Original Contribution GENE EXPRESSION PROFILE OF OXIDANT STRESS AND NEURODEGENERATION IN TRANSGENIC MICE DEFICIENT IN -TOCOPHEROL TRANSFER PROTEIN KISHORCHANDRA GOHIL,* BETTINA C. SCHOCK,* ABHISHEK A. CHAKRABORTY, † YUKO TERASAWA, ‡ JACOB RABER, ‡ ROBERT V. FARESE,JR., ‡ LESTER PACKER, § CARROLL E. CROSS,* and MARET G. TRABER *Center for Comparative Respiratory and Medicine, Department of Internal Medicine, University of California, Davis, CA, USA; † Department of Biochemistry, Maharaja Sayajirao University, Sayajigunj, Baroda, India; ‡ Cardiovascular Research Institute and Department of Medicine, University of California, San Franscisco, CA, USA; § Department of Molecular Pharmacology and Toxicology, University of Southern California, Los Angeles, CA, USA; and Linus Pauling Institute, Department of Nutrition and Food Management, Oregon State University, Corvallis, OR, USA (Received 14 April 2003; Revised 1 July 2003; Accepted 17 July 2003) Abstract—Alpha-tocopherol transfer protein (TTP) regulates the retention and secretion of -tocopherol (-T) by the liver. Deletion of the TTP gene (Ttpa) in mice results in systemic deficiency of -T and neurological dysfunctions described in patients with mutated Ttpa. We have explored genome-wide changes in mRNAs from brain cortex and liver of Ttpa-deficient (Ttpa / ) mice and wild-type (Ttpa / ) mice. Selective inductions of genes regulated by antioxidant response elements were detected in Ttpa / livers compared to Ttpa / livers, suggesting increased oxidant stress in Ttpa / livers. The activation of cell proliferation pathways in Ttpa / livers was indicated by the induction of genes that encode growth factor-binding proteins, mitogen-activated protein kinase kinase 3, and apoptosis inhibitor 6. The induction of synuclein- and repression of synuclein- genes was detected in Ttpa / cortex. This may predispose Ttpa / cortex to increased formation of synuclein- aggregates and Lewy body, often associated with oxidant stress. Cortex of Ttpa / mice revealed repression of genes encoding synaptic proteins, protein kinase C family members, and myelin proteins. A 13-fold decrease in the expression of retinoic acid receptor-related orphan receptor- mRNA predicts staggerer-like phenotype (ataxia and deficits of motor coordination) of Ttpa / mice. The repression of specific genes that determine synaptic plasticity and neuronal development may account for suppressed electrophysiological activities of cortex and impaired behavior in Ttpa / mice. © 2003 Elsevier Inc. Keywords—Long-term potentiation, Anxiety, Vitamin E, Antioxidant, Oligonucleotide arrays, Neurodegeneration, Synapse, Free radicals INTRODUCTION Vitamin E was discovered as a dietary substance essen- tial for normal reproduction in rats [1], and, subse- quently, its molecular structure as -tocopherol was elu- cidated. The precise mechanism of -T action in the prevention of fetal resorption or its potential actions in adults is unknown. However, numerous studies suggest that its activity as a scavenger of reactive metabolites of oxygen and as an inhibitor of lipid peroxidation in mem- branes may play important roles in its physiological functions in fetal implantation [2] and during postfetal life of mammals [3]. Nonantioxidant roles of -T have also been reported [4,5] and suggest that biological ac- tions of vitamin E (VE) are mediated through diverse molecular mechanisms that may include its actions as a regulator of gene transcription. Physiological concentrations of -T are primarily de- termined by the -tocopherol transfer protein (TTP) that preferentially binds to -T, compared to more abundant, naturally occurring isomers of VE with similar antioxi- dant properties [6]. TTP is a member of the SEC14 family of proteins that bind to specific lipids, such as -T to TTP [7–9], phosphatidyl inositol to SEC14 [10], retinaldehyde to retinaldehyde-binding protein, and Address correspondence to: Dr. Kishorchandra Gohil, University of California, Department of Internal Medicine, Center for Comparative Respiratory and Medicine, Surge 1, Room 1144, Davis, CA 95616, USA; Tel: (530) 752-0674; Fax: (530) 752-4374; E-Mail: [email protected]. Free Radical Biology & Medicine, Vol. 35, No. 11, pp. 1343–1354, 2003 Copyright © 2003 Elsevier Inc. Printed in the USA. All rights reserved 0891-5849/03/$–see front matter doi:10.1016/S0891-5849(03)00509-4 1343

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Original Contribution
GENE EXPRESSION PROFILE OF OXIDANT STRESS ANDNEURODEGENERATION IN TRANSGENIC MICE DEFICIENT IN
�-TOCOPHEROL TRANSFER PROTEIN
KISHORCHANDRA GOHIL,* BETTINA C. SCHOCK,* A BHISHEK A. CHAKRABORTY,† YUKO TERASAWA,‡
JACOB RABER,‡ ROBERT V. FARESE, JR.,‡ LESTER PACKER,§ CARROLL E. CROSS,* and MARET G. TRABER�
*Center for Comparative Respiratory and Medicine, Department of Internal Medicine, University of California, Davis, CA, USA;†Department of Biochemistry, Maharaja Sayajirao University, Sayajigunj, Baroda, India;‡Cardiovascular Research Institute and
Department of Medicine, University of California, San Franscisco, CA, USA;§Department of Molecular Pharmacology andToxicology, University of Southern California, Los Angeles, CA, USA; and�Linus Pauling Institute, Department of Nutrition and
Food Management, Oregon State University, Corvallis, OR, USA
(Received 14 April 2003;Revised 1 July 2003;Accepted 17 July 2003)
Abstract—Alpha-tocopherol transfer protein (TTP) regulates the retention and secretion of�-tocopherol (�-T) by theliver. Deletion of the TTP gene (Ttpa) in mice results in systemic deficiency of�-T and neurological dysfunctionsdescribed in patients with mutatedTtpa. We have explored genome-wide changes in mRNAs from brain cortex and liverof Ttpa-deficient (Ttpa�/�) mice and wild-type (Ttpa�/�) mice. Selective inductions of genes regulated by antioxidantresponse elements were detected inTtpa�/� livers compared toTtpa�/� livers, suggesting increased oxidant stress inTtpa�/� livers. The activation of cell proliferation pathways inTtpa�/� livers was indicated by the induction of genesthat encode growth factor-binding proteins, mitogen-activated protein kinase kinase 3, and apoptosis inhibitor 6. Theinduction of synuclein-� and repression of synuclein-� genes was detected inTtpa�/� cortex. This may predisposeTtpa�/� cortex to increased formation of synuclein-� aggregates and Lewy body, often associated with oxidant stress.Cortex ofTtpa�/� mice revealed repression of genes encoding synaptic proteins, protein kinase C family members, andmyelin proteins. A 13-fold decrease in the expression of retinoic acid receptor-related orphan receptor-� mRNA predictsstaggerer-like phenotype (ataxia and deficits of motor coordination) ofTtpa�/� mice. The repression of specific genesthat determine synaptic plasticity and neuronal development may account for suppressed electrophysiological activitiesof cortex and impaired behavior inTtpa�/� mice. © 2003 Elsevier Inc.
Keywords—Long-term potentiation, Anxiety, Vitamin E, Antioxidant, Oligonucleotide arrays, Neurodegeneration,Synapse, Free radicals
INTRODUCTION
Vitamin E was discovered as a dietary substance essen-tial for normal reproduction in rats [1], and, subse-quently, its molecular structure as�-tocopherol was elu-cidated. The precise mechanism of�-T action in theprevention of fetal resorption or its potential actions inadults is unknown. However, numerous studies suggestthat its activity as a scavenger of reactive metabolites ofoxygen and as an inhibitor of lipid peroxidation in mem-
branes may play important roles in its physiologicalfunctions in fetal implantation [2] and during postfetallife of mammals [3]. Nonantioxidant roles of�-T havealso been reported [4,5] and suggest that biological ac-tions of vitamin E (VE) are mediated through diversemolecular mechanisms that may include its actions as aregulator of gene transcription.
Physiological concentrations of�-T are primarily de-termined by the�-tocopherol transfer protein (TTP) thatpreferentially binds to�-T, compared to more abundant,naturally occurring isomers of VE with similar antioxi-dant properties [6]. TTP is a member of the SEC14family of proteins that bind to specific lipids, such as�-Tto TTP [7–9], phosphatidyl inositol to SEC14 [10],retinaldehyde to retinaldehyde-binding protein, and
Address correspondence to: Dr. Kishorchandra Gohil, University ofCalifornia, Department of Internal Medicine, Center for ComparativeRespiratory and Medicine, Surge 1, Room 1144, Davis, CA 95616,USA; Tel: (530) 752-0674; Fax: (530) 752-4374; E-Mail:[email protected].
Free Radical Biology & Medicine, Vol. 35, No. 11, pp. 1343–1354, 2003Copyright © 2003 Elsevier Inc.
Printed in the USA. All rights reserved0891-5849/03/$–see front matter
doi:10.1016/S0891-5849(03)00509-4
1343

squalene to supernatant factor [11,12]. The conservationof structural and functional properties of the SEC14family suggests that the proteins play an important rolein the assembly and functions of biological membranesand in secretory processes.
A need for �-T in postnatal humans remained contro-versial for decades because of the lack of a well-defineddeficiency disease [6]. Discovery of vitamin E deficiencyin patients with neurological diseases [13] in the past 10years has rekindled the interest in the importance of �-Tin humans. This group of patients showed mutations inthe TTP gene, demonstrating an essential role of TTP indetermining systemic concentrations of �-T and main-taining normal architecture of the nervous system andneurological functions. Transgenic mice rendered defi-cient in TTP gene lack TTP and have systemic deficiencyof �-T [14] that is attributed to failure in the secretion of�-T from the liver [6]. Ttpa�/� mice show many featuresof neurological disease seen in humans [15]. A commonfeature of the Ttpa�/� mice and human diseases associ-ated with �-T deficiency is the delayed onset of thedisease phenotype. This may be attributed to a gradualloss of neurons and neuronal functions caused bychanges in the expression of the genome imposed by thedeficiency of the TTP or �-T.
We hypothesized that the systemic deficiency of �-Tinflicted by the deletion of TTP gene would alter thetissue-specific transcriptional program through the po-tential actions of TTP and �-T on gene expression. Thishypothesis was tested in livers and brain cortex fromyoung Ttpa normal and Ttpa�/� mice using high-densityoligonucleotide arrays to identify genome-wide changesin the expression of mRNAs. These gene-profiling stud-ies in young mice describe cumulative effects of the TTPgene deletion and the consequent changes in vitamin Estatus that must have occurred during development invivo. The description of mRNA expression changes maybe relevant to the effects of chronic deficiency of vitaminE seen in patients with mutations in TTP gene. Thechanges in the behavior parameters such as performancein elevated water maze and exploratory behaviors wereanalyzed to correlate the changes in transcriptional pro-files with the changes in the biochemical and behavioralphenotypes. The DNA sequences of a selected group ofbrain cortex genes downregulated in Ttpa�/� mice wereanalyzed for transcription factor-binding sites.
MATERIALS AND METHODS
Male and female C57BL6J mice with a deletion in theTtpa gene (Ttpa�/�) and wild-type mice (Ttpa�/�) fromour colonies were used. The colony of Ttpa�/� mice inC57BL6/J genetic background was obtained by back-crossing the original colony [14] of mixed (50%
C57BL6/J and 50% 129/SvJae) mice heterozygous forthe deletion (Ttpa�/�) with C57BL6/J (Ttpa�/�) mice.The offsprings were genotyped using specific primers forTtpa, as described previously [14]. Additionally, thegenotype of each animal was confirmed by Western blotanalysis of TTP protein in liver using an anti-TTP anti-serum, as described previously [14]. The concentrationsof �-T in plasma, liver, and cortex were also determined[16] for confirmation of the predicted �-T status of thetwo distinct groups of mice.
Neurological behavior analysis
Examination of balance, coordination, strength, ex-ploratory behavior, and anxiety were performed in mice(12–16 weeks old) from our original colony (50%C57BL6J and 50% 129SvJae, n � 8–15), as describedpreviously [17]. To minimize the influences on behaviorbetween mice, mice were housed individually for 7 dbefore behavior tests under conditions of constant tem-perature (18°C), light (6:00 a.m. to 6:00 p.m.), and withaccess to food and water ad libitum. The apparatus wascleaned with 1 mM acetic acid to remove the animalodors before and after each test.
Open-field activity for assessment of exploratory behav-ior. Mice were placed individually into brightly lit auto-mated cages equipped with rows of infrared photocellsinterfaced with a computer (San Diego Instruments, SanDiego, CA, USA). After 1 min adaptation period, open-field activity was recorded for 10 min. Recorded beambreaks were used to calculate active times, path lengths,rearing times, and rearing events.
Elevated plus maze. Anxiety levels were assessed withan elevated, plus-shaped maze consisting of two openarms and two closed arms equipped with rows of infraredphotocells interfaced with a computer (Hamilton, Poway,CA, USA). Mice were placed individually in the centerof the maze and allowed free access to the arms for 10min. They could spend their time either in a closed safearea (closed arms) or in an open area (open arms).Recorded beam brakes were used to calculate the timespent, the distance moved, and entries in the closed andopen arms.
Rotorod. Motor performance was assessed with a rotorodtest (San Diego Instruments). After a 1 min adaptationperiod on the rod at rest, the rod was accelerated by 5revolutions/min every 15 s and the length of time miceremained on the rod (fall latency) was recorded.
Statistical analysis. Behavioral data are expressed asmean � SEM. The statistical significance of differencesbetween groups was determined by ANOVA followed
1344 K. GOHIL et al.

by Tukey-Kramer test when appropriate; p � .05 wasconsidered significant. Appropriate nonparametric testswere used when the data was not normally distributed.
RNA extraction and preparation of biotin-labeled RNAfor GeneChip analysis
These procedures were performed essentially as de-scribed previously [18]. Specifically, total RNA wasextracted from about 100 mg of liver or cortex from eachmouse (12–16 weeks old) by homogenization of thetissue in Trizol Reagent and purification and quantitationof RNA according to the manufacturer’s (Invitrogen)protocol. An equal aliquot (10 �g) of total RNA extractin RNAase- and DNAase-free water from each tissuefrom the group of mice (n � 4–6) was combined. Analiquot (20 �g) of pooled RNA solution was used forpreparation of biotin-labeled RNA for hybridization tohigh-density oligonucleotide arrays (Mu74Av2), as de-scribed in the Affymetrix (Santa Clara, CA, USA) pro-tocol for sample preparation [18]. Biotin-labeled RNAsamples were hybridized to murine GeneChips. Thescanned images of hybridization signals were analyzedwith the Affymetrix Microarray Analysis Suite (MAS)5.0.
Data analysis
Absolute mRNA expression (present or absent) anddifferential (Ttpa�/� vs. Ttpa�/�) mRNA expressionanalysis were obtained using the MAS 5.0 software.When the p value for the detection signal was � .039 (p� .0002–.039), the expression of the mRNA was clas-sified as present. All mRNAs with the p value for detec-tion � .039 were considered absent. The signal intensi-ties for transcripts classified as present ranged from5–7000 units.
To obtain the list of TTP-sensitive genes, the entiredata set from the two groups of mice was subjected to“batch analysis” that generated data with fold change foreach mRNA. The data that showed either an increase ora decrease of more than 2-fold were selected as TTP-sensitive genes. All other changes were excluded fromfurther analysis.
Results and discussions focus primarily on differ-entially (� 2-fold) expressed mRNAs that were de-tected with a high level of confidence (p � .039) in atleast one set of the pair of data used for differentialanalysis in liver or brain cortex. This conservativeanalysis excluded a large number of genes that wereidentified as significantly different between Ttpa�/�
and Ttpa�/� mice but whose detection was of lowconfidence (p � .039).
Validation of GeneChip data
We [18,19] and others [20,21] have shown that thechanges in the expression of mRNAs selected by theconservative analysis of hybridization data, as describedabove, could be confirmed by independent analysis suchas Northern Blot, polymerase chain reaction (PCR), real-time PCR, and in some cases by immunoblot analysis ofthe encoded proteins [18]. Therefore, in this study, fur-ther analysis of the changes in the expression of mRNAsdescribed below was not performed.
The expression data and their interpretations werevalidated by comparison of the specificity of the expres-sion levels of selected genes whose expression and ac-tivities of the encoded proteins are known to be tissuespecific.
Search for common DNA elements and transcriptionfactors for a selected group of differentially expressedgenes
The genome database was mined to identify conservedfeatures in the genes encoding 7 mRNAs that were detectedin the cortex with a high level of confidence. The analysisfor the presence of transcription factor-binding sites wasperformed on sequences upstream (up to 10 Kb) of thecoding sequence. The complete mRNA sequences of thegenes were obtained from GenBank at the National Centerfor Biotechnology Information (http://www.ncbi.nlm.nih-.gov) and aligned against the complete gene sequence onthe Mus musculus genome using BLAST tools at the Na-tional Center for Biotechnology Information. The completegenomic sequence was then located on the respective chro-mosome and the sequence of the complete SuperContig’swas retrieved and searched for the promoter elements usingthe Neural Network Promoter Prediction software at theBerkeley Drosophila Genome Program site (http://www-.fruitfly.org/seq_tools/promoter.html). Upstream sequence(up to 10 Kb) of the promoter site between the obtainedsequence and the original mRNA was confirmed by usingthe nucleotide-nucleotide BLAST tool. This confirmed se-quence was used to search for transcription factor-bindingsites using the MatInspector V2.2 software that uses matri-ces characterized by the TransFac 4.0 database (http://trans-fac.gbf.de/cgi-bin/matSearch/matsearch.pl) [22]. The datawas tabulated and analyzed for clustering of binding sites ofany particular transcription factor in the upstream regions ofthe seven selected genes.
RESULTS AND DISCUSSION
Systemic deficiency of �-T and TTP in Ttpa�/� mice
Plasma concentrations of �-T, expressed as mean �SD, for C57BL6/J mice (n � 4–6) were 4.5 � 0.8 �Mand 0.3 � 0.03 �M in Ttpa�/� and Ttpa�/� mice,
1345Gene expression profile of TTP-null mice
respectively. Liver �-T concentrations were 13.0 � 4.0nmoles/g wet weight and 6.1 � 2.2 nmoles/g wet weightin Ttpa�/� and Ttpa�/� mice, respectively. Cortex �-Tconcentrations were 24 � 2.5 nmoles/g wet weight and0.25 � 0.05 nmoles/g wet weight in Ttpa�/� andTtpa�/� mice, respectively. These data confirmed pre-dicted status of �-T in Ttpa�/� mice of C57BL6/J back-ground. Western blot analysis of TTP with anti-TTPantiserum showed an abundance of the 32 kD protein inlivers of Ttpa�/� mice but undetectable to trace amountsin Ttpa�/� mice (data not shown). These observationsconfirmed that our current colony of Ttpa�/� mice ofC57BL6J background is very similar, if not identical, toour original colony of Ttpa�/� (50% C57BL6/J and 50%129SvJae) mice [14] with respect to biochemical pheno-types. Mice (12–16 weeks old) from our current colonywere used for gene expression analysis.
Validation of analysis and interpretations of geneexpression profiles obtained by high-densityoligonucleotide arrays
Murine GeneChips detected �2000 mRNAs, withhigh statistical confidence (p � .0002–.039), in liver andcortex. All the mRNAs with the detection p � .039 wereconsidered undetectable. The mRNA expression profilesfrom the two tissues were interrogated with the knownactivities of liver- and cortex-specific proteins to estab-
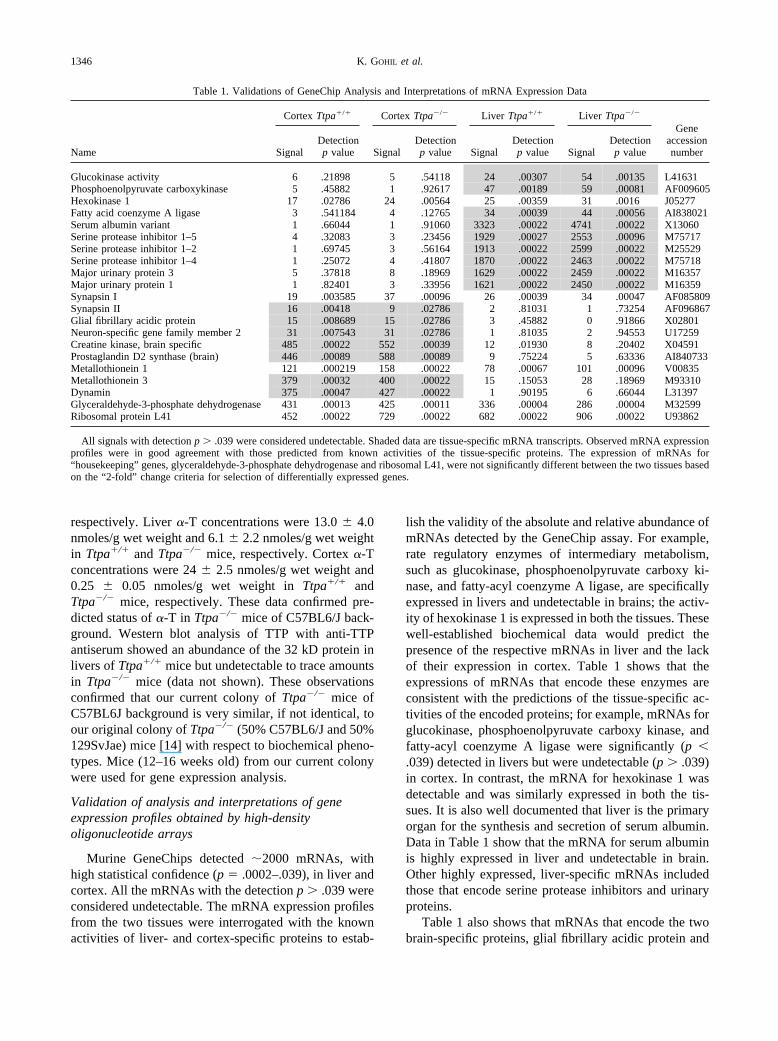
lish the validity of the absolute and relative abundance ofmRNAs detected by the GeneChip assay. For example,rate regulatory enzymes of intermediary metabolism,such as glucokinase, phosphoenolpyruvate carboxy ki-nase, and fatty-acyl coenzyme A ligase, are specificallyexpressed in livers and undetectable in brains; the activ-ity of hexokinase 1 is expressed in both the tissues. Thesewell-established biochemical data would predict thepresence of the respective mRNAs in liver and the lackof their expression in cortex. Table 1 shows that theexpressions of mRNAs that encode these enzymes areconsistent with the predictions of the tissue-specific ac-tivities of the encoded proteins; for example, mRNAs forglucokinase, phosphoenolpyruvate carboxy kinase, andfatty-acyl coenzyme A ligase were significantly (p �.039) detected in livers but were undetectable (p � .039)in cortex. In contrast, the mRNA for hexokinase 1 wasdetectable and was similarly expressed in both the tis-sues. It is also well documented that liver is the primaryorgan for the synthesis and secretion of serum albumin.Data in Table 1 show that the mRNA for serum albuminis highly expressed in liver and undetectable in brain.Other highly expressed, liver-specific mRNAs includedthose that encode serine protease inhibitors and urinaryproteins.
Table 1 also shows that mRNAs that encode the twobrain-specific proteins, glial fibrillary acidic protein and
Table 1. Validations of GeneChip Analysis and Interpretations of mRNA Expression Data
Name
Cortex Ttpa�/� Cortex Ttpa�/� Liver Ttpa�/� Liver Ttpa�/�
GeneaccessionnumberSignal
Detectionp value Signal
Detectionp value Signal
Detectionp value Signal
Detectionp value
Glucokinase activity 6 .21898 5 .54118 24 .00307 54 .00135 L41631Phosphoenolpyruvate carboxykinase 5 .45882 1 .92617 47 .00189 59 .00081 AF009605Hexokinase 1 17 .02786 24 .00564 25 .00359 31 .0016 J05277Fatty acid coenzyme A ligase 3 .541184 4 .12765 34 .00039 44 .00056 AI838021Serum albumin variant 1 .66044 1 .91060 3323 .00022 4741 .00022 X13060Serine protease inhibitor 1–5 4 .32083 3 .23456 1929 .00027 2553 .00096 M75717Serine protease inhibitor 1–2 1 .69745 3 .56164 1913 .00022 2599 .00022 M25529Serine protease inhibitor 1–4 1 .25072 4 .41807 1870 .00022 2463 .00022 M75718Major urinary protein 3 5 .37818 8 .18969 1629 .00022 2459 .00022 M16357Major urinary protein 1 1 .82401 3 .33956 1621 .00022 2450 .00022 M16359Synapsin I 19 .003585 37 .00096 26 .00039 34 .00047 AF085809Synapsin II 16 .00418 9 .02786 2 .81031 1 .73254 AF096867Glial fibrillary acidic protein 15 .008689 15 .02786 3 .45882 0 .91866 X02801Neuron-specific gene family member 2 31 .007543 31 .02786 1 .81035 2 .94553 U17259Creatine kinase, brain specific 485 .00022 552 .00039 12 .01930 8 .20402 X04591Prostaglandin D2 synthase (brain) 446 .00089 588 .00089 9 .75224 5 .63336 AI840733Metallothionein 1 121 .000219 158 .00022 78 .00067 101 .00096 V00835Metallothionein 3 379 .00032 400 .00022 15 .15053 28 .18969 M93310Dynamin 375 .00047 427 .00022 1 .90195 6 .66044 L31397Glyceraldehyde-3-phosphate dehydrogenase 431 .00013 425 .00011 336 .00004 286 .00004 M32599Ribosomal protein L41 452 .00022 729 .00022 682 .00022 906 .00022 U93862
All signals with detection p � .039 were considered undetectable. Shaded data are tissue-specific mRNA transcripts. Observed mRNA expressionprofiles were in good agreement with those predicted from known activities of the tissue-specific proteins. The expression of mRNAs for“housekeeping” genes, glyceraldehyde-3-phosphate dehydrogenase and ribosomal L41, were not significantly different between the two tissues basedon the “2-fold” change criteria for selection of differentially expressed genes.
1346 K. GOHIL et al.

neuron-specific protein, were detected in brain cortexand were undetectable in livers, providing further vali-dation of the GeneChip analysis. Also, the signal forbrain-specific metallothionein III was high in cortex andundetectable in liver; this is in good agreement withNorthern blot analysis of metallothionein III mRNAs inlivers and brain from C57BL6 mice [23]. A similarcomparison of mRNA profile of lungs with livers fromC57BL6 mice identified tissue-specific differences in theexpression profiles [24]. The data in Table 1 also showthat the assay is reliable over a wide dynamic range in thevariation of mRNA abundance; low abundance mRNAs,such as those for glucokinase and synapsins, and highabundance mRNAs, such as those for serum albumin andbrain-specific creatine kinase, were detected with a sim-ilarly high statistical confidence (p � .009). Equallynoteworthy are the similarities in the absolute expressionsignals for each of the mRNAs from the two geneticallydifferent groups of mice.
The GeneChip assay could reliably and predictablydistinguish between the two tissues and showed that theexpressions of most of the genes in liver (98%) and incortex (90%) were statistically indistinguishable fromthe two samples (n � 5–6) of genetically different
(Ttpa�/� vs. Ttpa�/�) populations of mice. The follow-ing paragraphs describe the genes whose expressionswere affected by the deletion of the TTP gene.
The effect of TTP gene deletion on expressions of livermRNAs
Differential analysis of �2000 liver mRNAs identi-fied 48 TTP/�-T-sensitive transcripts; the expressions of9 mRNAs were decreased and 39 mRNAs were in-creased. Table 2 shows a list of selected transcripts thatwere differentially expressed in livers of Ttpa�/� mice.
A strong indication for a state of neovascularization inlivers of Ttpa�/� mice was revealed by reciprocal regu-lation of two mRNAs that encode proteins essential inblood vessel formation. A 10-fold increase in mRNA forquaking and a 14-fold decrease in mRNA for NMDAreceptor-regulated gene 1 that encodes Tubedown-1were detected (Table 2). The former is essential in bloodvessel development [25] and the latter inhibits capillaryformation [26]. We predict that the expression of thesetranscripts is predominantly in smooth muscle and endo-thelial cells. This prediction is supported by a 20-foldinduction in the transcript for hyaluronidase 1 (Table 2),
Table 2. TTP- and �-T–Sensitive Genes in Liver: Indicators of Vasculogenesis and Cell Proliferation
Foldchange Name Function(s)
Gene accessionnumber
�13.9 NMDA receptor–regulated gene 1 Inhibits blood vessel development AI645561�3.7 (Asp-Glu-Ala-Asp/His) box poly peptide 19 ATP-dependent helicase L25125
2.0 Peroxisome proliferator–activated receptor � Nuclear receptors X576382.0 Nucleoside diphosphate kinase Synthesis of nucleoside triphosphates AV2164682.1 Inter-� trypsin inhibitor, heavy chain 3 Inhibitor of hyaluronidases X703932.1 Insulin-like growth factor–binding protein 2 Stimulator of cell growth and proliferation X815802.1 Aplysia ras-related homolog 9 (RhoC) Regulator of cell growth and proliferation AV0645022.1 Kruppel-like factor 9 Regulation of transcription AI8480502.1 Ornithine decarboxylase antizyme inhibitor Increases polyamine biosynthesis AF0321282.1 RAB9, member RAS oncogene family Regulator of cell growth and proliferation AB0272902.3 Cytochromes P450 and 2d9 Xenobiotic metabolism AV0038302.5 Metallothionein II Induced by oxidant stress K022362.5 Proteasome-related subunit Cell cycle regulator AF0713152.5 Proteasome 26S subunit, non-ATPase, 13 Cell cycle regulator AW0454512.5 Deiodinase, iodothyronine, type I Increases thyroxine T3, ligand for nuclear receptors U498612.8 Mitogen-activated protein kinase kinase 3 Regulates stress-induced cell proliferation AI8526363.0 Cytochrome, P450 and 1a2, aromatic compound inducible Xenobiotic metabolism X042833.2 Ubiquitin D Regulates cell cycle and proliferation AL0786303.7 Glutathione S-transferase, � 2 Stress response and cell proliferation X534513.7 Apoptosis inhibitory 6 Regulates progression of cell cycle AF0114284.0 Zinc fingers and homeoboxes protein 1 Transcription factor AV3260824.3 Initiation factor elF4-� Translation initiation factor AV3807939.8 Serine/threonine kinase 16 Differentiation and proliferation AF062076
10.6 Quaking Essential for blood vessel development U4494011.3 G-protein � WD 40 repeat Transduction of G-protein–mediated signalling AI85037812.1 Growth factor receptor–bound protein 10 Interacts with growth factor receptors AF02207219.7 Hyaluronidase 1 Regulates fluidity of extracellular matrix of proliferating
cellsAF011567
A selected list of differentially expressed mRNAs in livers of TTP-deficient and normal mice. Reciprocal relationship in the expression of twomRNAs that encode proteins of blood vessels (shown in bold) suggest a state of blood vessel cell proliferation in livers of TTP- and �-T–deficientmice.
1347Gene expression profile of TTP-null mice

because high activity of hyaluronidase is also associatedwith smooth muscle cell proliferation in response toinjury [27]. These processes are likely to be driven bymitogenic growth factors as suggested by the increase inthe transcripts for growth factor receptor adaptor protein(12-fold), mitogen-activated protein kinase kinase 3 (3-fold), and serine/threonine kinase 16 (10-fold). A previ-ous study that used cDNA arrays showed induction ofmRNA for a transducer of MAP kinase pathway andsuppression of genes that are proapoptotic in livers ofrats deficient in vitamin E and selenium [28]. Thesechanges were reversed when the diets of rats were sup-plemented with either vitamin E or selenium.
An indication for an increased oxidant stress in livers ofTtpa�/� mice imposed by decreased concentration of �-T,was suggested by the induction of transcripts that are reg-ulated by antioxidant response elements (ARE) or electro-phile responsive elements (EpRE) that define conservednucleotide sequences on the coding DNA strand to whichredox-sensitive transcription factors bind and activate genetranscription [29]. These included mRNAs for metallothio-nein 2 (2.5-fold), glutathione-S-transferase � (3.7-fold), andcytochromes P450, 1a2, and 2d9 (2- to 3-fold) [30].
Cytochrome P450s belong to a superfamily of hemeproteins that play an important role in detoxification ofxenobiotics and are components of phase I response [31].The expression of glutathione transferase family mem-bers are differentially regulated by reactive intermediates[32,33]. The induction of metallothionein I mRNA (3.6-fold) and the protein in livers of rats deficient in vitaminE and selenium, and reversal of this by dietary supple-mentation with vitamin E, has been reported previously[28]. Numerous studies have suggested that high activi-ties of metallothioneins and glutathione-S-transferase �are detected in proliferating and drug-resistant cells [34],and induction of these transcripts in livers of Ttpa-defi-cient mice suggests a state of oxidant-induced cellularproliferation in TTP and �-T-deficient livers.
The gene-profiling data were notable for the lack ofinduction in the genes for the primary antioxidant enzymescatalase, peroxidase, and superoxide dismutase in TTP-deficient livers. A previous study also reported a lack ofincrease in the activities of these primary antioxidant en-zymes in the presence of increased markers of oxidantstress in liver in vivo [35]. These observations suggestdifferential sensitivity of transcriptional processes thattransduce the changes in the vitamin E concentrations andits antioxidant and nonantioxidant effects.
Gene expression profiles of brain cortex from Ttpa�/�
mice
GeneChip analysis of cortex RNA detected �2000transcripts with high statistical confidence (p � .039).
This was comparable to that observed for liver. How-ever, unlike that in liver, 130 transcripts were differen-tially expressed between cortex from Ttpa�/� andTtpa�/� mice, showing increased sensitivity of braincortex genome compared to that of liver to the TTP/�-Tdeficiency. Another notable difference was in the ratio ofrepressed to induced gene expression; 90 genes wererepressed and 50 were induced in cortex of Ttpa�/�
mice. These data reveal distinct effects of TTP/�-T onthe same genome in metabolically and functionally dif-ferent organs. The effects of dietary vitamin E on theexpression of multiple genes has been demonstrated infetal rat brains [36]. It is likely that the differentialmRNA expression profile of cortex is primarily attrib-uted to differences in the �-tocopherol content of thecortex and not the expression of TTP and its mRNA,because the latter are almost undetectable in Ttpa�/�
cortex [6]. This is in contrast to the liver in which boththe TTP and its mRNA are highly expressed in Ttpa�/�
mice, raising the possibility that TTP, in addition to �-T,may affect the transcription of selected liver genes; thedeletion of Ttpa obliterates the expression of TTP but notthe liver �-tocopherol content.
Indicators of inhibition of neurogenesis andneurodegeneration in Ttpa�/� cortex
A 13-fold decrease in the abundance for mRNA en-coding retinoic acid receptor-related orphan receptor-(ROR-) � is an indictor of inhibited neurogenesis in thecortex of Ttpa�/� mice (Table 3). ROR-�-null mice aredeficient in differentiation and maturation of specificneurons [37] and deficient in numerous behaviors includ-ing impairment to react to novelty [38]. Behavioral anal-ysis of Ttpa�/� mice also indicates such impairments(see Figs. 2 and 3). A similar decrease in the expressionof hairless mRNA, which encodes a protein that interactswith thyroid hormone receptors [39], was detected inTtpa�/� cortex (Table 3). Mice deficient in hairlessmRNA show increased indices of aging in cerebellum[40]. The transcription of hairless gene is induced bythyroid hormone [39], and the repression of hairlessmRNA in Ttpa�/� cortex detected in this study suggeststhat �-T may be a cofactor for normal responses tothyroid hormone in cortex.
A possible interaction between TTP/�-T and thyroidhormone status is further indicated by a coordinatedrepression of genes encoding proteins essential for my-elination, a process that is also inhibited in hypothyroidbrain [39]. Table 3 shows 2- to 5-fold decreases inmRNAs for proteolipid protein, sialytransferases, andmyelin-associated oligodendrocytic basic protein inTtpa�/� mice. It is also noteworthy that the transcript forcontactin-associated protein 1 was decreased by 10-fold;
1348 K. GOHIL et al.

this protein has been suggested to be essential for inter-actions between axons and myelin forming oligodendro-cytes at nodes of Ranvier in myelinated fibers [41].
Collectively, these observations would predict the in-hibition of myelination in Ttpa�/� mice. This predictionis supported by histochemical analysis that showed adecrease in myelinated fibers in 1 year old Ttpa�/� micebut not in younger Ttpa�/� mice [15]. The data presentedhere were obtained from young mice (3 to 4 months old)and suggest that the molecular processes of neurodegen-eration are initiated long before the histochemical mark-ers of neurodegeneration are detectable in Ttpa�/� mice.
Reciprocal changes in the expressions of synuclein-�and synuclein-� genes (Table 3) provide additional sup-port for a neurodegenerative state of Ttpa�/� cortex. Theabsolute expression of mRNAs for synuclein-� was �20fold � synuclein-� in Ttpa�/� mice. Synuclein-� is apresynaptic protein and is a major fibrillar component ofLewy bodies [42,43]. Oxidative stress is one of severalfactors that are associated with the formation of Lewybodies [44]. It has been suggested that oxidative eventsmay also stimulate precipitation of pre-existingsynuclein. Aggregation of synuclein-� is inhibited bysynuclein-� in vitro [45] and in vivo [46]. Also, overex-pression of �-synuclein increases intracellular reactiveoxygen species [47].
Differential mRNA expression analysis in our studyshowed a 3-fold decrease in synuclein-� and a 2-foldincrease in synuclein-� mRNAs in Ttpa�/� mice, sug-gesting a neuronal state predisposed to increased levelsof potentially neurotoxic aggregates of synuclein-�. A13-fold increase in cytochrome c (Table 3), a coactivatorof “executioner” caspases 3, 6, and 7, suggests increasedapoptosis [48] in Ttpa�/� cortex and is another indica-tion of neurodegeneration. It is noteworthy that cyto-chrome c and apoptosis are also increased in another in
vivo model of defective myelination, the Jimpy mouse[49]. Furthermore, neuronal loss has been suggested toprecede disruptions of myelination in patients with vita-min E deficiency [50]. The changes in the expressionprofiles of these two distinct functional classes of genesin Ttpa�/� mice suggest close coupling of apoptoticneuronal loss with loss of myelin and may account forthe loss of myelinated fibers reported in vitamin E-defi-cient mice [15] and in humans [50].
Suppression of genes for synaptic plasticity in Ttpa�/�
mice
The abundance of at least 20 mRNAs that encodeproteins enriched in synapses was decreased by 2- to20-fold in Ttpa�/� mice (Table 4 and Fig. 1). Thisobservation suggests an essential role of brain �-T in theregulation of synaptic structure and functions. The tran-scripts for complexins I and II were decreased by 2- and28-fold, respectively, in Ttpa�/� mice compared to theirwild-type littermates. These proteins interact with othervesicle proteins such as vesicle-associated membraneprotein 2 (mRNA repressed by 20-fold) and are essentialfor calcium-dependent docking and fusion of synapticvesicles to presynaptic membranes and release of neuro-transmitters [51].
Regulated release of excitatory neurotransmitters suchas glutamate from presynaptic terminals plays a vital rolein the electrophysiologically defined process of long-term potentiation [52]. Differential gene expression anal-ysis identified a decrease (�2.6-fold) in the abundance ofmRNAs that code for subunits of two distinct types ofglutamate receptors that regulate calcium influx intopostsynaptic terminals (Table 4 and Fig. 1). A coordi-nated decrease (up to 5-fold) was also detected in theexpression of mRNAs that encode calcium-regulated
Table 3. Markers of Neurodegeneration in Cortex of Ttpa�/� Mice
Foldchange Name Function(s)
Gene accessionnumber
�13.9 Hairless Transgenic null mice show increased age markers Z32675�13.0 ROR-related orphan receptor � Transgenic null mice show behavior dysfunctions U53228�9.8 Contactin-associated protein 1 Formation of nodes of Ranvier AF039833�5.3 Insulin receptor substrate 1 Regulates differentiation X69722�4.6 Proteolipid protein (myelin) Essential for myelin M16472�3.2 Cyclin-dependent kinase 5, regulatory subunit (p35) Phosphorylates � and regulates axonal microtubules U89527�2.8 Synuclein, � Prevents aggregation of �-synuclein and Lewy bodies AI839708�2.6 Sialyltransferase 9 Synthesis of glycolipids and glycoproteins Y15003�2.5 Sialyltransferase 7 Synthesis of glycolipids and glycoproteins X94000�2.3 Myelin-associated oligodendrocytic basic protein Synthesis of myelin U81317�2.0 Huntingtin-associated protein 1 Neurite development and synaptic function AJ002272
2.0 Synuclein-�, synaptic protein (induced by oxidative stress) Major component of Lewy Bodies AF04467213.0 Cytochrome c Mitochondrial, proapoptotic X01756
A selected list of differentially expressed mRNAs in cortex of TTP-deficient and normal mice. The encoded proteins regulate neurogenesis,myelination and neurodegeration.
1349Gene expression profile of TTP-null mice

proteins, such as visinin-like 1 and protein kinase C. Theimportance of these proteins in learning and memory iswell established [53]. Hence, GeneChip analysis of TTP-deficient mice identified a “global” suppression of tran-scriptionally regulated genes that are important in elec-trical activity of cortex. Functional prediction of thismolecular profile would be suppression in the electro-physiological activity of cortex. This was experimentallydemonstrated in a genetically related strain of mice withthe systemic deficiency of �-T [15].
Additionally, the failure of neurogenesis, increasedneurodegeneration, and loss of synaptic function inyoung mice (12–16 weeks old) deficient in �-tocopherolmay render these mice unresponsive to �-tocopherolsupplementation in their adulthood. This experimentallytestable hypothesis is based on experimental data thatshowed limited capacity for neurogenesis of adult cortex[54] and may account for failure or modest therapeuticeffects of vitamin E supplementations in elderly humans[55,56].
Conserved transcription factor-binding sites inpromoter regions of repressed genes
Differential gene expression analysis of cortex fromTtpa�/� and Ttpa�/� mice identified at least 20 genesthat were repressed in Ttpa�/� mice (Table 4, Fig. 1).The coordinated repression of these genes may be me-
Table 4. Suppression of Genes Encoding Synaptic Proteins in Cortex of Ttpa�/� mice
Foldchange Name
Gene accessionnumber
Proteins of synaptic vesicles�27.9 Complexin 2 D38613�19.7 Vesicle-associated membrane protein 2 U60150�10.6 ADP ribosylation factor 3 AI838022�3.5 Synaptotagmin 11 AB026808�2.8 Synuclein, � AI839708�2.8 Kinesin-heavy chain AF053473�2.5 Myosin-heavy chain AW124250�2.0 Complexin 1 D38614
Calcium signal tranducing proteins�4.6 Protein kinase C, � AF028009�4.0 Protein kinase C, � L28035�3.0 Visinin-like 1, calcium-binding protein D21165�2.6 Glutamate receptor, ionotropic, NMDA1 (� 1) D10028�2.5 ATPase, Na�/K� transporting, � 2 polypeptide AI839697�2.5 Glutamate receptor, ionotropic, AMPA1 (� 1) AW048549�2.3 Neurogranin (protein kinase C substrate, RC3) AI841709�2.3 Mitogen-activated protein kinase kinase kinase 5 AB006787�2.1 Protein kinase, cAMP-dependent regulatory, type I � M20473�2.1 Neuronal tyrosine/threonine phosphatase 1 X95518�2.0 Hippocalcin AB015200�2.0 Ephrin A3, ligand for receptor tyrosine kinase U92885�2.0 Potassium inwardly-rectifying channel, subfamily J, member 4 U11075�2.0 Protein tyrosine phosphatase, nonreceptor type substrate 1 D85785�2.1 Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein D83037
Differential analysis of mRNAs in cortex of TTP-deficient and normal mice identified 23 suppressed mRNAs that encodesynaptic proteins.
Fig. 1. Schematic illustration of some of the synaptic proteins known tobe important in learning and memory. The mRNAs for most of theseproteins were downregulated in TTP-deficient mice. 1 � complexin 1;2 � complexin 2; 3 � vesicle-associated membrane protein 2; 4 �ADP ribosylation factor 3; 5 � synaptogamin 11; 6 � kinesin-heavychain; 7 � myosin-heavy chain; 8 � visinin-like 1, calcium-bindingprotein; 9 � hippocalcin; and, 10 � neurogranin, substrate for proteinkinase c.
1350 K. GOHIL et al.
diated through the actions of transcription factors thatbind to DNA sequences conserved in their promoterregions. Therefore, the gene database was searched, asdescribed in Materials and Methods, for transcriptionfactors binding sequences in DNA upstream (�10 kb)of the coding regions of seven genes of the twentygenes. The selection was based on the availability ofthe complete DNA sequences of the candidate genesand the ease of analysis within the constraints ofcomputation time to illustrate the feasibility of theanalytical approach.
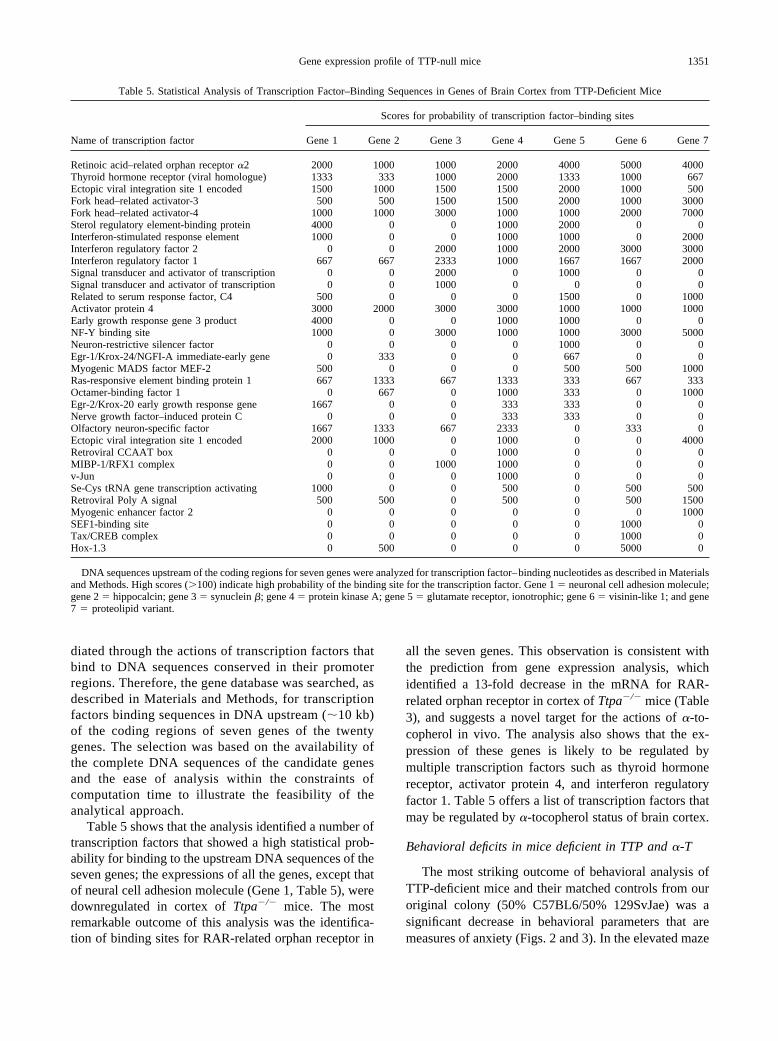
Table 5 shows that the analysis identified a number oftranscription factors that showed a high statistical prob-ability for binding to the upstream DNA sequences of theseven genes; the expressions of all the genes, except thatof neural cell adhesion molecule (Gene 1, Table 5), weredownregulated in cortex of Ttpa�/� mice. The mostremarkable outcome of this analysis was the identifica-tion of binding sites for RAR-related orphan receptor in
all the seven genes. This observation is consistent withthe prediction from gene expression analysis, whichidentified a 13-fold decrease in the mRNA for RAR-related orphan receptor in cortex of Ttpa�/� mice (Table3), and suggests a novel target for the actions of �-to-copherol in vivo. The analysis also shows that the ex-pression of these genes is likely to be regulated bymultiple transcription factors such as thyroid hormonereceptor, activator protein 4, and interferon regulatoryfactor 1. Table 5 offers a list of transcription factors thatmay be regulated by �-tocopherol status of brain cortex.
Behavioral deficits in mice deficient in TTP and �-T
The most striking outcome of behavioral analysis ofTTP-deficient mice and their matched controls from ouroriginal colony (50% C57BL6/50% 129SvJae) was asignificant decrease in behavioral parameters that aremeasures of anxiety (Figs. 2 and 3). In the elevated maze
Table 5. Statistical Analysis of Transcription Factor–Binding Sequences in Genes of Brain Cortex from TTP-Deficient Mice
Name of transcription factor
Scores for probability of transcription factor–binding sites
Gene 1 Gene 2 Gene 3 Gene 4 Gene 5 Gene 6 Gene 7
Retinoic acid–related orphan receptor �2 2000 1000 1000 2000 4000 5000 4000Thyroid hormone receptor (viral homologue) 1333 333 1000 2000 1333 1000 667Ectopic viral integration site 1 encoded 1500 1000 1500 1500 2000 1000 500Fork head–related activator-3 500 500 1500 1500 2000 1000 3000Fork head–related activator-4 1000 1000 3000 1000 1000 2000 7000Sterol regulatory element-binding protein 4000 0 0 1000 2000 0 0Interferon-stimulated response element 1000 0 0 1000 1000 0 2000Interferon regulatory factor 2 0 0 2000 1000 2000 3000 3000Interferon regulatory factor 1 667 667 2333 1000 1667 1667 2000Signal transducer and activator of transcription 0 0 2000 0 1000 0 0Signal transducer and activator of transcription 0 0 1000 0 0 0 0Related to serum response factor, C4 500 0 0 0 1500 0 1000Activator protein 4 3000 2000 3000 3000 1000 1000 1000Early growth response gene 3 product 4000 0 0 1000 1000 0 0NF-Y binding site 1000 0 3000 1000 1000 3000 5000Neuron-restrictive silencer factor 0 0 0 0 1000 0 0Egr-1/Krox-24/NGFI-A immediate-early gene 0 333 0 0 667 0 0Myogenic MADS factor MEF-2 500 0 0 0 500 500 1000Ras-responsive element binding protein 1 667 1333 667 1333 333 667 333Octamer-binding factor 1 0 667 0 1000 333 0 1000Egr-2/Krox-20 early growth response gene 1667 0 0 333 333 0 0Nerve growth factor–induced protein C 0 0 0 333 333 0 0Olfactory neuron-specific factor 1667 1333 667 2333 0 333 0Ectopic viral integration site 1 encoded 2000 1000 0 1000 0 0 4000Retroviral CCAAT box 0 0 0 1000 0 0 0MIBP-1/RFX1 complex 0 0 1000 1000 0 0 0v-Jun 0 0 0 1000 0 0 0Se-Cys tRNA gene transcription activating 1000 0 0 500 0 500 500Retroviral Poly A signal 500 500 0 500 0 500 1500Myogenic enhancer factor 2 0 0 0 0 0 0 1000SEF1-binding site 0 0 0 0 0 1000 0Tax/CREB complex 0 0 0 0 0 1000 0Hox-1.3 0 500 0 0 0 5000 0
DNA sequences upstream of the coding regions for seven genes were analyzed for transcription factor–binding nucleotides as described in Materialsand Methods. High scores (�100) indicate high probability of the binding site for the transcription factor. Gene 1 � neuronal cell adhesion molecule;gene 2 � hippocalcin; gene 3 � synuclein �; gene 4 � protein kinase A; gene 5 � glutamate receptor, ionotrophic; gene 6 � visinin-like 1; and gene7 � proteolipid variant.
1351Gene expression profile of TTP-null mice
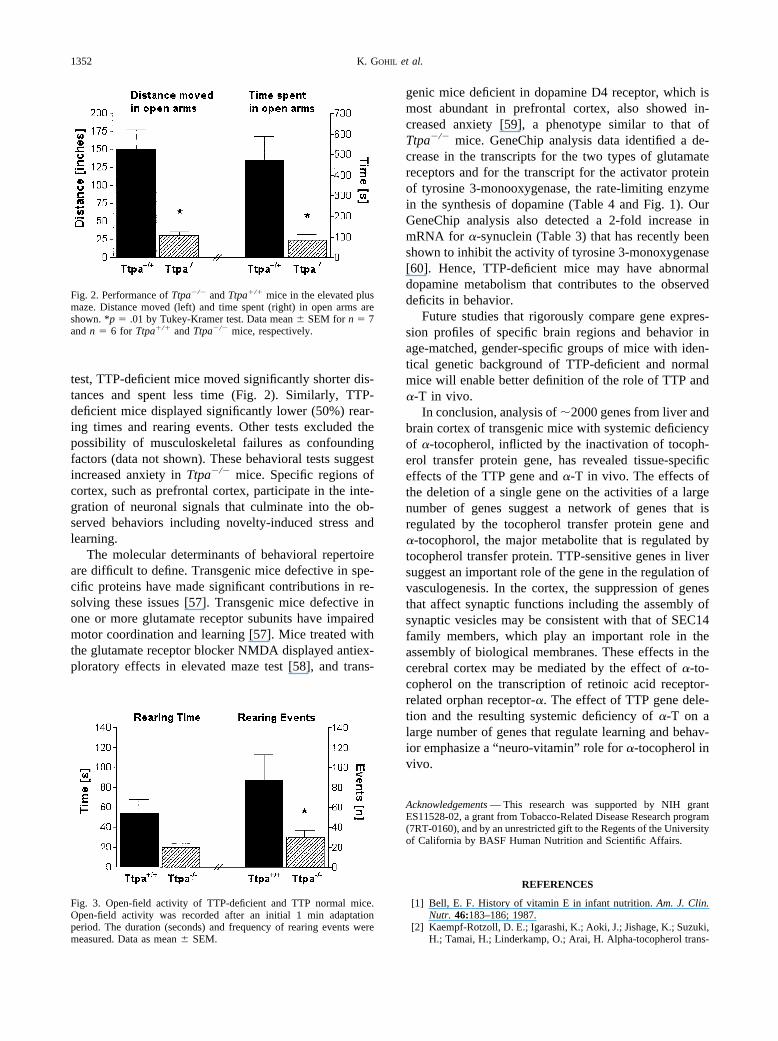
test, TTP-deficient mice moved significantly shorter dis-tances and spent less time (Fig. 2). Similarly, TTP-deficient mice displayed significantly lower (50%) rear-ing times and rearing events. Other tests excluded thepossibility of musculoskeletal failures as confoundingfactors (data not shown). These behavioral tests suggestincreased anxiety in Ttpa�/� mice. Specific regions ofcortex, such as prefrontal cortex, participate in the inte-gration of neuronal signals that culminate into the ob-served behaviors including novelty-induced stress andlearning.
The molecular determinants of behavioral repertoireare difficult to define. Transgenic mice defective in spe-cific proteins have made significant contributions in re-solving these issues [57]. Transgenic mice defective inone or more glutamate receptor subunits have impairedmotor coordination and learning [57]. Mice treated withthe glutamate receptor blocker NMDA displayed antiex-ploratory effects in elevated maze test [58], and trans-
genic mice deficient in dopamine D4 receptor, which ismost abundant in prefrontal cortex, also showed in-creased anxiety [59], a phenotype similar to that ofTtpa�/� mice. GeneChip analysis data identified a de-crease in the transcripts for the two types of glutamatereceptors and for the transcript for the activator proteinof tyrosine 3-monooxygenase, the rate-limiting enzymein the synthesis of dopamine (Table 4 and Fig. 1). OurGeneChip analysis also detected a 2-fold increase inmRNA for �-synuclein (Table 3) that has recently beenshown to inhibit the activity of tyrosine 3-monoxygenase[60]. Hence, TTP-deficient mice may have abnormaldopamine metabolism that contributes to the observeddeficits in behavior.
Future studies that rigorously compare gene expres-sion profiles of specific brain regions and behavior inage-matched, gender-specific groups of mice with iden-tical genetic background of TTP-deficient and normalmice will enable better definition of the role of TTP and�-T in vivo.
In conclusion, analysis of �2000 genes from liver andbrain cortex of transgenic mice with systemic deficiencyof �-tocopherol, inflicted by the inactivation of tocoph-erol transfer protein gene, has revealed tissue-specificeffects of the TTP gene and �-T in vivo. The effects ofthe deletion of a single gene on the activities of a largenumber of genes suggest a network of genes that isregulated by the tocopherol transfer protein gene and�-tocophorol, the major metabolite that is regulated bytocopherol transfer protein. TTP-sensitive genes in liversuggest an important role of the gene in the regulation ofvasculogenesis. In the cortex, the suppression of genesthat affect synaptic functions including the assembly ofsynaptic vesicles may be consistent with that of SEC14family members, which play an important role in theassembly of biological membranes. These effects in thecerebral cortex may be mediated by the effect of �-to-copherol on the transcription of retinoic acid receptor-related orphan receptor-�. The effect of TTP gene dele-tion and the resulting systemic deficiency of �-T on alarge number of genes that regulate learning and behav-ior emphasize a “neuro-vitamin” role for �-tocopherol invivo.
Acknowledgements — This research was supported by NIH grantES11528-02, a grant from Tobacco-Related Disease Research program(7RT-0160), and by an unrestricted gift to the Regents of the Universityof California by BASF Human Nutrition and Scientific Affairs.
REFERENCES
[1] Bell, E. F. History of vitamin E in infant nutrition. Am. J. Clin.Nutr. 46:183–186; 1987.
[2] Kaempf-Rotzoll, D. E.; Igarashi, K.; Aoki, J.; Jishage, K.; Suzuki,H.; Tamai, H.; Linderkamp, O.; Arai, H. Alpha-tocopherol trans-
Fig. 2. Performance of Ttpa�/� and Ttpa�/� mice in the elevated plusmaze. Distance moved (left) and time spent (right) in open arms areshown. *p � .01 by Tukey-Kramer test. Data mean � SEM for n � 7and n � 6 for Ttpa�/� and Ttpa�/� mice, respectively.
Fig. 3. Open-field activity of TTP-deficient and TTP normal mice.Open-field activity was recorded after an initial 1 min adaptationperiod. The duration (seconds) and frequency of rearing events weremeasured. Data as mean � SEM.
1352 K. GOHIL et al.

fer protein is specifically localized at the implantation site ofpregnant mouse uterus. Biol. Reprod. 67:599–604; 2002.
[3] Brigelius-Flohe, R.; Traber, M. G. Vitamin E: function and me-tabolism. FASEB J. 13:1145–1155; 1999.
[4] Azzi, A.; Ricciarelli, R.; Zingg, J. M. Nonantioxidant molecularfunctions of �-tocopherol (vitamin E). FEBS Lett. 519:8–10;2002.
[5] Yamauchi, J.; Iwamoto, T.; Kida, S.; Masushige, S.; Yamada, K.;Esashi, T. Tocopherol-associated protein is a ligand-dependenttranscriptional activator. Biochem. Biophys. Res. Commun. 285:295–299; 2001.
[6] Traber, M. G.; Arai, H. Molecular mechanisms of vitamin Etransport. Annu. Rev. Nutr. 19:343–355; 1999.
[7] Sato, Y.; Arai, H.; Miyata, A.; Tokita, S.; Yamamoto, K.; Tanabe,T.; Inoue, K. Primary structure of �-tocopherol transfer proteinfrom rat liver. Homology with cellular retinaldehyde-bindingprotein. J. Biol. Chem. 268:17705–17710; 1993.
[8] Arita, M.; Sato, Y.; Miyata, A.; Tanabe, T.; Takahashi, E.; Kay-den, H. J.; Arai, H.; Inoue, K. Human �-tocopherol transferprotein: cDNA cloning, expression, and chromosomal localiza-tion. Biochem. J. 306:437–443; 1995.
[9] Zimmer, S.; Stocker, A.; Sarbolouki, M. N.; Spycher, S. E.;Sassoon, J.; Azzi, A. A novel human tocopherol-associated pro-tein: cloning, in vitro expression, and characterization. J. Biol.Chem. 275:25672–25680; 2000.
[10] Sha, B.; Phillips, S. E.; Bankaitis, V. A.; Luo, M. Crystal structureof the Saccharomyces cerevisiae phosphatidylinositol-transferprotein. Nature 391:506–510; 1998.
[11] Shibata, N.; Arita, M.; Misaki, Y.; Dohmae, N.; Takio, K.; Ono,T.; Inoue, K.; Arai, H. Supernatant protein factor, which stimu-lates the conversion of squalene to lanosterol, is a cytosolicsqualene transfer protein and enhances cholesterol biosynthesis.Proc. Natl. Acad. Sci. USA 98:2244–2249; 2001.
[12] Singh, D. K.; Mokashi, V.; Elmore, C. L.; Porter, T. D. Phos-phorylation of supernatant protein factor enhances its ability tostimulate microsomal squalene monooxygenase. J. Biol. Chem.278:5646–5651; 2003.
[13] Yokota, T.; Shiojiri, T.; Gotoda, T.; Arita, M.; Arai, H.; Ohga, T.;Kanda, T.; Suzuki, J.; Imai, T.; Matsumoto, H.; Harino, S.;Kiyosawa, M.; Mizusawa, H.; Inoue, K. Friedreich-like ataxiawith retinitis pigmentosa caused by the His101Gln mutation ofthe �-tocopherol transfer protein gene. Ann. Neurol. 41:826–832;1997.
[14] Terasawa, Y.; Ladha, Z.; Leonard, S. W.; Morrow, J. D.; New-land, D.; Sanan, D.; Packer, L.; Traber, M. G.; Farese, R. V. JrIncreased atherosclerosis in hyperlipidemic mice deficient in�-tocopherol transfer protein and vitamin E. Proc. Natl. Acad.Sci. USA 97:13830–13834; 2000.
[15] Yokota, T.; Igarashi, K.; Uchihara, T.; Jishage, K.; Tomita, H.;Inaba, A.; Li, Y.; Arita, M.; Suzuki, H.; Mizusawa, H.; Arai, H.Delayed-onset ataxia in mice lacking �-tocopherol transfer pro-tein: model for neuronal degeneration caused by chronic oxidativestress. Proc. Natl. Acad. Sci. USA 98:15185–15190; 2001.
[16] Leonard, S. W.; Terasawa, Y.; Farese, R. V. Jr.; Traber, M. G.Incorporation of deuterated RRR- or all-rac-�-tocopherol inplasma and tissues of �-tocopherol transfer protein-null mice.Am. J. Clin. Nutr. 75:555–560; 2002.
[17] Raber, J.; Akana, S. F.; Bhatnagar, S.; Dallman, M. F.; Wong, D.;Mucke, L. Hypothalamic-pituitary-adrenal dysfunction inApoE(�/�) mice: possible role in behavioral and metabolicalterations. J. Neurosci. 20:2064–2071; 2000.
[18] Gohil, K.; Moy, R. K.; Farzin, S.; Maguire, J. J.; Packer, L.mRNA expression profile of a human cancer cell line in responseto Ginkgo biloba extract: induction of antioxidant response andthe Golgi system. Free Radic. Res. 33:831–849; 2000.
[19] Watanabe, C. M.; Wolffram, S.; Ader, P.; Rimbach, G.; Packer,L.; Maguire, J. J.; Schultz, P. G.; Gohil, K. The in vivo neuro-modulatory effects of the herbal medicine ginkgo biloba. Proc.Natl. Acad. Sci. USA 98:6577–6580; 2001.
[20] Lockhart, D. J.; Dong, H.; Byrne, M. C.; Follettie, M. T.; Gallo,M. V.; Chee, M. S.; Mittmann, M.; Wang, C.; Kobayashi, M.;
Horton, H.; Brown, E. L. Expression monitoring by hybridizationto high-density oligonucleotide arrays. Nat. Biotechnol. 14:1675–1680; 1996.
[21] Thibault, C.; Lai, C.; Wilke, N.; Duong, B.; Olive, M. F.; Rah-man, S.; Dong, H.; Hodge, C. W.; Lockhart, D. J.; Miles, M. F.Expression profiling of neural cells reveals specific patterns ofethanol-responsive gene expression. Mol. Pharmacol. 58:1593–1600; 2000.
[22] Quandt, K.; Frech, K.; Karas, H.; Wingender, E.; Werner, T.MatInd and MatInspector: new fast and versatile tools for detec-tion of consensus matches in nucleotide sequence data. NucleicAcids Res. 23:4878–4884; 1995.
[23] Zheng, H.; Berman, N. E.; Klaassen, C. D. Chemical modulationof metallothionein I and III mRNA in mouse brain. Neurochem.Int. 27:43–58; 1995.
[24] Gohil, K.; Cross, C. E.; Last, J. A. Ozone-induced disruptions oflung transcriptomes. Biochem. Biophys. Res. Commun. 305:719–728; 2003.
[25] Noveroske, J. K.; Lai, L.; Gaussin, V.; Northrop, J. L.; Nakamura,H.; Hirschi, K. K.; Justice, M. J. Quaking is essential for bloodvessel development. Genesis 32:218–230; 2002.
[26] Gendron, R. L.; Adams, L. C.; Paradis, H. Tubedown-1, a novelacetyltransferase associated with blood vessel development. Dev.Dyn. 218:300–315; 2000.
[27] Chajara, A.; Raoudi, M.; Delpech, B.; Levesque, H. The fibro-proliferative response of arterial smooth muscle cells to ballooncatheter injury is associated with increased hyaluronidase produc-tion and hyaluronan degradation. Atherosclerosis 157:293–300;2001.
[28] Fischer, A.; Pallauf, J.; Gohil, K.; Weber, S. U.; Packer, L.;Rimbach, G. Effect of selenium and vitamin E deficiency ondifferential gene expression in rat liver. Biochem. Biophys. Res.Commun. 285:470–475; 2001.
[29] Kong, A. N.; Owuor, E.; Yu, R.; Hebbar, V.; Chen, C.; Hu, R.;Mandlekar, S. Induction of xenobiotic enzymes by the MAPkinase pathway and the antioxidant or electrophile response ele-ment (ARE/EpRE). Drug Metab. Rev. 33:255–271; 2001.
[30] Buetler, T. M.; Gallagher, E. P.; Wang, C.; Stahl, D. L.; Hayes,J. D.; Eaton, D. L. Induction of phase I and phase II drug-metabolizing enzyme mRNA, protein, and activity by BHA,ethoxyquin, and oltipraz. Toxicol. Appl. Pharmacol. 135:45–57;1995.
[31] Danielson, P. B. The cytochrome P450 superfamily: biochemis-try, evolution, and drug metabolism in humans. Curr. DrugMetab. 3:561–597; 2002.
[32] Hayes, J. D.; Pulford, D. J. The glutathione S-transferase super-gene family: regulation of GST and the contribution of the isoen-zymes to cancer chemoprotection and drug resistance. Crit. Rev.Biochem. Mol. Biol. 30:445–600; 1995.
[33] Davis, W. Jr.; Ronai, Z.; Tew, K. D. Cellular thiols and reactiveoxygen species in drug-induced apoptosis. J. Pharmacol. Exp.Ther. 296:1–6; 2001.
[34] Volm, M. Multidrug resistance and its reversal. Anticancer Res.18:2905–2917; 1998.
[35] Ibrahim, W.; Lee, U. S.; Yeh, C. C.; Szabo, J.; Bruckner, G.;Chow, C. K. Oxidative stress and antioxidant status in mouseliver: effects of dietary lipid, vitamin E, and iron. J. Nutr. 127:1401–1406; 1997.
[36] Roy, S.; Lado, B. H.; Khanna, S.; Sen, C. K. Vitamin E sensitivegenes in the developing rat fetal brain: a high-density oligonucle-otide microarray analysis. FEBS Lett. 530:17–23; 2002.
[37] Steinmayr, M.; Andre, E.; Conquet, F.; Rondi-Reig, L.; Delhaye-Bouchaud, N.; Auclair, N.; Daniel, H.; Crepel, F.; Mariani, J.;Sotelo, C.; Becker-Andre, M. Staggerer phenotype in retinoid-related orphan receptor �-deficient mice. Proc. Natl. Acad. Sci.USA 95:3960–3965; 1998.
[38] Goodall, G.; Gheusi, G. Abnormal patterns of maze patrolling inthe mutant mouse staggerer. Behav. Neural Biol. 47:307–320;1987.
[39] Thompson, C. C.; Potter, G. B. Thyroid hormone action in neuraldevelopment. Cereb. Cortex 10:939–945; 2000.
1353Gene expression profile of TTP-null mice

[40] Garcia-Atares, N.; San Jose, I.; Cabo, R.; Vega, J. A.; Represa, J.Changes in the cerebellar cortex of hairless Rhino-J mice (hr-rh-j). Neurosci. Lett. 256:13–16; 1998.
[41] Gollan, L.; Sabanay, H.; Poliak, S.; Berglund, E. O.; Ranscht, B.;Peles, E. Retention of a cell adhesion complex at the paranodaljunction requires the cytoplasmic region of Caspr. J. Cell Biol.157:1247–1256; 2002.
[42] Spillantini, M. G.; Schmidt, M. L.; Lee, V. M.; Trojanowski,J. Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies.Nature 388:839–840; 1997.
[43] Goedert, M. Alpha-synuclein and neurodegenerative diseases.Nat. Rev. Neurosci. 2:492–501; 2001.
[44] Cummings, J. L. Lewy body diseases with dementia: pathophys-iology and treatment. Brain Cogn. 28:266–280; 1995.
[45] Uversky, V. N.; Li, J.; Souillac, P.; Millett, I. S.; Doniach, S.;Jakes, R.; Goedert, M.; Fink, A. L. Biophysical properties of thesynucleins and their propensities to fibrillate: inhibition of�-synuclein assembly by �- and �-synucleins. J. Biol. Chem.277:11970–11978; 2002.
[46] Hashimoto, M.; Rockenstein, E.; Mante, M.; Mallory, M.;Masliah, E. Beta-synuclein inhibits �-synuclein aggregation: apossible role as an anti-parkinsonian factor. Neuron 32:213–223;2001.
[47] Junn, E.; Mouradian, M. M. Human �-synuclein overexpressionincreases intracellular reactive oxygen species levels and suscep-tibility to dopamine. Neurosci. Lett. 320:146–150; 2002.
[48] Chandra, D.; Liu, J. W.; Tang, D. G. Early mitochondrial activa-tion and cytochrome c upregulation during apoptosis. J. Biol.Chem. 277:50842–50854; 2002.
[49] Cerghet, M.; Bessert, D. A.; Nave, K. A.; Skoff, R. P. Differentialexpression of apoptotic markers in jimpy and in Plp overexpres-sors: evidence for different apoptotic pathways. J. Neurocytol.30:841–855; 2001.
[50] Sokol, R. J.; Kayden, H. J.; Bettis, D. B.; Traber, M. G.; Neville,H.; Ringel, S.; Wilson, W. B.; Stumpf, D. A. Isolated vitamin Edeficiency in the absence of fat malabsorption—familial andsporadic cases: characterization and investigation of causes.J. Lab. Clin. Med. 111:548–559; 1988.
[51] Marz, K. E.; Hanson, P. I. Sealed with a twist: complexin and thesynaptic SNARE complex. Trends Neurosci. 25:381–383; 2002.
[52] Sheng, M.; Kim, M. J. Postsynaptic signaling and plasticity mech-anisms. Science 298:776–780; 2002.
[53] Shobe, J. The role of PKA, CaMKII, and PKC in avoidanceconditioning: permissive or instructive? Neurobiol. Learn Mem.77:291–312; 2002.
[54] Anderson, M. F.; Aberg, M. A.; Nilsson, M.; Eriksson, P. S.Insulin-like growth factor-I and neurogenesis in the adult mam-malian brain. Brain Res. Dev. Brain Res. 134:115–122; 2002.
[55] Floyd, R. A.; Hensley, K. Oxidative stress in brain aging. Impli-cations for therapeutics of neurodegenerative diseases. Neurobiol.Aging 23:795–807; 2002.
[56] Grodstein, F.; Chen, J.; Willett, W. C. High-dose antioxidantsupplements and cognitive function in community-dwelling el-derly women. Am. J. Clin. Nutr. 77:975–984; 2003.
[57] Chen, C.; Tonegawa, S. Molecular genetic analysis of synapticplasticity, activity-dependent neural development, learning, andmemory in the mammalian brain. Annu. Rev. Neurosci. 20:157–184; 1997.
[58] Vasar, E.; Harro, J.; Lang, A.; Soosaar, A.; Oopik, T.; Koks, S.;Sihver, S.; Volke, V. Anti-exploratory effect of N-methyl-D-aspartate in elevated plus-maze. Involvement of NMDA and CCKreceptors. Eur. Neuropsychopharmacol. 3:63–73; 1993.
[59] Falzone, T. L.; Gelman, D. M.; Young, J. I.; Grandy, D. K.; Low,M. J.; Rubinstein, M. Absence of dopamine D4 receptors resultsin enhanced reactivity to unconditioned, but not conditioned, fear.Eur. J. Neurosci. 15:158–164; 2002.
[60] Perez, R. G.; Waymire, J. C.; Lin, E.; Liu, J. J.; Guo, F.; Zigmond,M. J. A role for �-synuclein in the regulation of dopaminebiosynthesis. J. Neurosci. 22:3090–3099; 2002.
ABBREVIATIONS
�-T—�-tocopherolNMDA—N-methyl-D-aspartateTTP—tocopherol transfer proteinTtpa—gene for �-tocopherol transfer proteinTtpa�/�—normal mice with functional TTP geneTtpa�/�—mice deficient in TTP gene and TTPVE—vitamin E
1354 K. GOHIL et al.
Related Documents











