Thompson and Gilbert R. Upchurch, Jr P. Kaldjian, Guanyi Lu, Peter K. Henke, James C. Stanley, Stephen J. Weiss, Robert W. Gorav Ailawadi, Jonathan L. Eliason, Karen J. Roelofs, Indranil Sinha, Kevin K. Hannawa, Eric Gender Differences in Experimental Aortic Aneurysm Formation Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2004 American Heart Association, Inc. All rights reserved. Greenville Avenue, Dallas, TX 75231 is published by the American Heart Association, 7272 Arteriosclerosis, Thrombosis, and Vascular Biology doi: 10.1161/01.ATV.0000143386.26399.84 2004; 2004;24:2116-2122; originally published online August 26, Arterioscler Thromb Vasc Biol. http://atvb.ahajournals.org/content/24/11/2116 World Wide Web at: The online version of this article, along with updated information and services, is located on the http://atvb.ahajournals.org//subscriptions/ at: is online Arteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions: http://www.lww.com/reprints Information about reprints can be found online at: Reprints: document. Question and Answer Permissions and Rights page under Services. Further information about this process is available in the which permission is being requested is located, click Request Permissions in the middle column of the Web Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for can be obtained via RightsLink, a service of the Arteriosclerosis, Thrombosis, and Vascular Biology in Requests for permissions to reproduce figures, tables, or portions of articles originally published Permissions: by guest on November 24, 2014 http://atvb.ahajournals.org/ Downloaded from by guest on November 24, 2014 http://atvb.ahajournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Thompson and Gilbert R. Upchurch, JrP. Kaldjian, Guanyi Lu, Peter K. Henke, James C. Stanley, Stephen J. Weiss, Robert W.
Gorav Ailawadi, Jonathan L. Eliason, Karen J. Roelofs, Indranil Sinha, Kevin K. Hannawa, EricGender Differences in Experimental Aortic Aneurysm Formation
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2004 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.0000143386.26399.842004;
2004;24:2116-2122; originally published online August 26,Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/24/11/2116World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from

Gender Differences in Experimental AorticAneurysm Formation
Gorav Ailawadi, Jonathan L. Eliason, Karen J. Roelofs, Indranil Sinha, Kevin K. Hannawa,Eric P. Kaldjian, Guanyi Lu, Peter K. Henke, James C. Stanley, Stephen J. Weiss,
Robert W. Thompson, Gilbert R. Upchurch, Jr
Objective—It is hypothesized that a male predominance, similar to that in humans, persists in a rodent model ofexperimental abdominal aortic aneurysm (AAA) via alterations in matrix metalloproteinases (MMPs).
Methods and Results—Group I experiments were as follows: elastase perfusion of the infrarenal aorta was performed inmale (M) and female (F) rats. At 14 days, aortas were harvested for immunohistochemistry, real-time polymerase chainreaction (PCR), and zymography. Group II experiments were the following: abdominal aorta was transplanted from For M donors into F or M recipients. At 14 days, rodents that had undergone transplantation underwent elastase perfusion.In group III, male rats were given estradiol or sham 5 days before elastase perfusion. In group I, M rats had larger AAAswith higher frequency than did F rats. M rat aortas had more significant macrophage infiltrates and increased matrixmetalloproteinase (MMP)-9 production and activity. In group II, M-to-M aortic transplants uniformly developedaneurysms after elastase perfusion, whereas F-to-F aortic transplants remained resistant to aneurysm formation. F aortastransplanted into M recipients, however, lost aneurysm resistance. In group III, estradiol-treated rats demonstratedsmaller aneurysms and less macrophage infiltrate and MMP-9 compared with M controls after elastase.
Conclusions—These data provide evidence of gender-related differences in AAA development, which may reflect anestrogen-mediated reduction in macrophage MMP-9 production. (Arterioscler Thromb Vasc Biol. 2004;24:2116-2122.)
Key Words: aorta � aneurysm � genetic � estrogen � metalloproteinase
Abdominal aortic aneurysms (AAAs) are potentially life-threatening, accounting for 150 000 hospital admissions
yearly.1 Clear gender differences exist, with a prevalence inmen 4-times that in women.2,3 The diminished risk of AAAdevelopment is lost in women after menopause, suggestingthat reproductive events, including circulating estrogens, mayplay a protective role.4,5
Prominent local inflammatory cell infiltration, aortic wallcytokine production, medial wall destruction by proteinases,and smooth muscle cell depletion characterize most AAAs.Destruction of elastin and collagen in the media by variousmatrix metalloproteinases (MMPs) is considered an essentialelement of AAA formation.6–8 MMP-1, MMP-2, MMP-3,MMP-9, MMP-12, as well as tissue inhibitor ofmetalloproteinase-1, are all upregulated in the walls of humanAAAs.9–14 Two of these, MMP-2 and MMP-9, have beenextensively studied. MMP-9 has attracted particular attentionin that it is highly expressed in human AAA wall and ispresent in serum from AAA patients.14,15 Mice with deletionof the gene responsible for the MMP-9 protein are resistant to
the development of experimental AAAs.10 In addition,MMP-2, derived from aortic mesenchymal cells, appearsnecessary for experimental aneurysm formation.16
Many studies implicating MMPs in AAA evolution haveused a rat or mouse model with porcine pancreatic elastaseperfusion of the infrarenal aorta. This model causes an initialinflux of macrophages and lymphocytes leading to destruc-tion and remodeling of the aortic wall matrix, and subsequentaneurysm development.17 Atherosclerosis, only once consid-ered an essential to aneurysm development, is not thought tobe the mechanism responsible for AAA formation.18,19 Im-portantly, nearly all previous studies have been performedusing male rodents. To date, the influence of gender onexperimental AAA formation has received little attention.Furthermore, although estrogen is known to affect collagenand elastin matrix remodeling in rats,20 its role in AAAformation has not been studied. This investigation wasdesigned to determine the relevance of male and femalegender on experimental AAA formation and to define localand systemic events that might influence any anticipateddifferences related to gender.
Original received December 15, 2003; final version accepted August 10, 2004.From Jobst Vascular Research Laboratories, Section of Vascular Surgery (G.A., J.L.E., K.J.R., I.S., K.K.H., P.K.H., J.C.S., G.R.U.), Section of
Transplant Surgery (G.L.), Department of Pathology (E.P.K.), and Division of Molecular Medicine and Genetics, Department of Internal Medicine(S.J.W.), University of Michigan Medical School, Ann Arbor; and the Section of Vascular Surgery, Department of Surgery and Department of CellBiology and Physiology (R.W.T.), Washington University Medical School, St Louis, Missouri.
Correspondence to Dr Gilbert R. Upchurch Jr, University of Michigan Medical Center, 2210 Taubman Health Care Center, 1500 E Medical CenterDrive, Ann Arbor, MI 48109-0329. E-mail [email protected]
© 2004 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://www.atvbaha.org DOI: 10.1161/01.ATV.0000143386.26399.84
2116 by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from

MethodsSprague-Dawley rats (200 to 250 grams, age 8 to 10 weeks),obtained from Charles River Laboratories (Wilmington, Mass), wereused in all experiments. Procedures and experiments were approvedby the University of Michigan Universal Committee on the Use andCare of Animals (#8220 and #8314).
Elastase Perfusion Aneurysm ModelPancreatic porcine elastase perfusion of the rat aortas was performedas described previously.17 Male and female rats (n�15, each) wereanesthetized with 2 to 2.5% isoflurane inhalation, and the infrarenalabdominal aorta was isolated under sterile conditions. Digital videomicrometry was performed to directly measure outer aortic diameter.Specifically, images of the aorta were obtained using a Spot InsightColor Optical Camera (Diagnostic Instruments, Sterling Heights,Mich) attached to an operating microscope (Nikon, Melville, NY).Aortic diameters were then measured at the level of the left renalvein, the mid-infrarenal aorta, and the aortic bifurcation in triplicateusing Image Pro Express software (Media Cybernetics Inc, SilverSpring, Md). Temporary proximal and distal aortic control wasobtained using temporary 4-0 cotton suture loops, following whichan aortotomy was made near the aortic bifurcation with a 30-gaugeneedle. The infrarenal aorta was cannulated with PE-10 tubing andperfused with 12 U of porcine pancreatic elastase diluted to a totalvolume of 2 mL with sterile normal saline (Lot #032K7660 or Lot#102K685; Sigma, St. Louis, Mo) over 60 minutes. Subsequently,the tubing was removed and the aortotomy repaired with 10-0monofilament suture. Patency was assured in all cases. Aorticdiameter measurements were repeated immediately after perfusion.The intestines were replaced; the abdominal wall was closed; and therats were recovered. At 7 or 14 days, aortas were re-exposed andaortic diameters were re-measured in vivo. Aneurysm formation wasdefined as a 100% increase in an individual animal’s pre-elastaseperfusion aortic diameter. The infrarenal aorta was then removed andsubjected to histological study, immunohistochemistry, and quanti-tative polymerase chain reaction (PCR).
Aortic TransplantationIn additional rats, transplantation of the infrarenal rat aorta wasperformed as previously described.21 Briefly, male and female donorrats were anesthetized and the abdominal aortas isolated. Donor ratswere anticoagulated with 300 U of heparin and the abdominal aortawas rapidly removed and placed in cold 0.9% normal saline. Therecipient rats’ infrarenal abdominal aortas were similarly isolatedand proximal and distal aortic control was obtained with temporary4-0 cotton suture loops. The recipient abdominal aorta was excisedand donor abdominal aorta was transplanted into the infrarenalposition of a size-matched recipient using a running 10-0 monofil-ament suture in an end-to-end fashion. After aortic patency wasassured, the abdominal incision was closed and rats were recovered.
Female donor aortas were transplanted into male recipients (n�7)with controls including female aortas transplanted into femalerecipients (n�7) and male aortas transplanted into male recipients(n�9). Fourteen days after transplantation, the transplanted aortaswere subjected to pancreatic porcine elastase perfusion and harvestedafter 14 days as previously noted.
Estrogen Pellet ImplantationIn other experiments, male rats were randomized to implantation ofan estrogen pellet or sham implantation (n�13, each). The formerinvolved the subcutaneous implantation of a 21-day slow-release0.1-mg 17�-estradiol pellet (Innovative Research of America, Sara-sota, Fla) in the posterior neck. Sham rats underwent the sameimplantation procedure without insertion of any pellet. This partic-ular estradiol dose results in serum estradiol levels at 2 to 3 weeks of44.7�6.1 pg/mL compared with 15.0�2.2 pg/mL in control rats(P�0.05).22 Rat aortas from both groups were subjected to elastaseperfusion 5 days later. The aortas were then removed for study 7 or14 days after elastase perfusion.
Histological AnalysisAll excised aortas were fixed in 10% formalin for 18 hours, followedby immersion in 70% ethanol for 24 hours. Aortas were thenimbedded in paraffin and 4-�m sections were prepared with hema-toxylin and eosin and Verhoeff-Van Gieson stains.
Immunohistochemistry was undertaken after deparaffinization,rehydration, and unmasking using Trilogy (Cell Marque Corp, HotSprings, Ariz) in a Princess model pressure cooker (Cell Marque).Endogenous peroxidase activity was then blocked using 3% hydro-gen peroxide in methanol. To help ensure that rejection was notoccurring in elastase-perfused or transplanted animals, anti-T lym-phocyte immunohistochemistry was performed. Specifically, antiratCD3 monoclonal antibody (BD Pharmingen, San Diego, Calif) wasused as the primary antibody and mouse IgG Vectastain (VectorLaboratories, Burlingame, Calif) as the secondary antibody. Ratspleen was used as the positive control for anti-CD3 staining. ED-1macrophage staining was performed using mouse antirat ED-1primary antibody (Serotec, Raleigh, NC) and mouse IgG Vectastainsecondary antibody (Vector Laboratories). MMP-9 immunohisto-chemistry was performed using rabbit antirat MMP-9 polyclonalprimary antibody (Chemicon International, Temecula, Calif) andrabbit IgG Vectastain secondary antibody (Vector Laboratories).Staining for all these antibodies was performed using Vector Redalkaline phosphatase (Vector Laboratories) followed by hematoxylinQS counterstain (Vector Laboratories).
Colocalization studies involved deparaffinization, rehydration,and unmasking as previously described. Endogenous peroxidaseactivity was blocked using 3% hydrogen peroxide in methanol.Staining for ED-1 was performed using mouse antirat ED-1 primaryantibody (Serotec), mouse IgG Vectastain secondary antibody (Vec-tor Laboratories), and Vector Blue alkaline phosphatase. Sampleswere stored in PBS at 4°C overnight. The next day, staining forMMP-9 was performed using rabbit antirat MMP-9 polyclonalprimary antibody AB19016 (Chemicon International), rabbit IgGVectastain secondary antibody (Vector Laboratories), and VectorRed alkaline phosphatase stain.
Quantitative PCRExpression of MMP-9 and �-actin mRNA was determined usingquantitative PCR. Messenger RNA was isolated by exposure ofaortas to TRIzol reagent and reverse-transcribed by incubating witholigo-dT primer (Life Technologies, Grand Island, NY) and M-MLVReverse Transcriptase (Life Technologies, Grand Island, NY) at94°C for 3 minutes, followed by 40°C for 70 minutes. The resultantcDNA was amplified by Taq Polymerase (Promega, Madison, Wis)in a SmartCycler quantitative PCR system (Cepheid, Sunnyvale,Calif). SYBR intercalating dye (Roche, Indianapolis, Ind) was usedto monitor cDNA amplification for each gene. MMP-9 and �-actinprimer sequences were derived using Primer Premeir software(PREMIER Biosoft International, Palo Alto, Calif) based on primarycDNA sequences from GenBank (http://www.ncbi.nlm.nih.gov/Genbank/). Primer sequences are as follows: MMP-9 forwardprimer, CGC CAA CTA TGA CCA GGA TA; MMP-9 reverseprimer, GTT GCC CCC AGT TAC AGT; �-actin forward primer,ATG GGT CAG AAG GAT TCC TAT GTG; �-actin reverseprimer, CTT CAT GAG GTA GTC AGT CAG GTC. Results werenormalized using �-actin to account for variation in mRNA amounts.Quantification of mRNA levels used �Ct values, calculated by theformula:
�C t�Ct target gene�Ct �-actin.
Expression of the target gene in ratio to �-actin expression wascalculated by the formula:
target gene expression/�-actin expression�2�(�Ct)
Substrate Gel ZymographyMMP-9 distribution after elastase perfusion was determined byzymography as previously described.23 Gelatinase activity wasevident by clear bands against a dark blue background. The molec-
Ailawadi et al Gender Differences in Experimental AA Formation 2117
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from

ular weight of each band was determined by comparison of the bandsagainst samples containing human recombinant MMP-9 (Oncogene,Boston, Mass). In previous studies, these bands were inhibited byEDTA and are thus metalloproteinases.23 Semiquantitative measure-ments were performed using densitometry as described and normal-ized to total protein.
DensitometryGels were imaged with a FOTO/Analyst charge-coupled deviceCAMERA (Fotodyne, Hartland, Wis). Band strengths were quanti-fied using GEL-Pro Analyzer software version 3.1 (Media Cyber-netics, Silver Springs, Md).
Total Protein AssayTotal cellular protein was determined by a bicinchoninic acid proteinassay (Pierce, Rockford, Ill) in aortas on which MMP-9 activityassays were performed after they had been solubilized in 0.1%sodium dodecyl sulfate.
Data AnalysisData are represented as mean�SE. Data were assessed by nonpairedt test or ANOVA with statistical significance assigned as P�0.05.When significance was reached, post hoc Tukey test was used tocompare individual groups. Statistical analysis was performed usingPrism software (GraphPad Software, San Diego, Calif).
ResultsBaseline Histology in Male Versus Female RatsMale and female rats’ aortas not subjected to any interventionwere harvested and subjected to histological analysis. Maleand female aortas were nearly identical in wall thickness andaortic lamellar structure (data not shown) by Verhoeff-VanGieson stain. Aortas from both genders were indistinguish-able by an experienced pathologist (E.P.K). CD3 immuno-histochemistry demonstrated little to no lymphocytic infil-trate in the aortas of either males or females (data not shown).
Experimental AAA Formation in Male VersusFemale RatsPreperfusion baseline aortic diameters were not different(P�0.20) between male and female aortas (1.41�0.16 versus1.32�0.07, respectively). The mean increase in aortic dia-meter 14 days after elastase perfusion in male aortas was200�37.6%, whereas female aortas had a mean aortic dia-meter increase of 69.4�26.5% (P�0.0234; Figure 1a). Theincidence of AAAs defined as an increase in aortic diameter atleast 100% from preperfusion diameter was 82% in male ratscompared with 29% in female rats (P�0.0229; Figure 1a).
Male aneurysmal aortas exhibited circumferential disrup-tion of the elastic lamellae 14 days after elastase perfusion,whereas the aortas of female rats after elastase perfusion hadlargely intact elastin fibers (please see www.ahajournals.org).CD3 staining demonstrated minimal lymphocyte infiltrationin either male or female aortas (data not shown). ED-1–positive cells were located primarily in the adventitia andmedia, consistent with previous reports.10 Macrophage infil-tration was more prominent in the male aortas, where ED-1–positive cell counts were 6.2�1.0 cells/high-powered field(HPF) compared with 0.54�0.02 cells/HPF in female aortas(P�0.003; please see www.ahajournals.org).
MMP-9 staining was also more evident in the media andadventitia of male aortas compared with female aortas, as
evidenced by immunohistochemistry (Figure 1b and 1c).Colocalization of ED-1 and MMP-9 demonstrated increasedcostaining of macrophages and MMP-9 in male aortas versusfemale aortas (Figure 1d and 1e). Similarly, male aortasexhibited increased MMP-9 expression by quantitative PCRcompared with female aortas (males, 0.39�0.09 versus0.003�0.001 MMP-9 mRNA copies; P�0.001; please seewww.ahajournals.org). Total MMP-9 activity by zymographywas 369% greater in male than female aortas (P�0.022;please see www.ahajournals.org).
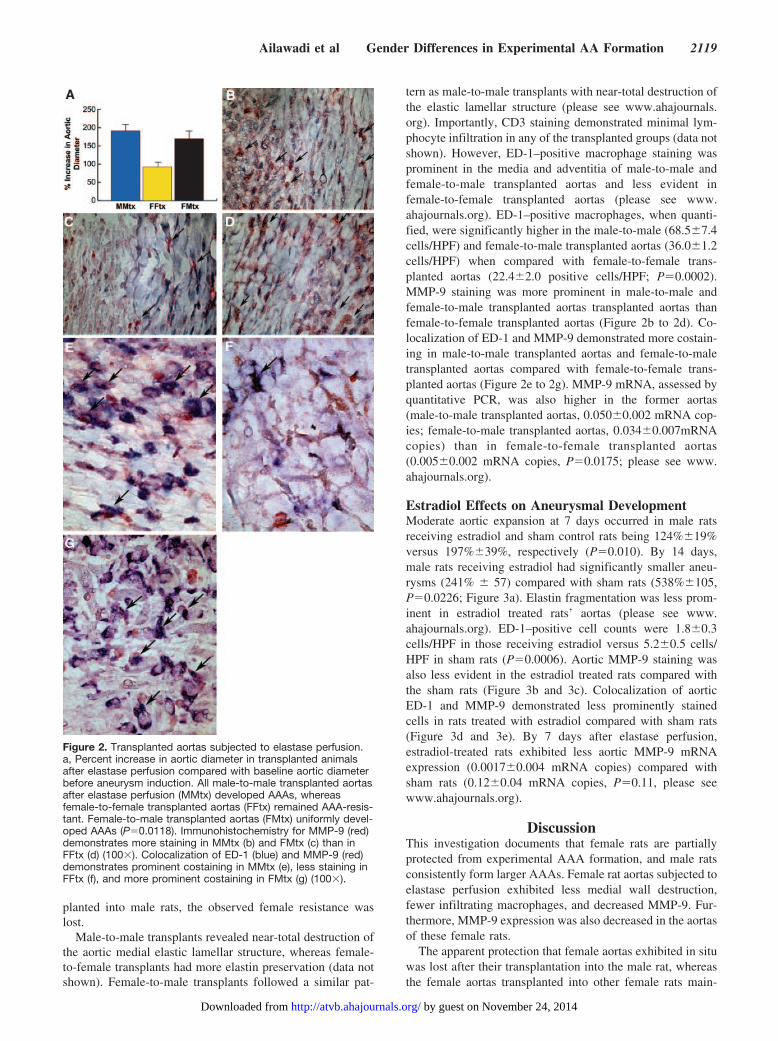
Aneurysm-Resistant Phenotype Is Lost AfterTransplantation of the Female Into the MaleAll male and female aortas, when transplanted into malerecipients and subsequently subjected to elastase perfusion,developed AAAs at 14 days, whereas only 17% of the femaleaortas transplanted into female recipients developed AAAs.Aortic dilations were similar among male-to-male andfemale-to-male transplanted aortas, but were significantlylower in the female-to-female transplanted aortas (male-to-male transplants, 189�22%; female-to-male transplants,197�34%; female-to-female transplants, 93�13%; P�0.0118;Figure 2a). Thus, although female aortas transplanted intofemale recipients remained resistant to AAA, when trans-
Figure 1. Male and female aortas 14 days after elastase perfu-sion. a, Percent increase in aortic diameter in male (M) andfemale (F) aortas 14 days after elastase perfusion comparedwith baseline aortic diameter. M had significantly larger AAAs(P�0.0234) and higher incidence of AAAs (P�0.0229) comparedwith F. MMP-9 immunohistochemical staining in M (b) and F (c)aorta after elastase perfusion (100�). Positive staining regions(arrows) are more prominent in M. Colocalization for ED-1 (blue)and MMP-9 (red) demonstrates more prominent costaining in M(d) and F (e) (100�). MMP-9 staining appeared associated withED-1–positive staining cells.
2118 Arterioscler Thromb Vasc Biol. November 2004
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from
planted into male rats, the observed female resistance waslost.
Male-to-male transplants revealed near-total destruction ofthe aortic medial elastic lamellar structure, whereas female-to-female transplants had more elastin preservation (data notshown). Female-to-male transplants followed a similar pat-
tern as male-to-male transplants with near-total destruction ofthe elastic lamellar structure (please see www.ahajournals.org). Importantly, CD3 staining demonstrated minimal lym-phocyte infiltration in any of the transplanted groups (data notshown). However, ED-1–positive macrophage staining wasprominent in the media and adventitia of male-to-male andfemale-to-male transplanted aortas and less evident infemale-to-female transplanted aortas (please see www.ahajournals.org). ED-1–positive macrophages, when quanti-fied, were significantly higher in the male-to-male (68.5�7.4cells/HPF) and female-to-male transplanted aortas (36.0�1.2cells/HPF) when compared with female-to-female trans-planted aortas (22.4�2.0 positive cells/HPF; P�0.0002).MMP-9 staining was more prominent in male-to-male andfemale-to-male transplanted aortas transplanted aortas thanfemale-to-female transplanted aortas (Figure 2b to 2d). Co-localization of ED-1 and MMP-9 demonstrated more costain-ing in male-to-male transplanted aortas and female-to-maletransplanted aortas compared with female-to-female trans-planted aortas (Figure 2e to 2g). MMP-9 mRNA, assessed byquantitative PCR, was also higher in the former aortas(male-to-male transplanted aortas, 0.050�0.002 mRNA cop-ies; female-to-male transplanted aortas, 0.034�0.007mRNAcopies) than in female-to-female transplanted aortas(0.005�0.002 mRNA copies, P�0.0175; please see www.ahajournals.org).
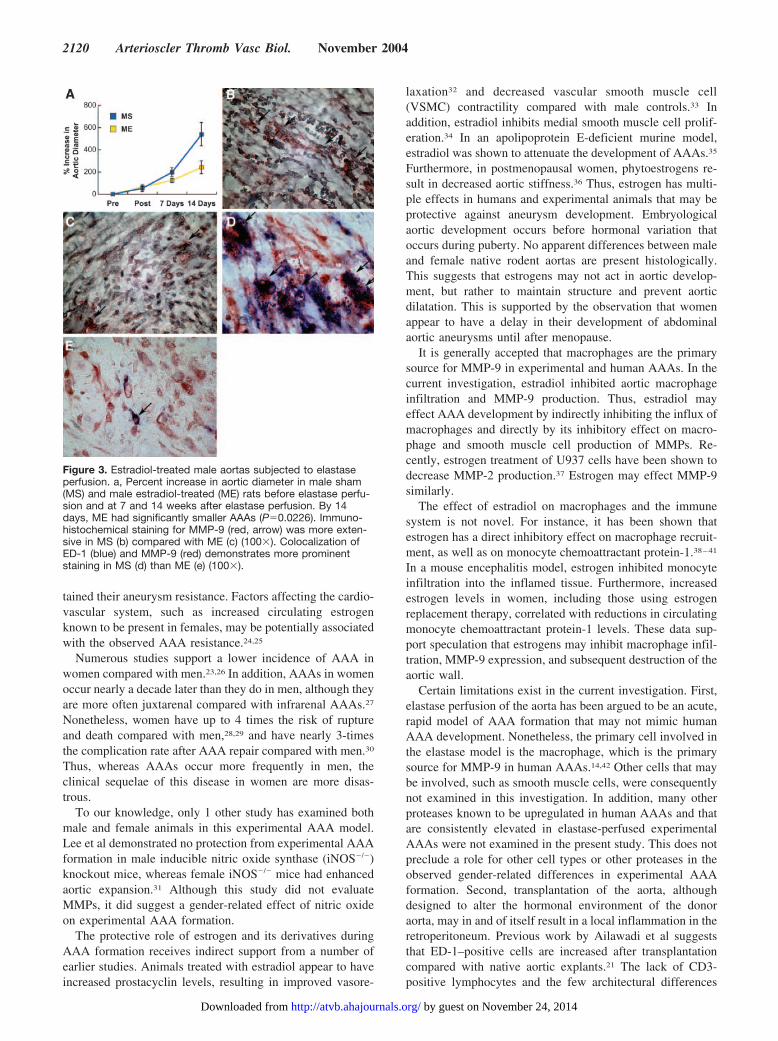
Estradiol Effects on Aneurysmal DevelopmentModerate aortic expansion at 7 days occurred in male ratsreceiving estradiol and sham control rats being 124%�19%versus 197%�39%, respectively (P�0.010). By 14 days,male rats receiving estradiol had significantly smaller aneu-rysms (241% � 57) compared with sham rats (538%�105,P�0.0226; Figure 3a). Elastin fragmentation was less prom-inent in estradiol treated rats’ aortas (please see www.ahajournals.org). ED-1–positive cell counts were 1.8�0.3cells/HPF in those receiving estradiol versus 5.2�0.5 cells/HPF in sham rats (P�0.0006). Aortic MMP-9 staining wasalso less evident in the estradiol treated rats compared withthe sham rats (Figure 3b and 3c). Colocalization of aorticED-1 and MMP-9 demonstrated less prominently stainedcells in rats treated with estradiol compared with sham rats(Figure 3d and 3e). By 7 days after elastase perfusion,estradiol-treated rats exhibited less aortic MMP-9 mRNAexpression (0.0017�0.004 mRNA copies) compared withsham rats (0.12�0.04 mRNA copies, P�0.11, please seewww.ahajournals.org).
DiscussionThis investigation documents that female rats are partiallyprotected from experimental AAA formation, and male ratsconsistently form larger AAAs. Female rat aortas subjected toelastase perfusion exhibited less medial wall destruction,fewer infiltrating macrophages, and decreased MMP-9. Fur-thermore, MMP-9 expression was also decreased in the aortasof these female rats.
The apparent protection that female aortas exhibited in situwas lost after their transplantation into the male rat, whereasthe female aortas transplanted into other female rats main-
Figure 2. Transplanted aortas subjected to elastase perfusion.a, Percent increase in aortic diameter in transplanted animalsafter elastase perfusion compared with baseline aortic diameterbefore aneurysm induction. All male-to-male transplanted aortasafter elastase perfusion (MMtx) developed AAAs, whereasfemale-to-female transplanted aortas (FFtx) remained AAA-resis-tant. Female-to-male transplanted aortas (FMtx) uniformly devel-oped AAAs (P�0.0118). Immunohistochemistry for MMP-9 (red)demonstrates more staining in MMtx (b) and FMtx (c) than inFFtx (d) (100�). Colocalization of ED-1 (blue) and MMP-9 (red)demonstrates prominent costaining in MMtx (e), less staining inFFtx (f), and more prominent costaining in FMtx (g) (100�).
Ailawadi et al Gender Differences in Experimental AA Formation 2119
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from
tained their aneurysm resistance. Factors affecting the cardio-vascular system, such as increased circulating estrogenknown to be present in females, may be potentially associatedwith the observed AAA resistance.24,25
Numerous studies support a lower incidence of AAA inwomen compared with men.23,26 In addition, AAAs in womenoccur nearly a decade later than they do in men, although theyare more often juxtarenal compared with infrarenal AAAs.27
Nonetheless, women have up to 4 times the risk of ruptureand death compared with men,28,29 and have nearly 3-timesthe complication rate after AAA repair compared with men.30
Thus, whereas AAAs occur more frequently in men, theclinical sequelae of this disease in women are more disas-trous.
To our knowledge, only 1 other study has examined bothmale and female animals in this experimental AAA model.Lee et al demonstrated no protection from experimental AAAformation in male inducible nitric oxide synthase (iNOS�/�)knockout mice, whereas female iNOS�/� mice had enhancedaortic expansion.31 Although this study did not evaluateMMPs, it did suggest a gender-related effect of nitric oxideon experimental AAA formation.
The protective role of estrogen and its derivatives duringAAA formation receives indirect support from a number ofearlier studies. Animals treated with estradiol appear to haveincreased prostacyclin levels, resulting in improved vasore-
laxation32 and decreased vascular smooth muscle cell(VSMC) contractility compared with male controls.33 Inaddition, estradiol inhibits medial smooth muscle cell prolif-eration.34 In an apolipoprotein E-deficient murine model,estradiol was shown to attenuate the development of AAAs.35
Furthermore, in postmenopausal women, phytoestrogens re-sult in decreased aortic stiffness.36 Thus, estrogen has multi-ple effects in humans and experimental animals that may beprotective against aneurysm development. Embryologicalaortic development occurs before hormonal variation thatoccurs during puberty. No apparent differences between maleand female native rodent aortas are present histologically.This suggests that estrogens may not act in aortic develop-ment, but rather to maintain structure and prevent aorticdilatation. This is supported by the observation that womenappear to have a delay in their development of abdominalaortic aneurysms until after menopause.
It is generally accepted that macrophages are the primarysource for MMP-9 in experimental and human AAAs. In thecurrent investigation, estradiol inhibited aortic macrophageinfiltration and MMP-9 production. Thus, estradiol mayeffect AAA development by indirectly inhibiting the influx ofmacrophages and directly by its inhibitory effect on macro-phage and smooth muscle cell production of MMPs. Re-cently, estrogen treatment of U937 cells have been shown todecrease MMP-2 production.37 Estrogen may effect MMP-9similarly.
The effect of estradiol on macrophages and the immunesystem is not novel. For instance, it has been shown thatestrogen has a direct inhibitory effect on macrophage recruit-ment, as well as on monocyte chemoattractant protein-1.38–41
In a mouse encephalitis model, estrogen inhibited monocyteinfiltration into the inflamed tissue. Furthermore, increasedestrogen levels in women, including those using estrogenreplacement therapy, correlated with reductions in circulatingmonocyte chemoattractant protein-1 levels. These data sup-port speculation that estrogens may inhibit macrophage infil-tration, MMP-9 expression, and subsequent destruction of theaortic wall.
Certain limitations exist in the current investigation. First,elastase perfusion of the aorta has been argued to be an acute,rapid model of AAA formation that may not mimic humanAAA development. Nonetheless, the primary cell involved inthe elastase model is the macrophage, which is the primarysource for MMP-9 in human AAAs.14,42 Other cells that maybe involved, such as smooth muscle cells, were consequentlynot examined in this investigation. In addition, many otherproteases known to be upregulated in human AAAs and thatare consistently elevated in elastase-perfused experimentalAAAs were not examined in the present study. This does notpreclude a role for other cell types or other proteases in theobserved gender-related differences in experimental AAAformation. Second, transplantation of the aorta, althoughdesigned to alter the hormonal environment of the donoraorta, may in and of itself result in a local inflammation in theretroperitoneum. Previous work by Ailawadi et al suggeststhat ED-1–positive cells are increased after transplantationcompared with native aortic explants.21 The lack of CD3-positive lymphocytes and the few architectural differences
Figure 3. Estradiol-treated male aortas subjected to elastaseperfusion. a, Percent increase in aortic diameter in male sham(MS) and male estradiol-treated (ME) rats before elastase perfu-sion and at 7 and 14 weeks after elastase perfusion. By 14days, ME had significantly smaller AAAs (P�0.0226). Immuno-histochemical staining for MMP-9 (red, arrow) was more exten-sive in MS (b) compared with ME (c) (100�). Colocalization ofED-1 (blue) and MMP-9 (red) demonstrates more prominentstaining in MS (d) than ME (e) (100�).
2120 Arterioscler Thromb Vasc Biol. November 2004
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from

other than those described does suggest that rejection is notinvolved in this process after transplantation or elastaseperfusion. Despite this lack of perceived differences, com-parisons between transplanted elastase-perfused aortas andnontransplanted elastase-perfused aortas cannot be made.Third, 2 different lots of elastase were used in the presentinvestigation and may have resulted in varied results. Thefirst lot was used for elastase perfusion of aortas in the firstand second group of experiments (intact and transplantedanimals), whereas the latter lot was used for the third group ofexperiments (those treated with estradiol). These elastase lotswere quite different, in as much as the nontransplanted andtransplanted elastase-perfused male aortas increased theiraortic diameter by �200%, whereas elastase-perfused maleaortas used in the estradiol-treatment experiments developedalmost 500% increases in their aortic diameter using adifferent lot of elastase. Such variation has been reported withdifferent lots of elastase despite uniform dose and activity.43
As a consequence, groups treated with different lots ofelastase should not be compared, with comparisons limitedonly to animals treated with the same lot of elastase.
Despite these limitations, this investigation supports thetheory that gender differences in experimental AAA forma-tion exist that may be related to estrogenic effects onmacrophages and MMPs. Gender differences in other celllines and proteases, as well as cytokines, must be better-evaluated to further completely characterize the disparitybetween men and women with regard to AAA formation. Theefficacy of estradiol will need to be better-delineated beforethe institution of this or related hormones as a potentialtherapeutic agent.
AcknowledgmentsFunded by an American College of Surgeons Resident ResearchScholarship and National Institutes of Health Training Grant inMolecular and Cellular Cardiology T32-HL07853-05 (to G.A.);Society of University Surgeons Junior Faculty Award (to P.K.H.);University of Michigan McKay Grant, National Institutes of HealthMentored Clinician Scientists Development Award (K08 HL67885-01), and the Lifeline Foundation Research Award (to G.R.U.), andJobst Vascular Research Fund (to P.K.H., J.C.S., and G.R.U.).
References1. National Hospital Discharge Survey Data. http://www.cdc.gov/nchs/data/
series/sr_13/sr13_153.pdf. Hyattsville, Md: National Center for HealthStatistics; Updated November, 2002.
2. Pleumeekers HJ, Hoes AW, van der DE, van Urk H, Hofman A, de JongPT, Grobbee DE. Aneurysms of the abdominal aorta in older adults. TheRotterdam Study. Am J Epidemiol. 1995;142:1291–1299.
3. Singh K, Bonaa KH, Jacobsen BK, Bjork L, Solberg S. Prevalence of andrisk factors for abdominal aortic aneurysms in a population-based study:The Tromso Study. Am J Epidemiol. 2001;154:236–244.
4. Bengtsson H, Sonesson B, Bergqvist D. Incidence and prevalence ofAAA. Ann NY Acad Sci. 1996;800:1–24.
5. La Vecchia C, Decarli A, Franceschi S, Gentile A, Negri E, Parazzini F.Menstrual and reproductive factors and the risk of myocardial infarctionin women under fifty-five years of age. Am J Obstet Gynecol. 1987;157:1108–1112.
6. Freestone T, Turner RJ, Coady A, Higman DJ, Greenhalgh RM, PowellJT. Inflammation and matrix metalloproteinases in the enlargingabdominal aortic aneurysm. Arterioscler Thromb Vasc Biol. 1995;15:1145–1151.
7. Tamarina NA, McMillan WD, Shively VP, Pearce WH. Expression ofmatrix metalloproteinases and their inhibitors in aneurysms and normalaorta. Surgery. 1997;122:264–271.
8. Thompson RW, Parks WC. Role of matrix metalloproteinases inabdominal aortic aneurysms. Ann NY Acad Sci. 1996;800:157–174.
9. Allaire E, Forough R, Clowes M, Starcher B, Clowes AW. Local over-expression of TIMP-1 prevents aortic aneurysm degeneration and rupturein a rat model. J Clin Invest. 1998;102:1413–1420.
10. Pyo R, Lee JK, Shipley JM, Curci JA, Mao D, Ziporin SJ, Ennis TL,Shapiro SD, Senior RM, Thompson RW. Targeted gene disruption ofmatrix metalloproteinase-9 (gelatinase B) suppresses development ofexperimental abdominal aortic aneurysms. J Clin Invest. 2000;105:1641–1649.
11. Curci JA, Liao S, Huffman MD, Shapiro SD, Thompson RW. Expressionand localization of macrophage elastase (matrix metalloproteinase-12) inabdominal aortic aneurysms. J Clin Invest. 1998;102:1900–1910.
12. Carrell TW, Burnand KG, Wells GM, Clements JM, Smith A.Stromelysin-1 (matrix metalloproteinase-3) and tissue inhibitor ofmetalloproteinase-3 are overexpressed in the wall of abdominal aorticaneurysms. Circulation. 2002;105:477–482.
13. Davis V, Persidskaia R, Baca-Regen L, Itoh Y, Nagase H, Persidsky Y,Ghorpade A, Baxter BT. Matrix metalloproteinase-2 production and itsbinding to the matrix are increased in abdominal aortic aneurysms. Arte-rioscler Thromb Vasc Biol. 1998;18:1625–1633.
14. Thompson RW, Holmes DR, Mertens RA, Liao S, Botney MD, MechamRP, Welgus HG, Parks WC. Production and localization of 92-kilodaltongelatinase in abdominal aortic aneurysms. An elastolytic metallopro-teinase expressed by aneurysm-infiltrating macrophages. J Clin Invest.1995;96:318–326.
15. McMillan WD, Pearce WH. Increased plasma levels ofmetalloproteinase-9 are associated with abdominal aortic aneurysms. JVasc Surg. 1999;29:122–127.
16. Longo GM, Xiong W, Greiner TC, Zhao Y, Fiotti N, Baxter BT. Matrixmetalloproteinases 2 and 9 work in concert to produce aortic aneurysms.J Clin Invest. 2002;110:625–632.
17. Anidjar S, Salzmann JL, Gentric D, Lagneau P, Camilleri JP, Michel JB.Elastase-induced experimental aneurysms in rats. Circulation. 1990;82:973–981.
18. Agmon Y, Khandheria BK, Meissner I, Schwartz GL, Sicks JD, FoughtAJ, O’Fallon WM, Wiebers DO, Tajik AJ. Is aortic dilatation an athero-sclerosis-related process? Clinical, laboratory, and transesophageal echo-cardiographic correlates of thoracic aortic dimensions in the populationwith implications for thoracic aortic aneurysm formation. J Am CollCardiol. 2003;42:1076–1083.
19. Shteinberg D, Halak M, Shapiro S, Kinarty A, Sobol E, Lahat N, KarmeliR. Abdominal aortic aneurysm and aortic occlusive disease: a comparisonof risk factors and inflammatory response. Eur J Vasc Endovasc Surg.2000;20:462–465.
20. Fischer GM, Swain ML. Influence of contraceptive and other sex steroidson aortic collagen and elastin. Exper Mol Pathol. 1980;33:15–24.
21. Ailawadi G, Knipp BS, Lu G, Roelofs KJ, Ford JW, Hannawa KK,Bishop K, Thanaporn P, Henke PK, Stanley JC, Upchurch GR, Jr. Anonintrinsic regional basis for increased infrarenal aortic MMP-9expression and activity. J Vasc Surg. 2003;37:1059–1066.
22. Brooks-Asplund EM, Shoukas AA, Kim SY, Burke SA, Berkowitz DE.Estrogen has opposing effects on vascular reactivity in obese, insulin-re-sistant male Zucker rats. J Appl Physiol. 2002;92:2035–2044.
23. Eagleton MJ, Peterson DA, Sullivan VV, Roelofs KJ, Ford JA, StanleyJC, Upchurch GR Jr. Nitric oxide inhibition increases aortic wall matrixmetalloproteinase-9 expression. J Surg Res. 2002;104:15–21.
24. Leinwand LA. Sex is a potent modifier of the cardiovascular system.J Clin Invest. 2003;112:302–307.
25. Mendelsohn ME, Karas RH. The protective effects of estrogen on thecardiovascular system. N Engl J Med. 1999;340:1801–1811.
26. Katz DJ, Stanley JC, Zelenock GB. Gender differences in abdominalaortic aneurysm prevalence, treatment, and outcome. J Vasc Surg. 1997;25:561–568.
27. Velazquez OC, Larson RA, Baum RA, Carpenter JP, Golden MA,Mitchell ME, Pyeron A, Barker CF, Fairman RM. Gender-related dif-ferences in infrarenal aortic aneurysm morphologic features: issuesrelevant to Ancure and Talent endografts. J Vasc Surg. 2001;33(suppl):84.
28. Brown PM, Zelt DT, Sobolev B. The risk of rupture in untreated aneu-rysms: the impact of size, gender, and expansion rate. J Vasc Surg.2003;37:280–284.
29. Dimick JB, Stanley JC, Axelrod DA, Kazmers A, Henke PK, Jacobs LA,Wakefield TW, Greenfield LJ, Upchurch GR Jr. Variation in death rate
Ailawadi et al Gender Differences in Experimental AA Formation 2121
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from

after abdominal aortic aneurysmectomy in the United States: impact ofhospital volume, gender, and age. Ann Surg. 2002;235:579–585.
30. Wolf YG, Arko FR, Hill BB, Olcott C, Harris EJ Jr, Fogarty TJ, ZarinsCK. Gender differences in endovascular abdominal aortic aneurysmrepair with the AneuRx stent graft. J Vasc Surg. 2002;35:882–886.
31. Lee JK, Borhani M, Ennis TL, Upchurch GR Jr, Thompson RW. Exper-imental abdominal aortic aneurysms in mice lacking expression ofinducible nitric oxide synthase. Arterioscler Thromb Vasc Biol. 2001;21:1393–1401.
32. Bolego C, Cignarella A, Ruzza R, Zaarour C, Messi E, Zanisi M, PuglisiL. Differential effects of low- and high-dose estrogen treatments onvascular responses in female rats. Life Sciences. 1997;60:2291–2302.
33. Murphy JG, Khalil RA. Gender-specific reduction in contractility and[Ca(2�)](i) in vascular smooth muscle cells of female rat. Am J PhysiolCell Physiol. 2000;278:C834–C844.
34. Sullivan TR Jr, Karas RH, Aronovitz M, Faller GT, Ziar JP, Smith JJ,O’Donnell TF Jr, Mendelsohn ME. Estrogen inhibits the response-to-injury in a mouse carotid artery model. J Clin Invest. 1995;96:2482–2488.
35. Martin-McNulty B, Tham DM, da C, V, Ho JJ, Wilson DW, Rutledge JC,Deng GG, Vergona R, Sullivan ME, Wang YX. 17 �-estradiol attenuatesdevelopment of angiotensin II-induced aortic abdominal aneurysm inapolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 2003;23:1627–1632.
36. van der Schouw YT, Pijpe A, Lebrun CE, Bots ML, Peeters PH, vanStaveren WA, Lamberts SW, Grobbee DE. Higher usual dietary intake of
phytoestrogens is associated with lower aortic stiffness in postmeno-pausal women. Arterioscler Thromb Vasc Biol. 2002;22:1316–1322.
37. Halpern V, Song X, Faust GR. Estrogen inhibits elastase secretion fromU937 cells via nitric oxide. Presented at the Lifeline Foundation ResearchForum, Society of Vascular Surgery. June 8, 2003.
38. Seli E, Pehlivan T, Selam B, Garcia-Velasco JA, Arici A. Estradioldown-regulates MCP-1 expression in human coronary artery endothelialcells. Fertil Steril. 2002;77:542–547.
39. Jilma B, Jilma-Stohlawetz P. Female gender, menstrual cycle andestradiol affect plasma levels of monocyte chemotactic protein-1(MCP-1) in humans. Cardiovasc Res. 2002;55:416.
40. Rodriguez E, Lopez R, Paez A, Masso F, Montano LF. 17�-estradiolinhibits the adhesion of leukocytes in TNF-� stimulated human endothe-lial cells by blocking IL-8 and MCP-1 secretion, but not its transcription.Life Sciences. 2002;71:2181–2193.
41. Yamada K, Hayashi T, Kuzuya M, Naito M, Asai K, Iguchi A. Physio-logical concentration of 17 �-estradiol inhibits chemotaxis of humanmonocytes in response to monocyte chemotactic protein 1. Artery. 1996;22:24–35.
42. Hibbs MS. Expression of 92 kDa phagocyte gelatinase by inflammatoryand connective tissue cells. Matrix Supplement. 1992;1:51–57.
43. Curci JA, Thompson RW. “Variable induction of experimental abdominalaortic aneurysms with different preparations of porcine pancreatic elas-tase”. J Vasc Surg. 1999;29:385.
2122 Arterioscler Thromb Vasc Biol. November 2004
by guest on November 24, 2014http://atvb.ahajournals.org/Downloaded from
Related Documents











