Molecular Immunology 44 (2007) 1015–1028 Gangliosides, Ab1 and Ab2 antibodies II. Light versus heavy chain: An idiotype-anti-idiotype case study Alejandro L ´ opez-Requena a , Mabel Rodr´ ıguez a , Cristina Mateo de Acosta a , Ernesto Moreno b , Yaquelin Puchades c , Majela Gonz´ alez a,c , Ariel Talavera b , Aisel Valle a , Tays Hern´ andez a , Ana Mar´ ıa V´ azquez a , Rolando P´ erez a,∗ a Department of Antibody Engineering, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba b Department of Computational and Structural Biology, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba c Pharmaceutical Division, Center for Genetic Engineering and Biotechnology, P.O. Box 6162, Havana 11600, Cuba Received 21 January 2006; received in revised form 4 March 2006; accepted 9 March 2006 Available online 18 April 2006 Abstract The antibody heavy chain is generally more important than the light chain for the interaction with the antigen, although many reports demonstrate the influence of the light chain in the antibody binding properties. The heavy chains of anti-N-glycolyl-ganglioside P3 mAb and anti-idiotypic 1E10 mAb display complementary charged residues in their H-CDRs, particularly in H-CDR3. A basic residue in P3 mAb H-CDR1 was shown to be crucial for the interaction with the antigen and 1E10 mAb. The immunogenetic features of three other P3 mAb anti-idiotypic mAbs are now analyzed. One of them bears the same heavy chain as 1E10 mAb and a different light chain, but differs in its binding to P3 mAb mutants where H-CDR basic residues were replaced and in the binding to 1E10-specific phagotopes. Chimeric hybrid antibodies with P3 and 1E10 mAb heavy chains and unrelated light chains were obtained to further determine the importance of heavy chains in P3 and 1E10 mAb binding properties. One of the P3 heavy chain hybrid antibodies retained the specificity of P3 mAb with slight affinity differences. The heavy chains appear to play the main role in these mAb interactions, with the light chains modulating the affinity to their ligands. © 2006 Elsevier Ltd. All rights reserved. Keywords: Heavy chain; Light chain; N-Glycolyl-gangliosides; Anti-idiotypic antibodies 1. Introduction Heavy chain is generally regarded as protagonist in antibody specificity. There are several examples of VH alone binding to the antigen, without the participation of any light chain variable region (VL) (Hamers-Casterman et al., 1993; Noel et al., 1996; Reiter et al., 1999; Tanha et al., 2001). Natural and engineered homodimers of VH domains with binding activity have recently been described (Sepulveda et al., 2003; Jin et al., 2003). The heavy chain variable region (VH) plays an important role for most of the molecular interactions with the antigen, as revealed by the crystal structures of several antibody–antigen ∗ Corresponding author. Tel.: +53 7 2716810; fax: +53 7 2720644. E-mail address: [email protected] (R. P´ erez). complexes (Morea et al., 1998; Xu and Davis, 2000). Among the few anti-carbohydrate antibodies with available crystal struc- tures, two of them display heavy chains having a preponderant role in antigen binding (Calarese et al., 2003; Krengel et al., 2003; Rojas et al., 2004). These two anti-carbohydrate anti- bodies have in common a long H-CDR3 (16 and 14 amino acids, respectively) that stands in the middle of the binding site dividing it into two regions. In both cases, the anchoring of the carbohydrate antigen happens in a pocket formed on the VH side. Heavy chains were considered to be the main responsible for the binding properties also for an anti-N-glycolyl-ganglioside mAb, named P3 (V´ azquez et al., 1995), and for its anti-idiotypic mAb, called 1E10 (V´ azquez et al., 1998), based on several experimental facts as well as theoretical analyses. P3 mAb recognizes N-glycolylated gangliosides (V´ azquez et al., 1995), 0161-5890/$ – see front matter © 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.molimm.2006.03.004

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A
t1taHcom©
K
1
strRhb
rr
0d
Molecular Immunology 44 (2007) 1015–1028
Gangliosides, Ab1 and Ab2 antibodiesII. Light versus heavy chain: An idiotype-anti-idiotype case study
Alejandro Lopez-Requena a, Mabel Rodrıguez a, Cristina Mateo de Acosta a,Ernesto Moreno b, Yaquelin Puchades c, Majela Gonzalez a,c,
Ariel Talavera b, Aisel Valle a, Tays Hernandez a,Ana Marıa Vazquez a, Rolando Perez a,∗
a Department of Antibody Engineering, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cubab Department of Computational and Structural Biology, Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba
c Pharmaceutical Division, Center for Genetic Engineering and Biotechnology, P.O. Box 6162, Havana 11600, Cuba
Received 21 January 2006; received in revised form 4 March 2006; accepted 9 March 2006Available online 18 April 2006
bstract
The antibody heavy chain is generally more important than the light chain for the interaction with the antigen, although many reports demonstratehe influence of the light chain in the antibody binding properties. The heavy chains of anti-N-glycolyl-ganglioside P3 mAb and anti-idiotypicE10 mAb display complementary charged residues in their H-CDRs, particularly in H-CDR3. A basic residue in P3 mAb H-CDR1 was showno be crucial for the interaction with the antigen and 1E10 mAb. The immunogenetic features of three other P3 mAb anti-idiotypic mAbs are nownalyzed. One of them bears the same heavy chain as 1E10 mAb and a different light chain, but differs in its binding to P3 mAb mutants where-CDR basic residues were replaced and in the binding to 1E10-specific phagotopes. Chimeric hybrid antibodies with P3 and 1E10 mAb heavy
hains and unrelated light chains were obtained to further determine the importance of heavy chains in P3 and 1E10 mAb binding properties. Onef the P3 heavy chain hybrid antibodies retained the specificity of P3 mAb with slight affinity differences. The heavy chains appear to play theain role in these mAb interactions, with the light chains modulating the affinity to their ligands.2006 Elsevier Ltd. All rights reserved.
eywords: Heavy chain; Light chain; N-Glycolyl-gangliosides; Anti-idiotypic antibodies
cttr2baso
. Introduction
Heavy chain is generally regarded as protagonist in antibodypecificity. There are several examples of VH alone binding tohe antigen, without the participation of any light chain variableegion (VL) (Hamers-Casterman et al., 1993; Noel et al., 1996;eiter et al., 1999; Tanha et al., 2001). Natural and engineeredomodimers of VH domains with binding activity have recentlyeen described (Sepulveda et al., 2003; Jin et al., 2003).
The heavy chain variable region (VH) plays an importantole for most of the molecular interactions with the antigen, asevealed by the crystal structures of several antibody–antigen
∗ Corresponding author. Tel.: +53 7 2716810; fax: +53 7 2720644.E-mail address: [email protected] (R. Perez).
V
tmmer
161-5890/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.molimm.2006.03.004
omplexes (Morea et al., 1998; Xu and Davis, 2000). Amonghe few anti-carbohydrate antibodies with available crystal struc-ures, two of them display heavy chains having a preponderantole in antigen binding (Calarese et al., 2003; Krengel et al.,003; Rojas et al., 2004). These two anti-carbohydrate anti-odies have in common a long H-CDR3 (16 and 14 aminocids, respectively) that stands in the middle of the bindingite dividing it into two regions. In both cases, the anchoringf the carbohydrate antigen happens in a pocket formed on theH side.Heavy chains were considered to be the main responsible for
he binding properties also for an anti-N-glycolyl-ganglioside
Ab, named P3 (Vazquez et al., 1995), and for its anti-idiotypicAb, called 1E10 (Vazquez et al., 1998), based on severalxperimental facts as well as theoretical analyses. P3 mAbecognizes N-glycolylated gangliosides (Vazquez et al., 1995),

1 lar Im
w(ga2Cabt(
t(wP
2
2
cRN4m(ss
2
akmGag1
2
usag(2mSpfiefilp
ec
2a
w
((
V
(
(
V
(
V
(
2
bc
2
Du
2o
PaoTrp
016 A. Lopez-Requena et al. / Molecu
hose expression has been associated to human malignanciesMalykh et al., 2001). Due to the low immunogenicity of theselycolipids, anti-idiotypic antibodies have been developed as anlternative for cancer therapy (Bhattacharya-Chatterjee et al.,000; Foon et al., 2000; Alfonso et al., 2002; Dıaz et al., 2003;hapman, 2003). 1E10, 3B11, 4F2 (Vazquez et al., 1998), 3F9nd 4C8 mAbs (our unpublished data) are anti-idiotypic anti-odies specific for P3 mAb. 1E10 mAb has been used in clinicalrials with melanoma (Alfonso et al., 2002) and breast cancerDıaz et al., 2003) patients, with promising results.
Previous results suggested that heavy chains play an impor-ant role in the idiotypic interactions of P3 and 1E10 mAbsPerez et al., 2001; Lopez-Requena et al., 2007). In the presentork, we assessed the contribution of heavy and light chain of3 and 1E10 mAbs to their molecular interactions.
. Materials and methods
.1. Cells
NS0 transfectomas expressing chimeric P3 antibody,himeric P3 light chain, chimeric 1E10 light chain (Lopez-equena et al., 2003) and chimeric T1 light chain (Roque-avarro et al., 2003), and hybridomas producing 1E10, 3B11,F2 (Vazquez et al., 1998), 4C8 and 3F9 (our unpublished data)Abs were cultured in Dulbecco’s modified Eagles medium
DMEM) supplemented with 10% heat inactivated fetal calferum (FCS), antibiotic mixtures of penicillin (100 U/mL) andtreptomycin (100 �g/mL), and 2 mM l-glutamine.
.2. Gangliosides
GM3(NeuGc) and GM2(NeuGc) gangliosides, namedccording to the nomenclature of Svennerholm (1964), wereindly provided by Dr. L.E. Fernandez, Vaccine Depart-ent, Center of Molecular Immunology (Havana, Cuba).M3(NeuGc) ganglioside was isolated from horse erythrocytes
s previously described (Stults et al., 1989). GM2(NeuGc) gan-lioside was purified from Balb/c mouse liver (Hashimoto et al.,983).
.3. Monoclonal antibodies (mAbs)
1E10, 3B11, 4F2 (Vazquez et al., 1998), 4C8 and 3F9 (ournpublished data) mAbs, all murine anti-idiotypic IgG (IgG1, �)pecific for P3 mAb, 4G9 mAb (Rodrıguez et al., 2003), murinenti-idiotypic IgG (IgG1, �) specific for anti-GM3(NeuGc) gan-lioside 14F7 mAb (Carr et al., 2000) and chimeric P3, 1E10Lopez-Requena et al., 2003) and T1 (Roque-Navarro et al.,003) antibodies were purified by Protein-A Affinity Chro-atography (Pharmacia, Uppsala, Sweden) and analyzed byDS-PAGE under reducing conditions. Murine antibodies wereurified from ascitic fluid, while chimeric antibodies were puri-ed from transfectoma culture supernatants. P3 mAb (Vazquez
t al., 1995) (IgM, �) was purified from ascitic fluid by gelltration chromatography using a Sephacryl S-300 high reso-ution column (Pharmacia, Uppsala, Sweden) equilibrated withhosphate-buffered saline (PBS) containing 0.5 M NaCl.
c5cP
munology 44 (2007) 1015–1028
The specificity of the purified antibodies was confirmed bynzyme-linked immunoadsorbent assay (ELISA) and proteinoncentration estimated by optical density at 280 nm.
.4. Primers for cDNA synthesis and variable regionsmplification by polymerase chain reaction (PCR)
As primers for cDNA synthesis, the chain-specific 3′ primersere used.V� region 5′ primers:
1) 5′-GAC ATT CAG CTG ACC CAG TCT CCA-3′;2) 5′-GGG GAT ATC CACC ATG GAT TTT CAG GTG CAG
ATT TTC AG-3′;
� region 3′ primers:
3) 5′-GGA AGC TTG AAG ATG GAT ACA GTT GGT GCAGC-3′;
4) 5′-AGC GTC GAC TTACG TTT (TG)AT TTC CA(GA)CTT (GT)GT-3′;
H region 5′ primer:
5) 5′-CAGG T(T/G/C)(A/C) A(A/G)C TGC AG(G/C) AGTC(A/T)G G-3′;
H region 3′ primer:
6) 5′-GGAAGCTTAGACAGATGGGGGTGTCGTTTTG-3′.
.5. Vectors
The pMOS-Blue vector for cloning and sequencing of anti-ody variable regions was purchased from Amersham Pharma-ia, UK.
.6. Selective medium
For the selection of whole antibody-producing transfectomas,MEM-F12 containing 10% FCS and histidinol at 10 mM wassed as selective medium.
.7. Purification of phagotopes recognized by chimeric P3r 1E10 antibodies from the pVIII-12aa displayed library
12mer peptides displayed on filamentous phage, specific for3 or 1E10 mAbs, were previously selected (Lopez-Requena etl., 2007). The phages were amplified by infecting 0.4 mL of anvernight culture of Escherichia coli TG1 cells with 104 AmpR-U of the phage, and superinfecting with 1011 kanamicine
esistance transducing unit (KanR-TU) M13K07 helper phagearticles. The cells were grown in Luria Bertani (LB) medium
ontaining 100 �g/mL Amp, 70 �g/mL Kan and 1 mM IPTG forh at 37 ◦C with vigorous shaking. The phage suspension wasentrifuged and the supernatant phages precipitated with 4%EG 8000/0.5 M NaCl.
lar Im
2
c(bs1shbmp
mR
2
w7rop(rmr
aDGcpbk
2
ccfpcbftR
2
Vgd
wcgbt5wActc(wo
bc2nscfcmiGwmcc
a(tiVPP
fio
2r
awnrwmt
A. Lopez-Requena et al. / Molecu
.8. Phage ELISAs
The ELISA assay used to measure the recognition of phagelones by the P3-specific Ab2 mAbs was previously describedLopez-Requena et al., 2007). In the case of chimeric hybrid anti-odies, plates were coated with goat anti-human IgG (�-chain-pecific) antibody (Sigma, St. Louis, MO) at a concentration of0 �g/mL. After blocking, plates were incubated for 1 h withupernatants from the transfectomas producing the chimericybrid antibodies or the chimeric “wild” antibodies, at an anti-ody concentration of 3 �g/mL. For the cross-recognition of P3Ab heavy chain by the 1E10-specific phagotopes, different
hage particle numbers/well were assayed.Inhibition of phage binding to P3 mAb by its anti-idiotypic
Abs was measured by ELISA, as previously described (Lopez-equena et al., 2007).
.9. Isolation of V-genes
Approximately 107 of antibody-producing hybridoma cellsere pelleted, washed twice with PBS, and quickly frozen at0 ◦C until use. Total cellular RNA was extracted with Trizoleagent (Gibco-BRL, Paisley, UK). cDNA synthesis from 5 �gf total RNA and PCR amplification of VH and V� genes wereerformed by using the Access RT-PCR System from PromegaMadison, WI). The primers for cDNA synthesis/PCR of V�egion of 3F9 mAb were 1 and 3; and 2 and 4 for 4C8 and 4F2Abs. Primers 5 and 6 were used for the amplification of VH
egions of the three mAbs.PCR products were verified by agarose gel electrophoresis in
1.5% gel stained with ethidium bromide. Amplified V� and VHNAs were purified using Qiaquick Gel Extraction Kit (QiagenmbH, Hilden, Germany). The purified PCRs products were
loned into pMOS-Blue vector, following instructions of therovider (Amersham Pharmacia, UK). Clones were sequencedy the dideoxy method (Sanger et al., 1977) using T7 DNA Polit (Pharmacia).
.10. Chimeric antibody expression
NS0 transfectoma cells expressing chimeric P3 light chain,himeric 1E10 light chain (Lopez-Requena et al., 2003) orhimeric T1 light chain (Roque-Navarro et al., 2003) were trans-ected by electroporation with 10 �g of maxiprep DNA fromAH4604 expression vector containing chimeric 1E10 heavyhain, chimeric P3 heavy chain (Lopez-Requena et al., 2003) oroth, respectively, linearized with Pvu I digestion. The trans-ection method and the selection of transfectomas secretinghe chimeric IgG were previously described in detail (Lopez-equena et al., 2007).
.11. Antibody binding assays
Binding of chimeric P3 antibody and chimericHP3/V�1E10 and VHP3/V�T1 hybrid antibodies to gan-liosides was determined using an ELISA assay as previouslyescribed (Alfonso et al., 1995; Lopez-Requena et al., 2007),
Bss1
munology 44 (2007) 1015–1028 1017
ith the diluted supernatants from transfectomas producing thehimeric antibodies and an alkaline phosphatase-conjugatedoat anti-human IgG (�-chain-specific) (Sigma) as second anti-ody. For the experiments of inhibition of P3 mAb binding tohe ganglioside by Ab2 mAbs, aliquots of murine P3 mAb with0% maximum reactivity against GM3(NeuGc) gangliosideere mixed with equal volumes of different concentrations ofb2 mAbs and the mixtures were added to microtiter plates
oated with GM3(NeuGc) ganglioside. The binding of P3 mAbo the ganglioside was detected using an alkaline phosphatase-onjugated goat anti-mouse IgM (�-chain-specific) antibodySigma) diluted 1:1000. Percent inhibition of P3 mAb bindingas calculated relative to the binding of P3 mAb in the absencef inhibitor.
Ab2 cross-blocking experiments were performed usingiotinylated anti-idiotypic mAbs. The protocol for mAb–biotinonjugation was previously described (Lopez-Requena et al.,007). Non-saturating amount of biotinylated Ab2 mAb andon-biotinylated blocking Ab2 mAb (serial four-fold dilutionstarting at 100 �g/mL) were added together to microtiter platesoated overnight at 4 ◦C with 10 �g/mL of P3 mAb and blockedor 1 h at 37 ◦C with PBS-T-BSA (phosphate-buffered salineontaining 0.05% Tween 20, pH 7.5, and 1% bovine serum albu-in). After 2 h of incubation at 37 ◦C, plates were washed and
ncubated with avidin–biotin–peroxidase complex (Dakopatts,lostrup, Denmark) for 1 h at 37 ◦C. Plates were developedith the o-phenylendiamine substrate solution and absorbanceonitored at 492 nm. The results were plotted, and the con-
entrations to achieve 50% inhibition of binding (IC50) werealculated.
Binding of P3 mAb H-CDR R → S mutant antibodies to P3nti-idiotypic mAbs was determined as previously describedLopez-Requena et al., 2007), with the diluted supernatants fromransfectomas producing the chimeric mutant antibodies. A sim-lar ELISA assay was used for determining reactivity of chimericH1E10/V�P3 and VH1E10/V�T1 hybrid antibodies against3 mAb, and chimeric VHP3/V�1E10 and VHP3/V�T1 against3 mAb Ab2 mAbs.
Three samples of each experiment were tested and the coef-cient of variation was <10% for all assays. Background valuesf absorbance were less than 0.1.
.12. Immunogenetic analysis of the antibody variableegions
Individual VH and VL nucleotide sequences were searchedgainst the EMBL/GenBank database for sequence similarityith known murine genes. The germ-line gene with the highestucleotide identity was analyzed as a possible origin of the rear-anged gene. The J� and JH regions sequenced were comparedith the five known murine J� genes and with the four knownurine JH genes (Kabat et al., 1991). The analysis of the junc-
ional sequences in VH sequences was performed as reported by
angs et al. (1991). D region nucleotides were identified afterequence alignment of heavy chain CDR3 (H-CDR3) nucleotideequences with known murine D minigenes (Kabat et al.,991).
1 lar Immunology 44 (2007) 1015–1028
2
fevP
3
34
bwi(rPui
bfcitwPm4fitbf
FAwwmrc
Table 1Cross-inhibition analyses of Ab2 mAb binding to Ab1 P3 mAba
IC50 inhibitor mAb/IC50 reference mAbb
Inhibitory unlabeled mAb Biotynilated mAbs
1E10 3F9 4C8 4F2 3B11
1E10 1.00 1.73 13.90 29.05 –3F9 0.60 1.00 4.57 17.77 –4C8 0.44 0.36 1.00 2.71 –4F2 0.06 0.22 1.19 1.00 –3B11 0.11 0.30 0.41 0.24 1.00
a Data from cross-inhibition experiments of 1E10, 4F2 and 3B11 mAbs witheach other were taken from Vazquez et al. (1998).
b The results are given as the ratio of IC50 observed for the inhibitor mAbdl
3h
2sBnod
iStabt
018 A. Lopez-Requena et al. / Molecu
.13. Model of antibody Fv fragments
Computer models of antibody variable regions were builtrom the amino acid sequences using the web antibody mod-lling (WAM) server (Whitelegg and Rees, 2000). Models wereisualized with the program Swiss PDB Viewer (Guex andeitsch, 1997).
. Results
.1. Immunochemical characterization of Ab2 3F9 andC8 mAbs
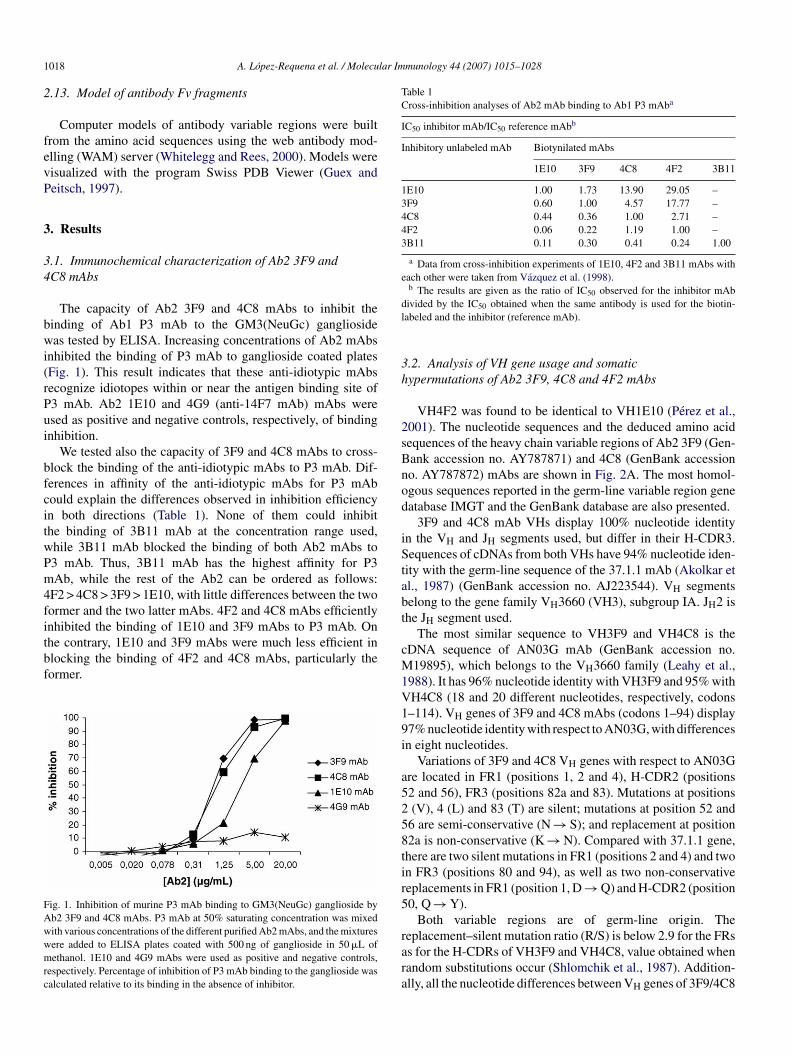
The capacity of Ab2 3F9 and 4C8 mAbs to inhibit theinding of Ab1 P3 mAb to the GM3(NeuGc) gangliosideas tested by ELISA. Increasing concentrations of Ab2 mAbs
nhibited the binding of P3 mAb to ganglioside coated platesFig. 1). This result indicates that these anti-idiotypic mAbsecognize idiotopes within or near the antigen binding site of3 mAb. Ab2 1E10 and 4G9 (anti-14F7 mAb) mAbs weresed as positive and negative controls, respectively, of bindingnhibition.
We tested also the capacity of 3F9 and 4C8 mAbs to cross-lock the binding of the anti-idiotypic mAbs to P3 mAb. Dif-erences in affinity of the anti-idiotypic mAbs for P3 mAbould explain the differences observed in inhibition efficiencyn both directions (Table 1). None of them could inhibithe binding of 3B11 mAb at the concentration range used,hile 3B11 mAb blocked the binding of both Ab2 mAbs to3 mAb. Thus, 3B11 mAb has the highest affinity for P3Ab, while the rest of the Ab2 can be ordered as follows:
F2 > 4C8 > 3F9 > 1E10, with little differences between the twoormer and the two latter mAbs. 4F2 and 4C8 mAbs efficiently
nhibited the binding of 1E10 and 3F9 mAbs to P3 mAb. Onhe contrary, 1E10 and 3F9 mAbs were much less efficient inlocking the binding of 4F2 and 4C8 mAbs, particularly theormer.ig. 1. Inhibition of murine P3 mAb binding to GM3(NeuGc) ganglioside byb2 3F9 and 4C8 mAbs. P3 mAb at 50% saturating concentration was mixedith various concentrations of the different purified Ab2 mAbs, and the mixturesere added to ELISA plates coated with 500 ng of ganglioside in 50 �L ofethanol. 1E10 and 4G9 mAbs were used as positive and negative controls,
espectively. Percentage of inhibition of P3 mAb binding to the ganglioside wasalculated relative to its binding in the absence of inhibitor.
cM1V19i
a5258tir5
rara
ivided by the IC50 obtained when the same antibody is used for the biotin-abeled and the inhibitor (reference mAb).
.2. Analysis of VH gene usage and somaticypermutations of Ab2 3F9, 4C8 and 4F2 mAbs
VH4F2 was found to be identical to VH1E10 (Perez et al.,001). The nucleotide sequences and the deduced amino acidequences of the heavy chain variable regions of Ab2 3F9 (Gen-ank accession no. AY787871) and 4C8 (GenBank accessiono. AY787872) mAbs are shown in Fig. 2A. The most homol-gous sequences reported in the germ-line variable region geneatabase IMGT and the GenBank database are also presented.
3F9 and 4C8 mAb VHs display 100% nucleotide identityn the VH and JH segments used, but differ in their H-CDR3.equences of cDNAs from both VHs have 94% nucleotide iden-
ity with the germ-line sequence of the 37.1.1 mAb (Akolkar etl., 1987) (GenBank accession no. AJ223544). VH segmentselong to the gene family VH3660 (VH3), subgroup IA. JH2 ishe JH segment used.
The most similar sequence to VH3F9 and VH4C8 is theDNA sequence of AN03G mAb (GenBank accession no.19895), which belongs to the VH3660 family (Leahy et al.,
988). It has 96% nucleotide identity with VH3F9 and 95% withH4C8 (18 and 20 different nucleotides, respectively, codons–114). VH genes of 3F9 and 4C8 mAbs (codons 1–94) display7% nucleotide identity with respect to AN03G, with differencesn eight nucleotides.
Variations of 3F9 and 4C8 VH genes with respect to AN03Gre located in FR1 (positions 1, 2 and 4), H-CDR2 (positions2 and 56), FR3 (positions 82a and 83). Mutations at positions(V), 4 (L) and 83 (T) are silent; mutations at position 52 and
6 are semi-conservative (N → S); and replacement at position2a is non-conservative (K → N). Compared with 37.1.1 gene,here are two silent mutations in FR1 (positions 2 and 4) and twon FR3 (positions 80 and 94), as well as two non-conservativeeplacements in FR1 (position 1, D → Q) and H-CDR2 (position0, Q → Y).
Both variable regions are of germ-line origin. Theeplacement–silent mutation ratio (R/S) is below 2.9 for the FRs
s for the H-CDRs of VH3F9 and VH4C8, value obtained whenandom substitutions occur (Shlomchik et al., 1987). Addition-lly, all the nucleotide differences between VH genes of 3F9/4C8
A. Lopez-Requena et al. / Molecular Immunology 44 (2007) 1015–1028 1019
Fig. 2. (A) Nucleotide and deduced amino acid sequences of the cDNA encoding the heavy chain variable region of anti-idiotypic 3F9 (VH3F9) and 4C8 (VH4C8)mAbs compared with the most closely related germ-line and cDNA sequences (see text). Amino acids are numbered according to Kabat. Spaces have been introducedto maximize alignment. Dashed lines indicate nucleotide sequence identities. The amino acid residue encoded by each codon is given above the nucleotide sequence.(B) Possible origins of heavy chain CDR3 of anti-idiotypic 3F9 and 4C8 mAbs. N nucleotides are represented with asterisk. Dashed lines indicate nucleotide sequenceidentities with known D and J genes. The germ-line sequences of DSP2.2 and J genes are from Kabat database.

1020 A. Lopez-Requena et al. / Molecular Immunology 44 (2007) 1015–1028
Fig. 3. Nucleotide and deduced amino acid sequences of the cDNA encoding the light chain variable region of anti-idiotypic 3F9 (A), and 4C8 and 4F2 (B) mAbscompared with the most closely related germ-line sequences (see text and legend from Fig. 2 for details). In the case of V�3F9 (A), the comparison with the mostclosely related cDNA sequence is also included.
lar Immunology 44 (2007) 1015–1028 1021
as
3
binotm(D9cBba
oi
3o
lA4F
naoatABiw3(
1ns
3a
aPFwm
Fig. 4. Reactivity of 1E10-specific phagotopes with other P3 mAb anti-idiotypicm1c
3b
3psi5ct7
3m
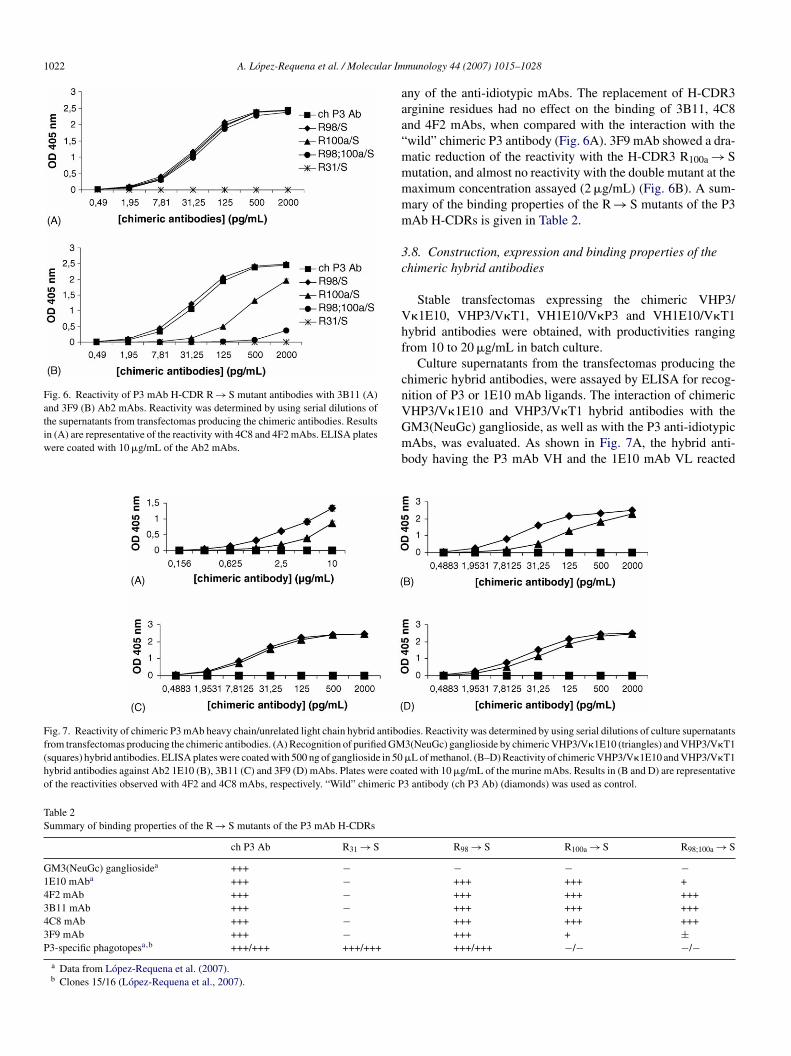
The recognition of P3 mAb H-CDR R → S mutant antibod-ies by the P3 anti-idiotypic mAbs was tested by ELISA (Fig. 6).The H-CDR1 R31 → S mutant antibody was not recognized by
Fig. 5. Inhibition of P3-specific phagotope binding to P3 mAb by P3 mAb
A. Lopez-Requena et al. / Molecu
nd 37.1.1 or AN03G are also present in several other similarequences of the GenBank database (not shown).
.3. Analysis of VH junctional regions
Nine (37%) out of 24 nucleotides coding for 3F9 H-CDR3elong to the D region, and 10 (42%) to JH2, while the remain-ng 5 (21%) are N nucleotides. For the 4C8 H-CDR3 (30ucleotides), the distribution is 11 (37%), 10 (33%, the samef 3F9 ones), 9 (30%), respectively (Fig. 2B). Both VHs usedhe DSP2.2 segment in the reading frame 3. The VDJ rearrange-
ent in VH3F9 uses 9 of the 17 nucleotides of DSP2.2 minigenenucleotides 4–12), while VH4C8 uses its final 11 nucleotides.SP2.2 minigene codes from the final two nucleotides of codon6 to the first nucleotide of codon 99 in VH3F9. In VH4C8, itodes from the two final nucleotides of codon 95 to codon 98.oth fragments are involved in the coding of four amino acids,ut integrally only in three (96–98) in VH4C8, and in two (97nd 98) in VH3F9.
TdT catalyzed-N additions correspond to four N5′ and onlyne N3′ nucleotide in VH3F9. The last ones are more frequentn VH4C8: eight, while it has only one N5′ nucleotide.
.4. Analysis of VL gene usage and somatic hypermutationsf Ab2 3F9, 4C8 and 4F2 mAbs
The nucleotide and deduced amino acid sequences ofight chain variable region (V�) 3F9 (GenBank accession no.Y803754), 4C8 (GenBank accession no. AY787869) andF2 (GenBank accession no. AY787870) mAbs are shown inig. 3.
V�3F9 belongs to the gene family V�23, and shares a 98.2%ucleotide identity with the germ-line gene 23–43 (GenBankccession no. AJ235973) (Schaeble et al., 1999). The changesbserved in the nucleotide sequence lead only to an aminocid replacement at position 3 (V → Q). The rest of the muta-ions are silent. The Naid 60 mAb VL (GenBank accession no.Y661814) (Park et al., unpublished), is 99% similar to V�3F9.oth sequences are identical, except for two amino acid changes
n L-CDR3. Nevertheless, these replacements are not observedhen comparing V�3F9 with 23–43 gene, which suggests thatF9 mAb light chain variable region is a new germ-line geneFig. 3A). The J� segment used is J�1.
V�4C8 is identical to V�4F2, from the V�4/5 family, and are00% identical to the am4 germ-line gene (GenBank accessiono. AJ231219) (Schaeble et al., 1999) (Fig. 3B). J�1 is the J�
egment used.
.5. Interaction of 1E10-specific phagotopes with P3nti-idiotypic mAbs
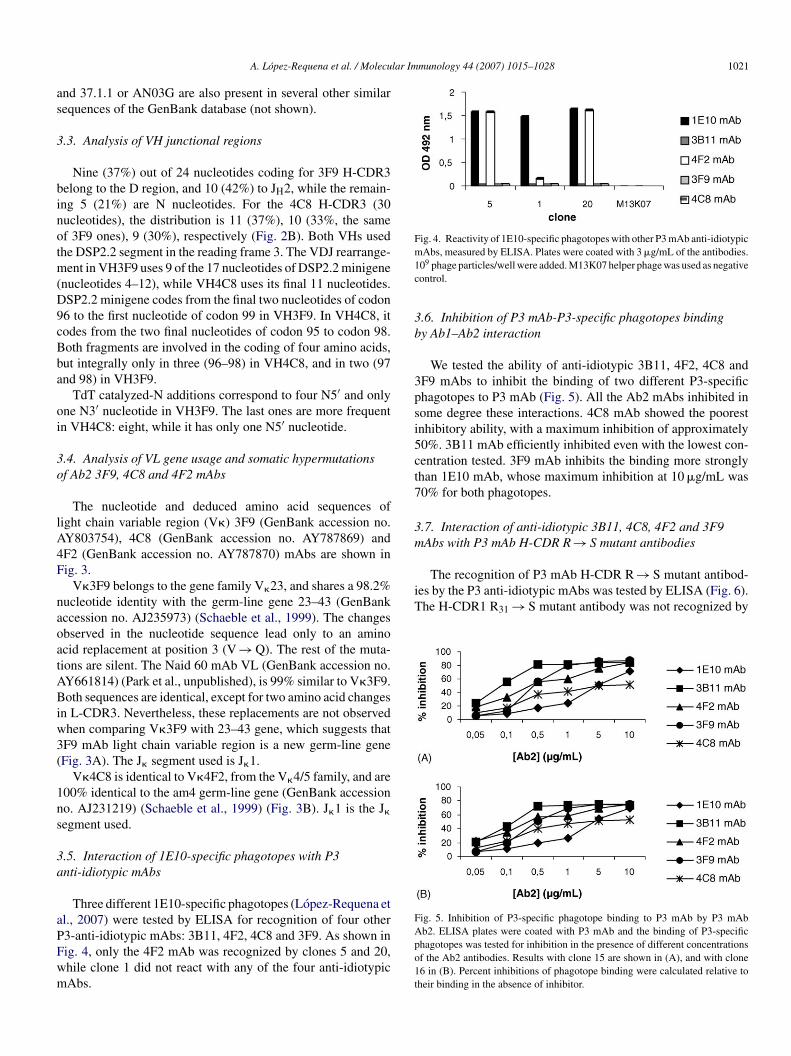
Three different 1E10-specific phagotopes (Lopez-Requena etl., 2007) were tested by ELISA for recognition of four other
3-anti-idiotypic mAbs: 3B11, 4F2, 4C8 and 3F9. As shown inig. 4, only the 4F2 mAb was recognized by clones 5 and 20,hile clone 1 did not react with any of the four anti-idiotypicAbs.Apo1t
Abs, measured by ELISA. Plates were coated with 3 �g/mL of the antibodies.09 phage particles/well were added. M13K07 helper phage was used as negativeontrol.
.6. Inhibition of P3 mAb-P3-specific phagotopes bindingy Ab1–Ab2 interaction
We tested the ability of anti-idiotypic 3B11, 4F2, 4C8 andF9 mAbs to inhibit the binding of two different P3-specifichagotopes to P3 mAb (Fig. 5). All the Ab2 mAbs inhibited inome degree these interactions. 4C8 mAb showed the poorestnhibitory ability, with a maximum inhibition of approximately0%. 3B11 mAb efficiently inhibited even with the lowest con-entration tested. 3F9 mAb inhibits the binding more stronglyhan 1E10 mAb, whose maximum inhibition at 10 �g/mL was0% for both phagotopes.
.7. Interaction of anti-idiotypic 3B11, 4C8, 4F2 and 3F9Abs with P3 mAb H-CDR R → S mutant antibodies
b2. ELISA plates were coated with P3 mAb and the binding of P3-specifichagotopes was tested for inhibition in the presence of different concentrationsf the Ab2 antibodies. Results with clone 15 are shown in (A), and with clone6 in (B). Percent inhibitions of phagotope binding were calculated relative toheir binding in the absence of inhibitor.
1022 A. Lopez-Requena et al. / Molecular Im
Fig. 6. Reactivity of P3 mAb H-CDR R → S mutant antibodies with 3B11 (A)and 3F9 (B) Ab2 mAbs. Reactivity was determined by using serial dilutions ofthe supernatants from transfectomas producing the chimeric antibodies. Resultsin (A) are representative of the reactivity with 4C8 and 4F2 mAbs. ELISA plateswere coated with 10 �g/mL of the Ab2 mAbs.
aaa“mmmmm
3c
Vhf
cnVGmb
Fig. 7. Reactivity of chimeric P3 mAb heavy chain/unrelated light chain hybrid antibofrom transfectomas producing the chimeric antibodies. (A) Recognition of purified GM(squares) hybrid antibodies. ELISA plates were coated with 500 ng of ganglioside in 50hybrid antibodies against Ab2 1E10 (B), 3B11 (C) and 3F9 (D) mAbs. Plates were coaof the reactivities observed with 4F2 and 4C8 mAbs, respectively. “Wild” chimeric P
Table 2Summary of binding properties of the R → S mutants of the P3 mAb H-CDRs
ch P3 Ab R31 → S
GM3(NeuGc) gangliosidea +++ −1E10 mAba +++ −4F2 mAb +++ −3B11 mAb +++ −4C8 mAb +++ −3F9 mAb +++ −P3-specific phagotopesa,b +++/+++ +++/+++
a Data from Lopez-Requena et al. (2007).b Clones 15/16 (Lopez-Requena et al., 2007).
munology 44 (2007) 1015–1028
ny of the anti-idiotypic mAbs. The replacement of H-CDR3rginine residues had no effect on the binding of 3B11, 4C8nd 4F2 mAbs, when compared with the interaction with thewild” chimeric P3 antibody (Fig. 6A). 3F9 mAb showed a dra-atic reduction of the reactivity with the H-CDR3 R100a → Sutation, and almost no reactivity with the double mutant at theaximum concentration assayed (2 �g/mL) (Fig. 6B). A sum-ary of the binding properties of the R → S mutants of the P3Ab H-CDRs is given in Table 2.
.8. Construction, expression and binding properties of thehimeric hybrid antibodies
Stable transfectomas expressing the chimeric VHP3/�1E10, VHP3/V�T1, VH1E10/V�P3 and VH1E10/V�T1ybrid antibodies were obtained, with productivities rangingrom 10 to 20 �g/mL in batch culture.
Culture supernatants from the transfectomas producing thehimeric hybrid antibodies, were assayed by ELISA for recog-ition of P3 or 1E10 mAb ligands. The interaction of chimeric
HP3/V�1E10 and VHP3/V�T1 hybrid antibodies with theM3(NeuGc) ganglioside, as well as with the P3 anti-idiotypicAbs, was evaluated. As shown in Fig. 7A, the hybrid anti-ody having the P3 mAb VH and the 1E10 mAb VL reacted
dies. Reactivity was determined by using serial dilutions of culture supernatants3(NeuGc) ganglioside by chimeric VHP3/V�1E10 (triangles) and VHP3/V�T1�L of methanol. (B–D) Reactivity of chimeric VHP3/V�1E10 and VHP3/V�T1ted with 10 �g/mL of the murine mAbs. Results in (B and D) are representative3 antibody (ch P3 Ab) (diamonds) was used as control.
R98 → S R100a → S R98;100a → S
− − −+++ +++ ++++ +++ ++++++ +++ ++++++ +++ ++++++ + ±+++/+++ −/− −/−
A. Lopez-Requena et al. / Molecular Immunology 44 (2007) 1015–1028 1023
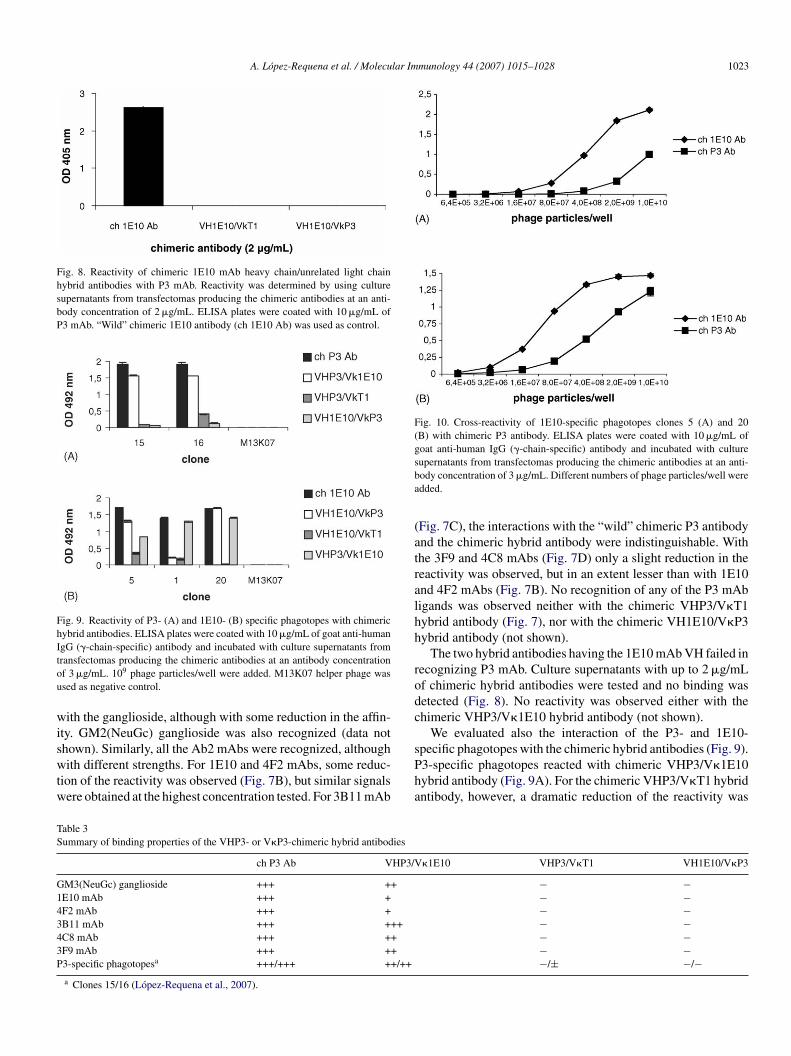
Fig. 8. Reactivity of chimeric 1E10 mAb heavy chain/unrelated light chainhybrid antibodies with P3 mAb. Reactivity was determined by using culturesupernatants from transfectomas producing the chimeric antibodies at an anti-body concentration of 2 �g/mL. ELISA plates were coated with 10 �g/mL ofP3 mAb. “Wild” chimeric 1E10 antibody (ch 1E10 Ab) was used as control.
Fig. 9. Reactivity of P3- (A) and 1E10- (B) specific phagotopes with chimerichybrid antibodies. ELISA plates were coated with 10 �g/mL of goat anti-humanIgG (�-chain-specific) antibody and incubated with culture supernatants fromtou
wiswtw
Fig. 10. Cross-reactivity of 1E10-specific phagotopes clones 5 (A) and 20(B) with chimeric P3 antibody. ELISA plates were coated with 10 �g/mL ofgoat anti-human IgG (�-chain-specific) antibody and incubated with culturesba
(atralhh
rodc
TS
G14343P
ransfectomas producing the chimeric antibodies at an antibody concentrationf 3 �g/mL. 109 phage particles/well were added. M13K07 helper phage wassed as negative control.
ith the ganglioside, although with some reduction in the affin-ty. GM2(NeuGc) ganglioside was also recognized (data not
hown). Similarly, all the Ab2 mAbs were recognized, althoughith different strengths. For 1E10 and 4F2 mAbs, some reduc-ion of the reactivity was observed (Fig. 7B), but similar signalsere obtained at the highest concentration tested. For 3B11 mAb
sPha
able 3ummary of binding properties of the VHP3- or V�P3-chimeric hybrid antibodies
ch P3 Ab VHP3/V
M3(NeuGc) ganglioside +++ ++E10 mAb +++ +F2 mAb +++ +B11 mAb +++ +++C8 mAb +++ ++F9 mAb +++ ++3-specific phagotopesa +++/+++ ++/++
a Clones 15/16 (Lopez-Requena et al., 2007).
upernatants from transfectomas producing the chimeric antibodies at an anti-ody concentration of 3 �g/mL. Different numbers of phage particles/well weredded.
Fig. 7C), the interactions with the “wild” chimeric P3 antibodynd the chimeric hybrid antibody were indistinguishable. Withhe 3F9 and 4C8 mAbs (Fig. 7D) only a slight reduction in theeactivity was observed, but in an extent lesser than with 1E10nd 4F2 mAbs (Fig. 7B). No recognition of any of the P3 mAbigands was observed neither with the chimeric VHP3/V�T1ybrid antibody (Fig. 7), nor with the chimeric VH1E10/V�P3ybrid antibody (not shown).
The two hybrid antibodies having the 1E10 mAb VH failed inecognizing P3 mAb. Culture supernatants with up to 2 �g/mLf chimeric hybrid antibodies were tested and no binding wasetected (Fig. 8). No reactivity was observed either with thehimeric VHP3/V�1E10 hybrid antibody (not shown).
We evaluated also the interaction of the P3- and 1E10-
pecific phagotopes with the chimeric hybrid antibodies (Fig. 9).3-specific phagotopes reacted with chimeric VHP3/V�1E10ybrid antibody (Fig. 9A). For the chimeric VHP3/V�T1 hybridntibody, however, a dramatic reduction of the reactivity was�1E10 VHP3/V�T1 VH1E10/V�P3
− −− −− −− −− −− −−/± −/−

1024 A. Lopez-Requena et al. / Molecular Immunology 44 (2007) 1015–1028
Table 4Summary of binding properties of the VH1E10- or V�1E10-chimeric hybrid antibodies
ch 1E10 Ab VH1E10/V�P3 VH1E10/V�T1 VHP3/V�1E10
P3 mAb +++ − − −1E10-specific phagotopesa +++/+++/+++ ++/−/+++ ±/−/− +b/+++/+b
a
ofa
VIm
Vtrtp14fa
c
4
amhaicmCetpt
FDb
Clones 5/1/20 (Lopez-Requena et al., 2007).b Cross-reactivity with P3 mAb heavy chain variable region (see Fig. 10).
bserved for clone 16, while no reactivity at all was detectedor clone 15. No recognition of chimeric VH1E10/V�P3 hybridntibody was observed.
1E10-specific clones 5 and 20 recognized chimericH1E10/V�P3 hybrid antibody, while clone 1 did not (Fig. 9B).
nteraction with chimeric VH1E10/V�T1 hybrid antibody wasarginal for clone 5, and practically absent for clones 1 and 20.The three 1E10-specific phagotopes reacted with chimeric
HP3/V�1E10 hybrid antibody (Fig. 9B). The recognition ofhis hybrid antibody by clones 5 and 20 was due to a cross-eaction with the P3 mAb heavy chain at high phage concentra-ions. As shown in Fig. 10, approximately 25-fold more phagearticles were needed for chimeric P3 antibody than for chimericE10 antibody, in order to obtain similar signal values. With× 108 phage particles/well for clone 5 (Fig. 10A) and 8 × 107
or clone 20 (Fig. 10B), only reactivity against chimeric 1E10ntibody could be detected.
The results from the binding assays for VHP3- and VH1E10-himeric hybrid antibodies are summarized in Tables 3 and 4.
cbwc
ig. 11. Amino acid sequences of the variable regions of (A) heavy chains from P3 anashed lines indicate nucleotide sequence identities. The CDRs are underlined and seen introduced to align CDRs with different number of amino acid residues.
. Discussion
In the present work we sequenced three more anti-idiotypicntibodies of Ab1 P3 mAb: 3F9, 4C8, and 4F2. 3F9 and 4C8Ab VHs differ only in their H-CDR3. This CDR in 4C8 mAb
as two amino acids more than in 3F9 mAb, one of which isn aspartate residue. Thus, 3F9 mAb has four aspartate residuesn the H-CDR3, while 4C8 mAb has five. The two amino acidhanges observed in VH3F9 and VH4C8 with respect to theirost similar germ-line sequence occur in FR1 (D1 → Q) andDR2 (Q50 → Y). Given the fact that these changes occur in sev-ral of the most similar sequences with VH3F9 and VH4C8, andhat the replacement to silent (R/S) ratio (0.5) is below the valueroposed to indicate a positive selection of nucleotide changeshat affect amino acid sequence (2.9) (Shlomchik et al., 1987), it
an be affirmed that the VH3F9 and VH4C8 regions are codifiedy germ-line genes, as VH1E10 (Perez et al., 2001). In contrastith these two antibodies, VH3B11 differs with respect to itslosest germ-line sequence in a silent mutation in FR2, three
ti-idiotypic mAbs and (B) light chains from P3 anti-idiotypic, P3 and T1 mAbs.eparated from the frameworks by spaces. Spaces within CDR sequences have

lar Immunology 44 (2007) 1015–1028 1025
crs
spismoDbtApebtimTaA3ppa
s(HNt
ttVcc
fabarioVf
itwb4Pi
Fig. 12. (A) Molecular surface calculated on the model of the Fv fragment of P3mAb, displaying in different colours those V� CDR residues that differ betweenP3 and 1E10 mAbs: yellow for L-CDR1, blue for L-CDR2 and magenta forL-CDR3. V� framework residues, as well as those L-CDR amino acids thatare identical in both antibodies, are shown in light blue. VH is displayed inred, with the H-CDR1 R31 residue shown in white. (B) Similar figure for 1E10mAb, showing the differences with 4F2 mAb in the V� CDRs. The same coloursi
cits
Rtrdnr
A. Lopez-Requena et al. / Molecu
onservative changes in FRs 1 and 3, and four non-conservativeeplacements in CDR2. These changes indicate a process ofomatic hypermutation (Perez et al., 2001).
Comparison of VH sequences of all anti-idiotypic mAbs,howed some interesting features. The heavy chain residue atosition 54 (D), which is exposed at the tip of the H-CDR2,s conserved in all antibodies. This is a relatively frequentpecificity-determining residue for protein antigens, as deter-ined by the analysis of several three-dimensional structures
f antigen–antibody complexes (Almagro, 2004). The use ofSP2.2 minigene in reading frame 3, as previously discussedy Perez et al. (2001), allows the presence of at least one aspar-ate residue in the H-CDR3 of all Ab1 P3 anti-idiotypic mAbs.s DSP2.2 codes for two aspartic acids in reading frame 3, therobability increases to have at least a complete codon afterxonuclease cut and random nucleotide addition during assem-ly of the VH region. The usage of the same D minigene byhese different mAbs is then relevant, considering the possiblemportance of electrostatic forces in the interaction of P3 Ab1
Ab with its anti-idiotypic mAbs (Perez et al., 2001, 2002).he acid amino acid motif found by Perez et al. (2001) for 1E10nd 3B11 mAbs, may now be redefined as follows for the fiveb2 mAbs: E/D95-D96-Y/D97-Y/D98. Additionally, except forB11 mAb, the other four Ab2 have another aspartate residue inosition 101. D101 is, however, a structurally conserved residueresent in most antibodies, located deep at the base of H-CDR3nd with little solvent exposure.
V� regions were all codified by germ-line genes. V�3F9 hasome nucleotide changes with respect to germ-line gene 23–43Schaeble et al., 1999), but only one amino acid replacement.owever, this change was also present in the most similar geneaid 60 (Park et al., unpublished), which prompted us to suggest
hat V�3F9 belongs to a new germ-line gene.The analysis of 4F2 mAb presented a more interesting situa-
ion. The VH regions of 4F2 and 1E10 mAb are identical, whilehe V� regions are different. V�4F2 was found to be identical to�4C8. Then, 4F2 mAb is a kind of a “natural hybrid” antibody,
omposed by the 1E10 mAb heavy chain and the 4C8 mAb lighthain.
The construction of chimeric hybrid antibodies allowed us tourther assess the importance of each antibody chain for the P3nd 1E10 mAb binding properties. Chimeric VHP3/V�1E10,ut no VHP3/V�T1, hybrid antibody recognized GM3(NeuGc)nd GM2(NeuGc) gangliosides, with a slight reduction in theeactivity. This result is a strong indication that the VHP3 regions much more important than the V�P3 region for the recognitionf its ganglioside antigens. As shown in Figs. 11 and 12A, the� residues exposed at the binding site surface are very different
or P3 and 1E10 light chains.In order to find a possible explanation for the loss of activ-
ty of the chimeric VHP3/V�T1 hybrid antibody, we comparedhe three-dimensional model of the Fv fragment of P3 mAbith the models of VHP3/V�1E10 and VHP3/V�T1 hybrid anti-
odies. We found that the smaller threonine residue at position6 in V�T1 (instead of the leucine residue in V�P3) allows3 mAb H-CDR3 W100e to flip into a cavity at the VH–V�nterface (Fig. 13). This side chain displacement affects the
stwm
cheme as in panel A was applied. “For interpretation of the references to colourn this figure legend, the reader is referred to the web version of the article”.
onformation of H-CDR3, which may result in loss of the bind-ng capability. This conformational change does not occur forhe VHP3/V�1E10 hybrid antibody, since V�1E10 displays theame leucine residue as V�P3 at position 46.
The study of the recognition of 1E10 mAb by the P3 mAb→ S mutant antibodies (Lopez-Requena et al., 2007) showed
hat none of the individual replacements of the H-CDR3 arginineesidues affected this interaction. The affinity of the H-CDR3ouble mutant antibody decreased, but it was still well recog-ized. In contrast with this, the substitution of the H-CDR1 R31esidue completely abolished the reactivity. This result, like theimilar one obtained for the ganglioside, points to the impor-
ance of the P3 mAb heavy chain also for Ab2 binding. Heree evaluated the interaction of four additional P3-anti-idiotypicAbs with the four P3 mAb H-CDR R → S mutant antibodies.
1026 A. Lopez-Requena et al. / Molecular Immunology 44 (2007) 1015–1028
Fig. 13. (A) Superimposed models of the Fv fragments of P3 mAb and chimeric VHP3/V�T1 hybrid antibody. As both structures are very similar, only the whole P3mAb model is shown. V� is shown in blue and VH in red. Hybrid antibody H-CDR3 is shown in magenta. (B) Zoom-in into the region of interest. “For interpretationo vers
AsmRCtA(rladrtwtawia
rtdmtbmahiii
iVfiattthtb
ttPtctap2
bact
nm
f the references to colour in this figure legend, the reader is referred to the web
ll the anti-idiotypic mAbs that were now tested showed someimilarities in binding to these mutants with respect to 1E10Ab, which were the absence of interaction with the H-CDR131 → S mutant antibody and the full reactivity with the H-DR3 R98 → S mutant antibody. For 3B11, 4C8 and 4F2 mAbs
he P3 mAb H-CDR3 mutations had no effect at all. These threeb2 mAbs have a greater affinity for P3 mAb than 1E10 mAb
Vazquez et al., 1998; our present results). It is interesting toemark that 4F2 mAb, whose heavy chain is identical but theight chain is different to those of 1E10 mAb, has different inter-ctions with P3 mAb (Vazquez et al., 1998) and the H-CDR3ouble mutant antibody, which indicates that the light chain isesponsible for this behavior. For 3F9 mAb, the substitution ofhe H-CDR3 R100a caused a strong reduction of the reactivity,hich was more dramatic with the double mutant. Thus, unlike
he rest of the Ab2 mAbs, 3F9 mAb seems to be critically inter-cting not only with the P3 mAb H-CDR1 R31 residue, but alsoith the H-CDR3 R100a residue, which are both relevant for the
nteraction of P3 mAb with the gangliosides (Lopez-Requena etl., 2007).
As with the ganglioside, the anti-idiotypic antibodies wereecognized only by the chimeric hybrid antibody composed byhe VHP3 and the V�1E10 regions. The interaction pattern wasifferent to the one obtained with the P3 mAb H-CDR3 R → Sutant antibodies. Firstly, 1E10 and 4F2 mAbs, which share
he same VH region, behaved identically against the hybrid anti-ody, despite their different light chains. Secondly, 3F9 and 4C8Abs, from the same VH family, equally reacted with the hybrid
ntibody, showing a smaller decrease in affinity. Thirdly, the only
ypermutated Ab2 mAb, 3B11 mAb, from the same VH fam-ly as 3F9 and 4C8 mAbs, showed no difference at all in itsnteraction with the hybrid antibody, when compared with thenteraction with the chimeric P3 antibody.12to
ion of the article”.
The interaction of phagotopes with the hybrid antibod-es was also evaluated. P3-specific phagotopes recognizedHP3/V�1E10 and one of them also VHP3/V�T1, which con-rms that the interaction occurs with the heavy chain of thentibody. This result, together with the inhibition of the phago-ope binding to P3 by all the Ab2 mAbs, is another indication ofhe prevalence of the heavy chain in the anti-idiotypic interac-ions. It is interesting to remark that the chimeric VHP3/V�T1ybrid antibody, which recognized neither the ganglioside norhe anti-idiotypic antibodies, reacted only with one phagotope,ut much more weakly than the VHP3/V�1E10 one.
The facts that individual mutations of the arginines at theip of H-CDR3 in the P3 antibody did not impair the bindingo its anti-idiotypic mAbs, and that replacing the light chain of3 by a totally different one did not affect either these interac-
ions, strongly suggest that the Ab2 mAbs recognize the heavyhain part of the binding site, where H-CDR1 R31 plays a cen-ral role (Lopez-Requena et al., 2007). In contrast with haptenntigens, it has been calculated that the residue at heavy chainosition 31 contacts very frequently protein antigens (Almagro,004).
For studying the role of the heavy chain of 1E10 mAb in itsinding properties, we constructed the chimeric VH1E10/V�P3nd VH1E10/V�T1 hybrid antibodies. They both, as well ashimeric VHP3/V�1E10 hybrid antibody, were unable to bindo P3 mAb.
Two out of the three 1E10-specific phagotopes also recog-ized 4F2 mAb, which bears the same heavy chain as 1E10Ab. Interestingly, only the binding of the other phagotope to
E10 mAb was inhibited by P3 mAb (Lopez-Requena et al.,007). These results indicate that even though two of the pep-ides clearly recognize the antibody heavy chain, and in spitef the observed similarity with P3 mAb H-CDR3 for one of

lar Im
tthi1rtmartPn
bcomctFbsaovavpc
mfnmmicscaadottp
A
t
R
A
A
A
A
B
B
C
C
C
D
F
G
H
H
J
K
K
L
L
A. Lopez-Requena et al. / Molecu
hem, these peptides react with zones not critically involved inhe Ab1–Ab2 interaction. As expected, chimeric VH1E10/V�P3ybrid antibody was recognized by the phagotopes that alsonteract with 4F2 mAb. The other peptide recognizes only theE10 mAb light chain variable region, as demonstrated with itseactivity with the chimeric VHP3/V�1E10 hybrid antibody andhe lack of interaction with the other hybrid antibodies and 4F2
Ab. Perez et al. (2002) also selected peptides displayed on fil-mentous phage that bound to 1E10 mAb. As in our previousesults (Lopez-Requena et al., 2007), the peptide selected withhe highest frequency has an RRPK motif (RRPKVFRQV) and3 mAb inhibited its binding to 1E10 mAb. This phagotope didot recognize 4F2 mAb (Perez et al., 2002).
The reactivity results for the VH1E10-chimeric hybrid anti-odies and the 1E10-specific phagotopes, suggest that the lighthain of the Ab2 1E10 mAb is important for the recognitionf the Ab1 P3 mAb. However, the fact that both 1E10 and 4F2Abs, which share the same heavy chain but have different light
hains, are anti-idiotypic mAbs of P3, clearly demonstrates thatheir heavy chains are the main responsible for their specificity.ig. 12B shows that the amino acid residues located at the VLinding site surface are very different in 1E10 and 4F2 mAbs. Aimilar picture was obtained with the chimeric VH1E10/V�P3nd VH1E10/V�T1 hybrid antibodies (not shown). In the casef these two antibodies, the lack of reactivity may be due to unfa-orable conformational changes of the H-CDR3 caused by themino acid side chains of the light chain at the interface. As pre-iously exemplified, even when only the heavy chain residuesarticipate in the interaction with the antigen, not all the VH/VLombinations are productive.
The heavy chains of P3 and 1E10 mAbs appear to play theain role in these antibodies binding properties. Experimental
acts presented here point to the conclusion that the recog-ition of both the ganglioside antigens and the anti-idiotypicAbs by P3 mAb, and the recognition of P3 mAb by 1E10Ab depend mainly on the heavy chain, with a minor partic-
pation of the light chain. However, the contribution of lighthains to the binding affinity of these antibodies was also clearlyhowed. Reports in the literature and our results demonstrate thatase-to-case analysis is necessary to assess the contribution ofntibody chains to their binding properties. Replacements of thecid amino acids in H-CDR3 of 1E10 anti-idiotypic mAb, toetermine their importance in Ab1–Ab2 interaction, are nowngoing. Crystallization of the P3–1E10 complex for struc-ure determination will be soon attempted, which will allow uso check the validity of part of the conclusions raised in thisaper.
cknowledgements
We thank Katya Sosa and Tania Grinan for technical assis-ance. This work was supported by the Cuban Government.
eferences
kolkar, P.N., Sikder, S.K., Bhattacharya, S.B., Liao, J., Gruezo, F., Morrison,S.L., Kabat, E.A., 1987. Different VL and VH germ-line genes are used to
L
munology 44 (2007) 1015–1028 1027
produce similar combining sites with specificity for alpha(1–6)dextrans. J.Immunol. 138, 4472–4479.
lfonso, M., Dıaz, A., Hernandez, A.M., Perez, A., Rodrıguez, E., Bitton, R.,Perez, R., Vazquez, A.M., 2002. An anti-idiotype vaccine elicits a specificresponse to N-glycolyl sialic acid residues of glycoconjugates in melanomapatients. J. Immunol. 168, 2523–2529.
lfonso, M., Vazquez, A.M., Carr, A., Haerslev, T., Fernandez, L.E., Lanio,M.E., Alvarez, C., Zeuthen, J., Perez, R., 1995. T-cell independent B-cellresponse to self monosialogangliosides: primary response monoclonal anti-bodies. Hybridoma 14, 209–216.
lmagro, J.C., 2004. Identification of differences in the specificity-determiningresidues of antibodies that recognize antigens of different size: implicationsfor the rational design of antibody repertoires. J. Mol. Recog. 17, 132–143.
angs, L.A., Sanz, I.E., Teale, J.M., 1991. Comparison of D, JH and junctionaldiversity in the fetal, adult, and aged B cell repertoires. J. Immunol. 146,1996–2004.
hattacharya-Chatterjee, M., Chatterjee, S.K., Foon, K.A., 2000. Anti-idiotypevaccine against cancer. Immunol. Lett. 74, 51–58.
alarese, D.A., Scanlan, C.N., Zwick, M.B., Deechongkit, S., Mimura, Y.,Kunert, R., Zhu, P., Wormald, M.R., Stanfield, R.L., Roux, K.H., Kelly,J.W., Rudd, P.M., Dwek, R.A., Katinger, H., Burton, D.R., Wilson, I.A.,2003. Antibody domain exchange is an immunological solution to carbohy-drate cluster recognition. Science 300, 2065–2071.
arr, A., Mullet, A., Mazorra, Z., Vazquez, A.M., Alfonso, M., Mesa, C.,Rengifo, E., Perez, R., Fernandez, L.E., 2000. A mouse IgG1 monoclonalantibody specific for N-glycolyl GM3 ganglioside recognized breast andmelanoma tumors. Hybridoma 19, 241–247.
hapman, P.B., 2003. Vaccinating against GD3 ganglioside using BEC2 anti-idiotypic monoclonal antibody. Curr. Opin. Investig. Drugs 4, 710–715.
ıaz, A., Alfonso, M., Alonso, R., Saurez, G., Troche, M., Catala, M., Dıaz,R.M., Perez, R., Vazquez, A.M., 2003. Immune responses in breast can-cer patients immunized with an anti-idiotype antibody mimicking NeuGc-containing gangliosides. Clin. Immunol. 107, 80–89.
oon, K.A., Lutzky, J., Baral, R.N., Yannelli, J.R., Hutchins, L., Teitelbaum, A.,Kashala, O.L., Das, R., Garrison, J., Reisfeld, R.A., Bhattacharya-Chatterjee,M., 2000. Clinical and immune responses in advanced melanoma patientsimmunized with an anti-idiotype antibody mimicking disialogangliosideGD2. J. Clin. Oncol. 18, 376–384.
uex, N., Peitsch, M.C., 1997. SWISS-MODEL and the Swiss-PdbViewer:an environment for comparative protein modeling. Electrophoresis 18,2714–2723, http://www.expasy.org/spdbv/.
amers-Casterman, C., Atarhouch, T., Muyldermans, S., Robinson, G., Hamers,C., Songa, E.B., Bendahman, N., Hamers, R., 1993. Naturally occurringantibodies devoid of light chains. Nature 363, 446–448.
ashimoto, Y., Otsuka, H., Sudo, K., Suzuki, K., Yamakawa, T., 1983. Geneticregulation of GM2 expression in liver mouse. J. Biochem. 93, 895–901.
in, H., Sepulveda, J., Burrone, O.R., 2003. Selection and characterisation ofbinders based on homodimerisation of immunoglobulin V(H) domains.FEBS Lett. 554, 323–329.
abat, E.A., Wu, T.T., Peery, H.M., Gottesmann, K.S., Foeller, C., 1991.Sequences of Proteins of Immunological Interest, fifth ed. U.S. Departmentof Health and Human Services, U.S. Government Printing Office, Washing-ton.
rengel, U., Olsson, L.-L., Martınez, C., Talavera, A., Rojas, G., Mier, E.,Angstrom, J., Moreno, E., 2003. Structure and molecular interactions ofa unique anti-tumor antibody specific for N-glycolyl GM3. J. Biol. Chem.279, 5597–5603.
eahy, D.J., Rule, G.S., Whittaker, M.M., McConnell, H.M., 1988. Sequencesof 12 monoclonal anti-dinitrophenyl spin-label antibodies for NMR studies.Proc. Natl. Acad. Sci. U.S.A. 85, 3661–3665.
opez-Requena, A., De Acosta, C.M., Moreno, E., Gonzalez, M., Puchades,Y., Talavera, A., Vispo, N.S., Vazquez, A.M., Perez, R., 2007. Ganglio-sides, Ab1 and Ab2 antibodies. I. Towards a molecular dissection of an
idiotype–anti-idiotype system. Mol. Immunol. 44, 423–433.opez-Requena, A., Mateo de Acosta, C., Perez, A., Valle, A., Lombardero,J., Sosa, K., Perez, R., Vazquez, A.M., 2003. Chimeric anti-N-glycolyl-ganglioside and its anti-idiotypic MAbs: immunodominance of their variableregions. Hybrid. Hybridomics 22, 235–243.

1 lar Im
M
M
N
P
P
R
R
R
R
S
S
S
S
S
ST
V
V
028 A. Lopez-Requena et al. / Molecu
alykh, Y.N., Schauer, R., Shaw, L., 2001. N-Glycolylneuraminic acid in humantumours. Biochimie 83, 623–634.
orea, V., Tramontano, A., Rustici, M., Chothia, C., Lesk, A.M., 1998. Confor-mations of the third hypervariable region in the VH domain of immunoglob-ulins. J. Mol. Biol. 275, 269–294.
oel, D., Bernardi, T., Navarro-Teulon, I., Marin, M., Martinetto, J.P., Ducan-cel, F., Mani, J.C., Pau, B., Piechaczyk, M., Biard-Piechaczyk, M., 1996.Analysis of the individual contributions of immunoglobulin heavy and lightchains to the binding of antigen using cell transfection and plasmon reso-nance analysis. J. Immunol. Methods 193, 177–187.
erez, A., Lombardero, J., Mateo, C., Mustelier, G., Alfonso, M., Vazquez,A.M., Perez, R., 2001. Immunogenetic analysis of variable regions encodingAb1 and �-type Ab2 antibodies from the NeuGc-containing gangliosidefamily. Hybridoma 20, 211–221.
erez, A., Mier, E.S., Santiago, N., Perez, R., 2002. A monoclonal anti-body against NeuGc-containing gangliosides contains a regulatory idiotopeinvolved in the interaction with B and T cells. Mol. Immunol. 39, 103–112.
eiter, Y., Schuck, P., Boyd, L.F., Plaksin, D., 1999. An antibody single-domainphage display library of a native heavy chain variable region: isolation offunctional single-domain VH molecules with a unique interface. J. Mol.Biol. 290, 685–698.
odrıguez, M., Llanes, L., Perez, A., Perez, R., Vazquez, A.M., 2003. Genera-tion and characterization of an anti-idiotype monoclonal antibody related toGM3(NeuGc) ganglioside. Hybrid. Hybridomics 22, 307–314.
ojas, G., Talavera, A., Munoz, Y., Rengifo, E., Krengel, U., Angstrom, J., Gav-ilondo, J., Moreno, E., 2004. Light chain shuffling results in successful phagedisplay of antibody fragments to N-glycolyl GM3 ganglioside. J. Immunol.
Methods 293, 71–83.oque-Navarro, L., Mateo, C., Lombardero, J., Mustelier, G., Fernandez, A.,Sosa, K., Morrison, S.L., Perez, R., 2003. Humanization of predicted T-cellepitopes reduces the immunogenicity of chimeric antibodies: new evidencesupporting a simple method. Hybrid. Hybridomics 22, 245–257.
W
X
munology 44 (2007) 1015–1028
anger, F., Nicklen, S., Coulson, A.R., 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74, 5463–5467.
chaeble, K.F., Thiebe, R., Bensch, A., Brensing-Kueppers, J., Heim, V.,Kirschbaum, T., Lamm, R., Ohnrich, M., Pourrajabi, S., Roeschenthaler,F., Schwendinger, J., Wichelhaus, D., Zocher, I., Zachau, H.G., 1999. Char-acteristics of the immunoglobulin Vkappa genes, pseudogenes, relics andorphons in the mouse genome. Eur. J. Immunol. 29, 2082–2086.
epulveda, J., Jin, H., Sblattero, D., Bradbury, A., Burrone, O.R., 2003. Bindersbased on dimerised immunoglobulin VH domains. J. Mol. Biol. 333,355–365.
hlomchik, M.J., Marshak-Rothstein, A., Wolfowicz, C.B., Rothstein, T.L.,Weigert, M.G., 1987. The role of clonal selection and somatic mutationin autoimmunity. Nature 328, 805–811.
tults, C.L.M., Sweeley, C.C., Matcher, B.A., 1989. Glycosphingolipids: struc-ture, biological source, and properties. Methods Enzymol. 179, 167–214.
vennerholm, L., 1964. The gangliosides. J. Lipid Res. 5, 145–155.anha, J., Xu, P., Chen, Z., Ni, F., Kaplan, H., Narang, S.A., MacKenzie, C.R.,
2001. Optimal design features of camelized human single-domain antibodylibraries. J. Biol. Chem. 276, 24774–24780.
azquez, A.M., Alfonso, M., Lanne, B., Karlsson, K.-A., Carr, A., Barroso,O., Fernandez, L.E., Rengifo, E., Lanio, M.E., Alvarez, C., Zeuthen, J.,Perez, R., 1995. Generation of a murine monoclonal antibody specific forN-glycolylneuraminic acid-containing gangliosides that also recognizes sul-fated glycolipids. Hybridoma 14, 551–556.
azquez, A.M., Perez, A., Hernandez, A.M., Macıas, A., Alfonso, M., Bombino,G., Perez, R., 1998. Syngeneic anti-idiotypic monoclonal antibodies to ananti-NeuGc-containing ganglioside monoclonal antibody. Hybridoma 17,527–534.
hitelegg, N.R.J., Rees, A.R., 2000. WAM—an improved algorithmfor modelling antibodies on the web. Prot. Eng. 13, 819–824,http://antibody.bath.ac.uk/.
u, J.L., Davis, M.M., 2000. Diversity in the CDR3 region of V(H) is sufficientfor most antibody specificities. Immunity 13, 37–45.
Related Documents











