Cell Tissue Rcs (1993) 271:537-544 Cell&Tissue Research Springer-Verlag 1993 Galanin-like immunoreactivity in extrinsic and intrinsic nerves to the gut of the Atlantic cod, Gadus morhua, and the effect of galanin on the smooth muscle of the gut Paul Karila, Ann-Cathrine J6nsson, J6rgen Jensen, Susanne Holmgren Comparative Neuroseience Unit, Department of Zoophysiology, University of G6teborg, Medieinaregatan 18, S-41390 G6teborg, Sweden Received: 11 June 1992 / Accepted: 4 October 1992 Abstract. The presence of galanin-like immunoreactivity in nerves to the stomach of the Atlantic cod has been investigated, by immunohistochemistry. The distribution of ganglion cells showing galanin-like immunoreactivity was compared with the total distribution in nerves and ganglia. Projection studies were made to determine the origin of the galanin neurons. The effect of galanin was studied in smooth muscle strip preparations of the gut wall and arteries. Galanin-like immunoreactive ganglion cells frequently occurred along the vagal branches to the stomach. Most of them projected cranially. Immuno- reactive nerve fibres were present in all layers of the gut and around arterial branches on the surface of the stomach. Ligations of the vagus and splanchnic nerves produced accumulations of immunoreactive material on both sides of the ligature. Galanin produced weak con- tractile effects unaffected by tetrodotoxin on the gut wall and on gut arteries. It is concluded that a population of the ganglion cells along the vagus nerve in the Atlantic cod contains a galanin-like peptide. Some of these cells may be parts of autonomic parasympathetic pathways innervating the gut of the Atlantic cod, having direct excitatory effects on the smooth muscles of the gut wall and gut arteries. Key words: Galanin - Gut - Arteries - Vagus nerve - Sympathetic ganglia - Gachts morhtta (Teleostei) Introduction Galanin is a 29 amino-acid-residue peptide originally isolated from porcine intestine (Tatemoto et al. 1983). The peptide has since been demonstrated in a number of species in a variety of central and peripheral nervous tissues, including a dense population of nerves in the gastrointestinal canal of many mammals (Ekblad et al. 1985; Bishop et al. 1986).In addition, galanin has been Correspondence to: P. Karila identified in sensory neurons, often co-localised with substance P and calcitonin gene-related peptide (Ju et al. 1987). A dense nerve net and high concentrations of galanin are also reported in amphibian and reptilian ganglia, gut, central nervous system and heart (Gibbins et al. 1989; Morris et al. 1989; McKeon et al. 1990). In sympathetic neurons, galanin often coexists with neu- ropeptide Y (Gibbins etal. 1989; Morris et al. 1989, 1992). The physiological effects of galanin in mammals are diverse. A wide ranging, but not unspecific, inhibition of postprandial release of hormones and neurotransmit- ters has been described; galanin also has a pronounced effect on acid secretion and affects gut motility (Dunning et al. 1986; Lindskog and Ahr6n 1987; Bauer et al. 1989; Schepp et al. 1990; u et al. 1990; Rattan 1991). The effect of galanin of the contractile activity of the gut is species-dependent, and may differ between circular and longitudinal muscle, and between different parts of the gut (Ekblad et al. 1985a; Fox et al. 1986, 1988). Ex- periments on the cardiac ganglion of the amphibian Nec- tttrtts macttlostts have shown that galanin may be both depolarising and hyperpolarising on postsynaptic gangli- on cells (Konopka and Parsons 1989; Parsons and Kon- opka 1990). The present study was initiated to investigate the presence of intrinsic and extrinsic pathways of galanin- containing neurons innervating the gut of the Atlantic cod and the possible effects of these neurons on the motility of the gut wall and arteries. Materials and methods Atlantic cod, Gadus mor]ma, of both sexes and with a body weight of 300-800 g were used. The fish were captured off the Swedish west coast by local fishermen, and transferred to the laboratory where they were kept in well aerated circulating sea water at 8- 10 ~ C. Prior to use, the fish were killed by a sharp blow to the head.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cell Tissue Rcs (1993) 271:537-544 Cell&Tissue
Research �9 Springer-Verlag 1993
Galanin-like immunoreactivity in extrinsic and intrinsic nerves to the gut of the Atlantic cod, Gadus morhua, and the effect of galanin on the smooth muscle of the gut Paul Karila, Ann-Cathrine J6nsson, J6rgen Jensen, Susanne Holmgren
Comparative Neuroseience Unit, Department of Zoophysiology, University of G6teborg, Medieinaregatan 18, S-41390 G6teborg, Sweden
Received: 11 June 1992 / Accepted: 4 October 1992
Abstract. The presence of galanin-like immunoreactivity in nerves to the stomach of the Atlantic cod has been investigated, by immunohistochemistry. The distribution of ganglion cells showing galanin-like immunoreactivity was compared with the total distribution in nerves and ganglia. Projection studies were made to determine the origin of the galanin neurons. The effect of galanin was studied in smooth muscle strip preparations of the gut wall and arteries. Galanin-like immunoreactive ganglion cells frequently occurred along the vagal branches to the stomach. Most of them projected cranially. Immuno- reactive nerve fibres were present in all layers of the gut and around arterial branches on the surface of the stomach. Ligations of the vagus and splanchnic nerves produced accumulations of immunoreactive material on both sides of the ligature. Galanin produced weak con- tractile effects unaffected by tetrodotoxin on the gut wall and on gut arteries. It is concluded that a population of the ganglion cells along the vagus nerve in the Atlantic cod contains a galanin-like peptide. Some of these cells may be parts of autonomic parasympathetic pathways innervating the gut of the Atlantic cod, having direct excitatory effects on the smooth muscles of the gut wall and gut arteries.
Key words: Galanin - Gut - Arteries - Vagus nerve - Sympathetic ganglia - Gachts morhtta (Teleostei)
Introduction
Galanin is a 29 amino-acid-residue peptide originally isolated from porcine intestine (Tatemoto et al. 1983). The peptide has since been demonstrated in a number of species in a variety of central and peripheral nervous tissues, including a dense population of nerves in the gastrointestinal canal of many mammals (Ekblad et al. 1985; Bishop et al. 1986).In addition, galanin has been
Correspondence to: P. Karila
identified in sensory neurons, often co-localised with substance P and calcitonin gene-related peptide (Ju et al. 1987). A dense nerve net and high concentrations of galanin are also reported in amphibian and reptilian ganglia, gut, central nervous system and heart (Gibbins et al. 1989; Morris et al. 1989; McKeon et al. 1990). In sympathetic neurons, galanin often coexists with neu- ropeptide Y (Gibbins etal . 1989; Morris et al. 1989, 1992).
The physiological effects of galanin in mammals are diverse. A wide ranging, but not unspecific, inhibition of postprandial release of hormones and neurotransmit- ters has been described; galanin also has a pronounced effect on acid secretion and affects gut motility (Dunning et al. 1986; Lindskog and Ahr6n 1987; Bauer et al. 1989; Schepp et al. 1990; u et al. 1990; Rattan 1991). The effect of galanin of the contractile activity of the gut is species-dependent, and may differ between circular and longitudinal muscle, and between different parts of the gut (Ekblad et al. 1985a; Fox et al. 1986, 1988). Ex- periments on the cardiac ganglion of the amphibian Nec- tttrtts macttlostts have shown that galanin may be both depolarising and hyperpolarising on postsynaptic gangli- on cells (Konopka and Parsons 1989; Parsons and Kon- opka 1990).
The present study was initiated to investigate the presence of intrinsic and extrinsic pathways of galanin- containing neurons innervating the gut of the Atlantic cod and the possible effects of these neurons on the motility of the gut wall and arteries.
Materials and methods
Atlantic cod, Gadus mor]ma, of both sexes and with a body weight of 300-800 g were used. The fish were captured off the Swedish west coast by local fishermen, and transferred to the laboratory where they were kept in well aerated circulating sea water at 8- 10 ~ C. Prior to use, the fish were killed by a sharp blow to the head.

538
Ligations/denervations
The fish were anaesthetized in MS222 (2,2,2-tricaine methane sul- phonate, 100 mg/l), and kept anaesthetized during the operation by a continuous flow of sea water containing MS222 (50 mg/l) over the gills. A 30-mm incision was made ventrally, behind the left pectoral fin, and the vagal and splanchnic nerve branches to the gut were identified. In one group of animals, the splanchnic nerve was ligated at a level immediately distal to the gallbladder 01 =7). In another group, the left vagus nerve was ligated proximal to the gallbladder (n=9). The incision was sutured, both groups were left for 3-5 days, killed by a blow to the head, and pieces of the nerves including 5 mm above and below the ligature were taken for immunohistochemistry.
For the denervations, the fish were anaesthetized as above, and the splanchnic nerves were located and cut immediately distal to the gallbladder. For the vagal denervation, a small incision was made behind the 4th gill arch on one or both sides and the vagal branches to the gut were cut. Some 5-7 days after the denervation, the fish were killed by a blow to the head, and the stomach and proximal intestine were dissected out for immunohistochcmistry.
Sham operations were performed following the procedure de- scribed above, but without sectioning of the nerves (splanchnic nerve n=4 , vagus nerve n=4) .
hmmmohistochemistry
For studies of the distribution of ganglion cells immunoreactive for neuron-specific enolase (NSE) [which approximately equals the total amount of ganglion cells (Marangos et al. 1981)] and galanin, the mesentery of the stomach including the splanchnic and vagus branches to the stomach, the coeliac ganglion and the nodose gan- glion were dissected out, pinned fiat to dental wax and fixed in a mixture of 15% picric acid and 2% formalin in 0.1 M phosphate buffer (Zamboni's fixative, pH=7.2) for 20 h. To improve immu- nostaining in thick preparations and to avoid non-specific back- ground fluorescence, the tissues were dehydrated, treated with xy- lene for 30rain and rehydrated (Costa ct al. 1980). The nerves were cut into l-cm pieces that were teased into thin threads before incubation with primary antiserum (60-74 h at 4 ~ C). Ganglia were snap-frozen in liquid nitrogen and sectioned on a cryostat.
From ligated or denervated fish, l-cm samples (in ligated fish including the ligature) from the vagus and splanchnic nerves, and the complete coeliac and nodose ganglia were dissected out, pinned carefully to dental wax to ensure a fiat preparation and fixed. The samples were fixed in either 0.4% parabenzoquinone (PBQ) for 4 h and transferred to sucrose 20% or in Zamboni's fixative for 20 h and dehydrated, treated with xylene for 30 min and rchyd- rated prior to treatment with 20% sucrose at least overnight. The preparations were quick-frozen in liquid N2, or in isopentane cooled by liquid N2. Sections (10 tam thick) were cut on a cryostat and thaw-mounted onto poly-L-lysine or chrome-alum-coated slides.
Small pieces from several locations along the gut were taken from control (sham-operated) and denervated fish, stretched slight- ly on dental wax and fixed floating in Zamboni's fixative for 20 h. The tissues were then dehydrated, xylene-treated and rehydrated. Half of the tissue pieces were peeled and the myenteric plexus was used for whole-mounts according to Costa et al. (1980), where- as the other half was left in 20% sucrose overnight to be used for sectioning.
Sections and whole-mounts were incubated with primary antise- rum for 16-20 h in a moist chamber, repeatedly rinsed for 30 rain in high salinity phosphate-buffered saline (PBS 0.1 M, NaCI 0.5 M, pH 7.2), incubated for l h with secondary antibody in a humid chamber.and repeatedly rinsed for 30 min in PBS. The staining samples were mounted in carbonate buffer:glycerol (1:1, pH 8.4) and viewed in a Leitz Dialux fluorescence microscope.
The distribution of ganglion cells in extrinsic nerves to the gut was derived by counting 5800 NSE-immunoreactive cells from 8
fish. The diameter of 536 cells in randomly chosen sections from 5 fish was measured using a microscope equipped with a microme- ter ocular to evaluate the size distribution of NSE-immunorcactive cells. The distribution and proportion of galanin-like-immunoreac- tive (GLI) cells was determined from a total of 1390 cells from another 8 fish. To compare the size with the NSE-immunoreactive cells, 50 GLI cells from 8 fish were measured in a similar way as the NSE-immunoreactive cells.
Strip preparations
The stomach and intestine were dissected out, opened longitudinal- ly and pinned to a wax plate. The mucosa was carefully removed from the stomach. Strips, 2 • 12 min, were cut from the cardiac stomach and the intestine, in both cases along the circular and the longitudinal axis of the gut. Segments (12-14 mm) of the coeliac and mesenteric arteries were cut out and opened longitudinally.
The strips were suspended in 5-ml or 10-ml organ baths in Atlantic cod Ringer's solution at 10 ~ C containing (in mmol !-~): NaCI, 150.1; KCI, 5.2; MgSO4, 1.8; CaCI2, 1.9; NaHzPO4, 1.9; Nat tCO3, 7.0; glucose, 5.6 [modified by R. Fritsche from Sundcll and Bj6rnsson (1988)]. The Ringer's solution was bubbled contin- uously with a mixture of O2/CO2 (99.7/0.3%) to maintain the pH at 7.8. The upper end of the strip was attached to a Grass FT03 transducer for the recording of tension on a Grass Polygraph Model 7 or 79. The gut wall strips were mounted with an initial tension of 10 milliNewton (mN) and vessels were mounted at the same length as measured in situ (12-14 ram). The preparations were left for 1 h before the experiment. During this time, the ten- sion of the gut strips was reduced to slightly above zero, and spon- taneous rhythmic activity often developed. Vessel preparations adopted a stable baseline at a tension ranging between l0 and 40 mN.
Antisera and drugs
Three primary galanin antisera were used for immunohistochemis- try: B49-1 (1:100-200, Milab, Sweden), RAS-7153 (1:100-200, Peninsula, UK) and CA250 [1:500-1000, Cambridge Research Bio- chemicals (CRB), UK]. Specificity of the reaction was tested by preabsorbtion for 3 days of CA250 (1:500) with galanin. The im- munoreaction was quenched in both ganglion cells and nerve fibres when galanin was added every 24 h (l nmol/ml). The secondary antiserum was in all cases swine-anti-rabbit IgG conjugated with fluorescein isothiocyanate (1:10; Dakopatts, Sweden). An antise- rum raised against NSE (A589, 1:200; Dakopatts) was used to assess the total population of ganglion cells. Synthetic galanin (CRB) was dissolved according to the suppliers recommendations, and was stored frozen in stock solutions o f l 0 -4 or 10 -3 M. Work- ing dilutions were made with cod Ringer's solution containing se- rum albumin (10 mg/ml). L-adrenaline bitartrate,.carbachol and tetrodotoxin (all from Sigma, USA) were dissolved in distilled water to give stock solutions of 1 to 10 mM and were further diluted in Ringer's solution to the final concentrations given below.
Calculations
Values are g iven_SE, unless otherwise stated. Wilcoxons Signed- ranks test was used for the statistical evaluation of data.
Resu l t s
Location of nerve cell bodies
G a n g l i o n cel ls ( s t a ined u s ing the N S E a n t i s e r u m ) w e r e s c a t t e r e d s ing ly o r in sma l l c lus te r s a l o n g the en t i r e
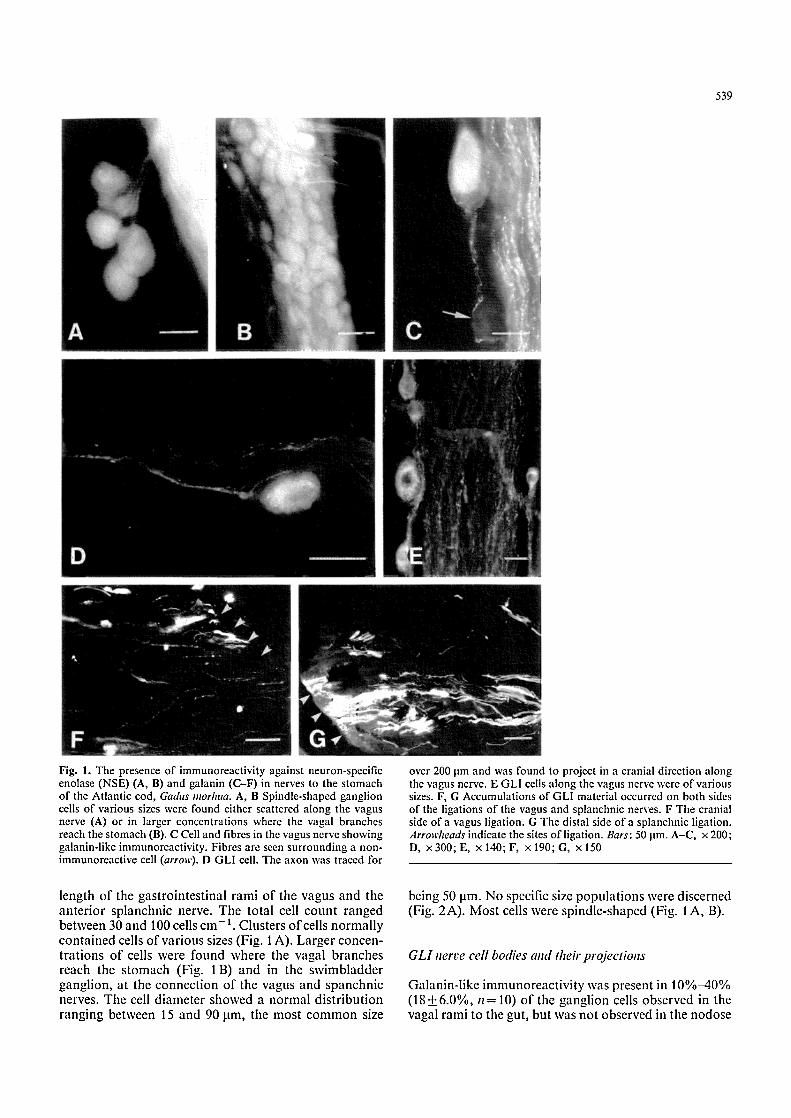
539
Fig. 1. The presence of immunoreactivity against neuron-specific enolase (NSE) (A, B) and galanin (C-F) in nerves to the stomach of the Atlantic cod, Gadus morhua. A, B Spindle-shaped ganglion cells of various sizes were found either scattered along the vagus nerve (A) or in larger concentrations where the vagal branches reach the stomach (B). C Cell and fibres in the vagus nerve showing galanin-like immunoreactivity. Fibres are seen surrounding a non- immunoreactive cell (arrow). D GLI cell. The axon was traced for
over 200 lam and was found to project in a cranial direction along the vagus nerve. E GLI cells along the vagus nerve were of various sizes. F, G Accumulations of GLI material occurred on both sides of the ligations of the vagus and splanchnic nerves. F The cranial side of a vagus ligation. G The distal side of a splanchnic ligation. Arrowheads indicate the sites ofligation. Bars: 50 lam. A-C, x 200; D, x300;E, x 140;F, • 190;G, x150
length o f the gas t ro in tes t ina l r ami o f the vagus and the a n t e r i o r sp lanchnic nerve. The to ta l cell coun t r anged be tween 30 and 100 cells c m - t. Clus ters o f cells n o r m a l l y con t a ined cells o f va r ious sizes (Fig. 1 A). La rge r concen- t r a t ions o f cells were found where the vagal b r anches reach the s t omach (Fig. 1 B) and in the s w i m b l a d d e r gangl ion , a t the connec t ion o f the vagus and spanchn ic nerves. The cell d i ame te r showed a n o r m a l d i s t r i bu t i on r ang ing be tween 15 and 90 lam, the mos t c o m m o n size
being 50 lam. N o specific size p o p u l a t i o n s were d iscerned (Fig. 2A) . M o s t cells were sp ind le - shaped (Fig. 1 A , B).
G L I nerve cell bodies and theh" projections
Galan in - l i ke i m m u n o r e a e t i v i t y was present in 1 0 % - 4 0 % ( 1 8 + 6 . 0 % , n = 10) o f the gang l ion cells obse rved in the vagal rami to the gut, bu t was no t obse rved in the nodose
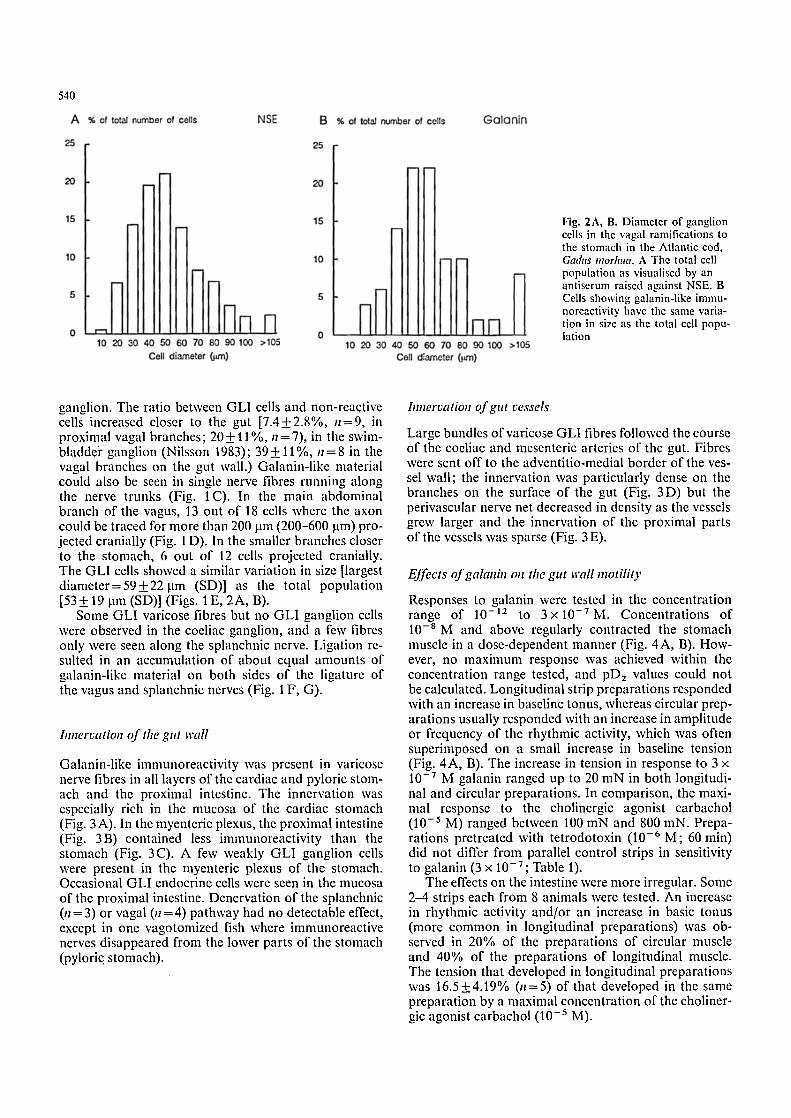
540
Fig. 2A, B. Diameter of ganglion cells in the vagal ramifications to the stomach in the Atlantic cod, Gadus morhua. A The total cell population as visualised by an antiserum raised against NSE. B Cells showing galanin-like immu- noreactivity have the same varia- tion in size as the total cell popu- lation
ganglion. The ratio between GLI cells and non-reactive cells increased closer to the gut [7.4+2.8%, n=9, in proximal vagal branches; 20+ 11%, n=7), in the swim- bladdei" ganglion (Nilsson 1983); 39 + 11%, n = 8 in the vagal branches on the gut wall.) Galanin-like material could also be seen in single nerve fibres running along the nerve trunks (Fig. 1 C). In the main abdominal branch of the vagus, 13 out of 18 cells where the axon could be traced for more than 200 jam (200-600 jam) pro- jected cranially (Fig. 1 D). In the smaller branches closer to the stomach, 6 out of 12 cells projected cranially. The GLI cells showed a similar variation in size [largest diameter= 59 + 22 ~tm (SD)] as the total population [53-1-19 jam (SD)] (Figs. | E, 2A, B).
Some GLI varicose fibres but no GLI ganglion cells were observed in the coeliac ganglion, and a few fibres only were seen along the splanchnic nerve. Ligation re- suited in an accumulation of about equal amounts of galanin-like material on both sides of the ligature of the vagus and splanchnic nerves (Fig. 1 F, G).
hmervation of the gut wall
Galanin-like immunoreactivity was present in varicose nerve fibres in all layers of the cardiac and pyloric stom- ach and the proximal intestine. The innervation was especially rich in the mucosa of the cardiac stomach (Fig. 3A). In the myenteric plexus, the proximal intestine (Fig. 3B) contained less immunoreactivity than the stomach (Fig. 3C). A few weakly GLI ganglion cells were present in the myenteric plexus of the stomach. Occasional GLI endocrine cells were seen in the mucosa of the proximal intestine. Denervation of the splanchnic (11= 3) or vagal (1l =4) pathway had no detectable effect, except in one vagotomized fish where immunoreactive nerves disappeared from the lower parts of the stomach (pyloric stomach).
bmervation of gut vessels
Large bundles of varicose GLI fibres followed the course of the coeliac and mesenteric arteries of the gut. Fibres were sent off to the adventitio-medial border of the ves- sel wall; the innervation was particularly dense on the branches on the surface of the gut (Fig. 3D) but the perivascular nerve net decreased in density as the vessels grew larger and the innervation of the proximal parts of the vessels was sparse (Fig. 3 E).
Effects of galanhl oll the gut wall motili O,
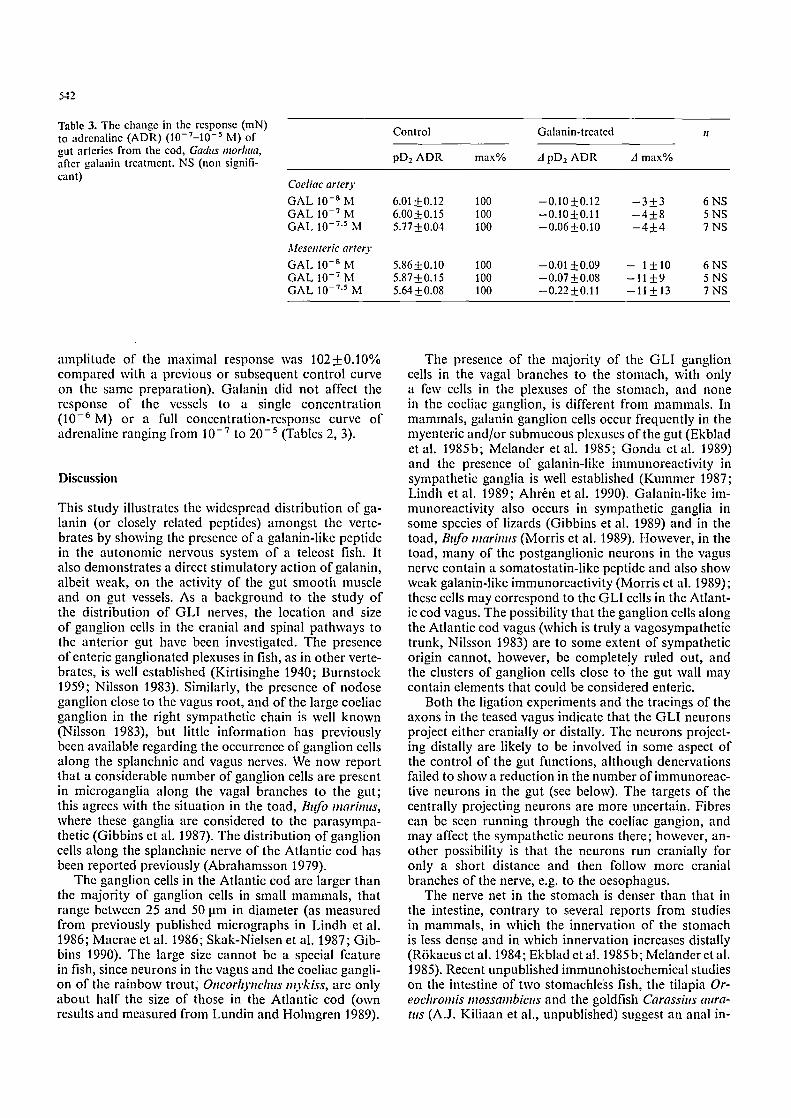
Responses to galanin were tested in the concentration range of 10 -12 to 3 • Concentrations of 10-SM and above regularly contracted the stomach muscle in a dose-dependent manner (Fig. 4A, B). How- ever, no maximum response was achieved within the concentration range tested, and pDz values could not be calculated. Longitudinal strip preparations responded with an increase in baseline tonus, whereas circular prep- arations usually responded with an increase in amplitude or frequency of the rhythmic activity, which was often superimposed on a small increase in baseline tension (Fig. 4A, B). The increase in tension in response to 3 x 10 -7 M galanin ranged up to 20 mN in both longitudi- nal and circular preparations. In comparison, the maxi- mal response to the cholinergic agonist carbachol (10 -5 M) ranged between 100 mN and 800 mN. Prepa- rations pretreated with tetrodotoxin (10 -6 M; 60 min) did not differ from parallel control strips in sensitivity to galanin (3 • 10-v; Table 1).
The effects on the intestine were more irregular. Some 2-4 strips each from 8 animals were tested. An increase in rhythmic activity and/or an increase in basic tonus (more common in longitudinal preparations) was ob- served in 20% of the preparations of circular muscle and 40% of the preparations of longitudinal muscle. The tension that developed in longitudinal preparations was 16.5+4.19% (n=5) of that developed in the same preparation by a maximal concentration of the choliner- gic agonist carbachol (10 -5 M).
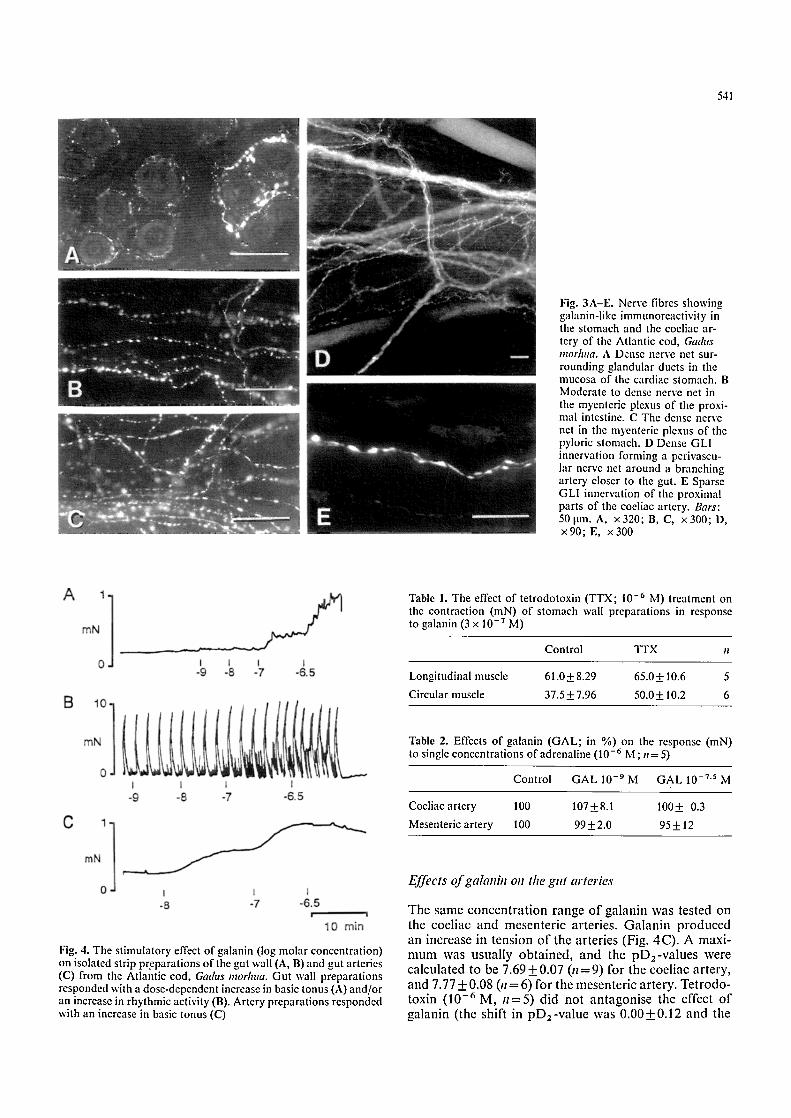
541
Fig. 3A-E. Nerve fibres showing galanin-like immunoreactivity in the stomach and the coeliac ar- tery of the Atlantic cod, Ga&ts mor]ma. A Dense nerve net sur- rounding glandular ducts in the mucosa of the cardiac stomach. B Moderate to dense nerve net in the myenteric plexus of the proxi- mal intestine. C The dense nerve net in the myenteric plexus of the pyloric stomach. D Dense GLI innervation forming a pcrivascu- lar nerve net around a branching artery closer to the gut. E Sparse GLI innervation of the proximal parts of the coeliac artery. Bars: 50 Itm. A, x320; B, C, x300; D, x90; E, x300
Table 1. The effect of tetrodotoxin (TTX; 10 -6 M) treatment on the contraction (mN) of stomach wall prcparations in response to galanin (3 x 10 -7 M)
Control TTX n
Longitudinal muscle 61.0 + 8.29 65.0 + 10.6 5
Circular muscle 37.5___7.96 50.0+__ 10.2 6
Table 2. Effects of galanin (GAL; in %) on the response (mN) to single concentrations of adrenaline (10 -6 M ; n = 5)
Control G A L 10 - 9 M GAL 10 -7.5 M
Coeliac artery 100 1074-8.1 1004- 0.3
Mesenteric artery 100 994-2.0 954-12
Fig. 4. The stimulatory effect of galanin (log molar concentration) on isolated strip preparations of the gut wall (A, B) and gut arteries (C) from the Atlantic cod, Gad, ts morhtta. Gut wall preparations responded with a dose-dependent increase in basic tonus (A) and/or an increase in rhythmic activity (B). Artery preparations responded with an increase in basic tonus (C)
Effects o f galanht oll the gut arteries
The same c o n c e n t r a t i o n range o f ga lan in was tested on the coel iac and mesente r ic ar ter ies . G a l a n i n p r o d u c e d an increase in tens ion o f the ar ter ies (Fig. 4C) . A maxi - m u m was usua l ly ob t a ined , and the p D z - v a l u e s were ca lcu la ted to be 7.69_+0.07 ( n = 9 ) for the coel iac a r te ry , and 7.77_+ 0.08 (n = 6) for the mesenter ic a r te ry . Te t rodo - toxin (10 - 6 M, n = 5 ) d id no t an t agon i se the effect o f ga lan in ( the shift in p D 2 - v a l u e was 0.00_+0.12 and the
542
Table 3. The change in the response (mN) to adrenaline (ADR) (10-7-10 -5 M) of gut arteries from the cod, Gadus morhua, after galanin treatment. NS (non signifi- cant)
Coeliac artery GAL 10 -8 M GAL 10 -7 M GAL 10 -7.5 M
Mesenteric artery GAL 10 -8 M G A L 10 -7 M G A L 10 -7.5 M
Control Galanin-treated n
pD2 ADR max% A pD2 ADR A max%
6.01+0.12 100 --0.10+0.12 --3+3 6NS 6.00_+0.15 100 -0.10_+0.11 --4-+8 5NS 5.77-+0.04 100 -0.06-t-0.10 -4-+4 7 NS
5.86-+0.10 100 -0.01_0.09 - 1_10 6NS 5.87-+0.15 100 -0.07-t-0.08 -11-+9 5NS 5.64-+0.08 100 -0.22_+0.11 -11_+13 7NS
amplitude of tile maximal response was 102+0.10% compared with a previous or subsequent control curve on the same preparation). Galanin did not affect the response of the vessels to a single concentration (10 -6 M) or a full concentration-response curve of adrenaline ranging from 10 - 7 to 20 - 5 (Tables 2, 3).
Discussion
This study illustrates the widespread distribution of ga- lanin (or closely related peptides) amongst the verte- brates by showing the presence of a galanin-like peptide in the autonomic nervous system of a teleost fish. It also demonstrates a direct stimulatory action of galanin, albeit weak, on the activity of the gut smooth muscle and on gut vessels. As a background to the study of the distribution of GLI nerves, the location and size of ganglion cells in the cranial and spinal pathways to the anterior gut have been investigated. The presence of enteric ganglionated plexuses in fish, as in other verte- brates, is well established (Kirtisinghe 1940; Burnstock 1959; Nilsson 1983). Similarly, the presence of nodose ganglion close to the vagus root, and of the large coeliae ganglion in the right sympathetic chain is well known (Nilsson 1983), but little information has previously been available regarding the occurrence of ganglion cells along the splanchnic and vagus nerves. We now report that a considerable number of ganglion cells are present in microganglia along the vagal branches to the gut; this agrees with the situation in the toad, Bttfo marhms, where these ganglia are considered to the parasympa- thetic (Gibbins et al. 1987). The distribution of ganglion cells along the splanchnic nerve of the Atlantic cod has been reported previously (Abrahamsson 1979).
The ganglion cells in the Atlantic cod are larger than the majority of ganglion cells in small mammals, that range between 25 and 50 ~tm in diameter (as measured from previously published micrographs in Lindh et al. 1986; Macrae et al. 1986; Skak-Nielsen et al. 1987; Gib- bins 1990). The large size cannot be a special feature in fish, since neurons in the vagus and the coeliac gangli- on of the rainbow trout, Oncorhynchus no'kiss, are only about half the size of those in the Atlantic cod (own results and measured from Lundin and Holmgren 1989).
The presence of the majority of the GLI ganglion cells in the vagal branches to the stomach, with only a few cells in the plexuses of the stomach, and none in the coeliac ganglion, is different from mammals. In mammals, galanin ganglion cells occur frequently in the myenteric and/or submucous plexuses of the gut (Ekblad et al. 1985b; Melander et al. 1985; Gonda et al. 1989) and the presence of galanin-like immunoreactivity in sympathetic ganglia is well established (Kummer 1987; Lindh et al. 1989; Ahr6n et al. 1990). Galanin-like im- munoreactivity also occurs in sympathetic ganglia in some species of lizards (Gibbins et al. 1989) and in the toad, Bufo marhms (Morris et al. 1989). However, in the toad, many of the postganglionic neurons in the vagus nerve contain a somatostatin-like peptide and also show weak galanin-like immunoreactivity (Morris et al. 1989); these cells may correspond to the GLI cells in the Atlant- ic cod vagus. The possibility that the ganglion cells along the Atlantic cod vagus (which is truly a vagosympathetic trunk, Nilsson 1983) are to some extent of sympathetic origin cannot, however, be complete!y ruled out, and the clusters of ganglion cells close to the gut wall may contain elements that could be considered enteric.
Both the ligation experiments and the tracings of the axons in the teased vagus indicate that the GLI neurons project either cranially or distally. The neurons project- ing distally are likely to be involved in some aspect of the control of the gut functions, although denervations failed to show a reduction in the number ofimmunoreac- tive neurons in the gut (see below). The targets of the centrally projecting neurons are more uncertain. Fibres can be seen running through the coeliac gangion, and may affect the sympathetic neurons there; however, an- other possibility is that the neurons run cranially for only a short distance and then follow more cranial branches of the nerve, e.g. to the oesophagus.
The nerve net in the stomach is denser than that in the intestine, contrary to several reports from studies in mammals, in which the innervation of the stomach is less dense and in which innervation increases distally (R6kaeus et al. 1984; Ekblad et al. 1985 b; Melander et al. 1985). Recent unpublished immunohistochemical studies on the intestine of two stomachless fish, the tilapia Or- eochromis nwssambicus and the goldfish Carassius aura- ttts (A.J. Kiliaan et al., unpublished) suggest an anal in-

543
crease of G L I fibres; this confirms the large variat ion in innervation patterns amongst fish observed in numer- ous other studies. Similarly, G L I nerves are observed in the mucosa o f the intestine in the tilapia and in the s tomach of the Atlantic cod, but arc absent in the gold- fish intestine.
The lack of effect in the denervation experiments on the innervation o f the gut wall may be at tr ibutable to the fact that most GLI ganglion cells projecting to the stomach are situated in the fine branches close to the gut wall, whereas the denervations were made further away from the stomach wall. Surgical ligations closer to the s tomach are technically difficult and cause bleed- ing. The ligation experiments, however, confirm the re- sults of the tracing studies, viz. that G L I neurons in the vagus project both centrally and distally.
Since the initial report of the discovery and function of galanin (Tatemoto et al. 1983), there have been nu- merous studies in mammals reporting both excitatory and inhibitory actions of galanin on smooth muscle ac- tivity of the gut, varying with species and part o f the gut. The excitatory effects are in general direct on the smooth mucle cells, whereas the inhibitory effects may be caused by hyperpolarisat ion of excitatory neurons (neuromodulat ion) or a direct effect on the muscle cells (Ekblad et al. 1985a; Fox et al. 1986, 1988; Yau et al. 1986; Gonda et al. 1989). The direct nature of the excit- a tory effect of galanin in the gut o f the Atlantic cod thus corresponds to the situation in mammals .
The innervation pattern of the Atlantic cod s tomach implies that action is exerted on mucosal events, as well as on gut motility. This has not been further investigated in the present study.
The contractile effects found in the arteries of the Atlantic cod gut in the present study agree with the pres- sure effect o f galanin reported in the toad, Bufo marinus (Courtice 1991), but is contrary to the vasodilator effect of galanin found in the arterioles of the small intestine of the guinea pig (Kotecha and Neild 1991). Galanin coexists with catecholamines in sympathet ic vascular neurons of several vertebrates, including placental mam- mals, marsupials, lizards and toads (Kummer 1987; Gib- bins et al. 1989; Lindh et al. 1989; Morris et al. 1989; Ahr~n et al. 1990), and an interaction or cofunction be- tween the adrenergic transmitter and galanin might be expected. It has been suggested that galanin augments the electrically induced contraction of the rat vas defer- ens by potentiating the release of noradrenaline f rom sympathetic nerves (Ohhashi and Jacobowitz 1985), whereas other studies (in the mammal ian gut) indicate postjunctional interactions with other hormones/ t rans- mitters ofgalanin in its action on smooth muscle (Rat tan 1991). However, galanin does not affect the adrenaline- induced contractions of the arteries of the Atlantic cod in the present experiments, indicating that the contractile effect of galanin on these arteries does not involve a potentiat ion of the adrenergic effect.
Acknowledgements. This study was supported by the Swedish Natu- ral Science Research Council and the Adlersbertska Research Foundation. We acknowledge the expert help of Mrs. Christina
Hagstr6m (immunohistochemistry), Ms. Inger Holmquist (photo- graphy) and Mrs. Ann Wikstr6m (muscle strip experiments).
References
Abrahamsson T (1979) Axonal transport of adrenaline, noradrcna- line and phenylethanolamine-N-methyl transferase (PNMT) in sympathetic neurons of the Atlantic cod, Gadus morhua. Acta Physiol Seand 105:316-325
Ahr6n B, B6ttcher G, Kowalyk S, Dunning BE, Sundler F, Ta- borsky GJ (1990) Galanin is co-localised with noradrenaline and neuropeptide-Y in dog pancreas_ and celiac ganglion. Ccll Tissue Res 261:49-58
Bauer FE, Zintel A, Kenny M J, Calder D, Ghatei MA, Bloom SR (1989) Inhibitory effect ofgalanin on postprandial gastroin- testinal motility and gut hormone release in humans. Gastroen- terology 97: 260-264
Bishop AE, Polak JM, Bauer FE, Christofidcs ND, Carlei F, Bloom SR (1986) Occurrence and distribution of a newly dis- covered peptide, galanin, in the mammalian enteric nervous system. Gut 27:849-857
Burnstock G (1959) The innervation of the gut of the brown trout, Sahno trutta. Q J Microsc Sci 100:199-220
Costa M, Buffa R, Furness JB, Solcia E (1980) Immunohistochemi- cal localization of polypeptidcs in peripheral autonomic nerves using whole mount preparations. Histochemistry 65:157-165
Courtice GP (1991) Effect of neuropeptide Y and galanin on auto- nomic control of heart rate in the toad, Bufo marhlus. J Auton Nerv Syst 33:231-238
Dunning BE, Ahr~n B, Veith RC, B6ttcher G, Sundler F, Taborsky GJ Jr (1986) Galanin: a novel pancreatic neuropeptide. Am J Physiol 251:E 127-E 133
Ekblad E, H,hkanson R, Sundler F, Wahlestedt C (1985a) Galanin: neuromodulatory and direct contractile effects on smooth mus- cle preparations. Br J Pharmacol 86:241-246
Ekblad E, R6kaeus/~, Hfikansson R, Sundler F (1985b) Galanin nerve fibers in the rat gut: distribution, origin and projections. Neuroscience 16:355-363
Fox JET, McDonald TJ, Kostolanska F, Tatemoto K (1986) Ga- lanin: an inhibitory peptidc of the canine small intestine. Life Sci 39:103-110
Fox JET, Brooks B, McDonald TJ, Barnett W, Kostolanska F, Yanaihara C, Yanaihara N, R6kaeus/~ (1988) Actions ofgalan- in fragments on rat, guinea-pig, and canine intestinal motility. Peptides 9:1183-1189
Gibbins IL (1990) Target-related patterns of co-existence of neu- ropeptide-Y, vasoactive intestinal peptide, enkephalin and sub- stance P in cranial parasympathetic neurons innervating the facial skin and exocrine glands of guinea-pigs. Neurosciencc 38: 541-560
Gibbins IL, Campbell G, Morris JL, Nilsson S, Murphy R (1987) Pathway-specific connections between pepide-containing pre- ganglionic and postganglionic neurons in the vagus nerve of the toad, Bttfo marbltts. J Auton Nerv Syst 20:43-55
Gibbins IL, Morris JL, Holmgrcn S (1989) Distribution of neu- ropeptide Y and galanin in cardiovascular sympathetic neurons of Australian lizards. Proc Int Union Physiol Sci 17:62
Gonda T, Daniel EE, McDonald TJ, Fox JET, Brooks BD, Oki M (1989) Distribution and function of enteric GAL-IR nerves in dogs: comparison with VIP. Am J Physiol 256:G 884-G 896
Ju G, H6kfelt T, Brodin E, Fahrenkrug J, Fischer MA, Frey P, Elde RP, Brown JC (1987) Primary sensory neurons of the rat showing calcitonin gene-related peptide immunorcactivity and their relation to substance P-, somatostatin-, galanin-, va- soaetive intestinal polypeptide- and cholecystokinin-immunorc- active ganglion cells. Cell Tissue Res 247:417-431
Kirtisinghe P (1940) The myenteric nerve plexus in some lower chordates. Q J Microsc Sci 81:521-539

544
Konopka LM, Parsons RL (1989) Characteristics of the galanin- induced depolarization of mudpuppy parasympathetic post- ganglionic neurons. Neurosci Lett 99:142-146
Kotccha N, Neild TO (1991) Effects of vasodilator peptides on arterioles of the small intestine. In: Perry MA, Garlick DG (eds) Progress in microciculation research. Proceedings of the 6th Australian and New Zealand Symposium, University of New South Wales, pp 78-80
Kummer W (1987) Galanin- and neuropeptide Y-like immunoreac- tivities coexist in paravertebral sympathetic neurones of the cat. Neurosci Lett 78 : 127-131
Lindh B, Hfkfelt T, Elfvin LG, Tercnius L, Fahrenkrug J, Elde R, Goldstein M (1986) Topography of NPY-, somatostatin- and VIP-immunoreactive, neuronal subpopulations in the guin- ea pig celiac-superior mesenteric ganglion and their projection to the pylorus. J Neurosci 6:2371-2383
Lindh B, Lundberg JM, Hrkfelt T (1989) NPY-, galanin-, VIP] PHI-, CGRP- and substance P-immunorcactive neuronal sub- populations in cat autonomic and sensory ganglia and their projections. Cell Tissue Res 256:259-273
Lindskog S, Ahrrn B (1987) Galanin: effects on basal and stimu- lated insulin and glucagon secretion in the mouse. Acta Physiol Scand 129:305-309
Lundin K, Holmgrcn S (1989) The occurrence and distribution of peptide- or 5-HT-containing nerves in the swimbladder of four different species of telcosts, Gadus morhua, Ctenolabrus rupestris, Anguilla anguilla, Sahno gairdneri. Cell Tissue Res 257: 641-647
Macrae LM, Furness JB, Costa M (1986) Distribution of sub- groups of noradrenaline neurons in the coeliac ganglion of the guinea-pig. Cell Tissue Res 244:173-180
Marangos PJ, Schmeichel DE, Oertel WH (1981) Neuron-specific enolase: a functional marker for the diffuse neuro-endocrine system. In: Bloom SR, Polak JM (eds) Gut hormones. Chur- chill Livingstone, Edinburgh London Melbourne, pp 101-106
McKeon TW, Carraway RE, Konopka LM, Parsons RL (1990) Distribution of galanin-like peptide in various tissues of Nectur- us maculosus. Cell Tissue Res 262:461-466
Melander T, Hrkfelt T, Rrkaeus/~, Fahrenkrug J, Tatcmoto K, Mutt V (1985) Distribution of galanin-like immunoreactivity in the gastro-intestinal tract of several mammalian species. Cell Tissue Res 239:253-270
Morris JL, Gibbins IL, Osborne PB (1989) Galanin-like immunore- activity in sympathetic and parasympathetic neurons of the toad, Bufo mariJms. Neurosci Lett 102:142-148
Morris JL, Gibbins IL, Holmgren S (1992) Galanin is more com- mon than NPY in vascular sympathetic neurons of the brush- tailed possum. Regul Pept 37:101-109
Nilsson S (1983) Autonomic nerve function in the vertebrates. Springer, Berlin Heidelberg New York
Ohhashi T, Jacobowitz DM (1985) Galanin potentiates electrical stimulation and exogenous norepinephrine-induced contrac- tions in the rat vas deferens. Regul Pept 12:163-171
Parsons RL, Konopka LM (1990) Galanin-induced hyperpolariza- tion of mudpuppy neurons is calcium dependent. Neurosci Lett 115:207-212
Rattan S (1991) Role of galanin in tiae gut. Gastroenterology 100:1762-1768
Rrkaeus/~, Melander T, H6kfelt T, Lundberg JM, Tatemoto K, Carlquist M, Mutt V (1984) A galanin-like peptide in the central nervous system and intestine of the rat. Neurosci Lett 47:161- 166
Schepp W, Prinz C, Tatge C, Hfikansson R, Schusdziarra V, Clas- sen M (1990) Galanin inhibits gastrin release from isolated rat gastric G-cells. Am J Physiol 258:G596-G602
Skak-Nielsen T, Poulsen SS, Hoist JJ (1987) Immunohistochemical detection of ganglia in the rat stomach serosa, containing neu- rons immunoreactive for gastrin-releasing peptide and vasoac- tive intestinal peptide. Histochemistry 87:47-52
Sundell K, Bjfrnsson BTh (1988) Kinetics of calcium fluxes across the intestinal mucosa and the marine teleost, Gadus morhua, measured using an in vitro perfusion method. J Exp Biol 140:171-186
Tatemoto K, Rrkaeus A, J6rnvall H, McDonald TJ, Mutt V (1983) Galanin - a novel biologically active peptide from porcine intes- tine. FEBS Lett 164:124-128
Yagci RV, Alptekin N, Rossowski W J, Brown A, Coy DH, Ertan A (1990) Inhibitory effect ofgalanin on basal and pentagastrin- stimulated gastric acid secretion in rats. Scand J Gastroenterol 25:853-858
Yau WM, Dorsett JA, Youther ML (1986) Evidence for galanin as an inhibitory neuropeptide on myenteric cholinergic neurons in the guinea pig small intestine. Neurosci Lett 72:305-308
Related Documents











