Gene expression profiling of normal and ruptured canine anterior cruciate ligaments D. N. Clements B.Sc., B.V.Sc.yz*, S. D. Carter Ph.D., Professory, J. F. Innes Ph.D., Professory, W. E. R. Ollier Ph.D., Professorz and P. J. R. Day Ph.D.zx y Musculoskeletal Research Group, Faculty of Veterinary Science, University of Liverpool, Liverpool, L69 3BX, UK z Centre for Integrated Genomic Medical Research, The Stopford Building, University of Manchester, Oxford Road, Manchester, M13 9PT, UK x ISASdInstitute for Analytical Sciences, Bunsen-Kirchhoff-Street 11, 44139 Dortmund, Germany Summary Objective: To identify genes which may be involved in the development of anterior cruciate ligament (ACL) laxity and rupture in a naturally occurring canine osteoarthritis (OA) model. Design: Three groups of dog were studied: (1) dogs with ACL rupture; (2) dogs with intact ACLs from a breed predisposed to ACL rupture; (3) dogs with intact ACLs from a breed at very low risk of rupture. The transcriptomes of the ACLs from each group were compared using a whole genome microarray and quantitative reverse transcriptase polymerase chain reaction. Differential gene expression in ruptured canine ACLs was compared with that published in the literature for ruptured human ACLs. Results: No significant differences were identified between the gene expression profiles of normal ACLs of a breed predisposed to ACL rup- ture when compared to a breed relatively resistant to ACL rupture. A general pattern of increased protease and extracellular structural matrix gene expression was identified in the ruptured ACLs when compared to intact ACLs. The gene expression profiles of ruptured canine ACLs demonstrate similar patterns to those previously reported for ruptured human ACLs. Conclusions: A transcriptomic basis to breed specific risk for the development of canine ACL rupture was not identified. Although changes in matrix associated gene expression in the ruptured ACL are similar between humans and dogs, the molecular events which may predispose to ACL laxity and rupture were not defined. ª 2007 Osteoarthritis Research Society International. Published by Elsevier Ltd. All rights reserved. Key words: Dog, Anterior cruciate ligament, Microarray, Gene expression. Introduction Joint laxity is hypothesised to be an important contributor to the pathogenesis of human knee OA, with laxity in the valgusevarus 1 and anterioreposterior 1,2 planes increasing with the severity of OA. The anterior cruciate ligament (ACL) is the primary stabiliser of the knee joint, and rupture of this ligament results in joint instability and the develop- ment of osteoarthritis (OA) 3e5 . ACL rupture is identified more commonly in patients with knee OA 6 , although this is not necessarily a sequel to a previous traumatic event 6 . Pathological changes to the ACL resulting in knee laxity may predispose patients to knee OA. This hypothesis is supported by spontaneous animal models of OA which highlight the association of ligament laxity, specifically of the ACL, and the development of OA 7 . A similar spontane- ous condition is recognised in dogs where disease of the canine anterior (cranial) cruciate ligament results in a pro- gressive pathological failure of the ligament (rupture), the development of joint instability and secondary OA 8 . Exper- imental transection of the ACL in normal dogs results in the spontaneous and progressive development of stifle OA 9 , and highlights the importance of joint stability, and par- ticularly ACL integrity, to prevent OA. Epidemiological studies reveal that dogs demonstrate a breed-associated risk to ACL rupture 10,11 , with ‘‘at-risk’’ breeds such as the Labrador Retriever (LR) demonstrating much higher levels of risk than ‘‘protected’’ breeds, such as the Greyhound (GH). Dogs from breeds predisposed to ACL rupture have reduced ligament stiffness and reduce load to ultimate failure when compared to dogs from breeds format low risk of ACL rupture 12e14 . This implies that the ge- netic susceptibility to the development of ACL rupture man- ifests itself through changes in the mechanical properties of the ACL. Increased levels of pro-matrix metalloproteinase-2 (pro-MMP-2) have been identified in normal ACLs of dogs with a high-risk of ACL rupture (LR) compared to dogs with a low risk of ACL rupture (GH) 14 . Similar changes in gross ACL biomechanical properties (increasing laxity) have been related to molecular differences (increased MMP-2 and pro-MMP-2 protein) in the ACL in an animal model of spontaneous knee OA 7 , further supporting the link between the development of knee OA, knee laxity and molecular changes at a cellular level. *Address correspondence and reprint requests to: Dylan Neil Clements, B.Sc., B.V.Sc., Musculoskeletal Research Group, Faculty of Veterinary Science, University of Liverpool, Liverpool, L69 3BX, UK. Tel: 44-7803203551; Fax: 44-1517944219; E-mail: [email protected] Received 21 December 2006; revision accepted 6 June 2007. Osteoarthritis and Cartilage (2008) 16, 195e203 ª 2007 Osteoarthritis Research Society International. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.joca.2007.06.013 International Cartilage Repair Society 195

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Osteoarthritis and Cartilage (2008) 16, 195e203
ª 2007 Osteoarthritis Research Society International. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.joca.2007.06.013
InternationalCartilageRepairSociety
Gene expression profiling of normal and ruptured canineanterior cruciate ligamentsD. N. Clements B.Sc., B.V.Sc.yz*, S. D. Carter Ph.D., Professory, J. F. Innes Ph.D.,Professory, W. E. R. Ollier Ph.D., Professorz and P. J. R. Day Ph.D.zxyMusculoskeletal Research Group, Faculty of Veterinary Science, University of Liverpool,Liverpool, L69 3BX, UKzCentre for Integrated Genomic Medical Research, The Stopford Building, University of Manchester,Oxford Road, Manchester, M13 9PT, UKx ISASdInstitute for Analytical Sciences, Bunsen-Kirchhoff-Street 11, 44139 Dortmund, Germany
Summary
Objective: To identify genes which may be involved in the development of anterior cruciate ligament (ACL) laxity and rupture in a naturallyoccurring canine osteoarthritis (OA) model.
Design: Three groups of dog were studied: (1) dogs with ACL rupture; (2) dogs with intact ACLs from a breed predisposed to ACL rupture;(3) dogs with intact ACLs from a breed at very low risk of rupture. The transcriptomes of the ACLs from each group were compared usinga whole genome microarray and quantitative reverse transcriptase polymerase chain reaction. Differential gene expression in ruptured canineACLs was compared with that published in the literature for ruptured human ACLs.
Results: No significant differences were identified between the gene expression profiles of normal ACLs of a breed predisposed to ACL rup-ture when compared to a breed relatively resistant to ACL rupture. A general pattern of increased protease and extracellular structural matrixgene expression was identified in the ruptured ACLs when compared to intact ACLs. The gene expression profiles of ruptured canine ACLsdemonstrate similar patterns to those previously reported for ruptured human ACLs.
Conclusions: A transcriptomic basis to breed specific risk for the development of canine ACL rupture was not identified. Although changes inmatrix associated gene expression in the ruptured ACL are similar between humans and dogs, the molecular events which may predispose toACL laxity and rupture were not defined.ª 2007 Osteoarthritis Research Society International. Published by Elsevier Ltd. All rights reserved.
Key words: Dog, Anterior cruciate ligament, Microarray, Gene expression.
Introduction
Joint laxity is hypothesised to be an important contributor tothe pathogenesis of human knee OA, with laxity in thevalgusevarus1 and anterioreposterior1,2 planes increasingwith the severity of OA. The anterior cruciate ligament(ACL) is the primary stabiliser of the knee joint, and ruptureof this ligament results in joint instability and the develop-ment of osteoarthritis (OA)3e5. ACL rupture is identifiedmore commonly in patients with knee OA6, although thisis not necessarily a sequel to a previous traumatic event6.
Pathological changes to the ACL resulting in knee laxitymay predispose patients to knee OA. This hypothesis issupported by spontaneous animal models of OA whichhighlight the association of ligament laxity, specifically ofthe ACL, and the development of OA7. A similar spontane-ous condition is recognised in dogs where disease of thecanine anterior (cranial) cruciate ligament results in a pro-gressive pathological failure of the ligament (rupture), the
*Address correspondence and reprint requests to: Dylan NeilClements, B.Sc., B.V.Sc., Musculoskeletal Research Group,Faculty of Veterinary Science, University of Liverpool, Liverpool,L69 3BX, UK. Tel: 44-7803203551; Fax: 44-1517944219; E-mail:[email protected]
Received 21 December 2006; revision accepted 6 June 2007.
195
development of joint instability and secondary OA8. Exper-imental transection of the ACL in normal dogs results inthe spontaneous and progressive development of stifleOA9, and highlights the importance of joint stability, and par-ticularly ACL integrity, to prevent OA.
Epidemiological studies reveal that dogs demonstratea breed-associated risk to ACL rupture10,11, with ‘‘at-risk’’breeds such as the Labrador Retriever (LR) demonstratingmuch higher levels of risk than ‘‘protected’’ breeds, such asthe Greyhound (GH). Dogs from breeds predisposed toACL rupture have reduced ligament stiffness and reduceload to ultimate failure when compared to dogs from breedsformat low risk of ACL rupture12e14. This implies that the ge-netic susceptibility to the development of ACL rupture man-ifests itself through changes in the mechanical properties ofthe ACL. Increased levels of pro-matrix metalloproteinase-2(pro-MMP-2) have been identified in normal ACLs of dogswith a high-risk of ACL rupture (LR) compared to dogswith a low risk of ACL rupture (GH)14. Similar changes ingross ACL biomechanical properties (increasing laxity)have been related to molecular differences (increasedMMP-2 and pro-MMP-2 protein) in the ACL in an animalmodel of spontaneous knee OA7, further supporting thelink between the development of knee OA, knee laxityand molecular changes at a cellular level.

196 D. N. Clements et al.: Gene expression of canine anterior cruciate ligaments
The histological appearance of the normal canine ACLis very similar to that of the normal human ACL15, althoughthe identification of other features such as chondroidmetaplasia cells is more common in the normal canineACL16. Following human ACL rupture, four histologicalphases are recognised, namely, inflammation (0e3weeks), epiligamentous regenerations (3e8 weeks), prolif-eration (8e32 weeks), and remodelling (1e2 years)17.Although gross histological features of ACL rupture arealso similar in both humans and dogs, the ruptured canineACL demonstrates more severe matrix degeneration,chondroid metaplasia and inflammation on histologicalevaluation than ruptured human ACL16. Inflammation isa component of naturally occurring ACL rupture, as recog-nised in histological sections of the synovial membrane18
and ligament16, although synovial fluid cytology profile inthe vast majority of cases is non-inflammatory19.
Gene expression within the normal and pathological ACLhas not been extensively studied in either humans or ca-nines. In man the ruptured ACL expresses higher quantitiesof messenger RNA (mRNA) coding for Type I collagen(COL1), Type III collagen (COL3), biglycan (BGN) and tis-sue inhibitor of metalloproteinase-1 (TIMP-1), than the nor-mal (non-ruptured) ACL20. In dog, the mRNA expression ofmatrix metalloproteinase-2 (MMP-2) and matrix metallopro-teinase-9 (MMP-9), tartrate-resistant acid phosphatase(TRAP) and cathepsin S (CTSS) have been reported tobe increased in the ruptured ACL when compared to normalACL21, although studies of proteins present suggest thatpro-MMP-2 is increased in the ruptured canine ACL, butthere are no changes in active MMP-2, MMP-9, TIMP-1 orTIMP-222.
Differences between the transcriptome of ruptured andnormal tissue can be identified by gene expression profiling.Canine specific microarray (MA) platforms are now avail-able to perform genome wide expression profiling of caninecells from dissected tissues23. MA analysis has allowed theidentification of differential gene expression pathways inconnective tissues, such as cartilage and tendon, which fur-ther improves our understanding of the molecular pathwaysinvolved in OA24,25 and tendon repair26.
We hypothesised that altered gene expression profileswould be identified when comparing the transcriptome ofthe normal ACL from dogs of breeds predisposed to ACLrupture (LR), and those at low risk from ACL rupture(GH). Secondly, we hypothesised that in breeds at risk ofACL rupture differential expression of genes would be iden-tified between the transcriptomes of normal ACL and theruptured ACL. Finally, we hypothesised that genes whichhave been previously reported as being differentially ex-pressed in the ruptured human ACL would demonstratesimilar patterns of expression to those we identified in rup-tured canine ACL.
Methods
RNA EXTRACTION
Normal canine ACLs were harvested by sharp dissectionand from the knees (stifles) of dogs (seven LRs [mean age5.4 years {standard deviation (SD)þ/� 3.3 years, range1e10 years}, three male neutered, two female neutered,one entire male and one entire female mean weight28.7 kg {SDþ/� 2.4 kg, range 25e32 kg}], five GH [meanage 3.8 years {SDþ/� 4.1 years, range 1.5e10 years},three entire female, two entire male, mean weight 33.0 kg{SDþ/� 3.8 kg, range 30e38 kg}]) without any evidence
of knee pathology, and which were euthanatized for rea-sons unrelated to orthopaedic disease. The central third ofthe ligament was preserved. Ruptured canine ACLs were ob-tained from five LRs (mean age 7.7 years [SDþ/� 1.6years, range 5.5e10 years], three neutered male, one neu-tered female, one male, mean weight 33.4 kg [SD þ/�3.0 kg,range 30e37 kg]) during routine surgical treatment for theACL rupture (medial parapatellar arthrotomy), and storedin RNAlater� as recommended by the manufacturer(Qiagen Ltd., Crawley, UK). All dogs with ACL rupturehad radiographic evidence of stifle OA (osteophytosis).
RNA EXTRACTION AND QUALITY ASSESSMENT
Tissue samples were removed from RNAlater� and totalRNA was extracted using phenol/guanidine HCl reagents(Trizol�, Invitrogen Ltd., Dorset, UK) and isolated as previ-ously described including the use of an on-column DNA di-gestion step27,28, RNA integrity was analysed by evaluatingthe capillary electrophoresis trace (Agilent 2100 Bioana-lyser, Agilent Technologies, California, USA) of the sampleusing the RNA integrity number (RIN) algorithm29, Degrada-tion factor (DF)30 and ribosomal peak ratio. The samplesdetermined to have no, or minimal, loss of integrity weredeemed suitable for use in experiments in accordancewith a previously developed quality algorithm28.
RNA AMPLIFICATION
mRNA was amplified for each sample, starting with200 ng total RNA using a commercially available kit(Ambion T7 MEGAscript� high yield transcription kit,Ambion (Europe) Ltd, Huntingdon, Cambridge, UK) as pre-viously described31. A second round of mRNA amplificationwas performed using MessageAmp� amplified RNA(aRNA) Amplification kit (Ambion (Europe) Ltd), as de-scribed by the manufacturer. The aRNA was quantified us-ing a spectrophotometer.
aRNA LABELLING
Two micrograms of aRNA were labelled with Cyanine-3dCTP (Cy3) or Cyanine-5dCTP (Cy5), using a fluorescentdye labelling kit (Agilent Technologies UK Ltd., SouthQueensferry, UK) as described by the manufacturer. Fluo-rescence incorporation was determined using a spectropho-tometer, ensuring that > 750 ng cRNA was labelled, andthat the label incorporation was >8 pmol per mg RNA. Sam-ples were stored at �80�C until use.
MA HYBRIDISATION, SLIDE READING, DATA NORMALISATION
AND STATISTICS
Samples hybridised to a canine specific, custom de-signed, whole genome 44219 spot 60mer oligonucleotideMA chip32 and read using an Agilent DNA MA Slide Reader,and fluorescence data extracted by employing the AgilentFeature Extraction 8.5 software (Agilent Technologies UKLtd). For full details see Supplementary material. Datawere imported into Genedata Expressionist Analyst (Gene-data AG, Basel, Switzerland), and the Cy3 and Cy5 fluores-cence intensities normalised using locally weighted linearregression (LOWESS)33. Expression data were then ex-ported into Excel 2003 and comparison between groupswas achieved using paired student’s t tests. Comparisonsof the number of genes up- or down-regulated in both the

197Osteoarthritis and Cartilage Vol. 16, No. 2
normal GH ACL and ruptured LR ACL when compared tothe normal LR ACL were made using Chi squared analysis(Minitab v14.1, Minitab Ltd, Coventry, UK). Correction formultiple hypothesis testing was performed using the Benja-mini and Hochberg false discovery rate (FDR)34. Correct Pvalues were calculated by dividing the true P value by theindividual correction factor and multiplying by 0.05.
QUANTITATIVE REVERSE TRANSCRIPTASE POLYMERASE
CHAIN REACTION (RT-QPCR)
The original (un-amplified) mRNA samples used for theMA experiments, and further suitable high grade RNA sam-ple collections from further patients with stifle OA and ACLrupture (totals, n¼ 21 ruptured ACL [mean age 6.3 years{SDþ/� 2.7 years, range 2e12 years}, seven femaleneutered, eight male neutered, three entire female, threeentire male, mean weight 32.7 kg {SDþ/� 6.9 kg, range24e55 kg}], n¼ 13 normal LR ACLs [mean age 4.9 years{SDþ/� 3.0 years, range 1e10 years}, two female neutered,two male neutered, five entire male, four entire female, meanweight 28.8 kg {SDþ/� 3.5 kg, range 21e34 kg}]) and n¼ 7normal GH ACLs [mean age 3.7 years {SDþ/� 3.0 years,range 1.5e10 years}, three entire female, four entire male,mean weight 31.7 kg {SDþ/� 2.0 kg, range 30e35 kg}])were obtained and used in RT-qPCR experiments. Reversetranscription was performed as previously described35. Theage and weights were compared by student’s t tests, and sexand neuter status were compared by Fisher’s exact tests.Significance was set at P< 0.05.
ASSAY DESIGN
Genes were selected for assaying on the basis of theirperceived relevance to the ACL extracellular matrix (from lit-erature review), and the results of the MA screen. Assayswere designed for quantification of expression of 23 genesof interest and five reference genes, as previously de-scribed35. The primer and probe sequences for each assayhave been previously published35 or are listed in Table I.The assays were used to quantify; three collagen genes(Type I collagen, alpha two chain (COL1A2), Type III colla-gen alpha 1 chain (COL3A1), Type V collagen alpha 1 chain(COL5A1)), seven extracellular matrix genes (aggrecan(AGC1), BGN, chondroitin sulphate proteoglycan 2 (Versi-can/CSPG2), decorin (DCN), lumican (LUM), tenascin C(TNC), vimentin (VIM)), proteases and their inhibitors (a dis-integrin and metalloproteinase with thrombospondin-likemotif-4 (ADAMTS-4), -5 (ADAMTS-5), cathepsin B (CTSB)and D (CTSD), MMP-2, MMP-9, and MMP-13, caspase-8(CASP-8), tissue inhibitors of metalloproteinase-1 (TIMP-1)and-2 (TIMP-2)), a growth factor (insulin like growth
TablePrimer and probe sequences fo
Gene Forward
ADAMTS-4 GACCAGTGCAAACTCACCTGB2M CCTTGCTCCTCATCCTCCTCASP-8 GAGCTTCAGATACCAGGCAGACOX2 AAATTGCTGGCAGGGTTGIGF-1 GGGGGTTCTACTTCAACAAGCMMP-2 ACCTGCAAGGCAGTGGTCMMP-9 CACGCATGACATCTTCCAGTNOS2A GGCTCAAATCACAACGGAAT
factor-1 (IGF-1)), prostaglandin-G, -H synthetase-2(PTGS-2), inducible nitric oxide synthetase 2A (NOS2A),genomic DNA (GEN), and five reference genes (selectedusing a previously published reference gene stability algo-rithm36; beta 2-microglobulin (B2M), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), TATA box bindingprotein (TBP), ribosomal protein L13a (RPL13A) and succi-nate dehydrogenase complex, subunit A (SDHA)). Lockednucleic acid probes with a 50 reporter dye FAM (6-carboxyfluorescein) and 30 quencher dye TAMRA (6-carboxytetra-methyl-rhodamine) were synthesised by Roche DiagnosticsLtd. (Lewes, West Sussex, UK).
The RT-qPCR assays were all performed in triplicate us-ing a TaqMan� ABI PRISM 7900 SDS (Applied Biosys-tems, California, USA) in 10 ml volumes in 384-well plateformat, as previously described35, and are further describedin Supplementary material.
RT-qPCR DATA ANALYSIS
Geometric means36 were calculated for the combinedthree reference genes (B2M, SDHA, TBP), and used tocalculate the deltaedelta CT values and the relative amountof each target gene37. Two reference genes (GAPDH andRPL13A) were not included in the calculations, becausethey had near-differential expression between the normaland ruptured ligament samples, and the normal LR andGH ACL samples respectively, even when included asa part of the normalisation calculation. Quantitative PCRdata for each group were compared with the calculationsof means, SDs, fold changes from normal and paired two-tailed t tests (body weight and age) performed in a spread-sheet program (Microsoft Excel 2003). Significance wasestablished at P< 0.05, and data were checked for errorsdue to multiple hypothesis testing using the Benjamini andHochberg false discovery rate (FDR)34.
CLUSTER ANALYSIS
The normalised MA data for 63 genes differentially ex-pressed between ruptured (LR) ACL when compared tonormal (LR) ACL and with complete annotation were loadedinto a gene clustering software (Cluster, Eisen Labs, http://rana.lbl.gov/EisenSoftware.htm). Data were log trans-formed and genes centered to the mean. Hierarchicalclustering of differentially expressed genes was thenperformed for arrays and genes using Spearman’s RankCorrelation and complete linkage link clustering. Clusteringof genes and arrays were visualised with publicly availablesoftware (TreeView, Eisen Labs, http://rana.lbl.gov/EisenSoftware.htm).
Ir quantitative PCR assays
Reverse Probe
CAGGGAGTCCCATCTACCAC GGCCCTGGTGGGTGTCGTGAGTACACTTG CAGCATCCTGAAATCTGAAAAAGCATGACC CTCTGCCTTCGAAGCTTTTGCTACTTGTTG GGTGGCAGTCATCCACGATGCCTGTCT CTCCAGCATCCAAATTTCACGCTTTTCA AGCTGGAGCGAGAATTCACACGCCAGTA CTTCTGCCAGAGCTCGACCAGGAGAGTG CCAGCCGC
198 D. N. Clements et al.: Gene expression of canine anterior cruciate ligaments
Results
COMPARISON OF PATIENT SIGNALMENT
A significant difference (P¼ 0.025) was noted in the pa-tient weight of tissues used in the MA experiment to com-pare the ruptured ACL LRs and the normal ACL LRs. Asecond significant difference (P¼ 0.024) was noted in neu-ter status of the patients used in the RT-qPCR experimentto compare the ruptured ACL LRs (more likely to be neu-tered) and the normal ACL LRs. No other differences inage, weight sex or neuter status were determined.
MA
In the normal GH ACL, compared to the normal LR ACL,925 transcripts were up-regulated (P< 0.05). Conversely1050 transcripts were down-regulated in the normal GHACL, compared to the normal LR ACL. Of the 925 tran-scripts up-regulated in the normal GH ACL, 455 were alsosignificantly up-regulated in the ruptured LR ACL when
Table IIdentification of genes up- or down-regulated (three fold or more) when c
analysi
Gene ID Name Ref seq numbe
NAP1L1 Nucleosome assemblyprotein 1-like 1
XM_847704
COL3A1 Collagen 3, alpha 1 XM_858055RGS10 Regulator of G-protein signalling 10 XM_535032MARCKS Myristoylated alanine-rich
protein kinase C substrateXM_850164
CDH11 Cadherin 11, type 2, OB-cadherin XM_859908SPARC Osteonectin XM_8499STMN1 Stathmin 1 XM_861033ZSWIM2 Zinc finger, SWIM domain
containing 2XM_535994
TMSB10 Thymosin beta-10 XM_849812LGALS1 Lectin, galactose
binding, soluble 1XM_549042
CENPH Centromere protein H XM_847537ALDH1L2 Aldehyde dehydrogenase 1 family,
member L2XM_531763
ANKRD10 Ankyrin repeatdomain 10
XM_843234
RET RET tyrosinekinase/camp proteinkinase A subunit RI
XM_543915
CAV1 Caveolin 1 NM_001003296TUBB Beta-tublin XM_532060TUBA Alpha-tubulin XM_857454ATP11B Atpase, class I, type 11B XM_535816WDR1 WD repeat
domain 1XM_848702
SNX6 Sorting nexin 6 XM_547770ACTB Beta actin XM_845524IGFBP7 Insulin-like growth
factor binding protein 7XM_856005
ADK Adenosine kinase,transcript variant 3
XM_858911
CCNB1 Cyclin B1 XM_544149SSR2 Signal sequence
receptor, betaNM_001003269
CIDEB Cell death-inducing DNA fragmentationfactor, alphasubunit-like effector B
XM_848904
FAT4 FAT tumoursuppressor homolog 4
XM_843601
ACTR3 Actin-related protein 3 XM_848216
compared to the normal LR ACL, and 450 of the 1070 tran-scripts were down-regulated in both the normal GH ACLand LR ACL when compared to the normal LR ACL. Thenumber of transcripts whose expression was increased ordecreased in both normal GH ACL and ruptured LR ACLwas significantly greater (P< 0.001) than by chance alone.The FDR determined that none of the transcripts were sig-nificantly up- or down-regulated when corrected for multiplehypothesis testing.
Four thousand and thirty-eight transcripts were up-regu-lated in ruptured LR ACL when compared to normal LRACL. Five thousand and nineteen transcripts were down-regulated in ruptured LR ACL when compared to the normalLR ACL. The FDR determined that 99 transcripts were sig-nificantly up-regulated and 17 transcripts were significantlydown-regulated when corrected for multiple hypothesis test-ing. Eighty-seven transcripts (of which 24 transcripts wererepeats) had a defined annotation and 29 transcripts hadno defined annotation. The annotated transcripts whose ex-pression was changed by three fold are listed in Table II.
Iompared between ruptured ACL and normal ACL in LRs in the MAs
r Function Fold change Corrected P value
Nucleic acid processing 19.0þ/� 7.4 0.0373
Structure 9.5þ/� 4.0 0.0328Signalling 8.2þ/� 2.5 0.0220Structure 7.8þ/� 2.2 0.0189
Structure 6.9þ/� 2.4 0.0476Signalling 6.3þ/� 2.1 0.0387Structure 6.2þ/� 1.7 0.0194Unknown 5.9þ/� 1.9 0.0328
Structure 5.7þ/� 1.9 0.0388Signalling 5.4þ/� 1.3 0.0195
Nucleic acid processing 5.4þ/� 1.8 0.0480Metabolism 5.3þ/� 1.6 0.0345
Nucleic acid processing 5.2þ/� 1.3 0.0207
Signalling 4.9þ/� 1.7 0.0478
Structure 4.8þ/� 0.9 0.0137Structure 4.8þ/� 1.4 0.0255Structure 4.7þ/� 1.0 0.0195Transport 4.7þ/� 1.5 0.0381Structure 4.4þ/� 1.3 0.0263
Signalling 4.4 þ/� 0.9 0.0158Structure 3.8þ/� 0.8 0.0188Signalling 3.7þ/� 0.8 0.0196
Signalling 3.6þ/� 1.1 0.0472
Structure 3.6þ/� 0.4 0.0009Transport 3.5þ/� 0.9 0.0342
Signalling 3.2þ/� 0.9 0.0477
Signalling 3.0þ/� 0.7 0.0341
Structure 0.3 þ/� 0.1 0.0379
199Osteoarthritis and Cartilage Vol. 16, No. 2
RT-qPCR
The results of the RT-qPCR are presented in Table III.Two genes (COL5A1 and RPL13A) were determined tobe up-regulated (P< 0.05) in the normal LR ACL whencompared to the normal GH ACL. The FDR determinedthat neither gene was significantly up-regulated.
Sixteen genes were significantly up-regulated (n¼ 14) ordown-regulated (n¼ 2) in the ruptured LR ACL, when com-pared to the normal LR ACL. The FDR determined that 14of these genes were significantly up-regulated, after cor-recting for multiple hypothesis testing (AGC, CASP-8,COL1A2, COL3A1, COL5A1, CTSB, CTSD, IGF-1, LUM,MMP-2, MMP-9, COX2, TIMP-1, and TNC) and two weresignificantly down-regulated (DCN, TIMP-2).
When the RT-qPCR results for the sample subsets usedfor MA analysis were examined separately, 12 genes weresignificantly up-regulated in the ruptured LR ACL whencompared to the normal LR ACL, of which eight geneswere still significantly up-regulated after FDR correction(AGC, CASP-8, COL1A2, COL3A1, CTSB, MMP9, TIMP-1,and TNC). One gene was determined as being up-regulatedin the normal LR ACL when compared to the normal GHACL (IGF-1), although this was not significant when cor-rected for multiple hypothesis testing.
When FDR correction was applied to MA data, but onlyto the 24 candidate genes evaluated by RT-qPCR, then 11genes were determined to be significantly up-regulated(CASP-8, COL1A2, COL3A1, COL5A1, CTSB, GAPDH,IGF-1, LUM, MMP-13, TIMP-1, and TNC). Nine of thesegenes (with the exceptions being MMP-13 and GAPDH)were confirmed to be significantly up-regulated in thelarger cohort of samples evaluated the RT-qPCR. WhenFDR correction was applied to the complete MA data set(41623 genes), only two genes evaluated by RT-qPCR
Table IGene expression folds change and significance (uncorrected P values)(GH) as determined by MA, RT-PCR of MA samples (MA RT-PCR), and R
(to 24 genes) ar
MA
Ratio LDvs LN
P value Ratio GHvs LN
P value Ratio LDvs LN
P value
ADAMTS-4 1.0 0.982 1.1 0.408 0.9 0.881ADAMTS-5 0.8 0.605 1.6 0.286 0.3 0.248AGC 3.9 0.043 0.5 0.126 13.9 0.007BGN 1.7 0.086 0.9 0.795 0.9 0.916CASP-8 1.8 0.000 0.9 0.479 2.1 0.003COL1A2 12.8 0.001 0.7 0.306 14.3 0.004COL3A1 9.5 0.000 0.6 0.086 18.7 0.004COL5A1 3.9 0.004 0.9 0.727 6.4 0.025CSPG2 0.8 0.078 0.8 0.818 1.0 0.893CTSB 6.0 0.012 0.7 0.293 5.6 0.010CTSD 1.0 0.475 1.0 0.756 1.9 0.201DCN 0.7 0.100 0.9 0.742 0.5 0.053GAPDH 2.6 0.011 0.6 0.124 1.4 0.327IGF-1 2.2 0.001 1.3 0.029 3.4 0.086LUM 3.5 0.012 0.4 0.035 3.5 0.025MMP-13 2.2 0.023 1.2 0.420 13.5 0.051MMP-2 36.1 0.042 1.5 0.345 34.1 0.032MMP-9 1.5 0.068 1.1 0.758 336.4 0.013NOS2 0.7 0.515 1.8 0.450 0.0 0.324PTGS-2 1.0 0.990 1.3 0.228 4.9 0.033TIMP-1 6.2 0.020 0.9 0.791 3.0 0.002TIMP-2 1.0 0.889 0.6 0.143 0.5 0.107TNC 2.4 0.008 1.1 0.136 4.6 0.008VIM 1.1 0.607 0.7 0.125 1.1 0.601
were differentially expressed (CASP-8 and COL3A1).Both these genes were also differentially expressed inthe larger cohort of samples evaluated by RT-qPCR.Overall, for genes significantly up- or down-regulated byeither MA (n¼ 2), RT-qPCR of a larger cohort of samples(n¼ 7) or both techniques (n¼ 9), the direction foldchange in expression agreed for all except one gene(TIMP-2, down-regulated in RT-qPCR samples). A graph-ical representation of the fold changes in expression ofgenes up-regulated in ruptured LR ACL, as determinedby MA, is illustrated in Fig. 1.
CLUSTER ANALYSIS
The corresponding hierarchical cluster analysis plot ispresented in Fig. 2. Clustering of the arrays illustrates clearseparation of the ruptured and normal samples, which is un-surprising as the list of genes clustered was filtered on sig-nificance of differential expression between normal andruptured. Samples of three of the four dogs from a breedat low risk of ACL rupture (GH) demonstrated patterns ofgene expression which clustered together, and whichwere more closely related to ruptured ACL than thosefrom dogs at high-risk of ACL rupture. Clustering of thegenes demonstrated alignment of genes known to havesimilar function, such as alpha-tubulin (TUBA), beta-tubulin(TUBB), alpha-actin (ACTA) and beta-actin (ACTB) withinclosely related branches, supports both the method of anal-ysis and the validity of the data.
Discussion
Differential gene expression was identified in rupturedACL when compared to normal LR ACL using both
IIin ruptured LR ACL (D), normal LR ACL (N) and normal GH ACLT-qPCR of all samples (significant results, surviving FDR correctione in bold)
MA RT ALL RT
Ratio GH vs LN P value Ratio LDvs LN
P value Ratio GHvs LN
P value
51.7 0.365 2.7 0.084 3.4 0.5584.4 0.278 0.6 0.217 1.9 0.4270.9 0.759 14.3 0.000 1.1 0.8141.3 0.702 1.6 0.133 1.2 0.6520.9 0.630 1.5 0.028 0.9 0.5740.9 0.849 11.5 0.000 1.0 0.9050.9 0.839 16.0 0.000 1.1 0.8540.5 0.117 9.9 0.003 0.5 0.0190.7 0.498 1.0 0.910 0.7 0.3190.9 0.499 4.7 0.000 1.0 0.9202.1 0.115 2.9 0.001 1.3 0.3890.7 0.175 0.5 0.000 0.8 0.4621.1 0.606 1.6 0.043 1.2 0.2044.3 0.042 3.8 0.003 2.0 0.0710.7 0.226 2.4 0.002 0.8 0.2383.3 0.449 21.9 0.038 2.7 0.4021.2 0.612 24.5 0.000 1.1 0.7931.2 0.826 491.9 0.018 1.5 0.526
20.1 0.395 0.2 0.225 9.6 0.4451.4 0.622 6.0 0.028 0.7 0.4741.5 0.492 5.3 0.001 1.0 0.9520.6 0.180 0.6 0.006 0.7 0.1320.9 0.897 4.7 0.000 0.9 0.6500.9 0.567 1.1 0.418 1.1 0.593

Fig. 1. Fold change in gene expression patterns of candidate genesin ruptured ACL when compared to normal ACL from high-risk dogs(LR) as determined by MA, RT-qPCR of MA samples (MA RT-qPCR), and RT-qPCR of all samples. The candidate genesselected for comparison are those differentially expressed by MA
evaluation.
200 D. N. Clements et al.: Gene expression of canine anterior cruciate ligaments
expression profiles generated by MA and RT-qPCR. Theoverall pattern of gene expression reported in the rupturedACL suggests that both catabolism (MMP and CTS produc-tion) and repair (collagen and extracellular matrix produc-tion)38 are increased in the ruptured canine ACL whencompared to the normal ACL. The changes in gene expres-sion are consistent with both histological features of colla-gen disruption and epiligamentous repair39 and molecularchanges21 reported in the ruptured canine ACL. CompleteACL rupture was diagnosed in all joints from which dis-eased tissues were obtained. To date, the results of primaryrepair of ruptured ACLs have been poor both in man40 andanimal models38. Thus the anabolic response of the rup-tured ACL, which has lost its ability to resist mechanicalload, would appear to be futile.
A transcriptomic basis for breed risk to ACL rupture wasnot identified. Over 40% of genes up- or down-regulated inthe GH ACLs (before correction for multiple hypothesis test-ing) showed a similar differential expression as observed inthe transcriptome of ruptured LR ACLs, suggesting that ex-pression profiles reported for normal GH ACLs were moreakin to those identified in the ruptured LR ACLs. Further-more, hierarchical cluster analysis of the most differentiallyexpressed genes in ruptured ACL demonstrated clusteringand separation of the expression profiles of three of thefour low risk (GH) normal ACL samples from the high-risk(LR) normal ACL samples. Thus, a transcriptomic basis tothe breed specific risk may exist, but our methods werenot sensitive enough to characterise it. Alternatively thesmall sample size evaluated by MA may have dictatedthat individuals in the high-risk group were simply not atrisk of ACL rupture, although the breed itself is, becauseACL rupture does not affect all individuals in the breed.Conversely, one may interpret the results to indicate thata transcriptomic risk to the development of ACL rupture trulydoes not exist, despite differences in relative risk of ACLrupture between dog breeds10,11, and biomechanical12,14
and biochemical14,41 differences in the intact ACL betweendog breeds protected or at risk of ACL rupture. Clearly,there are advantages in using laboratory animal models ofdisease, which demonstrate a consistent phenotype, thusproviding more homogeneous information as to the molec-ular basis to the disease. However, such models may not
always reflect polygenetic nature to a disease, or the asso-ciated environmental influences, which the canine popula-tion shares with its human counterparts.
Although no pathways were consistently represented inthe differentially expressed genes as determined by MA,a number of interesting and functionally related rupture as-sociated genes were up-regulated. ACTA, ACTB, TUBBand TUBA are all genes encoding intracellular structuralmolecules, whose up-regulation suggests and increase incytoplasmic activity in ruptured LR ACLs. SPARC (osteo-nectin), an extracellular matrix protein which is involved inligament development, remodelling and repair42, was alsoincreased in ruptured LR ACLs, suggesting that the genemay have a key role in the response to ACL rupture. Themajority of the genes identified as being differentially ex-pressed in ruptured ACL have no previous known associa-tion with OA or ligament pathology, which makes theirprecise role in the ruptured ACL difficult to define.
The normal resorption of ACL matrix collagen has beenhypothesised to occur by fibroblast phagocytosis and intra-cellular digestion with lysosomal CTSs, whereas inflamma-tory remodelling of collagen is thought to be mediated byMMPs 43. Our results suggest that both processes are ac-tive, as there were increases in the expression of bothCTSB and CTSD and MMP-2 and MMP-9 expressionswere identified in the ruptured ACL, although the relative im-portance of expression changes in each of the moleculesrequires further study. Ligament CTSB expression increaseswith mechanical stress44, and the experimental inductionof immune-mediated synovitis increases CTSD in synovialfluid, with concomitant reduction in the mechanical proper-ties of the ACL45. At the protein level, pro-MMP-2 is raisedin ruptured ACLs in dogs22. Increased TIMP-1 expressionwas identified in ruptured ACLs, which is consistent withevaluation of gene expression in ruptured ACLs20, althoughthis increase does not reflect enzyme activity in rupturedcanine ACLs22. The reduced expression of TIMP-2 in ACLrupture is interesting, because this is contrary to the changeone may expect on the basis of its biological activity yet isconsistent with similar reductions in the expression of thisgene reported in osteoarthritic articular cartilage35,46 andreductions in the level of protein of this gene reported inthe synovial fluid of dogs with ACL rupture47.
Up-regulation of extracellular matrix gene expression(COL1A2, COL3A1, COL5A1, AGR, LUM and TNC) sug-gests increased extracellular matrix production as part ofan attempted reparative process. This concurs with reportsdocumenting an increased expression of COL1 and COL3in the ruptured human ACL when compared to normal(non-ruptured) ACLs20 although we did not record an in-creased expression of BGN in ruptured canine ACLs, ashas been reported for ruptured human ACLs20. Reducedexpression of a number of these genes (COL1, COL3,DCN and LUM) is reported in the ACL of rabbits duringpregnancy, and is associated with a concurrent increasein ACL laxity48. We did not record a detectable differencein the expression of these genes in the normal ACL oftwo breeds known to demonstrate different laxity measure-ment14. Interestingly a similar pattern of gene expressionchanges (increased COL1A2, COL3A1, COL5A1, LUMand TNC gene expression) is also reported in end-stage ca-nine hip OA cartilage35, which suggests that these changesreflect a more primordial mesenchymal tissue response inOA. TNC expression is directly related to mechanical loadin fibroblasts49, thus it increased expression in the rupturedACL and may represent a response to increased loadingprior to rupture, or an attempt at ligament repair, and
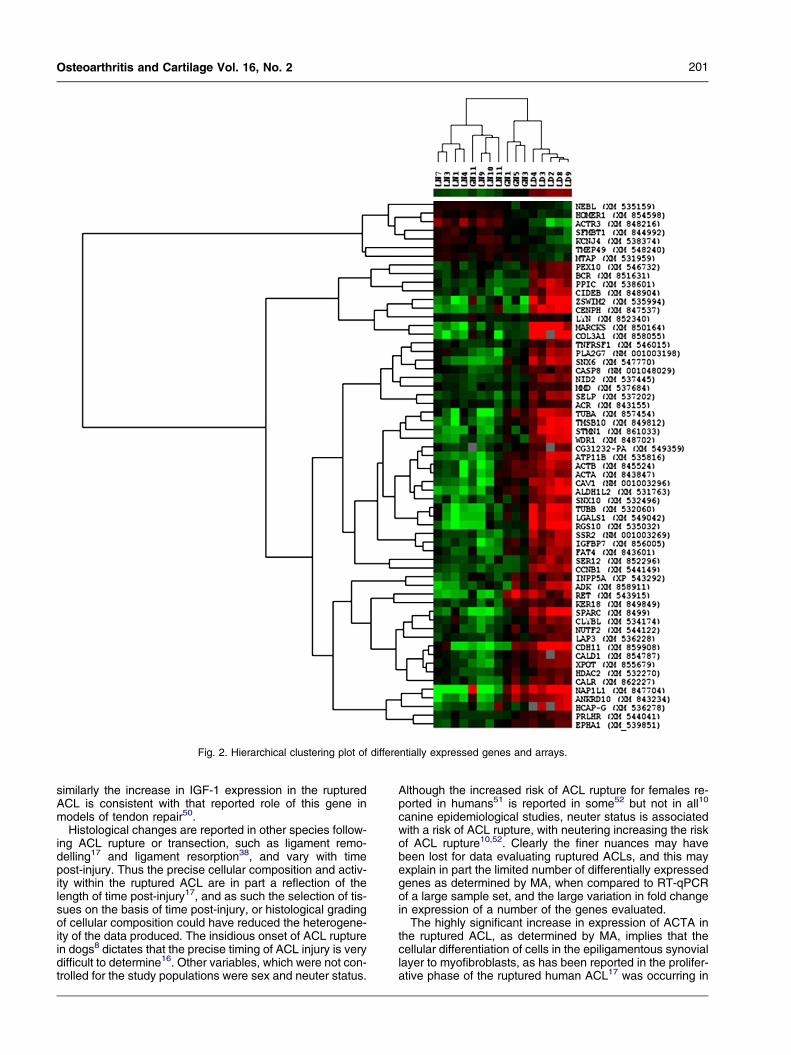
Fig. 2. Hierarchical clustering plot of differentially expressed genes and arrays.
201Osteoarthritis and Cartilage Vol. 16, No. 2
similarly the increase in IGF-1 expression in the rupturedACL is consistent with that reported role of this gene inmodels of tendon repair50.
Histological changes are reported in other species follow-ing ACL rupture or transection, such as ligament remo-delling17 and ligament resorption38, and vary with timepost-injury. Thus the precise cellular composition and activ-ity within the ruptured ACL are in part a reflection of thelength of time post-injury17, and as such the selection of tis-sues on the basis of time post-injury, or histological gradingof cellular composition could have reduced the heterogene-ity of the data produced. The insidious onset of ACL rupturein dogs8 dictates that the precise timing of ACL injury is verydifficult to determine16. Other variables, which were not con-trolled for the study populations were sex and neuter status.
Although the increased risk of ACL rupture for females re-ported in humans51 is reported in some52 but not in all10
canine epidemiological studies, neuter status is associatedwith a risk of ACL rupture, with neutering increasing the riskof ACL rupture10,52. Clearly the finer nuances may havebeen lost for data evaluating ruptured ACLs, and this mayexplain in part the limited number of differentially expressedgenes as determined by MA, when compared to RT-qPCRof a large sample set, and the large variation in fold changein expression of a number of the genes evaluated.
The highly significant increase in expression of ACTA inthe ruptured ACL, as determined by MA, implies that thecellular differentiation of cells in the epiligamentous synoviallayer to myofibroblasts, as has been reported in the prolifer-ative phase of the ruptured human ACL17 was occurring in

202 D. N. Clements et al.: Gene expression of canine anterior cruciate ligaments
the ruptured canine ACL. Furthermore, the significant in-creases in other matrix components suggested a prolifera-tive response, whilst an epiligamentous inflammationrecorded on histological studies of ruptured canine ACL53
is indicated by the increase in COX2 expression.In summary, we could not identify a transcriptomic basis
to the breed specific risk for the development of canine ACLrupture, although a large number of new genes were iden-tified as being differentially expressed in ruptured ACL.The expression profiles of ruptured ACLs were similar tothose previously reported for ruptured human ACLs20.
Acknowledgements
This project was funded by the PetPlan Charitable Trustand the Biotechnology and Biological Sciences ResearchCouncil.
Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.joca.2007.06.013.
References
1. Wada M, Imura S, Baba H, Shimada S. Knee laxity inpatients with osteoarthritis and rheumatoid arthritis.Br J Rheumatol 1996;35:560e3.
2. Sharma L, Lou C, Felson DT, Dunlop DD, Kirwan-Mellis G, Hayes KW, et al. Laxity in healthy and oste-oarthritic knees. Arthritis Rheum 1999;42:861e70.
3. von Porat A, Roos EM, Roos H. High prevalence ofosteoarthritis 14 years after an anterior cruciate liga-ment tear in male soccer players: a study of radio-graphic and patient relevant outcomes. Ann RheumDis 2004;63:269e73.
4. Kannus P, Jarvinen M. Posttraumatic anterior cruciateligament insufficiency as a cause of osteoarthritis ina knee joint. Clin Rheumatol 1989;8:251e60.
5. Jacobsen K. Osteoarthrosis following insufficiency ofthe cruciate ligaments in man. A clinical study. ActaOrthop Scand 1977;48:520e6.
6. Hill CL, Seo GS, Gale D, Totterman S, Gale ME,Felson DT. Cruciate ligament integrity in osteoarthritisof the knee. Arthritis Rheum 2005;52:794e9.
7. Quasnichka HL, Anderson-MacKenzie JM, Tarlton JF,Sims TJ, Billingham ME, Bailey AJ. Cruciate ligamentlaxity and femoral intercondylar notch narrowing inearly-stage knee osteoarthritis. Arthritis Rheum 2005;52:3100e9.
8. Bennett D, Tennant B, Lewis DG, Baughan J, May C,Carter S. A reappraisal of anterior cruciate ligamentdisease in the dog. J Small Anim Pract 1988;29:275e97.
9. Brandt KD, Braunstein EM, Visco DM, O’Connor B,Heck D, Albrecht M. Anterior (cranial) cruciate liga-ment transection in the dog: a bona fide model of os-teoarthritis, not merely of cartilage injury and repair.J Rheumatol 1991;18:436e46.
10. Whitehair JG, Vasseur PB, Willits NH. Epidemiology ofcranial cruciate ligament rupture in dogs. J Am VetMed Assoc 1993;203:1016e9.
11. Duval JM, Budsberg SC, Flo GL, Sammarco JL. Breed,sex, and body weight as risk factors for rupture of the
cranial cruciate ligament in young dogs. J Am Vet MedAssoc 1999;215:811e4.
12. Wingfield C, Amis AA, Stead AC, Law HT. Comparisonof the biomechanical properties of rottweiler and rac-ing greyhound cranial cruciate ligaments. J SmallAnim Pract 2000;41:303e7.
13. Wingfield C, Amis AA, Stead AC, Law HT. Cranial cru-ciate stability in the rottweiler and racing greyhound:an in vitro study. J Small Anim Pract 2000;41:193e7.
14. Comerford EJ, Tarlton JF, Innes JF, Johnson KA,Amis AA, Bailey AJ. Metabolism and composition ofthe canine anterior cruciate ligament relate to differ-ences in knee joint mechanics and predisposition toligament rupture. J Orthop Res 2005;23:61e6.
15. Murray MM, Weiler A, Spindler KP. Interspecies varia-tion in the fibroblast distribution of the anterior cruciateligament. Am J Sports Med 2004;32:1484e91.
16. Barrett JG, Hao Z, Graf BK, Kaplan LD, Heiner JP,Muir P. Inflammatory changes in ruptured canine cra-nial and human anterior cruciate ligaments. Am J VetRes 2005;66:2073e80.
17. Murray MM, Martin SD, Martin TL, Spector M. Histolog-ical changes in the human anterior cruciate ligamentafter rupture. J Bone Joint Surg Am 2000;82:1387.
18. Lemburg AK, Meyer-Lindenberg A, Hewicker-Trautwein M. Immunohistochemical characterizationof inflammatory cell populations and adhesion mole-cule expression in synovial membranes from dogswith spontaneous cranial cruciate ligament rupture.Vet Immunol Immunopathol 2004;97:231e40.
19. Griffin DW, Vasseur PB. Synovial fluid analysis in dogswith cranial cruciate ligament rupture. J Am AnimHosp Assoc 1992;28:277e81.
20. Lo IK, Marchuk LL, Hart DA, Frank CB. Comparison ofmRNA levels for matrix molecules in normal and disru-pted human anterior cruciate ligaments using reversetranscription-polymerase chain reaction. J OrthopRes 1998;16:421e8.
21. Muir P, Danova NA, Argyle DJ, Manley PA, Hao Z. Col-lagenolytic protease expression in cranial cruciate lig-ament and stifle synovial fluid in dogs with cranialcruciate ligament rupture. Vet Surg 2005;34:482e90.
22. Comerford EJ, Innes JF, Tarlton JF, Bailey AJ. Investi-gation of the composition, turnover, and thermal prop-erties of ruptured cranial cruciate ligaments of dogs.Am J Vet Res 2004;65:1136e41.
23. Burton-Wurster N, Mateescu RG, Todhunter RJ,Clements KM, Sun Q, Scarpino V, et al. Genes incanine articular cartilage that respond to mechanicalinjury: gene expression studies with affymetrix caninegenechip. J Hered 2005;96:821e8.
24. Aigner T, Zien A, Gehrsitz A, Gebhard PM, McKenna L.Anabolic and catabolic gene expression pattern analy-sis in normal versus osteoarthritic cartilage using com-plementary DNA-array technology. Arthritis Rheum2001;44:2777e89.
25. Aigner T, Fundel K, Saas J, Gebhard PM, Haag J, Weiss T,et al. Large-scale gene expression profiling reveals majorpathogenetic pathways of cartilage degeneration in oste-oarthritis. Arthritis Rheum 2006;54:3533e44.
26. Molloy TJ, Wang Y, Horner A, Skerry TM, Murrell GA.Microarray analysis of healing rat Achilles tendon: ev-idence for glutamate signaling mechanisms and em-bryonic gene expression in healing tendon tissue.J Orthop Res 2006;24:842e55.
27. Reno C, Marchuk L, Sciore P, Frank CB, Hart DA.Rapid isolation of total RNA from small samples of

203Osteoarthritis and Cartilage Vol. 16, No. 2
hypocellular, dense connective tissues. Biotechniques1997;22:1082e6.
28. Clements DN, Vaughan-Thomas A, Peansukmanee S,Carter SD, Innes JF, Ollier WER, et al. Assessment ofthe use of RNA quality metrics for the screening of nor-mal and pathological canine articular cartilage sam-ples. Am J Vet Res 2006;67:1438e44.
29. Imbeaud S, Graudens E, Boulanger V, Barlet X,Zaborski P, Eveno E, et al. Towards standardizationof RNA quality assessment using user-independentclassifiers of microcapillary electrophoresis traces. Nu-cleic Acids Res 2005;33:e56.
30. Auer H, Lyianarachchi S, Newsom D, Klisovic MI,Marcucci U, Kornacker K. Chipping away at the chipbias: RNA degradation in microarray analysis. NatGenet 2003;35:292e3.
31. Goff LA, Bowers J, Schwalm J, Howerton K, Getts RC,Hart RP. Evaluation of sense-strand mRNA amplifica-tion by comparative quantitative PCR. BMC Genomics2004;5:76.
32. Jones P, Jones C, Fretwell N, Martin A, Soloviev M. De-sign and production of a whole genome dog oligonu-cleotide microarray. 2004.
33. Workman C, Jensen LJ, Jarmer H, Berka R, Gautier L,Nielser HB, et al. A new non-linear normalizationmethod for reducing variability in DNA microarray ex-periments. Genome Biology 2002;3. research0048.
34. Benjamini Y, Hochberg Y. Controlling the false discov-ery rate; a practical and powerful approach to multipletesting. J Royal Stat Soc Ser B 1995;57:289e300.
35. Clements DN, Carter SD, Innes JF, Ollier WE, Day PJ.Analysis of normal and osteoarthritic canine cartilagemRNA expression by quantitative-PCR. Arthritis ResTher 2006;8:R158.
36. Vandesompele J, De Preter K, Pattyn F, Poppe B, VanRoy N, De Paepe A, et al. Accurate normalization ofreal-time quantitative RT-PCR data by geometric aver-aging of multiple internal control genes. Genome Biol-ogy 2002;3:0034.1e0034.11.
37. Livak KJ, Schmittgen TD. Analysis of relative gene ex-pression data using real-time quantitative PCR and the2-[Delta][Delta]CT method. Methods 2001;25:402e8.
38. Amiel D, Ishizue KK, Harwood FL, Kitabayashi L,Akeson WH. Injury of the anterior cruciate ligament:the role of collagenase in ligament degeneration.J Orthop Res 1989;7:486e93.
39. Hayashi K, Frank JD, Dubinsky C, Hao Z, Markel MD,Manley PA, et al. Histologic changes in rupturedcanine cranial cruciate ligament. Vet Surg 2003;32:269e77.
40. Feagin JA Jr, Curl WW. Isolated tear of the anterior cru-ciate ligament: 5-year follow-up study. Am J SportsMed 1976;4:95e100.
41. Comerford EJ, Tarlton JF, Avery NC, Bailey AJ, InnesJF. Distal femoral intercondylar notch dimensionsand their relationship to composition and metabolismof the canine anterior cruciate ligament. OsteoarthritisCartilage 13;14:273e8.
42. Tremble PM, Lane TF, Sage EH, Werb Z. SPARC, asecreted protein associated with morphogenesis andtissue remodeling, induces expression of metallopro-teinases in fibroblasts through a novel extracellularmatrix-dependent pathway. J Cell Biol 1993;121:1433e44.
43. Everts V, van der ZE, Creemers L, Beertsen W. Phago-cytosis and intracellular digestion of collagen, its rolein turnover and remodelling. Histochem J 1996;28:229e45.
44. Yamaguchi M, Ozawa Y, Nogimura A, Aihara N,Kojima T, Hirayama Y, et al. Cathepsins B and Lincreased during response of periodontal ligamentcells to mechanical stress in vitro. Connect TissueRes 2004;45:181e9.
45. Goldberg VM, Burstein A, Dawson M. The influence ofan experimental immune synovitis on the failure modeand strength of the rabbit anterior cruciate ligament.J Bone Joint Surg Am 1982;64:900e6.
46. Stoker A, Cook J, Kuroki K, Fox D. Site-specific anal-ysis of gene expression in early osteoarthritis usingthe Pond-Nuki model in dogs. J Orthop Surg Res2006;1:8.
47. Salinardi BJ, Roush JK, Schermerhorn T,Mitchell KE. Matrix metalloproteinase and tissue in-hibitor of metalloproteinase in serum and synovialfluid of osteoarthritic dogs. Vet Comp Orth Trauma2006;19:49e55.
48. Hart DA, Sciore P, Boykiw R, Reno C. Pregnancy in-duces complex changes in the pattern of mRNA ex-pression in knee ligaments of the adolescent rabbit.Matrix Biol 1998;17:21e34.
49. Sarasa-Renedo A, Chiquet M. Mechanical signals reg-ulating extracellular matrix gene expression in fibro-blasts. Scand J Med Sci Sports 2005;15:223e30.
50. Molloy T, Wang Y, Murrell G. The roles of growth fac-tors in tendon and ligament healing. Sports Med2003;33:381e94.
51. Ireland ML. The female ACL: why is it more prone to in-jury? Orthop Clin North Am 2002;33:637e51.
52. Slauterbeck JR, Pankratz K, Xu KT, Bozeman SC,Hardy DM. Canine ovariohysterectomy and orchiec-tomy increases the prevalence of ACL injury. ClinOrthop Relat Res 2004;301e5.
53. Muir P, Schamberger GM, Manley PA, Hao Z. Localiza-tion of cathepsin k and tartrate-resistant acid phospha-tase in synovium and cranial cruciate ligament in dogswith cruciate disease. Vet Surg 2005;34:239e46.
Related Documents