1 Incidence, diversity and molecular epidemiology of sapoviruses in swine across Europe 1 Running title: Porcine sapoviruses in Europe 2 Gábor Reuter, 1 * Janet Zimšek-Mijovski, 2 Mateja Poljšak-Prijatelj, 2 Ilaria Di Bartolo, 3 Franco 3 Maria Ruggeri, 3 Tuija Kantala, 4 Leena Maunula, 4 István Kiss, 5 Sándor Kecskeméti, 5 Nabil 4 Halaihel, 6 Javier Buesa, 7 Christina Johnsen, 8 Charlotte K Hjulsager, 9 Lars E Larsen, 9 Marion 5 Koopmans, 10 Blenda Böttiger 8 6 7 ÁNTSZ Regional Institute of State Public Health Service, Pécs, Hungary 1 8 University of Ljubljana, Ljubljana, Slovenia 2 9 Istituto Superiore di Sanita, Rome, Italy 3 10 University of Helsinki, Helsinki, Finland 4 11 Central Agricultural Office, Veterinary Diagnostic Directorate, Debrecen, Hungary 5 12 University of Zaragoza, Zaragoza, Spain 6 13 University of Valencia, Valencia, Spain 7 14 Statens Serum Institut, Copenhagen, Denmark 8 15 Technical University of Denmark, National Veterinary Institute, Copenhagen, Denmark 9 16 National Institute for Public Health and the Environment, Bilthoven, The Netherlands 10 17 * Address for correspondence: 18 Dr Gábor Reuter 19 Regional Laboratory of Virology, 20 ÁNTSZ Regional Institute of State Public Health Service 21 Szabadság út 7. H-7623 Pécs, Hungary 22 Telephone: +36 06 (72) 514 979 23 Fax: +36 06 (72) 514 949 24 Email: [email protected] 25 Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. J. Clin. Microbiol. doi:10.1128/JCM.01279-09 JCM Accepts, published online ahead of print on 25 November 2009 on June 26, 2018 by guest http://jcm.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
Incidence, diversity and molecular epidemiology of sapoviruses in swine across Europe 1
Running title: Porcine sapoviruses in Europe 2
Gábor Reuter,1* Janet Zimšek-Mijovski,2 Mateja Poljšak-Prijatelj,2 Ilaria Di Bartolo,3 Franco 3
Maria Ruggeri,3 Tuija Kantala,4 Leena Maunula,4 István Kiss,5 Sándor Kecskeméti,5 Nabil 4
Halaihel,6 Javier Buesa,7 Christina Johnsen,8 Charlotte K Hjulsager,9 Lars E Larsen,9 Marion 5
Koopmans,10 Blenda Böttiger8 6
7
ÁNTSZ Regional Institute of State Public Health Service, Pécs, Hungary1 8
University of Ljubljana, Ljubljana, Slovenia2 9
Istituto Superiore di Sanita, Rome, Italy3 10
University of Helsinki, Helsinki, Finland4 11
Central Agricultural Office, Veterinary Diagnostic Directorate, Debrecen, Hungary5 12
University of Zaragoza, Zaragoza, Spain6 13
University of Valencia, Valencia, Spain7 14
Statens Serum Institut, Copenhagen, Denmark8 15
Technical University of Denmark, National Veterinary Institute, Copenhagen, Denmark9 16
National Institute for Public Health and the Environment, Bilthoven, The Netherlands10 17
* Address for correspondence: 18
Dr Gábor Reuter 19
Regional Laboratory of Virology, 20
ÁNTSZ Regional Institute of State Public Health Service 21
Szabadság út 7. H-7623 Pécs, Hungary 22
Telephone: +36 06 (72) 514 979 23
Fax: +36 06 (72) 514 949 24
Email: [email protected] 25
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Clin. Microbiol. doi:10.1128/JCM.01279-09 JCM Accepts, published online ahead of print on 25 November 2009
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

2
Abstract 26
Porcine sapovirus is an enteric calicivirus in domestic pigs belonging to the family 27
Caliciviridae. Some porcine sapoviruses are genetically related to human caliciviruses, which 28
has raised public health concerns of animal reservoirs and potential cross-species transmission 29
for sapoviruses. In this comprehensive study we report the incidence, genetic diversity and 30
molecular epidemiology of sapoviruses detected in domestic pigs in six European countries, 31
Denmark, Finland, Hungary, Italy, Slovenia and Spain, between 2004 and 2007. A total of 32
1050 swine fecal samples from 88 pig farms were collected and tested by reverse 33
transcription-polymerase chain reaction for sapoviruses, and positive findings were confirmed 34
by sequencing. Sapoviruses were detected in 80 (7.6%) samples collected on 39 (44.3%) 35
farms, and in every country. The highest prevalence was seen among piglets aged 2-8 weeks 36
and – where tested (Spain and Denmark) - there was no significant difference in the 37
proportion of sapovirus positive findings in healthy animals or animals with diarrhea. Based 38
upon the RNA-polymerase region highly heterogeneous populations of viruses were identified 39
representing 6 different genogroups (III, VI, VII, and VIII) including potential new 40
genogroups (IX and X) with a predominance of GIII (50.6%). Genogroup VIII, found in 5 of 41
the 6 countries, had the highest homology (up to 66% at amino acid level) to human sapovirus 42
strains. Sapoviruses are commonly circulating and endemic agents in swine herds Europe-43
wide. Highly heterogeneous, and potential new genogroups of sapoviruses were found in pigs, 44
however, any “human-like” sapovirus was not detected. 45
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

3
Introduction 46
Caliciviruses (family Caliciviridae) are small, non-enveloped viruses with single-47
stranded, positive-sense genomic RNA which are classified – at present – into five genera: 48
Vesivirus, Lagovirus, Norovirus, Nebovirus and Sapovirus (14, 49
http://talk.ictvonline.org/media/p/1203.aspx). Viruses within one calicivirus genus are 50
phylogenetically related, and have the same genomic organization (5). The sapovirus genome 51
is 7.3 to 7.5 kb in length and contains two main open reading frames (ORFs). Sapoviruses are 52
important enteric pathogens that can cause diarrhea in humans, pigs and mink (5, 6, 16). 53
Based upon phylogenetic clustering of the capsid gene (ORF1) and protein sequences, 54
sapoviruses have been classified into five distinct genogroups (GI to GV) (3). Human 55
sapoviruses belong to GI, GII, GIV and GV, whereas porcine sapovirus belongs to GIII. 56
Recently, new porcine sapovirus genogroups (GVI, GVII, GVIII) were proposed (15, 25, 27). 57
Recombinant sapoviruses - both in human and swine hosts - have also been described (8, 24). 58
Each genogroup can be further divided into genetically diverse genotypes. 59
Porcine sapovirus (historically called porcine enteric calicivirus – PEC) was first 60
identified in the United States by electron microscopy in 1980 (21) and genetically 61
characterized as a sapovirus in 1999 (5, 24). Porcine sapoviruses have been reported in only a 62
few additional countries: in The Netherlands (van der Heide et al., available only in the 63
GenBank database), in South Korea (11), Venezuela (17), Hungary (20) and recently in Italy 64
(16), Brazil (1) and Canada (13). The porcine sapovirus strains Cowden and LL14/02/US, 65
representatives of sapovirus GIII, were detected in diarrheic piglets and have been shown to 66
induce enteric diseases and lesions in experimentally infected pigs (4, 7). 67
A public health concern of potential cross-species transmission and animal reservoirs 68
for sapoviruses has been raised. In this study, we report the incidence, genetic diversity and 69
molecular epidemiology of sapoviruses detected in domestic pigs in six European countries. 70
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

4
Materials and Methods 71
Stool samples. 72
A total of 1050 fecal samples from domestic pigs (Sus scrofa domestica) collected at 73
88 swine farms in six European countries (Denmark, Finland, Hungary, Italy, Slovenia and 74
Spain) from January 2004 to December 2007 were tested for sapoviruses (Table 1). Detailed 75
information of sampling periods, age groups, number of tested farms and animals per country 76
is summarized in Table 1. Farms and sampled animals were randomly selected. Clinically 77
healthy animals, without signs of gastroenteritis, were tested in Finland, Hungary, Italy and 78
Slovenia; however, both healthy and diarrheic animals were tested in Denmark and Spain. 79
Fresh fecal samples were placed into sterile containers and stored frozen at -20ºC until tested. 80
RNA extraction and RT-PCR. 81
The RNA was extracted from fecal suspensions according to the standard RNA 82
extraction methods used by each laboratory. RT-PCR was also performed by the participating 83
laboratories using the generic calicivirus primer pair p290/p289 (10), a degenerated version of 84
these primers p290D/p289D (3) or a specific sapovirus primer pair SR80/JV33 (23), all 85
designed for the RNA-dependent RNA polymerase region (Table 1). PCR-products of primer-86
pair p289/p290, p290D/p289D and SR80/JV33 are 331-nt (95-aa), 331-nt and 320-nt for 87
sapovirus, respectively. 88
Sequence and phylogenetic analysis. 89
PCR-products were sequenced directly in both directions using the PCR primers. 90
Sequences were analyzed using the GeneDoc version 2.6 program (19). A dendrogram was 91
constructed using the UPGMA method by MEGA version 3.1 (12). Maintaining the 92
continuity of the current nomenclature for sapovirus genogrouping, we used the previously 93
proposed taxonomic names (25). 94
Statistics. 95
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

5
To test the differences in proportions of double and single infections in animals with 96
diarrhea, Fischer’s exact test was used, and a single-tailed p-value of < 0.5 was considered a 97
significant difference. The association between the incidence of sapovirus infection and 98
diarrhea was tested by chi square analysis. 99
Nucleotide sequence accession numbers. 100
Sapovirus sequences from pigs were submitted – in batch sets for each country - to 101
GenBank under accession numbers FJ854507-FJ854541 (Denmark); FJ861075-FJ861076 and 102
FJ866501-FJ866503 (Finland); DQ383274, FJ808729-FJ808731 (Hungary); GQ228085-103
GQ228090 (Italy); FJ715777-FJ715805 (Slovenia); FN397821 (Spain). 104
Results 105
A total of 117 (11.1%) samples from 1050 pigs yielded a PCR fragment with the 106
expected size for sapoviruses following RT-PCR and gel-electrophoresis. Two thirds (N=80) 107
of these (68% of gel-positives; 7.6% of samples tested) was confirmed as sapovirus specific 108
by sequence analysis of the PCR-products (Table 1). The low amount of DNA obtained for 109
some samples did not allow performing sequence reactions for all samples. In addition, 110
bacterial and host DNA sequences with expected size (false positive) were also amplified 111
using p289/p290 primers. Double infections were detected in 7 pigs increasing the total 112
number of sapovirus sequences available for study to 87. All these sequences contained the 113
conserved amino acid motif GLPSG of the calicivirus RNA-dependent RNA polymerase. The 114
incidence of the sapovirus-positive samples ranged between 1.6% (Hungary) and 49.1% 115
(Denmark). Samples from 39 (44.3%) of the 88 farms included were sapovirus PCR-positive, 116
ranging from 7% in Spain to 87.5% in Slovenia. 117
The number of the fecal samples positive by age-group and the incidence of sapovirus-118
positive animals by age-groups were summarized in Table 1. The sapovirus incidence ranged 119
between 0% and 66.7% within each age group (Table 1). In Finland, Hungary, Italy and 120
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

6
Slovenia all samples – both sapovirus positive and negative - were obtained from clinically 121
healthy pigs, while in Denmark and Spain samples from both, with and without diarrhea, were 122
tested. In Denmark, sapoviruses were detected in 14/30 (47%) and in 14/27 (52%) samples 123
from animals with and without diarrhea, respectively. The corresponding proportions in Spain 124
were 13/113 (12%) and in 14/108 (13%), respectively. No significant differences in the 125
proportion of sapovirus positive samples among pigs with or without diarrhea were seen. 126
A total of 81 out of 87 sapovirus polymerase sequences could be used for further 127
analysis: 35 (43.2%) sequences from Denmark, 5 (6.2%) from Finland, 7 (8.6%) from 128
Hungary, 5 (6.2%) from Italy, 28 (34.6%) from Slovenia and 1 (1.2%) from Spain. The 129
phylogenetic analysis divided the sapovirus sequences into six different genogroups (Figure 130
1). Most sapoviruses belonged to the genogroup GIII (N=41; 50.6%) but GVI (N=1; 1.2%), 131
GVII (N=11; 13.6%) and GVIII (N=6; 7.4%) were also confirmed. Furthermore, strains 132
belonging to two potentially new sapovirus genogroups, tentatively classified as GIX (N=8; 133
9.9%) and GX (N=14; 17.3%) were also detected (Figure 1). By phylogenetic analysis, two 134
lineages (A and B) of GIII strains were found (Figure 1). All six genogroups were identified 135
in Denmark. Three genogroups, GIII, GVIII and GX were found in at least four of the 136
countries studied, whereas GVI, GVII and GIX were detected only in one or two countries. 137
For comparison of sapovirus strains found in pigs and in humans, we compared 138
nucleotide and amino acid sequence identities of the RNA-polymerase gene between the 139
study strains in pigs and the prototype sapovirus genogroups in humans (Table 2). The highest 140
sequence identities (up to 66% in amino acid) were found between genogroup GVIII and all 141
human sapovirus genogroups. GVIII strains in pigs are related to each other with 63-87% 142
nucleotide and 76-98% amino acid identity, respectively. The lowest sequence identity was 143
seen between genogroup IX and human sapoviruses. Between the two GIII lineages, GIIIA 144
and GIIIB, the nucleotide and amino acid identities were 68-84% and 75-91%, respectively. 145
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

7
The distribution of sapovirus genogroups over time (figure not shown) and by age 146
groups (Figure 2) was further analysed. In the first 2 years of the study, genogroup GIII 147
strains were detected most commonly (86%), but this data was from Hungary and Slovenia 148
only, whereas strains belonging to six genogroups (GIIIA and B, GVI, GVII, GVIII, GIX and 149
GX) were found in 5 countries in the last two years (2006 and 2007). Clustering of genotypes 150
by age showed that the dominant GIII strains were detected primarily in young pigs (78%, 151
49% and 0% of strains from pigs in age groups less than 1 month, 1-3 months and more than 152
3 months, respectively). GVI and GVI genotype strains were identified only in age group 1-3 153
months. 154
Double infections with two sapoviruses from different genogroups were identified in 7 155
animals (Vet4 and Vet4B, GVII/GIX; VET19 and Vet19B, GVIII/GX; Vet21A and Vet21B, 156
GIII/GVII; Vet23 and Vet23B, GIII/GVII; Vet25 and Vet25B, GIX/GIII; Vet31 and Vet31B, 157
GVII/GX, Vet33A and Vet33B, GIX/GX) under age of 12 weeks. The different virus types 158
were detected by the use of different diagnostic primer pairs. Six of the seven animals with a 159
double infection had diarrhoea, which was significantly more than seen in the 21 animals 160
infected with a single sapovirus and where only 8 animals had diarrhoea in Denmark. 161
Furthermore, identical sapovirus sequences were detected within the same herds in 6 animal 162
pairs in Denmark (N=4) and Hungary (N=2). 163
Discussion 164
Only few studies have been done to investigate the molecular epidemiology of 165
sapoviruses in swine, since the discovery of porcine sapovirus in 1980 (21). In this study, we 166
describe a large, integrated analysis related to the incidence, genetic diversity and molecular 167
epidemiology of porcine sapoviruses in six European countries. 168
Based upon our data, approximately 8% of tested animals were infected with 169
sapoviruses and shedding the virus in the feces, and sapoviruses were present in more than 170
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

8
40% of the selected swine farms in Europe, indicating that sapoviruses are circulating as 171
endemic agents in swine herds. However, the variation of sapoviruses incidence in individual 172
pigs, as well as among farms, in the participating countries was considerable (data not shown). 173
This could be a reflection of the different possibilities to obtain samples in each of the 174
participating countries. The overall sapovirus positivity rate in our study is lower than in some 175
other studies showing a prevalence of sapoviruses of at least 62% in 621 pigs from 7 different 176
farms in the USA (26), of 30% among 113 piglets from 34 farms in Brazil (1), and of 23% in 177
53 post-weaning pigs from 5 farms in Korea (28), but considerable ranges have been 178
published as well (25). The primers used and selection of samples tested regarding age and 179
state of health differed also in these studies making direct comparisons between them difficult, 180
but it can be concluded that sapovirus infections among pigs are common events. However, 181
our data confirms that pigs are infected with sapoviruses early in life periods in line with the 182
studies in Venezuela, Brazil, Korea and USA (17, 1, 9, 26). As also described in the papers 183
from Venezuela, Brazil and Belgium (18) we found sapoviruses in nearly equal numbers in 184
pigs with and without diarrhea as well. However, diarrhea was significantly associated with 185
co-infection with different sapovirus strains. This does not necessarily imply that the double 186
infection itself causes diarrhea, but might reflect lower hygienic standard in the pens of these 187
pigs, and other pathogens could therefore also be present. 188
It should be also noted that double infections were detected only with using different 189
primer-sets indicating that there is a diagnostic gap in sapovirus detection. Against this, 190
extremely diverse groups of sapoviruses were detected among swine in this study. Porcine 191
strains clustered into a total of 4 known and 2 - previously unknown - potential new 192
genogroups (GIX and GX). On the other hand, genotype III strains were detected most 193
frequently. Their predominance in a limited time period and in young animals suggests that 194
the pigs were sampled during a GIII epidemic, but coordinated studies across regions or 195
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

9
countries are needed to better understand the dynamics of genogroup(s) changes and the 196
possibilities of drift and international spread of sapoviruses among pigs. 197
Recently, it was shown that in some cases there was no sharp demarcation of host-198
species for some calicivirus infections; some caliciviruses may have zoonotic potential and 199
animals such as domestic pigs may act as a reservoir for norovirus or vesivirus (24, 2, 22). By 200
phylogenetic analysis, a higher genetic diversity was seen among porcine sapoviruses than 201
within the known human sapoviruses indicating a longer co-evolution of sapoviruses in swine. 202
One sapovirus genogroup, GVIII, occupy a special position among sapoviruses. GVIII is 203
genetically more closely related to human sapoviruses (especially GV and GI) than to other 204
sapovirus genogroups in swine (25, 16). A previous study suggested that this strain circulates 205
infrequently and has low levels of virus copies in the feces of pigs and therefore probably 206
originates from a non-porcine host (25). Interestingly, in our study, these unique sapovirus 207
strains were found in 5 of the 6 countries constituting 7.4% of the positive sapovirus samples. 208
This indicates a Europe-wide circulation and a relatively frequent incidence of this strain in 209
pigs. Until now, GVIII or GVIII-like strains have not been detected in humans. On the other 210
hand, among the known sapoviruses in swine, the GVIII strains have the closest evolutionary 211
cross-point between human and porcine sapoviruses, as shown by sequence and phylogenetic 212
analysis. 213
Based on this comprehensive study no evidence for zoonotic sapovirus could be found 214
in pigs, but a surprisingly diverse profile of sapoviruses was observed in European swine 215
herds. While the clinical relevance remains to be determined, it reflects how little we know 216
about the viral pathogens present in these food animals. Knowledge about their potential for 217
spread, including contribution to recombination events as was described for sapoviruses 218
belonging to different genogroups is important to understand the role of swine as a potential 219
reservoir. 220
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

10
Acknowledgements 221
This work was supported by grants from the Hungarian Scientific Research Fund 222
(OTKA, F048433) and the project „Enteric Virus Emergence, New Tools” (EVENT, SP22-223
CT-2004-502571) funded by the European Union. 224
References 225
1. Barry, A.F., A.F. Alfieri, and A.A. Alfieri. 2008. High genetic diversity in RdRp gene 226
of Brazilian porcine sapovirus strains. Vet. Microbiol. 131:185-191. 227
2. Farkas, T., S. Nakajima, M. Sugieda, X. Deng, W. Zhong, and X. Jiang. 2005. 228
Seroprevalence of noroviruses in swine. J. Clin. Microbiol.; 43:657-661. 229
3. Farkas, T., W. M. Zhong, Y. Jing, P.W. Huang, S.M. Espinosa, N. Martinez, A.L. 230
Morrow, G.M. Ruiz-Palacios, L.K. Pickering, and X. Jiang. 2004. Genetic diversity 231
among sapovirus. Arch. Virol. 149:1309-1323. 232
4. Flynn, W. T., L. J. Saif, and P. D. Moorhead. 1988. Pathogenesis of porcine enteric 233
calicivirus-like virus in four-day-old gnotobiotic pigs. Am. J. Vet. Res. 49:819-825. 234
5. Guo, M., K.O. Chang, M. E. Hardy, Q. Zhang, A. V. Parwani, and L. J. Saif. 1999. 235
Molecular characterization of a porcine enteric calicivirus genetically related to Sapporo-236
like human caliciviruses. J. Virol. 73:9625-9631. 237
6. Guo, M., J. F. Evermann, and L. J. Saif. 2001. Detection and molecular 238
characterization of cultivable caliciviruses from clinically normal mink and enteric 239
caliciviruses associated with diarrhea in mink. Arch Virol. 146:479-493. 240
7. Guo, M. J., K. O. Hayes, A. Cho, V. Parwani, L. M. Lucas, and L. J. Saif. 2001. 241
Comparative pathogenesis of tissue culture-adapted and wild-type Cowden porcine 242
enteric calicivirus (PEC) in gnotobiotic pigs and induction of diarrhea by intravenous 243
inoculation of wild-type PEC. J. Virol. 75:9239-9251. 244
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

11
8. Hansman, G. S., N. Takeda, T. Oka, M. Oseto, K. O. Hedlund, and K. Katayama. 245
2005. Intergenogroup recombination in sapoviruses. Emerg. Infect. Dis. 11:1916-1920. 246
9. Jeong, C., S. I. Park, S. H. Park, H. H. Kim, S. J. Park, J. H. Jeong, H. E. Choy, L. J. 247
Saif, S. K. Kim, M. I. Kang, B. H. Hyun, and K. O. Cho. 2007. Genetic diversity of 248
porcine sapoviruses. Vet. Microbiol. 122:246-257. 249
10. Jiang, X., P. W. Huang, W. M. Zhong, T. Farkas, D. W. Cubitt, and D. O. Matson. 250
1999. Design and evaluation of a primer pair that detects both Norwalk- and Sapporo-like 251
caliciviruses by RT-PCR. J. Virol. Methods 83:145-154. 252
11. Kim, H. J., H. S. Cho, K. O. Cho, and N. Y. Park. 2006. Detection and molecular 253
characterization of porcine enteric calicivirus in Korea, genetically related to sapoviruses. 254
J. Vet. Med. B 53:155-159. 255
12. Kumar, S., K. Tamura, I. B. Jakobsen, and M. Nei. 2001. MEGA2: Molecular 256
Evolutionary Genetics Analysis software. Bioinformatics 17:1244-1245. 257
13. L’Homme, Y., R. Sanregret, E. Plante-Fortier, A. M. Lamontagne, G. Lacroix, M. 258
Ouardani, J. Deschamps, G. Simard, and C. Simard. 2009. Genetic diversity of 259
porcine norovirus and sapovirus: Canada, 2005-2007. Arch. Virol. 154:581-93. 260
14. Mayo MA. Virus Taxonomy – Houston 2002. 2002. Arch. Virol. 147:1071-1076. 261
15. Martella, V., K. Bányai, E. Lorusso, A. L. Bellacicco, N. Decaro, V. Mari, L. Saif, V. 262
Costantini, S. De Grazia, G. Pezzotti, A. Lavazza, and C. Buonavoglia. 2008. Genetic 263
heterogeneity of porcine enteric caliciviruses identified from diarrhoeic piglets. Virus 264
Genes 36:365-373. 265
16. Martella, V., E. Lorusso, K. Banyai, N. Decaro, M. Corrente, G. Elia, A. Cavalli, A. 266
Radogna, V. Constantini, L. J. Saif, A. Lavazza, L. Di Trani, and C. Bounavoglia. 267
2008. Identification of a porcine calicivirus related genetically to human sapoviruses. J 268
Clin. Microbiol. 46:1907-1913. 269
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

12
17. Martinez, M. A., A. C. Alcala, G. Carruyo, L. Botero, F. Liprandi, and J. E. Ludert. 270
2006. Molecular detection of porcine enteric caliciviruses in Venezuelan farms. Vet. 271
Microbiol. 116:77-84. 272
18. Mauroy, A., A. Scipioni, E. Mathijs, C. Miry, D. Ziant, C. Thys, and E. Thiry. 273
2008. Noroviruses and sapoviruses in pigs in Belgium. Arch. Virol. 153:1927-1931. 274
19. Nicholas, K. B., and H. B. Nicholas, Jr. 1997. GeneDoc: a tool for editing and 275
annotating multiple sequence alignments. Distributed by the author. 276
http://www.psc.edu/biomed/genedoc 277
20. Reuter, G., H. Bíró, and Gy. Szőcs. 2007. Enteric caliciviruses in domestic pigs in 278
Hungary. Arch. Virol. 152:611-614. 279
21. Saif, L. J., E. H. Bohl, K. W. Theil, R. F. Cross, and J. A. House. 1980. Rotavirus-like, 280
calicivirus-like, and 23-nm virus-like particles associated with diarrhea in young pigs. J. 281
Clin. Microbiol. 12:105-111. 282
22. Smith, A. W., P. L. Iversen, D. E. Skilling, D. A. Stein, K. Bok, and D. O. Matson. 283
2006. Vesivirus viremia and seroprevalence in humans. J. Med. Virol. 78:693-701. 284
23. Vinjé, J., H. Deijl, R. van der Heide, D. C. Lewis, K-O. Hedlund, L. Svensson, and 285
M. P. G. Koopmans. 2000. Molecular detection and epidemiology of “Sapporo-like 286
viruses”. J. Clin. Microbiol. 38:530-536. 287
24. Wang, Q-H., M. G. Han, S. Cheetham, M. Souza, J. A. Funk, and L. J. Saif. 2005. 288
Porcine noroviruses related to human noroviruses. Emerg. Infect. Dis. 12:1874-1881. 289
25. Wang, Q-H., M. G. Han, J. A. Funk, G. Bowman, D. A. Janies, and L. J. Saif. 290
2005. Genetic diversity and recombination of porcine sapoviruses. J. Clin. Microbiol. 291
43:5963-5972. 292
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

13
26. Wang, Q-H., M. Souza, J. A. Funk, W. Zhang, and L. J. Saif. 2006. Prevalence of 293
noroviruses and sapoviruses in swine of various ages determined by reverse transcription-294
PCR and microwell hybridization assays. J. Clin. Microbiol. 44:2057-2062. 295
27. Yin, Y., Y. Tohya, Y. Ogawa, D. Numazawa, K. Kato, and H. Akashi. 2006. Genetic 296
analysis of calicivirus genomes detected in intestinal contents of piglets in Japan. Arch. 297
Virol. 151:1749-1759. 298
28. Yu, J. N., M. Y. Kim, D. G. Kim, S. E. Kim, J. B. Lee, S. Y. Park, C. S. Song, H. V. 299
Shin, K. H. Seo, and I. S. Choi. 2008. Prevalence of hepatitis E virus and sapovirus in 300
post-weaning pigs and identification of their genetic diversity. Arch. Virol. 153:739-742.301
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

14
Figure legends 302
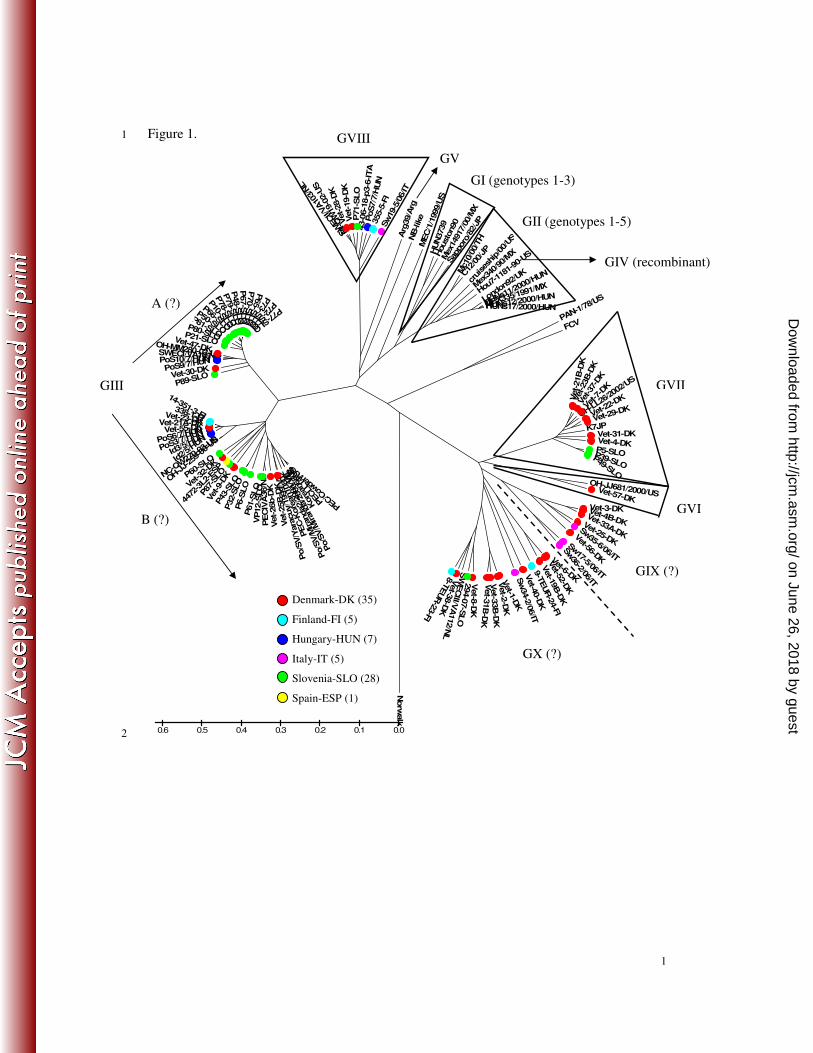
Figure 1. Phylogenetic analysis of sapoviruses including porcine sapoviruses detected in this 303
study in Europe based upon the 286-nt long RNA-dependent RNA polymerase region of 304
caliciviruses. A dendrogram was constructed using the UPGMA method by MEGA (version 305
3.1). The country-origin of the strains is indicated by colour coding (Denmark: red, Finland: 306
light-blue, Hungary: blue, Italy: purple, Slovenia: green, Spain: yellow). To maintain the 307
continuity of the current nomenclature for sapovirus genogrouping we used the previously 308
proposed taxonomic names as a basis (Wang et al., 2005, Martella et al., 2008). Sapoviruses 309
in GI, GII, GIV and GV are detected in humans. Swine-origin sapoviruses belongs to GIII, 310
GVI, GVII, GVIII, GIX and GX. Reference strains were obtained from GenBank: 311
PEC/Cowden/US (AF182760), PECLL14/US (AY425671), PECIVA36/NL (AY615805), 312
Korean6802 (AY289186), London/92/UK (U95645), HUNs11/2000/HUN (AF488717), 313
HUNs12/2000/HUN (AF488718), HUNs17/2000/HUN (AF488720), HUN3739/2008/HUN 314
(FJ844411), Sapporo/82/JP (U77903), Houston/90/US (U95644), Po/SV/Yaracuy/1999/VE 315
(AY633966), Po/SV/Miranda/2000/VE (AY633963), Po/SV/Miranda2/2001/VE (AY633965), 316
PEC-Korean10802 (AY289188), OH-JJ259-00-US (AY826423), NC-QW270-03-US 317
(AY826426), SWECI/VA10/NL (AY615807), OH-MM280-03-US (AY823308), 318
MEX335/1991/MX (AY157869), PAN-1/78/US (AF091736), 43-06-18-p-3-6-ITA 319
(AB221477), SWECIII/VA112/NL (AY615814), OH-LL26/2002/US (AY974195), OH-320
JJ681/2000/US (AY974192), Norwalk (M87661), Mex14917/00/MX (AF435810), 321
Mex340/90/MX (AF435809), cruiseship/00/US (AY157863), Mc10/00/TH (AY237420), 322
C12/00/JP (AY603425), Hou7-1181-90-US (AF435814), Arg39/Arg (AF405715), 323
MEC/1/1999/US (AF338404), FCV (M86379), NB-like (AY082891), K7JP (AB221130), 324
SWECII/VA103/NL (AY615811), MI-QW19-02-US (AY826424). 325
326
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

15
Figure 2. Prevalence of sapovirus genogroups by age group based upon 78 sapovirus strains. 327
Y axis represents the prevalence of genogroups in percentage (%). Count numbers in each bar 328
slice represents sample size for genogroup. Total sample sizes per each age group are at the 329
top of the bars. 330
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

1
Table 1. Incidence of sapoviruses in fecal samples collected from domestic pigs from 88 randomly selected swine farms in six European 1
countries between January 2004 and December 2007. Information of sampling periods, age groups, primers, number of tested and positive farms 2
and animals per country are shown. Samples from healthy animals were collected in Finland, Hungary, Italy, and Slovenia, whereas samples 3
from both healthy and diarrheic animals were collected in Denmark and Spain. 4
5
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

1
Table 2. Nucleotide and amino acid sequence identities (range for all strains sequenced) based 1
upon the RNA-polymerase region between the study strains detected in pigs (rows) and the 2
sapovirus genogroup prototype of human origin (columns). Bold numbers indicate the highest 3
nucleotide and amino acid identities. 4
5
human
swinea
GI
Sapporo/82/JP
(U77903)
GII
London/92/UK
(U95645)
GIV
Hou-7-1181-90-US
(AF435814)
GV
Arg39/Arg
(AF405715)
51-55%
51-55%
52-57%
50-54%
54-58%
50-54%
52-56%
49-52%
GIII A (N=20)b
nt
aa
GIII B (N=21)
nt
aa
50-59%
52-62%
54-59%
50-60%
55-60%
53-57%
53-55%
46-49%
GVI (N=1)
nt
aa
49%
41%
50%
42%
52%
43%
45%
32%
GVII (N=11)
nt
aa
42-46%
36-38%
44-48%
35-41%
44-47%
36-42%
41-45%
33-36%
GVIII (N=6)
nt
aa
50-62%
52-63%
53-64%
53-65%
54-65%
55-66%
54-60%
52-55%
GIX (N=8)
nt
aa
40-42%
33-36%
39-46%
33-37%
40-47%
36-41%
41-44%
29-30%
GX (N=14)
nt
aa
47-52%
37-41%
44-52%
37-43%
44-56%
38-43%
41-48%
30-35%
6
aall study strains (N=81) are included for sequence analysis in each genogroup as separated in 7
Figure 1. For homology comparison, percentage results of the lowest and the highest nt 8
(nucleotide) and aa (amino acid) strain identities are indicated in each box as a percentage 9
range in each genogroup. 10
bnumber of sequences analyzed 11
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
1
Figure 1. 1
PEC/Cow
den/
US
PECLL
14/U
S
Kore
an68
02
Po/S
V/M
iranda/2
000/V
E
Po/S
V/M
iranda2/2
001/V
E
PEC-K
ore
an10802
Po/S
V/Y
ara
cuy/1
999/V
EVet-25B
-DK
Vet-26b-D
KPE
CIV
A36/N
LV
P12-S
LO
P61-S
LO
P6-S
LO
P32-S
LO
P43
-SLO
Vet
-9-D
K
P87
-SLO
4472
-3L2
-ESP
Vet-3
2-DK
P60-S
LO
OH-J
J-25
9-00
-US
NC-QW
270-
03-U
S
Id2/
5/HUN
Id3/5/HUN
PoS3/7/HUNPoS6/7/HUNVet-55-DKVet-21A-DK
Vet-23-DK
338-1-FI
14-351-3-FI
P89-SLOVet-30-DKPoS9/7/HUNPoS10/7/HUNSWECI/VA10/NL
OH-MM280-03-US
Vet-47-DK
P21-SLO
P80-SLO
P81-SLO
P50-SLO
P66-SLO
P10-S
LO
P78
-SLO
P79-S
LO
P48-S
LO
P67-S
LO
P70-S
LO
P65-S
LO
P73-S
LO
P72-S
LO
P77-SLO
SW
ECII/VA10
3/NL
MI-Q
W19
-02-
US
Vet-28-D
K
Vet-19-D
K
P71-S
LO
43-0
6-1
8-p
3-6
-ITA
PoS7/7
/HU
N355-5
-FI
Sw
19-5
/06/IT
Arg
39/A
rgN
B-lik
eM
EC
/1/1
999/U
SH
UN
3739
Housto
n90
Mex1
491
7/0
0/M
X
Sap
poro
/82/
JP
Mc1
0/00/
TH
C12
/00/JP
cruise
ship/00/US
Mex
340/90
/MX
Hou7-
1181
-90-
US
London92
/UK
HUNs11/2000/H
UN
MEX335/1991/MX
HUNs12/2000/HUN
HUNs17/2000/HUN
PAN-1/78/U
S
FCV
Vet-21B
-DK
Vet-23B-D
K
Vet
-37-
DK
Vet-7-D
K
OH-LL26
/2002
/US
Vet-22
-DK
Vet-29-DK
K7JPVet-31-DKVet-4-DKP5-SLOP39-SLO
P49-SLO
OH-JJ681/2000/USVet-57-DKVet-3-DKVet-4B-DK
Vet-33A-DK
Vet-25-DK
Sw35-6/06/IT
Vet-56-DK
Sw17-5/06/IT
Sw36-2/06/IT
Vet-6-DK
Vet-52-D
K
Vet-1
9B-D
K
9-T
EUR-2
4-F
I
Vet-4
0-D
K
Sw
34-2
/06/IT
Vet-1
-DK
Vet-2
-DK
Vet-3
3B
-DK
Vet-3
1B
-DK
Vet-8
-DK
294-0
7-S
LO
SW
EC
III/VA
112/N
LVet-3
8-D
K8-T
EUR-2
3-FI
Norw
alk
0.00.10.20.30.40.50.6 2
GII (genotypes 1-5)
Denmark-DK (35)
Finland-FI (5)
Hungary-HUN (7)
Italy-IT (5)
Slovenia-SLO (28)
Spain-ESP (1)
B (?)
A (?)
GVIII
GV
GI (genotypes 1-3)
GIV (recombinant)
GVII
GVI
GIII
GIX (?)
GX (?)
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

Figure 2.
4 15
13
71
11
1
5
3
22
3
101
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
<1 month 1-3 months >3 months
GIIIA GIIIB GVI GVII GVIII GIX GX
N=57 N=14 N=7
on June 26, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
Related Documents











