Furcation, field-splitting, and the evolutionary origins of novelty in arthropod photoreceptors Todd H. Oakley * , David C. Plachetzki, Ajna S. Rivera Ecology Evolution and Marine Biology, University of CaliforniadSanta Barbara, Santa Barbara, CA 93106, USA Received 7 May 2007; accepted 28 August 2007 Abstract Arthropod photoreceptor evolution is a prime example of how evolution has used existing components in the origin of new structures. Here, we outline a comparative approach to understanding the mutational origins of novel structures, describing multiple examples from arthropod photoreceptor evolution. We suggest that developmental mechanisms have often split photoreceptors during evolution (field-splitting) and we introduce ‘‘co-duplication’’ as a null model for the mutational origins of photoreceptor components. Under co-duplication, gene duplication events coincide with the origin of a higher level structure like an eye. If co-duplication is rejected for a component, that component probably came to be used in a new photoreceptor through regulatory mutations. If not rejected, a gene duplication mutation may have allowed the com- ponent to be used in a new structure. In multiple case studies in arthropod photoreceptor evolution, we consistently reject the null hypothesis of co-duplication of genetic components and photoreceptors. Nevertheless, gene duplication events have in some cases occurred later, allowing divergence of photoreceptors. These studies provide a new perspective on the evolution of arthropod photoreceptors and provide a comparative approach that generalizes to the study of any evolutionary novelty. Ó 2007 Elsevier Ltd. All rights reserved. Keywords: Duplication; Co-option; Photoreceptor; Evolution; Eye; Bricolage 1. Introduction One of the most surprising generalities of the last quarter century of biological research is the extensive conservation of the molecular machinery of life. Biologists have learned that genes and often gene functions can be conserved for hundreds of millions of years of evolutionary time (e.g. McGinnis et al., 1984a,b; Halder et al., 1995). How evolution utilizes common building blocks to generate the riotous diversity of lifedsome- times termed ‘‘the novelty problem’’dhas thus become a cen- tral question in the field of evolutionary biology (Mu ¨ller and Wagner, 1991; West-Eberhard, 2003; Carroll et al., 2005). Few traits illustrate this tension between shared history and divergence better than arthropod eyes. Arthropods exhibit the widest diversity of eye designs in the animal kingdom. In addi- tion to pit and lens eye designs common to other animal groups, some arthropods possess exotic designs like refracting or re- flecting superposition compound eyes (Land and Nilsson, 2002). Yet coupled with this amazing display of diversity are uncanny similarities in genes, development, and morphology. For example, vertebrate Pax-6 genes can rescue function of mu- tations of the homologous arthropod gene eyeless (Halder et al., 1995). In addition, similar patterns of development are ob- served in the compound eyes of divergent taxa (e.g. Melzer et al., 2000). How evolution has used common building blocks to generate the diversity and complexity of arthropod eyes is the subject of this contribution. Two interdependent mutational mechanisms generally are implicated in the evolutionary origins of increased complexity or biodiversity. The first mechanism is gene duplication plus divergence (e.g. Ohno, 1970; Li, 1997; Taylor and Raes, 2004). Here, ancestral genes duplicate and one or both copies * Corresponding author. Fax: þ1 805 893 4724. E-mail address: [email protected] (T.H. Oakley). 1467-8039/$ - see front matter Ó 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.asd.2007.08.002 Arthropod Structure & Development 36 (2007) 386e400 www.elsevier.com/locate/asd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Arthropod Structure amp Development 36 (2007) 386e400wwwelseviercomlocateasd
Furcation field-splitting and the evolutionary origins ofnovelty in arthropod photoreceptors
Todd H Oakley David C Plachetzki Ajna S Rivera
Ecology Evolution and Marine Biology University of CaliforniadSanta Barbara Santa Barbara CA 93106 USA
Received 7 May 2007 accepted 28 August 2007
Abstract
Arthropod photoreceptor evolution is a prime example of how evolution has used existing components in the origin of new structures Herewe outline a comparative approach to understanding the mutational origins of novel structures describing multiple examples from arthropodphotoreceptor evolution We suggest that developmental mechanisms have often split photoreceptors during evolution (field-splitting) and weintroduce lsquolsquoco-duplicationrsquorsquo as a null model for the mutational origins of photoreceptor components Under co-duplication gene duplicationevents coincide with the origin of a higher level structure like an eye If co-duplication is rejected for a component that component probablycame to be used in a new photoreceptor through regulatory mutations If not rejected a gene duplication mutation may have allowed the com-ponent to be used in a new structure In multiple case studies in arthropod photoreceptor evolution we consistently reject the null hypothesis ofco-duplication of genetic components and photoreceptors Nevertheless gene duplication events have in some cases occurred later allowingdivergence of photoreceptors These studies provide a new perspective on the evolution of arthropod photoreceptors and provide a comparativeapproach that generalizes to the study of any evolutionary novelty 2007 Elsevier Ltd All rights reserved
Keywords Duplication Co-option Photoreceptor Evolution Eye Bricolage
1 Introduction
One of the most surprising generalities of the last quartercentury of biological research is the extensive conservation ofthe molecular machinery of life Biologists have learned thatgenes and often gene functions can be conserved for hundredsof millions of years of evolutionary time (eg McGinnis et al1984ab Halder et al 1995) How evolution utilizes commonbuilding blocks to generate the riotous diversity of lifedsome-times termed lsquolsquothe novelty problemrsquorsquodhas thus become a cen-tral question in the field of evolutionary biology (Muller andWagner 1991 West-Eberhard 2003 Carroll et al 2005)Few traits illustrate this tension between shared history anddivergence better than arthropod eyes Arthropods exhibit the
Corresponding author Fax thorn1 805 893 4724
E-mail address oakleylifesciucsbedu (TH Oakley)
1467-8039$ - see front matter 2007 Elsevier Ltd All rights reserved
doi101016jasd200708002
widest diversity of eye designs in the animal kingdom In addi-tion to pit and lens eye designs common to other animal groupssome arthropods possess exotic designs like refracting or re-flecting superposition compound eyes (Land and Nilsson2002) Yet coupled with this amazing display of diversity areuncanny similarities in genes development and morphologyFor example vertebrate Pax-6 genes can rescue function of mu-tations of the homologous arthropod gene eyeless (Halder et al1995) In addition similar patterns of development are ob-served in the compound eyes of divergent taxa (eg Melzeret al 2000) How evolution has used common building blocksto generate the diversity and complexity of arthropod eyes is thesubject of this contribution
Two interdependent mutational mechanisms generally areimplicated in the evolutionary origins of increased complexityor biodiversity The first mechanism is gene duplication plusdivergence (eg Ohno 1970 Li 1997 Taylor and Raes2004) Here ancestral genes duplicate and one or both copies
387TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
subsequently diverge in a descendant Whereas the ancestralgenome had a single gene copy the descendant has two differ-ent paralogs that may diverge to increase genomic complexity(eg Hughes 1994) A second genetic mechanism is regula-tory mutation where ancestral genes are used in new combi-nations in a descendant by altering the regulation of thosegenes (eg Britten and Davidson 1969 Wray et al 2003Wray 2007) In order to increase complexity through regula-tory mutation descendants must use an ancestral gene innew spatial or temporal contexts that generate novel interac-tions thereby increasing the total diversity of interactions Be-fore discussing how these processes might have contributed toarthropod visual sense organ evolution we will first introducea phylogenetic perspective on identifying these processesfocusing on how these mutations affect different levels ofbiological organization
11 lsquolsquoTree thinkingrsquorsquo and evolutionary novelty
Here we present the beginnings of a conceptual frameworkfor understanding the evolutionary origins of complexity andbiodiversity that is rooted in phylogenetic or lsquolsquotree thinkingrsquorsquoapproaches Phylogenetic methodology reconstructs patternsof common ancestry by cladistic analyses of components ofhigher level units For example phylogenies of species are es-timated by numerical analyses of components of those speciessuch as morphological traits or DNA sequences Similarlyphylogenies of genes (gene trees) are estimated by analyzingtheir components namely nucleotides or amino acids In addi-tion to the familiar patterns of common ancestry of species andgenes we are exploring phylogenetic approaches to under-standing the common ancestry of traits at other levels of bio-logical organization such as gene networks cell types andorgans (Oakley 2003a Plachetzki et al 2005 Serb andOakley 2005 Plachetzki and Oakley 2007)
Phylogenetic trees represent patterns of common ancestryLineages at one level of organization are composed of lower-level lineages For example species lineages are composed ofgene lineages and gene lineages are composed of protein do-main lineages (Maddison 1997) Phylogenetic patterns resultfrom evolutionary processes that split lineages usually by fis-sion or duplication Species originate in speciation events andgenes originate in duplication events Additionally analogousprocesses occur at other levels of biological organization theseprocesses will be discussed herein To refer generally to pro-cesses of lineage splitting we introduce the term lsquolsquofurcationrsquorsquodefined as the lsquolsquoformation of a fork or division into branchesrsquorsquo
A new term is preferred because no single established term isquite general enoughdduplication implies that the entities areidentical at conception which is not always the case lsquolsquoBifurca-tionrsquorsquo is not a precise term either because it precludes simulta-neous splitting of lineages into more than two descendants andit also has a different meaning in mathematics The conceptuallysimilar term lsquolsquocladogenesisrsquorsquo historically refers primarily tospecies lineages Unlike these established terms lsquolsquofurcationrsquorsquois meant to generalize to any level of organization especiallyto include processes that split cell type gene network organ
and tissue lineages during evolution Furcations must arise asmutations leading to variation in the number of biological unitswithin a species which may or may not be fixed by populationlevel processes Once fixed furcated biological units may re-main the same diverge or be lost The concept of furcation al-lows lsquolsquotree thinkingrsquorsquo (OrsquoHara 1997) and the applied statisticsof phylogenetic methodology to extend beyond species- andgene-level perspectives to all levels of biology because any bi-ological units that undergo furcation will exhibit nested patternsof common ancestry Just as a focus on speciation allowed newinsights into the origin of species (Dobzhansky 1937 Mayr1942) we propose that a focus on furcation will lead to new in-sights into the origin of novel traits during evolution
Here we present a phylogenetic perspective on the historyof arthropod photoreceptive structures We suggest that devel-opmental processes namely regulatory genetic mutations mayoften have split photoreceptive structures during arthropodevolution allowing for the origins of lsquolsquonewrsquorsquo receptors thatlater diverged from each other in structure andor functionWe find no evidence of gene duplication mutations contribut-ing directly to the origins of novel arthropod photoreceptivestructures However in some cases the process of gene dupli-cation plus divergence has contributed to the differentiation ofstructures that probably originally furcated by regulatory mu-tations perhaps through splitting developmental fields
12 Comparative approaches
Throughout this article we will use comparative ap-proaches to understand the history of arthropod photoreceptivestructures and their components often using three specificsteps We first identify a trait of interest and address the ques-tion of when that trait originated This first step involves typ-ical methods of mapping the trait of interest on a phylogenetictree When sister species share a trait their common ancestorlikely had that trait a logic that is formalized for more com-plex histories through parsimony and statistical model-basedcharacter analyses (Swofford and Maddison 1987 Pagel1994 Cunningham et al 1998 Oakley 2003b Ronquist2004) In the current context outgroup analysis can identifyputative cases where developmental processes split photore-ceptive structures in ancestral species ingroup species willhave multiple structures that develop from a single field andoutgroups will possess a single receptor that develops froma homologous field By scoring complex traits as binary states(present or absent) such phylogenetic techniques make thesimplifying assumption that all components of that trait simul-taneously are gained or lost Here we argue that this simplify-ing discretizing approach may be considered a null hypothesisfor the mutations involved in the origins of novelty which wecall lsquolsquoco-duplicationrsquorsquo Under the co-duplication null hypothe-sis the genetic components of a trait were all duplicated si-multaneously along with the furcation of the higher-leveltrait (arthropod photoreceptive structures in the present case)
After trait mapping our second step is to test the null hypoth-esis by examining when the genetic components of the furcatedstructures were last duplicated Any genetic components that
388 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
are shared between the furcated photoreceptive structures rejectthe null-hypothesis of co-duplication Shared components musthave become part of new photoreceptive structures through reg-ulatory mutations Namely components of a single ancestral re-ceptor became components of multiple furcated receptors ina descendant Therefore the genetic components gained usagein a new context and were not duplicated Although in somecases components may be shared between furcated photorecep-tive structures other components may be unique to one of thefurcated structures In these cases we take a third step to datethe origin of the components and test the co-duplication null hy-pothesis We provide examples below describing how the originof components can be dated using phylogenetic techniques likereconciled tree analysis and relaxed molecular clock dating ofgene duplication events Any genetic components that do not re-ject the co-duplication hypothesis might have contributed tonovelty through gene duplication Below we will first sketchsome developmental mechanisms that split photoreceptor fieldsspatiotemporally to allow for the origins of novel features Wethen discuss multiple case studies in arthropod photoreceptorevolution that reject co-duplication in favor of regulatory muta-tions This leads us to hypothesize that furcation of photorecep-tors by regulatory mutations not including gene duplicationmutations has been a major engine in the origins of arthropodphotoreceptor diversity
2 Mechanisms of field splitting and furcation inspace and time
Although studied in a few cases like segmentation and ap-pendage formation (Panganiban et al 1997 Seaver 2003) themechanisms of trait furcation are understudied and poorly un-derstood compared to furcation of species (speciation) andgenes (gene duplication) Yet a more comprehensive under-standing of these mechanisms would lead to insight on the or-igins of novelty during evolution because like the duplicationof a gene furcation of any biological unit with modifiablecomponents (eg photoreceptors) can lead to evolutionary di-versity when one unit gains a new function (neofunctionaliza-tion) or each retains a different partition of ancestral functions(subfunctionalization as defined by Force et al 1999) Whilethe mechanisms of gene duplication involve genetic muta-tional events and the mechanisms of speciation involve geo-graphic and population-level processes the mechanisms offurcation at the level of traits such as photoreceptors involvedevelopmental processes Next we discuss some specific ex-amples of how developmental processes may have been in-volved in furcation of photoreceptors followed by spatialandor temporal divergence of the paralogous traits These pro-cesses highlight ways that regulatory mutations may havecaused furcation of photoreceptors without the co-duplicationof associated genetic components
21 Furcation and spatial divergence
A simple example of gene regulatory changes causing furca-tion is the serial repetition of an existing structure sometimes
termed heterotopy (Haeckel 1866 Zelditch and Fink 1996West-Eberhard 2003) One example is the addition of omma-tidia to a compound eye which does not necessarily requirenew genes or cell-types to be invented instead a reuse of exist-ing genetic networks allows repetition (furcation) of a structureand its components While this simple case does not necessarilylead to a more complex eye larger numbers of ommatidia maylead to increased visual ability for example by decreasing theinter-ommatidial angle to raise visual acuity (Hecht and Wolf1929) or by increasing the region of space the eye can observeat one time One specific developmental mechanism of increas-ing ommatidial number could be the expansion of the develop-mental field by either a permissive signal (eg decapentaplegic)or the suppression of a restrictive signal (eg wingless) Thesevariationsdwhich could also lead to loss of ommatidiadarisewithin species leading to polymorphisms that may or may notbecome fixed by population level processes like selection ordrift By these mechanisms the alteration of ommatidial num-ber could originate without adding duplicated genes (eg opsin)or novel cell types In our terminology increasing ommatidialnumber would represent a furcation event The observation ofshared components of the new ommatidia would reject a nullhypothesis of co-duplication of those components and theommatidia thus highlighting the role of gene regulatory muta-tions as opposed to gene duplications in the origin of novelommatidia Even after originating by regulatory mutationsduplicated genes could still become expressed in furcatedommatidia later
22 Furcation and temporal divergence
Once furcated the paralogous structures may diverge in var-ious ways such as in their timing of differentiation which maybe considered heterochrony A focus on furcation raises an in-teresting new perspective on heterochrony because we areviewing the traits themselves as comprising evolutionary line-ages Heterochrony then becomes a comparison of ancestraland descendent developmental trajectories in the context ofthe traits rather than the more standard context of the organismIn other words heterochrony can involve a comparison betweenancestral and descendent traits as well as ancestral and descen-dent species This idea is further explored after describingchanges in timing in the Drosophila visual system develop-ment which we will argue may be considered heterochrony
The Drosophila visual system is comprised of three devel-opmentally related unitsdthe ocelli paired Bolwigrsquos organs(larval stemmata) and compound eyes (adult eyes) Here wefocus on the larval stemmata and adult eyes About a thirdof the way through embryogenesis the visual anlage un-dergoes a series of divisions (field splitting) to form laterallysymmetric precursors of the three eye types (Daniel et al1999) The larval eye is innervated and expresses opsin proteinby the end of embryogenesis (Pollock and Benzer 1988Meinertzhagen and Hanson 1993) but later undergoes furtherdifferentiation to become the adult extra-retinal eyelet(Helfrich-Forster et al 2002) In contrast the adult compoundeyes do not complete development until the final larval stage
389TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Instead they remain as undifferentiated tissue throughout em-bryonic and most of larval development (Wolff and Ready1993)
We submit that aspects of fly photoreceptor organ evolutionmay be viewed as heterochrony when focusing on the recep-tors themselves as a phylogenetic lineage The ancestor toboth fly Bolwigrsquos organs and compound eyes is a single con-tinuous developmental program where adult eyes developfrom juvenile eyes Comparisons of a hypothetical ancestorand the two paralogous descendants reveal changes in timingof differentiation (Fig 1) For example relative to an ancestralstate the fly eyelet (the adult form of Bolwigrsquos organ) mightbe considered neotonous In addition the trajectory of flycompound eye development contains a period of slow or tem-porarily arrested development relative to the ancestral stateWe note that these changes may not be called heterochronyunder the mathematical definition of Rice (1997) which re-quires uniform rate changes in developmental programsWhether or not we use the term heterochrony paralogous flyvisual organs provide an example where furcated structureshave diverged in the timing of their differentiation relative toeach other and their ancestral developmental program Belowwe apply the phylogenetic approaches mentioned above togain greater understanding of the evolutionary origins of thisdual receptor system
221 When did the novelty originateThe evolution and development of insect eyes is fairly well
understood Structural and developmental studies comparingthe larval and adult eyes of Drosophila with those of other in-sects including outgroup species where the larval eye givesrise to the adult eye have shown that compound eyes andstemmata are paralogous in insects and likely emerged from
Eye
Diff
eren
tiatio
n(n
ot to
sca
le)
Juvenile Adult
Developm(not to
A
Fig 1 Comparison of timing of development for insect photoreceptors The Y-axi
specifically defined) The X-axis represents developmental time (units not specifica
eyes develop directly from juvenile eyes In many cases eye differentiation ceases i
Circle represents developmental field splitting Dashed line is compound eye onto
togeny both stemmata and compound eyes underwent changes in timing which m
the furcation of a single ancestral eye field (Friedrich2006a) This ancestral field first developed into an embryoniceye which persisted in development through larval stages tobecome the adult compound eye as is the case in ametabolousand hemimetabolous insects (Meinertzhagen 1973 Bate1978) In holometabolous insects larval and adult eye devel-opment is decoupleddthese two eye types develop from sep-arate primordia each derived from the same embryonic anlage(Paulus 1979 Green et al 1993 Friedrich and Benzer 2000Liu and Friedrich 2004) Thus the evolutionary furcation ofthe eye field producing both larval eyes and adult compoundeyes likely occurred within the same phylogenetic interval asthe split between hemi- and holometabolous insects (Fig 2)
222 What are the differences in components of adultand larval eyes
The differences in larval and adult fly eyes extend to theirstructure and function as well as their developmental originThe adult eye is the familiar compound eye a large complexstructure made of hundreds of ommatidia and thousands ofcells including specialized cell types called retinular (R-cells)cone cells and pigment cells (Paulus 1979) It is an image form-ing eye and has a large associated nerve network The larval eyeby comparison is quite simple It is made of few photoreceptorsand has roles in circadian rhythm entrainment and in mediat-ing phototaxis in foraging and pupating larvae (Kaneko et al1997 Helfrich-Forster et al 2002)
Interestingly in this furcation of eyes and the subsequentadoption of disparate functions there is evidence of regulatorygenetic mutations but not of gene duplication mutations Forexample Drosophila adult and larval eyes both express someof the same opsins (Malpel et al 2002) In addition many ofthe same developmental genes are expressed in both larval
ental Time scale)
Adult
CompoundEye
Stemmataembr
yoge
nesi
s
met
amor
phos
is
Eyelet
larv
al B
s represents a heuristic sketch of the amount of differentiation (with units not
lly defined) (A) The ontogeny of eyes in a hypothetical ancestral insect Adult
n adults (not shown) (B) Ontogeny of fly photoreceptors (see text for details)
geny solid line is stemmataeyelet ontogeny Relative to the ancestral eye on-
ay be considered heterochrony but not by all definitions (eg Rice 1997)
Ametabola Hemimetabola Holometabola
Reducedlarval eyes
Separate larvaland adult eyes
Fig 2 Phylogeny of the insects based on Giribet et al (2001) In the ancestral
single eye field adult eyes arose from larval eyes This can be seen in hemi-
metabolous and ametabolous insects as well as in some crustaceans In the lin-
eage leading to the Holometabola this field furcated to form adult and larval
eyes with paralogous tissue origins (gray circle) Later within the Holometa-
bola the larval eyes were reduced to produce novel photosensory organs
(black circle) These are exemplified by Bolwigrsquos organ in Drosophila
toy
toyey
otd
eya
so
en
en
zen
ato
gl
Kr
Mu
hh
dpp
wg
N
Egfr
Spi
vn
Compound Eye Bolwigrsquos Organ Ocellus
Fig 3 Evolutionary history of Drosophila photoreceptor organs compiled by
Friedrich (2006b) Photoreceptors lsquolsquofurcatersquorsquo (see text for definition) during
evolution leading to patterns of common ancestry that can be conceived as
phylogenetic trees (see also Geeta 2003 Serb and Oakley 2005) Bolwigrsquos
organ and compound eyes likely share a more recent common ancestor than
either does with ocelli (reviewed in Friedrich 2006a) Assuming those rela-
tionships we mapped presence or absence of expression of different genes
on the photoreceptor phylogeny using parsimony as implemented in Mesquite
(Maddison and Maddison 2004) Black ovals represent gain of gene expres-
sion white ovals represent loss of gene expression Gene abbreviations ey frac14eyeless Kr frac14 Kruppel zen frac14 zerknullt en frac14 engrailed toy frac14 twin-of-eyeless
vn frac14 vein Spi frac14 spitz Egfr frac14 Epidermal growth factor receptor N frac14 Notch
dpp frac14 decapentaplegic hh frac14 hedgehog Mu frac14 Munster gl frac14 glass ato frac14atonal so frac14 sine oculis otd frac14 orthodenticle eya frac14 eyes absent
390 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
and adult eyes Hedgehog and decapentaplegic specify the eyeprimordium and initiate eye development in both eye types (He-berlein et al 1993 Suzuki and Saigo 2000 Chang et al 2001)This is followed by expression of many of the same lsquolsquoretinal de-termination network genesrsquorsquo (reviewed in Friedrich 2006a) Infact 13 genes are known to be expressed in all three fly photo-receptor organs which may be taken as evidence for their com-mon ancestry (Fig 3) In addition to many similaritiesdifferences in gene expression also exist between fly compoundeyes and Bolwigrsquos organs (Friedrich 2006a) Kruppel and zenare involved in Bolwigrsquos organ development but not in that ofcompound eyes or ocelli Furthermore engrailed and toy expres-sion is absent in Bolwigrsquos organ but present in the other two vi-sual structures although engrailed has no detectable function inthe adult eye (Strutt and Mlodzik 1996) Five additional genesare known to differ in presenceabsence of expression betweencompound eye and Bolwigrsquos organ (reviewed in Friedrich2006a) Because no evidence supports the hypothesis that theseexpression differences owe their origins to gene duplicationevents concomitant with the origin of Bolwigrsquos organ a co-duplication hypothesis is rejected for all known components
223 SummaryAlthough future work on the mechanisms responsible for
the initial field splitting of fly visual organs could implicatethe involvement of duplicated genes available evidence indi-cates that the origin of separate larval and adult photoreceptororgans in insects was due primarily to regulatory genetic mu-tations that altered the timing of expression of existing geneswhereby photoreceptive organ differentiation started to occurat two different times during ontogeny Despite the fact thatgene duplication mutations can be involved in the origin ofnew photoreceptive structures as evidenced by vertebraterods and cones (Nordstrom et al 2004 Serb and Oakley2005 Plachetzki and Oakley 2007) the origin of distinct flyphotoreceptive organs produces no such evidence leading usto reject the null hypothesis of co-duplication As such fly
photoreceptive organs provide an example of how a tissue fur-cation can result in structures expressed at different times aswell as in different places Although we know that differentgene expression patterns are present in the two visual struc-tures we do not know which specific regulatory mutationscause the furcation of these organs Identifying such mutationswould be a major contribution to understanding this system
3 Additional case studies in furcation
Although we have the most genetic information availableregarding the origin of insect stemmata described abovea number of additional case studies suggest similar patterns
391TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
for the origins of other arthropod photoreceptive structures Inall cases we find no evidence for the involvement of geneduplication mutations in the origins of the photoreceptorsNevertheless in three casesdostracod median and compoundeyes insect laminar and medullar-projecting photoreceptorsand Limulus median ocelli and compound eyesdsomeevidence suggests that gene duplication mutations weresubsequently involved in the differentiation of furcated eyetypes
31 Ostracod compound eyes
311 When did the novelty originateThe origin of ostracod compound eyes provides another ex-
ample of how co-duplication can be used as a phylogeneticnull model for the origins of evolutionary complexity Molec-ular phylogenies of Ostracoda indicate that myodocopidsdtheonly Ostracoda with compound eyesdare a monophyleticgroup whose closest relatives lack compound eyes (Oakleyand Cunningham 2002 Yamaguchi and Endo 2003 Oakley2005) Although support for myodocopid monophyly is verystrong outstanding issues in ostracod phylogeny include thepossibilities that ostracods are not monophyletic (Wakayamain press) and near outgroupsdoften called lsquolsquoMaxillopodarsquorsquodarenot monophyletic (Regier et al 2005) such that understand-ing ostracod phylogeny requires understanding phylogeny ofearly branching crustaceans Crustacean and arthropod phy-logeny are notoriously difficult with different data often yield-ing different results (eg Boore et al 1995 Friedrich andTautz 1995 Regier and Shultz 1997 Giribet and Ribera2000 Giribet et al 2001 Regier et al 2005) What currentlyis clear for ostracods is that myodocopids with compound eyesare monophyletic (including Silurian specimens (Siveter et al2003 2007)) and they are closely related to some groups thatpossess lateral eyes (some juvenile thecostracans and bran-chiurans) and multiple groups that lack compound lateral
m Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
c
c
m
m
m
x
m
c
m
m
A
m
Stemc
m m
m
c
m
B
Fig 4 Inferred ancestral states null hypothesis and inferred history of myodocopid
pound and median eyes of myodocopid ostracods and relatives A species phylogen
enclosing lsquolsquomrsquorsquo represent median eyes circles enclosing lsquolsquocrsquorsquo represent compound ey
with a circle enclosing an lsquolsquoxrsquorsquo Ancestral myodocopids likely had compound and m
median eyes (Oakley and Cunningham 2002 Oakley 2005) (B) A graphical illustr
pound eyes Illustrated is a phylogenetic tree of species that includes one living and
gene duplication events are predicted to coincide in time with the origin of compoun
such as opsin (blackwhite phylogeny inside grey species phylogeny) show a speci
ralogs one expressed in compound eyes the other in median eyes Halocyprida m
duplication could hold for any component of myodocopid compound eyes rejecting
of the trait (C) Based on opsin molecular clock analyses (Fig 5) a gene duplicatio
the origin of myodocopids and their compound eyes
eyes (copepods mystacocarids many thecostracans penta-stomes and tantulocarids)
Assuming the currently available molecular phylogenetichypotheses for Ostracoda (Oakley and Cunningham 2002 Ya-maguchi and Endo 2003 Oakley 2005) statistical consider-ations indicate that compound eyes were more likely absentin the ancestor of ostracods than the alternative hypothesisthat compound eyes were lost numerous times in relatives ofnon-myodocopids However there are two important consider-ations First inclusion of additional outgroups could changethe phylogeny andor statistics but since numerous additionaloutgroups lack lateral compound eyes including more out-groups is most likely to increase support for ancestral absenceof lateral eyes Second an unambiguous discrimination of thealternatives depends on the unknown relative rate of gain ver-sus loss of lateral eyes during macroevolution which can onlybe estimated (Oakley and Cunningham 2002 Oakley 2005)
With these considerations in mind available analyses donot support lsquolsquophylogenetic homologyrsquorsquo of ostracod compoundeyes with compound eyes of other arthropods Phylogenetichomology is an explicit hypothesis about the presence of a par-ticular trait (in this case a compound eye) in a common ances-tor (Butler and Saidel 2000) Current phylogenies suggest thatcompound eyes were absent in the common ancestor of all os-tracods and later originated at or somewhat before the ances-tor of myodocopids (Fig 4A) Compound eyes could havebeen present in an earlier arthropod ancestor and regained inmyodocopids perhaps indicating lsquolsquogenerative homologyrsquorsquowhere the same developmental program allows for multiplephylogenetic origins of compound eyes (Meyer 1999 Butlerand Saidel 2000) The process of re-evolving generatively ho-mologous structures has been termed lsquolsquoswitchback evolutionrsquorsquo(Van Valen 1979) and the multifunctionality of most genes(Piatigorsky 2007) suggests genes may often be retainedeven after the loss of a particular structure (Marshall et al1994)
Halocyprida
Myodocopida
Copepoda
Myodocopida
c
m
m
x Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
x
m
C
m
m
mc
m
c
c
m
eyes and components (A) Summary of ancestral state reconstruction of com-
y in grey shows relationships of myodocopid Ostracoda and relatives Circles
es Halocyprida are ostracods that lack compound and median eyes illustrated
edian eyes whereas the ancestor of Copepoda thornMyodocopa likely had only
ation of a lsquolsquoco-duplicationrsquorsquo null hypothesis for the origin of myodocopid com-
one stem group myodocopid species and two outgroups Under co-duplication
d eyes at or before the origin of myodocopids Under such a model gene trees
fic pattern where myodocopids possess duplicated and monophyletic opsin pa-
ay or may not have lost opsins illustrated with lsquolsquorsquorsquo A null hypothesis of co-
that null indicates that regulatory genetic mutations were involved in the origin
n event (white circle) led to compound and median eye-specific paralogs after
392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

387TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
subsequently diverge in a descendant Whereas the ancestralgenome had a single gene copy the descendant has two differ-ent paralogs that may diverge to increase genomic complexity(eg Hughes 1994) A second genetic mechanism is regula-tory mutation where ancestral genes are used in new combi-nations in a descendant by altering the regulation of thosegenes (eg Britten and Davidson 1969 Wray et al 2003Wray 2007) In order to increase complexity through regula-tory mutation descendants must use an ancestral gene innew spatial or temporal contexts that generate novel interac-tions thereby increasing the total diversity of interactions Be-fore discussing how these processes might have contributed toarthropod visual sense organ evolution we will first introducea phylogenetic perspective on identifying these processesfocusing on how these mutations affect different levels ofbiological organization
11 lsquolsquoTree thinkingrsquorsquo and evolutionary novelty
Here we present the beginnings of a conceptual frameworkfor understanding the evolutionary origins of complexity andbiodiversity that is rooted in phylogenetic or lsquolsquotree thinkingrsquorsquoapproaches Phylogenetic methodology reconstructs patternsof common ancestry by cladistic analyses of components ofhigher level units For example phylogenies of species are es-timated by numerical analyses of components of those speciessuch as morphological traits or DNA sequences Similarlyphylogenies of genes (gene trees) are estimated by analyzingtheir components namely nucleotides or amino acids In addi-tion to the familiar patterns of common ancestry of species andgenes we are exploring phylogenetic approaches to under-standing the common ancestry of traits at other levels of bio-logical organization such as gene networks cell types andorgans (Oakley 2003a Plachetzki et al 2005 Serb andOakley 2005 Plachetzki and Oakley 2007)
Phylogenetic trees represent patterns of common ancestryLineages at one level of organization are composed of lower-level lineages For example species lineages are composed ofgene lineages and gene lineages are composed of protein do-main lineages (Maddison 1997) Phylogenetic patterns resultfrom evolutionary processes that split lineages usually by fis-sion or duplication Species originate in speciation events andgenes originate in duplication events Additionally analogousprocesses occur at other levels of biological organization theseprocesses will be discussed herein To refer generally to pro-cesses of lineage splitting we introduce the term lsquolsquofurcationrsquorsquodefined as the lsquolsquoformation of a fork or division into branchesrsquorsquo
A new term is preferred because no single established term isquite general enoughdduplication implies that the entities areidentical at conception which is not always the case lsquolsquoBifurca-tionrsquorsquo is not a precise term either because it precludes simulta-neous splitting of lineages into more than two descendants andit also has a different meaning in mathematics The conceptuallysimilar term lsquolsquocladogenesisrsquorsquo historically refers primarily tospecies lineages Unlike these established terms lsquolsquofurcationrsquorsquois meant to generalize to any level of organization especiallyto include processes that split cell type gene network organ
and tissue lineages during evolution Furcations must arise asmutations leading to variation in the number of biological unitswithin a species which may or may not be fixed by populationlevel processes Once fixed furcated biological units may re-main the same diverge or be lost The concept of furcation al-lows lsquolsquotree thinkingrsquorsquo (OrsquoHara 1997) and the applied statisticsof phylogenetic methodology to extend beyond species- andgene-level perspectives to all levels of biology because any bi-ological units that undergo furcation will exhibit nested patternsof common ancestry Just as a focus on speciation allowed newinsights into the origin of species (Dobzhansky 1937 Mayr1942) we propose that a focus on furcation will lead to new in-sights into the origin of novel traits during evolution
Here we present a phylogenetic perspective on the historyof arthropod photoreceptive structures We suggest that devel-opmental processes namely regulatory genetic mutations mayoften have split photoreceptive structures during arthropodevolution allowing for the origins of lsquolsquonewrsquorsquo receptors thatlater diverged from each other in structure andor functionWe find no evidence of gene duplication mutations contribut-ing directly to the origins of novel arthropod photoreceptivestructures However in some cases the process of gene dupli-cation plus divergence has contributed to the differentiation ofstructures that probably originally furcated by regulatory mu-tations perhaps through splitting developmental fields
12 Comparative approaches
Throughout this article we will use comparative ap-proaches to understand the history of arthropod photoreceptivestructures and their components often using three specificsteps We first identify a trait of interest and address the ques-tion of when that trait originated This first step involves typ-ical methods of mapping the trait of interest on a phylogenetictree When sister species share a trait their common ancestorlikely had that trait a logic that is formalized for more com-plex histories through parsimony and statistical model-basedcharacter analyses (Swofford and Maddison 1987 Pagel1994 Cunningham et al 1998 Oakley 2003b Ronquist2004) In the current context outgroup analysis can identifyputative cases where developmental processes split photore-ceptive structures in ancestral species ingroup species willhave multiple structures that develop from a single field andoutgroups will possess a single receptor that develops froma homologous field By scoring complex traits as binary states(present or absent) such phylogenetic techniques make thesimplifying assumption that all components of that trait simul-taneously are gained or lost Here we argue that this simplify-ing discretizing approach may be considered a null hypothesisfor the mutations involved in the origins of novelty which wecall lsquolsquoco-duplicationrsquorsquo Under the co-duplication null hypothe-sis the genetic components of a trait were all duplicated si-multaneously along with the furcation of the higher-leveltrait (arthropod photoreceptive structures in the present case)
After trait mapping our second step is to test the null hypoth-esis by examining when the genetic components of the furcatedstructures were last duplicated Any genetic components that
388 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
are shared between the furcated photoreceptive structures rejectthe null-hypothesis of co-duplication Shared components musthave become part of new photoreceptive structures through reg-ulatory mutations Namely components of a single ancestral re-ceptor became components of multiple furcated receptors ina descendant Therefore the genetic components gained usagein a new context and were not duplicated Although in somecases components may be shared between furcated photorecep-tive structures other components may be unique to one of thefurcated structures In these cases we take a third step to datethe origin of the components and test the co-duplication null hy-pothesis We provide examples below describing how the originof components can be dated using phylogenetic techniques likereconciled tree analysis and relaxed molecular clock dating ofgene duplication events Any genetic components that do not re-ject the co-duplication hypothesis might have contributed tonovelty through gene duplication Below we will first sketchsome developmental mechanisms that split photoreceptor fieldsspatiotemporally to allow for the origins of novel features Wethen discuss multiple case studies in arthropod photoreceptorevolution that reject co-duplication in favor of regulatory muta-tions This leads us to hypothesize that furcation of photorecep-tors by regulatory mutations not including gene duplicationmutations has been a major engine in the origins of arthropodphotoreceptor diversity
2 Mechanisms of field splitting and furcation inspace and time
Although studied in a few cases like segmentation and ap-pendage formation (Panganiban et al 1997 Seaver 2003) themechanisms of trait furcation are understudied and poorly un-derstood compared to furcation of species (speciation) andgenes (gene duplication) Yet a more comprehensive under-standing of these mechanisms would lead to insight on the or-igins of novelty during evolution because like the duplicationof a gene furcation of any biological unit with modifiablecomponents (eg photoreceptors) can lead to evolutionary di-versity when one unit gains a new function (neofunctionaliza-tion) or each retains a different partition of ancestral functions(subfunctionalization as defined by Force et al 1999) Whilethe mechanisms of gene duplication involve genetic muta-tional events and the mechanisms of speciation involve geo-graphic and population-level processes the mechanisms offurcation at the level of traits such as photoreceptors involvedevelopmental processes Next we discuss some specific ex-amples of how developmental processes may have been in-volved in furcation of photoreceptors followed by spatialandor temporal divergence of the paralogous traits These pro-cesses highlight ways that regulatory mutations may havecaused furcation of photoreceptors without the co-duplicationof associated genetic components
21 Furcation and spatial divergence
A simple example of gene regulatory changes causing furca-tion is the serial repetition of an existing structure sometimes
termed heterotopy (Haeckel 1866 Zelditch and Fink 1996West-Eberhard 2003) One example is the addition of omma-tidia to a compound eye which does not necessarily requirenew genes or cell-types to be invented instead a reuse of exist-ing genetic networks allows repetition (furcation) of a structureand its components While this simple case does not necessarilylead to a more complex eye larger numbers of ommatidia maylead to increased visual ability for example by decreasing theinter-ommatidial angle to raise visual acuity (Hecht and Wolf1929) or by increasing the region of space the eye can observeat one time One specific developmental mechanism of increas-ing ommatidial number could be the expansion of the develop-mental field by either a permissive signal (eg decapentaplegic)or the suppression of a restrictive signal (eg wingless) Thesevariationsdwhich could also lead to loss of ommatidiadarisewithin species leading to polymorphisms that may or may notbecome fixed by population level processes like selection ordrift By these mechanisms the alteration of ommatidial num-ber could originate without adding duplicated genes (eg opsin)or novel cell types In our terminology increasing ommatidialnumber would represent a furcation event The observation ofshared components of the new ommatidia would reject a nullhypothesis of co-duplication of those components and theommatidia thus highlighting the role of gene regulatory muta-tions as opposed to gene duplications in the origin of novelommatidia Even after originating by regulatory mutationsduplicated genes could still become expressed in furcatedommatidia later
22 Furcation and temporal divergence
Once furcated the paralogous structures may diverge in var-ious ways such as in their timing of differentiation which maybe considered heterochrony A focus on furcation raises an in-teresting new perspective on heterochrony because we areviewing the traits themselves as comprising evolutionary line-ages Heterochrony then becomes a comparison of ancestraland descendent developmental trajectories in the context ofthe traits rather than the more standard context of the organismIn other words heterochrony can involve a comparison betweenancestral and descendent traits as well as ancestral and descen-dent species This idea is further explored after describingchanges in timing in the Drosophila visual system develop-ment which we will argue may be considered heterochrony
The Drosophila visual system is comprised of three devel-opmentally related unitsdthe ocelli paired Bolwigrsquos organs(larval stemmata) and compound eyes (adult eyes) Here wefocus on the larval stemmata and adult eyes About a thirdof the way through embryogenesis the visual anlage un-dergoes a series of divisions (field splitting) to form laterallysymmetric precursors of the three eye types (Daniel et al1999) The larval eye is innervated and expresses opsin proteinby the end of embryogenesis (Pollock and Benzer 1988Meinertzhagen and Hanson 1993) but later undergoes furtherdifferentiation to become the adult extra-retinal eyelet(Helfrich-Forster et al 2002) In contrast the adult compoundeyes do not complete development until the final larval stage
389TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Instead they remain as undifferentiated tissue throughout em-bryonic and most of larval development (Wolff and Ready1993)
We submit that aspects of fly photoreceptor organ evolutionmay be viewed as heterochrony when focusing on the recep-tors themselves as a phylogenetic lineage The ancestor toboth fly Bolwigrsquos organs and compound eyes is a single con-tinuous developmental program where adult eyes developfrom juvenile eyes Comparisons of a hypothetical ancestorand the two paralogous descendants reveal changes in timingof differentiation (Fig 1) For example relative to an ancestralstate the fly eyelet (the adult form of Bolwigrsquos organ) mightbe considered neotonous In addition the trajectory of flycompound eye development contains a period of slow or tem-porarily arrested development relative to the ancestral stateWe note that these changes may not be called heterochronyunder the mathematical definition of Rice (1997) which re-quires uniform rate changes in developmental programsWhether or not we use the term heterochrony paralogous flyvisual organs provide an example where furcated structureshave diverged in the timing of their differentiation relative toeach other and their ancestral developmental program Belowwe apply the phylogenetic approaches mentioned above togain greater understanding of the evolutionary origins of thisdual receptor system
221 When did the novelty originateThe evolution and development of insect eyes is fairly well
understood Structural and developmental studies comparingthe larval and adult eyes of Drosophila with those of other in-sects including outgroup species where the larval eye givesrise to the adult eye have shown that compound eyes andstemmata are paralogous in insects and likely emerged from
Eye
Diff
eren
tiatio
n(n
ot to
sca
le)
Juvenile Adult
Developm(not to
A
Fig 1 Comparison of timing of development for insect photoreceptors The Y-axi
specifically defined) The X-axis represents developmental time (units not specifica
eyes develop directly from juvenile eyes In many cases eye differentiation ceases i
Circle represents developmental field splitting Dashed line is compound eye onto
togeny both stemmata and compound eyes underwent changes in timing which m
the furcation of a single ancestral eye field (Friedrich2006a) This ancestral field first developed into an embryoniceye which persisted in development through larval stages tobecome the adult compound eye as is the case in ametabolousand hemimetabolous insects (Meinertzhagen 1973 Bate1978) In holometabolous insects larval and adult eye devel-opment is decoupleddthese two eye types develop from sep-arate primordia each derived from the same embryonic anlage(Paulus 1979 Green et al 1993 Friedrich and Benzer 2000Liu and Friedrich 2004) Thus the evolutionary furcation ofthe eye field producing both larval eyes and adult compoundeyes likely occurred within the same phylogenetic interval asthe split between hemi- and holometabolous insects (Fig 2)
222 What are the differences in components of adultand larval eyes
The differences in larval and adult fly eyes extend to theirstructure and function as well as their developmental originThe adult eye is the familiar compound eye a large complexstructure made of hundreds of ommatidia and thousands ofcells including specialized cell types called retinular (R-cells)cone cells and pigment cells (Paulus 1979) It is an image form-ing eye and has a large associated nerve network The larval eyeby comparison is quite simple It is made of few photoreceptorsand has roles in circadian rhythm entrainment and in mediat-ing phototaxis in foraging and pupating larvae (Kaneko et al1997 Helfrich-Forster et al 2002)
Interestingly in this furcation of eyes and the subsequentadoption of disparate functions there is evidence of regulatorygenetic mutations but not of gene duplication mutations Forexample Drosophila adult and larval eyes both express someof the same opsins (Malpel et al 2002) In addition many ofthe same developmental genes are expressed in both larval
ental Time scale)
Adult
CompoundEye
Stemmataembr
yoge
nesi
s
met
amor
phos
is
Eyelet
larv
al B
s represents a heuristic sketch of the amount of differentiation (with units not
lly defined) (A) The ontogeny of eyes in a hypothetical ancestral insect Adult
n adults (not shown) (B) Ontogeny of fly photoreceptors (see text for details)
geny solid line is stemmataeyelet ontogeny Relative to the ancestral eye on-
ay be considered heterochrony but not by all definitions (eg Rice 1997)
Ametabola Hemimetabola Holometabola
Reducedlarval eyes
Separate larvaland adult eyes
Fig 2 Phylogeny of the insects based on Giribet et al (2001) In the ancestral
single eye field adult eyes arose from larval eyes This can be seen in hemi-
metabolous and ametabolous insects as well as in some crustaceans In the lin-
eage leading to the Holometabola this field furcated to form adult and larval
eyes with paralogous tissue origins (gray circle) Later within the Holometa-
bola the larval eyes were reduced to produce novel photosensory organs
(black circle) These are exemplified by Bolwigrsquos organ in Drosophila
toy
toyey
otd
eya
so
en
en
zen
ato
gl
Kr
Mu
hh
dpp
wg
N
Egfr
Spi
vn
Compound Eye Bolwigrsquos Organ Ocellus
Fig 3 Evolutionary history of Drosophila photoreceptor organs compiled by
Friedrich (2006b) Photoreceptors lsquolsquofurcatersquorsquo (see text for definition) during
evolution leading to patterns of common ancestry that can be conceived as
phylogenetic trees (see also Geeta 2003 Serb and Oakley 2005) Bolwigrsquos
organ and compound eyes likely share a more recent common ancestor than
either does with ocelli (reviewed in Friedrich 2006a) Assuming those rela-
tionships we mapped presence or absence of expression of different genes
on the photoreceptor phylogeny using parsimony as implemented in Mesquite
(Maddison and Maddison 2004) Black ovals represent gain of gene expres-
sion white ovals represent loss of gene expression Gene abbreviations ey frac14eyeless Kr frac14 Kruppel zen frac14 zerknullt en frac14 engrailed toy frac14 twin-of-eyeless
vn frac14 vein Spi frac14 spitz Egfr frac14 Epidermal growth factor receptor N frac14 Notch
dpp frac14 decapentaplegic hh frac14 hedgehog Mu frac14 Munster gl frac14 glass ato frac14atonal so frac14 sine oculis otd frac14 orthodenticle eya frac14 eyes absent
390 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
and adult eyes Hedgehog and decapentaplegic specify the eyeprimordium and initiate eye development in both eye types (He-berlein et al 1993 Suzuki and Saigo 2000 Chang et al 2001)This is followed by expression of many of the same lsquolsquoretinal de-termination network genesrsquorsquo (reviewed in Friedrich 2006a) Infact 13 genes are known to be expressed in all three fly photo-receptor organs which may be taken as evidence for their com-mon ancestry (Fig 3) In addition to many similaritiesdifferences in gene expression also exist between fly compoundeyes and Bolwigrsquos organs (Friedrich 2006a) Kruppel and zenare involved in Bolwigrsquos organ development but not in that ofcompound eyes or ocelli Furthermore engrailed and toy expres-sion is absent in Bolwigrsquos organ but present in the other two vi-sual structures although engrailed has no detectable function inthe adult eye (Strutt and Mlodzik 1996) Five additional genesare known to differ in presenceabsence of expression betweencompound eye and Bolwigrsquos organ (reviewed in Friedrich2006a) Because no evidence supports the hypothesis that theseexpression differences owe their origins to gene duplicationevents concomitant with the origin of Bolwigrsquos organ a co-duplication hypothesis is rejected for all known components
223 SummaryAlthough future work on the mechanisms responsible for
the initial field splitting of fly visual organs could implicatethe involvement of duplicated genes available evidence indi-cates that the origin of separate larval and adult photoreceptororgans in insects was due primarily to regulatory genetic mu-tations that altered the timing of expression of existing geneswhereby photoreceptive organ differentiation started to occurat two different times during ontogeny Despite the fact thatgene duplication mutations can be involved in the origin ofnew photoreceptive structures as evidenced by vertebraterods and cones (Nordstrom et al 2004 Serb and Oakley2005 Plachetzki and Oakley 2007) the origin of distinct flyphotoreceptive organs produces no such evidence leading usto reject the null hypothesis of co-duplication As such fly
photoreceptive organs provide an example of how a tissue fur-cation can result in structures expressed at different times aswell as in different places Although we know that differentgene expression patterns are present in the two visual struc-tures we do not know which specific regulatory mutationscause the furcation of these organs Identifying such mutationswould be a major contribution to understanding this system
3 Additional case studies in furcation
Although we have the most genetic information availableregarding the origin of insect stemmata described abovea number of additional case studies suggest similar patterns
391TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
for the origins of other arthropod photoreceptive structures Inall cases we find no evidence for the involvement of geneduplication mutations in the origins of the photoreceptorsNevertheless in three casesdostracod median and compoundeyes insect laminar and medullar-projecting photoreceptorsand Limulus median ocelli and compound eyesdsomeevidence suggests that gene duplication mutations weresubsequently involved in the differentiation of furcated eyetypes
31 Ostracod compound eyes
311 When did the novelty originateThe origin of ostracod compound eyes provides another ex-
ample of how co-duplication can be used as a phylogeneticnull model for the origins of evolutionary complexity Molec-ular phylogenies of Ostracoda indicate that myodocopidsdtheonly Ostracoda with compound eyesdare a monophyleticgroup whose closest relatives lack compound eyes (Oakleyand Cunningham 2002 Yamaguchi and Endo 2003 Oakley2005) Although support for myodocopid monophyly is verystrong outstanding issues in ostracod phylogeny include thepossibilities that ostracods are not monophyletic (Wakayamain press) and near outgroupsdoften called lsquolsquoMaxillopodarsquorsquodarenot monophyletic (Regier et al 2005) such that understand-ing ostracod phylogeny requires understanding phylogeny ofearly branching crustaceans Crustacean and arthropod phy-logeny are notoriously difficult with different data often yield-ing different results (eg Boore et al 1995 Friedrich andTautz 1995 Regier and Shultz 1997 Giribet and Ribera2000 Giribet et al 2001 Regier et al 2005) What currentlyis clear for ostracods is that myodocopids with compound eyesare monophyletic (including Silurian specimens (Siveter et al2003 2007)) and they are closely related to some groups thatpossess lateral eyes (some juvenile thecostracans and bran-chiurans) and multiple groups that lack compound lateral
m Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
c
c
m
m
m
x
m
c
m
m
A
m
Stemc
m m
m
c
m
B
Fig 4 Inferred ancestral states null hypothesis and inferred history of myodocopid
pound and median eyes of myodocopid ostracods and relatives A species phylogen
enclosing lsquolsquomrsquorsquo represent median eyes circles enclosing lsquolsquocrsquorsquo represent compound ey
with a circle enclosing an lsquolsquoxrsquorsquo Ancestral myodocopids likely had compound and m
median eyes (Oakley and Cunningham 2002 Oakley 2005) (B) A graphical illustr
pound eyes Illustrated is a phylogenetic tree of species that includes one living and
gene duplication events are predicted to coincide in time with the origin of compoun
such as opsin (blackwhite phylogeny inside grey species phylogeny) show a speci
ralogs one expressed in compound eyes the other in median eyes Halocyprida m
duplication could hold for any component of myodocopid compound eyes rejecting
of the trait (C) Based on opsin molecular clock analyses (Fig 5) a gene duplicatio
the origin of myodocopids and their compound eyes
eyes (copepods mystacocarids many thecostracans penta-stomes and tantulocarids)
Assuming the currently available molecular phylogenetichypotheses for Ostracoda (Oakley and Cunningham 2002 Ya-maguchi and Endo 2003 Oakley 2005) statistical consider-ations indicate that compound eyes were more likely absentin the ancestor of ostracods than the alternative hypothesisthat compound eyes were lost numerous times in relatives ofnon-myodocopids However there are two important consider-ations First inclusion of additional outgroups could changethe phylogeny andor statistics but since numerous additionaloutgroups lack lateral compound eyes including more out-groups is most likely to increase support for ancestral absenceof lateral eyes Second an unambiguous discrimination of thealternatives depends on the unknown relative rate of gain ver-sus loss of lateral eyes during macroevolution which can onlybe estimated (Oakley and Cunningham 2002 Oakley 2005)
With these considerations in mind available analyses donot support lsquolsquophylogenetic homologyrsquorsquo of ostracod compoundeyes with compound eyes of other arthropods Phylogenetichomology is an explicit hypothesis about the presence of a par-ticular trait (in this case a compound eye) in a common ances-tor (Butler and Saidel 2000) Current phylogenies suggest thatcompound eyes were absent in the common ancestor of all os-tracods and later originated at or somewhat before the ances-tor of myodocopids (Fig 4A) Compound eyes could havebeen present in an earlier arthropod ancestor and regained inmyodocopids perhaps indicating lsquolsquogenerative homologyrsquorsquowhere the same developmental program allows for multiplephylogenetic origins of compound eyes (Meyer 1999 Butlerand Saidel 2000) The process of re-evolving generatively ho-mologous structures has been termed lsquolsquoswitchback evolutionrsquorsquo(Van Valen 1979) and the multifunctionality of most genes(Piatigorsky 2007) suggests genes may often be retainedeven after the loss of a particular structure (Marshall et al1994)
Halocyprida
Myodocopida
Copepoda
Myodocopida
c
m
m
x Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
x
m
C
m
m
mc
m
c
c
m
eyes and components (A) Summary of ancestral state reconstruction of com-
y in grey shows relationships of myodocopid Ostracoda and relatives Circles
es Halocyprida are ostracods that lack compound and median eyes illustrated
edian eyes whereas the ancestor of Copepoda thornMyodocopa likely had only
ation of a lsquolsquoco-duplicationrsquorsquo null hypothesis for the origin of myodocopid com-
one stem group myodocopid species and two outgroups Under co-duplication
d eyes at or before the origin of myodocopids Under such a model gene trees
fic pattern where myodocopids possess duplicated and monophyletic opsin pa-
ay or may not have lost opsins illustrated with lsquolsquorsquorsquo A null hypothesis of co-
that null indicates that regulatory genetic mutations were involved in the origin
n event (white circle) led to compound and median eye-specific paralogs after
392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

388 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
are shared between the furcated photoreceptive structures rejectthe null-hypothesis of co-duplication Shared components musthave become part of new photoreceptive structures through reg-ulatory mutations Namely components of a single ancestral re-ceptor became components of multiple furcated receptors ina descendant Therefore the genetic components gained usagein a new context and were not duplicated Although in somecases components may be shared between furcated photorecep-tive structures other components may be unique to one of thefurcated structures In these cases we take a third step to datethe origin of the components and test the co-duplication null hy-pothesis We provide examples below describing how the originof components can be dated using phylogenetic techniques likereconciled tree analysis and relaxed molecular clock dating ofgene duplication events Any genetic components that do not re-ject the co-duplication hypothesis might have contributed tonovelty through gene duplication Below we will first sketchsome developmental mechanisms that split photoreceptor fieldsspatiotemporally to allow for the origins of novel features Wethen discuss multiple case studies in arthropod photoreceptorevolution that reject co-duplication in favor of regulatory muta-tions This leads us to hypothesize that furcation of photorecep-tors by regulatory mutations not including gene duplicationmutations has been a major engine in the origins of arthropodphotoreceptor diversity
2 Mechanisms of field splitting and furcation inspace and time
Although studied in a few cases like segmentation and ap-pendage formation (Panganiban et al 1997 Seaver 2003) themechanisms of trait furcation are understudied and poorly un-derstood compared to furcation of species (speciation) andgenes (gene duplication) Yet a more comprehensive under-standing of these mechanisms would lead to insight on the or-igins of novelty during evolution because like the duplicationof a gene furcation of any biological unit with modifiablecomponents (eg photoreceptors) can lead to evolutionary di-versity when one unit gains a new function (neofunctionaliza-tion) or each retains a different partition of ancestral functions(subfunctionalization as defined by Force et al 1999) Whilethe mechanisms of gene duplication involve genetic muta-tional events and the mechanisms of speciation involve geo-graphic and population-level processes the mechanisms offurcation at the level of traits such as photoreceptors involvedevelopmental processes Next we discuss some specific ex-amples of how developmental processes may have been in-volved in furcation of photoreceptors followed by spatialandor temporal divergence of the paralogous traits These pro-cesses highlight ways that regulatory mutations may havecaused furcation of photoreceptors without the co-duplicationof associated genetic components
21 Furcation and spatial divergence
A simple example of gene regulatory changes causing furca-tion is the serial repetition of an existing structure sometimes
termed heterotopy (Haeckel 1866 Zelditch and Fink 1996West-Eberhard 2003) One example is the addition of omma-tidia to a compound eye which does not necessarily requirenew genes or cell-types to be invented instead a reuse of exist-ing genetic networks allows repetition (furcation) of a structureand its components While this simple case does not necessarilylead to a more complex eye larger numbers of ommatidia maylead to increased visual ability for example by decreasing theinter-ommatidial angle to raise visual acuity (Hecht and Wolf1929) or by increasing the region of space the eye can observeat one time One specific developmental mechanism of increas-ing ommatidial number could be the expansion of the develop-mental field by either a permissive signal (eg decapentaplegic)or the suppression of a restrictive signal (eg wingless) Thesevariationsdwhich could also lead to loss of ommatidiadarisewithin species leading to polymorphisms that may or may notbecome fixed by population level processes like selection ordrift By these mechanisms the alteration of ommatidial num-ber could originate without adding duplicated genes (eg opsin)or novel cell types In our terminology increasing ommatidialnumber would represent a furcation event The observation ofshared components of the new ommatidia would reject a nullhypothesis of co-duplication of those components and theommatidia thus highlighting the role of gene regulatory muta-tions as opposed to gene duplications in the origin of novelommatidia Even after originating by regulatory mutationsduplicated genes could still become expressed in furcatedommatidia later
22 Furcation and temporal divergence
Once furcated the paralogous structures may diverge in var-ious ways such as in their timing of differentiation which maybe considered heterochrony A focus on furcation raises an in-teresting new perspective on heterochrony because we areviewing the traits themselves as comprising evolutionary line-ages Heterochrony then becomes a comparison of ancestraland descendent developmental trajectories in the context ofthe traits rather than the more standard context of the organismIn other words heterochrony can involve a comparison betweenancestral and descendent traits as well as ancestral and descen-dent species This idea is further explored after describingchanges in timing in the Drosophila visual system develop-ment which we will argue may be considered heterochrony
The Drosophila visual system is comprised of three devel-opmentally related unitsdthe ocelli paired Bolwigrsquos organs(larval stemmata) and compound eyes (adult eyes) Here wefocus on the larval stemmata and adult eyes About a thirdof the way through embryogenesis the visual anlage un-dergoes a series of divisions (field splitting) to form laterallysymmetric precursors of the three eye types (Daniel et al1999) The larval eye is innervated and expresses opsin proteinby the end of embryogenesis (Pollock and Benzer 1988Meinertzhagen and Hanson 1993) but later undergoes furtherdifferentiation to become the adult extra-retinal eyelet(Helfrich-Forster et al 2002) In contrast the adult compoundeyes do not complete development until the final larval stage
389TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Instead they remain as undifferentiated tissue throughout em-bryonic and most of larval development (Wolff and Ready1993)
We submit that aspects of fly photoreceptor organ evolutionmay be viewed as heterochrony when focusing on the recep-tors themselves as a phylogenetic lineage The ancestor toboth fly Bolwigrsquos organs and compound eyes is a single con-tinuous developmental program where adult eyes developfrom juvenile eyes Comparisons of a hypothetical ancestorand the two paralogous descendants reveal changes in timingof differentiation (Fig 1) For example relative to an ancestralstate the fly eyelet (the adult form of Bolwigrsquos organ) mightbe considered neotonous In addition the trajectory of flycompound eye development contains a period of slow or tem-porarily arrested development relative to the ancestral stateWe note that these changes may not be called heterochronyunder the mathematical definition of Rice (1997) which re-quires uniform rate changes in developmental programsWhether or not we use the term heterochrony paralogous flyvisual organs provide an example where furcated structureshave diverged in the timing of their differentiation relative toeach other and their ancestral developmental program Belowwe apply the phylogenetic approaches mentioned above togain greater understanding of the evolutionary origins of thisdual receptor system
221 When did the novelty originateThe evolution and development of insect eyes is fairly well
understood Structural and developmental studies comparingthe larval and adult eyes of Drosophila with those of other in-sects including outgroup species where the larval eye givesrise to the adult eye have shown that compound eyes andstemmata are paralogous in insects and likely emerged from
Eye
Diff
eren
tiatio
n(n
ot to
sca
le)
Juvenile Adult
Developm(not to
A
Fig 1 Comparison of timing of development for insect photoreceptors The Y-axi
specifically defined) The X-axis represents developmental time (units not specifica
eyes develop directly from juvenile eyes In many cases eye differentiation ceases i
Circle represents developmental field splitting Dashed line is compound eye onto
togeny both stemmata and compound eyes underwent changes in timing which m
the furcation of a single ancestral eye field (Friedrich2006a) This ancestral field first developed into an embryoniceye which persisted in development through larval stages tobecome the adult compound eye as is the case in ametabolousand hemimetabolous insects (Meinertzhagen 1973 Bate1978) In holometabolous insects larval and adult eye devel-opment is decoupleddthese two eye types develop from sep-arate primordia each derived from the same embryonic anlage(Paulus 1979 Green et al 1993 Friedrich and Benzer 2000Liu and Friedrich 2004) Thus the evolutionary furcation ofthe eye field producing both larval eyes and adult compoundeyes likely occurred within the same phylogenetic interval asthe split between hemi- and holometabolous insects (Fig 2)
222 What are the differences in components of adultand larval eyes
The differences in larval and adult fly eyes extend to theirstructure and function as well as their developmental originThe adult eye is the familiar compound eye a large complexstructure made of hundreds of ommatidia and thousands ofcells including specialized cell types called retinular (R-cells)cone cells and pigment cells (Paulus 1979) It is an image form-ing eye and has a large associated nerve network The larval eyeby comparison is quite simple It is made of few photoreceptorsand has roles in circadian rhythm entrainment and in mediat-ing phototaxis in foraging and pupating larvae (Kaneko et al1997 Helfrich-Forster et al 2002)
Interestingly in this furcation of eyes and the subsequentadoption of disparate functions there is evidence of regulatorygenetic mutations but not of gene duplication mutations Forexample Drosophila adult and larval eyes both express someof the same opsins (Malpel et al 2002) In addition many ofthe same developmental genes are expressed in both larval
ental Time scale)
Adult
CompoundEye
Stemmataembr
yoge
nesi
s
met
amor
phos
is
Eyelet
larv
al B
s represents a heuristic sketch of the amount of differentiation (with units not
lly defined) (A) The ontogeny of eyes in a hypothetical ancestral insect Adult
n adults (not shown) (B) Ontogeny of fly photoreceptors (see text for details)
geny solid line is stemmataeyelet ontogeny Relative to the ancestral eye on-
ay be considered heterochrony but not by all definitions (eg Rice 1997)
Ametabola Hemimetabola Holometabola
Reducedlarval eyes
Separate larvaland adult eyes
Fig 2 Phylogeny of the insects based on Giribet et al (2001) In the ancestral
single eye field adult eyes arose from larval eyes This can be seen in hemi-
metabolous and ametabolous insects as well as in some crustaceans In the lin-
eage leading to the Holometabola this field furcated to form adult and larval
eyes with paralogous tissue origins (gray circle) Later within the Holometa-
bola the larval eyes were reduced to produce novel photosensory organs
(black circle) These are exemplified by Bolwigrsquos organ in Drosophila
toy
toyey
otd
eya
so
en
en
zen
ato
gl
Kr
Mu
hh
dpp
wg
N
Egfr
Spi
vn
Compound Eye Bolwigrsquos Organ Ocellus
Fig 3 Evolutionary history of Drosophila photoreceptor organs compiled by
Friedrich (2006b) Photoreceptors lsquolsquofurcatersquorsquo (see text for definition) during
evolution leading to patterns of common ancestry that can be conceived as
phylogenetic trees (see also Geeta 2003 Serb and Oakley 2005) Bolwigrsquos
organ and compound eyes likely share a more recent common ancestor than
either does with ocelli (reviewed in Friedrich 2006a) Assuming those rela-
tionships we mapped presence or absence of expression of different genes
on the photoreceptor phylogeny using parsimony as implemented in Mesquite
(Maddison and Maddison 2004) Black ovals represent gain of gene expres-
sion white ovals represent loss of gene expression Gene abbreviations ey frac14eyeless Kr frac14 Kruppel zen frac14 zerknullt en frac14 engrailed toy frac14 twin-of-eyeless
vn frac14 vein Spi frac14 spitz Egfr frac14 Epidermal growth factor receptor N frac14 Notch
dpp frac14 decapentaplegic hh frac14 hedgehog Mu frac14 Munster gl frac14 glass ato frac14atonal so frac14 sine oculis otd frac14 orthodenticle eya frac14 eyes absent
390 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
and adult eyes Hedgehog and decapentaplegic specify the eyeprimordium and initiate eye development in both eye types (He-berlein et al 1993 Suzuki and Saigo 2000 Chang et al 2001)This is followed by expression of many of the same lsquolsquoretinal de-termination network genesrsquorsquo (reviewed in Friedrich 2006a) Infact 13 genes are known to be expressed in all three fly photo-receptor organs which may be taken as evidence for their com-mon ancestry (Fig 3) In addition to many similaritiesdifferences in gene expression also exist between fly compoundeyes and Bolwigrsquos organs (Friedrich 2006a) Kruppel and zenare involved in Bolwigrsquos organ development but not in that ofcompound eyes or ocelli Furthermore engrailed and toy expres-sion is absent in Bolwigrsquos organ but present in the other two vi-sual structures although engrailed has no detectable function inthe adult eye (Strutt and Mlodzik 1996) Five additional genesare known to differ in presenceabsence of expression betweencompound eye and Bolwigrsquos organ (reviewed in Friedrich2006a) Because no evidence supports the hypothesis that theseexpression differences owe their origins to gene duplicationevents concomitant with the origin of Bolwigrsquos organ a co-duplication hypothesis is rejected for all known components
223 SummaryAlthough future work on the mechanisms responsible for
the initial field splitting of fly visual organs could implicatethe involvement of duplicated genes available evidence indi-cates that the origin of separate larval and adult photoreceptororgans in insects was due primarily to regulatory genetic mu-tations that altered the timing of expression of existing geneswhereby photoreceptive organ differentiation started to occurat two different times during ontogeny Despite the fact thatgene duplication mutations can be involved in the origin ofnew photoreceptive structures as evidenced by vertebraterods and cones (Nordstrom et al 2004 Serb and Oakley2005 Plachetzki and Oakley 2007) the origin of distinct flyphotoreceptive organs produces no such evidence leading usto reject the null hypothesis of co-duplication As such fly
photoreceptive organs provide an example of how a tissue fur-cation can result in structures expressed at different times aswell as in different places Although we know that differentgene expression patterns are present in the two visual struc-tures we do not know which specific regulatory mutationscause the furcation of these organs Identifying such mutationswould be a major contribution to understanding this system
3 Additional case studies in furcation
Although we have the most genetic information availableregarding the origin of insect stemmata described abovea number of additional case studies suggest similar patterns
391TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
for the origins of other arthropod photoreceptive structures Inall cases we find no evidence for the involvement of geneduplication mutations in the origins of the photoreceptorsNevertheless in three casesdostracod median and compoundeyes insect laminar and medullar-projecting photoreceptorsand Limulus median ocelli and compound eyesdsomeevidence suggests that gene duplication mutations weresubsequently involved in the differentiation of furcated eyetypes
31 Ostracod compound eyes
311 When did the novelty originateThe origin of ostracod compound eyes provides another ex-
ample of how co-duplication can be used as a phylogeneticnull model for the origins of evolutionary complexity Molec-ular phylogenies of Ostracoda indicate that myodocopidsdtheonly Ostracoda with compound eyesdare a monophyleticgroup whose closest relatives lack compound eyes (Oakleyand Cunningham 2002 Yamaguchi and Endo 2003 Oakley2005) Although support for myodocopid monophyly is verystrong outstanding issues in ostracod phylogeny include thepossibilities that ostracods are not monophyletic (Wakayamain press) and near outgroupsdoften called lsquolsquoMaxillopodarsquorsquodarenot monophyletic (Regier et al 2005) such that understand-ing ostracod phylogeny requires understanding phylogeny ofearly branching crustaceans Crustacean and arthropod phy-logeny are notoriously difficult with different data often yield-ing different results (eg Boore et al 1995 Friedrich andTautz 1995 Regier and Shultz 1997 Giribet and Ribera2000 Giribet et al 2001 Regier et al 2005) What currentlyis clear for ostracods is that myodocopids with compound eyesare monophyletic (including Silurian specimens (Siveter et al2003 2007)) and they are closely related to some groups thatpossess lateral eyes (some juvenile thecostracans and bran-chiurans) and multiple groups that lack compound lateral
m Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
c
c
m
m
m
x
m
c
m
m
A
m
Stemc
m m
m
c
m
B
Fig 4 Inferred ancestral states null hypothesis and inferred history of myodocopid
pound and median eyes of myodocopid ostracods and relatives A species phylogen
enclosing lsquolsquomrsquorsquo represent median eyes circles enclosing lsquolsquocrsquorsquo represent compound ey
with a circle enclosing an lsquolsquoxrsquorsquo Ancestral myodocopids likely had compound and m
median eyes (Oakley and Cunningham 2002 Oakley 2005) (B) A graphical illustr
pound eyes Illustrated is a phylogenetic tree of species that includes one living and
gene duplication events are predicted to coincide in time with the origin of compoun
such as opsin (blackwhite phylogeny inside grey species phylogeny) show a speci
ralogs one expressed in compound eyes the other in median eyes Halocyprida m
duplication could hold for any component of myodocopid compound eyes rejecting
of the trait (C) Based on opsin molecular clock analyses (Fig 5) a gene duplicatio
the origin of myodocopids and their compound eyes
eyes (copepods mystacocarids many thecostracans penta-stomes and tantulocarids)
Assuming the currently available molecular phylogenetichypotheses for Ostracoda (Oakley and Cunningham 2002 Ya-maguchi and Endo 2003 Oakley 2005) statistical consider-ations indicate that compound eyes were more likely absentin the ancestor of ostracods than the alternative hypothesisthat compound eyes were lost numerous times in relatives ofnon-myodocopids However there are two important consider-ations First inclusion of additional outgroups could changethe phylogeny andor statistics but since numerous additionaloutgroups lack lateral compound eyes including more out-groups is most likely to increase support for ancestral absenceof lateral eyes Second an unambiguous discrimination of thealternatives depends on the unknown relative rate of gain ver-sus loss of lateral eyes during macroevolution which can onlybe estimated (Oakley and Cunningham 2002 Oakley 2005)
With these considerations in mind available analyses donot support lsquolsquophylogenetic homologyrsquorsquo of ostracod compoundeyes with compound eyes of other arthropods Phylogenetichomology is an explicit hypothesis about the presence of a par-ticular trait (in this case a compound eye) in a common ances-tor (Butler and Saidel 2000) Current phylogenies suggest thatcompound eyes were absent in the common ancestor of all os-tracods and later originated at or somewhat before the ances-tor of myodocopids (Fig 4A) Compound eyes could havebeen present in an earlier arthropod ancestor and regained inmyodocopids perhaps indicating lsquolsquogenerative homologyrsquorsquowhere the same developmental program allows for multiplephylogenetic origins of compound eyes (Meyer 1999 Butlerand Saidel 2000) The process of re-evolving generatively ho-mologous structures has been termed lsquolsquoswitchback evolutionrsquorsquo(Van Valen 1979) and the multifunctionality of most genes(Piatigorsky 2007) suggests genes may often be retainedeven after the loss of a particular structure (Marshall et al1994)
Halocyprida
Myodocopida
Copepoda
Myodocopida
c
m
m
x Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
x
m
C
m
m
mc
m
c
c
m
eyes and components (A) Summary of ancestral state reconstruction of com-
y in grey shows relationships of myodocopid Ostracoda and relatives Circles
es Halocyprida are ostracods that lack compound and median eyes illustrated
edian eyes whereas the ancestor of Copepoda thornMyodocopa likely had only
ation of a lsquolsquoco-duplicationrsquorsquo null hypothesis for the origin of myodocopid com-
one stem group myodocopid species and two outgroups Under co-duplication
d eyes at or before the origin of myodocopids Under such a model gene trees
fic pattern where myodocopids possess duplicated and monophyletic opsin pa-
ay or may not have lost opsins illustrated with lsquolsquorsquorsquo A null hypothesis of co-
that null indicates that regulatory genetic mutations were involved in the origin
n event (white circle) led to compound and median eye-specific paralogs after
392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699
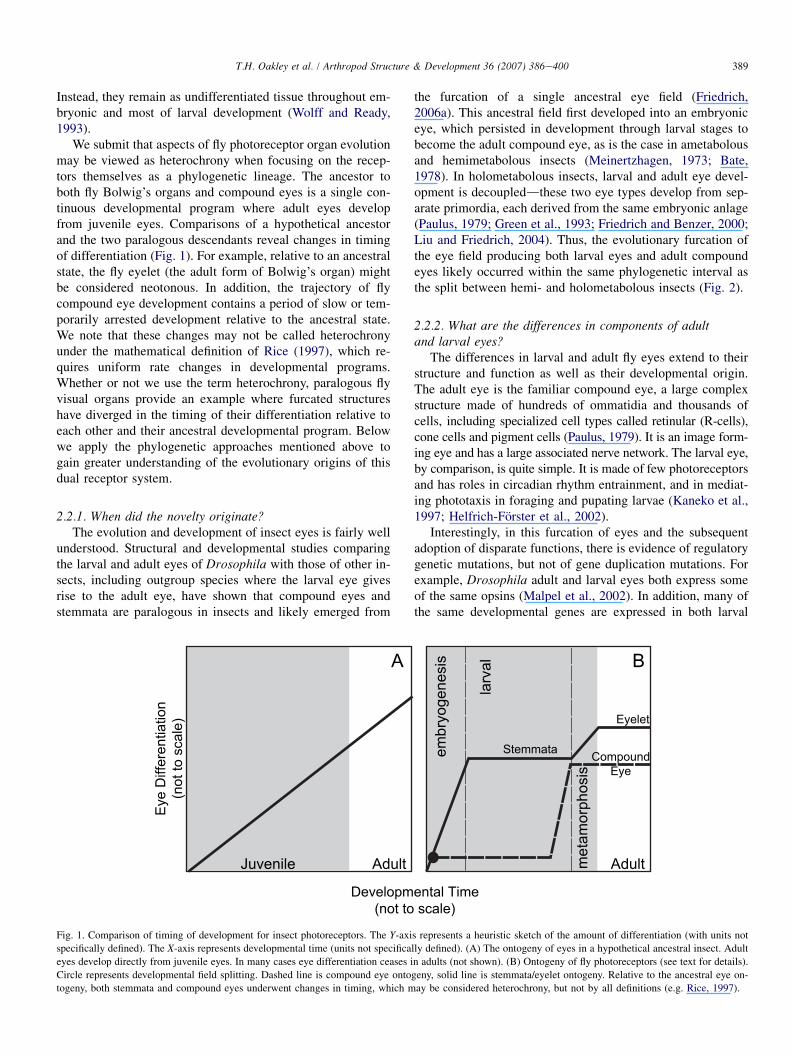
389TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Instead they remain as undifferentiated tissue throughout em-bryonic and most of larval development (Wolff and Ready1993)
We submit that aspects of fly photoreceptor organ evolutionmay be viewed as heterochrony when focusing on the recep-tors themselves as a phylogenetic lineage The ancestor toboth fly Bolwigrsquos organs and compound eyes is a single con-tinuous developmental program where adult eyes developfrom juvenile eyes Comparisons of a hypothetical ancestorand the two paralogous descendants reveal changes in timingof differentiation (Fig 1) For example relative to an ancestralstate the fly eyelet (the adult form of Bolwigrsquos organ) mightbe considered neotonous In addition the trajectory of flycompound eye development contains a period of slow or tem-porarily arrested development relative to the ancestral stateWe note that these changes may not be called heterochronyunder the mathematical definition of Rice (1997) which re-quires uniform rate changes in developmental programsWhether or not we use the term heterochrony paralogous flyvisual organs provide an example where furcated structureshave diverged in the timing of their differentiation relative toeach other and their ancestral developmental program Belowwe apply the phylogenetic approaches mentioned above togain greater understanding of the evolutionary origins of thisdual receptor system
221 When did the novelty originateThe evolution and development of insect eyes is fairly well
understood Structural and developmental studies comparingthe larval and adult eyes of Drosophila with those of other in-sects including outgroup species where the larval eye givesrise to the adult eye have shown that compound eyes andstemmata are paralogous in insects and likely emerged from
Eye
Diff
eren
tiatio
n(n
ot to
sca
le)
Juvenile Adult
Developm(not to
A
Fig 1 Comparison of timing of development for insect photoreceptors The Y-axi
specifically defined) The X-axis represents developmental time (units not specifica
eyes develop directly from juvenile eyes In many cases eye differentiation ceases i
Circle represents developmental field splitting Dashed line is compound eye onto
togeny both stemmata and compound eyes underwent changes in timing which m
the furcation of a single ancestral eye field (Friedrich2006a) This ancestral field first developed into an embryoniceye which persisted in development through larval stages tobecome the adult compound eye as is the case in ametabolousand hemimetabolous insects (Meinertzhagen 1973 Bate1978) In holometabolous insects larval and adult eye devel-opment is decoupleddthese two eye types develop from sep-arate primordia each derived from the same embryonic anlage(Paulus 1979 Green et al 1993 Friedrich and Benzer 2000Liu and Friedrich 2004) Thus the evolutionary furcation ofthe eye field producing both larval eyes and adult compoundeyes likely occurred within the same phylogenetic interval asthe split between hemi- and holometabolous insects (Fig 2)
222 What are the differences in components of adultand larval eyes
The differences in larval and adult fly eyes extend to theirstructure and function as well as their developmental originThe adult eye is the familiar compound eye a large complexstructure made of hundreds of ommatidia and thousands ofcells including specialized cell types called retinular (R-cells)cone cells and pigment cells (Paulus 1979) It is an image form-ing eye and has a large associated nerve network The larval eyeby comparison is quite simple It is made of few photoreceptorsand has roles in circadian rhythm entrainment and in mediat-ing phototaxis in foraging and pupating larvae (Kaneko et al1997 Helfrich-Forster et al 2002)
Interestingly in this furcation of eyes and the subsequentadoption of disparate functions there is evidence of regulatorygenetic mutations but not of gene duplication mutations Forexample Drosophila adult and larval eyes both express someof the same opsins (Malpel et al 2002) In addition many ofthe same developmental genes are expressed in both larval
ental Time scale)
Adult
CompoundEye
Stemmataembr
yoge
nesi
s
met
amor
phos
is
Eyelet
larv
al B
s represents a heuristic sketch of the amount of differentiation (with units not
lly defined) (A) The ontogeny of eyes in a hypothetical ancestral insect Adult
n adults (not shown) (B) Ontogeny of fly photoreceptors (see text for details)
geny solid line is stemmataeyelet ontogeny Relative to the ancestral eye on-
ay be considered heterochrony but not by all definitions (eg Rice 1997)
Ametabola Hemimetabola Holometabola
Reducedlarval eyes
Separate larvaland adult eyes
Fig 2 Phylogeny of the insects based on Giribet et al (2001) In the ancestral
single eye field adult eyes arose from larval eyes This can be seen in hemi-
metabolous and ametabolous insects as well as in some crustaceans In the lin-
eage leading to the Holometabola this field furcated to form adult and larval
eyes with paralogous tissue origins (gray circle) Later within the Holometa-
bola the larval eyes were reduced to produce novel photosensory organs
(black circle) These are exemplified by Bolwigrsquos organ in Drosophila
toy
toyey
otd
eya
so
en
en
zen
ato
gl
Kr
Mu
hh
dpp
wg
N
Egfr
Spi
vn
Compound Eye Bolwigrsquos Organ Ocellus
Fig 3 Evolutionary history of Drosophila photoreceptor organs compiled by
Friedrich (2006b) Photoreceptors lsquolsquofurcatersquorsquo (see text for definition) during
evolution leading to patterns of common ancestry that can be conceived as
phylogenetic trees (see also Geeta 2003 Serb and Oakley 2005) Bolwigrsquos
organ and compound eyes likely share a more recent common ancestor than
either does with ocelli (reviewed in Friedrich 2006a) Assuming those rela-
tionships we mapped presence or absence of expression of different genes
on the photoreceptor phylogeny using parsimony as implemented in Mesquite
(Maddison and Maddison 2004) Black ovals represent gain of gene expres-
sion white ovals represent loss of gene expression Gene abbreviations ey frac14eyeless Kr frac14 Kruppel zen frac14 zerknullt en frac14 engrailed toy frac14 twin-of-eyeless
vn frac14 vein Spi frac14 spitz Egfr frac14 Epidermal growth factor receptor N frac14 Notch
dpp frac14 decapentaplegic hh frac14 hedgehog Mu frac14 Munster gl frac14 glass ato frac14atonal so frac14 sine oculis otd frac14 orthodenticle eya frac14 eyes absent
390 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
and adult eyes Hedgehog and decapentaplegic specify the eyeprimordium and initiate eye development in both eye types (He-berlein et al 1993 Suzuki and Saigo 2000 Chang et al 2001)This is followed by expression of many of the same lsquolsquoretinal de-termination network genesrsquorsquo (reviewed in Friedrich 2006a) Infact 13 genes are known to be expressed in all three fly photo-receptor organs which may be taken as evidence for their com-mon ancestry (Fig 3) In addition to many similaritiesdifferences in gene expression also exist between fly compoundeyes and Bolwigrsquos organs (Friedrich 2006a) Kruppel and zenare involved in Bolwigrsquos organ development but not in that ofcompound eyes or ocelli Furthermore engrailed and toy expres-sion is absent in Bolwigrsquos organ but present in the other two vi-sual structures although engrailed has no detectable function inthe adult eye (Strutt and Mlodzik 1996) Five additional genesare known to differ in presenceabsence of expression betweencompound eye and Bolwigrsquos organ (reviewed in Friedrich2006a) Because no evidence supports the hypothesis that theseexpression differences owe their origins to gene duplicationevents concomitant with the origin of Bolwigrsquos organ a co-duplication hypothesis is rejected for all known components
223 SummaryAlthough future work on the mechanisms responsible for
the initial field splitting of fly visual organs could implicatethe involvement of duplicated genes available evidence indi-cates that the origin of separate larval and adult photoreceptororgans in insects was due primarily to regulatory genetic mu-tations that altered the timing of expression of existing geneswhereby photoreceptive organ differentiation started to occurat two different times during ontogeny Despite the fact thatgene duplication mutations can be involved in the origin ofnew photoreceptive structures as evidenced by vertebraterods and cones (Nordstrom et al 2004 Serb and Oakley2005 Plachetzki and Oakley 2007) the origin of distinct flyphotoreceptive organs produces no such evidence leading usto reject the null hypothesis of co-duplication As such fly
photoreceptive organs provide an example of how a tissue fur-cation can result in structures expressed at different times aswell as in different places Although we know that differentgene expression patterns are present in the two visual struc-tures we do not know which specific regulatory mutationscause the furcation of these organs Identifying such mutationswould be a major contribution to understanding this system
3 Additional case studies in furcation
Although we have the most genetic information availableregarding the origin of insect stemmata described abovea number of additional case studies suggest similar patterns
391TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
for the origins of other arthropod photoreceptive structures Inall cases we find no evidence for the involvement of geneduplication mutations in the origins of the photoreceptorsNevertheless in three casesdostracod median and compoundeyes insect laminar and medullar-projecting photoreceptorsand Limulus median ocelli and compound eyesdsomeevidence suggests that gene duplication mutations weresubsequently involved in the differentiation of furcated eyetypes
31 Ostracod compound eyes
311 When did the novelty originateThe origin of ostracod compound eyes provides another ex-
ample of how co-duplication can be used as a phylogeneticnull model for the origins of evolutionary complexity Molec-ular phylogenies of Ostracoda indicate that myodocopidsdtheonly Ostracoda with compound eyesdare a monophyleticgroup whose closest relatives lack compound eyes (Oakleyand Cunningham 2002 Yamaguchi and Endo 2003 Oakley2005) Although support for myodocopid monophyly is verystrong outstanding issues in ostracod phylogeny include thepossibilities that ostracods are not monophyletic (Wakayamain press) and near outgroupsdoften called lsquolsquoMaxillopodarsquorsquodarenot monophyletic (Regier et al 2005) such that understand-ing ostracod phylogeny requires understanding phylogeny ofearly branching crustaceans Crustacean and arthropod phy-logeny are notoriously difficult with different data often yield-ing different results (eg Boore et al 1995 Friedrich andTautz 1995 Regier and Shultz 1997 Giribet and Ribera2000 Giribet et al 2001 Regier et al 2005) What currentlyis clear for ostracods is that myodocopids with compound eyesare monophyletic (including Silurian specimens (Siveter et al2003 2007)) and they are closely related to some groups thatpossess lateral eyes (some juvenile thecostracans and bran-chiurans) and multiple groups that lack compound lateral
m Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
c
c
m
m
m
x
m
c
m
m
A
m
Stemc
m m
m
c
m
B
Fig 4 Inferred ancestral states null hypothesis and inferred history of myodocopid
pound and median eyes of myodocopid ostracods and relatives A species phylogen
enclosing lsquolsquomrsquorsquo represent median eyes circles enclosing lsquolsquocrsquorsquo represent compound ey
with a circle enclosing an lsquolsquoxrsquorsquo Ancestral myodocopids likely had compound and m
median eyes (Oakley and Cunningham 2002 Oakley 2005) (B) A graphical illustr
pound eyes Illustrated is a phylogenetic tree of species that includes one living and
gene duplication events are predicted to coincide in time with the origin of compoun
such as opsin (blackwhite phylogeny inside grey species phylogeny) show a speci
ralogs one expressed in compound eyes the other in median eyes Halocyprida m
duplication could hold for any component of myodocopid compound eyes rejecting
of the trait (C) Based on opsin molecular clock analyses (Fig 5) a gene duplicatio
the origin of myodocopids and their compound eyes
eyes (copepods mystacocarids many thecostracans penta-stomes and tantulocarids)
Assuming the currently available molecular phylogenetichypotheses for Ostracoda (Oakley and Cunningham 2002 Ya-maguchi and Endo 2003 Oakley 2005) statistical consider-ations indicate that compound eyes were more likely absentin the ancestor of ostracods than the alternative hypothesisthat compound eyes were lost numerous times in relatives ofnon-myodocopids However there are two important consider-ations First inclusion of additional outgroups could changethe phylogeny andor statistics but since numerous additionaloutgroups lack lateral compound eyes including more out-groups is most likely to increase support for ancestral absenceof lateral eyes Second an unambiguous discrimination of thealternatives depends on the unknown relative rate of gain ver-sus loss of lateral eyes during macroevolution which can onlybe estimated (Oakley and Cunningham 2002 Oakley 2005)
With these considerations in mind available analyses donot support lsquolsquophylogenetic homologyrsquorsquo of ostracod compoundeyes with compound eyes of other arthropods Phylogenetichomology is an explicit hypothesis about the presence of a par-ticular trait (in this case a compound eye) in a common ances-tor (Butler and Saidel 2000) Current phylogenies suggest thatcompound eyes were absent in the common ancestor of all os-tracods and later originated at or somewhat before the ances-tor of myodocopids (Fig 4A) Compound eyes could havebeen present in an earlier arthropod ancestor and regained inmyodocopids perhaps indicating lsquolsquogenerative homologyrsquorsquowhere the same developmental program allows for multiplephylogenetic origins of compound eyes (Meyer 1999 Butlerand Saidel 2000) The process of re-evolving generatively ho-mologous structures has been termed lsquolsquoswitchback evolutionrsquorsquo(Van Valen 1979) and the multifunctionality of most genes(Piatigorsky 2007) suggests genes may often be retainedeven after the loss of a particular structure (Marshall et al1994)
Halocyprida
Myodocopida
Copepoda
Myodocopida
c
m
m
x Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
x
m
C
m
m
mc
m
c
c
m
eyes and components (A) Summary of ancestral state reconstruction of com-
y in grey shows relationships of myodocopid Ostracoda and relatives Circles
es Halocyprida are ostracods that lack compound and median eyes illustrated
edian eyes whereas the ancestor of Copepoda thornMyodocopa likely had only
ation of a lsquolsquoco-duplicationrsquorsquo null hypothesis for the origin of myodocopid com-
one stem group myodocopid species and two outgroups Under co-duplication
d eyes at or before the origin of myodocopids Under such a model gene trees
fic pattern where myodocopids possess duplicated and monophyletic opsin pa-
ay or may not have lost opsins illustrated with lsquolsquorsquorsquo A null hypothesis of co-
that null indicates that regulatory genetic mutations were involved in the origin
n event (white circle) led to compound and median eye-specific paralogs after
392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

Ametabola Hemimetabola Holometabola
Reducedlarval eyes
Separate larvaland adult eyes
Fig 2 Phylogeny of the insects based on Giribet et al (2001) In the ancestral
single eye field adult eyes arose from larval eyes This can be seen in hemi-
metabolous and ametabolous insects as well as in some crustaceans In the lin-
eage leading to the Holometabola this field furcated to form adult and larval
eyes with paralogous tissue origins (gray circle) Later within the Holometa-
bola the larval eyes were reduced to produce novel photosensory organs
(black circle) These are exemplified by Bolwigrsquos organ in Drosophila
toy
toyey
otd
eya
so
en
en
zen
ato
gl
Kr
Mu
hh
dpp
wg
N
Egfr
Spi
vn
Compound Eye Bolwigrsquos Organ Ocellus
Fig 3 Evolutionary history of Drosophila photoreceptor organs compiled by
Friedrich (2006b) Photoreceptors lsquolsquofurcatersquorsquo (see text for definition) during
evolution leading to patterns of common ancestry that can be conceived as
phylogenetic trees (see also Geeta 2003 Serb and Oakley 2005) Bolwigrsquos
organ and compound eyes likely share a more recent common ancestor than
either does with ocelli (reviewed in Friedrich 2006a) Assuming those rela-
tionships we mapped presence or absence of expression of different genes
on the photoreceptor phylogeny using parsimony as implemented in Mesquite
(Maddison and Maddison 2004) Black ovals represent gain of gene expres-
sion white ovals represent loss of gene expression Gene abbreviations ey frac14eyeless Kr frac14 Kruppel zen frac14 zerknullt en frac14 engrailed toy frac14 twin-of-eyeless
vn frac14 vein Spi frac14 spitz Egfr frac14 Epidermal growth factor receptor N frac14 Notch
dpp frac14 decapentaplegic hh frac14 hedgehog Mu frac14 Munster gl frac14 glass ato frac14atonal so frac14 sine oculis otd frac14 orthodenticle eya frac14 eyes absent
390 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
and adult eyes Hedgehog and decapentaplegic specify the eyeprimordium and initiate eye development in both eye types (He-berlein et al 1993 Suzuki and Saigo 2000 Chang et al 2001)This is followed by expression of many of the same lsquolsquoretinal de-termination network genesrsquorsquo (reviewed in Friedrich 2006a) Infact 13 genes are known to be expressed in all three fly photo-receptor organs which may be taken as evidence for their com-mon ancestry (Fig 3) In addition to many similaritiesdifferences in gene expression also exist between fly compoundeyes and Bolwigrsquos organs (Friedrich 2006a) Kruppel and zenare involved in Bolwigrsquos organ development but not in that ofcompound eyes or ocelli Furthermore engrailed and toy expres-sion is absent in Bolwigrsquos organ but present in the other two vi-sual structures although engrailed has no detectable function inthe adult eye (Strutt and Mlodzik 1996) Five additional genesare known to differ in presenceabsence of expression betweencompound eye and Bolwigrsquos organ (reviewed in Friedrich2006a) Because no evidence supports the hypothesis that theseexpression differences owe their origins to gene duplicationevents concomitant with the origin of Bolwigrsquos organ a co-duplication hypothesis is rejected for all known components
223 SummaryAlthough future work on the mechanisms responsible for
the initial field splitting of fly visual organs could implicatethe involvement of duplicated genes available evidence indi-cates that the origin of separate larval and adult photoreceptororgans in insects was due primarily to regulatory genetic mu-tations that altered the timing of expression of existing geneswhereby photoreceptive organ differentiation started to occurat two different times during ontogeny Despite the fact thatgene duplication mutations can be involved in the origin ofnew photoreceptive structures as evidenced by vertebraterods and cones (Nordstrom et al 2004 Serb and Oakley2005 Plachetzki and Oakley 2007) the origin of distinct flyphotoreceptive organs produces no such evidence leading usto reject the null hypothesis of co-duplication As such fly
photoreceptive organs provide an example of how a tissue fur-cation can result in structures expressed at different times aswell as in different places Although we know that differentgene expression patterns are present in the two visual struc-tures we do not know which specific regulatory mutationscause the furcation of these organs Identifying such mutationswould be a major contribution to understanding this system
3 Additional case studies in furcation
Although we have the most genetic information availableregarding the origin of insect stemmata described abovea number of additional case studies suggest similar patterns
391TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
for the origins of other arthropod photoreceptive structures Inall cases we find no evidence for the involvement of geneduplication mutations in the origins of the photoreceptorsNevertheless in three casesdostracod median and compoundeyes insect laminar and medullar-projecting photoreceptorsand Limulus median ocelli and compound eyesdsomeevidence suggests that gene duplication mutations weresubsequently involved in the differentiation of furcated eyetypes
31 Ostracod compound eyes
311 When did the novelty originateThe origin of ostracod compound eyes provides another ex-
ample of how co-duplication can be used as a phylogeneticnull model for the origins of evolutionary complexity Molec-ular phylogenies of Ostracoda indicate that myodocopidsdtheonly Ostracoda with compound eyesdare a monophyleticgroup whose closest relatives lack compound eyes (Oakleyand Cunningham 2002 Yamaguchi and Endo 2003 Oakley2005) Although support for myodocopid monophyly is verystrong outstanding issues in ostracod phylogeny include thepossibilities that ostracods are not monophyletic (Wakayamain press) and near outgroupsdoften called lsquolsquoMaxillopodarsquorsquodarenot monophyletic (Regier et al 2005) such that understand-ing ostracod phylogeny requires understanding phylogeny ofearly branching crustaceans Crustacean and arthropod phy-logeny are notoriously difficult with different data often yield-ing different results (eg Boore et al 1995 Friedrich andTautz 1995 Regier and Shultz 1997 Giribet and Ribera2000 Giribet et al 2001 Regier et al 2005) What currentlyis clear for ostracods is that myodocopids with compound eyesare monophyletic (including Silurian specimens (Siveter et al2003 2007)) and they are closely related to some groups thatpossess lateral eyes (some juvenile thecostracans and bran-chiurans) and multiple groups that lack compound lateral
m Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
c
c
m
m
m
x
m
c
m
m
A
m
Stemc
m m
m
c
m
B
Fig 4 Inferred ancestral states null hypothesis and inferred history of myodocopid
pound and median eyes of myodocopid ostracods and relatives A species phylogen
enclosing lsquolsquomrsquorsquo represent median eyes circles enclosing lsquolsquocrsquorsquo represent compound ey
with a circle enclosing an lsquolsquoxrsquorsquo Ancestral myodocopids likely had compound and m
median eyes (Oakley and Cunningham 2002 Oakley 2005) (B) A graphical illustr
pound eyes Illustrated is a phylogenetic tree of species that includes one living and
gene duplication events are predicted to coincide in time with the origin of compoun
such as opsin (blackwhite phylogeny inside grey species phylogeny) show a speci
ralogs one expressed in compound eyes the other in median eyes Halocyprida m
duplication could hold for any component of myodocopid compound eyes rejecting
of the trait (C) Based on opsin molecular clock analyses (Fig 5) a gene duplicatio
the origin of myodocopids and their compound eyes
eyes (copepods mystacocarids many thecostracans penta-stomes and tantulocarids)
Assuming the currently available molecular phylogenetichypotheses for Ostracoda (Oakley and Cunningham 2002 Ya-maguchi and Endo 2003 Oakley 2005) statistical consider-ations indicate that compound eyes were more likely absentin the ancestor of ostracods than the alternative hypothesisthat compound eyes were lost numerous times in relatives ofnon-myodocopids However there are two important consider-ations First inclusion of additional outgroups could changethe phylogeny andor statistics but since numerous additionaloutgroups lack lateral compound eyes including more out-groups is most likely to increase support for ancestral absenceof lateral eyes Second an unambiguous discrimination of thealternatives depends on the unknown relative rate of gain ver-sus loss of lateral eyes during macroevolution which can onlybe estimated (Oakley and Cunningham 2002 Oakley 2005)
With these considerations in mind available analyses donot support lsquolsquophylogenetic homologyrsquorsquo of ostracod compoundeyes with compound eyes of other arthropods Phylogenetichomology is an explicit hypothesis about the presence of a par-ticular trait (in this case a compound eye) in a common ances-tor (Butler and Saidel 2000) Current phylogenies suggest thatcompound eyes were absent in the common ancestor of all os-tracods and later originated at or somewhat before the ances-tor of myodocopids (Fig 4A) Compound eyes could havebeen present in an earlier arthropod ancestor and regained inmyodocopids perhaps indicating lsquolsquogenerative homologyrsquorsquowhere the same developmental program allows for multiplephylogenetic origins of compound eyes (Meyer 1999 Butlerand Saidel 2000) The process of re-evolving generatively ho-mologous structures has been termed lsquolsquoswitchback evolutionrsquorsquo(Van Valen 1979) and the multifunctionality of most genes(Piatigorsky 2007) suggests genes may often be retainedeven after the loss of a particular structure (Marshall et al1994)
Halocyprida
Myodocopida
Copepoda
Myodocopida
c
m
m
x Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
x
m
C
m
m
mc
m
c
c
m
eyes and components (A) Summary of ancestral state reconstruction of com-
y in grey shows relationships of myodocopid Ostracoda and relatives Circles
es Halocyprida are ostracods that lack compound and median eyes illustrated
edian eyes whereas the ancestor of Copepoda thornMyodocopa likely had only
ation of a lsquolsquoco-duplicationrsquorsquo null hypothesis for the origin of myodocopid com-
one stem group myodocopid species and two outgroups Under co-duplication
d eyes at or before the origin of myodocopids Under such a model gene trees
fic pattern where myodocopids possess duplicated and monophyletic opsin pa-
ay or may not have lost opsins illustrated with lsquolsquorsquorsquo A null hypothesis of co-
that null indicates that regulatory genetic mutations were involved in the origin
n event (white circle) led to compound and median eye-specific paralogs after
392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

391TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
for the origins of other arthropod photoreceptive structures Inall cases we find no evidence for the involvement of geneduplication mutations in the origins of the photoreceptorsNevertheless in three casesdostracod median and compoundeyes insect laminar and medullar-projecting photoreceptorsand Limulus median ocelli and compound eyesdsomeevidence suggests that gene duplication mutations weresubsequently involved in the differentiation of furcated eyetypes
31 Ostracod compound eyes
311 When did the novelty originateThe origin of ostracod compound eyes provides another ex-
ample of how co-duplication can be used as a phylogeneticnull model for the origins of evolutionary complexity Molec-ular phylogenies of Ostracoda indicate that myodocopidsdtheonly Ostracoda with compound eyesdare a monophyleticgroup whose closest relatives lack compound eyes (Oakleyand Cunningham 2002 Yamaguchi and Endo 2003 Oakley2005) Although support for myodocopid monophyly is verystrong outstanding issues in ostracod phylogeny include thepossibilities that ostracods are not monophyletic (Wakayamain press) and near outgroupsdoften called lsquolsquoMaxillopodarsquorsquodarenot monophyletic (Regier et al 2005) such that understand-ing ostracod phylogeny requires understanding phylogeny ofearly branching crustaceans Crustacean and arthropod phy-logeny are notoriously difficult with different data often yield-ing different results (eg Boore et al 1995 Friedrich andTautz 1995 Regier and Shultz 1997 Giribet and Ribera2000 Giribet et al 2001 Regier et al 2005) What currentlyis clear for ostracods is that myodocopids with compound eyesare monophyletic (including Silurian specimens (Siveter et al2003 2007)) and they are closely related to some groups thatpossess lateral eyes (some juvenile thecostracans and bran-chiurans) and multiple groups that lack compound lateral
m Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
c
c
m
m
m
x
m
c
m
m
A
m
Stemc
m m
m
c
m
B
Fig 4 Inferred ancestral states null hypothesis and inferred history of myodocopid
pound and median eyes of myodocopid ostracods and relatives A species phylogen
enclosing lsquolsquomrsquorsquo represent median eyes circles enclosing lsquolsquocrsquorsquo represent compound ey
with a circle enclosing an lsquolsquoxrsquorsquo Ancestral myodocopids likely had compound and m
median eyes (Oakley and Cunningham 2002 Oakley 2005) (B) A graphical illustr
pound eyes Illustrated is a phylogenetic tree of species that includes one living and
gene duplication events are predicted to coincide in time with the origin of compoun
such as opsin (blackwhite phylogeny inside grey species phylogeny) show a speci
ralogs one expressed in compound eyes the other in median eyes Halocyprida m
duplication could hold for any component of myodocopid compound eyes rejecting
of the trait (C) Based on opsin molecular clock analyses (Fig 5) a gene duplicatio
the origin of myodocopids and their compound eyes
eyes (copepods mystacocarids many thecostracans penta-stomes and tantulocarids)
Assuming the currently available molecular phylogenetichypotheses for Ostracoda (Oakley and Cunningham 2002 Ya-maguchi and Endo 2003 Oakley 2005) statistical consider-ations indicate that compound eyes were more likely absentin the ancestor of ostracods than the alternative hypothesisthat compound eyes were lost numerous times in relatives ofnon-myodocopids However there are two important consider-ations First inclusion of additional outgroups could changethe phylogeny andor statistics but since numerous additionaloutgroups lack lateral compound eyes including more out-groups is most likely to increase support for ancestral absenceof lateral eyes Second an unambiguous discrimination of thealternatives depends on the unknown relative rate of gain ver-sus loss of lateral eyes during macroevolution which can onlybe estimated (Oakley and Cunningham 2002 Oakley 2005)
With these considerations in mind available analyses donot support lsquolsquophylogenetic homologyrsquorsquo of ostracod compoundeyes with compound eyes of other arthropods Phylogenetichomology is an explicit hypothesis about the presence of a par-ticular trait (in this case a compound eye) in a common ances-tor (Butler and Saidel 2000) Current phylogenies suggest thatcompound eyes were absent in the common ancestor of all os-tracods and later originated at or somewhat before the ances-tor of myodocopids (Fig 4A) Compound eyes could havebeen present in an earlier arthropod ancestor and regained inmyodocopids perhaps indicating lsquolsquogenerative homologyrsquorsquowhere the same developmental program allows for multiplephylogenetic origins of compound eyes (Meyer 1999 Butlerand Saidel 2000) The process of re-evolving generatively ho-mologous structures has been termed lsquolsquoswitchback evolutionrsquorsquo(Van Valen 1979) and the multifunctionality of most genes(Piatigorsky 2007) suggests genes may often be retainedeven after the loss of a particular structure (Marshall et al1994)
Halocyprida
Myodocopida
Copepoda
Myodocopida
c
m
m
x Halocyprida
Myodocopida
Copepoda
Stem Myodocopida
x
m
C
m
m
mc
m
c
c
m
eyes and components (A) Summary of ancestral state reconstruction of com-
y in grey shows relationships of myodocopid Ostracoda and relatives Circles
es Halocyprida are ostracods that lack compound and median eyes illustrated
edian eyes whereas the ancestor of Copepoda thornMyodocopa likely had only
ation of a lsquolsquoco-duplicationrsquorsquo null hypothesis for the origin of myodocopid com-
one stem group myodocopid species and two outgroups Under co-duplication
d eyes at or before the origin of myodocopids Under such a model gene trees
fic pattern where myodocopids possess duplicated and monophyletic opsin pa-
ay or may not have lost opsins illustrated with lsquolsquorsquorsquo A null hypothesis of co-
that null indicates that regulatory genetic mutations were involved in the origin
n event (white circle) led to compound and median eye-specific paralogs after
392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

392 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Unlike compound eyes multiple lines of evidence indicatethat myodocopid median eyes are homologous to median eyesof close relatives (Oakley and Cunningham 2002 Oakley2003c) Ostracod median eyes show variation among majorgroups of Podocopa (Tanaka 2005) but are generally verysimilar to those in related crustacean groups including somefeatures considered diagnostic like the presence of tapetalcells (Elofsson 1966 Andersson and Nilsson 1981 Oakleyand Cunningham 2002 Oakley 2003c) Both morphologyand phylogenetic distribution are consistent with homologyof myodocopid podocopid and some non-ostracod (ie cope-pod and thecostracan) median eyes Based on these obser-vations and the eyesrsquo phylogenetic distributions Oakley(2003c) proposed that median eyes were present and com-pound eyes were absent in ancestral ostracods (whether ornot compound eyes were present in a more distant arthropodancestor whereby the myodocopid compound eye wouldrepresent a re-gain) Oakley (2003c) speculated that genesused for compound eyes could be maintained even if theeyes themselves were lost such that myodocopid lateraleyes are generatively but not phylogenetically homologousBelow we hypothesize a possible mechanism in furcation bydevelopmental field splitting for such a (re)gain of convergentcompound eyes Perhaps many genetic components of com-pound eyes are identical to or recent duplicates of the compo-nents of median eyes
312 What are the components of myodocopidcompound eyes
By definition complex traits like compound eyes consist ofnumerous components For example thousands of genes areexpressed in eye development of Drosophila melanogasterand 371 were specifically up-regulated in response to ectopicexpression of eyeless in leg imaginal disks (Michaut et al2003) Although little is known about the specific componentsof compound eyes in non-model organisms like Ostracoda wecan make some initial hypotheses
First we hypothesize that like fly photoreceptive organs(Figs 1e3) ostracod photoreceptive organs (compound andmedian eyes) develop from a single anlage Although myodo-copid eye development in particular has not been studied yetsome researchers have hypothesized that multiple photorecep-tive organs within other crustacean species develop from sin-gle anlagen (reviewed in Elofsson 2006) In Penaeusduorarum a U-shaped anlage gives rise to both compoundand frontal eyes (Dobkin 1961) Similarly in Artemia salinaeyes and brain develop from a V-shaped anlage The frontaleyes develop from the base of the V shape and the compoundeyes and brain from the arms of the V-shaped anlage (Benesch1969 reviewed in Elofsson 2006) Finally in Triops a compli-cated developmental pattern holds Elofsson (2006) summa-rizes the work of Dahl (1959) by stating that dorsolaterallobes of one anlage lsquolsquosplit longitudinally into the anlage sup-plying frontal eyes [which are of dual developmental origin]and the one intended for the compound eyesrsquorsquo In contrast re-cent studies using cell proliferation markers have found thatthe cells of lateral and median eyes of Artemia and Limulus
proliferate at different times (Wildt and Harzsch 2002Harzsch et al 2006) which does not directly support their or-igin from single anlagen Nevertheless we speculate that com-pound and median eyes of myodocopids and other arthropodspecies may develop from single anlagen
Similarly we hypothesize that like Drosophila ostracodsrsquomultiple photoreceptive organs share a common ancestrywhich should be evidenced by shared expression of manygenes The specific identity of shared genes between ostracodeye types is predicted by the conservation of many geneticand developmental processes across animal eye developmentFor example retinal determination genes are highly conservedacross animals (Gehring and Ikeo 1999) and opsin-mediatedphototransduction is a process with conserved components inmost animals (Arendt 2003 Plachetzki and Oakley 2007)Understanding how numerous existing components came tobe used in phylogenetically non-homologousdbut potentiallyparalogousdcompound eyes represents a major research pro-gram for the future To initiate such a research program ourlab has examined one critical component of eyes namely opsinthe protein that forms light reactive visual pigments of animaleyes (Oakley and Huber 2004)
313 When did the components originateEvolutionary noveltiesdeven those as complex as eyesd
originate by either structural or regulatory mutations that be-come fixed in populations (but see West-Eberhard 2003) Giventhe broad evidence for shared genes and developmental pro-cesses involved in ontogeny of all animal eyes and other traitsit is highly unlikely that the numerous components of a complextrait like a myodocopid eye would arise de novo Instead com-ponents of a new trait should usually be assembled from exist-ing components From this perspective new alternatives can beproposed did the components of new myodocopid eyes arise bygene duplications or by regulatory mutations that caused exist-ing components to be expressed in a new location (bricolage)These components are used for vision so components of newcompound eyes were co-opted or duplicated from existing me-dian eye components Of these alternatives the possibility ofgene duplications provides a testable null hypothesis to inves-tigate the origins of myodocopid compound eyes Namelybased on phylogenetic analyses indicating that compoundeyes (re-)originated in myodocopids a null hypothesis isthat all components of that eye duplicated coincident withthe origin of the eye itself
How do we test this null hypothesis and determine whencompound eye components originated We will explain twophylogenetic approaches First reconciled tree analysis(RTA) (Page 1993 Page and Charleston 1998) can be usedto determine when gene duplications occurred Under thenull hypothesis of co-duplication gene duplication eventsshould date to the time of inferred origin of the eye structuresThis null hypothesis makes three testable predictions with re-spect to any component of the myodocopid compound eye(Fig 4B) (1) Components (genes) exist that are used onlyin the compound eye of myodocopids (2) Those genes areparalogs (duplicates) of genes expressed elsewhere in
393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

393TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
myodocopids probably in the median eye (3) Non-myodoco-pids possess a single copy of the gene in question such thatthe gene duplication occurred after the speciation event lead-ing to myodocopids on one branch and the nearest outgroupson the other branch If any of these hypotheses are not metco-duplication of that component and the eye itself is rejectedin favor of an origin by regulatory mutation
We illustrate the RTA approach with existing data from os-tracod opsins where all three hypotheses outlined are metthus initially supporting the null hypothesis of co-duplicationover bricolage First opsin genes exist that are used only inthe compound eyes of myodocopids We previously used re-verse transcriptaseePCR to amplify opsins expressed in myo-docopid compound eyes and uncovered a clade of compound
=1000mya
gt520 mya
First FossilMyodocopidCompound Eye
425 mya
gt520 mya
Fig 5 A chronogram of various arthropod opsin genes The phylogenetic relation
duplication (Fig 3B) because a gene duplication event leading to compound an
to copepods thorn myodocopids and myodocopids themselves However the chronogr
copid median eye and compound eye opsins is substantially younger than the firs
illustrated opsin chronogram we first obtained opsin amino acid sequences from Ge
the best-fit model using ProtTest (Abascal et al 2005) to be RtRev thorn I thorn G and as
phyml (Guindon and Gascuel 2003) The ML tree with branch lengths was used fo
plemented in r8s using an empirically determined smoothing parameter (Sanderson
bilaterians to be 1 billion years and second setting a minimum divergence for Cop
estimate for bilaterian divergence times to show that we still estimate relatively yo
timated origins of the eyes where those paralogs are expressed Accession numbers
AAI13559 AAL37512 AAL37523 AAL37548 AAP49025 AAP49026 AAS88
ABI48892 CAI47588 CAI47588 CAI47589 CAI47590 NP_477096 NP_524
Q25157 Q25158 and Q94741 The Ixodes sequence was obtained from EST G
genome traces Two sequences were previously unpublished Full length Tigrio30- and 50-RACE (Frohman et al 1988) Full details for isolation of those two se
eye-specific opsins that were not expressed in median eyesor other tissues (Oakley and Huber 2004) In addition thecompound eye opsins were found to be paralogs (duplicates)of opsins expressed elsewhere in median eyes Finallya non-myodocopid (the copepod Tigriopus californicus lack-ing compound eyes) possesses a single ortholog of the dupli-cated myodocopid genes Phylogenetic and RTA analysesindicate that the gene duplication giving rise to compoundeye specific opsins occurred after the split of copepods and os-tracods but before the split of two myodocopid ostracods(Fig 5) Although this is a broad phylogenetic interval thuslessening the power of the test we cannot reject the null hy-pothesis of duplication of opsin at the origin of myodocopidcompound eyes using RTA alone
206 mya 20 mya
Limulus polyphemus Comp EyeIxodes scapularis
Homo sapiens melanopsinDrosophila melanogaster Rh7Hemigrapsus sanguinus One
Tigriopus californicus Median Eye
Drosophila melanogaster Rh5
Limulus polyphemus Ocellus
Drosophila melanogaster Rh4Drosophila melanogaster Rh3
Drosophila melanogaster Rh2
Drosophila melanogaster Rh1
Drosophila melanogaster Rh6
Calliphora vicina Rh1
Calliphora vicina Rh3
Calliphora vicina Rh5
Calliphora vicina Rh6
Vanessa cardui Blue
Vanessa cardui UV
Hemigrapsus sanguinus Two
Vargula hilgendorfii Median Eye
Vargula tsujii Comp EyeSkogsbergia lerneri Median Eye
Skogsbergia lerneri Comp Eye
Schistocerca gregariaOpsin2Papilio glaucus
Bactrocera dorsalis
Colias philodice UV
Archaeomysis grebnitzkii
Neomysis americana
Homarus gammarus
Neogonodactylus oerstedii
Procambarus clarkii
Mysis diluviana
Euphausia superba
ships of myodocopid and copepod (Tigriopus) opsins alone do not reject co-
d median eye-specific paralogs occurs between the speciation events leading
am offers a further test that rejects co-duplication The divergence of myodo-
t myodocopid compound eye preserved in the fossil record To construct the
nBank and aligned them using tcoffee (Notredame et al 2000) We determined
sumed that model to estimate the maximum likelihood (ML) phylogeny using
r a penalized likelihood relaxed molecular clock divergence time analysis im-
2003) We report results using two calibration points first fixing the origin of
epoda and Ostracoda to be 520 million years We deliberately picked a high
ung ages for the myodocopid and Limulus opsin duplications compared to es-
for sequences used in this analysis are AAB31031 AAB31032 AAD29445
872 AAY16532 ABG37009 ABI48869 ABI48874 ABI48884 ABI48885
035 NP_524368 NP_524398 P17646 P22269 P35356 P35360 P35361
893P57RK12 whose full sequence was confirmed by comparison to public
pus californicus and Vargula tsujii cDNA sequences were obtained using
quences will be published elsewhere
394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

394 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
A second related way to test the null hypothesis of co-duplication is by explicitly estimating divergence times forthe origin of components of myodocopid compound eyesand comparing them to each other and to the estimated originof the compound eye itself Under a co-duplication nullhypothesis origination times of eye components and thosefor the eye itself should be statistically indistinguishableAgain we illustrate this concept using opsin genes andmyodocopid compound eyes
In the case of myodocopid compound eyes the fossil recordprovides a direct estimate for the timing of origin The earliestfossil myodocopids are exquisitely preserved specimens com-plete with lateral compound eyes from the Silurian(w425 million years ago) of Herefordshire United Kingdom(Siveter et al 2003 2007) These specimens have some fea-tures unique to living myodocopids like compound eyes and7th limbs but are likely stem group myodocopids represent-ing extinct sister taxa of living myodocopids (Siveter et al2007) Whether they are stem or crown group myodocopidsthese fossils represent a minimum age estimate for the originof myodocopid compound eyes as 425 million years ago(mya) Given this age estimate we can compare explicit esti-mates of the origin of compound eye-specific opsins as a fur-ther test of the null hypothesis that opsin gene duplication wasinvolved in the origin of myodocopid compound eyes Withcurrently available data we are able to reject the null hypo-thesis of the coincident timing of opsin gene duplication andcompound eye origins in myodocopids Namely we usedPenalized Likelihood (Sanderson 2002 2003) to generate a re-laxed molecular clock estimate for the gene duplication eventleading to myodocopid compound eye opsins Even takinga conservative approach of using a very old estimate of bilat-erian origins of 1 billion years to calibrate arthropod opsinphylogeny (Wray et al 1996 Benton and Donoghue 2007)the origin of myodocopid compound eye-specific opsins is es-timated to be much younger than the Silurian compound eyeapproximately 206 mya (Fig 5) The relaxed molecular clockanalysis therefore rejects the null hypothesis of co-duplicationof opsins and myodocopid eyes
314 Summary and implicationsMyodocopid compound eyes are likely not phylogeneti-
cally homologous to compound eyes of other arthropods(Oakley and Cunningham 2002 Oakley 2005) Ostracodcompound eyes express a unique opsin gene with no evidenceof expression anywhere else (Oakley and Huber 2004) Thecompound eye opsins are most closely related to paralogousopsins expressed in the median eye of myodocopids Thesefacts are consistent with the possibility of opsin gene duplica-tion associated with the origin of ostracod compound eyesHowever a relaxed molecular clock analysis that estimatedthe origin of compound eye opsins indicates an age ofw200 mya much younger than the first fossilized ostracodcompound eye from w425 mya (Siveter et al 2003) Therelaxed clock analysis thus rejects the null hypothesis ofco-duplication in favor of regulatory mutations in opsin atthe origin of myodocopid compound eyes These results
indicate that early in myodocopid history ostracod compoundeyes may have used the same opsin as the median eye Lateropsin duplicated allowing different paralogs to be expressedin each eye type This scenario of gene duplication leadingto dual functional specialization of an ancestral gene hasbeen discussed in the field of molecular evolution (Piatigorskyand Wistow 1989 Hughes and Hughes 1993 Proulx andPhillips 2006)
A major challenge for the hypothesized evolutionary eventsoutlined above is the morphological similarity of ostracod com-pound eyes with other arthropod compound eyes Althoughhighly detailed homoplasies are rather common during evolu-tion (Conway Morris 2003) convergence on the cellular pat-terns in ostracod eyes would be surprising If ostracodcompound eyes represent convergence how are the detailedmorphological similarities with other arthropods explained ge-netically Namely although the number of cells in ostracod om-matidia differs from other arthropods (reviewed in Oakley2003c) ostracod ommatidia possess retinular crystallinecone and pigment cells like other arthropod compound eyesOne possibility is switchback evolution described above wheregenetic programs are maintained in the absence of a structureThe difficulty with switchback evolution is that not only genesbut also developmental processes and protein interactions thatlead to compound eye morphology must be (re)gained The rea-son for the maintenance of interactions may be more difficult toexplain than the simple maintenance of the genes themselvesCurrently there is no clear answer to this difficulty however ex-amination of genes known to be involved in arthropod com-pound eye morphogenesis such as spacemaker (Zelhof et al2006 Osorio 2007) and many others (reviewed in Tepass andHarris 2007) could provide some clues especially if thesegenesdand more importantly their interactionsdare used inmultiple developmental processes
32 Pancrustacean outer photoreceptor cells R1eR6
A subset of the photoreceptor cells (retinular or lsquolsquoRrsquorsquo cells)of hexapod and crustacean compound eyes provides anothertrait that illustrates how the null hypothesis of co-duplicationcan be used to understand character evolution Many pancrus-taceans possess eight R-cells (Paulus 1979) In Drosophilamelanogaster these cells are named R1eR8 in each ommatid-ium the basic unit that makes up the faceted (compound)lateral eyes found in each of the major arthropod divisions(for alternative R-cell nomenclature see Meinertzhagen1976) Based on the neuroanatomy of crustaceansdmostlystudied in Malacostraca (Bullock and Horridge 1965) andbranchiopods (Melzer et al 1997)dand the fruitfly Drosoph-ila melanogaster (Morante and Desplan 2004) R-cells may begrouped into separate homologous evolutionary modulescells R1eR6 and cells R7 and R8 In flies R1eR6 project tothe lamina whereas R7 and R8 project to the medulla twostructures that are elaborated from the protocerebrum (Bullockand Horridge 1965) Here we use some of the phylogeneticapproaches outlined above to examine the evolutionary originsof the R1eR6dlaminar projectingdphotoreceptor cells
395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

395TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
321 When did the R1e6 receptors originateUnderstanding when laminar-projecting photoreceptors
originated requires homologizing R-cell types in different ar-thropods in relationship to arthropod phylogenydsubjectsthat require additional research (Paulus 2000 Yu et al2004) Nevertheless some preliminary hypotheses can bemade For example the order of R-cell specification providesclues to homology In the ontogeny of the Drosophila omma-tidium R8 is specified first R1e6 are then specified in cou-plets through a series of lateral inductions and R7 isspecified last (Morante and Desplan 2004) Similar patternsof retinular cell specification during ommatidial ontogenyhave been observed in the notostracans Triops and Lepidurus(Melzer et al 1997 2000 Harzsch and Walossek 2001)and the decapods Procambarus (Hafner and Tokarski 1998)and Homarus (Hafner and Tokarski 2001) Namely thesame progression of R-cell specification noted above for Dro-sophila (ie R8 first then R1e6 in couplets and R7 last) isevident in the notostracan ommatidia that have been studiedFurther ommatidia from insects and crustaceans are specifiedin the morphogenetic furrow a wave of cell differentiation thatsweeps across the retina For the purpose of explication wetentatively place the origin of the outer retinular cell evolution-ary module at the base of Hexapoda and the Malacostraca thornBranchiopoda (Fig 6A) Because the monophyly of the
Procambarus
Drosophila
Myriapoda
Limulus
Rh1
Rh2
Rh6
Ocellar
CE
HexapodaCrustaceaMyriapodaChelicerata
A
B
CE
Fig 6 Phylogenetic depiction of the evolution of the R1e6 module (A) Avail-
able evidence suggests that the R1e6 module (gray circle) originated prior to
Pancrustacea (see text) (B) Phylogenetic relationships (see Fig 5) of the clade
of arthropod opsins that includes Rh1 reconciled to a conservative arthropod
species tree suggesting evolution by bricolage for the R1e6 module Inconsis-
tent with the null hypothesis of co-duplication Rh1 opsin evolved by gene du-
plication within the hexapod lineage Rh1 opsin orthologs do not exist in other
non-hexapod taxa but the origin of the larger clade of opsins that includes Rh1
evolved prior to the major arthropod lineages No opsin data currently exist
from myriapods CE compound eye Rh rhodopsin
Crustacea is often not supported by molecular data (Regierand Shultz 1997 Lavrov et al 2004 Cook et al 2005)and because R-cell ontogeny is unstudied in major crustaceangroups like Ostracoda and Thecostraca our placement of theR1e6 origination must be considered tenuous Neverthelessthis placement is conservative with respect to the presentdiscussion
322 What are the components of R1e6 photoreceptorsAs discussed above for compound eyes in general compo-
nents of laminar projecting cells are numerous and best knownin Drosophila Seven opsin genes have been identified inDrosophila (Rh1e7) six of which are expressed in the eightR-cells of fly ommatidia The R1e6 group of cells expressesa single opsin (Rh1) whereas R7 expresses either Rh3 orRh4 and R8 expresses either Rh3 or Rh5 (Morante and Des-plan 2004) In addition to opsin expression a suite of regula-tory genes has been shown to coordinate the fates of differentsets of R cells and include atonal for R8 (Jarman et al 1994)rough for R2 and R5 (Heberlein et al 1991) seven-up whichprevents R7 fate in R1 R2 R3 and R4 Bar and lozenge for R1and R6 (Mlodzik et al 1990 Higashijima et al 1992) andsevenless (Harris et al 1976) bride of sevenless (Reinkeand Zipursky 1988) prospero (Cook et al 2003) and Notch(Cooper and Bray 2000) for R7 In many cases these factorswork in a combinatorial fashion in specifying R-cell fate andmay effect the specification of other cell types as well(Nagaraj and Banerjee 2004)
323 When did the components originateIn our phylogenetic scheme for elucidating the mechanisms
of novelty origination (ie gene duplication plus divergenceand regulatory mutation) the next step is to ask what arethe evolutionary histories of the components that comprisethe trait in question and do their origins coincide with thatof the higher-level trait There is no evidence for duplicationwithin pancrustaceans of the regulatory components listedabove Therefore these components must have become in-volved in development of the two different R-cell modulesby regulatory mutations In contrast different opsin genesare expressed in the R1eR6 cells compared to R7eR8 cellsDoes the origin of these furcated photoreceptor modules coin-cide with the duplication of opsin genes
One way to address this issue is to conduct reconciled treeanalyses using a species phylogeny and a phylogeny of the dif-ferently expressed opsin genes Our null hypothesis predictsthat both the components of the trait (opsin) and the trait itself(the R1e6 module) will appear at the same phylogenetic inter-val We have already placed the origin of the R1e6 module atthe last common ancestor of the Hexapoda and the Malacostra-ca thorn Branchiopoda Does the history of the opsins expressedin the laminar-projecting photoreceptors also fit this pattern
Fig 6B shows the results of reconciling arthropod Rh1-clade opsin phylogeny with the species phylogeny Our analy-sis suggests that the Rh1 opsin specific to R1e6 moduleexpression has evolved by gene duplication within the hexa-pod lineage more recently than a minimum estimate for the
396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

396 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
origin of dual photoreceptor modules Furthermore searches ofsequence databases did not reveal Rh1 opsin orthologs fromany taxon outside of Hexapoda Further bioinformatic screensof the genome trace data from Daphnia pulex did not reveal anyRh1 opsins leading us to believe that the apparent absence ofRh1 from crustaceans is not due to loss (data not shown) Thusin the case of Rh1 expression we can reject the null hypothesisof duplication at the origination of the R1e6 module as well
324 SummaryHere we have described how our phylogenetic scheme can
be used to elucidate the mutational mechanisms (ie gene du-plication or regulatory mutations) involved with the origin ofretinular cell morphologies We conceive of retinal cell clus-ters as furcated units R1eR6 (laminar projecting) are oneunit and R7eR8 (medullar projecting) are another unit Aco-duplication null hypothesis where gene duplication eventswere associated with the origin of these units is rejected withcurrently available data Even in the case of opsin where dif-ferential expression of paralogous genes is known reconciledtree analysis indicates that this opsin gene duplication is morerecent than the origin of the separate photoreceptor cell clus-ters Not unlike ostracod eyes the opsin data suggest thata gene duplication event with subsequent partitioning of ex-pression might have served to further differentiate the laminarand medullar projecting photoreceptor clusters after theseclusters originally furcated by regulatory mutations
33 Additional examples
We have presented multiple examples aimed at understand-ing the origins of evolutionary novelties in arthropod photore-ceptors using phylogenetic comparative approaches Thisapproach suggests a research program in the comparative biol-ogy of arthropod photoreceptors highlighting the need for ad-ditional developmental and molecular data especially fromnon-model groups We believe the patterns illustrated by ourexamples will apply more broadly across arthropods and webriefly indicate here a few additional examples of potentialfurcation events of arthropod photoreceptors
Limulus and relatives possess median ocelli and compoundeyes which express different opsins (Smith et al 1993 butsee Dalal et al 2003) Were these opsin paralogs co-duplicatedwith the eye types themselves Similar to ostracod opsins theanswer based on relaxed molecular clock analysis of Limulusopsin (Fig 5) and the fossil record appears to be lsquolsquonorsquorsquo A diver-gence time estimate for the Limulus paralogs is 20 mya faryounger than known occurrences of limulid xiphosuran andeurypterid median ocelli in the fossil record (Stormer 1952)which we assume to be homologous to Limulus ocelli Hereagain the null hypothesis of co-duplication is rejected suchthat opsin duplication was probably not involved in the originof xiphosuran eyes Rather the gene duplication leading to dif-ferential expression in the two eye types apparently came later
As another example hyperiid amphipods exhibit a wide ar-ray of compound eyes generally correlated with the differingvisual requirements and light regimes of the diverse ocean
depths at which they live (Land 1989a) In the mid-water do-main of hyperiids down-welling light from the surface is muchgreater than the reflected light from objects below Yet as smallcarnivores hyperiids must be able to see objects both aboveand below While some midwater fish have solved this problemby evolving two very different types of eyes (eg Lockett1977) hyperiid eyes are split to see effectively in both condi-tions by using an upper and lower field of ommatidia with di-vergent morphological properties The extent of the differencesrange from distinct facet sizes in upper and lower portions ofa single eye to a double retina in a single eye to two distincteyes (Land 1989a) Interestingly the retinas of one double-eyed hyperiid (Phronima sedentaria) originate from a singleembryonic retinal field (Minkiewicz 1909) therefore repre-senting a furcation event The morphological similarity of com-ponents namely the various cell types making up the eye andcommon developmental origin suggests that the double eyesare made largely of the same genetic components perhapsspecified by the same developmental networks in two spatiallocations Furthermore there is no evidence for separate opsinsin the two eye-types Electroretinograms showed only one peakof spectral sensitivity although it is unclear whether both eyeswere included in each sample (Frank and Widder 1999) If thegenetic components are indeed identical the null hypothesis ofco-duplication would be once again rejected in favor of regula-tory genetic changes that allowed a field splitting event allow-ing the origination of the novel eyes of P sedentaria and closerelatives
Other potential examples exist in arthropods The multipleeyes of arachnids could be the result of furcationdwere geneduplications involved in their origins Other arthropods alsopossess many eyes such as the 12 found in the larvae of thediving beetle Thermonectus marmoratus (Mandapaka et al2006) Even the multiple receptors of ocelli median and fron-tal eyes (Elofsson 2006) could be the result of furcationevents The split compound eyes of gyrinid beetles one por-tion looking into the water and one portion out would makean interesting case study of an lsquolsquoincipient furcationrsquorsquo Many ar-thropods also possess specializations in part of the retina suchas highly acute zones in predatorrsquos eyes (Land 1989b) or spec-tral sensitivity differences linked to behavior or habitat suchas honey bee drones specialized to detect the queen againstthe sky (Stavenga 1992) In one ostracod species a radicalsexual dimorphism develops by furcation In Euphilomedescarcharodonta females burrow in the sand and have tiny rudi-mentary eyes while adult males are free swimming and havelarge compound eyes This dimorphism occurs via a furcationof the developing larval eye field This furcation happens onlyin the males where one of the duplicated fields goes on to formthe large compound eyes (Rivera and Oakley in preparation)In all these cases it is interesting to ponder whether gene du-plications were required for the specializations or splits tooriginate Did gene duplications come later to differentiatethe specialized modules that possess different functional de-mands These are just a few of many possible examplesfrom the complex evolutionary history of arthropod light re-ceptive organs
397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

397TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
4 Summary and discussion
Progress in science is often driven by the tension betweenalternative themes that color researchersrsquo perceptions of nature(Shermer 2002) Here we use the evolutionary history of ar-thropod compound eyes to explore a thematic pair that hasrisen to the forefront of current evolutionary thinkingdthe rel-ative role of structural (gene duplication) versus regulatory(co-option) mutations in the origins of novelty Not unlikethe paired concepts of neutral and adaptive evolution thestructuralregulatory dichotomy has produced advocates forboth alternatives (Ohno 1970 King and Wilson 1975 Jacob1977 Carroll et al 2005) As with any thematic pair reality isa mixture of both alternativesdstructural and regulatory ge-netic changes have both had critical impacts on the origin ofnovelties As with some thematic pairsdlike neutral versusadaptive molecular changedone theme may be used asa null hypothesis that when rejected provides support for theother Analogous to how neutral theory provides a null hypo-thesis for adaptive molecular evolution (Kreitman 1996) wesuggest a means by which the co-occurrence in time of geneduplication mutations and the origin of higher level traits(lsquolsquoco-duplicationrsquorsquo) may be considered as a null hypothesisthat when rejected favors a role for regulatory mutations inthe origins of novelties This approach involves explicit com-parative phylogenetic approaches and tools to provide a valu-able null model for the evolutionary origins of novel traits Assuch even if co-duplication is rarely true it provides a testablenull hypothesis If in fact true those components for whichco-duplication cannot be rejected provide interesting subjectsfor further study Does natural selection act on co-duplicatedunits as a whole or individually How did specific co-duplica-tional events originatedwere they genome duplications orsegmental duplications of co-regulated complexes Has selec-tion favored the assembly of genetic units amenable to co-duplication
In the case of arthropod eye evolution we follow Frie-drich (2006a) and Oakley (2003a) and conceive of photore-ceptors as phylogenetic lineages that may lsquolsquofurcatersquorsquo duringevolution by developmental mechanisms While gene dupli-cations could be involved in these furcation events as theyseem to be in vertebrate rodcone origins (Nordstrom et al2004 Serb and Oakley 2005 Plachetzki and Oakley2007) we find little evidence for gene duplication events as-sociated with the origins of novel arthropod photoreceptivestructures Rejecting this null hypothesis of lsquolsquoco-duplica-tionrsquorsquodthe duplication of genes associated with higher-levelfurcation eventsdpoints instead to regulatory mutations asa primary driver of the furcation of arthropod photoreceptivestructures Following furcation the subsequent diversificationof the photoreceptive organs has involved both regulatorychanges as evidenced by differential expression of non-duplicated genes in fly stemmata and compound eyes andby differential expression of duplicated genes in myodocopidostracod compound and median eyes hexapod laminar andmedullar photoreceptors and Limulus compound eyes andocelli
The conceptual approach outlined here raises issues thatmust be developed further For example one concern maybe ambiguity over what exactly is splitting Does splitting in-volve morphological structures or gene regulatory mecha-nisms We maintain that the answer is both We takefurcation to mean lineage splitting of any lsquolsquomodulersquorsquo at anylevel of biological organization (Serb and Oakley 2005)This highlights the need for a clear definition of a modulea subject of entire books (Wagner 2001 Schlosser and Wag-ner 2004) Just as the study of speciation was propelled for-ward by a usable definition of a species (Mayr 1942)a pragmatic definition of modules is required for understand-ing novelty We advocatedand practiced here as much as pos-sibledthe explicit definition of a module of interest asa collection of specific interacting components This allowsa pragmatic fluidity to the module concept For example thesplit eyes of P sedentaria or gyrinid beetles discussed abovecould be defined as one module or two depending on the ques-tion We seek to understand how these lsquolsquodualrsquorsquo retinas origi-nated such that we define each species as having twoseparate modules From a phylogenetic perspective the simi-larity of modules indicates common ancestry indicating thatmechanisms exist to have split them Similarly we can definemodules as gene regulatory networks again by specificallyenumerating their components The perspective advocatedhere is an approach to understanding the relative history ofthe specifically defined components of modules New modulesdo not often evolve from nothing Therefore we maintain theyusually have a shared ancestry with diversification driven byfurcational processes
This comparative perspective hints at the possibility ofa complicated evolutionary history of arthropod photoreceptiveorgans If the histories of fly stemmata and myodocopid com-pound eyes sketched here hold up to further scrutiny the re-sults might generalize and indicate common switchingbetween compound and simple eyes following furcationevents This is a non-conventional hypothesis Neverthelesssuch a hypothesis is consistent with many observations de-scribed above such as the common development of differenteye types from single anlage the common genetic programsof different types of eyes and the complicated phylogeneticdistributions of frontal and lateral eyes But how does suchlsquolsquoswitchingrsquorsquo occur morphologically The simplification ofcompound eyes is not controversial for example in the originof stemmata The primary challenge to this hypothesis is that itrequires multiple origins of compound eye morphogenesis anapparently complicated set of developmental events that oncelost seems difficult to regain
Under this hypothesis origin of arthropod photoreceptorsby furcation indicates the possibility of generating a phyloge-netic tree for numerousdperhaps mostdparalogous photore-ceptive organs of arthropods (and outgroups) Alreadya phylogenetic tree of Drosophila photoreceptors has beenconceived (Friedrich 2006ab) and gene expression changescan be mapped onto that tree (Fig 3) As analogous infor-mation becomes available outside flies we will be able tobuild trees of numerous arthropod photoreceptors and test
398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

398 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
interesting questions such as the homology andor paralogy ofmedian and lateral photoreceptors and the homology andorparalogy of ommatidial types in different arthropod groups
Acknowledgments
Thanks to two anonymous referees for valuable criticismsand A Syme and A Wilson for comments and ideas Thiswork was supported by NSF grants to THO and DCP
References
Abascal F Zardoya R Posada D 2005 ProtTest selection of best-fit
models of protein evolution Bioinformatics 21 2104e2105
Andersson A Nilsson DE 1981 Fine-structure and optical-properties of an
ostracode (Crustacea) nauplius eye 107 361e374
Arendt D 2003 Evolution of eyes and photoreceptor cell types International
Journal of Developmental Biology 47 563e571
Bate CM 1978 Development of sensory systems in Arthropods In
Jacobson M (Ed) Handbook of Sensory Physiology Vol IX Springer
Heidelberg pp 1e53
Benesch R 1969 Zur Ontogenie und Morphologie von Artemia salina Zoo-
logische Jahrbucher Abteilung fur Anatomie und Ontogenie der Tiere 86
307e458
Benton MJ Donoghue PC 2007 Paleontological evidence to date the tree
of life Molecular Biology and Evolution 24 26e53
Boore JL Collins TM Stanton D Daehler LL Brown WM 1995 De-
ducing the pattern of arthropod phylogeny from mitochondrial DNA rear-
rangements Nature 376 163e165
Britten RJ Davidson EH 1969 Gene regulation for higher cells a theory
Science 165 349e357
Bullock TH Horridge GA 1965 Structure and function in the nervous sys-
tems of invertebrates Freeman San Francisco
Butler AB Saidel WM 2000 Defining sameness historical biological
and generative homology Bioessays 22 846e853
Carroll SB Grenier JK Weatherbee SD 2005 From DNA to Diversity
Molecular Genetics and the Evolution of Animal Design Blackwell
Oxford
Chang T Mazotta J Dumstrei K Dumitrescu A Hartenstein V 2001
Dpp and Hh signaling in the Drosophila embryonic eye field Develop-
ment 128 4691e4704
Conway Morris S 2003 Lifersquos Solution Inevitable Humans in a Lonely Uni-
verse Cambridge University Press Cambridge
Cook T Pichaud F Sonneville R Papatsenko D Desplan C 2003 Dis-
tinction between color photoreceptor cell fates is controlled by Prospero in
Drosophila Developmental Cell 4 853e864
Cook CE Yue Q Akam M 2005 Mitochondrial genomes suggest that
hexapods and crustaceans are mutually paraphyletic Proceedings of the
Royal Society of London Biological Sciences 272 1295e1304
Cooper MT Bray SJ 2000 R7 photoreceptor specification requires Notch
activity Current Biology 10 1507e1510
Cunningham CW Omland KO Oakley TH 1998 Reconstructing ances-
tral character states a critical reappraisal Trends in Ecology and Evolution
13 361e366
Dahl E 1959 The ontogeny and comparative anatomy of some protocerebral
sense organs in notostracan phyllopods Quarterly Journal of Microscopi-
cal Science 100 445e462
Dalal JS Jinks RN Cacciatore C Greenberg RM Battelle BA 2003
Limulus opsins diurnal regulation of expression Visual Neuroscience 20
523e534
Daniel A Dumstrei K Lengyel JA Hartenstein V 1999 The control of
cell fate in the embryonic visual system by atonal tailless and EGFR sig-
naling Development 126 2945e2954
Dobkin S 1961 Early developmental stages of pink shrimp Penaeus duorarum
from Florida Waters US Fish and Wildlife Service US Govt Print Office
Dobzhansky T 1937 Genetics and the origin of species Columbia University
Press New York
Elofsson R 1966 The nauplius eye and frontal organs in Malacostraca (Crus-
tacea) Sarsia 25 1e25
Elofsson R 2006 The frontal eyes of crustaceans Arthropod Structure and
Development 35 275e291
Force A Lynch M Pickett FB Amores A Yan YL Postlethwait J
1999 Preservation of duplicate genes by complementary degenerative mu-
tations Genetics 151 1531e1545
Frank TM Widder EA 1999 Comparative study of the spectral sensitiv-
ities of mesopelagic crustaceans Journal of Comparative Physiology A
185 255e265
Friedrich M 2006a Continuity versus split and reconstitution exploring the
molecular developmental corollaries of insect eye primordium evolution
Developmental Biology 299 310e329
Friedrich M 2006b Ancient mechanisms of visual sense organ development
based on comparison of the gene networks controlling larval eye ocellus
and compound eye specification in Drosophila Arthropod Structure and
Development 35 357e378
Friedrich M Benzer S 2000 Divergent decapentaplegic expression patterns
in compound eye development and the evolution of insect metamorphosis
Journal of Experimental Zoology 288 39e55
Friedrich M Tautz D 1995 Ribosomal DNA phylogeny of the major extant
arthropod classes and the evolution of myriapods Nature 376 165e167
Frohman MA Dush MK Martin GR 1988 Rapid production of full-
length cDNAs from rare transcripts amplification using a single gene-spe-
cific oligonucleotide primer Proceedings of the National Academy of Sci-
ences USA 85 8998e9002
Geeta R 2003 Structure trees and species trees what they say about morpho-
logical development and evolution Evolution and Development 5 609e621
Gehring WJ Ikeo K 1999 Pax 6 Mastering eye morphogenesis and eye
evolution Trends in Genetics 15 371e377
Giribet G Ribera C 2000 A review of arthropod phylogeny New data
based on ribosomal DNA sequences and direct character optimization Cla-
distics 16 204e231
Giribet G Edgecombe GD Wheeler WC 2001 Arthropod phylogeny
based on eight molecular loci and morphology Nature 413 157e161
Green P Hartenstein A Hartenstein V 1993 The embryonic development
of the Drosophila visual system Cell and Tissue Research 273 583e598
Guindon S Gascuel O 2003 A simple fast and accurate algorithm to es-
timate large phylogenies by maximum likelihood Systematic Biology 52
696e704
Haeckel E 1866 Generelle Morphologie der Organismen Allgemeine
Grundzuge der organischen Formen-Wissenschaft mechanisch begrundet
durch die von Charles Darwin reformierte Descendenz-Theorie Georg
Reimer Berlin
Hafner GS Tokarski TR 1998 Morphogenesis and pattern formation in
the retina of the crayfish Procambarus clarkii Cell and Tissue Research
293 535e550
Hafner GS Tokarski TR 2001 Retinal development in the lobster Homa-
rus americanusdComparison with compound eyes of insects and other
crustaceans Cell and Tissue Research 305 147e158
Halder G Callaerts P Gehring WJ 1995 Induction of ectopic eyes by tar-
geted expression of the eyeless gene in Drosophila Science 267 1788e
1792
Harris WA Stark WS Walker JA 1976 Genetic dissection of photore-
ceptor system in compound eye of Drosophila melanogaster Journal of
Physiology-London 256 415
Harzsch S Walossek D 2001 Neurogenesis in the developing visual system
of the branchiopod crustacean Triops longicaudatus (LeConte 1846) cor-
responding patterns of compound-eye formation in Crustacea and Insecta
Development Genes and Evolution 211 37e43
Harzsch S Vilpoux K Blackburn DC Plachetzki DC Brown NL
Melzer R Kempler KE Batelle BA 2006 Evolution of arthropod vi-
sual systems development of the eyes and central visual pathways in the
horseshoe crab Limulus polyphemus Linnaeus 1758 (Chelicerata Xipho-
sura) Developmental Dynamics 235 2641e2655
399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

399TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Heberlein U Mlodzik M Rubin GM 1991 Cell-fate determination in the
developing Drosophila eye role of the rough gene Development 112
703e712
Heberlein U Wolff T Rubin GM 1993 The TGF beta homolog dpp and
the segment polarity gene hedgehog are required for propagation of a mor-
phogenetic wave in the Drosophila retina Cell 75 913e926
Hecht S Wolf E 1929 The visual acuity of the honey bee Journal of Gen-
eral Physiology 12 727e760
Helfrich-Forster C Edwards T Yasuyama K Wisotzki B Schneuwly S
Stanewsky R Meinertzhagen IA et al 2002 The extraretinal eyelet of
Drosophila development ultrastructure and putative circadian function
Journal of Neuroscience 22 9255e9266
Higashijima S Kojima T Michiue T Ishimaru S Emori Y Saigo K
1992 Dual Bar homeobox genes of Drosophila required in two photore-
ceptor cells R1 and R6 and primary pigment cells for normal eye devel-
opment Genes and Development 6 50e60
Hughes AL 1994 The evolution of functionally novel proteins after gene
duplicaton Philosophical Transactions of the Royal Society of London
B 256 119e124
Hughes MK Hughes AL 1993 Evolution of duplicated genes in a tetra-
ploid animal Xenopus laevis Molecular Biology and Evolution 10
1360e1369
Jacob F 1977 Evolution and tinkering Science 196 1161e1166
Jarman AP Grell EH Ackerman L Jan LY Jan YN 1994 Atonal is
the proneural gene for Drosophila photoreceptors Nature 369 398e400
Kaneko M Helfrich-Forster C Hall JC 1997 Spatial and temporal ex-
pression of the period and timeless genes in the developing nervous system
of Drosophila newly identified pacemaker candidates and novel features
of clock gene product cycling Journal of Neuroscience 17 6745e6760
King MC Wilson AC 1975 Evolution at two levels in humans and chim-
panzees Science 188 107e116
Kreitman M 1996 The neutral theory is dead Long live the neutral theory
Bioessays 18 678e683
Land MF 1989a The eyes of hyperiid amphipods relations of optical struc-
ture to depth Journal of Comparative Physiology A 164 751e762
Land M 1989b Variations in the structure and design of compound eyes In
Hardie RC Stavenga DG (Eds) Facets of Vision Springer Berlin
Heidelberg New York pp 471e492
Land MF Nilsson D-E 2002 Animal Eyes Oxford University Press
Oxford
Lavrov DV Brown WM Boore JL 2004 Phylogenetic position of the
Pentastomida and (pan)crustacean relationships Proceedings of the Royal
Society of London B 271 537e544
Li W-H 1997 Molecular Evolution Sinauer Sunderland MA
Liu Z Friedrich M 2004 The Tribolium homologue of glass and the evo-
lution of insect larval eyes Developmental Biology 269 36e54
Lockett NA 1977 Adaptations to the deep-sea environment In
Crescitelli F (Ed) Handbook of sensory physiology Vol VII5 Springer
Berlin pp 67e192
Maddison WP 1997 Gene trees in species trees Systematic Biology 46
523e536
Maddison WP Maddison DR 2004 Mesquite a modular system for evo-
lutionary analysis version 105
Malpel S Klarsfeld A Rouyer F 2002 Larval optic nerve and adult extra-
retinal photoreceptors sequentially associate with clock neurons during
Drosophila brain development Development 129 1443e1453
Mandapaka K Morgan RC Buschbeck EK 2006 Twenty-eight retinas
but only twelve eyes an anatomical analysis of the larval visual system
of the diving beetle Thermonectus marmoratus (Coleoptera Dytiscidae)
Journal of Comparative Neurology 497 166e181
Marshall CR Raff EC Raff RA 1994 Dollorsquos law and the death and re-
surrection of genes Proceedings of the National Academies of Sciences
USA 91 12283e12287
Mayr E 1942 Systematics and the Origin of Species Columbia University
Press New York
McGinnis W Garber RL Wirz J Kuroiwa A Gehring WJ 1984a A
homologous protein-coding sequence in Drosophila homeotic genes and
its conservation in other metazoans Cell 37 403e408
McGinnis W Hart CP Gehring WJ Ruddle FH 1984b Molecular clon-
ing and chromosome mapping of a mouse DNA sequence homologous to
homeotic genes of Drosophila Cell 38 675e680
Meinertzhagen IA 1973 Development of the compound eye and optic lobe
of insects In Young D (Ed) Developmental Neurobiology of Arthro-
pods Cambridge University Press Cambridge pp 51e104
Meinertzhagen IA 1976 The organization of perpendicular fibre pathways
in the insect optic lobe Philosophical Transactions of the Royal Society
London B 274 555e594
Meinertzhagen IA Hanson TE 1993 The development of the optic
lobe In Bate M Martinez-Arias A (Eds) The Development of Dro-
sophila melanogaster Cold Spring Harbor Laboratory Press pp 1363e1491
Melzer RR Diersch R Nicastro D Smola U 1997 Compound eye evo-
lution Highly conserved retinula and cone cell patterns indicate a common
origin of the insect and crustacean ommatidium Naturwissenschaften 84
542e544
Melzer RR Michalke C Smola U 2000 Walking on insect paths Early
ommatidial development in the compound eye of the ancestral crustacean
Triops cancriformis Naturwissenschaften 87 308e311
Meyer A 1999 Homology and homoplasy the retention of genetic pro-
grammes Novartis Foundation Symposia 222 141e153
Michaut L Flister S Neeb M White KP Certa U Gehring WJ 2003
Analysis of the eye developmental pathway in Drosophila using DNA mi-
croarrays Proceedings of the National Academies of Sciences USA 100
4024e4029
Minkiewicz R 1909 Memoire sur la biologie du Tonnelier de mer (Phronimasedentaria Forsk) I La coloration normale des Phronimes et son devel-
oppement par migration progressive des chromatophores Bulletin de LrsquoIn-
stitut Oceanographique Monaco 146 1e21
Mlodzik M Hiromi Y Weber U Goodman CS Rubin GM 1990 The
Drosophila seven-up gene a member of the steroid receptor gene super-
family controls photoreceptor cell fates Cell 60 211e224
Morante J Desplan C 2004 Building a projection map for photoreceptor
neurons in the Drosophila optic lobes Seminars in Cell and Developmen-
tal Biology 15 137e143
Muller GB Wagner GP 1991 Novelty in evolution Restructuring the
concept Annual Review of Ecology and Systematics 22 229e256
Nagaraj R Banerjee U 2004 The little R cell that could International Jour-
nal of Developmental Biology 48 755e760
Nordstrom K Larsson TA Larhammar D 2004 Extensive duplications of
phototransduction genes in early vertebrate evolution correlate with block
(chromosome) duplications Genomics 83 852e872
Notredame C Higgins DG Heringa J 2000 T-Coffee A novel method
for fast and accurate multiple sequence alignment Journal of Molecular
Biology 302 205e217
OrsquoHara RJ 1997 Population thinking and tree thinking in systematics
Zoologica Scripta 26 323e329
Oakley TH 2003a The eye as a replicating and diverging modular develop-
mental unit Trends in Ecology and Evolution 18 623e627
Oakley TH 2003b Maximum likelihood models of character trait evolution
Comments on Theoretical Biology 8 1e17
Oakley TH 2003c On homology of arthropod compound eyes Integrative
and Comparative Biology 43 522e530
Oakley TH 2005 Myodocopa (Crustacea Ostracoda) as models for evolu-
tionary studies of light and vision Multiple origins of bioluminescence
and extreme sexual dimorphism Hydrobiologia 538 179e192
Oakley TH Cunningham CW 2002 Molecular phylogenetic evidence for
the independent evolutionary origin of an arthropod compound eye
Proceedings of the National Academy of Sciences USA 99 1426e1430
Oakley TH Huber DR 2004 Differential expression of duplicated opsin
genes in two eye types of ostracod crustaceans Journal of Molecular Evo-
lution 59 239e249
Ohno S 1970 Evolution by Gene Duplication Springer New York
Osorio D 2007 Spam and the evolution of the flyrsquos eye Bioessays 29
111e115
Page RDM 1993 Parasites phylogeny and cospeciation International Jour-
nal for Parasitology 23 499e506
400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699

400 TH Oakley et al Arthropod Structure amp Development 36 (2007) 386e400
Page RDM Charleston MA 1998 Trees within trees phylogeny and his-
torical associations Trends in Ecology and Evolution 13 356e359
Pagel MD 1994 Detecting correlated evolution on phylogenies A general
method for the comparative analysis of discrete characters Proceedings
of the Royal Society London B 255 37e45
Panganiban G Irvine SM Lowe C Roehl H Corley LS Sherbon B
Grenier JK Fallon JF Kimble J Walker M Wray GA
Swalla BJ Martindale MQ Carroll SB 1997 The origin and evolu-
tion of animal appendages Proceedings of the National Academies of
Sciences USA 94 5162e5166
Paulus HF 1979 Eye structure and the monophyly of the Arthropoda In
Gupta AP (Ed) Arthropod Phylogeny Van Nostrand Reinhold New
York pp 299e383
Paulus HF 2000 Phylogeny of the Myriapoda-Crustacea-Insecta A new at-
tempt using photoreceptor structure Journal of Zoological Systematics and
Evolutionary Research 38 189e208
Piatigorsky J 2007 Gene Sharing and Evolution Harvard University Press
Cambridge MA
Piatigorsky J Wistow GJ 1989 Enzymecrystallins gene sharing as an
evolutionary strategy Cell 57 197e199
Plachetzki DC Oakley TH 2007 Key transitions during animal eye evo-
lution Novelty tree thinking co-option and co-duplication Integrative
and Comparative Biology 47 759e769
Plachetzki DC Serb JM Oakley TH 2005 New insights into the evolu-
tionary history of photoreceptor cells Trends in Ecology and Evolution 20
465e467
Pollock JA Benzer S 1988 Transcript localization of four opsin genes in
the three visual organs of Drosophila RH2 is ocellus specific Nature 333
779e782
Proulx SR Phillips PC 2006 Allelic divergence precedes and promotes
gene duplication Evolution 60 881e892
Regier JC Shultz JW 1997 Molecular phylogeny of the major arthropod
groups indicates polyphyly of crustaceans and a new hypothesis for the or-
igin of hexapods Molecular Biology and Evolution 14 902e913
Regier JC Shultz JW Kambic RE 2005 Pancrustacean phylogeny
hexapods are terrestrial crustaceans and maxillopods are not monophyletic
272 395e401
Reinke R Zipursky SL 1988 Cell-cell interaction in the Drosophila retina
the bride of sevenless gene is required in photoreceptor cell R8 for R7 cell
development Cell 55 321e220
Rice SH 1997 The analysis of ontogenetic trajectories when a change in
size or shape is not heterochrony Proceedings of the National Academies
of Sciences USA 94 907e912
Ronquist F 2004 Bayesian inference of character evolution Trends in Eco-
logy and Evolution 19 475e481
Sanderson MJ 2002 Estimating absolute rates of molecular evolution and
divergence times a penalized likelihood approach Molecular Biology
and Evolution 19 101e109
Sanderson MJ 2003 r8s inferring absolute rates of molecular evolution and
divergence times in the absence of a molecular clock Bioinformatics 19
301e302
Schlosser G Wagner GP 2004 Modularity in Development and Evolution
University of Chicago Press Chicago
Seaver EC 2003 Segmentation mono- or polyphyletic International Jour-
nal of Developmental Biology 47 583e595
Serb JM Oakley TH 2005 Hierarchical phylogenetics as a quantitative
analytical framework for evolutionary developmental biology Bioessays
27 1158e1166
Shermer M 2002 In Darwinrsquos Shadow The Life and Science of Alfred Rus-
sel Wallace Oxford University Press Oxford
Siveter DJ Sutton MD Briggs DE 2003 An ostracode crustacean with
soft parts from the Lower Silurian Science 302 1749e1751
Siveter DJ Siveter DJ Sutton MD Briggs DE 2007 Brood care
in a Silurian ostracod Proceedings of the Royal Society B 274
465e469
Smith WC Price DA Greenberg RM Battelle BA 1993 Opsins from
the lateral eyes and ocelli of the horseshoe crab Limulus polyphemus Pro-
ceedings of the National Academy of Sciences USA 90 6150e6154
Stavenga DG 1992 Eye regionalization and spectral tuning of retinal pig-
ments in insects Trends in Neuroscience 15 213e218
Stormer L 1952 Phylogeny and taxonomy of fossil horseshoe crabs Journal
of Paleontology 26 630e639
Strutt DI Mlodzik M 1996 The regulation of hedgehog and decapenta-
plegic during Drosophila eye imaginal disc development Mechanisms
of Development 58 39e50
Suzuki T Saigo K 2000 Transcriptional regulation of atonal required for
Drosophila larval eye development by concerted action of eyes absent
sine oculis and hedgehog signaling independent of fused kinase and cubitus
interruptus Development 127 1531e1540
Swofford DL Maddison WP 1987 Reconstructing ancestral character
states under Wagner parsimony Mathematical Biosciences 87 199e229
Tanaka G 2005 Morphological design and fossil record of the podocopid
ostracod naupliar eye Hydrobiologia 538 231e242
Taylor JS Raes J 2004 Duplication and divergence the evolution of new
genes and old ideas Annual Review of Genetics 38 615e643
Tepass U Harris KP 2007 Adherens junctions in Drosophila retinal mor-
phogenesis Trends in Cell Biology 17 26e35
Van Valen L 1979 Switchback evolution and photosynthesis in angiosperms
Evolutionary Theory 4 143e146
Wagner GP 2001 The Character Concept in Evolutionary Biology Aca-
demic Press San Diego
Wakayama N Embryonic development clarifies polyphyly in ostracod crusta-
ceans Journal of Zoology in press doi101111j1469-7998200700344x
West-Eberhard MJ 2003 Developmental Plasticity and Evolution Oxford
University Press Oxford
Wildt M Harzsch S 2002 A new look at an old visual system Structure
and development of the compound eyes and optic ganglia of the brine
shrimp Artemia salina Linnaeus 1758 (Branchiopoda Anostraca) Journal
of Neurobiology 52 117e132
Wolff T Ready DF 1993 Pattern formation in the Drosophila retina In
Bate M Martinez-Arias A (Eds) The Development of Drosophila mel-
anogaster Cold Spring Harbor Laboratory Press pp 1277e1325
Wray GA 2007 The evolutionary significance of cis-regulatory mutations
Nature Reviews Genetics 8 206e216
Wray GA Hahn MW Abouheif E Balhoff JP Pizer M
Rockman MV Romano LA 2003 The evolution of transcriptional reg-
ulation in eukaryotes Molecular Biology and Evolution 20 1377e1419
Wray GA Levinton JS Shapiro LH 1996 Molecular evidence for deep
Precambrian divergences among Metazoan phyla Science 274 568e573
Yamaguchi S Endo K 2003 Molecular phylogeny of Ostracoda (Crusta-
cea) inferred from 18S ribosomal DNA sequences implication for its
origin and diversification Marine Biology 143 23e38
Yu YL Khan J Khanna C Helman L Meltzer PS Merlino G 2004
Expression profiling identifies the cytoskeletal organizer ezrin and the de-
velopmental homeoprotein Six-1 as key metastatic regulators Nature Med-
icine 10 175e181
Zelditch ML Fink WL 1996 Heterochrony and heterotopy stability and
innovation in the evolution of form Paleobiology 22 241e254
Zelhof AC Hardy RW Becker A Zuker CS 2006 Transforming the
architecture of compound eyes Nature 443 696e699
Related Documents











