Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com Original Research Article Dement Geriatr Cogn Disord 2009;27:59–68 DOI: 10.1159/000191203 Genetic Analysis of Alzheimer’s Disease in the Uppsala Longitudinal Study of Adult Men Vilmantas Giedraitis a Lena Kilander a Malin Degerman-Gunnarsson a Johan Sundelöf a Tomas Axelsson b Ann-Christine Syvänen b Lars Lannfelt a Anna Glaser a a Molecular Geriatrics, Department of Public Health and Caring Sciences, and b Molecular Medicine, Department of Medical Sciences, Uppsala University, Uppsala, Sweden SNP at p ! 0.05 with association to AD. Only 20 genes re- mained significant after a permutation test to correct for multiple comparisons within individual genes. Using pub- licly available data from 2 genome-wide association (GWA) studies and linkage disequilibrium data from HapMap, we attempted to replicate the AD association identified in UL- SAM. In addition to apolipoprotein E, we were able to repli- cate 5 other genes in both GWA studies at p ! 0.05. Copyright © 2009 S. Karger AG, Basel Introduction Genetic factors influencing common complex condi- tions have proven difficult to identify, and data from nu- merous investigations have provided incomplete conclu- sions as to the identity of these genes. So far, only a few genes have been reproducibly associated to common dis- eases. The association of apolipoprotein E (APOE) to Alz- heimer’s disease (AD) has been convincingly and repeat- edly demonstrated in numerous studies [1] . Additional susceptibility genes for AD have been more elusive and studies aimed at identifying such genes have, prior to whole-genome approaches, mainly been focused on genes with a demonstrated or implied function related to de- Key Words Dementia Genetics of complex disorders Linkage and association analysis Alzheimer’s disease Apolipoprotein E Association study Gene polymorphism Longitudinal study Abstract Background/Aims: Genetic factors influencing common complex conditions have proven difficult to identify, and data from numerous investigations have provided incom- plete conclusions as to the identity of these genes. Here we aimed to identify susceptibility genes for late-onset Alz- heimer’s disease (AD). Methods: The case-control analysis included samples from 86 AD patients and 404 cognitively healthy controls selected from the Uppsala Longitudinal Study of Adult Men (ULSAM). In the incidence analysis, all 1,088 genotyped ULSAM participants were included. DNA samples from ULSAM participants were analyzed for 2,578 single nucleotide polymorphisms (SNP) within 368 genes. The selection of genes tested for association to AD within this cohort was based on genes previously implicated in conditions with relevance to ULSAM, such as dementia, car- diovascular disease, diabetes and metabolic syndrome, os- teoporosis, and cancer. Results/Conclusion: Association analysis revealed 82 genes containing at least 1 significant Accepted after revision: October 6, 2008 Published online: January 14, 2009 Vilmantas Giedraitis Public Health and Caring Sciences, Molecular Geriatrics, Uppsala University Dag Hammarskjölds väg 20, SE–751 85 Uppsala (Sweden) Tel. +46 18 471 5038, Fax +46 18 471 4808 E-Mail [email protected] © 2009 S. Karger AG, Basel 1420–8008/09/0271–0059$26.00/0 Accessible online at: www.karger.com/dem

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Fax +41 61 306 12 34E-Mail [email protected]
Original Research Article
Dement Geriatr Cogn Disord 2009;27:59–68 DOI: 10.1159/000191203
Genetic Analysis of Alzheimer’sDisease in the Uppsala Longitudinal Study of Adult Men
Vilmantas Giedraitis a Lena Kilander a Malin Degerman-Gunnarsson a
Johan Sundelöf a Tomas Axelsson b Ann-Christine Syvänen b Lars Lannfelt a
Anna Glaser a
a Molecular Geriatrics, Department of Public Health and Caring Sciences, and b Molecular Medicine,Department of Medical Sciences, Uppsala University, Uppsala , Sweden
SNP at p ! 0.05 with association to AD. Only 20 genes re-mained significant after a permutation test to correct for multiple comparisons within individual genes. Using pub-licly available data from 2 genome-wide association (GWA) studies and linkage disequilibrium data from HapMap, we attempted to replicate the AD association identified in UL-SAM. In addition to apolipoprotein E, we were able to repli-cate 5 other genes in both GWA studies at p ! 0.05.
Copyright © 2009 S. Karger AG, Basel
Introduction
Genetic factors influencing common complex condi-tions have proven difficult to identify, and data from nu-merous investigations have provided incomplete conclu-sions as to the identity of these genes. So far, only a few genes have been reproducibly associated to common dis-eases. The association of apolipoprotein E (APOE) to Alz-heimer’s disease (AD) has been convincingly and repeat-edly demonstrated in numerous studies [1] . Additional susceptibility genes for AD have been more elusive and studies aimed at identifying such genes have, prior to whole-genome approaches, mainly been focused on genes with a demonstrated or implied function related to de-
Key Words
Dementia � Genetics of complex disorders � Linkage and association analysis � Alzheimer’s disease � Apolipoprotein E � Association study � Gene polymorphism � Longitudinal study
Abstract
Background/Aims: Genetic factors influencing common complex conditions have proven difficult to identify, and data from numerous investigations have provided incom-plete conclusions as to the identity of these genes. Here we aimed to identify susceptibility genes for late-onset Alz-heimer’s disease (AD). Methods: The case-control analysis included samples from 86 AD patients and 404 cognitively healthy controls selected from the Uppsala Longitudinal Study of Adult Men (ULSAM). In the incidence analysis, all 1,088 genotyped ULSAM participants were included. DNA samples from ULSAM participants were analyzed for 2,578 single nucleotide polymorphisms (SNP) within 368 genes. The selection of genes tested for association to AD within this cohort was based on genes previously implicated in conditions with relevance to ULSAM, such as dementia, car-diovascular disease, diabetes and metabolic syndrome, os-teoporosis, and cancer. Results/Conclusion: Association analysis revealed 82 genes containing at least 1 significant
Accepted after revision: October 6, 2008 Published online: January 14, 2009
Vilmantas Giedraitis Public Health and Caring Sciences, Molecular Geriatrics, Uppsala University Dag Hammarskjölds väg 20, SE–751 85 Uppsala (Sweden) Tel. +46 18 471 5038, Fax +46 18 471 4808E-Mail [email protected]
© 2009 S. Karger AG, Basel1420–8008/09/0271–0059$26.00/0
Accessible online at:www.karger.com/dem

Giedraitis et al.
Dement Geriatr Cogn Disord 2009;27:59–68 60
mentia development or genes chromosomally positioned within linkage peaks correlated to AD [2, 3] . However, genes implicated in other complex conditions could also potentially be relevant to dementia. The APOE gene, for example, shows association to both cardiovascular dis-ease and AD [1, 4, 5] . There are also reports suggesting that body mass index (BMI) [6–8] and type 2 diabetes [9, 10] can be risk factors for AD. Although the molecular mechanisms underlying this correlation are largely un-known, one possible pathway could include insulin-de-grading enzyme [11] , which is involved in the degrada-tion of both insulin and amyloid- � (A � ), the latter being a central component in plaques that form in the brains of AD patients [12] . Furthermore, the Wnt signaling path-way, involving genes such as LRP1, 5 and 6, has been im-plicated in osteoporosis, cancer, neurodevelopment and neurodegenerative disorders, suggesting that these genes might be involved in mechanisms common to different complex conditions [13–15] .
The Uppsala Longitudinal Study of Adult Men (UL-SAM) was based on men born between 1920 and 1924 and living in Uppsala, Sweden, at the time of their 50th birthday [16] . These men have been regularly invited for health examinations over the years. The initial focus of ULSAM was on the identification of metabolic risk fac-tors for cardiovascular disease, but examinations have been expanded into other areas of complex traits. This includes significant efforts made to identify individuals diagnosed with different forms of dementia.
In the present study, we investigated the potential association of a large number of genes to AD within ULSAM. The analyzed genes were selected on the basis of being potentially relevant to dementia or to other com-plex conditions investigated within ULSAM. These con-ditions include dementia, diabetes and metabolic syn-drome, cardiovascular disease, osteoporosis, and can-cer.
Methods
Study Population We analyzed samples and data collected from ULSAM. This
study was initiated in 1970–1974, when all 50-year-old men living in Uppsala, Sweden, were invited for a health survey, initially fo-cusing on risk factors for cardiovascular disease. Out of a total of 2,841 men born in 1920–1924, 2,322 (82%) agreed to participate. Since then, the cohort has been re-examined at ages 60, 70, 77 and 82 years. With the aim of identifying men with AD and other cog-nitive disorders at an early point, ULSAM participants have so far been invited for cognitive testing at 72 (n = 999), 77 (n = 804) and 82 (n = 523) years of age. Subjects with low test performances were
encouraged to visit the Geriatric Memory Clinic at the Uppsala University Hospital for a thorough clinical assessment. To iden-tify dementia cases, all available medical records from the Upp-sala University Hospital, the general practitioners in Uppsala,the community nursing homes and dementia group members were reviewed up to December 31, 2005. Most reports on medical care of the ULSAM participants are provided within these set-tings. The records of all possible cases of dementia or cognitive impairment were reviewed and diagnoses were set by 2 experi-enced geriatricians, independently of each other. In case of dis-agreement, a third geriatrician reviewed the case and the diagno-sis was determined by consensus decision. The diagnosis of AD was defined according to the NINCDS-ADRDA criteria [17] and the DSM-IV criteria [18] . In the majority of AD cases, the original diagnostic workup was made after referral to the Geriatric Mem-ory Clinic based on cognitive testing in ULSAM. CT scans were available from 76 AD cases and were consistent with an AD diag-nosis, i.e. were normal or showed atrophy or mild to moderate white matter changes. CT findings of ̂ 2 brain infarctions that had been clinically silent, i.e. onset and progression were slow and gradual, were considered compatible with the diagnosis of AD, and were present in 3 AD cases. In total, 150 dementia cases were identified in ULSAM and 111 of them were diagnosed with AD. DNA samples were available from 86 AD cases. Individuals who at the 82-year follow-up had a Mini Mental State Examination score 6 26 and no dementia diagnoses were considered cogni-tively healthy. In total, 404 individuals met these criteria and had available DNA samples; these were used as a healthy control group. This study was approved by the ethics committee at Upp-sala University and informed consent for all parts of the study was obtained.
DNA Extraction Blood samples for DNA preparation were obtained from 944
men at examination at 70 years of age. In addition, DNA was also extracted from blood samples from examination around 77 years of age and from archived muscle biopsies, yielding a total of 1,088 DNA samples.
Gene Selection and Genotyping In total, 370 candidate genes with demonstrated or implied
involvement in dementia, diabetes, stroke, cancer and other com-mon complex diseases were investigated in this study. Specifical-ly for dementia, 80 genes involved in 3 major pathways known to affect brain function were selected: A � /lipid metabolism, oxida-tive stress and inflammation/apoptosis [2] . In addition, genes po-sitioned within previously identified AD linkage peaks were also selected. For most of the genes, SNP were distributed across the gene with an average distance of 5 kb. For some of the genes, spe-cific SNP associated with disease in previous studies were select-ed. A majority of the SNP were analyzed with the Illumina Gold-enGate � Assay [19] , but other minisequencing-based methods were also used [20, 21] .
Statistical Analysis Allelic frequency and Hardy-Weinberg equilibrium of SNP
were evaluated using the HaploView software [22] . Case-control association analysis was performed using unconditional logistic regression implemented in the Unphased software [23] . The sig-nificance of the results was evaluated by permutation analysis, by

AD Association Analysis in ULSAM Dement Geriatr Cogn Disord 2009;27:59–68 61
running 1,000 permutations per gene. Incidence and survival analysis was performed using Kaplan-Meier curves and extension of Gehan’s generalized Wilcoxon test in the Statistica software.
In silico replication analysis was performed by extracting data from 2 publicly available GWA studies [24, 25] . Using HapMap (www.hapmap.org) data, out of 146 SNP showing association to
AD in ULSAM, we could identify 1,911 SNP within linkage dis-equilibrium (LD; D � = 0.8) in both GWA studies. Reported asso-ciation from the identified SNP within the respective GWA study was examined as unadjusted data and used to confirm or disprove replication. As the association measure from the Li et al. [25] study, we used Fisher exact p values as published by the authors. From Coon et al. [24] , we calculated disease-association p values by the Plink software using original genotyping data available from the authors’ website.
Results
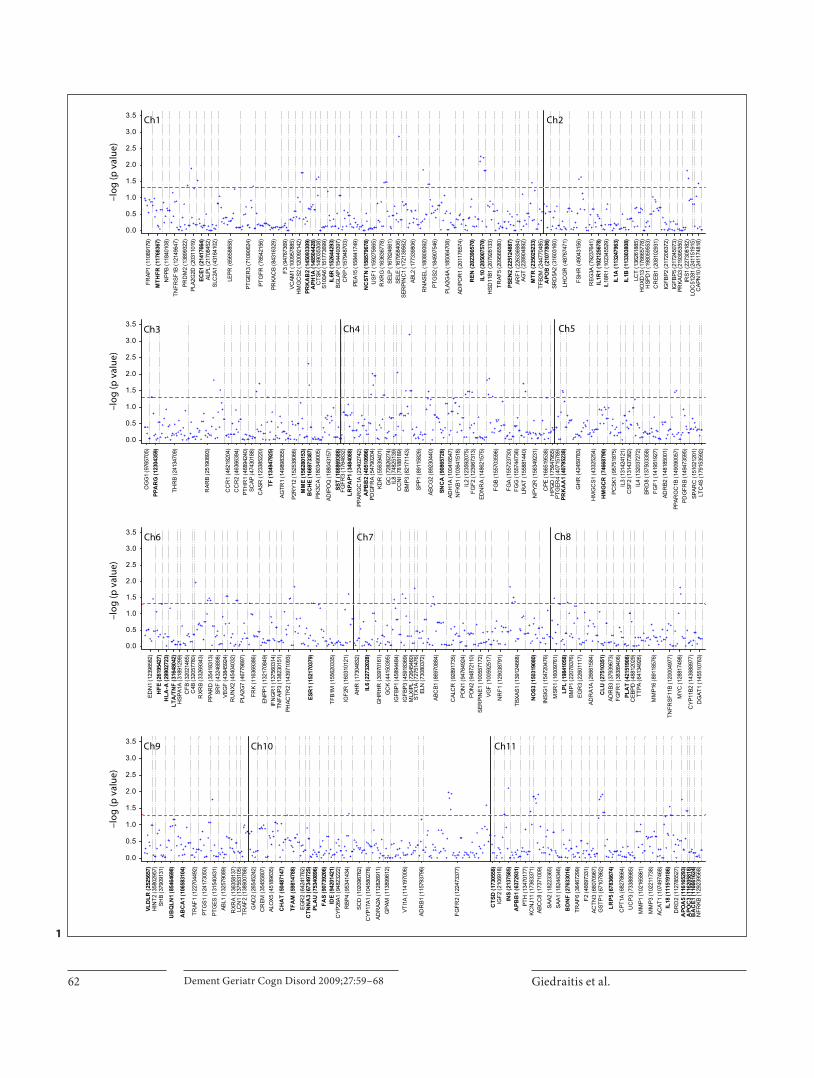
All AD patients and cognitively healthy control (HC) individuals with available DNA samples from ULSAM were included in the present investigation. This resulted in a total of 86 AD and 404 HC samples ( table 1 ). The ini-tial gene selection was based on genes implicated in bio-logical processes with relevance to dementia or genes that have for other reasons been previously investigated in AD association studies. In addition, genes associated with other traits of interest within ULSAM were also included. Altogether, the selected AD and HC groups were geno-typed for 3,118 SNP within 370 genes. The identities and chromosomal localizations of the selected genes are shown in figure 1 . For the majority of genes, SNP were chosen across the entire gene region with an average dis-tance of 5 kb, but, in a few cases, only SNPs of special in-terest based on literature reports were selected (informa-tion on selected SNP is available on request). Prior to the statistical analysis of the data, SNP with a minor allele frequency of ! 1% or with deviations from Hardy-Wein-
berg equilibrium were excluded, which resulted in analy-sis of a total of 2,578 SNP within 368 genes. Association analysis was performed using the Unphased software, which is based on standard unconditional logistic regres-sion. This program also includes a permutation test to establish a gene significance threshold level, taking into account multiple testing and maintaining linkage dis-equilibrium (LD) [23] .
The results of the association analysis revealed a total of 146 SNP within 82 genes with p values ̂ 0.05 ( fig. 1 ). After correcting for multiple testing (1,000 permuta-tions), 29 SNP within 20 genes remained significant atp ̂ 0.05 ( table 2 ). As expected, the APOE (MIM 107741) gene showed the strongest association to AD, with a p value of 10 –5 after permutation analysis (100,000 permu-tations; � 4 allele frequency was 33.1% in the AD group compared to 14.8% in the HC group). Other genes with significant associations to AD both prior and subsequent to permutation analysis included genes with previously described dementia correlations, as well as genes previ-ously associated with other traits. The genes showing the most significant p values both before and after permuta-tion were bone morphogenic protein 4 (BMP4; MIM 112262), bone morphogenic protein 3 (BMP3; MIM 112262) and selectin E (SELE; MIM 131210). In ULSAM, BMP3 and BMP4 were primarily thought of as candidate genes for osteoporosis, and SELE was initially selected as a gene involved in cardiovascular disease and inflamma-tion. The group of 10 genes with the most significant p values both prior and subsequent to permutation includ-ed butyrylcholinesterase (BCHE; MIM 177400) and pre-senilin 1 (PSEN1; MIM 104311), both previously reported to show association to AD [26–28] .
Three genes, BMP4, ESR2 and PSEN1, located in close proximity of each other on chromosome 14q all showed significant association with AD. Therefore, we performed
Table 1. Description of cases (AD) and controls (HC) from ULSAM included in the association study
Group AD HC
Number of individuals 86 404
Average age and range at onset(AD) or examination (HC)
80.2 (72–85) 81.8 (80–84)
Selection criteria AD according to NINCDS-ADRDAand DSM-IV; CT scan
attended examination at 82 years; no dementia diagnosis; MMSE ≥26
MMSE = Mini Mental State Examination.
Giedraitis et al.
Dement Geriatr Cogn Disord 2009;27:59–68 62
FRAP
1 (1
1089
179)
MTH
FR (1
1768
367)
NPP
B (1
1840
108)
TNFR
SF1B
(121
4964
7)PR
DM
2 (1
3899
322)
PLA2
G2D
(203
1101
9)EC
E1 (2
1417
664)
ALPL
(217
0845
2)SL
C2A
1 (4
3164
102)
LEPR
(656
5885
8)
PTG
ER3
(710
9062
4)
PTG
FR (7
8542
156)
PRKA
CB
(843
1632
9)
F3 (9
4767
369)
VCAM
1 (1
0095
7885
)H
MG
CS2
(120
0921
42)
PRK
AB
2 (1
4509
3309
)A
PH1A
(148
5044
28)
CTS
K (1
4903
5308
)S1
00A6
(151
7736
99)
IL6R
(152
6442
93)
BGLA
P (1
5444
9397
)C
RP
(157
9487
03)
PEA1
5 (1
5844
1749
)
NC
STN
(158
5796
78)
USF
1 (1
5927
5665
)R
XRG
(163
6367
78)
SELP
(167
8246
61)
SELE
(167
9584
06)
SER
PIN
C1
(172
1395
62)
ABL2
(177
3398
06)
RN
ASEL
(180
8093
92)
PTG
S2 (1
8490
7546
)
PLA2
G4A
(185
0647
08)
ADIP
OR
1 (2
0117
6574
)
REN
(202
3905
70)
IL10
(205
0075
70)
HSD
11B1
(207
9261
33)
TRAF
5 (2
0956
6580
)
PSEN
2 (2
2512
4887
)AR
F1 (2
2633
6984
)AG
T (2
2890
4892
)
MTR
(235
0252
33)
TFB2
M (2
4477
0485
)A
POB
(210
7780
6)SR
D5A
2 (3
1603
160)
LHC
GR
(487
6747
1)
FSH
R (4
9043
156)
REG
3A (7
9237
641)
IL1R
1 (1
0212
5678
)IL
18R
1 (1
0234
5529
)
IL1A
(113
2479
63)
IL1B
(113
3038
08)
LCT
(136
2618
85)
HO
XD13
(176
6657
78)
HSP
D1
(198
0595
53)
CR
EB1
(208
1029
31)
IGFB
P2 (2
1720
6372
)IG
FBP5
(217
2450
73)
PRKA
G3
(219
3953
50)
IRS1
(227
3081
82)
LOC
5128
1 (2
4115
1915
)C
APN
10 (2
4117
4818
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5–l
og (p
val
ue)
–log
(p v
alue
)–l
og (p
val
ue)
–log
(p v
alue
)Ch1 Ch2
OG
G1
(976
5705
)PP
AR
G (1
2304
359)
THR
B (2
4134
709)
RAR
B (2
5190
893)
CC
R1
(462
1820
4)C
CR
2 (4
6366
394)
PTH
R1
(468
9424
0)SC
AP (4
7430
188)
CAS
R (1
2338
5220
)
TF (1
3494
7925
)
AGTR
1 (1
4989
8355
)
P2R
Y12
(152
5380
66)
MM
E (1
5628
0153
)B
CH
E (1
6697
3387
)PI
K3C
A (1
8034
9005
)
ADIP
OQ
(188
0431
57)
SST
(188
8693
88)
FGFR
3 (1
7648
32)
LRPA
P1 (3
4840
88)
PPAR
GC
1A (2
3402
742)
APB
B2
(405
1099
6)PD
GFR
A (5
4790
204)
KDR
(556
3940
1)G
C (7
2826
274)
IL8
(748
2513
9)C
CN
I (78
1881
89)
BMP3
(821
7114
3)
SPP1
(891
1582
6)
ABC
G2
(892
3044
0)
SNC
A (9
0865
728)
ADH
1A (1
0041
6547
)N
FKB1
(103
6415
18)
IL2
(123
5920
75)
FGF2
(123
9673
13)
EDN
RA
(148
6215
75)
FGB
(155
7035
96)
FGA
(155
7237
30)
FGG
(155
7447
36)
LRAT
(155
8814
40)
NPY
2R (1
5634
9231
)
CPE
(166
5195
38)
HPG
D (1
7564
7955
)PT
GER
4 (4
0715
789)
PRK
AA
1 (4
0795
238)
GH
R (4
2459
783)
HM
GC
S1 (4
3325
254)
HM
GC
R (7
4668
790)
PCSK
1 (9
5751
875)
IL3
(131
4241
21)
CSF
2 (1
3143
7382
)IL
4 (1
3203
7272
)BR
D8
(137
5033
58)
FGF1
(141
9519
27)
ADR
B2 (1
4818
5001
)
PPAR
GC
1B (1
4909
0057
)PD
GFR
B (1
4947
3595
)
SPAR
C (1
5102
1201
)LT
C4S
(179
1535
92)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5 Ch5Ch3 Ch4
EDN
1 (1
2398
582)
HFE
(261
9542
7)H
LA-A
(299
0272
3)LT
A/T
NF
(316
4804
2)H
SPA1
A ( 3
1891
299)
CFB
(320
2146
5)C
4B (3
2057
780)
RXR
B (3
3269
343)
PPAR
D (3
5418
313)
SRF
(432
4689
8)VE
GF
(438
4592
4)R
UN
X2 (4
5404
032)
PLA2
G7
(467
7989
7)
FRK
(116
3693
86)
ENPP
1 (1
3217
0849
)IF
NG
R1
(137
5603
14)
TNFA
IP3
(138
2301
51)
PHAC
TR2
(143
9710
93)
ESR
1 (1
5217
0379
)
TFB1
M (1
5562
0335
)
IGF2
R (1
6031
0121
)
AHR
(173
0483
2)
IL6
(227
3202
8)
GH
RH
R (3
0970
161)
GC
K (4
4150
395)
IGFB
P1 (4
5894
484)
IGFB
P3 (4
5918
369)
MLX
IPL
(726
4546
0)ST
X1A
(727
5147
6)EL
N (7
3080
372)
ABC
B1 (8
6970
884)
CAL
CR
(928
9173
5)
PON
1 (9
4764
924)
PON
2 (9
4872
110)
SER
PIN
E1 (1
0055
7172
)VG
F (1
0059
2517
)
NR
F1 (1
2903
8791
)
TBXA
S1 ( 1
3912
4668
)
NO
S3 (1
5031
9080
)
INSI
G1
(154
7204
76)
MSR
1 (1
6009
761)
LPL
(198
4105
8)BM
P1 (2
2078
376)
EGR
3 (2
2601
117)
ADR
A1A
(266
6158
4)
CLU
(275
1035
1)AD
RB3
(379
3967
3)FG
FR1
(383
8940
6)PL
AT
(421
5190
8)C
EBPD
(488
1202
9)TT
PA (6
4134
926)
MM
P16
(891
1857
6)
TNFR
SF11
B (1
2000
4977
)
MYC
(128
8174
98)
CYP
11B2
(143
9889
77)
DG
AT1
(145
5107
63)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5 Ch8Ch6 Ch7
VLD
LR (2
5256
57)
HIN
T2 (3
5802
957)
SHB
(379
0913
1)
UB
QLN
1 (8
5464
698)
AB
CA
1 (1
0658
3104
)
TRAF
1 (1
2270
4492
)
PTG
S1 (1
2417
3050
)
PTG
ES (1
3154
0431
)AB
L1 (1
3257
9089
)
RXR
A (1
3635
8137
)LC
N1
(137
5531
05)
TRAF
2 (1
3890
0786
)G
AD2
(265
4524
2)C
REM
(354
5580
7)AL
OX5
(451
8963
5)
CH
AT
(504
8714
7)
TFA
M (5
9814
788)
EGR
2 (6
4241
762)
CTN
NA
3 (6
7349
725)
PLA
U (7
5340
896)
FAS
(907
3920
6)ID
E (9
4201
421)
CYP
26A1
(948
2322
2)R
BP4
(953
4143
4)
SCD
(102
0967
62)
CYP
17A1
(104
5802
78)
ADR
A2A
(112
8269
11)
GPA
M (1
1389
9612
)
VTI1
A (1
1419
7006
)
ADR
B1 (1
1579
3796
)
FGFR
2 (1
2247
3377
)
CTS
D (1
7305
58)
IGF2
(210
6918
)IN
S (2
1375
85)
APB
B1
(637
2931
)PT
H (1
3470
177)
KCN
J11
(173
6337
1)AB
CC
8 (1
7371
009)
SAA2
(182
2336
5)SA
A1 (1
8244
346)
BD
NF
(276
3301
6)TR
AF6
(364
6729
9)F2
(466
9733
1)AC
TN3
(660
7096
7)G
STP1
(671
0786
2)
LRP5
(678
3667
4)
CPT
1A (6
8278
664)
UC
P3 (7
3388
985)
MM
P1 (1
0216
5861
)
MM
P3 (1
0221
1738
)AC
AT1
(107
4974
68)
IL18
(111
5191
86)
DR
D2
(112
7855
27)
APO
A5
(116
1652
93)
APO
C3
(116
2058
18)
BA
CE1
(116
6616
24)
NFR
KB (1
2923
9568
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5Ch11Ch10Ch9
1

AD Association Analysis in ULSAM Dement Geriatr Cogn Disord 2009;27:59–68 63
haplotype analysis of this region and could demonstrate that the haplotype with the strongest AD association(p = 1.0 ! 10 –5 ) within this region included the most sig-nificantly associated SNP within BMP4 and ESR2 and 1 SNP from PSEN1, which was also significantly associated with AD.
As a supplement to the case-control comparison, we used all 1,088 available samples from ULSAM and per-formed incidence by genotype analysis, using AD diag-nosis as outcome. This analysis included 86 cases (AD) and 404 controls (HC) from the original study and an ad-ditional 598 samples. These 598 samples did not include any patients with AD diagnosis, but did contain samples from ULSAM participants who had been excluded from the control group due to circumstances such as other
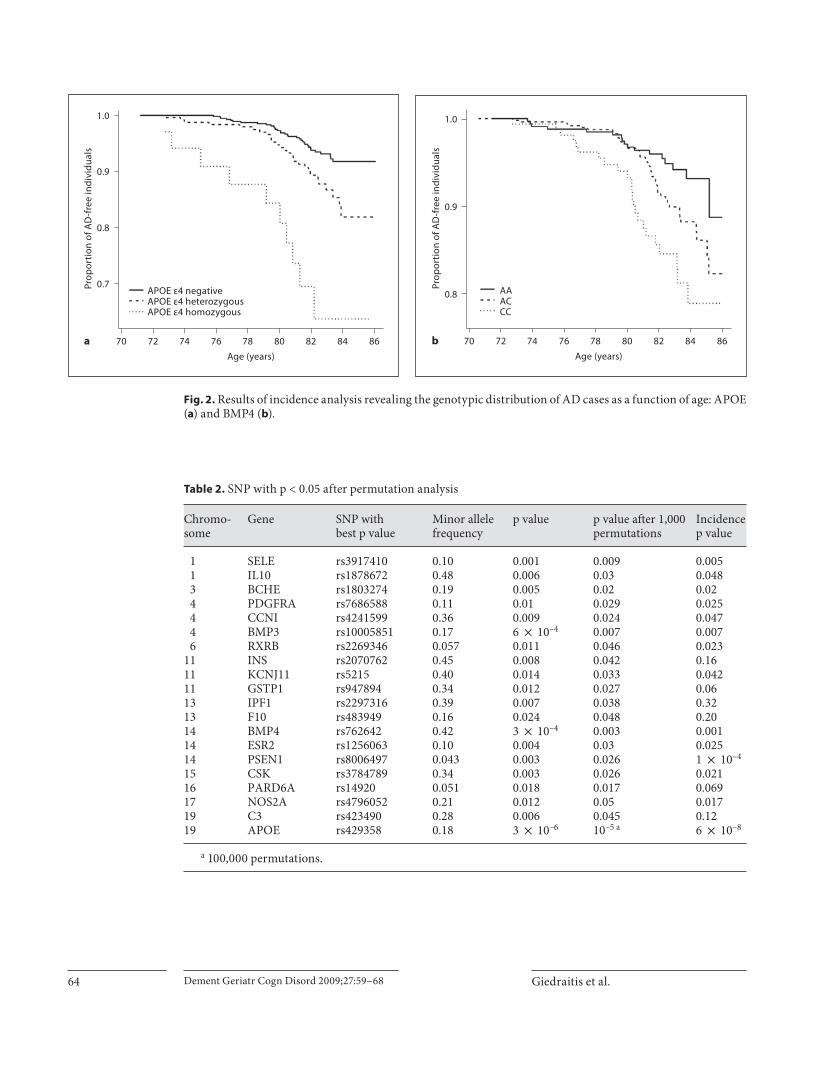
forms of dementia or not attending examination around 82 years of age. By this type of incidence analysis, the lon-gitudinal data from ULSAM can be explored in more de-tail than in a case-control study. We performed incidence analysis of all SNP with p ! 0.05 from the initial case-control comparison, and these results were consistent with the results from the case-control comparison ( ta-ble 2 ). Details from 2 SNP with the most significant as-sociation to AD are shown ( fig. 2 ) and the results revealed p values of 5.8 ! 10 –8 and 0.001 for APOE and BMP4, respectively. These results confirmed the initial effect in the case-control comparison and demonstrated the age effect of APOE, as � 4 homozygotes showed an increased AD frequency at an earlier age than other genotypic vari-ants. This effect is not evident for rs762642 from BMP4,
Co
lor v
ersi
on
avai
lab
le o
nlin
e
–log
(p v
alue
)–l
og (p
val
ue)
ADIP
OR
2 (1
6704
08)
FGF2
3 (4
3476
54)
VWF
(592
8301
)
CD
9 (6
1791
34)
TNFR
SF1A
(630
8184
)G
APD
H (6
5138
72)
A2M
(910
2453
)
VDR
(465
2158
7)
TEG
T (4
8421
607)
TFC
P2 (4
9774
887)
IGFB
P6 (5
1777
703)
RAR
G (5
1890
621)
MM
P19
(545
1548
1)LR
P1 (5
5808
543)
NR
1H4
(993
9181
0)
IGF1
(101
3138
06)
ACAC
B (1
0803
8783
)
PRK
AB
1 (1
1859
0144
)TC
F1 (1
1990
0932
)
IPF1
(273
9215
7)AL
OX5
AP (3
0207
645)
POST
N (3
7034
722)
TNFS
F11
(420
3487
2)TP
T1 (4
4673
747)
KLF5
(725
3093
1)ED
NR
B (7
7367
617)
IRS2
(109
2041
85)
F7 (1
1280
8106
)F1
0 (1
1282
5114
)LT
B4R
(238
3890
8)PT
GER
2 (5
1850
863)
BMP4
(534
8620
7)
ESR
2 (6
3621
388)
VTI1
B (6
7186
985)
PSEN
1 (7
2672
915)
DLS
T (7
4418
372)
SER
PIN
A3
(941
4846
7)
CYP
46A
1 (9
9220
407)
TRAF
3 (1
0231
3569
)TN
FAIP
2 (1
0266
2417
)AK
T1 (1
0430
6731
)C
YP19
A1
(492
8896
1)
LIPC
(565
1146
7)
APH
1B (6
1356
844)
CSK
(728
6148
9)
IGF1
R (9
7010
288)
CR
EBBP
(371
6568
)C
IITA
(108
7855
8)SO
CS1
(112
5577
5)VK
OR
C1
(310
0967
6)M
MP2
(540
7058
9)
CET
P (5
5553
263)
CX3
CL1
(559
6390
0)
CD
H11
(635
3818
4)
PAR
D6A
(662
5235
2)LC
AT (6
6531
154)
HP
(706
4600
9)M
LYC
D (8
2490
231)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
ALO
X15
(448
0963
)AL
OX1
2 (6
8401
08)
SLC
2A4
(712
5777
)VA
MP2
(800
3189
)EL
AC2
(128
3643
3)
SREB
F1 (1
7655
794)
NO
S2A
(231
0791
9)TN
FAIP
1 (2
3686
913)
TRAF
4 (2
4095
166)
SLC
6A4
(255
4903
2)B
LMH
(255
9934
9)C
CL2
(296
0640
9)PI
P5K2
B (3
4177
324)
THR
A (3
5472
589)
RAR
A (3
5718
972)
IGFB
P4 (3
5853
202)
CN
P (3
7372
285)
MLX
(379
7260
4)IT
GA2
B (3
9805
076)
MA
PT (4
1327
624)
CO
L1A1
(456
1645
6)M
PO (5
3702
201)
AC
E (5
8908
166)
GH
1 (5
9348
294)
GR
B2
(708
2575
2)TI
MP2
(743
6065
4)
NPC
1 (1
9365
461)
TNFR
SF11
A (5
8143
500)
PTBP
1 (7
4841
1)ST
K11
(115
6798
)TB
XA2R
(354
5504
)C
3 (6
6288
78)
ICAM
1 (1
0242
765)
LDLR
(110
6113
2)
CAC
NA1
A (1
3179
088)
PTG
ER1
(144
4427
8)G
DF1
5 (1
8357
968)
CO
MP
(187
5458
3)C
EBPA
(384
8277
6)U
SF2
(404
5172
1)PS
ENEN
(409
2833
4)TG
FB1
(465
2825
4)A
POE
(501
0087
9)PT
GIR
(518
1556
5)N
R1H
2 (5
5571
515)
TNN
I3 (6
0354
950)
IL11
(605
6756
9)TR
IB3
(309
273)
PRN
P (4
6149
96)
SNAP
25 (1
0147
477)
PCSK
2 (1
7154
752)
THBD
(229
7427
0)C
ST3
(235
6229
4)G
HR
H (3
5312
899)
HN
F4A
(424
1785
5)
MM
P9 (4
4070
954)
PTG
IS (4
7553
818)
PTPN
1 (4
8560
294)
BMP7
(551
7721
1)TN
FRSF
6B (6
1759
607)
BA
CE2
(414
6159
8)
CB
S (4
3346
370)
HM
OX1
(341
0705
7)C
YP2D
6 (4
0852
445)
PPAR
A (4
4925
163)
CPT
1B (4
9354
156)
PHEX
(219
6048
0)
TIM
P1 (4
7326
634)
AR (6
6680
599)
TAF1
(705
0283
9)IR
S4 (1
0786
2368
)AG
TR2
(115
2160
03)
CD
40LG
(135
5580
02)
FGF1
3 (1
3754
1401
)SO
X3 (1
3941
2818
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5 5.5 Ch21Ch22 ChX Ch20
Ch12 Ch13 Ch14 Ch15 Ch16
Ch17 Ch18 Ch19
Fig. 1. The complete results from the case-control study showing the distribution of SNP within genes positioned along the chro-mosomes (Ch) and the corresponding p values of AD association. Genes are evenly placed along the chromosomes with no exact chromosomal localization. The gene start positions based on the GeneCards database are shown in parentheses. If the gene posi-
tion is not available, the position of the first SNP in the gene (based on the Single Nucleotide Polymorphism Database) is shown. Horizontal dashed lines show the p value level of 0.05. Vertical dotted lines are separators between genes. Genes primar-ily selected as candidate genes to dementia are shown in bold.
Giedraitis et al.
Dement Geriatr Cogn Disord 2009;27:59–68 64
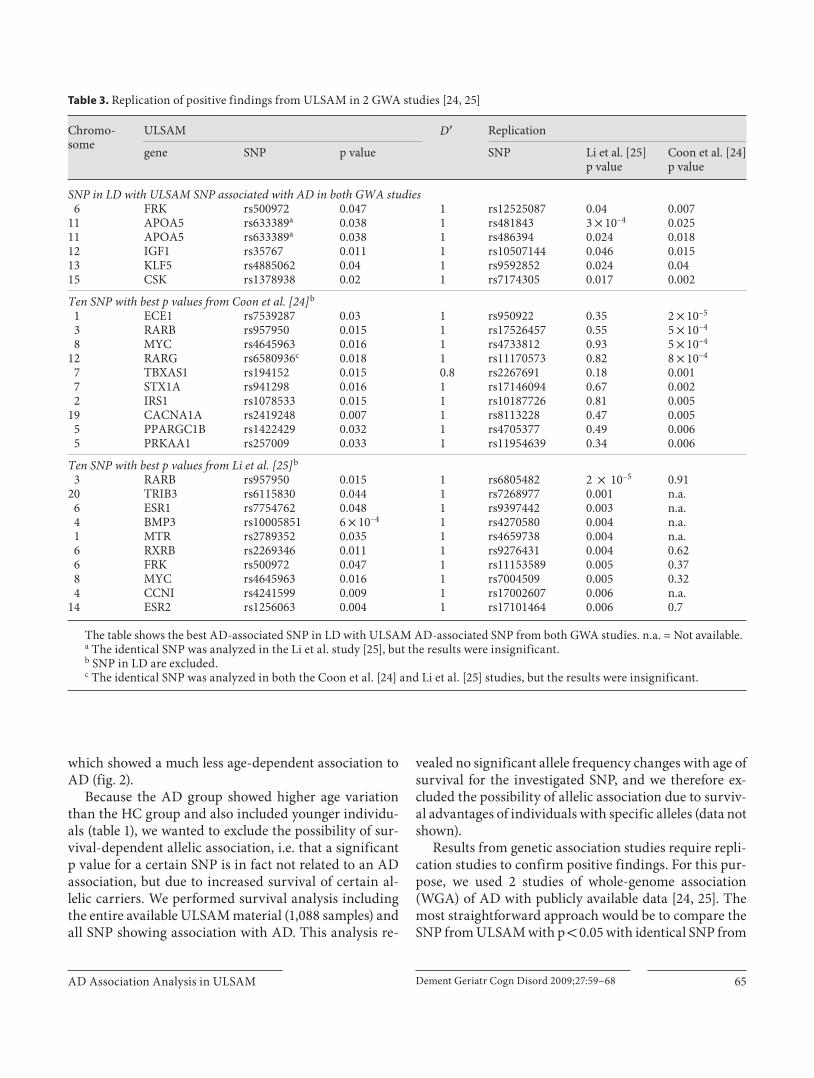
Table 2. SNP with p < 0.05 after permutation analysis
Chromo-some
Gene SNP withbest p value
Minor allelefrequency
p value p value after 1,000permutations
Incidencep value
1 SELE rs3917410 0.10 0.001 0.009 0.0051 IL10 rs1878672 0.48 0.006 0.03 0.0483 BCHE rs1803274 0.19 0.005 0.02 0.024 PDGFRA rs7686588 0.11 0.01 0.029 0.0254 CCNI rs4241599 0.36 0.009 0.024 0.0474 BMP3 rs10005851 0.17 6 ! 10–4 0.007 0.0076 RXRB rs2269346 0.057 0.011 0.046 0.023
11 INS rs2070762 0.45 0.008 0.042 0.1611 KCNJ11 rs5215 0.40 0.014 0.033 0.04211 GSTP1 rs947894 0.34 0.012 0.027 0.0613 IPF1 rs2297316 0.39 0.007 0.038 0.3213 F10 rs483949 0.16 0.024 0.048 0.2014 BMP4 rs762642 0.42 3 ! 10–4 0.003 0.00114 ESR2 rs1256063 0.10 0.004 0.03 0.02514 PSEN1 rs8006497 0.043 0.003 0.026 1 ! 10–4
15 CSK rs3784789 0.34 0.003 0.026 0.02116 PARD6A rs14920 0.051 0.018 0.017 0.06917 NOS2A rs4796052 0.21 0.012 0.05 0.01719 C3 rs423490 0.28 0.006 0.045 0.1219 APOE rs429358 0.18 3 ! 10–6 10–5 a 6 ! 10–8
a 100,000 permutations.
70
0.7
0.8
Prop
orti
on o
f AD
-fre
e in
div
idua
ls
0.9
1.0
a 72 74 76
APOE ε4 negativeAPOE ε4 heterozygousAPOE ε4 homozygous
78
Age (years)
80 82 84 86 70
0.8
0.9
Prop
orti
on o
f AD
-fre
e in
div
idua
ls
1.0
b 72 74 76
AAACCC
78
Age (years)
80 82 84 86
Fig. 2. Results of incidence analysis revealing the genotypic distribution of AD cases as a function of age: APOE ( a ) and BMP4 ( b ).
AD Association Analysis in ULSAM Dement Geriatr Cogn Disord 2009;27:59–68 65
which showed a much less age-dependent association to AD ( fig. 2 ).
Because the AD group showed higher age variation than the HC group and also included younger individu-als ( table 1 ), we wanted to exclude the possibility of sur-vival-dependent allelic association, i.e. that a significant p value for a certain SNP is in fact not related to an AD association, but due to increased survival of certain al-lelic carriers. We performed survival analysis including the entire available ULSAM material (1,088 samples) and all SNP showing association with AD. This analysis re-
vealed no significant allele frequency changes with age of survival for the investigated SNP, and we therefore ex-cluded the possibility of allelic association due to surviv-al advantages of individuals with specific alleles (data not shown).
Results from genetic association studies require repli-cation studies to confirm positive findings. For this pur-pose, we used 2 studies of whole-genome association (WGA) of AD with publicly available data [24, 25] . The most straightforward approach would be to compare the SNP from ULSAM with p ! 0.05 with identical SNP from
Table 3. Replication of positive findings from ULSAM in 2 GWA studies [24, 25]
Chromo-some
ULSAM D� Replication
gene SNP p value SNP Li et al. [25]p value
Coon et al. [24]p value
SNP in LD with ULSAM SNP associated with AD in both GWA studies6 FRK rs500972 0.047 1 rs12525087 0.04 0.007
11 APOA5 rs633389a 0.038 1 rs481843 3!10–4 0.02511 APOA5 rs633389a 0.038 1 rs486394 0.024 0.01812 IGF1 rs35767 0.011 1 rs10507144 0.046 0.01513 KLF5 rs4885062 0.04 1 rs9592852 0.024 0.0415 CSK rs1378938 0.02 1 rs7174305 0.017 0.002
Ten SNP with best p values from Coon et al. [24] b
1 ECE1 rs7539287 0.03 1 rs950922 0.35 2!10–5
3 RARB rs957950 0.015 1 rs17526457 0.55 5!10–4
8 MYC rs4645963 0.016 1 rs4733812 0.93 5!10–4
12 RARG rs6580936c 0.018 1 rs11170573 0.82 8!10–4
7 TBXAS1 rs194152 0.015 0.8 rs2267691 0.18 0.0017 STX1A rs941298 0.016 1 rs17146094 0.67 0.0022 IRS1 rs1078533 0.015 1 rs10187726 0.81 0.005
19 CACNA1A rs2419248 0.007 1 rs8113228 0.47 0.0055 PPARGC1B rs1422429 0.032 1 rs4705377 0.49 0.0065 PRKAA1 rs257009 0.033 1 rs11954639 0.34 0.006
Ten SNP with best p values from Li et al. [25] b
3 RARB rs957950 0.015 1 rs6805482 2 ! 10–5 0.9120 TRIB3 rs6115830 0.044 1 rs7268977 0.001 n.a.
6 ESR1 rs7754762 0.048 1 rs9397442 0.003 n.a.4 BMP3 rs10005851 6!10–4 1 rs4270580 0.004 n.a.1 MTR rs2789352 0.035 1 rs4659738 0.004 n.a.6 RXRB rs2269346 0.011 1 rs9276431 0.004 0.626 FRK rs500972 0.047 1 rs11153589 0.005 0.378 MYC rs4645963 0.016 1 rs7004509 0.005 0.324 CCNI rs4241599 0.009 1 rs17002607 0.006 n.a.
14 ESR2 rs1256063 0.004 1 rs17101464 0.006 0.7
The table shows the best AD-associated SNP in LD with ULSAM AD-associated SNP from both GWA studies. n.a. = Not available.a The identical SNP was analyzed in the Li et al. study [25], but the results were insignificant. b SNP in LD are excluded. c The identical SNP was analyzed in both the Coon et al. [24] and Li et al. [25] studies, but the results were insignificant.

Giedraitis et al.
Dement Geriatr Cogn Disord 2009;27:59–68 66
the 2 GWA studies. However, as ULSAM was analyzed with SNP from Illumina and the Coon et al. [24] and Li et al. [25] studies used Affymetrix technology, most SNP are not identical. From 146 SNP within 82 genes associ-ated with AD in ULSAM, only 13 were identical to SNP analyzed in both GWA studies, and an additional 9 were studied in Li et al. [25] . As the number of identical SNP between studies was so limited, we decided not to use identity as a selection criteria for replication, but rather compared SNP from the 2 GWA studies in LD with SNP from ULSAM found to be associated with AD. In both GWA studies, 1 SNP (rs4420638) close to the APOE locus showed a strong association with AD. This SNP is in LD with the 2 SNP (rs429358 and rs7412) identifying the APOE isoforms with D � = 0.86 and 0.90, respectively [24] . Therefore, we chose a threshold level for defining LD at D � = 0.8 for this replication study. Using HapMap data, we found 1,911 SNP from both GWA studies in LD with 114 SNP to be associated in ULSAM. Out of these, 6 SNP within 5 genes were associated with AD in both GWA studies ( table 3 ). An additional 113 SNP from Li et al. [25] and 102 SNP from Coon et al. [24] were found to be as-sociated in the respective study (p ! 0.05). This left 32 SNP from ULSAM that were not available from HapMap or did not show LD to any other SNP, and which could therefore not be assessed.
Discussion
For this association study of AD, we used all available genotypes from cases and controls generated within UL-SAM. In addition to genes with previously reported de-mentia correlations, genes selected for analysis of other complex conditions within ULSAM have also been in-cluded in the investigation. Although not strictly ‘hy-pothesis driven’, this approach could potentially indi-cate genes not previously shown to be associated with de-mentia.
This study did, in addition to the expected association of APOE, also point to several other genes with a poten-tial effect on the risk of developing AD. Amongst these, 2 genes (BMP4 and BMP3) attracted our attention. Al-though not previously reported to be involved in demen-tia development, this group of proteins regulates various biological processes, including cell growth and cell dif-ferentiation, and they play an important role in bone in-duction and fracture repair [29, 30] . Furthermore, BMP signaling and Wnt pathways both contribute to chondro-genesis and osteogenesis, and Wnt pathways have been
proposed to be involved in several neurodegenerative dis-orders, including AD [31] . This might suggest a common molecular mechanism by which the BMPs could also play a role in AD development. Another gene from this study with association to AD is SELE, which belongs to a group of adhesion molecules expressed by endothelial cells and involved in inflammation processes that could be rele-vant to AD pathogenesis. We could also detect associa-tion of 2 genes that in the past have repeatedly been im-plicated in AD association studies, BCHE and PSEN1 [26–28] . The products of these genes are involved in the metabolic pathways of acetylcholine [32] and amyloid precursor protein (APP) [33, 34] , respectively. A reduc-tion in cholinergic neurons is characteristic of AD, and the reduction in cholinergic activity that follows has pre-viously been associated with increased levels of BCHE [35, 36] . PSEN1 plays an important role in the cleavage of A � peptide from its precursor form APP [33, 34] , and A � is a major component in the plaques that form in the brains of AD patients.
Noticeably, on chromosome 14q, haplotype analysis including the genes BMP4, ESR1 and PSEN1, revealed a 3-marker haplotype clearly associated with AD. This haplotype includes 1 SNP from each gene, which are all intronic and not likely to be functional. However, they might be in LD with some functional variation within this region which has still to be identified. Interestingly, 1 SNP (rs165932) within PSEN1, included in this haplo-type, has been analyzed in other association studies with varying results [26, 27] .
Due to the limited number of cases in our study, sepa-rate association analyses for APOE � 4 carriers and non-carriers were not possible.
A problematic aspect of investigating associations be-tween such a large number of genes, as has been done here, is the substantial number of comparisons carried out. We dealt with this problem by permutation tests car-ried out within the Unphased software. This permutation takes into account LD between SNP and multiple com-parisons within individual genes. The method of adjust-ment is therefore fairly strict within genes, but does not take into consideration the number of investigated genes. The haplotype analysis of a 3-marker haplotype on chro-mosome 14q has not been adjusted for multiple compari-sons, but was based on individual SNP that showed sig-nificant AD associations.
When this study was initiated, WGA studies were not practically feasible. There have been rapid developments in WGA examinations in the past few years, and there are now reports on a number of genetic studies of complex

AD Association Analysis in ULSAM Dement Geriatr Cogn Disord 2009;27:59–68 67
traits employing this technology [37–39] . The advantages are obvious, and there have indeed been several reports of solid reproducible results generated from such investi-gations. Within the field of AD there are, to our knowl-edge, 2 such reports with publicly available data [24, 25] . We used data from these 2 GWA studies in an attempt to replicate association results from ULSAM. Given the large number of observations made within a GWA study, the possibility of finding any chosen SNP with an asso-ciation p value ! 0.05 that is due to pure chance is high. However, the risk of a random finding when investigating the same SNP within 2 independent GWA investigations is substantially decreased. We therefore only considered replication successful if confirmed in both GWA studies. By extracting information from these databases, we were able to replicate only 5 of the associations of AD to spe-cific genes detected within ULSAM. Within the APOA5 gene, an identical SNP (rs633389) that showed associa-tion in ULSAM was studied in Li et al. [25] with no sig-nificant association to AD. However, SNP in LD with rs633389 were associated with AD in the GWA studies by both Li et al. [25] and Coon et al. [24] ( table 3 ). This, of course, might represent just a chance finding, but could also be due to haplotype structure differences between populations. These SNP might be in LD with putative functional SNP, but the level of LD and haplotype struc-ture might differ between populations. Therefore, differ-ent SNP within the same gene show association with dis-ease. The small number of replicable associations is not surprising, given the limited number of AD cases avail-able from ULSAM and the possibility of random associa-
tion effects. Nonetheless, not being able to replicate the best association results from ULSAM is disappointing. However, the in silico replication analysis that was car-ried out here, where different genotyping platforms have been used, can be troublesome. Perhaps the most signifi-cant problem is that very few identical SNP could be iden-tified, and comparisons were based on LD between mark-ers as indicated by HapMap. Sometimes this method may not accurately reflect the real association value, and, more importantly, on a few occasions no SNP in LD with ULSAM-associated SNP could be identified from the GWA studies. In these instances, information on replica-tion is inconclusive.
In summary, we have identified 82 genes that can be associated with AD within ULSAM. Performing an in silico replication analysis using publicly available data from 2 GWA studies confirmed only 5 of these AD-as-sociated genes. However, we cannot exclude the possibil-ity that further attempts at replication using identical SNP from ULSAM and population-matched sample col-lection could support more of the association data sug-gested in this study.
Acknowledgments
This study was supported by Wallenberg Consortium North, Swedish Brain Fund, Alzheimerfonden, Swedish Research Coun-cil (2003-5546), APOPIS (Contract No. LSHM-CT-2003-503330), Stiftelsen Gamla Tjänarinnor, Capios Forskningsstiftelse, Erik, Karin och Gösta Selanders stiftelse and Swedish Lions Research Foundation.
References
1 Corder EH, Saunders AM, Strittmatter WJ, Schmechel DE, Gaskell PC, Small GW, Roses AD, Haines JL, Pericak-Vance MA: Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset fam-ilies. Science 1993; 261: 921–923.
2 Combarros O, Alvarez-Arcaya A, Sanchez-Guerra M, Infante J, Berciano J: Candidate gene association studies in sporadic Alz-heimer’s disease. Dement Geriatr Cogn Dis-ord 2002; 14: 41–54.
3 Bertram L, McQueen MB, Mullin K, Blacker D, Tanzi RE: Systematic meta-analyses ofAlzheimer disease genetic association stud-ies: the AlzGene database. Nat Genet 2007; 39: 17–23.
4 Schmitz F, Mevissen V, Krantz C, Kimmel M, Erdmann J, Hoffmann R, Zerres K, Ortlepp JR: Robust association of the APOE epsilon4 allele with premature myocardial infarction especially in patients without hy-percholesterolaemia: the Aachen study. Eur J Clin Invest 2007; 37: 106–108.
5 Song Y, Stampfer MJ, Liu S: Meta-analysis: apolipoprotein E genotypes and risk for cor-onary heart disease. Ann Intern Med 2004; 141: 137–147.
6 Knopman DS, Edland SD, Cha RH, Petersen RC, Rocca WA: Incident dementia in women is preceded by weight loss by at least a decade. Neurology 2007; 69: 739–746.
7 Whitmer RA, Gunderson EP, Quesenberry CP Jr, Zhou J, Yaffe K: Body mass index in midlife and risk of Alzheimer disease and vascular dementia. Curr Alzheimer Res 2007; 4: 103–109.
8 Gustafson D, Rothenberg E, Blennow K, Steen B, Skoog I: An 18-year follow-up of overweight and risk of Alzheimer disease. Arch Intern Med 2003; 163: 1524–1528.
9 Ott A, Stolk RP, van Harskamp F, Pols HA, Hofman A, Breteler MM: Diabetes mellitus and the risk of dementia: The Rotterdam Study. Neurology 1999; 53: 1937–1942.
10 Leibson CL, Rocca WA, Hanson VA, Cha R, Kokmen E, O’Brien PC, Palumbo PJ: The risk of dementia among persons with diabe-tes mellitus: a population-based cohort study. Ann NY Acad Sci 1997; 826: 422–427.
11 Qiu WQ, Walsh DM, Ye Z, Vekrellis K, Zhang J, Podlisny MB, Rosner MR, Safavi A, Hersh LB, Selkoe DJ: Insulin-degrading en-zyme regulates extracellular levels of amy-loid beta-protein by degradation. J Biol Chem 1998; 273: 32730–32738.

Giedraitis et al.
Dement Geriatr Cogn Disord 2009;27:59–68 68
12 Farris W, Mansourian S, Chang Y, Lindsley L, Eckman EA, Frosch MP, Eckman CB, Tanzi RE, Selkoe DJ, Guenette S: Insulin-de-grading enzyme regulates the levels of insu-lin, amyloid beta-protein, and the beta-amy-loid precursor protein intracellular domain in vivo. Proc Nat Acad Sci USA 2003; 100: 4162–4167.
13 Klaus A, Birchmeier W: Wnt signalling and its impact on development and cancer. Nat Rev Cancer 2008; 8: 387–398.
14 Johnson ML, Kamel MA: The Wnt signaling pathway and bone metabolism. Curr Opin Rheumatol 2007; 19: 376–382.
15 De Ferrari GV, Moon RT: The ups and downs of Wnt signaling in prevalent neurological disorders. Oncogene 2006; 25: 7545–7553.
16 Hedstrand H: A study of middle-aged men with particular reference to risk factors for cardiovascular disease. Ups J Med Sci Suppl 1975; 19: 1–61.
17 McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan EM: Clinical di-agnosis of Alzheimer’s disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Hu-man Services Task Force on Alzheimer’s Dis-ease. Neurology 1984; 34: 939–944.
18 APA: Diagnostic and Statistical Manual of Mental Disorders: DSM-IV. Washington, American Psychiatric Association, 1994.
19 Fan JB, Oliphant A, Shen R, Kermani BG, Garcia F, Gunderson KL, Hansen M, Steem-ers F, Butler SL, Deloukas P, Galver L, Hunt S, McBride C, Bibikova M, Rubano T, Chen J, Wickham E, Doucet D, Chang W, Camp-bell D, Zhang B, Kruglyak S, Bentley D, Haas J, Rigault P, Zhou L, Stuelpnagel J, Chee MS: Highly parallel SNP genotyping. Cold Spring Harb Symp Quant Biol 2003; 68: 69–78.
20 Bell PA, Chaturvedi S, Gelfand CA, Huang CY, Kochersperger M, Kopla R, Modica F, Pohl M, Varde S, Zhao R, Zhao X, Boyce-Ja-cino MT, Yassen A: SNPstream UHT: ultra-high throughput SNP genotyping for phar-macogenomics and drug discovery. Bio-techniques 2002;(suppl):70–72, 74, 76–77.
21 Chen X, Levine L, Kwok PY: Fluorescence polarization in homogeneous nucleic acid analysis. Genome Res 1999; 9: 492–498.
22 Barrett JC, Fry B, Maller J, Daly MJ: Haplo-view: analysis and visualization of LD and haplotype maps. Bioinformatics 2005; 21: 263–265.
23 Dudbridge F: Pedigree disequilibrium tests for multilocus haplotypes. Genet Epidemiol 2003; 25: 115–121.
24 Coon KD, Myers AJ, Craig DW, Webster JA, Pearson JV, Lince DH, Zismann VL, Beach TG, Leung D, Bryden L, Halperin RF, Mar-lowe L, Kaleem M, Walker DG, Ravid R, Heward CB, Rogers J, Papassotiropoulos A, Reiman EM, Hardy J, Stephan DA: A high-density whole-genome association study re-veals that APOE is the major susceptibility gene for sporadic late-onset Alzheimer’s dis-ease. J Clin Psychiatry 2007; 68: 613–618.
25 Li H, Wetten S, Li L, St Jean PL, Upmanyu R, Surh L, Hosford D, Barnes MR, Briley JD, Borrie M, Coletta N, Delisle R, Dhalla D, Ehm MG, Feldman HH, Fornazzari L, Gauthier S, Goodgame N, Guzman D, Ham-mond S, Hollingworth P, Hsiung GY, John-son J, Kelly DD, Keren R, Kertesz A, King KS, Lovestone S, Loy-English I, Matthews PM, Owen MJ, Plumpton M, Pryse-Phillips W, Prinjha RK, Richardson JC, Saunders A, Slater AJ, St George-Hyslop PH, Stinnett SW, Swartz JE, Taylor RL, Wherrett J, Williams J, Yarnall DP, Gibson RA, Irizarry MC, Mid-dleton LT, Roses AD: Candidate single-nu-cleotide polymorphisms from a genomewide association study of Alzheimer disease. Arch Neurol 2008; 65: 45–53.
26 Tilley L, Morgan K, Grainger J, Marsters P, Morgan L, Lowe J, Xuereb J, Wischik C, Har-rington C, Kalsheker N: Evaluation of poly-morphisms in the presenilin-1 gene and the butyrylcholinesterase gene as risk factors in sporadic Alzheimer’s disease. Eur J Hum Genet 1999; 7: 659–663.
27 van Duijn CM, Cruts M, Theuns J, Van Gas-sen G, Backhovens H, van den Broeck M, Wehnert A, Serneels S, Hofman A, Van Broeckhoven C: Genetic association of the presenilin-1 regulatory region with early-onset Alzheimer’s disease in a population-based sample. Eur J Hum Genet 1999; 7: 801–806.
28 Wiebusch H, Poirier J, Sevigny P, Schappert K: Further evidence for a synergistic associa-tion between APOE epsilon4 and BCHE-K in confirmed Alzheimer’s disease. Human Gen 1999; 104: 158–163.
29 Bandyopadhyay A, Tsuji K, Cox K, Harfe BD, Rosen V, Tabin CJ: Genetic analysis of the roles of BMP2, BMP4, and BMP7 in limb patterning and skeletogenesis. PLoS genetics 2006; 2:e216.
30 Engstrand T: Molecular biologic aspects of cartilage and bone: potential clinical appli-cations. Ups J Med Sci 2003; 108: 25–35.
31 Ille F, Atanasoski S, Falk S, Ittner LM, Marki D, Buchmann-Moller S, Wurdak H, Suter U, Taketo MM, Sommer L: Wnt/BMP signal in-tegration regulates the balance between pro-liferation and differentiation of neuroepi-thelial cells in the dorsal spinal cord. Dev Biol 2007; 304: 394–408.
32 Ballard CG, Greig NH, Guillozet-Bongaarts AL, Enz A, Darvesh S: Cholinesterases: roles in the brain during health and disease. Curr Alzheimer Res 2005; 2: 307–318.
33 Brunkan AL, Goate AM: Presenilin function and gamma-secretase activity. J Neurochem 2005; 93: 769–792.
34 Chyung JH, Raper DM, Selkoe DJ: Gamma-secretase exists on the plasma membrane as an intact complex that accepts substrates and effects intramembrane cleavage. J Biol Chem 2005; 280: 4383–4392.
35 Mesulam MM, Geula C: Butyrylcholinester-ase reactivity differentiates the amyloid plaques of aging from those of dementia. Ann Neurol 1994; 36: 722–727.
36 Moran MA, Mufson EJ, Gomez-Ramos P: Colocalization of cholinesterases with beta amyloid protein in aged and Alzheimer’s brains. Acta Neuropathol 1993; 85: 362–369.
37 Wellcome Trust Case Control Consortium: Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 2007; 447: 661–678.
38 Frayling TM, Timpson NJ, Weedon MN, Zeggini E, Freathy RM, Lindgren CM, Perry JR, Elliott KS, Lango H, Rayner NW, Shields B, Harries LW, Barrett JC, Ellard S, Groves CJ, Knight B, Patch AM, Ness AR, Ebrahim S, Lawlor DA, Ring SM, Ben-Shlomo Y, Jar-velin MR, Sovio U, Bennett AJ, Melzer D, Ferrucci L, Loos RJ, Barroso I, Wareham NJ, Karpe F, Owen KR, Cardon LR, Walker M, Hitman GA, Palmer CN, Doney AS, Morris AD, Smith GD, Hattersley AT, McCarthy MI: A common variant in the FTO gene is associated with body mass index and predis-poses to childhood and adult obesity. Science 2007; 316: 889–894.
39 Gudmundsson J, Sulem P, Manolescu A, Amundadottir LT, Gudbjartsson D, Helga-son A, Rafnar T, Bergthorsson JT, Agnarsson BA, Baker A, Sigurdsson A, Benediktsdottir KR, Jakobsdottir M, Xu J, Blondal T, Kostic J, Sun J, Ghosh S, Stacey SN, Mouy M, Sae-mundsdottir J, Backman VM, Kristjansson K, Tres A, Partin AW, Albers-Akkers MT, Godino-Ivan Marcos J, Walsh PC, Swinkels DW, Navarrete S, Isaacs SD, Aben KK, Graif T, Cashy J, Ruiz-Echarri M, Wiley KE, Suarez BK, Witjes JA, Frigge M, Ober C, Jonsson E, Einarsson GV, Mayordomo JI, Kiemeney LA, Isaacs WB, Catalona WJ, Barkardottir RB, Gulcher JR, Thorsteinsdot-tir U, Kong A, Stefansson K: Genome-wide association study identifies a second prostate cancer susceptibility variant at 8q24. Nat Genet 2007; 39: 631–637.
Related Documents











