Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the book Methods in Enzymology, Vol. 521 published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at: http://www.elsevier.com/locate/permissionusematerial From: Dasiel O. Borroto-Escuela, Wilber Romero-Fernandez, Pere Garriga, Francisco Ciruela, Manuel Narvaez, Alexander O. Tarakanov, Miklós Palkovits, Luigi F. Agnati, Kjell Fuxe, G Protein-Coupled Receptor Heterodimerization in the Brain. In Michael Conn, editor: Methods in Enzymology, Vol. 521, Burlington: Academic Press, 2013, pp. 281-294. ISBN: 978-0-12-391862-8 © Copyright 2013 Elsevier Inc. Academic Press

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use.
This chapter was originally published in the book Methods in Enzymology, Vol. 521 published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at:
http://www.elsevier.com/locate/permissionusematerial
From: Dasiel O. Borroto-Escuela, Wilber Romero-Fernandez, Pere Garriga, Francisco Ciruela, Manuel Narvaez, Alexander O. Tarakanov, Miklós Palkovits,
Luigi F. Agnati, Kjell Fuxe, G Protein-Coupled Receptor Heterodimerization in the Brain. In Michael Conn, editor: Methods in Enzymology, Vol. 521, Burlington:
Academic Press, 2013, pp. 281-294. ISBN: 978-0-12-391862-8
© Copyright 2013 Elsevier Inc. Academic Press

Author's personal copy
CHAPTER FIFTEEN
G Protein–Coupled ReceptorHeterodimerization in the BrainDasiel O. Borroto-Escuela*, Wilber Romero-Fernandez*,Pere Garriga†, Francisco Ciruela‡, Manuel Narvaez},Alexander O. Tarakanov}, Miklós Palkovits||, Luigi F. Agnati#,Kjell Fuxe*,1*Department of Neuroscience, Karolinska Institutet, Stockholm, Sweden†Departament d’Enginyeria Quımica, Universitat Politecnica de Catalunya, Barcelona, Spain‡Unitat de Farmacologia, Departament Patologia i Terapeutica Experimental, Universitatde Barcelona, Barcelona, Spain}Department of Physiology, School of Medicine, University of Malaga, Malaga, Spain}Russian Academy of Sciences, St. Petersburg Institute for Informatics and Automation,Saint Petersburg, Russia||Human Brain Tissue Bank, Semmelweis University, Budapest, Hungary#IRCCS Lido, Venice, Italy1Corresponding author: e-mail address: [email protected]
Contents
1.
MetISShttp
Introduction
hods in Enzymology, Volume 521 # 2013 Elsevier Inc.N 0076-6879 All rights reserved.://dx.doi.org/10.1016/B978-0-12-391862-8.00015-6
282
2. In Situ PLA for Demonstrating Receptor Heteromers and Their Receptor–ReceptorInteractions in Brain Tissue
283 3. Brain Tissue Preparation 283 4. Proximity Probes: Conjugation of Oligonucleotides to Antibodies 287 5. PLA Reactions, Reagents, and Solutions 287 6. Quantitative PLA Image Analysis 289 7. Advantages and Disadvantages of the PLA Method 289 8. Application 291 Acknowledgments 293 References 293Abstract
G protein–coupled receptors (GPCRs) play critical roles in cellular processes and signal-ing and have been shown to form heteromers with diverge biochemical and/or phar-macological activities that are different from those of the corresponding monomers orhomomers. However, despite extensive experimental results supporting the formationof GPCR heteromers in heterologous systems, the existence of such receptor hetero-complexes in the brain remains largely unknown, mostly because of the lack of appro-priate methodology. Herein, we describe the in situ proximity ligation assay procedureunderlining its high selectivity and sensitivity to image GPCR heteromers with confocal
281

282 Dasiel O. Borroto-Escuela et al.
Author's personal copy
microscopy in brain sections. We describe here how the assay is performed and discussadvantages and disadvantages of this method compared with other availabletechniques.
1. INTRODUCTION
Gprotein–coupled receptors (GPCRs) play critical roles in cellular pro-
cesses and signaling and have been shown to form heteromers with diverge
biochemical and/or pharmacological activities that are different from those
of the corresponding monomers or homomers. The idea of the existence
of direct interactions between two different GPCRs at the level of the plasma
membrane has its origin in 1980/1981 on the basis of the discovery that pep-
tides like cholecystokinin-8 (CCK-8) and substance P could modulate the
density, and especially the affinity, of distinct monoamine receptors in mem-
brane preparations from the CNS with in vivo functional correlates (Agnati,
Fuxe, Zini, Lenzi, & Hokfelt, 1980; Fuxe et al., 1981). These initial findings
were in line with the previous discovery of negative cooperativity between
b adrenergic receptors in 1974/1975 by Lefkowitz and colleagues, indicating
the possible existence of homodimers of b adrenergic receptors leading to
site–site interactions in recognition (Limbird, Meyts, & Lefkowitz, 1975).
Nevertheless, despite extensive experimental results supporting the for-
mation of GPCR heteromers in heterologous systems, the existence of such
receptor heterocomplexes in their native environment remains largely un-
known, mostly because of the lack of appropriate methodology. For in-
stance, until recent years, the methods that have been developed to study
receptor–receptor interactions in heteromers require that genetic constructs
be expressed in the cells to enable detection of the receptor interactions, thus
excluding the use of tissue samples (Borroto-Escuela, Garcia-Negredo,
Garriga, Fuxe, & Ciruela, 2010; Ferre et al., 2009; Fuxe et al., 2012).
In order to demonstrate in native tissue the existence of GPCR
heteromers, especially in a manner that can be generally applicable to differ-
ent receptor pairs, a well-characterized in situ proximity ligation assay (in situ
PLA) has been adapted to confirm the existence of GPCR heteromers in
brain slices ex vivo.
In situ PLA is based on a pair of antibodies that can bind to target proteins
and to which oligonucleotides have been attached. When the so-called
proximity probes recognize a target, for example, if the two target receptors
interact, the attached oligonucleotides are brought into a sufficiently close

283G Protein–Coupled Receptor Heterodimerization in the Brain
Author's personal copy
spatial proximity to allow them to join followed by ligation of the two linear
oligonucleotides into a circular DNA molecule. This newly formed DNA
circle strand can serve as a template for rolling circle amplification (RCA),
resulting in a long single-stranded rolling circle product (RCP) attached to
one of the proximity probes. As the RCP is linked to the proximity probe, it
is attached at the site where the proximity probe bound, which means that it
can be used to reveal the location of the receptor complex (Soderberg et al.,
2006, 2007). The RCPs can then be detected and quantified by hybridizing
fluorescent oligonucleotides to the repeated sequences of the RCPs, render-
ing them visible by fluorescence microscopy (Fig. 15.1).
Herein, we describe the in situ PLA procedure as a high selectivity and
sensitivity assay to image GPCR heteromers in brain sections by confocal
microscopy. We describe how the assay is performed and discuss advantages
and disadvantages of this method compared with other available techniques.
2. In Situ PLA FOR DEMONSTRATING RECEPTORHETEROMERS AND THEIR RECEPTOR–RECEPTOR
INTERACTIONS IN BRAIN TISSUEIn situ PLA has previously been performed to confirm the existence of
striatal A2AR–D2R heteromers (Trifilieff et al., 2011). The PLA technique
involved the use of two primary antibodies of different species directed to
either D2R or to A2AR (Fig. 15.2). We recommend to use in situ PLA also
to indicate the ratio between heteromers versus total number of the two par-
ticipating receptor populations, using in addition to Western blots, receptor
autoradiography, and biochemical binding methods, the two latter methods
showing the densities and affinities of the two functional receptor populations.
This will also help to normalize the heteromer values for comparison between
groups in addition to evaluating the potential changes in the total number of
the two receptor populations. The person doing the PLA measurements
should be blind to the code of the experimental groups studied.
3. BRAIN TISSUE PREPARATION
As for all antibody-based staining methods, the samples should be suf-
ficiently pretreated to fit the primary antibodies with respect to fixation,
permeabilization, and antigen retrieval of the tissue to be investigated. As
the protocols provided in this section are general, it is highly suggested to
apply the same protocol that has been working previously for your receptor
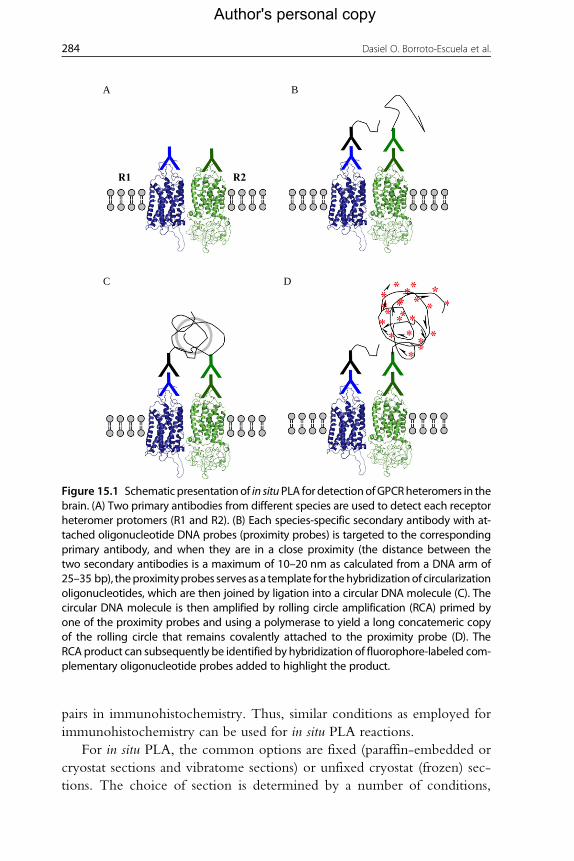
A B
C D
R1 R2
Figure 15.1 Schematic presentationof in situPLA for detectionofGPCRheteromers in thebrain. (A) Two primary antibodies from different species are used to detect each receptorheteromer protomers (R1 and R2). (B) Each species-specific secondary antibody with at-tached oligonucleotide DNA probes (proximity probes) is targeted to the correspondingprimary antibody, and when they are in a close proximity (the distance between thetwo secondary antibodies is a maximum of 10–20 nm as calculated from a DNA arm of25–35 bp), theproximityprobes serves asa template for thehybridizationof circularizationoligonucleotides, which are then joined by ligation into a circular DNA molecule (C). Thecircular DNA molecule is then amplified by rolling circle amplification (RCA) primed byone of the proximity probes and using a polymerase to yield a long concatemeric copyof the rolling circle that remains covalently attached to the proximity probe (D). TheRCA product can subsequently be identified by hybridization of fluorophore-labeled com-plementary oligonucleotide probes added to highlight the product.
284 Dasiel O. Borroto-Escuela et al.
Author's personal copy
pairs in immunohistochemistry. Thus, similar conditions as employed for
immunohistochemistry can be used for in situ PLA reactions.
For in situ PLA, the common options are fixed (paraffin-embedded or
cryostat sections and vibratome sections) or unfixed cryostat (frozen) sec-
tions. The choice of section is determined by a number of conditions,
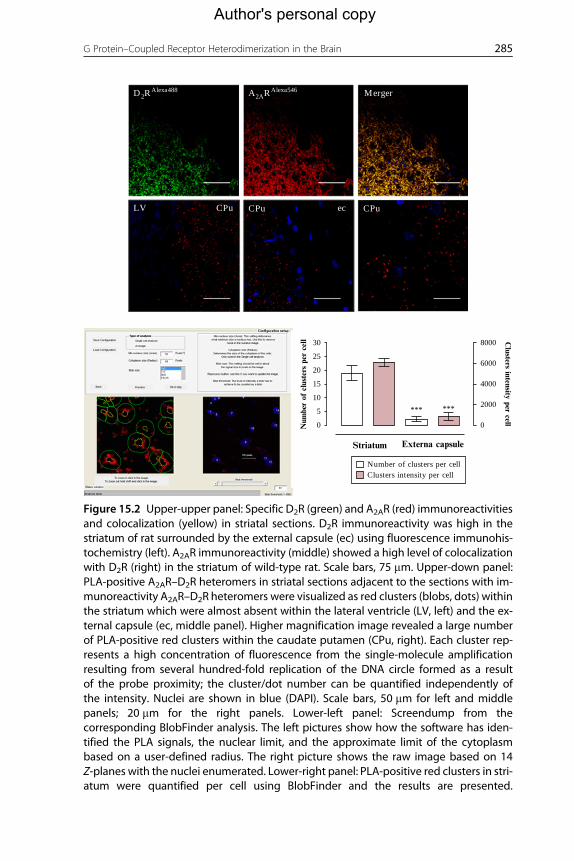
D2RAlexa488 A2ARAlexa546 Merger
LV CPu CPuecCPu
Striatum
0 0
2000
4000
6000
8000
***
Number of clusters per cellClusters intensity per cell
***5
10
15
20
25
30
Num
ber
of c
lust
ers
per
cell C
lusters intensity per cell
Externa capsule
Figure 15.2 Upper-upper panel: Specific D2R (green) and A2AR (red) immunoreactivitiesand colocalization (yellow) in striatal sections. D2R immunoreactivity was high in thestriatum of rat surrounded by the external capsule (ec) using fluorescence immunohis-tochemistry (left). A2AR immunoreactivity (middle) showed a high level of colocalizationwith D2R (right) in the striatum of wild-type rat. Scale bars, 75 mm. Upper-down panel:PLA-positive A2AR–D2R heteromers in striatal sections adjacent to the sections with im-munoreactivity A2AR–D2R heteromers were visualized as red clusters (blobs, dots) withinthe striatum which were almost absent within the lateral ventricle (LV, left) and the ex-ternal capsule (ec, middle panel). Higher magnification image revealed a large numberof PLA-positive red clusters within the caudate putamen (CPu, right). Each cluster rep-resents a high concentration of fluorescence from the single-molecule amplificationresulting from several hundred-fold replication of the DNA circle formed as a resultof the probe proximity; the cluster/dot number can be quantified independently ofthe intensity. Nuclei are shown in blue (DAPI). Scale bars, 50 mm for left and middlepanels; 20 mm for the right panels. Lower-left panel: Screendump from thecorresponding BlobFinder analysis. The left pictures show how the software has iden-tified the PLA signals, the nuclear limit, and the approximate limit of the cytoplasmbased on a user-defined radius. The right picture shows the raw image based on 14Z-planes with the nuclei enumerated. Lower-right panel: PLA-positive red clusters in stri-atum were quantified per cell using BlobFinder and the results are presented.
285G Protein–Coupled Receptor Heterodimerization in the Brain
Author's personal copy

286 Dasiel O. Borroto-Escuela et al.
Author's personal copy
including the time and skill of the investigator. However, careful consider-
ation of the fixation protocol is especially necessary to ensure the optimal
preservation of the morphology of the specimen and target antigen (recep-
tors). Incorrect specimen preparation can block or impede antigen labeling
in the tissue. Unfortunately, the methods that are best for the preservation of
tissue structure do so by modifying proteins, thereby reducing the efficiency
of antigen detection. In cases of failure, it is important to try with multiple
different conditions before you give up the in situ PLA.
1. Because of the ease of use, fixed frozen free-floating sections are often
employed in most of the in situ PLA experiments. First, animals are anes-
thetized by an intraperitoneal injection of, for example, pentobarbital
(60 mg/ml, [0.1 ml/100 g]) and perfused intracardially with 30–50 ml
of ice-cold 4% paraformaldehyde (PFA) in 0.1M phosphate-buffered sa-
line (PBS), pH 7.4, solution. After perfusion, brains are collected and
transferred into well-labeled glass vials filled with 4% PFA fixative solu-
tion for 6 h. Then, the brain pieces are placed in sucrose 20% in PBS and
incubated for 24 h until sections (10–30 mm thick) are generated and
serially collected using a cryostat. Alternatively to the use of fixed free-
floating sections, we can use tissue fixed frozen sections attached to
microscopy slides. Mounted sections on slides must be kept at �20 �Cuntil use. Encircle the tissue section on the glass slide by creating a hydro-
phobic barrier using a grease pen or a silicon mask and proceed as follows.
2. Wash the fixed free-floating sections four times with PBS, then incubate
with the blocking solution (10% fetal bovine serum [FBS] and 0.5% Tri-
ton X-100 or Tween-20 in Tris buffer saline [TBS], pH 7.4) for 2 h at
room temperature or 1 h at 37 �C and then follow the Protocol step 5.
To reduce the likelihood of unspecific binding of the antibodies to the tissue,
the tissue needs to be blocked by a blocking agent, such as bovine serum
albumin (BSA; by adding 1 ml BSA (10 mg/ml) and 1 ml sonicated salmon
sperm DNA (0.1 mg/ml) to 38 ml of 0.5% Triton X-100 or Tween-20 in
TBS, pH 7.4; Leuchowius,Weibrecht, & Soderberg, 2011) or animal serum
like 10% FBS (if animal serum is used, make sure that it is sterile filtered, as
unfiltered serum may increase the amount of background signals). Use the
blocking agent best suited for the antibodies used. Each time must be
Quantification of A2AR–D2R heteromers demonstrates highly significant differences inPLA clusters per cell between caudate putamen and external capsule (***P<0.001by Student's t-test).

287G Protein–Coupled Receptor Heterodimerization in the Brain
Author's personal copy
checked that the reaction should never become dry as this will cause high
background.
4. PROXIMITY PROBES: CONJUGATION OFOLIGONUCLEOTIDES TO ANTIBODIES
Proximity probes are created through the attachment of oligonucle-
otides to antibodies. The oligonucleotide component of the proximity pro-
bes can be covalently coupled to an antigen-binding component or attached
to secondary antibodies specific for antibodies raised in different species.
This approach avoids the need to conjugate the oligonucleotide components
to each primary antibody pair.
Several different types of chemistry can be used for the conjugation
of oligonucleotides to antibodies. Mainly three methods have been used
extensively in recent years: the maleimide/NHS-esther chemistry (SMCC;
Soderberg et al., 2006), the succinimidyl 4-hydrazinonicotinate acetone
hydrazone (SANH; Leuchowius et al., 2011), or the commercially available
Antibody-Oligonucleotide All-in-One Conjugation Kit from Solulink
company (http://www.solulink.com/), based on two complementary het-
erobifunctional linkers (Sulfo-S-4FB (formylbenzamide) and S-HyNic
(hydrazino-nicotinamide)). Because the act of conjugation can severely af-
fect the ability of some antibodies to bind antigen, it may be necessary to
analyze different antibodies, conjugation chemistries, and reaction condi-
tions to obtain suitable proximity probes.
Another possibility, not less useful, is to buy directly proximity probes
from specialized companies on antibody-oligonucleotide conjugation, for
example, Duolink (Uppsala, Sweden; http://www.olink.com/).
5. PLA REACTIONS, REAGENTS, AND SOLUTIONS
1. If primary antibodies directly labeled with oligonucleotides or primary
antibodies are used in combination with secondary proximity probes
(see Protocol step 4), the conditions for incubation with the primary
antibodies should be chosen according to the manufacturer’s recom-
mendations or will have to be identified by the users. For instance, in-
cubate the tissue with the primary antibodies diluted into a suitable
concentration in the wash buffer (0.5% Triton X-100 or Tween-20
in TBS, pH 7.4) at 1–2 h at 37 �C or þ4 �C overnight.

288 Dasiel O. Borroto-Escuela et al.
Author's personal copy
2. After incubation of the primary antibodies at conditions specified above
or determined by the user, excess antibody should be removed. Wash
the slides four times, 5 min each time, with wash buffer.
3. In the mean time, if primary antibodies are used in combination with
secondary proximity probes, dilute the proximity probes to a suitable
concentration in the wash buffer. It is important to use the same buffer
as those for the primary antibody to avoid background staining.
Apply the proximity probe mixture to the sample and incubate for
1 h at 37 �C in a humidity chamber. Do not allow the samples to
dry, as this will cause also artifacts. To remove unbound proximity pro-
bes, wash the slides four times, 5 min each time, with wash buffer.
4. Prepare the hybridization–ligation solution. To ensure optimal condi-
tions for the enzymatic reactions, the sections should be soaked for
1 min in 1� ligation buffer (10 mM Tris–acetate, 10 mM magnesium
acetate, 50 mM potassium acetate, pH 7.5; Soderberg et al., 2008),
prior to addition of the hybridization–ligation solution (final con-
centration: BSA (250 mg/ml), 1� T4 DNA ligase buffer, Tween-20
(0.05%), NaCl 250 mM, ATP 1 mM, and the circularization or con-
nector oligonucleotides 125–250 nM). Circularization or connector
oligonucleotides can be designed and synthesized as described previ-
ously (Soderberg et al., 2008). Remove the soaking solution (ligation
buffer) and add T4 DNA ligase at a final concentration of 0.05 U/ml tothe hybridization–ligation solution. Vortex briefly to mix the ligase
with the solution. Apply the mixture immediately to the sections
and slides in a humidity chamber for 30 min at 37 �C.5. Wash the sections three times with wash buffer in a washing jar for
5 min to remove excess connector oligonucleotides.
6. Prepare the RCA mixture. Soak the sections in 1� RCA buffer
(50 mM Tris–HCl, 10 mM MgCl2, 10 mM (NH4)2SO4, pH 7.5
adjusted with HCl) for 1 min. Remove the soaking solution and
add the RCA solution (final concentration: phi-29 polymerase
0.125–0.200 U/ml, BSA (250 mg/ml), 1� RCA buffer, Tween-20
(0.05%), and dNTP (250 mM for each)). Vortex briefly the RCA solu-
tion and incubate in a humidity chamber for 100 min at 37 �C.7. Wash the sections three times with wash buffer in a washing jar for
5 min. Prepare the detection solution (final concentration: BSA
(250 mg/ml), 2� sodium citrate, sodium chloride buffer, and the fluo-
rescence detection (e.g., Texas Red or Alexa 555)-oligonucleotide
strand (6.25 nM)), see Soderberg et al., 2008) and incubate the sections
in a humidity chamber for 30 min at 37 �C. Keep the detection

289G Protein–Coupled Receptor Heterodimerization in the Brain
Author's personal copy
solution in the dark to prevent fluorophore bleaching. From now on,
all reactions and wash steps should be performed in the dark.
8. Wash the sections twice, each time with wash buffer in a washing jar for
5 min in the dark.
9. Wash the sections once with TBS in a washing jar for 5 min in the dark.
10. Dry and mount sections with the appropriate mounting media (e.g.,
VectaShield or Dako). The sections should be protected against
light and can be stored for several days at 4 �C or for several months
at �20 �C.
6. QUANTITATIVE PLA IMAGE ANALYSIS
1. Visualize the sections with fluorescence microscopy equipped with
excitation/emission filters compatible with the fluorophores used.
The in situ PLA signals have a very characteristic appearance that is easily
recognized once you know what to look for. The PLA detection reac-
tion products are seen as bright fluorescent puncta of submicrometer size
(see Fig. 15.2). By moving the focus up and down in your sample tissue,
you should note appearance and disappearance of PLA signals. Up to a
certain density of PLA signals, they appear as discrete dots (puncta, blobs)
that can be easily enumerated using image analysis software.
2. Analyze the captured images by image techniques to quantify the num-
ber of dots (Fig. 15.2, lower panel). Many commercial image analysis
software packages can also be used in addition to free software packages,
such as BlobFinder (Allalou &Wahlby, 2009) or Cellprofiler (Carpenter
et al., 2006). The BlobFinder is a free software tool developed by the
Centre for Image Analysis-Uppsala University for such objectives (the
freeware is available for download from http://www.cb.uu.se/�amin/BlobFinder/; Fig. 15.2, lower panel). At higher densities per num-
ber of nuclei, the dots start to coalesce, thus making it more difficult to
resolve and enumerate individual signals. It is important to use the same
settings for image acquisition for all images in a series.
7. ADVANTAGES AND DISADVANTAGES OF THE PLAMETHOD
Comparison with other methods to study receptor–receptor interactions in
heteromers, such as FRET, BRET, and bimolecular fluorescence complementation:
In situ PLA can offer advantages by permitting analyses of interactions

290 Dasiel O. Borroto-Escuela et al.
Author's personal copy
among any receptors for which suitable antibodies are available without
using genetic constructs. Such constructs can perturb cellular function
and cannot always be used, for example, in cells and tissues from subjects.
In situ PLA can be performed in all samples of cells and tissues, and the
method is highly suited to investigate human specimens collected from
biobanks, in order to investigate patterns of changes in heteromers that
could provide insights on the role of basic heteromer mechanisms or have
a diagnostic value (Nilsson et al., 2010). The method has also proven useful
to monitor the effects of different compounds like agonists and antagonists
or their combined treatment on the receptor heteromers in cells and tissue
(Borroto-Escuela et al., 2011). The information is obtained at a resolution
of individual cells or even of subcellular compartments, providing pro-
found insights into cellular heterogeneity in tissues. The method also
provides an enhanced sensitivity and selectivity compared with many other
methods as powerful RCA and dual target recognition are used (Clausson
et al., 2011).
As with any method there are limitations, for instance, in situ PLA cannot
be used with live cells, as it requires cell fixation and, in some cases, perme-
abilized cells. When studying receptor–receptor interactions, it is important
to remember that the method, like many other methods for studying
protein–protein interactions, can show that two proteins are in close prox-
imity and likely directly interact. Proteins can also interact indirectly
through an adapter protein. The maximal distance between two epitopes
to give rise to a signal with in situ PLA is 10–30 nm with direct-conjugated
proximity probes, and slightly longer when secondary proximity probes are
used. By changing the length of the oligonucleotides, the maximal distance
limits can be reduced or increased.
Other critical parameters for achieving good results is the use of
excellent antibodies. The antibodies must also be used under optimal
conditions taking into consideration parameters such as antibody concen-
tration, epitopes targeted by the antibodies, fixation, antigen retrieval,
blocking conditions, etc. A range of controls both positive and negative
ones should be used to guarantee the specificity of the PLA signal.
Positive controls can include cells where the protein is known to be
expressed, such as in certain cells or tissues or in cells transfected to ex-
press the protein. Negative controls include cells or tissues that do not
express the protein or where the protein has been knocked out or down-
regulated by, for example, siRNA.
291G Protein–Coupled Receptor Heterodimerization in the Brain
Author's personal copy
8. APPLICATION
In situ PLA has been used to study proteins and protein–protein inte-
ractions in a range of applications (Leuchowius et al., 2010; Nilsson et al.,
2010). In2011, themethodwas employed to studyGPCRsheteromers,mainly
adenosine A2A and dopamine D2 receptor heteromers in striatal sections
(Trifilieff et al., 2011) and dopamine D2R and D4R in transiently transfected
HEK293T cells (Borroto-Escuela et al., 2011). In addition, the methods were
employed to demonstrate for the first time the existence of FGFR1 and
5-HT1A receptor heterocomplexes in the rat hippocampus and dorsal and
median raphe in the midbrain (Fig. 15.3A; Borroto-Escuela et al., 2012).
In the analysis using in situ PLA, it is also important to determine the ratio
between heteromers versus total number of the two participating receptor
populations, using in addition to Western blots, receptor autoradiography,
and biochemical binding methods, the two latter methods showing the den-
sities and affinities of the two functional receptor populations. The relation-
ship between these parameters will help to normalize the heteromer values
for comparison between groups in addition to evaluating the potential
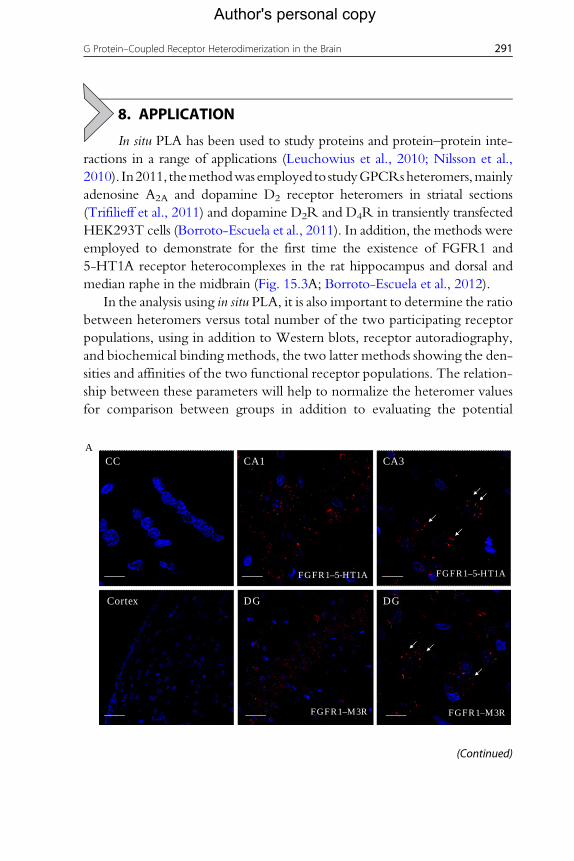
CC CA1
FGFR1–5-HT1A
FGFR1–M3R FGFR1–M3R
Cortex DG DG
FGFR1–5-HT1A
CA3A
(Continued)
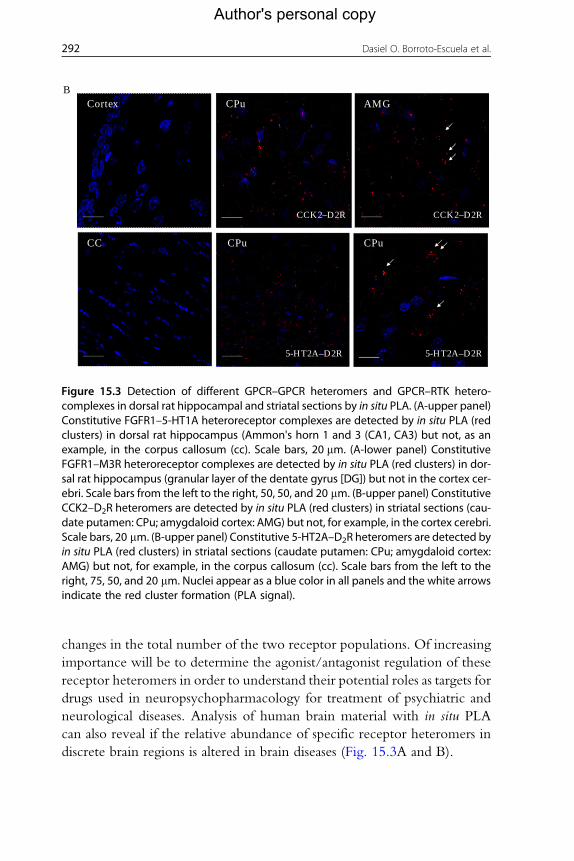
Cortex CPu
CPu CPu
BAMG
CCK2–D2R CCK2–D2R
5-HT2A–D2R5-HT2A–D2R
CC
Figure 15.3 Detection of different GPCR–GPCR heteromers and GPCR–RTK hetero-complexes in dorsal rat hippocampal and striatal sections by in situ PLA. (A-upper panel)Constitutive FGFR1–5-HT1A heteroreceptor complexes are detected by in situ PLA (redclusters) in dorsal rat hippocampus (Ammon's horn 1 and 3 (CA1, CA3) but not, as anexample, in the corpus callosum (cc). Scale bars, 20 mm. (A-lower panel) ConstitutiveFGFR1–M3R heteroreceptor complexes are detected by in situ PLA (red clusters) in dor-sal rat hippocampus (granular layer of the dentate gyrus [DG]) but not in the cortex cer-ebri. Scale bars from the left to the right, 50, 50, and 20 mm. (B-upper panel) ConstitutiveCCK2–D2R heteromers are detected by in situ PLA (red clusters) in striatal sections (cau-date putamen: CPu; amygdaloid cortex: AMG) but not, for example, in the cortex cerebri.Scale bars, 20 mm. (B-upper panel) Constitutive 5-HT2A–D2R heteromers are detected byin situ PLA (red clusters) in striatal sections (caudate putamen: CPu; amygdaloid cortex:AMG) but not, for example, in the corpus callosum (cc). Scale bars from the left to theright, 75, 50, and 20 mm. Nuclei appear as a blue color in all panels and the white arrowsindicate the red cluster formation (PLA signal).
292 Dasiel O. Borroto-Escuela et al.
Author's personal copy
changes in the total number of the two receptor populations. Of increasing
importance will be to determine the agonist/antagonist regulation of these
receptor heteromers in order to understand their potential roles as targets for
drugs used in neuropsychopharmacology for treatment of psychiatric and
neurological diseases. Analysis of human brain material with in situ PLA
can also reveal if the relative abundance of specific receptor heteromers in
discrete brain regions is altered in brain diseases (Fig. 15.3A and B).

293G Protein–Coupled Receptor Heterodimerization in the Brain
Author's personal copy
ACKNOWLEDGMENTSThis work has been supported by the Swedish Medical Research Council (04X-715) Torsten
andRagnar Soderberg Foundation (MN 65/08), TelethonTV3’s LaMarato Foundation 2008,
andM.M.Wallenberg Foundation to K. F., and Karolinska Institutets Forskningsstiftelser 2010
and 2011 toD.O. B.- E. TelethonTV3’s LaMarato Foundation 2008 to P.G. and K.F. Also by
grants SAF2008-01462 and Consolider-Ingenio CSD2008-00005 from Ministerio de Ciencia
e Innovacion to F. C. A. O. T. has not received any support for this work.
REFERENCESAgnati, L. F., Fuxe, K., Zini, I., Lenzi, P., & Hokfelt, T. (1980). Aspects on receptor reg-
ulation and isoreceptor identification. Medical Biology, 58, 182–187.Allalou, A., & Wahlby, C. (2009). BlobFinder, a tool for fluorescence microscopy image
cytometry. Computer Methods and Programs in Biomedicine, 94, 58–65.Borroto-Escuela, D. O., Garcia-Negredo, G., Garriga, P., Fuxe, K., & Ciruela, F. (2010).
The M(5) muscarinic acetylcholine receptor third intracellular loop regulates receptorfunction and oligomerization. Biochimica et Biophysica Acta, 1803, 813–825.
Borroto-Escuela, D. O., Romero-Fernandez, W., Mudo, G., Perez-Alea, M., Ciruela, F.,Tarakanov, A. O., et al. (2012). Fibroblast growth factor receptor 1–5-hydroxytryptamine1A heteroreceptor complexes and their enhancement of hippocampal plasticity. BiologicalPsychiatry, 71, 84–91.
Borroto-Escuela, D. O., Van Craenenbroeck, K., Romero-Fernandez, W., Guidolin, D.,Woods, A. S., Rivera, A., et al. (2011). Dopamine D2 and D4 receptor heteromerizationand its allosteric receptor-receptor interactions. Biochemical and Biophysical Research Com-munications, 404, 928–934.
Carpenter, A. E., Jones, T. R., Lamprecht, M. R., Clarke, C., Kang, I. H., Friman, O., et al.(2006). Cell Profiler: Image analysis software for identifying and quantifying cell pheno-types. Genome Biology, 7, R100.
Clausson, C. M., Allalou, A., Weibrecht, I., Mahmoudi, S., Farnebo, M., Landegren, U.,et al. (2011). Increasing the dynamic range of in situ PLA. Nature Methods, 8, 892–893.
Ferre, S., Baler, R., Bouvier, M., Caron, M. G., Devi, L. A., Durroux, T., et al. (2009).Building a new conceptual framework for receptor heteromers. Nature Chemical Biology,5, 131–134.
Fuxe, K., Agnati, L. F., Benfenati, F., Cimmino, M., Algeri, S., Hokfelt, T., et al. (1981).Modulation by cholecystokinins of 3H-spiroperidol binding in rat striatum: Evidencefor increased affinity and reduction in the number of binding sites. Acta PhysiologicaScandinavica, 113, 567–569.
Fuxe, K., Borroto-Escuela, D. O., Marcellino, D., Romero-Fernandez, W., Frankowska, M.,Guidolin, D., et al. (2012). GPCR heteromers and their allosteric receptor-receptor inter-actions. Current Medicinal Chemistry, 19, 356–363.
Leuchowius, K. J., Jarvius, M., Wickstrom, M., Rickardson, L., Landegren, U., Larsson, R.,et al. (2010). High content screening for inhibitors of protein interactions and post-translational modifications in primary cells by proximity ligation.Molecular & Cellular Pro-teomics, 9, 178–183.
Leuchowius, K. J., Weibrecht, I., & Soderberg, O. (2011). In situ proximity ligation assay formicroscopy and flow cytometry. Current Protocols in Cytometry, 36 chapter 9, unit 9.
Limbird, L. E., Meyts, P. D., & Lef kowitz, R. J. (1975). Beta-adrenergic receptors: Evidencefor negative cooperativity. Biochemical and Biophysical Research Communications, 64,1160–1168.

294 Dasiel O. Borroto-Escuela et al.
Author's personal copy
Nilsson, I., Bahram, F., Li, X., Gualandi, L., Koch, S., Jarvius, M., et al. (2010). VEGF re-ceptor 2/-3 heterodimers detected in situ by proximity ligation on angiogenic sprouts.The EMBO Journal, 29, 1377–1388.
Soderberg, O., Gullberg, M., Jarvius, M., Ridderstrale, K., Leuchowius, K. J., Jarvius, J.,et al. (2006). Direct observation of individual endogenous protein complexes in situby proximity ligation. Nature Methods, 3, 995–1000.
Soderberg, O., Leuchowius, K. J., Gullberg, M., Jarvius, M., Weibrecht, I., Larsson, L. G.,et al. (2008). Characterizing proteins and their interactions in cells and tissues using thein situ proximity ligation assay. Methods, 45, 227–232.
Soderberg, O., Leuchowius, K. J., Kamali-Moghaddam, M., Jarvius, M., Gustafsdottir, S.,Schallmeiner, E., et al. (2007). Proximity ligation: A specific and versatile tool for theproteomic era. Genetic Engineering, 28, 85–93.
Trifilieff, P., Rives, M. L., Urizar, E., Piskorowski, R. A., Vishwasrao, H. D., Castrillon, J.,et al. (2011). Detection of antigen interactions ex vivo by proximity ligation assay:Endogenous dopamine D2-adenosine A2A receptor complexes in the striatum.BioTechniques, 51, 111–118.
Related Documents











