred software (http://www.ch.embnet.org/software/TMPRED_form.html) predicted that ORF2 andORF3 consist each of six transmembrane domains.
Expression of the ABC transporter protein in M. mycoidessubsp. mycoides SC. To assess the ABC cassette protein en-coded on ORF2, total antigen preparations of M. mycoidessubsp. mycoides SC strain Afade (African cluster) and strain L2(European cluster) were analyzed on immunoblots usingmonospecific polyclonal antibodies from rabbit serum 89-d32directed against synthetic peptide 1002 derived from a strong
VOL. 8, 2001 GLYCEROL UPTAKE IN M. MYCOIDES SUBSP. MYCOIDES SC 87
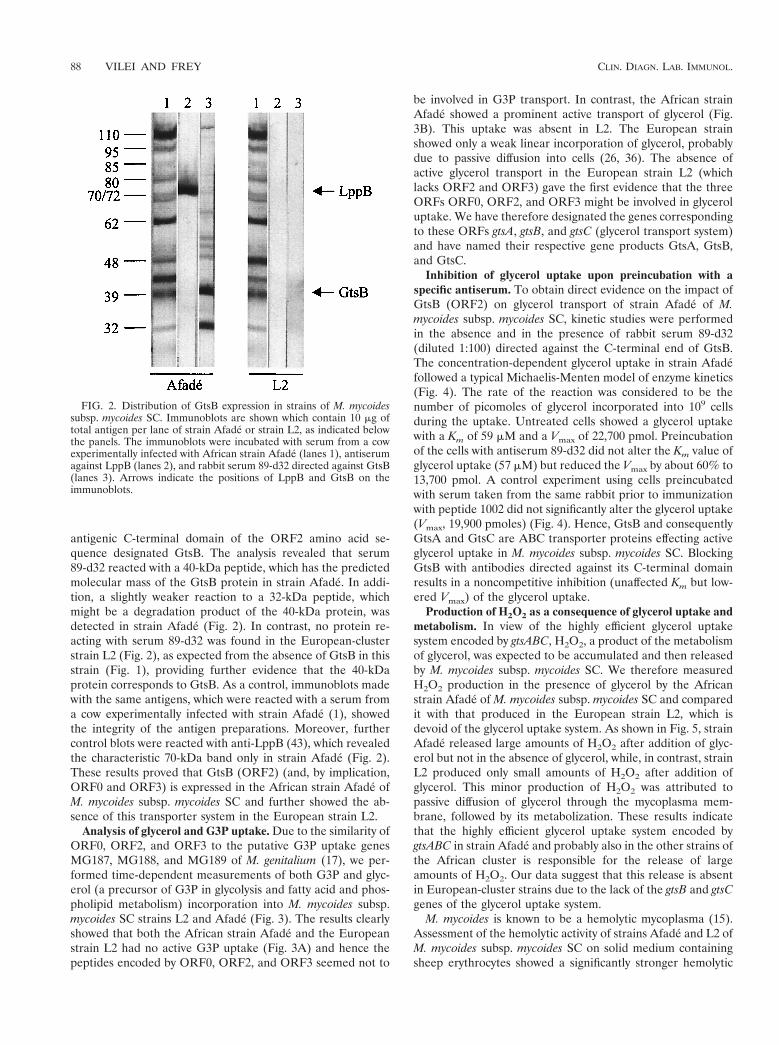
antigenic C-terminal domain of the ORF2 amino acid se-quence designated GtsB. The analysis revealed that serum89-d32 reacted with a 40-kDa peptide, which has the predictedmolecular mass of the GtsB protein in strain Afade. In addi-tion, a slightly weaker reaction to a 32-kDa peptide, whichmight be a degradation product of the 40-kDa protein, wasdetected in strain Afade (Fig. 2). In contrast, no protein re-acting with serum 89-d32 was found in the European-clusterstrain L2 (Fig. 2), as expected from the absence of GtsB in thisstrain (Fig. 1), providing further evidence that the 40-kDaprotein corresponds to GtsB. As a control, immunoblots madewith the same antigens, which were reacted with a serum froma cow experimentally infected with strain Afade (1), showedthe integrity of the antigen preparations. Moreover, furthercontrol blots were reacted with anti-LppB (43), which revealedthe characteristic 70-kDa band only in strain Afade (Fig. 2).These results proved that GtsB (ORF2) (and, by implication,ORF0 and ORF3) is expressed in the African strain Afade ofM. mycoides subsp. mycoides SC and further showed the ab-sence of this transporter system in the European strain L2.
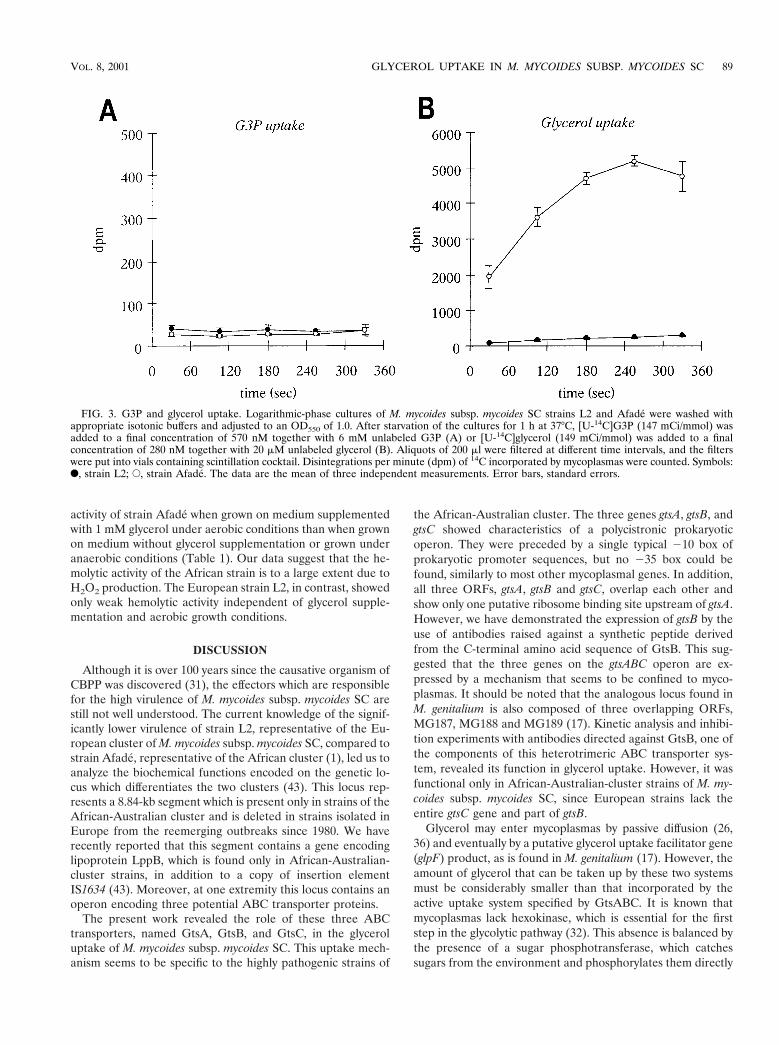
Analysis of glycerol and G3P uptake. Due to the similarity ofORF0, ORF2, and ORF3 to the putative G3P uptake genesMG187, MG188, and MG189 of M. genitalium (17), we per-formed time-dependent measurements of both G3P and glyc-erol (a precursor of G3P in glycolysis and fatty acid and phos-pholipid metabolism) incorporation into M. mycoides subsp.mycoides SC strains L2 and Afade (Fig. 3). The results clearlyshowed that both the African strain Afade and the Europeanstrain L2 had no active G3P uptake (Fig. 3A) and hence thepeptides encoded by ORF0, ORF2, and ORF3 seemed not to
be involved in G3P transport. In contrast, the African strainAfade showed a prominent active transport of glycerol (Fig.3B). This uptake was absent in L2. The European strainshowed only a weak linear incorporation of glycerol, probablydue to passive diffusion into cells (26, 36). The absence ofactive glycerol transport in the European strain L2 (whichlacks ORF2 and ORF3) gave the first evidence that the threeORFs ORF0, ORF2, and ORF3 might be involved in glyceroluptake. We have therefore designated the genes correspondingto these ORFs gtsA, gtsB, and gtsC (glycerol transport system)and have named their respective gene products GtsA, GtsB,and GtsC.
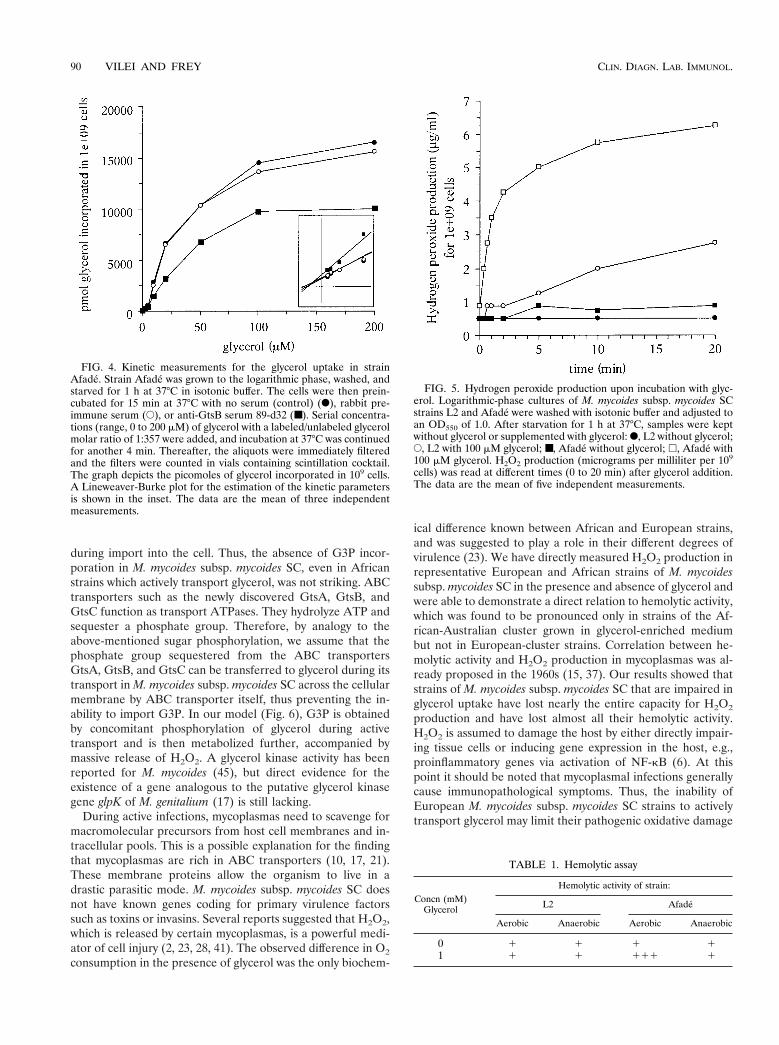
Inhibition of glycerol uptake upon preincubation with aspecific antiserum. To obtain direct evidence on the impact ofGtsB (ORF2) on glycerol transport of strain Afade of M.mycoides subsp. mycoides SC, kinetic studies were performedin the absence and in the presence of rabbit serum 89-d32(diluted 1:100) directed against the C-terminal end of GtsB.The concentration-dependent glycerol uptake in strain Afadefollowed a typical Michaelis-Menten model of enzyme kinetics(Fig. 4). The rate of the reaction was considered to be thenumber of picomoles of glycerol incorporated into 109 cellsduring the uptake. Untreated cells showed a glycerol uptakewith a Km of 59 mM and a Vmax of 22,700 pmol. Preincubationof the cells with antiserum 89-d32 did not alter the Km value ofglycerol uptake (57 mM) but reduced the Vmax by about 60% to13,700 pmol. A control experiment using cells preincubatedwith serum taken from the same rabbit prior to immunizationwith peptide 1002 did not significantly alter the glycerol uptake(Vmax, 19,900 pmoles) (Fig. 4). Hence, GtsB and consequentlyGtsA and GtsC are ABC transporter proteins effecting activeglycerol uptake in M. mycoides subsp. mycoides SC. BlockingGtsB with antibodies directed against its C-terminal domainresults in a noncompetitive inhibition (unaffected Km but low-ered Vmax) of the glycerol uptake.
Production of H2O2 as a consequence of glycerol uptake andmetabolism. In view of the highly efficient glycerol uptakesystem encoded by gtsABC, H2O2, a product of the metabolismof glycerol, was expected to be accumulated and then releasedby M. mycoides subsp. mycoides SC. We therefore measuredH2O2 production in the presence of glycerol by the Africanstrain Afade of M. mycoides subsp. mycoides SC and comparedit with that produced in the European strain L2, which isdevoid of the glycerol uptake system. As shown in Fig. 5, strainAfade released large amounts of H2O2 after addition of glyc-erol but not in the absence of glycerol, while, in contrast, strainL2 produced only small amounts of H2O2 after addition ofglycerol. This minor production of H2O2 was attributed topassive diffusion of glycerol through the mycoplasma mem-brane, followed by its metabolization. These results indicatethat the highly efficient glycerol uptake system encoded bygtsABC in strain Afade and probably also in the other strains ofthe African cluster is responsible for the release of largeamounts of H2O2. Our data suggest that this release is absentin European-cluster strains due to the lack of the gtsB and gtsCgenes of the glycerol uptake system.
M. mycoides is known to be a hemolytic mycoplasma (15).Assessment of the hemolytic activity of strains Afade and L2 ofM. mycoides subsp. mycoides SC on solid medium containingsheep erythrocytes showed a significantly stronger hemolytic
FIG. 2. Distribution of GtsB expression in strains of M. mycoidessubsp. mycoides SC. Immunoblots are shown which contain 10 mg oftotal antigen per lane of strain Afade or strain L2, as indicated belowthe panels. The immunoblots were incubated with serum from a cowexperimentally infected with African strain Afade (lanes 1), antiserumagainst LppB (lanes 2), and rabbit serum 89-d32 directed against GtsB(lanes 3). Arrows indicate the positions of LppB and GtsB on theimmunoblots.
88 VILEI AND FREY CLIN. DIAGN. LAB. IMMUNOL.
activity of strain Afade when grown on medium supplementedwith 1 mM glycerol under aerobic conditions than when grownon medium without glycerol supplementation or grown underanaerobic conditions (Table 1). Our data suggest that the he-molytic activity of the African strain is to a large extent due toH2O2 production. The European strain L2, in contrast, showedonly weak hemolytic activity independent of glycerol supple-mentation and aerobic growth conditions.
DISCUSSION
Although it is over 100 years since the causative organism ofCBPP was discovered (31), the effectors which are responsiblefor the high virulence of M. mycoides subsp. mycoides SC arestill not well understood. The current knowledge of the signif-icantly lower virulence of strain L2, representative of the Eu-ropean cluster of M. mycoides subsp. mycoides SC, compared tostrain Afade, representative of the African cluster (1), led us toanalyze the biochemical functions encoded on the genetic lo-cus which differentiates the two clusters (43). This locus rep-resents a 8.84-kb segment which is present only in strains of theAfrican-Australian cluster and is deleted in strains isolated inEurope from the reemerging outbreaks since 1980. We haverecently reported that this segment contains a gene encodinglipoprotein LppB, which is found only in African-Australian-cluster strains, in addition to a copy of insertion elementIS1634 (43). Moreover, at one extremity this locus contains anoperon encoding three potential ABC transporter proteins.
The present work revealed the role of these three ABCtransporters, named GtsA, GtsB, and GtsC, in the glyceroluptake of M. mycoides subsp. mycoides SC. This uptake mech-anism seems to be specific to the highly pathogenic strains of
the African-Australian cluster. The three genes gtsA, gtsB, andgtsC showed characteristics of a polycistronic prokaryoticoperon. They were preceded by a single typical 210 box ofprokaryotic promoter sequences, but no 235 box could befound, similarly to most other mycoplasmal genes. In addition,all three ORFs, gtsA, gtsB and gtsC, overlap each other andshow only one putative ribosome binding site upstream of gtsA.However, we have demonstrated the expression of gtsB by theuse of antibodies raised against a synthetic peptide derivedfrom the C-terminal amino acid sequence of GtsB. This sug-gested that the three genes on the gtsABC operon are ex-pressed by a mechanism that seems to be confined to myco-plasmas. It should be noted that the analogous locus found inM. genitalium is also composed of three overlapping ORFs,MG187, MG188 and MG189 (17). Kinetic analysis and inhibi-tion experiments with antibodies directed against GtsB, one ofthe components of this heterotrimeric ABC transporter sys-tem, revealed its function in glycerol uptake. However, it wasfunctional only in African-Australian-cluster strains of M. my-coides subsp. mycoides SC, since European strains lack theentire gtsC gene and part of gtsB.
Glycerol may enter mycoplasmas by passive diffusion (26,36) and eventually by a putative glycerol uptake facilitator gene(glpF) product, as is found in M. genitalium (17). However, theamount of glycerol that can be taken up by these two systemsmust be considerably smaller than that incorporated by theactive uptake system specified by GtsABC. It is known thatmycoplasmas lack hexokinase, which is essential for the firststep in the glycolytic pathway (32). This absence is balanced bythe presence of a sugar phosphotransferase, which catchessugars from the environment and phosphorylates them directly
FIG. 3. G3P and glycerol uptake. Logarithmic-phase cultures of M. mycoides subsp. mycoides SC strains L2 and Afade were washed withappropriate isotonic buffers and adjusted to an OD550 of 1.0. After starvation of the cultures for 1 h at 37°C, [U-14C]G3P (147 mCi/mmol) wasadded to a final concentration of 570 nM together with 6 mM unlabeled G3P (A) or [U-14C]glycerol (149 mCi/mmol) was added to a finalconcentration of 280 nM together with 20 mM unlabeled glycerol (B). Aliquots of 200 ml were filtered at different time intervals, and the filterswere put into vials containing scintillation cocktail. Disintegrations per minute (dpm) of 14C incorporated by mycoplasmas were counted. Symbols:F, strain L2; E, strain Afade. The data are the mean of three independent measurements. Error bars, standard errors.
VOL. 8, 2001 GLYCEROL UPTAKE IN M. MYCOIDES SUBSP. MYCOIDES SC 89
during import into the cell. Thus, the absence of G3P incor-poration in M. mycoides subsp. mycoides SC, even in Africanstrains which actively transport glycerol, was not striking. ABCtransporters such as the newly discovered GtsA, GtsB, andGtsC function as transport ATPases. They hydrolyze ATP andsequester a phosphate group. Therefore, by analogy to theabove-mentioned sugar phosphorylation, we assume that thephosphate group sequestered from the ABC transportersGtsA, GtsB, and GtsC can be transferred to glycerol during itstransport in M. mycoides subsp. mycoides SC across the cellularmembrane by ABC transporter itself, thus preventing the in-ability to import G3P. In our model (Fig. 6), G3P is obtainedby concomitant phosphorylation of glycerol during activetransport and is then metabolized further, accompanied bymassive release of H2O2. A glycerol kinase activity has beenreported for M. mycoides (45), but direct evidence for theexistence of a gene analogous to the putative glycerol kinasegene glpK of M. genitalium (17) is still lacking.
During active infections, mycoplasmas need to scavenge formacromolecular precursors from host cell membranes and in-tracellular pools. This is a possible explanation for the findingthat mycoplasmas are rich in ABC transporters (10, 17, 21).These membrane proteins allow the organism to live in adrastic parasitic mode. M. mycoides subsp. mycoides SC doesnot have known genes coding for primary virulence factorssuch as toxins or invasins. Several reports suggested that H2O2,which is released by certain mycoplasmas, is a powerful medi-ator of cell injury (2, 23, 28, 41). The observed difference in O2
consumption in the presence of glycerol was the only biochem-
ical difference known between African and European strains,and was suggested to play a role in their different degrees ofvirulence (23). We have directly measured H2O2 production inrepresentative European and African strains of M. mycoidessubsp. mycoides SC in the presence and absence of glycerol andwere able to demonstrate a direct relation to hemolytic activity,which was found to be pronounced only in strains of the Af-rican-Australian cluster grown in glycerol-enriched mediumbut not in European-cluster strains. Correlation between he-molytic activity and H2O2 production in mycoplasmas was al-ready proposed in the 1960s (15, 37). Our results showed thatstrains of M. mycoides subsp. mycoides SC that are impaired inglycerol uptake have lost nearly the entire capacity for H2O2
production and have lost almost all their hemolytic activity.H2O2 is assumed to damage the host by either directly impair-ing tissue cells or inducing gene expression in the host, e.g.,proinflammatory genes via activation of NF-kB (6). At thispoint it should be noted that mycoplasmal infections generallycause immunopathological symptoms. Thus, the inability ofEuropean M. mycoides subsp. mycoides SC strains to activelytransport glycerol may limit their pathogenic oxidative damage
FIG. 4. Kinetic measurements for the glycerol uptake in strainAfade. Strain Afade was grown to the logarithmic phase, washed, andstarved for 1 h at 37°C in isotonic buffer. The cells were then prein-cubated for 15 min at 37°C with no serum (control) (F), rabbit pre-immune serum (E), or anti-GtsB serum 89-d32 (■). Serial concentra-tions (range, 0 to 200 mM) of glycerol with a labeled/unlabeled glycerolmolar ratio of 1:357 were added, and incubation at 37°C was continuedfor another 4 min. Thereafter, the aliquots were immediately filteredand the filters were counted in vials containing scintillation cocktail.The graph depicts the picomoles of glycerol incorporated in 109 cells.A Lineweaver-Burke plot for the estimation of the kinetic parametersis shown in the inset. The data are the mean of three independentmeasurements.
FIG. 5. Hydrogen peroxide production upon incubation with glyc-erol. Logarithmic-phase cultures of M. mycoides subsp. mycoides SCstrains L2 and Afade were washed with isotonic buffer and adjusted toan OD550 of 1.0. After starvation for 1 h at 37°C, samples were keptwithout glycerol or supplemented with glycerol: F, L2 without glycerol;E, L2 with 100 mM glycerol; ■, Afade without glycerol; h, Afade with100 mM glycerol. H2O2 production (micrograms per milliliter per 109
cells) was read at different times (0 to 20 min) after glycerol addition.The data are the mean of five independent measurements.
TABLE 1. Hemolytic assay
Concn (mM)Glycerol
Hemolytic activity of strain:
L2 Afade
Aerobic Anaerobic Aerobic Anaerobic
0 1 1 1 11 1 1 111 1
90 VILEI AND FREY CLIN. DIAGN. LAB. IMMUNOL.

by H2O2 production and could explain the reduced morbidityand mortality of CBPP in Europe. Glycerol is metabolized viaG3P oxidase (a-GPO) in a reaction which yields 1 mol of H2O2
per mol of G3P oxidized. This previously led to speculationsthat the lack of the H2O2-producing enzyme a-GPO was re-sponsible for the reduced pathogenicity of some M. mycoidessubsp. mycoides SC strains (28, 45). The present work shows,however, that the lack of H2O2 production by certain strains ofM. mycoides subsp. mycoides SC such as the European-clusterstrains was due to the lack of an active glycerol uptake systemencoded on the gtsABC operon.
Summarizing, our results show that the Australian-African-cluster strains of M. mycoides subsp. mycoides SC are able toimport glycerol, a precursor of H2O2 while the less virulentEuropean cluster strains are devoid of this function, due to adeletion in the operon encoding the glycerol uptake system.Although the present results do not permit a direct comparisonof isogenic strains and complementation experiments whichare technically not possible in this mycoplasma species, weconclude that the lack of H2O2 production in the Europeancluster could be one of the reasons for its lower virulence.Consequently, the highly efficient glycerol uptake system al-lowing the production of H2O2, which damages host cells,might be a potential virulence attribute of M. mycoides subsp.mycoides SC.
ACKNOWLEDGMENTS
We are grateful to J. Nicolet for support and numerous discussionson this project, to M. Krawinkler for expert help with preparations ofmycoplasma cultures, and to Y. Schlatter for technical assistance with
DNA sequence analysis. We thank F. Santini (Istituto ZooprofilatticoSperimentale dell’Abruzzo e del Molise, Teramo, Italy) and F. Thiau-court (CIRAD-EMVT, Montpellier, France) for gifts of strains.
This study is part of the European COST action 826 on ruminantmycoplasmoses and was supported by grant C96.0073 from the SwissMinistry of Education and Science and by the Swiss Federal VeterinaryOffice.
REFERENCES
1. Abdo, E.-M., J. Nicolet, R. Miserez, R. Goncalves, J. Regalla, C. Griot, A.Bensaide, M. Krampe, and J. Frey. 1998. Humoral and bronchial immuneresponses in cattle experimentally infected with Mycoplasma mycoides subsp.mycoides small colony type. Vet. Microbiol. 59:109–122.
2. Abu-Groun, E. A., R. R. Taylor, H. Varsani, B. J. Wadher, R. H. Leach, andR. Miles. 1994. Biochemical diversity within the “Mycoplasma mycoides”cluster. Microbiology 140:2033–2042.
3. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
4. Atkin, C. L., S. Wei, and B. C. Cole. 1994. The Mycoplasma arthritidissuperantigen MAM: purification and identification of an active peptide.Infect. Immun. 62:5367–5375.
5. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A.Smith, and K. Struhl. 1999. Current protocols in molecular biology. JohnWiley & Sons, Inc., New York, N.Y.
6. Baeuerle, P. A., and T. Henkel. 1994. Function and activation of NF-kB inthe immune system. Annu. Rev. Immunol. 12:141–179.
7. Bairoch, A., P. Bucher, and K. Hofmann. 1995. The PROSITE database, itsstatus in 1995. Nucleic Acids Res. 24:189–196.
8. Bannerman, E. S., and J. Nicolet. 1971. Isolation and identification of por-cine Mycoplasma in Switzerland. Schweiz. Arch. Tierheilkd. 113:697–710.
9. Baseman, J. B., R. M. Cole, D. C. Krause, and D. K. Leith. 1982. Molecularbasis for cytadsorption of Mycoplasma pneumoniae. J. Bacteriol. 151:1514–1522.
10. Bork, P., C. Ouzounis, G. Casari, R. Schneider, C. Sander, M. Dolan, W.Gilbert, and P. M. Gillevet. 1995. Exploring the Mycoplasma capricolumgenome: a minimal cell reveals its physiology. Mol. Microbiol. 16:955–967.
11. Brenner, C., H. Wroblewski, M. Le Henaff, L. Montagnier, and A. Blan-chard. 1997. Spiralin, a mycoplasmal membrane lipoprotein, induces T-cell-independent B-cell blastogenesis and secretion of proinflammatory cyto-kines. Infect. Immun. 65:4322–4329.
12. Brzoska, P., M. Rimmele, K. Brzostek, and W. Boos. 1994. The pho regulon-dependent Ugp uptake system for glycerol-3-phosphate in Escherichia coli istrans inhibited by P. J. Bacteriol. 176:15–20.
13. Buttery, S. H., G. S. Cottew, and L. C. Lloyd. 1980. Effect of soluble factorsfrom Mycoplasma mycoides subsp. mycoides on the collagen content of bo-vine connective tissue. J. Comp. Pathol. 90:303–314.
14. Cheng, X., J. Nicolet, F. Poumarat, J. Regalla, F. Thiaucourt, and J. Frey.1995. Insertion element IS1296 in Mycoplasma mycoides subsp. mycoidessmall colony identifies a European clonal line distinct from African andAustralian strains. Microbiology 141:3221–3228.
15. Cole, B. C., J. R. Ward, and C. H. Martin. 1968. Hemolysin and peroxideactivity of Mycoplasma species. J. Bacteriol. 95:2022–2030.
16. Egwu, G. O., R. A. J. Nicholas, J. A. Ameh, and J. B. Bashiruddin. 1996.Contagious bovine pleuropneumonia: an update. Vet. Bull. 66:875–888.
17. Fraser, C. M., J. D. Gocayne, O. White, et al. 1995. The minimal genecomplement of Mycoplasma genitalium. Science 270:397–403.
18. Frey, J., X. Cheng, P. Kuhnert, and J. Nicolet. 1995. Identification andcharacterization of IS1296 in Mycoplasma mycoides subsp. mycoides SC andpresence in related mycoplasmas. Gene 160:95–100.
19. Herbelin, A., E. Ruuth, D. Delorme, C. Michel Herbelin, and F. Praz. 1994.Mycoplasma arginini TUH-14 membrane lipoproteins induce production ofinterleukin-1, interleukin-6, and tumor necrosis factor alpha by humanmonocytes. Infect. Immun. 62:4690–4694.
20. Higgins, C. F. 1992. ABC transporters: from microorganisms to man. Annu.Rev. Cell Biol. 8:67–113.
21. Himmelreich, R., H. Hilbert, H. Plagens, E. Pirkl, B. C. Li, and R. Herr-mann. 1996. Complete sequence analysis of the genome of the bacteriumMycoplasma pneumoniae. Nucleic Acids Res. 24:4420–4449.
22. Hofmann, K., and W. Stoffel. 1993. TMbase—a database of membranespanning proteins segments. Biol. Chem. Hoppe-Seyler 347:166.
23. Houshaymi, B. M., R. J. Miles, and R. A. Nicholas. 1997. Oxidation ofglycerol differentiates African from European isolates of Mycoplasma my-coides subspecies mycoides SC (small colony). Vet. Rec. 140:182–183.
24. Krause, D. C., and J. B. Baseman. 1983. Inhibition of mycoplasma pneu-moniae hemadsorption and adherence to respiratory epithelium by antibod-ies to a membrane protein. Infect. Immun. 39:1180–1186.
25. Lloyd, L. C., S. H. Buttery, and J. R. Hudson. 1971. The effect of the galactanand other antigens of Mycoplasma mycoides var. Mycoides on experimentalinfection with that organism in cattle. J. Med. Microbiol. 4:425–439.
26. McElhaney, R. N., J. de Gier, and E. C. van der Neut-Kok. 1973. The effectof alterations in fatty acid composition and cholesterol content on the non-
FIG. 6. Model for glycerol incorporation and H2O2 production inM. mycoides subsp. mycoides SC strain Afade. This model is a modi-fication of the model for sugar and central intermediate metabolism inM. pneumoniae reported by Himmelreich et al. (21). Glycerol is incor-porated into M. mycoides subsp. mycoides SC (African-Australian-cluster strains) through the proteins GtsA, GtsB, and GtsC encoded bythe gtsABC operon and phosphorylated during uptake. In the cyto-plasm, G3P is used as substrate by a-GPO to produce H2O2 from O2.The other product, dihydroxyacetone phosphate (DHAP), is metabo-lized in the glycolytic pathway in combination with glyceraldehyde-3-phosphate (GA3P).
VOL. 8, 2001 GLYCEROL UPTAKE IN M. MYCOIDES SUBSP. MYCOIDES SC 91

electrolyte permeability of Acholeplasma laidlawii B cells and derived lipo-somes. Biochim. Biophys. Acta 298:500–512.
27. Meier, B., and G. G. Habermehl. 1990. Evidence for superoxide dismutaseand catalase in mollicutes and release of reactive oxygen species. Arch.Biochem. Biophys. 277:74–79.
28. Miles, R. J., R. R. Taylor, and H. Varsani. 1991. Oxygen uptake and H2O2production by fermentative Mycoplasma spp. J. Med. Microbiol. 34:219–223.
29. Niang, M., R. F. Rosenbusch, M. C. Debey, Y. Niyo, J. J. Andrews, and M. L.Kaeberle. 1998. Field isolates of Mycoplasma ovipneumoniae exhibit distinctcytopathic effects in ovine tracheal organ cultures. J. Vet. Med. Ser. A45:29–40.
30. Nicholas, R. A. J., F. G. Santini, K. M. Clark, N. M. A. Palmer, P. DeSantis,and J. B. Bashiruddin. 1996. A comparison of serological tests and grosslung pathology for detecting contagious bovine pleuropneumonia in twogroups of Italian cattle. Vet. Rec. 139:89–93.
31. Nocard, E. I. E., and E. Roux. 1898. Le microbe de la peripneumonie. Ann.Inst. Pasteur 12:240–262.
32. Pollack, J. D. 1997. Mycoplasma genes: a case for reflective annotation.Trends Microbiol. 5:413–419.
33. Rawadi, G., and S. Roman-Roman. 1996. Mycoplasma membrane lipopro-teins induce proinflammatory cytokines by a mechanism distinct from that oflipopolysaccharide. Infect. Immun. 64:637–643.
34. Razin, S. 1997. The minimal cellular genome of Mycoplasma. Indian J. Bio-chem. Biophys. 34:124–130.
35. Razin, S., D. Yogev, and Y. Naot. 1998. Molecular biology and pathogenicityof mycoplasmas. Microbiol. Mol. Biol. Rev. 62:1094–1156.
36. Romijn, J. C., L. M. van Golde, R. N. McElhaney, and L. L. van Deenen.1972. Some studies on the fatty acid composition of total lipids and phos-phatidylglycerol from Acholeplasma laidlawii B and their relation to thepermeability of intact cells of this organism. Biochim. Biophys. Acta 280:22–32.
37. Somerson, N. L., B. E. Walls, and R. M. Chanock. 1965. Hemolysin ofMycoplasma pneumoniae: tentative identification as a peroxide. Science 150:226–228.
38. Taylor, R. R., K. Mohan, and R. J. Miles. 1996. Diversity of energy-yieldingsubstrates and metabolism in avian mycoplasmas. Vet. Microbiol. 51:291–304.
39. ter Laak, E. A. 1992. Contagious bovine pleuropneumonia. A review. Vet. Q.14:104–110.
40. Truniger, V., and W. Boos. 1993. Glycerol uptake in Escherichia coli issensitive to membrane lipid composition. Res. Microbiol. 144:565–574.
41. Tryon, V. V. and J. B. Baseman. 1992. Pathogenic determinants and mech-anisms, p. 457–471. In J. Maniloff, R. N. McElhaney, L. R. Finch, and J. B.Baseman (ed.), Mycoplasmas: molecular biology and pathogenesis. Ameri-can Society for Microbiology, Washington, D.C.
42. Valdivieso Garcia, A., S. Rosendal, O. B. Allen, C. M. Thompson, and S.Watson. 1989. Cytotoxicity of Mycoplasma mycoides subspecies mycoides forcultured endothelial cells. Int. J. Med. Microbiol. 272:202–209.
43. Vilei, E. M., E.-M. Abdo, J. Nicolet, A. Botelho, R. Goncalves, and J. Frey.2000. Genomic and antigenic differences between the European and African/Australian clusters of Mycoplasma mycoides subsp. mycoides SC. Microbiol-ogy 146:477–486.
44. Vilei, E. M., J. Nicolet, and J. Frey. 1999. IS1634, a novel insertion elementcreating long, variable-length direct repeats which is specific for Mycoplasmamycoides subsp. mycoides small-colony type. J. Bacteriol. 181:1319–1323.
45. Wadher, B. J., C. L. Henderson, R. J. Miles, and H. Varsani. 1990. A mutantof Mycoplasma mycoides subsp. mycoides lacking the H2O2-producing en-zyme L-a-glycerophosphate oxidase. FEMS Microbiol. Lett. 72:127–130.
46. Wise, K. S., D. Yogev, and R. Rosengarten. 1992. Antigenic variation, p.473–489. In J. Maniloff, R. N. McElhaney, L. R. Finch, and J. B. Baseman(ed.), Mycoplasmas: molecular biology and pathogenesis. American Societyfor Microbiology, Washington, D.C.
92 VILEI AND FREY CLIN. DIAGN. LAB. IMMUNOL.
Related Documents











