Depto. de Entomologia, Fitopatologia e Zoologia Agrı´cola da Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de Sa ˜o Paulo, Piracicaba, Brazil Gas Exchange and Emission of Chlorophyll Fluorescence during the Monocycle of Rust, Angular Leaf Spot and Anthracnose on Bean Leaves as a Function of their Trophic Characteristics R. B. BASSANEZI 1 , L. AMORIM 1 , A. BERGAMIN FILHO 1 and R. D. BERGER 2 Authors’ addresses: 1 Depto. de Entomologia, Fitopatologia e Zoologia Agrı´cola da Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de Sa˜o Paulo, C. Postal 9, Piracicaba, 13418–900, SP, Brazil; 2 Plant Pathology Department, University of Florida, Gainesville 32611–0680, USA (correspondence to L. Amorim. E-mail: [email protected]) With 4 figures Received July 23, 2001; accepted October 19, 2001 Keywords: Phaseolus vulgaris, Uromyces appendiculatus, Phaeoisariopsis griseola, Colletotrichum lindemuthianum, photosynthesis, respiration, transpiration Abstract Measurements related to gas exchange and chlorophyll fluorescence emission were taken from healthy and diseased bean leaves with rust, angular leaf spot, and anthracnose during lesion development for each disease. The experiments were performed at different tempera- tures of plant incubation, and using two bean cultivars. The main effect of temperature of plant incubation was in disease development. There was no significant differ- ence between cultivars in relation to disease develop- ment and in magnitude of physiological alterations when disease severity was the same for each cultivar. These diseases reduced the net photosynthetic rate and increased the dark respiration of infected leaves after the appearance of visible symptoms and the differences between healthy and diseased leaves increased with disease development. The transpiration rate and stoma- tal conductance were stable during the monocycle of rust, however, these two variables decreased in leaves with angular leaf spot and anthracnose beginning with symptom appearance and continuing until lesion devel- opment was complete. Carboxylation resistance was probably the main factor related to reduction of photosynthetic rate of the apparently healthy area of leaves with rust and angular leaf spot. Reduction of the intercellular concentration of CO 2 , due to higher stomatal resistance, was probably the main factor for leaves with anthracnose. Chlorophyll fluorescence assessments suggested that there was no change in electron transport capacity and generation of ATP and NADPH in apparently healthy areas of diseased leaves, but decreases in chlorophyll fluorescence emission occurred on visibly lesioned areas for all diseases. Minimal fluorescence was remarkably reduced in leaves with angular leaf spot. Maximal fluorescence and optimal quantum yield of photosystem II of leaves were reduced for all three diseases. Bean rust, caused by a biotrophic pathogen, induced less damage to the regu- lation mechanisms of the physiological processes of the remaining green area of diseased leaves than did bean angular leaf spot or anthracnose, caused by hemibio- trophic pathogens. The magnitude of photosynthesis reduction can be related to the host–pathogen trophic relationships. Introduction The disease process that is a result of the interaction between a pathogen and its host leads to changes in several physiological processes of the host plant, inclu- ding dark respiration, photosynthesis, translocation of water and nutrients, transpiration, and regulation of growth (Lucas, 1998). Fungal infection generally redu- ces the photosynthetic rate by decreasing functional leaf area and reducing the photosynthetic efficiency of the remaining green leaf area (Livne, 1964; Raggi, 1978; Boote et al., 1980; Bastiaans, 1991; Shtienberg, 1992; Bassanezi et al., 1997, 2000; 2001). Dark respiration of leaves usually rises after infection (Daly et al., 1961; Raggi, 1978; Owera et al., 1981; Martin, 1986; Bassanezi et al., 2001) in consequence of increased metabolic activity of cells of the diseased leaf (Lucas, 1998). The transpiration rate can go up or down during the infection process, depending on the pathosystem (Duni- way and Durbin, 1971; Owera et al., 1981; Martin, 1986; Ephrath et al., 1989; McGrath and Pennypacker, 1990; Shtienberg, 1992; Bassanezi et al., 1997). Water loss from an infected leaf can increase due to destruction of part of the leaf cuticle, increased permeability of leaf J. Phytopathology 150, 37–47 (2002) Ó 2002 Blackwell Wissenschafts-Verlag, Berlin ISSN 0931-1785 U. S. Copyright Clearance Centre Code Statement: 0931–1785/2002/5000–0037 $ 15.00/0 www.blackwell.de/synergy

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Depto. de Entomologia, Fitopatologia e Zoologia Agrıcola da Escola Superior de Agricultura ‘Luiz de Queiroz’,Universidade de Sao Paulo, Piracicaba, Brazil
Gas Exchange and Emission of Chlorophyll Fluorescence during
the Monocycle of Rust, Angular Leaf Spot and Anthracnose
on Bean Leaves as a Function of their Trophic Characteristics
R. B. BASSANEZI1, L. AMORIM
1, A. BERGAMIN FILHO1 and R. D. BERGER
2
Authors’ addresses: 1Depto. de Entomologia, Fitopatologia e Zoologia Agrıcola da Escola Superior de Agricultura ‘Luiz deQueiroz’, Universidade de Sao Paulo, C. Postal 9, Piracicaba, 13418–900, SP, Brazil; 2 Plant Pathology Department,
University of Florida, Gainesville 32611–0680, USA (correspondence to L. Amorim. E-mail: [email protected])
With 4 figures
Received July 23, 2001; accepted October 19, 2001
Keywords: Phaseolus vulgaris, Uromyces appendiculatus, Phaeoisariopsis griseola, Colletotrichum lindemuthianum,
photosynthesis, respiration, transpiration
AbstractMeasurements related to gas exchange and chlorophyllfluorescence emission were taken from healthy anddiseased bean leaves with rust, angular leaf spot, andanthracnose during lesion development for each disease.The experiments were performed at different tempera-tures of plant incubation, and using two bean cultivars.The main effect of temperature of plant incubation wasin disease development. There was no significant differ-ence between cultivars in relation to disease develop-ment and in magnitude of physiological alterationswhen disease severity was the same for each cultivar.These diseases reduced the net photosynthetic rate andincreased the dark respiration of infected leaves after theappearance of visible symptoms and the differencesbetween healthy and diseased leaves increased withdisease development. The transpiration rate and stoma-tal conductance were stable during the monocycle ofrust, however, these two variables decreased in leaveswith angular leaf spot and anthracnose beginning withsymptom appearance and continuing until lesion devel-opment was complete. Carboxylation resistance wasprobably the main factor related to reduction ofphotosynthetic rate of the apparently healthy area ofleaves with rust and angular leaf spot. Reduction of theintercellular concentration of CO2, due to higherstomatal resistance, was probably the main factor forleaves with anthracnose. Chlorophyll fluorescenceassessments suggested that there was no change inelectron transport capacity and generation of ATP andNADPH in apparently healthy areas of diseased leaves,but decreases in chlorophyll fluorescence emissionoccurred on visibly lesioned areas for all diseases.Minimal fluorescence was remarkably reduced in leaves
with angular leaf spot. Maximal fluorescence andoptimal quantum yield of photosystem II of leaves werereduced for all three diseases. Bean rust, caused by abiotrophic pathogen, induced less damage to the regu-lation mechanisms of the physiological processes of theremaining green area of diseased leaves than did beanangular leaf spot or anthracnose, caused by hemibio-trophic pathogens. The magnitude of photosynthesisreduction can be related to the host–pathogen trophicrelationships.
IntroductionThe disease process that is a result of the interactionbetween a pathogen and its host leads to changes inseveral physiological processes of the host plant, inclu-ding dark respiration, photosynthesis, translocation ofwater and nutrients, transpiration, and regulation ofgrowth (Lucas, 1998). Fungal infection generally redu-ces the photosynthetic rate by decreasing functional leafarea and reducing the photosynthetic efficiency of theremaining green leaf area (Livne, 1964; Raggi, 1978;Boote et al., 1980; Bastiaans, 1991; Shtienberg, 1992;Bassanezi et al., 1997, 2000; 2001). Dark respiration ofleaves usually rises after infection (Daly et al., 1961;Raggi, 1978; Owera et al., 1981; Martin, 1986; Bassaneziet al., 2001) in consequence of increased metabolicactivity of cells of the diseased leaf (Lucas, 1998). Thetranspiration rate can go up or down during theinfection process, depending on the pathosystem (Duni-way and Durbin, 1971; Owera et al., 1981; Martin, 1986;Ephrath et al., 1989; McGrath and Pennypacker, 1990;Shtienberg, 1992; Bassanezi et al., 1997). Water lossfrom an infected leaf can increase due to destruction ofpart of the leaf cuticle, increased permeability of leaf
J. Phytopathology 150, 37–47 (2002)Ó 2002 Blackwell Wissenschafts-Verlag, BerlinISSN 0931-1785
U. S. Copyright Clearance Centre Code Statement: 0931–1785/2002/5000–0037 $ 15.00/0 www.blackwell.de/synergy

cell membranes, or inhibition of stomatal closure.Reduction of transpiration may result from stomatalclosure, reduction of air space by hyphae or hypertro-phy of mesophyll cells, obstruction of conducting tissueand stomata, and defoliation.
Although it is well known that plant pathogens alterthe physiological processes of their hosts, comparativestudies involving biotrophic, hemibiotrophic and nec-rotrophic pathogens and their changes in the hostphysiological processes are not very common and aremade at a specific stage of disease development(Shtienberg, 1992; Bassanezi et al., 2000, 2001). Therelative photosynthetic rate per unit of wheat leaf areawere smaller for rust and powdery mildew, causedbiotrophic pathogens, than for Septoria leaf blotch,caused by a necrotrophic pathogen (Shtienberg, 1992).The greatest impact on photosynthesis of the necro-trophic pathogens could be explained by the secretionof enzymes or phytotoxic compounds that diffuse touncolonized portions of the leaf (Shtienberg, 1992).Necrotrophic pathogens that cause vein infection caninduce a systemic stomatal closure in green areas of theleaf (Meyer et al., 2001), reducing the photosyntheticrate of the whole leaf considerably even when diseaseseverity is low. For biotrophic pathogens, however, thegreatest decline in photosynthesis occurs in fungallyinvaded regions of the leaf or in the areas adjacent tothe fungal mycelium (Scholes and Rolfe, 1995). Thereduction in the rate of transpiration of plants infectedby necrotrophic pathogens was proportional to thediseased area (Shtienberg, 1992). The destruction ofcells were directly related to the presence of the fungus.Plants with rust, however, had a decrease in transpir-ation that was proportionally smaller than the corres-ponding reduction of healthy leaf area due to disease.At low levels of rust infection, the transpiration rateexceeded that of healthy leaves. This has been attrib-uted to the epidermal rupture after pustule formation(Shtienberg, 1992).
The methods commonly used for in vivo measure-ments of photosynthesis are gas exchange and chloro-phyll fluorescence emission (Hall et al., 1993). Variablesusually obtained by gas exchange systems are CO2
assimilation and release rates, transpiration rate,intercellular concentration of CO2, and stomatal con-ductance. Measurements of emission of fluorescence bychlorophyll a provide information about photosystem II(PSII) activity and changes in photosynthetic meta-bolism of diseased leaves (Scholes and Farrar, 1985;Moll et al., 1995; Peterson and Aylor, 1995; Wrightet al., 1995; Schnabel et al., 1998).
The aim of this study was to compare the alterationsin photosynthesis, dark respiration, and transpiration inleaves infected with biotrophic and hemibiotrophicpathogens during lesion development. The pathosystemsstudied were common bean (Phaseolus vulgaris L) withrust, angular leaf spot and anthracnose. Uromycesappendiculatus (Pers.) Unger. is a biotrophic pathogenthat causes bean rust and produces circular uredinialpustules on leaves that rupture the epidermis to produce
abundant urediniospores. Phaeoisariopsis griseola(Sacc.) Ferraris is a hemibiotrophic pathogen and thecausal agent of bean angular leaf spot producingnecrotic and angular lesions, chlorosis and prematureabscission of affected leaf. Colletotrichum lindemuthia-num (Sacc. and Magn.) Br. and Cav. is a hemibiotrophicpathogen that establishes infection through a briefbiotrophic phase and switches to a necrotrophic phase,when symptoms appear (O’Connell et al., 2000). Col-letotrichum lindemuthianum causes bean anthracnoseinfecting the lower surfaces of leaves and leaf veinsand produces necrotic lesions by cell wall enzymaticdegradation (Hall, 1991).
Materials and MethodsCommon bean cultivars and growth conditions
Two repetitions of experiments for each disease werecarried out in controlled environment chambers: R1and R2 for rust, S1 and S2 for angular leaf spot, andA1 and A2 for anthracnose. The influence of differenttemperatures on disease development and onphysiological alterations caused by the pathogens inleaves of the common bean cultivars Rosinha G-2and Carioca Comum were assessed in all experiments.The plants were grown in aluminium pots with 2 l ofa mixture of clay soil, sand, and cow manure in a2 : 2 : 1 proportion, and kept in a glasshouse untilinoculation.
Inoculation and incubation
Uromyces appendiculatus were cultivated on cv. RosinhaG-2 at 21°C and urediniospores were collected soonafter the onset of sporulation and stored until inocula-tion at )8°C in capsules put in a closed flask containingsilica gel. Phaeoisariopsis griseola was cultivated ontomato juice medium at 24°C in the dark. Conidia werecollected after 14 days, then used for inoculation.Colletotrichum lindemuthianum, race kappa, was culti-vated on sterilized bean pods at 24°C in the dark.Conidia were collected after 21 days and used forinoculation.There were four treatments distributed in a com-
pletely randomized design (with four replications) foreach experiment: inoculated plants of cv. Rosinha G-2,inoculated plants of cv. Carioca Comum, non-inocula-ted plants of cv. Rosinha G-2, and non-inoculatedplants of cv. Carioca Comum. Both leaf surfaces wereinoculated using an atomizer to spray a spore suspen-sion until run-off. For experiments with rust (R1and R2), the second trifoliate leaf was inoculated witha suspension of 5 · 104 urediniospores/ml of U. appendi-culatus + surfactant (Tween 20 at 0.1% v/v), 16 daysafter sowing (das). For angular leaf spot, the firsttrifoliate leaf was inoculated on the 14th das with asuspension of 2.8 · 105 conidia/ml of P. griseola +surfactant (experiment S1), and 1.4 · 106 conidia/ml + surfactant (experiment S2). For anthracnose(experiments A1 and A2), the first trifoliate leaf wasinoculated at 16 das with a suspension of 2.7 · 105
conidia/ml of C. lindemuthianum + surfactant. Plants
38 BASSANEZI et al.

of control treatments were sprayed with water + surf-actant only.
Plants were placed in a growth chamber (model E-7;Conviron, Winnipeg, Canada) immediately after inocu-lation. Pots were kept in the dark and enclosed in humidplastic bags for 24 h at 19°C for rust and anthracnose,and for 30 h at 24°C for angular leaf spot. When thehumid chambers were removed, the plants were kept atan irradiance (PAR) of 180–230 lE/m2s at the top of theplants during the 14 h photoperiod. The incubationtemperatures were 15, 19, and 23°C for rust andanthracnose, and 19, 22, and 25°C for angular leaf spot.
Assessment of gas exchange
The assessments of net photosynthetic rate (An, lmolCO2/m
2 per s), dark respiration (Rd, lmol CO2/m2 per s),
stomatal conductance to water vapour (gse, mol H2O/m2
per s), transpiration rate (E, mmol H2O/m2 per s), andintercellular concentration of CO2 (Ci, lmol CO2) weremade with a Portable Photosynthesis System (LI-6400;Licor, Lincoln, NB, USA) on an area of 6 cm2 on thecentral leaflet of inoculated leaves and correspondingleaves of non-inoculated plants. For plants with rust,five assessments were made at 5, 7, 9, 11 and 13 daysafter inoculation (dai). Six assessments were made at 3,5, 7, 9, 11, and 13 dai for angular leaf spot, and from thethird to eighth day after inoculation in the case ofanthracnose. The conditions during the assessmentswere: leaf temperature at 25°C; irradiance (PAR) of400 lE/m2 s provided by a red LED light source (6400–02; Licor); air flux of 500 lmol/s; CO2 concentration inthe air coming into the chamber ranged from 360 to380 ll/l; relative humidity in the chamber ranged from30 to 50%. The central leaflet was kept under light untilthe stabilization of An, E, gse, and Ci values, then thecentral leaflet was kept in the dark for 3 min to measureRd.
Assessment of variables of chlorophyll fluorescence
The minimal fluorescence (F0), maximal fluorescence(Fm), and optimal quantum yield of PSII (Fv/Fm)were assessed using a PAM-2000 fluorometer (Walz,Effeltrich, Germany) in the leaflets of non-inoculatedplants, and in two regions of inoculated leaves: lesionedand apparently healthy areas. These fluorescence vari-ables were assessed only on plants kept at 19°C inexperiments F1, S1, and A2.
The assessed leaf areas (38.5 mm2) were kept in thedark for 20 min with the use of a dark leaf clip (DLC-8;Walz) which cover an area of 350 mm2. The F0 wasassessed after the emission of a modulated red lightbeam of 600 Hz and very low intensity. The Fm wasobtained with a saturating light pulse for 0.6 s. The valueof Fv/Fm was calculated using Fv/Fm¼ (Fm – F0)/Fm.
Assessment of disease severity
The area assessed for gas exchange on the central leafletwas marked so that the next assessments could be madein the same area. This marked area (6 cm2) wasphotographed with a zoom digital camera (DC120:
Kodak DS, Rochester, NY, USA) at a distance of20 cm, illuminated by four fluorescent 20 W lamps. Thedigital images were transferred to a computer andprocessed with the software SIARCS 3.0 (EMBRAPA,Sao Carlos, Brazil) to estimate the severity of eachdisease. The necrotic area as well as the yellowish haloand the soaked, faded or dried areas that surrounded thelesions were all included in the assessment of diseaseseverity. The disease severity of the leaf region where thevariables of chlorophyll fluorescence were assessed wasestimated by counting the number of lesions andmultiplying this number by a visual estimate of averagelesion size with the aid of standard area diagrams, thendividing by assessed leaf area (38.5 mm2).
Data analysis
Data from the two repetitions of experiments for eachdisease were analysed together. The results wereexpressed as a percentage of the average of the healthyleaves. Gas exchange variables were related to eachother by the Pearson coefficient (r) and to diseaseseverity and stomatal conductance by non-linear corre-lation coefficients (R). Values of F0 and Fm were fitted bylinear regression, and values of Fv/Fm, by non-linearregression relative to the disease severity.
ResultsRust
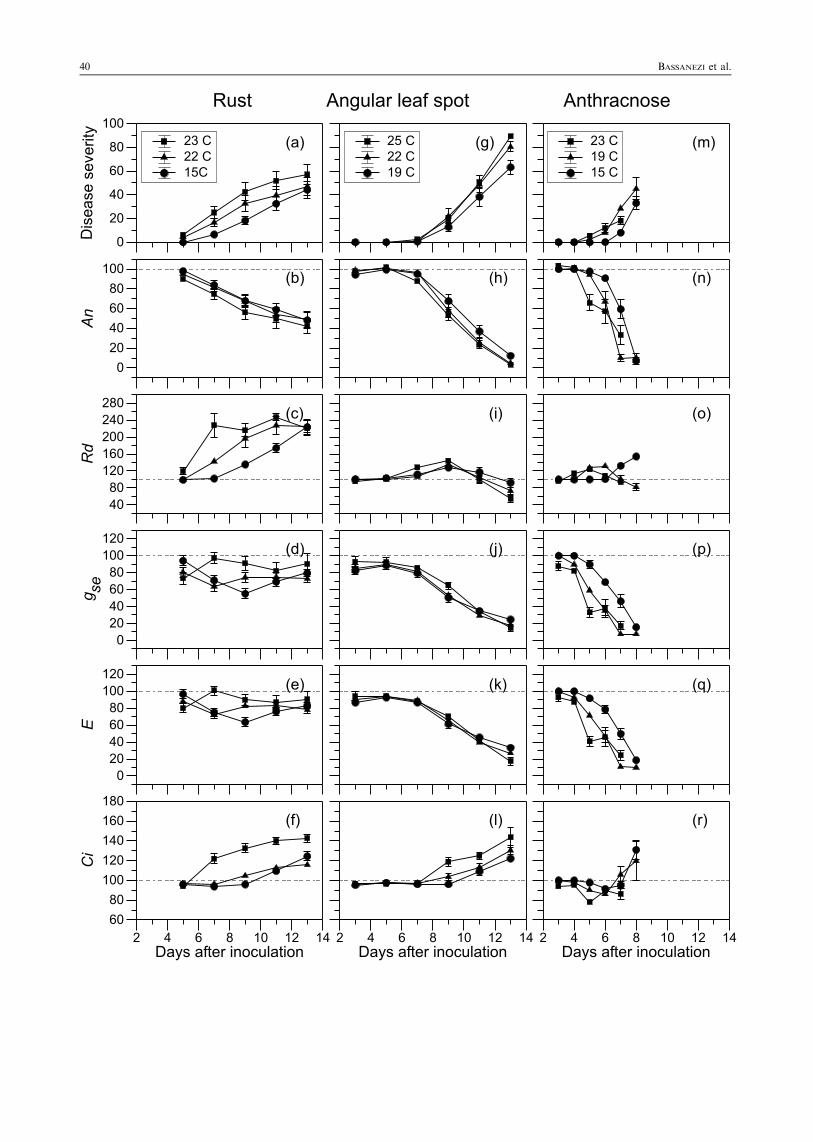
Disease developmentDisease development was faster at 19 and 23°C than at15°C. The first symptoms appeared at 4 dai in plantsthat were kept at 19 and 23°C and at 5 dai in those thatwere incubated at 15°C. Complete sporulation (100% ofsporulating pustules) was observed at 9 dai (19 and23°C) and 11 dai (15°C). There were no significantdifferences between bean cultivars in time of symptomappearance, number of lesions per leaf area, and rate oflesion growth (data not shown). On the 13th dai, theaverage rust severities for cv. Rosinha G-2 at 23, 19 and15°C were 60, 48, and 42%, respectively, whereas for cv.Carioca Comum the severities were 50, 45 and 43%. Ascultivar effects were insignificant, data for the twocultivars were combined in Fig. 1a–f. Disease severityincreased at each assessment due to lesion growth(Fig. 1a).
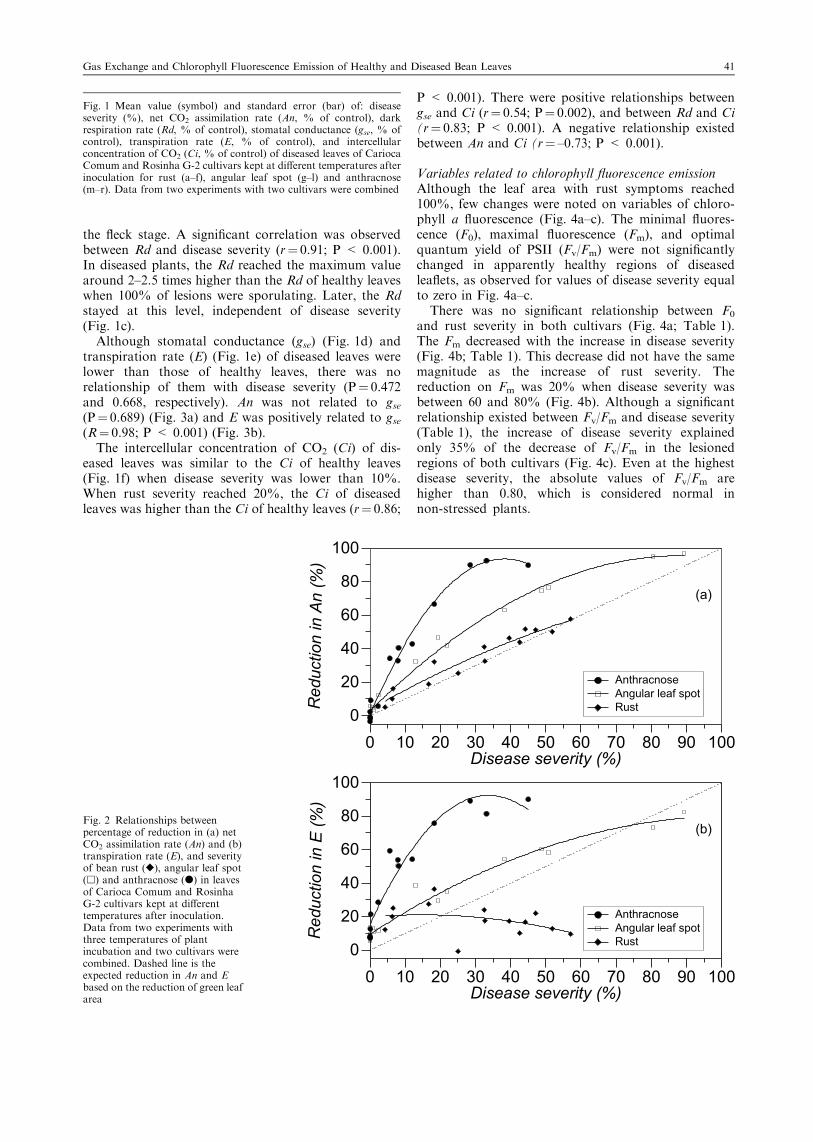
Gas exchange variablesThe net photosynthetic rate (An) of diseased leaves,expressed as a percentage of the rate of healthy leaves,decreased with the disease development for the threetemperatures (Fig. 1b). As temperature did not affectthe interaction between pathogens and the physiolo-gical processes, data from the three temperatures werecombined in Fig. 2. Reduction in An was positivelyrelated to disease severity (R¼ 0.97; P < 0.001)(Fig. 2a). This reduction was slightly greater thanexpected on the basis of leaf area covered by rustsymptoms.
There was an increase in dark respiration rate (Rd ) indiseased leaves compared with the healthy leaves after
Gas Exchange and Chlorophyll Fluorescence Emission of Healthy and Diseased Bean Leaves 39
40 BASSANEZI et al.
the fleck stage. A significant correlation was observedbetween Rd and disease severity (r¼ 0.91; P < 0.001).In diseased plants, the Rd reached the maximum valuearound 2–2.5 times higher than the Rd of healthy leaveswhen 100% of lesions were sporulating. Later, the Rdstayed at this level, independent of disease severity(Fig. 1c).
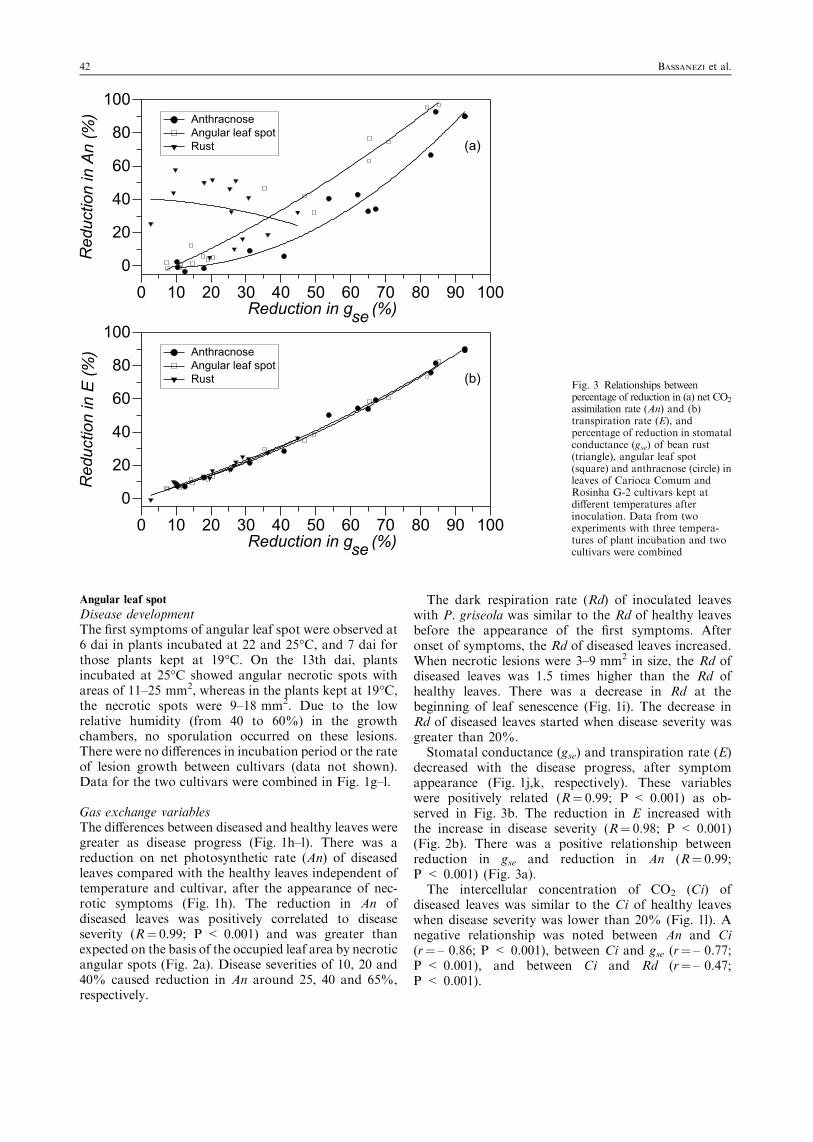
Although stomatal conductance (gse) (Fig. 1d) andtranspiration rate (E) (Fig. 1e) of diseased leaves werelower than those of healthy leaves, there was norelationship of them with disease severity (P¼ 0.472and 0.668, respectively). An was not related to gse(P¼ 0.689) (Fig. 3a) and E was positively related to gse(R¼ 0.98; P < 0.001) (Fig. 3b).
The intercellular concentration of CO2 (Ci) of dis-eased leaves was similar to the Ci of healthy leaves(Fig. 1f) when disease severity was lower than 10%.When rust severity reached 20%, the Ci of diseasedleaves was higher than the Ci of healthy leaves (r¼ 0.86;
P < 0.001). There were positive relationships betweengse and Ci (r¼ 0.54; P¼ 0.002), and between Rd and Ci(r¼ 0.83; P < 0.001). A negative relationship existedbetween An and Ci (r¼ –0.73; P < 0.001).
Variables related to chlorophyll fluorescence emissionAlthough the leaf area with rust symptoms reached100%, few changes were noted on variables of chloro-phyll a fluorescence (Fig. 4a–c). The minimal fluores-cence (F0), maximal fluorescence (Fm), and optimalquantum yield of PSII (Fv/Fm) were not significantlychanged in apparently healthy regions of diseasedleaflets, as observed for values of disease severity equalto zero in Fig. 4a–c.
There was no significant relationship between F0
and rust severity in both cultivars (Fig. 4a; Table 1).The Fm decreased with the increase in disease severity(Fig. 4b; Table 1). This decrease did not have the samemagnitude as the increase of rust severity. Thereduction on Fm was 20% when disease severity wasbetween 60 and 80% (Fig. 4b). Although a significantrelationship existed between Fv/Fm and disease severity(Table 1), the increase of disease severity explainedonly 35% of the decrease of Fv/Fm in the lesionedregions of both cultivars (Fig. 4c). Even at the highestdisease severity, the absolute values of Fv/Fm arehigher than 0.80, which is considered normal innon-stressed plants.
Fig. 2 Relationships betweenpercentage of reduction in (a) netCO2 assimilation rate (An) and (b)transpiration rate (E), and severityof bean rust (r), angular leaf spot(h) and anthracnose (d) in leavesof Carioca Comum and RosinhaG-2 cultivars kept at differenttemperatures after inoculation.Data from two experiments withthree temperatures of plantincubation and two cultivars werecombined. Dashed line is theexpected reduction in An and Ebased on the reduction of green leafarea
Fig. 1 Mean value (symbol) and standard error (bar) of: diseaseseverity (%), net CO2 assimilation rate (An, % of control), darkrespiration rate (Rd, % of control), stomatal conductance (gse, % ofcontrol), transpiration rate (E, % of control), and intercellularconcentration of CO2 (Ci, % of control) of diseased leaves of CariocaComum and Rosinha G-2 cultivars kept at different temperatures afterinoculation for rust (a–f), angular leaf spot (g–l) and anthracnose(m–r). Data from two experiments with two cultivars were combined
Gas Exchange and Chlorophyll Fluorescence Emission of Healthy and Diseased Bean Leaves 41
Angular leaf spot
Disease developmentThe first symptoms of angular leaf spot were observed at6 dai in plants incubated at 22 and 25°C, and 7 dai forthose plants kept at 19°C. On the 13th dai, plantsincubated at 25°C showed angular necrotic spots withareas of 11–25 mm2, whereas in the plants kept at 19°C,the necrotic spots were 9–18 mm2. Due to the lowrelative humidity (from 40 to 60%) in the growthchambers, no sporulation occurred on these lesions.There were no differences in incubation period or the rateof lesion growth between cultivars (data not shown).Data for the two cultivars were combined in Fig. 1g–l.
Gas exchange variablesThe differences between diseased and healthy leaves weregreater as disease progress (Fig. 1h–l). There was areduction on net photosynthetic rate (An) of diseasedleaves compared with the healthy leaves independent oftemperature and cultivar, after the appearance of nec-rotic symptoms (Fig. 1h). The reduction in An ofdiseased leaves was positively correlated to diseaseseverity (R¼ 0.99; P < 0.001) and was greater thanexpected on the basis of the occupied leaf area by necroticangular spots (Fig. 2a). Disease severities of 10, 20 and40% caused reduction in An around 25, 40 and 65%,respectively.
The dark respiration rate (Rd) of inoculated leaveswith P. griseola was similar to the Rd of healthy leavesbefore the appearance of the first symptoms. Afteronset of symptoms, the Rd of diseased leaves increased.When necrotic lesions were 3–9 mm2 in size, the Rd ofdiseased leaves was 1.5 times higher than the Rd ofhealthy leaves. There was a decrease in Rd at thebeginning of leaf senescence (Fig. 1i). The decrease inRd of diseased leaves started when disease severity wasgreater than 20%.Stomatal conductance (gse) and transpiration rate (E)
decreased with the disease progress, after symptomappearance (Fig. 1j,k, respectively). These variableswere positively related (R¼ 0.99; P < 0.001) as ob-served in Fig. 3b. The reduction in E increased withthe increase in disease severity (R¼ 0.98; P < 0.001)(Fig. 2b). There was a positive relationship betweenreduction in gse and reduction in An (R¼ 0.99;P < 0.001) (Fig. 3a).The intercellular concentration of CO2 (Ci) of
diseased leaves was similar to the Ci of healthy leaveswhen disease severity was lower than 20% (Fig. 1l). Anegative relationship was noted between An and Ci(r¼ – 0.86; P < 0.001), between Ci and gse (r¼ – 0.77;P < 0.001), and between Ci and Rd (r¼ – 0.47;P < 0.001).
Fig. 3 Relationships betweenpercentage of reduction in (a) net CO2
assimilation rate (An) and (b)transpiration rate (E), andpercentage of reduction in stomatalconductance (gse) of bean rust(triangle), angular leaf spot(square) and anthracnose (circle) inleaves of Carioca Comum andRosinha G-2 cultivars kept atdifferent temperatures afterinoculation. Data from twoexperiments with three tempera-tures of plant incubation and twocultivars were combined
42 BASSANEZI et al.
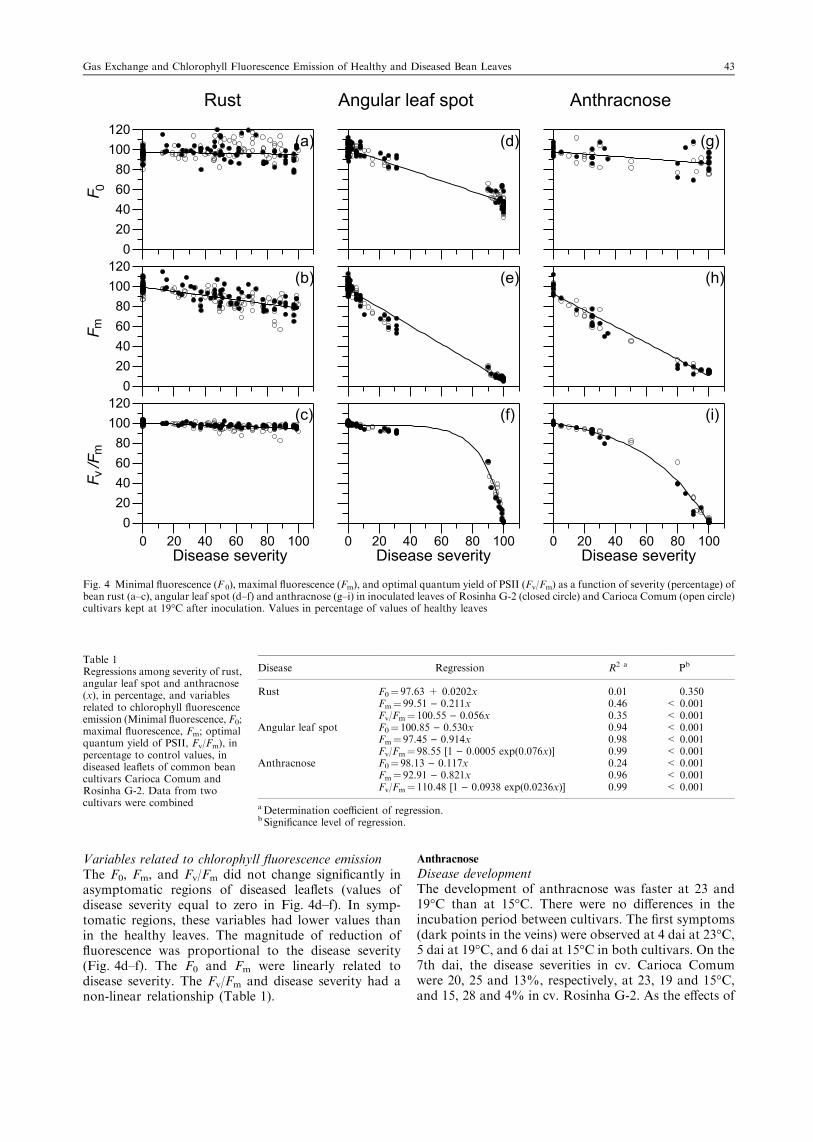
Variables related to chlorophyll fluorescence emissionThe F0, Fm, and Fv/Fm did not change significantly inasymptomatic regions of diseased leaflets (values ofdisease severity equal to zero in Fig. 4d–f). In symp-tomatic regions, these variables had lower values thanin the healthy leaves. The magnitude of reduction offluorescence was proportional to the disease severity(Fig. 4d–f). The F0 and Fm were linearly related todisease severity. The Fv/Fm and disease severity had anon-linear relationship (Table 1).
Anthracnose
Disease developmentThe development of anthracnose was faster at 23 and19°C than at 15°C. There were no differences in theincubation period between cultivars. The first symptoms(dark points in the veins) were observed at 4 dai at 23°C,5 dai at 19°C, and 6 dai at 15°C in both cultivars. On the7th dai, the disease severities in cv. Carioca Comumwere 20, 25 and 13%, respectively, at 23, 19 and 15°C,and 15, 28 and 4% in cv. Rosinha G-2. As the effects of
0m
mv
Fig. 4 Minimal fluorescence (F 0), maximal fluorescence (Fm), and optimal quantum yield of PSII (Fv/Fm) as a function of severity (percentage) ofbean rust (a–c), angular leaf spot (d–f) and anthracnose (g–i) in inoculated leaves of Rosinha G-2 (closed circle) and Carioca Comum (open circle)cultivars kept at 19°C after inoculation. Values in percentage of values of healthy leaves
Table 1Regressions among severity of rust,angular leaf spot and anthracnose(x), in percentage, and variablesrelated to chlorophyll fluorescenceemission (Minimal fluorescence, F0;maximal fluorescence, Fm; optimalquantum yield of PSII, Fv/Fm), inpercentage to control values, indiseased leaflets of common beancultivars Carioca Comum andRosinha G-2. Data from twocultivars were combined
Disease Regression R2 a Pb
Rust F0¼ 97.63 + 0.0202x 0.01 0.350Fm¼ 99.51 ) 0.211x 0.46 < 0.001Fv/Fm¼ 100.55 ) 0.056x 0.35 < 0.001
Angular leaf spot F0¼ 100.85 ) 0.530x 0.94 < 0.001Fm¼ 97.45 ) 0.914x 0.98 < 0.001Fv/Fm¼ 98.55 [1 ) 0.0005 exp(0.076x)] 0.99 < 0.001
Anthracnose F0¼ 98.13 ) 0.117x 0.24 < 0.001Fm¼ 92.91 ) 0.821x 0.96 < 0.001Fv/Fm¼ 110.48 [1 ) 0.0938 exp(0.0236x)] 0.99 < 0.001
aDetermination coefficient of regression.b Significance level of regression.
Gas Exchange and Chlorophyll Fluorescence Emission of Healthy and Diseased Bean Leaves 43

cultivars were insignificant, data for the two cultivarswere combined in Fig. 1m–r.
Gas exchange variablesRegardless of temperature and cultivar, the An ofdiseased leaves decreased in relation to the healthyleaves following the disease development (Fig. 1n). Thereduction in net photosynthetic rate (An) was observedsoon after the appearance of brown-red lesions in theveins with water soaking around the lesions (between 4and 5 dai at 23°C, 5 and 6 dai at 19°C and 6 dai at15°C). The proportion of reduction in An was positivelyrelated to anthracnose severity (R¼ 0.99; P < 0.001)and was higher than the proportion of leaf area withanthracnose symptoms (Fig. 2a). Only 10% of leaf areawith disease symptoms reduced An in almost 50% andwhen anthracnose severity reached values above 30%,the An was almost nil.
An increase in dark respiration rate (Rd ) of diseasedleaves in relation to the healthy leaves was observedfrom the appearance of the first symptoms to the stageof brown-red lesions in the veins (Fig. 1o). A reductionin Rd was observed in wilted leaves of plants incubatedat 19 and 23°C.
Stomatal conductance (gse) and transpiration rate(E) decreased during disease progress (Fig. 1p,q,respectively). There were positive relationshipsbetween gse and E (R¼ 0.99; P < 0.001) (Fig. 3b)and between anthracnose severity and reduction in E(R¼ 0.96; P < 0.001) (Fig. 2b). There was also pos-itive relationship between reduction in gse and reduc-tion in An (R¼ 0.97; P < 0.001) in the leaves withanthracnose.
The intercellular concentration of CO2 (Ci) inleaves with anthracnose was reduced after the appear-ance of first symptoms (Fig. 1r). The average reduc-tion in Ci in diseased leaves was 23, 17 and 10% at23, 19 and 15°C, respectively. There was a positivecorrelation between Ci and gse (r¼ 0.88; P < 0.001),and no significant relationship between Ci and Rd(P¼ 0.704).
Variables related to chlorophyll fluorescence emissionThe effect of anthracnose severity on fluorescencevariables of bean cultivars is shown in Fig. 4g–i.Significant linear regressions were observed between F0
and disease severity (Fig. 4g) but the coefficients ofdetermination (R2) were lower than 0.30 for bothcultivars (Table 1). The Fm was significantly reducedwith the increase in anthracnose severity (Fig. 4h;Table 1). The Fv/Fm and disease severity had a non-linear relationship for both cultivars (Fig. 4i; Table 1).While the anthracnose symptoms were limited to theinitial vein necrosis, the values of Fv/Fm were between0.75 and 0.85 (normal values for non-stressed plants). Inthe regions with vein necrosis and initial wilting (diseaseseverity higher than 50%), the reduction in Fv/Fm wasgreat. The fluorescence variables from apparentlyhealthy regions of diseased leaflets were similar to thevalues from healthy leaflets.
DiscussionAlthough the changes in physiological processes of aninfected host (decreased net photosynthetic rate andincreased dark respiration rate) occurred for the threediseases, each disease affected the physiological pro-cesses of infected leaves in its own way. The pattern ofresponses and the amount of reduction or increase ineach process were related to the type of host–pathogentrophic relationships. Biotrophy and necrotrophy donot represent absolute categories; in reality there is acontinuous gradation between the two types of para-sitism (Lucas, 1998). Uromyces appendiculatus,P. griseola and C. lindemuthianum are examples ofdifferent categories from this large diversity of trophicrelationships. Uromyces appendiculatus is a biotrophicfungus, P. griseola, a hemibiotrofic fungus, andC. lindemuthianum can be considered a necrotrophicfungus because its brief biotrophic phase had alreadybeen finished when photosynthesis assessments weremade. There was a gradually increasing damage on thephotosynthetic processes caused by these pathogensaccording to their trophic relationship; that is, lowerdamage caused by U. appendiculatus and higher, byC. lindemuthianum. The extent of the pathogen enzy-matic activity seems to be correlated to the reduction inthe host photosynthesis not only for bean pathogens. Ithas been reported that stem rust of wheat (McGrathand Pennypacker, 1990) and corn rust (Shtienberg,1992) caused a small reduction in the photosyntheticrate of the host plants. Pea spot caused by Mycosp-haerella pinodes (Garry et al., 1998), a necrotrophicfungus, however, caused significant reduction in CO2
assimilation.The effects of infection by U. appendiculatus on the
photosynthetic activity of common bean leaves havebeen described by many researchers. The decrease in netphotosynthetic rate (An) after symptom appearance(Livne, 1964; Raggi, 1978) has been associated withchloroplast alterations (Sziraki et al., 1984) and reduc-tion in chlorophyll content in diseased leaves (Szirakiet al., 1984; Moll et al., 1995; Wagner and Boyle, 1995;Stangarlin and Pascholati, 2000; Bassanezi et al., 2001;Lopes and Berger, 2001). In this study, the CO2
assimilation decreased with the evolution of disease,but this decrease was not totally explained by thereduction in green leaf area, which was interpreted asthe photosynthetic efficiency of the remaining appar-ently healthy leaf area was lightly affected. The rise instomatal resistance in the regions between pustulescould be responsible for part of the reduction inphotosynthetic rate, but during disease development,gse was stable, Ci increased, and An decreased. Appar-ently, An was not limited by a rise in the resistance ofCO2 uptake into the diseased leaf. Other alterations,such as changes in translocation patterns of photo-assimilates, carbohydrate accumulation (Zaki and Dur-bin, 1965; Livne and Daly, 1966; Sziraki et al., 1984;Wagner and Boyle, 1995), and an increase in acidinvertase activity in the tissues surrounding the pustules,
44 BASSANEZI et al.

would cause a down-regulation of the Calvin cycle and areduction of Rubisco activity (Wagner and Boyle, 1995;Stangarlin and Pascholati, 2000). These alterations canincrease the carboxylation resistance, and contribute tothe reduction of the photosynthetic rate in theseasymptomatic regions. The analysis of A/Ci curves ofrusted bean leaves suggested that reduction in Rubiscoactivity was the major cause of reduction in An ofdiseased leaves, whereas stomatal limitation was insig-nificant (Bassanezi et al., 2000).
In the pathosystem P. griseola–common bean, inwhich there is necrosis of foliar tissues, the decreased netphotosynthetic rate (An) was a consequence of loss ofchlorophyll. Stangarlin et al. (2000) and Bassanezi et al.(2001) observed a decreased chlorophyll content in thetissue that surrounded the lesions. In other pathosys-tems with necrotrophic or hemibiotrophic pathogens,the reduction in chlorophyll content had resulted fromthe action of enzymes that degraded organelles, ortoxins that were produced and released by the pathogenduring the colonization of host tissue (Fulton et al.,1965; Rasmussen and Scheffer, 1988). Until now, therehas been no evidence of such activities of enzymes ortoxins for the pathosystem P. griseola–common bean.The decrease in An was caused by the reduction inradiation interception (necrotic tissues) and by thedecreased photosynthetic efficiency of remaining greenfoliar tissue. This situation has been described in manypathosystems including hemibiotrophic or necrotrophicpathogens such as Cercospora arachidicola and Cercos-poridium personatum–peanut (Boote et al., 1980), Alter-naria alternata–cotton (Ephrath et al., 1989), andPyricularia grisea–rice (Bastiaans, 1991). Although apositive relationship between gse and An has been noted,the stomatal closure does not seem to be the mainlimiting factor for CO2 assimilation, because there was arise in Ci. Therefore, factors that lead to the increase incarboxylation resistance would be the main cause for thedecrease of An in the tissues between angular leaf spotlesions (Bassanezi et al., 2000).
The impact of C. lindemuthianum on the physiologicalprocesses of infected leaves is associated to the necro-trophic phase of fungal infection. An extensive dissolu-tion of cell walls occurs at the time that the pathogenadvances in the vascular tissue of the leaf (Wijesunderaet al., 1989). Low net photosynthetic rate (An) in leafletswith anthracnose was noticed when the first necroticsymptoms appeared. At this stage, host cells are killedrapidly in advance of infection (O’Connell et al., 2000).The An was drastically reduced as disease severityincreased. Wong and Thrower (1978a) had similarresults in leaves of Vigna sesquipedalis infected withC. lindemuthianum. Similar to the results from Lopesand Berger (2001) and Meyer et al. (2001), the decreasein An was higher then that expected by the reduction ofgreen leaf area. The lower photosynthetic efficiency ofasymptomatic leaf tissues could be explained by theincrease in stomatal and carboxylation resistances.There was no evidence that carbohydrate accumulationwas the cause of decreased CO2 assimilation in leaves
with anthracnose (Wong and Thrower, 1978b), but thereduction of Rubisco activity was significant in leaveswith this disease (Bassanezi et al., 2000).
The Fv/Fm parameter provides information on thepotential photosynthetic efficiency of PSII, and it isfrequently discussed as a vitality index (Moll et al.,1995). In this study, a slight reduction was observed inFv/Fm in the rusted regions. This occurred because thediseased region assessed by the fluorometer probecontained lesioned and non-lesioned tissues. Otherauthors (Moll et al., 1995; Lopes, 1999) reported signi-ficant reductions in Fv/Fm in the areas totally covered bybean rust pustules. There was a reduction in chlorophyllcontent and damage to the thylakoid membrane ofchloroplasts in the tissues infected by P. griseola. As aconsequence, reductions on Fv/Fm may have been from areduced quantum efficiency of PSII, including a lowercapacity of electron transport, and generation of ATPand NADPH. The F0 did not change in leaves withanthracnose because of the maintenance of chlorophyllcontent during the first stages of disease development, asobserved by Lopes (1999). However, Fm was signifi-cantly reduced at the appearance of first symptoms andcontinued to decrease as anthracnose severity increased.There was a clear imbalance between the drasticinhibition in An and the moderate decrease in photo-chemical reactions in bean leaves with anthracnose. Inthe same way for rust and angular leaf spot, apparentlyhealthy tissues of diseased leaflets were quite similar tohealthy leaflets for the fluorescence variables.
Few changes were observed on transpiration rate (E)of bean rusted leaves. Usually the fungi that cause rustwill rupture the leaf epidermis during sporulation,therefore E is increased (Duniway and Durbin, 1971;Owera et al., 1981). However, the E of rusted leaves ofwheat was smaller than that of the healthy leaves(McGrath and Pennypacker, 1990). The decreasedtranspiration rate (E) of leaves with angular leaf spotand anthracnose was directly related to decreased gse,and therefore it was associated with stomatal closure.For anthracnose, the reduction in gse was responsible fora lower Ci in diseased leaves and this reduction becamethe main limiting factor for the leaf capacity for CO2
assimilation (Bassanezi et al., 2000).Increased dark respiration rate (Rd) of rusted leaves
after the first symptom has already been reported (Dalyet al., 1961; Livne, 1964; Raggi, 1978; Lopes, 1999;Bassanezi et al., 2001). The respiratory rate reached amaximal value and stayed high during all the sporula-tion process, which meant that intense metabolic activityof the pathogen and host cells occurred at this stage.This fact is supported by the observations of Wagnerand Boyle (1995) who noted an increased acid invertaseactivity, starch hydrolysis, and glucose mobilizationduring the sporulation process of bean rust. Accordingto these researchers, production of uredosporesdemands a high efflux of metabolites and energy.Infected leaves with hemibiotrophic or necrotrophicpathogens often exhibit an increase in dark respirationrate (Rd) to supply the demand caused by accelerated
Gas Exchange and Chlorophyll Fluorescence Emission of Healthy and Diseased Bean Leaves 45

metabolic activity of host cells (Lucas, 1998). The Rd ofleaves with angular leaf spot rose rapidly soon aftersymptom appearance and then decreased. Similar resultswere observed in other pathosystems including hemi-biotrophic and necrotrophic fungi, such as Pyrenophorateres (Smedegaard-Petersen, 1984) and Rynchosporiumsecalis (Martin, 1986) in barley. An increase in Rd atinitial stages of disease was mainly a consequence ofhost-accelerated respiration, although the extension offungal participation is not clear (Lucas, 1998).
Knowledge of plant physiological processes and theway they are affected by a pathogen can be used toanalyse and predict the effect of the disease on cropgrowth and yield (Boote et al., 1980; Bastiaans, 1993).The contrasting effects of biotrophic and necrotrophicfungus on the reduction of photosynthesis have to betaken into account in the development of models thatforecast the damage caused by diseases. The use ofdisease severity as a predictor variable for crop damagemay not be accurate, especially for necrotrophic path-ogens. The effect of these pathogens on photosynthesisare proportionally greater than the correspondingreduction of healthy leaf area due to disease, with adirect impact on crop yield.
Acknowledgements
This research was supported by FAPESP (97/2308–8). Referees of
Journal of Phytopathology made valuable suggestions.
LiteratureBassanezi, R. B., L. Amorim, A. Bergamin Filho (2000): Analise das
trocas gasosas em feijoeiro com ferrugem, mancha angular eantracnose. Fitopatologia Brasileira 25, 643–650.
Bassanezi, R. B., L. Amorim, A. Bergamin Filho (2001): Eficienciafotossintetica de folhas de feijoeiro infectadas com o vırus domosaico-em-desenho, Uromyces appendiculatus e Phaeoisariopsisgriseola. Summa Phytopathologica 27, 5–11.
Bassanezi, R. B., M. C. Martins, C. V. Godoy, L. Amorim,A. Bergamin Filho (1997): Efeito da antracnose na eficienciafotossintetica do feijoeiro. Fitopatologia Brasileira 22, 520–524.
Bastiaans, L. (1991): Ratio between virtual and visual lesion size as ameasure to describe reduction in leaf photosynthesis of rice due toleaf blast. Phytopathology 81, 611–615.
Bastiaans, L. (1993): Effects of leaf blast on photosynthesis of rice.1. Leaf photosynthesis. Neth. J. Pl. Path. 99, 197–203.
Boote, K. J., J. W. Jones, G. H. Smerage, C. S. Barfield, R. D. Berger(1980): Photosynthesis of peanut canopies as affected by leafspotand artificial defoliation. Agron J. 72, 247–252.
Daly, J. M., A. A. Bell, L. R. Krupka (1961): Respiratory changesduring development of rust diseases. Phytopathology 51, 461–471.
Duniway, J. M., R. D. Durbin (1971): Some effects of Uromycesphaseoli on the transpiration rate and stomatal conductance ofbeans leaves. Phytopathology 61, 114–119.
Ephrath, J. E., D. Shtienberg, J. Drieshpoun, A. Dinoor, A. Marani(1989): Alternaria alternata in cotton (Gossypium hirsutum) cv.Acala: effects on gas exchange, yield components and yieldaccumulation. Neth. J. Plant Pathol. 95, 157–166.
Fulton, N. D., K. Bollenbacher, G. E. Templeton (1965): A metabolicfrom Alternaria tenuis that inhibits chlorophyll production. Phyto-pathology 55, 49–51.
Garry, G., M. H. Jeuffroy, B. Ney, B. Tivoli (1998): Effects ofAscochyta blight (Mycosphaerella pinodes) on the photosynthesizingleaf area and the photosynthetic efficiency of green leaf area ofdried-pea (Pisum sativum). Plant Pathol 47, 473–479.
Hall, R. (1991): Compendium of Bean Disease. APS Press, St.Paul, MN.
Hall, D. O., J. M. O. Scurlock, H. R. Bolhr-Nordenkampf, R. C.Leegood, S. P. Long (1993): Photosynthesis and Production in aChanging Environment: a Field and Laboratory Manual. Chapman& Hall, London.
Livne, A. (1964): Photosynthesis in healthy and rust affected plants.Plant Physiol. 39, 614–621.
Livne, A., J. M. Daly (1966): Translocation in healthy and rust-affected beans. Phytopathology 56, 170–175.
Lopes, D. B. (1999): Photosynthetic Competence of Bean Leaves withRust and Anthracnose. University of Florida, Gainesville, FL.
Lopes, D. B., R. D. Berger (2001): The effects of rust and anthracnoseon the photosynthetic competence of diseased bean leaves. Phyto-pathology 91, 212–220.
Lucas, J. A. (1998): Plant Pathology and Plant Pathogens. BlackwellScience, Oxford, UK.
Martin, P. J. (1986): Gaseous exchange studies of barley leaves infectedwith Rhynchosporium secalis (Oudem). J. J. Davis. Physiol. Mol.Plant Pathol. 28, 3–14.
McGrath, M. T., S. P. Pennypacker (1990): Alteration of physiologicalprocesses in wheat flag leaves caused by stem rust and leaf rust.Phytopathology 80, 677–686.
Meyer, S., K. Saccardy-Adji, F. Rizza, B. Genty (2001): Inhibition ofphotosynthesis by Colletotrichum lindemuthianum in bean leavesdetermined by chlorophyll fluorescence imaging. Plant, Cell Envi-ron. 24, 947–956.
Moll, S., P. Serrano, C. Boyle (1995): In vivo chlorophyll fluorescencein rust-infected bean plants. Angewandte Bot. 69, 163–168.
O’Connell, R. S. B. Perfect, R. Hughes, J. Carzaniga, J. Bailey (Green.(2000): Dissecting the cell biology of Colletotrichum infectionprocesses. In: Prusky, D., S. Freeman, M. B. Dickman (eds),Colletotrichum Host Specificity, Pathology, and Host–pathogenInteraction, pp. 57–77. APS Press, St. Paul, MN.
Owera, S. A. P., J. F. Farrar, R. Whitbread (1981): Growth andphotosynthesis in barley infected with brown rust. Physiol. PlantPath. 18, 79–90.
Peterson, R. B., D. E. Aylor (1995): Chlorophyll fluorescenceinduction in leaves of Phaseolus vulgaris infected with bean rust(Uromyces appendiculatus). Plant Physiol. 108, 163–171.
Raggi, V. (1978): The CO2 compensation point, photosynthesisand respiration in rust infected bean leaves. Physiol. Plant Path.13, 135–139.
Rasmussen, J. B., R. P. Scheffer (1988): Effects of selective toxin fromHelminthosporium carbonum on chlorophyll synthesis in maize.Physiol. Mol. Plant Pathol. 32, 283–291.
Schnabel, G., G. Strittmatter, G. Noga (1998): Changes in photosyn-thetic electron transport in potato cultivars with different fieldresistance after infection with Phytophthora infestans. J. Phyto-pathol. 146, 205–210.
Scholes, J. D., J. F. Farrar (1985): Photosynthesis and chloroplastfunctioning within individual pustules of Uromyces muscari onbluebell leaves. Physiol. Plant Path. 27, 387–400.
Scholes, J. D., S. A. Rolfe (1995): How do biotrophic pathogens affectthe photosynthetic metabolism of their hosts? Aspects Appl. Biol.42, 91–99.
Shtienberg, D. (1992): Effects of foliar diseases on gas exchangeprocesses: a comparative study. Phytopathology 82, 760–765.
Smedegaard-Petersen, V. (1984): The role of respiration and energygeneration in diseased and disease-resistant plants. In: Wood,R. K. S., J. G. Jellis (eds), Plant Diseases: Infection, Damage andLoss, pp. 73–85. Blackwell Scientific, Oxford, UK.
Stangarlin, J. R., S. F. Pascholati (2000): Atividades de ribulose-1,5-bifosfato carboxilase-oxigenase (rubisco), clorofilase, b-1,3-glucan-ase e quitinase e conteudo de clorofila em cultivares de feijoeiro(Phaseolus vulgaris) infectados com Uromyces appendiculatus.Summa Phytopathologica 26, 34–42.
Stangarlin, J. R., S. F. Pascholati, C. A. Labate (2000): Efeito dePhaeoisariopsis griseola na atividade de ribulose-1,5-bifosfato car-boxilase-oxigenase, clorofilase, b-1,3-glucanase e quitinase em cul-tivares de Phaseolus vulgaris. Fitopatologia Brasileira 25, 59–66.
Sziraki, I., L. A. Mustardy, A. Faludi-Daniel, Z. Kiraly (1984):Alterations in chloroplast ultrastructure and chlorophyll content in
46 BASSANEZI et al.

rust-infected Pinto Beans at different stages of disease development.Phytopathology 74, 77–84.
Wagner, S., C. Boyle (1995): Changes in carbohydrate, protein andchlorophyll content, and enzyme activity during switch fromuredinio- to teliospore sporulation in the bean-rust fungus Uromycesappendiculatus (Pers.) Link. J. Phytopathol. 143, 633–638.
Wijesundera, R. L. C., J. A. Bailey, R. J. W. Byrde, A. H. Fielding(1989): Cell wall degrading enzymes of Colletotrichum lindemuthia-num: their role in the development of bean anthracnose. Physiol.Mol. Plant Pathol. 34, 403–413.
Wong, P. Y. O., L. B. Thrower (1978a): Effect of Colletotrichumlindemuthianum on photosynthesis and respiration of Vignasesquipedalis. J. Plant Path. 92, 88–94.
Wong, P. Y. O., L. B. Thrower (1978b): Sugar metabolism andtranslocation in Vigna sesquipedalis infected by Colletotrichumlindemuthianum. J. Plant Path. 92, 102–112.
Wright, D. P., B. C. Baldwin, M. C. Shephard, J. D. Scholes (1995):Source-sink relationships in wheat leaves infected with powderymildew. II. Changes in the regulation of the Calvin cycle. Physiol.Mol. Plant Pathol. 47, 255–267.
Zaki, A. I., R. D. Durbin (1965): The effect of bean rust on thetranslocation of photosynthetic products from diseased leaves.Phytopathology 55, 528–529.
Gas Exchange and Chlorophyll Fluorescence Emission of Healthy and Diseased Bean Leaves 47
Related Documents











