Functional Metabotropic Glutamate Receptors on Nuclei from Brain and Primary Cultured Striatal Neurons ROLE OF TRANSPORTERS IN DELIVERING LIGAND* Received for publication, February 16, 2005, and in revised form, June 8, 2005 Published, JBC Papers in Press, June 15, 2005, DOI 10.1074/jbc.M501775200 Yuh-Jiin I. Jong‡, Vikas Kumar‡, Ann E. Kingston§, Carmelo Romano¶, and Karen L. O’Malley‡ From the ‡Departments of Anatomy and Neurobiology and ¶Ophthalmology, Washington University School of Medicine, St. Louis, Missouri 63110 and §Eli Lilly & Co. Ltd., Indianapolis, Indiana 46285-1533 G-protein-coupled receptors are well known for con- verting an extracellular signal into an intracellular re- sponse. Here we showed that the metabotropic gluta- mate receptor 5 (mGlu5) plays a dynamic intracellular role in signal transduction. Activation of endogenously expressed mGlu5 on striatal nuclear membranes leads to rapid, sustained calcium (Ca 2 ) responses within the nucleoplasm that can be blocked by receptor-specific antagonists. Extracellular ligands such as glutamate and quisqualate reach nuclear receptors via both sodium- dependent transporters and cystine glutamate exchang- ers. Inhibition of either transport system blocks radio- labeled agonist uptake as well as agonist-induced nuclear Ca 2 changes. Impermeable antagonists like LY393053 and LY367366 not only blocked [ 3 H]quisqual- ate binding but also prevented nontransported agonists such as (RS)-3,5-dihydroxyphenylglycine from inducing intracellular Ca 2 changes in heterologous cells. In con- trast, neither LY compound prevented quisqualate or glutamate from activating intracellular receptors lead- ing to Ca 2 responses. Inasmuch as Ca 2 can enter the nucleoplasm via the nuclear pore complex or from the nuclear lumen, the presence of nuclear mGlu5 receptors appeared to amplify the latter process generating a faster nuclear response in heterologous cells. In isolated striatal nuclei, nuclear receptor activation results in the de novo appearance of phosphorylated CREB protein. Thus, activation of nuclear mGlu5 receptors initiates a signaling cascade that is known to alter gene transcrip- tion and regulate many paradigms of synaptic plasticity. These studies demonstrated that mGlu5 receptors play a dynamic role in signaling both on and off the plasma membrane. The structure and function of G-protein-coupled receptors have received intense scrutiny over the past decades. These studies point to a dynamic environment in which receptors are not static but rather move on and off the plasma membrane according to environmental stimuli, specific targeting informa- tion, protein-protein interactions, etc. In this model, intracel- lular receptors are considered transitional, i.e. receptors that are either ready to be inserted into the plasma membrane or that have just been sequestered from such a site. Emerging data, however, suggest that some intracellular receptors may have intracellular functions as well. For example, a number of G-protein-coupled receptors such as the apelin, angiotensin AT1 and ATII, bradykinin B2, and lysophosphatidate LP1 re- ceptors have been localized within the nucleoplasm itself (1–3). In contrast, prostaglandin E 2 receptors have been found on the nuclear envelope together with their ligand-generating en- zymes (4, 5). Similarly, endothelin receptors A and B are also localized to the perinuclear region of cardiac ventricular myo- cytes where they mediate nuclear Ca 2 levels and activate nuclear protein kinases (6). Finally, we have shown that the metabotropic glutamate receptor, mGlu5, 1 can be expressed on nuclear membranes where it can couple with endogenous sig- naling components to induce changes in nuclear Ca 2 (7). Be- cause most studies investigating the properties of nuclear re- ceptors have been performed in heterologous cell types with overexpressed receptors, the question arises as to whether such phenomena are physiologically relevant and, in the case of mGlu5, how a ligand such as glutamate has access to this receptor. Widely expressed throughout the central nervous system, mGlu5 receptors play an important role in modulating neuro- nal excitability during critical processes such as development, synaptic plasticity, and learning. Typically, mGlu5 receptors couple to G q/11 leading to phosphoinositide signaling and re- lease of calcium from intracellular stores (8). In heterologous cell types, mGlu5 activation leads to marked oscillatory re- sponses (9), whereas more variable responses are seen in physiological environments, ranging from oscillations to ini- tial transient increases with sustained plateaus (10 –13). Re- sponses vary depending upon the neuronal subtype express- ing the receptor and/or with the level of agonist stimulation (12). Numerous morphological and physiological studies have sug- gested that mGlu5 receptors are enriched in the striatum where they are expressed on postsynaptic neurons, subsets of interneu- rons, as well as at presynaptic sites (14). Both at the light and electron microscopic level, large amounts of mGlu5 are intracel- lularly localized (40 –70%) where receptors are predominantly * This work was supported by National Institutes of Health Grants MH57817 and MH069646. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. To whom correspondence should be addressed: Dept. of Anatomy and Neurobiology, Washington University School of Medicine, 660 S. Euclid Ave., St. Louis, MO 63110. Tel.: 314-362-7087; Fax: 314-362- 3446; E-mail: [email protected]. 1 The abbreviations used are: mGlu5, metabotropic glutamate recep- tor 5; CREB, cAMP-response element-binding protein; xCT, cystine/ glutamate exchanger; EAAT, excitatory amino acid transporter; P1, postnatal day 1; DIV9, days in vitro 9; CPCCOEt, 7-(hydroxyimino)cy- clopropan[b]chromen-1a-carboxylate ethyl ester; MPEP, 2-methyl-6- (phenylethynyl)-pyridine; TBOA, threo--benzyloxyaspartate; DHPG, (RS)-3,5-dihydroxyphenylglycine; ER, endoplasmic reticulum; BAPTA, 1,2-bis-(2-aminophenoxy)ethane-N,N,N,N-tetraacetic acid; AMPA, - amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; HEK, human em- bryonic kidney cells; GABA, -aminobutyric acid; NMDG, N-methyl-D- glucamine; IP 3 , inositol 1,4,5-trisphosphate; HA, hemagglutinin. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 34, Issue of August 26, pp. 30469 –30480, 2005 © 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. This paper is available on line at http://www.jbc.org 30469 by guest on June 19, 2016 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Functional Metabotropic Glutamate Receptors on Nuclei from Brainand Primary Cultured Striatal NeuronsROLE OF TRANSPORTERS IN DELIVERING LIGAND*
Received for publication, February 16, 2005, and in revised form, June 8, 2005Published, JBC Papers in Press, June 15, 2005, DOI 10.1074/jbc.M501775200
Yuh-Jiin I. Jong‡, Vikas Kumar‡, Ann E. Kingston§, Carmelo Romano¶, and Karen L. O’Malley‡�
From the ‡Departments of Anatomy and Neurobiology and ¶Ophthalmology, Washington University School of Medicine,St. Louis, Missouri 63110 and §Eli Lilly & Co. Ltd., Indianapolis, Indiana 46285-1533
G-protein-coupled receptors are well known for con-verting an extracellular signal into an intracellular re-sponse. Here we showed that the metabotropic gluta-mate receptor 5 (mGlu5) plays a dynamic intracellularrole in signal transduction. Activation of endogenouslyexpressed mGlu5 on striatal nuclear membranes leadsto rapid, sustained calcium (Ca2�) responses within thenucleoplasm that can be blocked by receptor-specificantagonists. Extracellular ligands such as glutamateandquisqualatereachnuclearreceptorsviabothsodium-dependent transporters and cystine glutamate exchang-ers. Inhibition of either transport system blocks radio-labeled agonist uptake as well as agonist-inducednuclear Ca2� changes. Impermeable antagonists likeLY393053 and LY367366 not only blocked [3H]quisqual-ate binding but also prevented nontransported agonistssuch as (RS)-3,5-dihydroxyphenylglycine from inducingintracellular Ca2� changes in heterologous cells. In con-trast, neither LY compound prevented quisqualate orglutamate from activating intracellular receptors lead-ing to Ca2� responses. Inasmuch as Ca2� can enter thenucleoplasm via the nuclear pore complex or from thenuclear lumen, the presence of nuclear mGlu5 receptorsappeared to amplify the latter process generating afaster nuclear response in heterologous cells. In isolatedstriatal nuclei, nuclear receptor activation results in thede novo appearance of phosphorylated CREB protein.Thus, activation of nuclear mGlu5 receptors initiates asignaling cascade that is known to alter gene transcrip-tion and regulate many paradigms of synaptic plasticity.These studies demonstrated that mGlu5 receptors play adynamic role in signaling both on and off the plasmamembrane.
The structure and function of G-protein-coupled receptorshave received intense scrutiny over the past decades. Thesestudies point to a dynamic environment in which receptors arenot static but rather move on and off the plasma membraneaccording to environmental stimuli, specific targeting informa-tion, protein-protein interactions, etc. In this model, intracel-lular receptors are considered transitional, i.e. receptors thatare either ready to be inserted into the plasma membrane or
that have just been sequestered from such a site. Emergingdata, however, suggest that some intracellular receptors mayhave intracellular functions as well. For example, a number ofG-protein-coupled receptors such as the apelin, angiotensinAT1 and ATII, bradykinin B2, and lysophosphatidate LP1 re-ceptors have been localized within the nucleoplasm itself (1–3).In contrast, prostaglandin E2 receptors have been found on thenuclear envelope together with their ligand-generating en-zymes (4, 5). Similarly, endothelin receptors A and B are alsolocalized to the perinuclear region of cardiac ventricular myo-cytes where they mediate nuclear Ca2� levels and activatenuclear protein kinases (6). Finally, we have shown that themetabotropic glutamate receptor, mGlu5,1 can be expressed onnuclear membranes where it can couple with endogenous sig-naling components to induce changes in nuclear Ca2� (7). Be-cause most studies investigating the properties of nuclear re-ceptors have been performed in heterologous cell types withoverexpressed receptors, the question arises as to whether suchphenomena are physiologically relevant and, in the case ofmGlu5, how a ligand such as glutamate has access to thisreceptor.
Widely expressed throughout the central nervous system,mGlu5 receptors play an important role in modulating neuro-nal excitability during critical processes such as development,synaptic plasticity, and learning. Typically, mGlu5 receptorscouple to Gq/11 leading to phosphoinositide signaling and re-lease of calcium from intracellular stores (8). In heterologouscell types, mGlu5 activation leads to marked oscillatory re-sponses (9), whereas more variable responses are seen inphysiological environments, ranging from oscillations to ini-tial transient increases with sustained plateaus (10–13). Re-sponses vary depending upon the neuronal subtype express-ing the receptor and/or with the level of agonist stimulation(12).
Numerous morphological and physiological studies have sug-gested that mGlu5 receptors are enriched in the striatum wherethey are expressed on postsynaptic neurons, subsets of interneu-rons, as well as at presynaptic sites (14). Both at the light andelectron microscopic level, large amounts of mGlu5 are intracel-lularly localized (40–70%) where receptors are predominantly
* This work was supported by National Institutes of Health GrantsMH57817 and MH069646. The costs of publication of this article weredefrayed in part by the payment of page charges. This article musttherefore be hereby marked “advertisement” in accordance with 18U.S.C. Section 1734 solely to indicate this fact.
� To whom correspondence should be addressed: Dept. of Anatomyand Neurobiology, Washington University School of Medicine, 660 S.Euclid Ave., St. Louis, MO 63110. Tel.: 314-362-7087; Fax: 314-362-3446; E-mail: [email protected].
1 The abbreviations used are: mGlu5, metabotropic glutamate recep-tor 5; CREB, cAMP-response element-binding protein; xCT, cystine/glutamate exchanger; EAAT, excitatory amino acid transporter; P1,postnatal day 1; DIV9, days in vitro 9; CPCCOEt, 7-(hydroxyimino)cy-clopropan[b]chromen-1a-carboxylate ethyl ester; MPEP, 2-methyl-6-(phenylethynyl)-pyridine; TBOA, threo-�-benzyloxyaspartate; DHPG,(RS)-3,5-dihydroxyphenylglycine; ER, endoplasmic reticulum; BAPTA,1,2-bis-(2-aminophenoxy)ethane-N,N,N�,N�-tetraacetic acid; AMPA, �-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; HEK, human em-bryonic kidney cells; GABA, �-aminobutyric acid; NMDG, N-methyl-D-glucamine; IP3, inositol 1,4,5-trisphosphate; HA, hemagglutinin.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 34, Issue of August 26, pp. 30469–30480, 2005© 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 30469
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from

associated with the endoplasmic reticulum (ER) or nuclear mem-branes (14). Immunofluorescent staining of embryonic or neona-tal striatal cultures shows widespread mGlu5 expression; agonistexposure elicits an intracellular rise in Ca2� typified by an initialincrease followed by a sustained plateau. This response can beblocked by application of the mGlu5-specific antagonist MPEP(15). Moreover, activation of striatal mGlu5 receptors leads tophosphorylation of the CREB transcription factor and subse-quent activation of several immediate early genes (16). Thus,neonatal striatal cultures seem to be an ideal preparation to testthe notion that endogenous mGlu5 receptors are expressed onnuclear membranes where they are coupled to signaling mecha-nisms within the nuclear membrane.
For intracellular mGlu5 receptors to be of functional rele-vance, there must be a way to activate them. By using taggedreceptor molecules, we have shown previously that the mGlu5receptor topology is oriented such that ligand binding domainsare within the lumen of the nuclear envelope. Thus, a ligandsource from the extracellular milieu must traverse both theplasma membrane as well as an intracellular membrane. Intheory, some combination of plasma membrane and intracellu-lar transporter could transfer glutamate into the nuclear lu-men. For example, there are five sodium-dependent glutamatetransporter proteins that are present on the plasma membraneof glial and neuronal cells as well as many peripheral tissues(17). Many other glutamate carriers exist, however, includingthe cystine-glutamate exchanger. Ubiquitously expressedthroughout the body, the exchanger is a sodium-independent,anionic amino acid transporter composed of two separate pro-teins, xCT and 4F2 (18). The former confers substrate specific-ity, and the latter is common to many amino acid transporters.Because the mGlu5 agonist, quisqualate, is a substrate for thexCT exchanger (19), it too is a plausible candidate for intracel-lular ligand delivery.
Here we show that mGlu5 receptors are expressed on the cellsurface and intracellular membranes of neonatal striatal neu-rons. Activation of nuclear receptors leads to increased nucle-oplasmic Ca2�, which can be blocked by receptor-specific an-tagonists. Ligand delivery is via both sodium-dependent and-independent mechanisms. Impermeable antagonists can blockcellular responses to agonists that are not transported intocells but cannot block those that are. Finally, agonist-inducednuclear receptor activation leads to phosphorylation of CREB.Taken together, these data challenge existing paradigms bysuggesting a new mode of intracellular signal transductionmediated in situ by G-protein-coupled receptors.
EXPERIMENTAL PROCEDURES
Materials—2-Amino-2-(4-carboxyphenyl)-3-(9H-thioxanthen-9-yl) propa-noic acid (LY367366) and 2-amino-2-(3-cis and trans-carboxycyclobutyl)-3-(9H-thioxanthen-9-yl)propionic acid (LY393053) were obtained from Lilly.Quisqualate and (RS)-3,5-dihydroxyphenylglycine (DHPG) were purchasedfrom Tocris Cookson Inc. (Ellisville, MO). Unless otherwise indicated allother chemicals were from Sigma.
Cell Culture and Immunocytochemistry—The mGlu5/HEK stable cellline was generated and maintained as described (20). The mGlu5/GAB-ABR1 chimera (mG5C1) as well as its mutant (mG5C1ASA) were kind giftsfrom Dr. J. P. Pin (Institute of Functional Genomics, CNRS, France).Primary striatal neuronal cultures using neonatal 1-day-old rat pupswere prepared and maintained as detailed by Mao and Wang (15). Fixa-tion, blocking, and antibody incubation were as described previously (20).Primary antibodies included affinity-purified polyclonal anti-C-terminalmGlu5 (1:300) (21), monoclonal anti-lamin B2 (1:100; Zymed LaboratoriesInc.), monoclonal anti-GABA (1:1000; Sigma), polyclonal anti-phospho-CREB antibody (1:3000; Cell Signaling Technology, Inc., Beverly, MA),polyclonal anti-EAAT3 (1:1000; Chemicon International, Temecula, CA),monoclonal anti-Na�,K�-ATPase (�-6F, 1:1000; Developmental StudiesHybridoma Bank, University of Iowa, Iowa City), and polyclonal anti-quisqualate antibody (1:250; Dr. J. Koerner, University of Minnesota).Secondary antibodies included goat anti-rabbit and/or anti-mouse Cy3
(both at 1:300; Jackson ImmunoResearch, West Grove, PA) and goatanti-mouse Alexa 488 (1:300; Molecular Probes, Eugene, OR) or goatanti-rabbit Alexa 488 (1:750; Molecular Probes).
Tissue Isolation, Subcellular Fractionation, and Western Blot Anal-ysis—Postnatal day 1, 3, or 10 rodent striata were isolated and pro-cessed by mincing on ice with a razor blade followed by resuspension in20 volumes of Buffer “A” medium containing 2.0 mM MgCl2, 25 mM KCl,10 mM HEPES (pH 7.5), and protease inhibitors (Complete Tablets;Roche Applied Science). Cells were homogenized, and nuclei andplasma membranes were prepared as described previously (7). Aliquotsfrom each fraction were used for gel electrophoresis as well as mem-brane binding. Protein concentrations were determined using the Brad-ford assay (Bio-Rad). Fractionated proteins were separated by SDS-PAGE, blotted, and probed with anti-mGlu5 (1:1500) (21), monoclonalanti-lamin B2 (1:1000), or monoclonal anti-Na�,K�-ATPase. Anti-EAAT3 and xCT antibodies (Dr. J. Rothstein, The Johns Hopkins Uni-versity) were used at dilutions of 1:200 and 1:150, respectively. Ahorseradish peroxidase conjugated with goat anti-rabbit IgG (1:2000;Cell Signaling Technology, Inc.) or anti-mouse IgG (1:2500; Sigma) wasused in conjunction with enhanced chemiluminescence (Amersham Bio-sciences) to detect the signal. Densitometric analyses of mGlu5 proteinswere performed by using the Storm 860 Imager (Amersham Bio-sciences) together with associated software.
Extra- and Intracellular Buffers—Ca2� and uptake measurementsperformed on intact cells were done in Krebs-Ringer solution (KRS,containing the following (in mM): 137 NaCl, 5.1 KCl, 0.77 KH2PO4, 0.71MgSO4�7H2O, 1.1 CaCl2, 10 D-glucose, and 10 HEPES). To achievechloride-free conditions (Cl�-free), the buffer contained the following (inmM): 130 sodium gluconate, 5 potassium gluconate, 1.1 calcium gluco-nate, 0.77 KH2PO4, 0.71 MgSO4�7H2O, 10 D-glucose, and 10 HEPES. Toachieve sodium-free conditions (Na�-free) the buffer contained the fol-lowing (in mM): 137.5 choline chloride, 5.36 KCl, 0.77 KH2PO4, 0.71MgSO4�7H2O, 1.1 CaCl2, 10 D-glucose, and 10 HEPES. Sodium- andchloride-free conditions (Na�,Cl�-free) were achieved by using a buffercontaining the following (in mM): 274 N-methyl-D-glucamine, 5 potas-sium gluconate, 1.1 calcium gluconate, 0.77 KH2PO4, 0.71MgSO4�7H2O, 10 D-glucose, and 10 HEPES (pH 7.4 adjusted with 50%gluconic acid). Experiments using purified nuclei were done using in-tracellular buffer conditions. Intracellular buffers contained the follow-ing (in mM): 125 KCl, 2 KH2PO4, 2 MgCl2, 0.3 CaCl2, 10 D-glucose, 1ATP, and 40 HEPES (pH 7.0).
3H-Labeled Agonist Uptake—[3H]Quisqualate (26.0 Ci/mmol) andL-[3H]glutamate (43.0 Ci/mmol) were obtained from Amersham Bio-sciences. In some cases, radiolabeled agonists (3.25 �Ci/ml [3H]quis-qualate or 43.08 �Ci/ml L-[3H]glutamate) were added together withunlabeled drug to achieve a total quisqualate concentration of 120 and500 nM, 1 or 10 �M, or a total concentration of 10 �M or 1 mM glutamate.The mGlu5/HEK stable cell line (5 � 104 cells/well) or primary striatalneurons (3 � 105 cells/well) were cultured at 37 °C for 3 or 9 days,respectively, before use. Purified nuclei were washed three times in theappropriate buffer and then incubated at 37 °C in the presence orabsence of various concentrations of L-cystine or TBOA for 15 minbefore adding labeled agonist. Uptake was terminated after 15 min.Samples were rapidly rinsed three times with ice-cold phosphate-buff-ered saline, solubilized in 150 �l of 1% Triton X-100/phosphate-bufferedsaline, and then analyzed by liquid scintillation.
[3H]Quisqualate Binding Assay—Plasma membrane fractions wereprepared as described above from mGlu5-stable HEK cells. Membraneswere subsequently washed three times in 2 mM EDTA, 2 mM HEPES(pH 7.5), with protease inhibitors followed by centrifugation at 17,000 �g for 35 min. The final pellets were resuspended in buffer containing 40mM HEPES (pH 7.5), 2.5 mM Ca2�, and protease inhibitors. Binding wasperformed as described (20).
Fluorescent Measurements of Intracellular Ca2�—For whole cellmeasurements, dissociated striatal neurons were grown on 35-mmdishes with glass grids (1.2 � 105cells/grid), washed with Neurobasalmedium (NB; Invitrogen), and incubated with 5 �M Oregon Green 488BAPTA-AM, 0.00l% pluronic acid (Molecular Probes, Eugene, OR) inNB for 30 min at 37 °C. Cells were washed three times with NB andobserved using a laser confocal microscope (Olympus BX 50WI) with anOlympus LUMPlanFl/lR 40�/0.80w or 60�/0.90w objectives. The realtime images were captured by an Olympus Fluoview FVX confocal laserscanning system using Fluoview 2.0 acquisition software. Images wereconverted to TIF files for processing with MetaMorph (version 5.0.7)Professional Image Analysis software, produced by Universal Imaging.Drugs at 100� concentrations were added to the side of the dish andallowed to diffuse over the cells at room temperature. Quisqualate wasadded at a final concentration of 10 �M together with 5 �M GYKI53655
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium30470
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
(Research Biochemicals Inc., Natick, MA) and 20 �M 7-(hydroxyimino)-cyclopropan[b]chromen-1a-carboxylate ethyl ester (CPCCOEt; Tocris).The mGlu5-specific antagonist 2-methyl-6-(phenylethynyl)pyridine(MPEP; Tocris) was added at 1 �M.
Initiation of Ca2� responses was analyzed using MATLAB® 7.0.4.Specifically, agonist-mediated initiation was defined as the first detecta-ble change in fluorescence 3 S.D. above the mean of 5–6 s of base-linefluorescence.
Fluorescent Measurements of Ca2� in Isolated Nuclei—To measureCa2� changes in individual nuclei, nuclei were prepared as describedabove using Buffer A medium containing 125 mM sucrose. Isolatednuclei, plated on poly-D-lysine-coated 35-mm dishes with glass grids,were loaded with 20 �M Oregon Green 488 BAPTA-1AM, 0.00l% plu-ronic acid prepared in intracellular buffer together with 1 mM CaCl2, 1mM ATP, and protease inhibitors for 20–30 min on ice. After loading,nuclei were washed three times with the same buffer minus the Ca2�
indicator and imaged immediately thereafter. Drug treatments wereidentical to whole cells.
Reverse Transcription-PCR—RNA was extracted from P10 striataltissue with Trizol (Invitrogen) according to the manufacturer’s instruc-tions, and reverse transcription was performed. Equal amounts of RNAwere used for each reaction along with transporter-specific primersdesigned to cross intron junctions obtained from NCBI data bases.Resulting cDNA transcripts were amplified by PCR, fractionated byPAGE, and analyzed with Vistra Green (Amersham Biosciences) andquantitative fluorimaging.
RESULTS
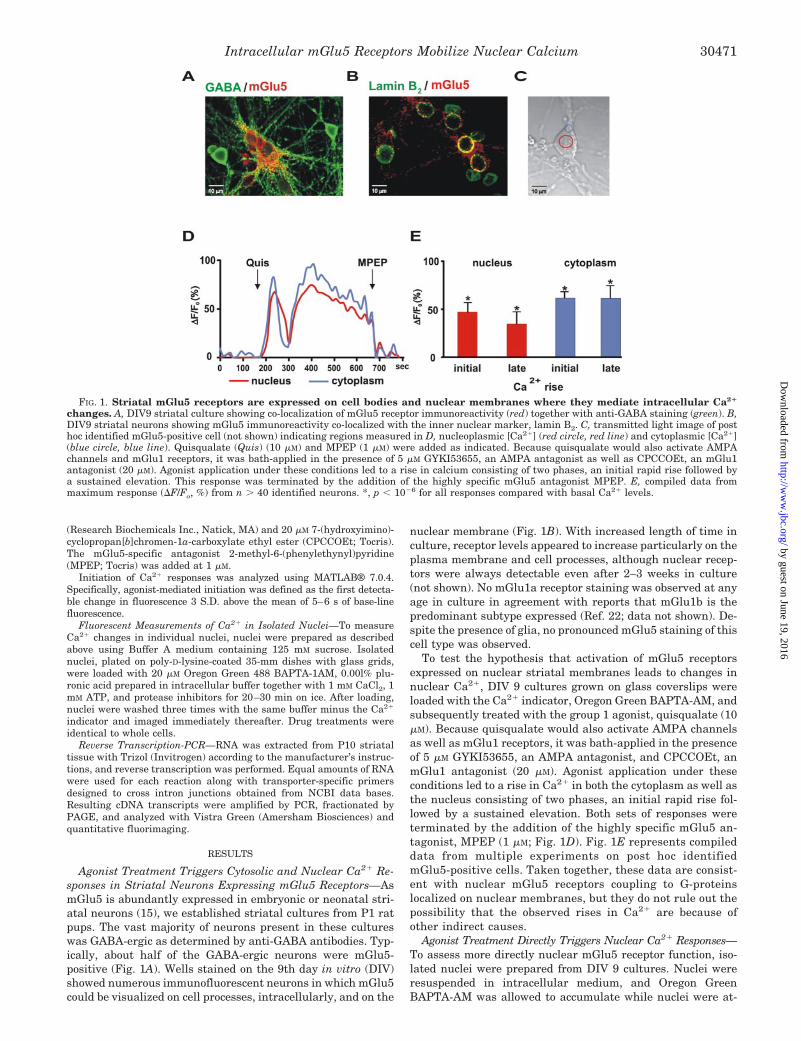
Agonist Treatment Triggers Cytosolic and Nuclear Ca2� Re-sponses in Striatal Neurons Expressing mGlu5 Receptors—AsmGlu5 is abundantly expressed in embryonic or neonatal stri-atal neurons (15), we established striatal cultures from P1 ratpups. The vast majority of neurons present in these cultureswas GABA-ergic as determined by anti-GABA antibodies. Typ-ically, about half of the GABA-ergic neurons were mGlu5-positive (Fig. 1A). Wells stained on the 9th day in vitro (DIV)showed numerous immunofluorescent neurons in which mGlu5could be visualized on cell processes, intracellularly, and on the
nuclear membrane (Fig. 1B). With increased length of time inculture, receptor levels appeared to increase particularly on theplasma membrane and cell processes, although nuclear recep-tors were always detectable even after 2–3 weeks in culture(not shown). No mGlu1a receptor staining was observed at anyage in culture in agreement with reports that mGlu1b is thepredominant subtype expressed (Ref. 22; data not shown). De-spite the presence of glia, no pronounced mGlu5 staining of thiscell type was observed.
To test the hypothesis that activation of mGlu5 receptorsexpressed on nuclear striatal membranes leads to changes innuclear Ca2�, DIV 9 cultures grown on glass coverslips wereloaded with the Ca2� indicator, Oregon Green BAPTA-AM, andsubsequently treated with the group 1 agonist, quisqualate (10�M). Because quisqualate would also activate AMPA channelsas well as mGlu1 receptors, it was bath-applied in the presenceof 5 �M GYKI53655, an AMPA antagonist, and CPCCOEt, anmGlu1 antagonist (20 �M). Agonist application under theseconditions led to a rise in Ca2� in both the cytoplasm as well asthe nucleus consisting of two phases, an initial rapid rise fol-lowed by a sustained elevation. Both sets of responses wereterminated by the addition of the highly specific mGlu5 an-tagonist, MPEP (1 �M; Fig. 1D). Fig. 1E represents compileddata from multiple experiments on post hoc identifiedmGlu5-positive cells. Taken together, these data are consist-ent with nuclear mGlu5 receptors coupling to G-proteinslocalized on nuclear membranes, but they do not rule out thepossibility that the observed rises in Ca2� are because ofother indirect causes.
Agonist Treatment Directly Triggers Nuclear Ca2� Responses—To assess more directly nuclear mGlu5 receptor function, iso-lated nuclei were prepared from DIV 9 cultures. Nuclei wereresuspended in intracellular medium, and Oregon GreenBAPTA-AM was allowed to accumulate while nuclei were at-
FIG. 1. Striatal mGlu5 receptors are expressed on cell bodies and nuclear membranes where they mediate intracellular Ca2�
changes. A, DIV9 striatal culture showing co-localization of mGlu5 receptor immunoreactivity (red) together with anti-GABA staining (green). B,DIV9 striatal neurons showing mGlu5 immunoreactivity co-localized with the inner nuclear marker, lamin B2. C, transmitted light image of posthoc identified mGlu5-positive cell (not shown) indicating regions measured in D, nucleoplasmic [Ca2�] (red circle, red line) and cytoplasmic [Ca2�](blue circle, blue line). Quisqualate (Quis) (10 �M) and MPEP (1 �M) were added as indicated. Because quisqualate would also activate AMPAchannels and mGlu1 receptors, it was bath-applied in the presence of 5 �M GYKI53655, an AMPA antagonist as well as CPCCOEt, an mGlu1antagonist (20 �M). Agonist application under these conditions led to a rise in calcium consisting of two phases, an initial rapid rise followed bya sustained elevation. This response was terminated by the addition of the highly specific mGlu5 antagonist MPEP. E, compiled data frommaximum response (�F/Fo, %) from n � 40 identified neurons. *, p � 10�6 for all responses compared with basal Ca2� levels.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium 30471
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
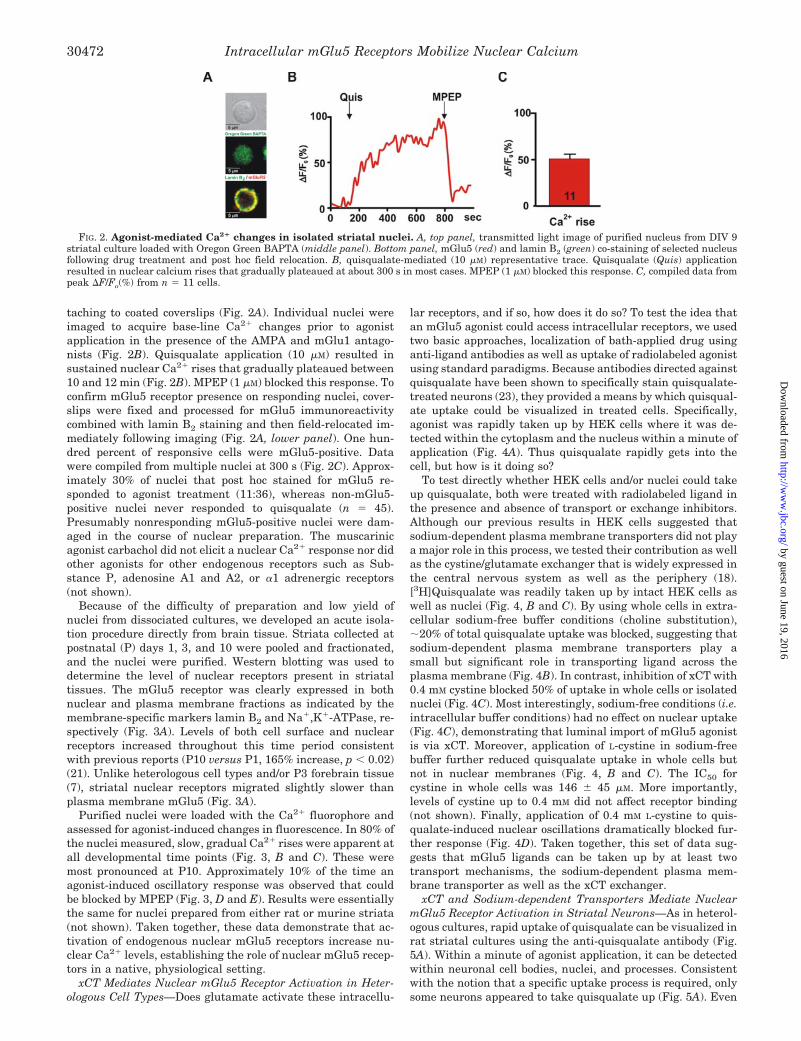
taching to coated coverslips (Fig. 2A). Individual nuclei wereimaged to acquire base-line Ca2� changes prior to agonistapplication in the presence of the AMPA and mGlu1 antago-nists (Fig. 2B). Quisqualate application (10 �M) resulted insustained nuclear Ca2� rises that gradually plateaued between10 and 12 min (Fig. 2B). MPEP (1 �M) blocked this response. Toconfirm mGlu5 receptor presence on responding nuclei, cover-slips were fixed and processed for mGlu5 immunoreactivitycombined with lamin B2 staining and then field-relocated im-mediately following imaging (Fig. 2A, lower panel). One hun-dred percent of responsive cells were mGlu5-positive. Datawere compiled from multiple nuclei at 300 s (Fig. 2C). Approx-imately 30% of nuclei that post hoc stained for mGlu5 re-sponded to agonist treatment (11:36), whereas non-mGlu5-positive nuclei never responded to quisqualate (n � 45).Presumably nonresponding mGlu5-positive nuclei were dam-aged in the course of nuclear preparation. The muscarinicagonist carbachol did not elicit a nuclear Ca2� response nor didother agonists for other endogenous receptors such as Sub-stance P, adenosine A1 and A2, or �1 adrenergic receptors(not shown).
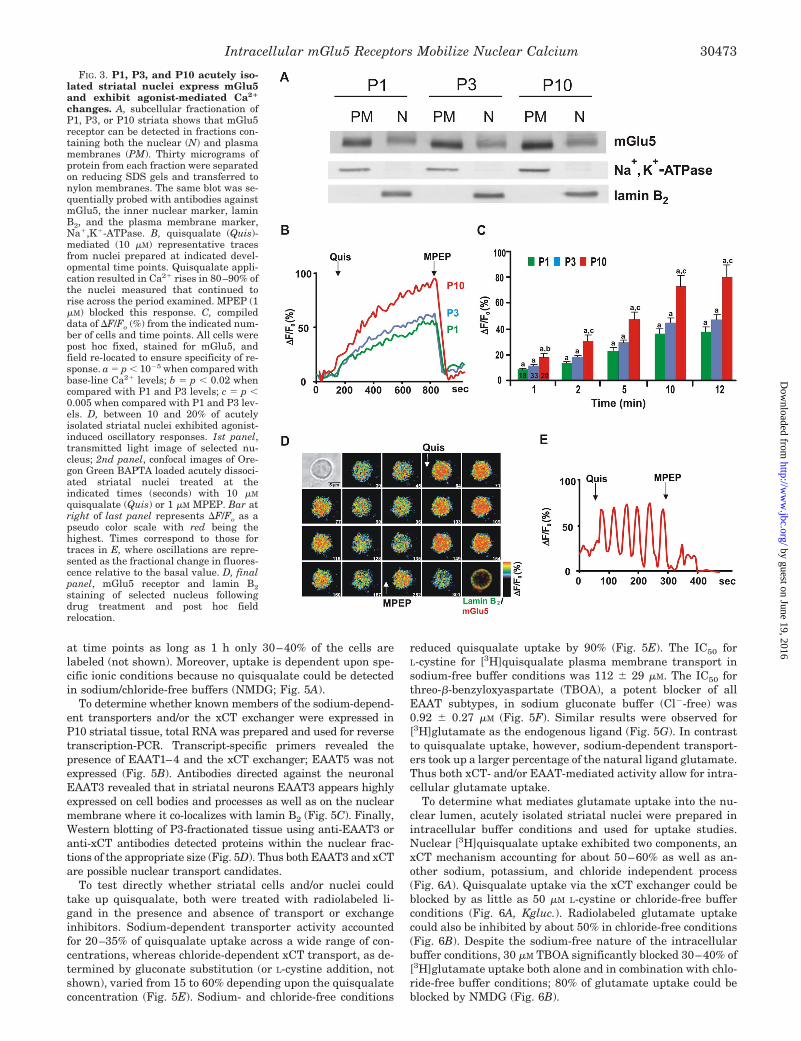
Because of the difficulty of preparation and low yield ofnuclei from dissociated cultures, we developed an acute isola-tion procedure directly from brain tissue. Striata collected atpostnatal (P) days 1, 3, and 10 were pooled and fractionated,and the nuclei were purified. Western blotting was used todetermine the level of nuclear receptors present in striataltissues. The mGlu5 receptor was clearly expressed in bothnuclear and plasma membrane fractions as indicated by themembrane-specific markers lamin B2 and Na�,K�-ATPase, re-spectively (Fig. 3A). Levels of both cell surface and nuclearreceptors increased throughout this time period consistentwith previous reports (P10 versus P1, 165% increase, p � 0.02)(21). Unlike heterologous cell types and/or P3 forebrain tissue(7), striatal nuclear receptors migrated slightly slower thanplasma membrane mGlu5 (Fig. 3A).
Purified nuclei were loaded with the Ca2� fluorophore andassessed for agonist-induced changes in fluorescence. In 80% ofthe nuclei measured, slow, gradual Ca2� rises were apparent atall developmental time points (Fig. 3, B and C). These weremost pronounced at P10. Approximately 10% of the time anagonist-induced oscillatory response was observed that couldbe blocked by MPEP (Fig. 3, D and E). Results were essentiallythe same for nuclei prepared from either rat or murine striata(not shown). Taken together, these data demonstrate that ac-tivation of endogenous nuclear mGlu5 receptors increase nu-clear Ca2� levels, establishing the role of nuclear mGlu5 recep-tors in a native, physiological setting.
xCT Mediates Nuclear mGlu5 Receptor Activation in Heter-ologous Cell Types—Does glutamate activate these intracellu-
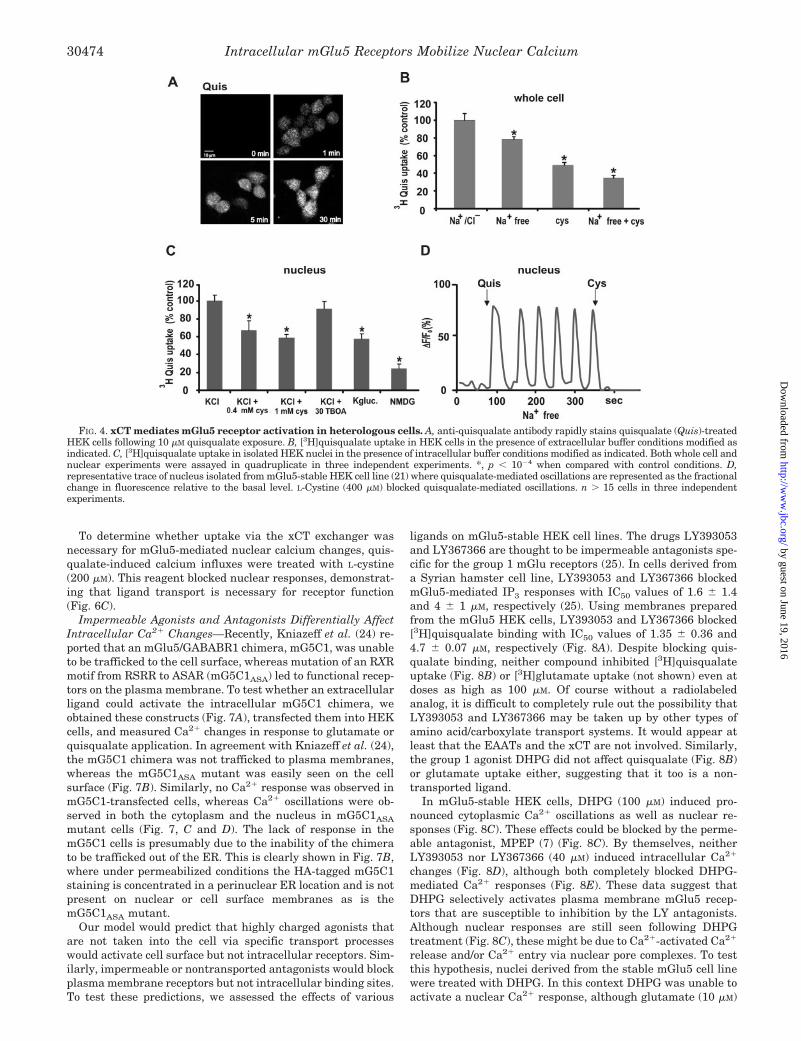
lar receptors, and if so, how does it do so? To test the idea thatan mGlu5 agonist could access intracellular receptors, we usedtwo basic approaches, localization of bath-applied drug usinganti-ligand antibodies as well as uptake of radiolabeled agonistusing standard paradigms. Because antibodies directed againstquisqualate have been shown to specifically stain quisqualate-treated neurons (23), they provided a means by which quisqual-ate uptake could be visualized in treated cells. Specifically,agonist was rapidly taken up by HEK cells where it was de-tected within the cytoplasm and the nucleus within a minute ofapplication (Fig. 4A). Thus quisqualate rapidly gets into thecell, but how is it doing so?
To test directly whether HEK cells and/or nuclei could takeup quisqualate, both were treated with radiolabeled ligand inthe presence and absence of transport or exchange inhibitors.Although our previous results in HEK cells suggested thatsodium-dependent plasma membrane transporters did not playa major role in this process, we tested their contribution as wellas the cystine/glutamate exchanger that is widely expressed inthe central nervous system as well as the periphery (18).[3H]Quisqualate was readily taken up by intact HEK cells aswell as nuclei (Fig. 4, B and C). By using whole cells in extra-cellular sodium-free buffer conditions (choline substitution),20% of total quisqualate uptake was blocked, suggesting thatsodium-dependent plasma membrane transporters play asmall but significant role in transporting ligand across theplasma membrane (Fig. 4B). In contrast, inhibition of xCT with0.4 mM cystine blocked 50% of uptake in whole cells or isolatednuclei (Fig. 4C). Most interestingly, sodium-free conditions (i.e.intracellular buffer conditions) had no effect on nuclear uptake(Fig. 4C), demonstrating that luminal import of mGlu5 agonistis via xCT. Moreover, application of L-cystine in sodium-freebuffer further reduced quisqualate uptake in whole cells butnot in nuclear membranes (Fig. 4, B and C). The IC50 forcystine in whole cells was 146 45 �M. More importantly,levels of cystine up to 0.4 mM did not affect receptor binding(not shown). Finally, application of 0.4 mM L-cystine to quis-qualate-induced nuclear oscillations dramatically blocked fur-ther response (Fig. 4D). Taken together, this set of data sug-gests that mGlu5 ligands can be taken up by at least twotransport mechanisms, the sodium-dependent plasma mem-brane transporter as well as the xCT exchanger.
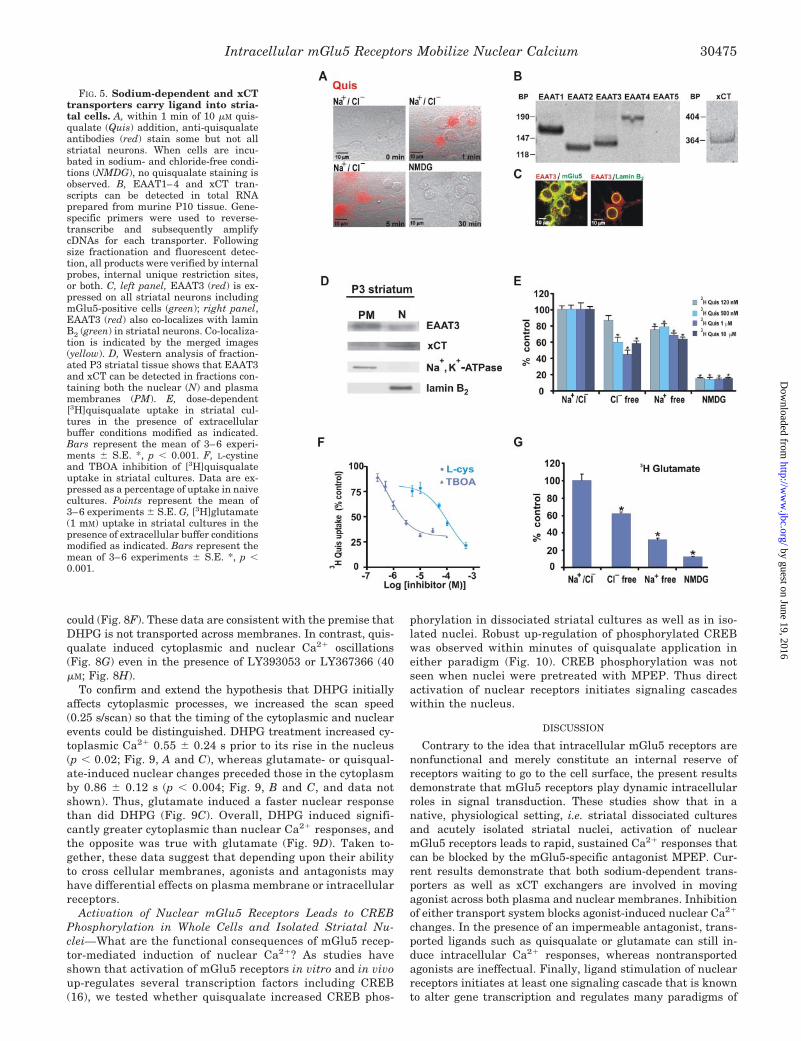
xCT and Sodium-dependent Transporters Mediate NuclearmGlu5 Receptor Activation in Striatal Neurons—As in heterol-ogous cultures, rapid uptake of quisqualate can be visualized inrat striatal cultures using the anti-quisqualate antibody (Fig.5A). Within a minute of agonist application, it can be detectedwithin neuronal cell bodies, nuclei, and processes. Consistentwith the notion that a specific uptake process is required, onlysome neurons appeared to take quisqualate up (Fig. 5A). Even
FIG. 2. Agonist-mediated Ca2� changes in isolated striatal nuclei. A, top panel, transmitted light image of purified nucleus from DIV 9striatal culture loaded with Oregon Green BAPTA (middle panel). Bottom panel, mGlu5 (red) and lamin B2 (green) co-staining of selected nucleusfollowing drug treatment and post hoc field relocation. B, quisqualate-mediated (10 �M) representative trace. Quisqualate (Quis) applicationresulted in nuclear calcium rises that gradually plateaued at about 300 s in most cases. MPEP (1 �M) blocked this response. C, compiled data frompeak �F/Fo(%) from n � 11 cells.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium30472
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
at time points as long as 1 h only 30–40% of the cells arelabeled (not shown). Moreover, uptake is dependent upon spe-cific ionic conditions because no quisqualate could be detectedin sodium/chloride-free buffers (NMDG; Fig. 5A).
To determine whether known members of the sodium-depend-ent transporters and/or the xCT exchanger were expressed inP10 striatal tissue, total RNA was prepared and used for reversetranscription-PCR. Transcript-specific primers revealed thepresence of EAAT1–4 and the xCT exchanger; EAAT5 was notexpressed (Fig. 5B). Antibodies directed against the neuronalEAAT3 revealed that in striatal neurons EAAT3 appears highlyexpressed on cell bodies and processes as well as on the nuclearmembrane where it co-localizes with lamin B2 (Fig. 5C). Finally,Western blotting of P3-fractionated tissue using anti-EAAT3 oranti-xCT antibodies detected proteins within the nuclear frac-tions of the appropriate size (Fig. 5D). Thus both EAAT3 and xCTare possible nuclear transport candidates.
To test directly whether striatal cells and/or nuclei couldtake up quisqualate, both were treated with radiolabeled li-gand in the presence and absence of transport or exchangeinhibitors. Sodium-dependent transporter activity accountedfor 20–35% of quisqualate uptake across a wide range of con-centrations, whereas chloride-dependent xCT transport, as de-termined by gluconate substitution (or L-cystine addition, notshown), varied from 15 to 60% depending upon the quisqualateconcentration (Fig. 5E). Sodium- and chloride-free conditions
reduced quisqualate uptake by 90% (Fig. 5E). The IC50 forL-cystine for [3H]quisqualate plasma membrane transport insodium-free buffer conditions was 112 29 �M. The IC50 forthreo-�-benzyloxyaspartate (TBOA), a potent blocker of allEAAT subtypes, in sodium gluconate buffer (Cl�-free) was0.92 0.27 �M (Fig. 5F). Similar results were observed for[3H]glutamate as the endogenous ligand (Fig. 5G). In contrastto quisqualate uptake, however, sodium-dependent transport-ers took up a larger percentage of the natural ligand glutamate.Thus both xCT- and/or EAAT-mediated activity allow for intra-cellular glutamate uptake.
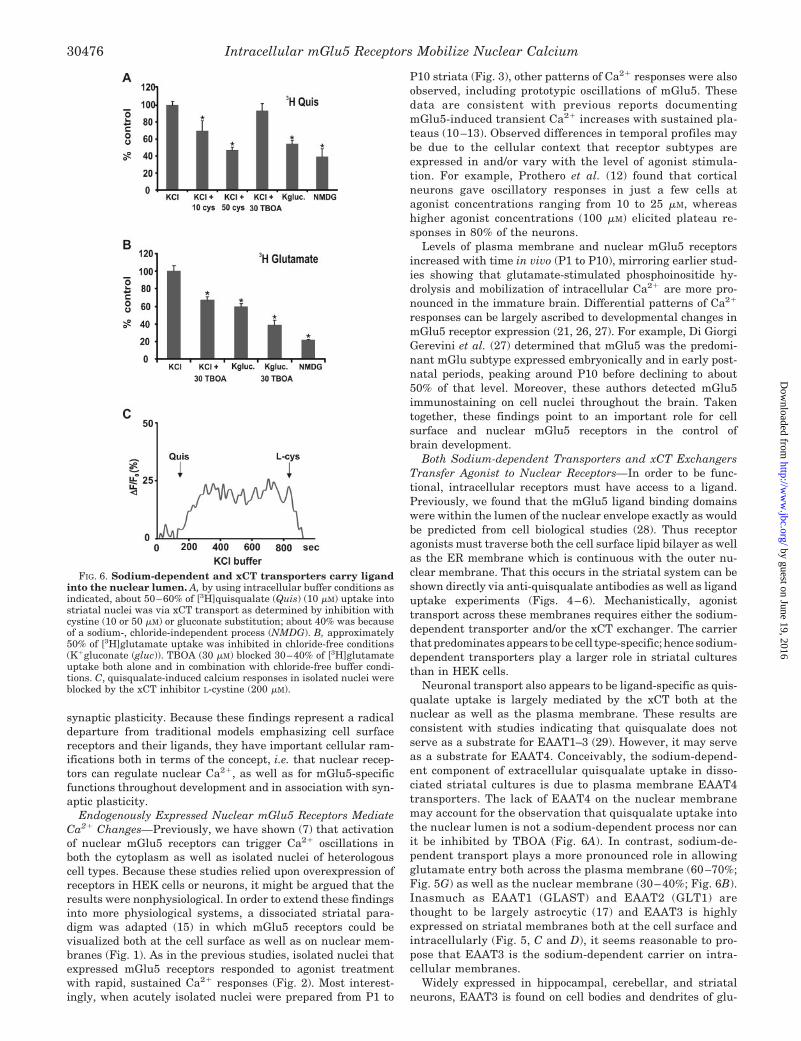
To determine what mediates glutamate uptake into the nu-clear lumen, acutely isolated striatal nuclei were prepared inintracellular buffer conditions and used for uptake studies.Nuclear [3H]quisqualate uptake exhibited two components, anxCT mechanism accounting for about 50–60% as well as an-other sodium, potassium, and chloride independent process(Fig. 6A). Quisqualate uptake via the xCT exchanger could beblocked by as little as 50 �M L-cystine or chloride-free bufferconditions (Fig. 6A, Kgluc.). Radiolabeled glutamate uptakecould also be inhibited by about 50% in chloride-free conditions(Fig. 6B). Despite the sodium-free nature of the intracellularbuffer conditions, 30 �M TBOA significantly blocked 30–40% of[3H]glutamate uptake both alone and in combination with chlo-ride-free buffer conditions; 80% of glutamate uptake could beblocked by NMDG (Fig. 6B).
FIG. 3. P1, P3, and P10 acutely iso-lated striatal nuclei express mGlu5and exhibit agonist-mediated Ca2�
changes. A, subcellular fractionation ofP1, P3, or P10 striata shows that mGlu5receptor can be detected in fractions con-taining both the nuclear (N) and plasmamembranes (PM). Thirty micrograms ofprotein from each fraction were separatedon reducing SDS gels and transferred tonylon membranes. The same blot was se-quentially probed with antibodies againstmGlu5, the inner nuclear marker, laminB2, and the plasma membrane marker,Na�,K�-ATPase. B, quisqualate (Quis)-mediated (10 �M) representative tracesfrom nuclei prepared at indicated devel-opmental time points. Quisqualate appli-cation resulted in Ca2� rises in 80–90% ofthe nuclei measured that continued torise across the period examined. MPEP (1�M) blocked this response. C, compileddata of �F/Fo (%) from the indicated num-ber of cells and time points. All cells werepost hoc fixed, stained for mGlu5, andfield re-located to ensure specificity of re-sponse. a � p � 10�5 when compared withbase-line Ca2� levels; b � p � 0.02 whencompared with P1 and P3 levels; c � p �0.005 when compared with P1 and P3 lev-els. D, between 10 and 20% of acutelyisolated striatal nuclei exhibited agonist-induced oscillatory responses. 1st panel,transmitted light image of selected nu-cleus; 2nd panel, confocal images of Ore-gon Green BAPTA loaded acutely dissoci-ated striatal nuclei treated at theindicated times (seconds) with 10 �M
quisqualate (Quis) or 1 �M MPEP. Bar atright of last panel represents �F/Fo as apseudo color scale with red being thehighest. Times correspond to those fortraces in E, where oscillations are repre-sented as the fractional change in fluores-cence relative to the basal value. D, finalpanel, mGlu5 receptor and lamin B2staining of selected nucleus followingdrug treatment and post hoc fieldrelocation.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium 30473
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
To determine whether uptake via the xCT exchanger wasnecessary for mGlu5-mediated nuclear calcium changes, quis-qualate-induced calcium influxes were treated with L-cystine(200 �M). This reagent blocked nuclear responses, demonstrat-ing that ligand transport is necessary for receptor function(Fig. 6C).
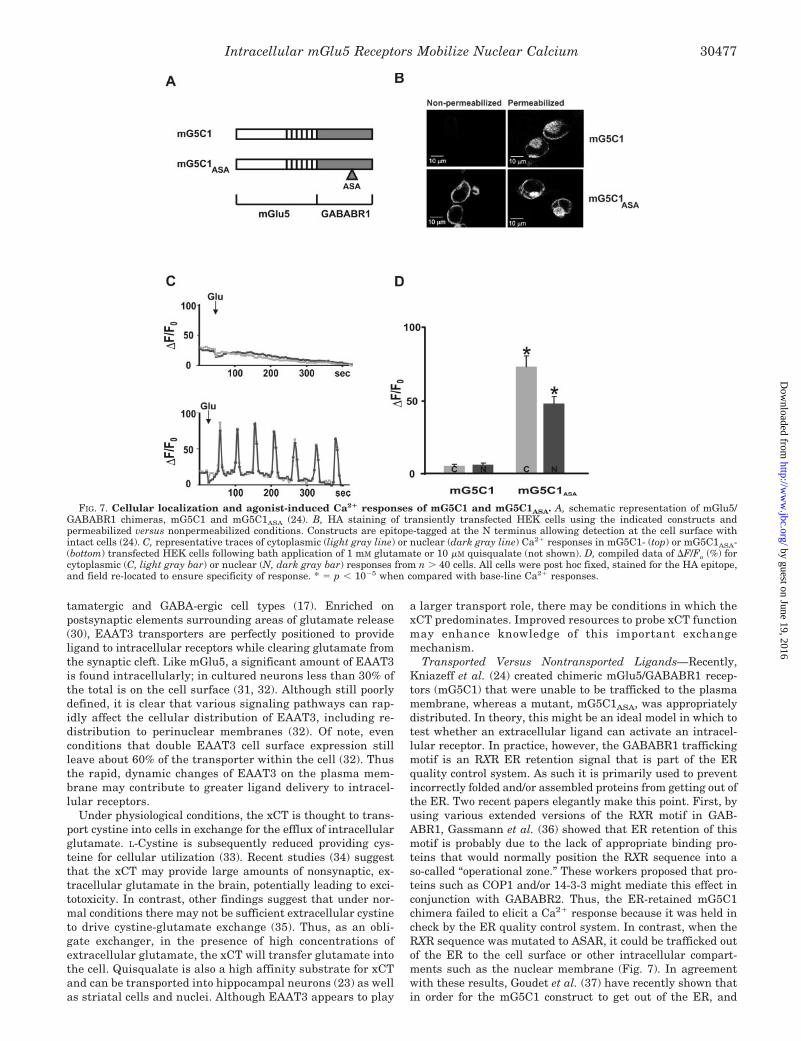
Impermeable Agonists and Antagonists Differentially AffectIntracellular Ca2� Changes—Recently, Kniazeff et al. (24) re-ported that an mGlu5/GABABR1 chimera, mG5C1, was unableto be trafficked to the cell surface, whereas mutation of an RXRmotif from RSRR to ASAR (mG5C1ASA) led to functional recep-tors on the plasma membrane. To test whether an extracellularligand could activate the intracellular mG5C1 chimera, weobtained these constructs (Fig. 7A), transfected them into HEKcells, and measured Ca2� changes in response to glutamate orquisqualate application. In agreement with Kniazeff et al. (24),the mG5C1 chimera was not trafficked to plasma membranes,whereas the mG5C1ASA mutant was easily seen on the cellsurface (Fig. 7B). Similarly, no Ca2� response was observed inmG5C1-transfected cells, whereas Ca2� oscillations were ob-served in both the cytoplasm and the nucleus in mG5C1ASA
mutant cells (Fig. 7, C and D). The lack of response in themG5C1 cells is presumably due to the inability of the chimerato be trafficked out of the ER. This is clearly shown in Fig. 7B,where under permeabilized conditions the HA-tagged mG5C1staining is concentrated in a perinuclear ER location and is notpresent on nuclear or cell surface membranes as is themG5C1ASA mutant.
Our model would predict that highly charged agonists thatare not taken into the cell via specific transport processeswould activate cell surface but not intracellular receptors. Sim-ilarly, impermeable or nontransported antagonists would blockplasma membrane receptors but not intracellular binding sites.To test these predictions, we assessed the effects of various
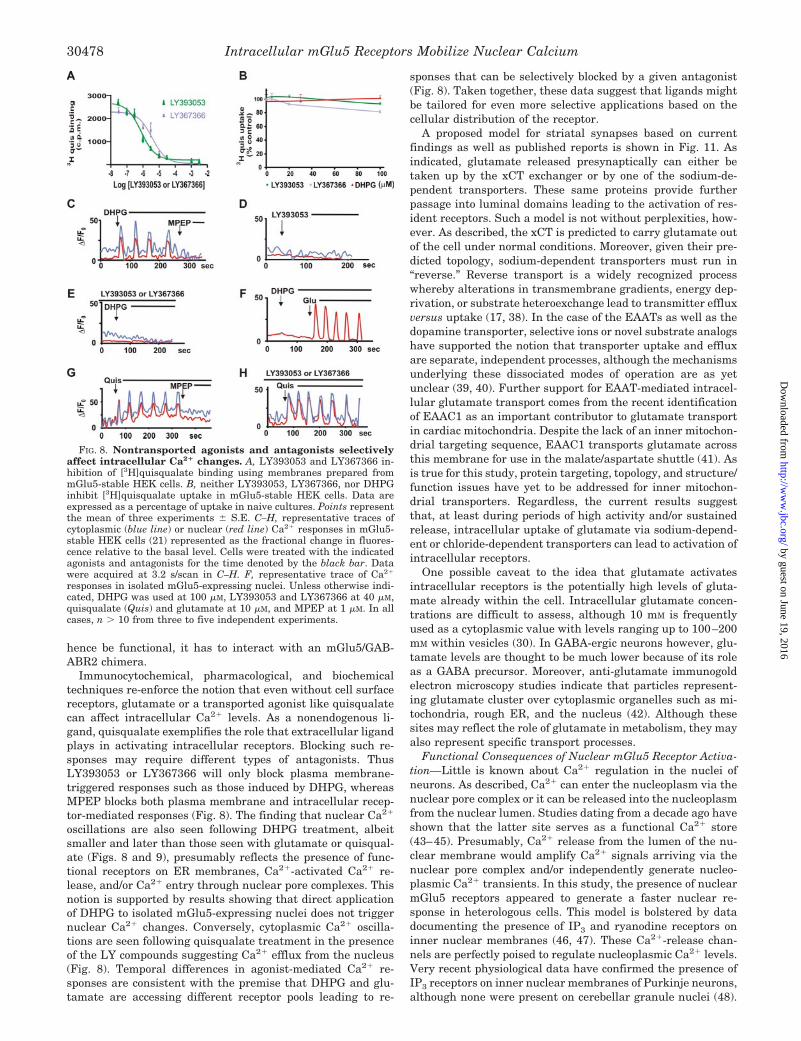
ligands on mGlu5-stable HEK cell lines. The drugs LY393053and LY367366 are thought to be impermeable antagonists spe-cific for the group 1 mGlu receptors (25). In cells derived froma Syrian hamster cell line, LY393053 and LY367366 blockedmGlu5-mediated IP3 responses with IC50 values of 1.6 1.4and 4 1 �M, respectively (25). Using membranes preparedfrom the mGlu5 HEK cells, LY393053 and LY367366 blocked[3H]quisqualate binding with IC50 values of 1.35 0.36 and4.7 0.07 �M, respectively (Fig. 8A). Despite blocking quis-qualate binding, neither compound inhibited [3H]quisqualateuptake (Fig. 8B) or [3H]glutamate uptake (not shown) even atdoses as high as 100 �M. Of course without a radiolabeledanalog, it is difficult to completely rule out the possibility thatLY393053 and LY367366 may be taken up by other types ofamino acid/carboxylate transport systems. It would appear atleast that the EAATs and the xCT are not involved. Similarly,the group 1 agonist DHPG did not affect quisqualate (Fig. 8B)or glutamate uptake either, suggesting that it too is a non-transported ligand.
In mGlu5-stable HEK cells, DHPG (100 �M) induced pro-nounced cytoplasmic Ca2� oscillations as well as nuclear re-sponses (Fig. 8C). These effects could be blocked by the perme-able antagonist, MPEP (7) (Fig. 8C). By themselves, neitherLY393053 nor LY367366 (40 �M) induced intracellular Ca2�
changes (Fig. 8D), although both completely blocked DHPG-mediated Ca2� responses (Fig. 8E). These data suggest thatDHPG selectively activates plasma membrane mGlu5 recep-tors that are susceptible to inhibition by the LY antagonists.Although nuclear responses are still seen following DHPGtreatment (Fig. 8C), these might be due to Ca2�-activated Ca2�
release and/or Ca2� entry via nuclear pore complexes. To testthis hypothesis, nuclei derived from the stable mGlu5 cell linewere treated with DHPG. In this context DHPG was unable toactivate a nuclear Ca2� response, although glutamate (10 �M)
FIG. 4. xCT mediates mGlu5 receptor activation in heterologous cells. A, anti-quisqualate antibody rapidly stains quisqualate (Quis)-treatedHEK cells following 10 �M quisqualate exposure. B, [3H]quisqualate uptake in HEK cells in the presence of extracellular buffer conditions modified asindicated. C, [3H]quisqualate uptake in isolated HEK nuclei in the presence of intracellular buffer conditions modified as indicated. Both whole cell andnuclear experiments were assayed in quadruplicate in three independent experiments. *, p � 10�4 when compared with control conditions. D,representative trace of nucleus isolated from mGlu5-stable HEK cell line (21) where quisqualate-mediated oscillations are represented as the fractionalchange in fluorescence relative to the basal level. L-Cystine (400 �M) blocked quisqualate-mediated oscillations. n � 15 cells in three independentexperiments.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium30474
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
could (Fig. 8F). These data are consistent with the premise thatDHPG is not transported across membranes. In contrast, quis-qualate induced cytoplasmic and nuclear Ca2� oscillations(Fig. 8G) even in the presence of LY393053 or LY367366 (40�M; Fig. 8H).
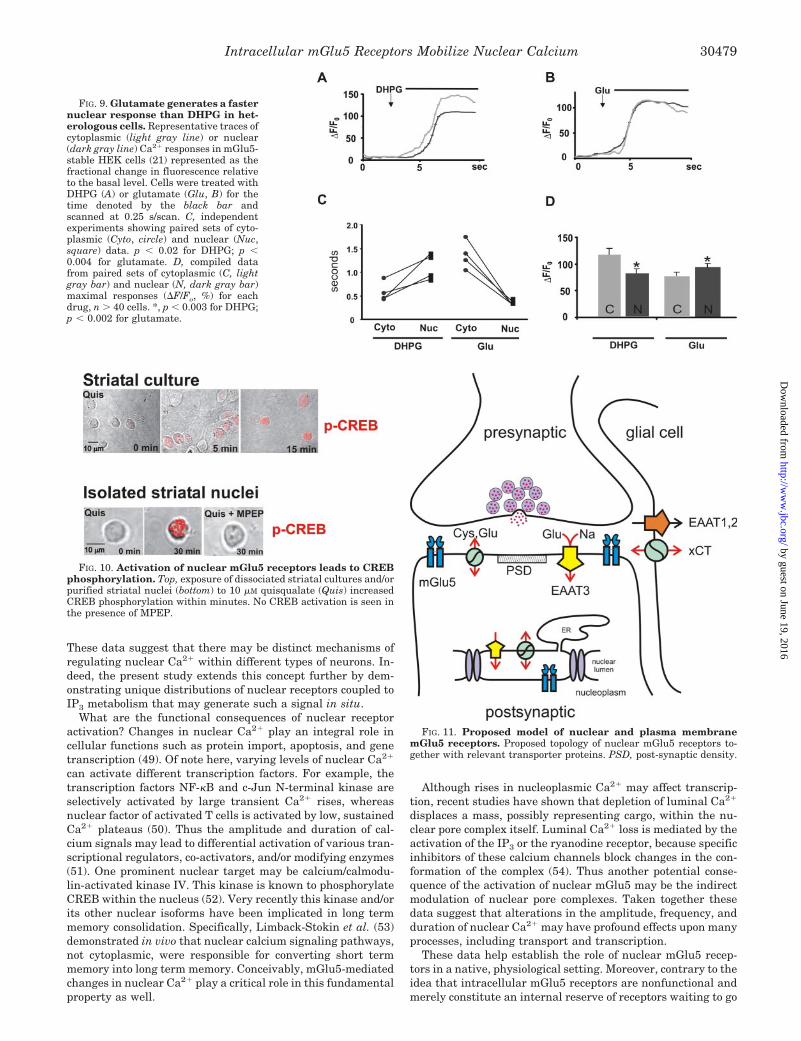
To confirm and extend the hypothesis that DHPG initiallyaffects cytoplasmic processes, we increased the scan speed(0.25 s/scan) so that the timing of the cytoplasmic and nuclearevents could be distinguished. DHPG treatment increased cy-toplasmic Ca2� 0.55 0.24 s prior to its rise in the nucleus(p � 0.02; Fig. 9, A and C), whereas glutamate- or quisqual-ate-induced nuclear changes preceded those in the cytoplasmby 0.86 0.12 s (p � 0.004; Fig. 9, B and C, and data notshown). Thus, glutamate induced a faster nuclear responsethan did DHPG (Fig. 9C). Overall, DHPG induced signifi-cantly greater cytoplasmic than nuclear Ca2� responses, andthe opposite was true with glutamate (Fig. 9D). Taken to-gether, these data suggest that depending upon their abilityto cross cellular membranes, agonists and antagonists mayhave differential effects on plasma membrane or intracellularreceptors.
Activation of Nuclear mGlu5 Receptors Leads to CREBPhosphorylation in Whole Cells and Isolated Striatal Nu-clei—What are the functional consequences of mGlu5 recep-tor-mediated induction of nuclear Ca2�? As studies haveshown that activation of mGlu5 receptors in vitro and in vivoup-regulates several transcription factors including CREB(16), we tested whether quisqualate increased CREB phos-
phorylation in dissociated striatal cultures as well as in iso-lated nuclei. Robust up-regulation of phosphorylated CREBwas observed within minutes of quisqualate application ineither paradigm (Fig. 10). CREB phosphorylation was notseen when nuclei were pretreated with MPEP. Thus directactivation of nuclear receptors initiates signaling cascadeswithin the nucleus.
DISCUSSION
Contrary to the idea that intracellular mGlu5 receptors arenonfunctional and merely constitute an internal reserve ofreceptors waiting to go to the cell surface, the present resultsdemonstrate that mGlu5 receptors play dynamic intracellularroles in signal transduction. These studies show that in anative, physiological setting, i.e. striatal dissociated culturesand acutely isolated striatal nuclei, activation of nuclearmGlu5 receptors leads to rapid, sustained Ca2� responses thatcan be blocked by the mGlu5-specific antagonist MPEP. Cur-rent results demonstrate that both sodium-dependent trans-porters as well as xCT exchangers are involved in movingagonist across both plasma and nuclear membranes. Inhibitionof either transport system blocks agonist-induced nuclear Ca2�
changes. In the presence of an impermeable antagonist, trans-ported ligands such as quisqualate or glutamate can still in-duce intracellular Ca2� responses, whereas nontransportedagonists are ineffectual. Finally, ligand stimulation of nuclearreceptors initiates at least one signaling cascade that is knownto alter gene transcription and regulates many paradigms of
FIG. 5. Sodium-dependent and xCTtransporters carry ligand into stria-tal cells. A, within 1 min of 10 �M quis-qualate (Quis) addition, anti-quisqualateantibodies (red) stain some but not allstriatal neurons. When cells are incu-bated in sodium- and chloride-free condi-tions (NMDG), no quisqualate staining isobserved. B, EAAT1–4 and xCT tran-scripts can be detected in total RNAprepared from murine P10 tissue. Gene-specific primers were used to reverse-transcribe and subsequently amplifycDNAs for each transporter. Followingsize fractionation and fluorescent detec-tion, all products were verified by internalprobes, internal unique restriction sites,or both. C, left panel, EAAT3 (red) is ex-pressed on all striatal neurons includingmGlu5-positive cells (green); right panel,EAAT3 (red) also co-localizes with laminB2 (green) in striatal neurons. Co-localiza-tion is indicated by the merged images(yellow). D, Western analysis of fraction-ated P3 striatal tissue shows that EAAT3and xCT can be detected in fractions con-taining both the nuclear (N) and plasmamembranes (PM). E, dose-dependent[3H]quisqualate uptake in striatal cul-tures in the presence of extracellularbuffer conditions modified as indicated.Bars represent the mean of 3–6 experi-ments S.E. *, p � 0.001. F, L-cystineand TBOA inhibition of [3H]quisqualateuptake in striatal cultures. Data are ex-pressed as a percentage of uptake in naivecultures. Points represent the mean of3–6 experiments S.E. G, [3H]glutamate(1 mM) uptake in striatal cultures in thepresence of extracellular buffer conditionsmodified as indicated. Bars represent themean of 3–6 experiments S.E. *, p �0.001.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium 30475
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
synaptic plasticity. Because these findings represent a radicaldeparture from traditional models emphasizing cell surfacereceptors and their ligands, they have important cellular ram-ifications both in terms of the concept, i.e. that nuclear recep-tors can regulate nuclear Ca2�, as well as for mGlu5-specificfunctions throughout development and in association with syn-aptic plasticity.
Endogenously Expressed Nuclear mGlu5 Receptors MediateCa2� Changes—Previously, we have shown (7) that activationof nuclear mGlu5 receptors can trigger Ca2� oscillations inboth the cytoplasm as well as isolated nuclei of heterologouscell types. Because these studies relied upon overexpression ofreceptors in HEK cells or neurons, it might be argued that theresults were nonphysiological. In order to extend these findingsinto more physiological systems, a dissociated striatal para-digm was adapted (15) in which mGlu5 receptors could bevisualized both at the cell surface as well as on nuclear mem-branes (Fig. 1). As in the previous studies, isolated nuclei thatexpressed mGlu5 receptors responded to agonist treatmentwith rapid, sustained Ca2� responses (Fig. 2). Most interest-ingly, when acutely isolated nuclei were prepared from P1 to
P10 striata (Fig. 3), other patterns of Ca2� responses were alsoobserved, including prototypic oscillations of mGlu5. Thesedata are consistent with previous reports documentingmGlu5-induced transient Ca2� increases with sustained pla-teaus (10–13). Observed differences in temporal profiles maybe due to the cellular context that receptor subtypes areexpressed in and/or vary with the level of agonist stimula-tion. For example, Prothero et al. (12) found that corticalneurons gave oscillatory responses in just a few cells atagonist concentrations ranging from 10 to 25 �M, whereashigher agonist concentrations (100 �M) elicited plateau re-sponses in 80% of the neurons.
Levels of plasma membrane and nuclear mGlu5 receptorsincreased with time in vivo (P1 to P10), mirroring earlier stud-ies showing that glutamate-stimulated phosphoinositide hy-drolysis and mobilization of intracellular Ca2� are more pro-nounced in the immature brain. Differential patterns of Ca2�
responses can be largely ascribed to developmental changes inmGlu5 receptor expression (21, 26, 27). For example, Di GiorgiGerevini et al. (27) determined that mGlu5 was the predomi-nant mGlu subtype expressed embryonically and in early post-natal periods, peaking around P10 before declining to about50% of that level. Moreover, these authors detected mGlu5immunostaining on cell nuclei throughout the brain. Takentogether, these findings point to an important role for cellsurface and nuclear mGlu5 receptors in the control ofbrain development.
Both Sodium-dependent Transporters and xCT ExchangersTransfer Agonist to Nuclear Receptors—In order to be func-tional, intracellular receptors must have access to a ligand.Previously, we found that the mGlu5 ligand binding domainswere within the lumen of the nuclear envelope exactly as wouldbe predicted from cell biological studies (28). Thus receptoragonists must traverse both the cell surface lipid bilayer as wellas the ER membrane which is continuous with the outer nu-clear membrane. That this occurs in the striatal system can beshown directly via anti-quisqualate antibodies as well as liganduptake experiments (Figs. 4–6). Mechanistically, agonisttransport across these membranes requires either the sodium-dependent transporter and/or the xCT exchanger. The carrierthat predominates appears to be cell type-specific; hence sodium-dependent transporters play a larger role in striatal culturesthan in HEK cells.
Neuronal transport also appears to be ligand-specific as quis-qualate uptake is largely mediated by the xCT both at thenuclear as well as the plasma membrane. These results areconsistent with studies indicating that quisqualate does notserve as a substrate for EAAT1–3 (29). However, it may serveas a substrate for EAAT4. Conceivably, the sodium-depend-ent component of extracellular quisqualate uptake in disso-ciated striatal cultures is due to plasma membrane EAAT4transporters. The lack of EAAT4 on the nuclear membranemay account for the observation that quisqualate uptake intothe nuclear lumen is not a sodium-dependent process nor canit be inhibited by TBOA (Fig. 6A). In contrast, sodium-de-pendent transport plays a more pronounced role in allowingglutamate entry both across the plasma membrane (60–70%;Fig. 5G) as well as the nuclear membrane (30–40%; Fig. 6B).Inasmuch as EAAT1 (GLAST) and EAAT2 (GLT1) arethought to be largely astrocytic (17) and EAAT3 is highlyexpressed on striatal membranes both at the cell surface andintracellularly (Fig. 5, C and D), it seems reasonable to pro-pose that EAAT3 is the sodium-dependent carrier on intra-cellular membranes.
Widely expressed in hippocampal, cerebellar, and striatalneurons, EAAT3 is found on cell bodies and dendrites of glu-
FIG. 6. Sodium-dependent and xCT transporters carry ligandinto the nuclear lumen. A, by using intracellular buffer conditions asindicated, about 50–60% of [3H]quisqualate (Quis) (10 �M) uptake intostriatal nuclei was via xCT transport as determined by inhibition withcystine (10 or 50 �M) or gluconate substitution; about 40% was becauseof a sodium-, chloride-independent process (NMDG). B, approximately50% of [3H]glutamate uptake was inhibited in chloride-free conditions(K�gluconate (gluc)). TBOA (30 �M) blocked 30–40% of [3H]glutamateuptake both alone and in combination with chloride-free buffer condi-tions. C, quisqualate-induced calcium responses in isolated nuclei wereblocked by the xCT inhibitor L-cystine (200 �M).
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium30476
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
tamatergic and GABA-ergic cell types (17). Enriched onpostsynaptic elements surrounding areas of glutamate release(30), EAAT3 transporters are perfectly positioned to provideligand to intracellular receptors while clearing glutamate fromthe synaptic cleft. Like mGlu5, a significant amount of EAAT3is found intracellularly; in cultured neurons less than 30% ofthe total is on the cell surface (31, 32). Although still poorlydefined, it is clear that various signaling pathways can rap-idly affect the cellular distribution of EAAT3, including re-distribution to perinuclear membranes (32). Of note, evenconditions that double EAAT3 cell surface expression stillleave about 60% of the transporter within the cell (32). Thusthe rapid, dynamic changes of EAAT3 on the plasma mem-brane may contribute to greater ligand delivery to intracel-lular receptors.
Under physiological conditions, the xCT is thought to trans-port cystine into cells in exchange for the efflux of intracellularglutamate. L-Cystine is subsequently reduced providing cys-teine for cellular utilization (33). Recent studies (34) suggestthat the xCT may provide large amounts of nonsynaptic, ex-tracellular glutamate in the brain, potentially leading to exci-totoxicity. In contrast, other findings suggest that under nor-mal conditions there may not be sufficient extracellular cystineto drive cystine-glutamate exchange (35). Thus, as an obli-gate exchanger, in the presence of high concentrations ofextracellular glutamate, the xCT will transfer glutamate intothe cell. Quisqualate is also a high affinity substrate for xCTand can be transported into hippocampal neurons (23) as wellas striatal cells and nuclei. Although EAAT3 appears to play
a larger transport role, there may be conditions in which thexCT predominates. Improved resources to probe xCT functionmay enhance knowledge of this important exchangemechanism.
Transported Versus Nontransported Ligands—Recently,Kniazeff et al. (24) created chimeric mGlu5/GABABR1 recep-tors (mG5C1) that were unable to be trafficked to the plasmamembrane, whereas a mutant, mG5C1ASA, was appropriatelydistributed. In theory, this might be an ideal model in which totest whether an extracellular ligand can activate an intracel-lular receptor. In practice, however, the GABABR1 traffickingmotif is an RXR ER retention signal that is part of the ERquality control system. As such it is primarily used to preventincorrectly folded and/or assembled proteins from getting out ofthe ER. Two recent papers elegantly make this point. First, byusing various extended versions of the RXR motif in GAB-ABR1, Gassmann et al. (36) showed that ER retention of thismotif is probably due to the lack of appropriate binding pro-teins that would normally position the RXR sequence into aso-called “operational zone.” These workers proposed that pro-teins such as COP1 and/or 14-3-3 might mediate this effect inconjunction with GABABR2. Thus, the ER-retained mG5C1chimera failed to elicit a Ca2� response because it was held incheck by the ER quality control system. In contrast, when theRXR sequence was mutated to ASAR, it could be trafficked outof the ER to the cell surface or other intracellular compart-ments such as the nuclear membrane (Fig. 7). In agreementwith these results, Goudet et al. (37) have recently shown thatin order for the mG5C1 construct to get out of the ER, and
FIG. 7. Cellular localization and agonist-induced Ca2� responses of mG5C1 and mG5C1ASA. A, schematic representation of mGlu5/GABABR1 chimeras, mG5C1 and mG5C1ASA (24). B, HA staining of transiently transfected HEK cells using the indicated constructs andpermeabilized versus nonpermeabilized conditions. Constructs are epitope-tagged at the N terminus allowing detection at the cell surface withintact cells (24). C, representative traces of cytoplasmic (light gray line) or nuclear (dark gray line) Ca2� responses in mG5C1- (top) or mG5C1ASA-(bottom) transfected HEK cells following bath application of 1 mM glutamate or 10 �M quisqualate (not shown). D, compiled data of �F/Fo (%) forcytoplasmic (C, light gray bar) or nuclear (N, dark gray bar) responses from n � 40 cells. All cells were post hoc fixed, stained for the HA epitope,and field re-located to ensure specificity of response. * � p � 10�5 when compared with base-line Ca2� responses.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium 30477
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
hence be functional, it has to interact with an mGlu5/GAB-ABR2 chimera.
Immunocytochemical, pharmacological, and biochemicaltechniques re-enforce the notion that even without cell surfacereceptors, glutamate or a transported agonist like quisqualatecan affect intracellular Ca2� levels. As a nonendogenous li-gand, quisqualate exemplifies the role that extracellular ligandplays in activating intracellular receptors. Blocking such re-sponses may require different types of antagonists. ThusLY393053 or LY367366 will only block plasma membrane-triggered responses such as those induced by DHPG, whereasMPEP blocks both plasma membrane and intracellular recep-tor-mediated responses (Fig. 8). The finding that nuclear Ca2�
oscillations are also seen following DHPG treatment, albeitsmaller and later than those seen with glutamate or quisqual-ate (Figs. 8 and 9), presumably reflects the presence of func-tional receptors on ER membranes, Ca2�-activated Ca2� re-lease, and/or Ca2� entry through nuclear pore complexes. Thisnotion is supported by results showing that direct applicationof DHPG to isolated mGlu5-expressing nuclei does not triggernuclear Ca2� changes. Conversely, cytoplasmic Ca2� oscilla-tions are seen following quisqualate treatment in the presenceof the LY compounds suggesting Ca2� efflux from the nucleus(Fig. 8). Temporal differences in agonist-mediated Ca2� re-sponses are consistent with the premise that DHPG and glu-tamate are accessing different receptor pools leading to re-
sponses that can be selectively blocked by a given antagonist(Fig. 8). Taken together, these data suggest that ligands mightbe tailored for even more selective applications based on thecellular distribution of the receptor.
A proposed model for striatal synapses based on currentfindings as well as published reports is shown in Fig. 11. Asindicated, glutamate released presynaptically can either betaken up by the xCT exchanger or by one of the sodium-de-pendent transporters. These same proteins provide furtherpassage into luminal domains leading to the activation of res-ident receptors. Such a model is not without perplexities, how-ever. As described, the xCT is predicted to carry glutamate outof the cell under normal conditions. Moreover, given their pre-dicted topology, sodium-dependent transporters must run in“reverse.” Reverse transport is a widely recognized processwhereby alterations in transmembrane gradients, energy dep-rivation, or substrate heteroexchange lead to transmitter effluxversus uptake (17, 38). In the case of the EAATs as well as thedopamine transporter, selective ions or novel substrate analogshave supported the notion that transporter uptake and effluxare separate, independent processes, although the mechanismsunderlying these dissociated modes of operation are as yetunclear (39, 40). Further support for EAAT-mediated intracel-lular glutamate transport comes from the recent identificationof EAAC1 as an important contributor to glutamate transportin cardiac mitochondria. Despite the lack of an inner mitochon-drial targeting sequence, EAAC1 transports glutamate acrossthis membrane for use in the malate/aspartate shuttle (41). Asis true for this study, protein targeting, topology, and structure/function issues have yet to be addressed for inner mitochon-drial transporters. Regardless, the current results suggestthat, at least during periods of high activity and/or sustainedrelease, intracellular uptake of glutamate via sodium-depend-ent or chloride-dependent transporters can lead to activation ofintracellular receptors.
One possible caveat to the idea that glutamate activatesintracellular receptors is the potentially high levels of gluta-mate already within the cell. Intracellular glutamate concen-trations are difficult to assess, although 10 mM is frequentlyused as a cytoplasmic value with levels ranging up to 100–200mM within vesicles (30). In GABA-ergic neurons however, glu-tamate levels are thought to be much lower because of its roleas a GABA precursor. Moreover, anti-glutamate immunogoldelectron microscopy studies indicate that particles represent-ing glutamate cluster over cytoplasmic organelles such as mi-tochondria, rough ER, and the nucleus (42). Although thesesites may reflect the role of glutamate in metabolism, they mayalso represent specific transport processes.
Functional Consequences of Nuclear mGlu5 Receptor Activa-tion—Little is known about Ca2� regulation in the nuclei ofneurons. As described, Ca2� can enter the nucleoplasm via thenuclear pore complex or it can be released into the nucleoplasmfrom the nuclear lumen. Studies dating from a decade ago haveshown that the latter site serves as a functional Ca2� store(43–45). Presumably, Ca2� release from the lumen of the nu-clear membrane would amplify Ca2� signals arriving via thenuclear pore complex and/or independently generate nucleo-plasmic Ca2� transients. In this study, the presence of nuclearmGlu5 receptors appeared to generate a faster nuclear re-sponse in heterologous cells. This model is bolstered by datadocumenting the presence of IP3 and ryanodine receptors oninner nuclear membranes (46, 47). These Ca2�-release chan-nels are perfectly poised to regulate nucleoplasmic Ca2� levels.Very recent physiological data have confirmed the presence ofIP3 receptors on inner nuclear membranes of Purkinje neurons,although none were present on cerebellar granule nuclei (48).
FIG. 8. Nontransported agonists and antagonists selectivelyaffect intracellular Ca2� changes. A, LY393053 and LY367366 in-hibition of [3H]quisqualate binding using membranes prepared frommGlu5-stable HEK cells. B, neither LY393053, LY367366, nor DHPGinhibit [3H]quisqualate uptake in mGlu5-stable HEK cells. Data areexpressed as a percentage of uptake in naive cultures. Points representthe mean of three experiments S.E. C–H, representative traces ofcytoplasmic (blue line) or nuclear (red line) Ca2� responses in mGlu5-stable HEK cells (21) represented as the fractional change in fluores-cence relative to the basal level. Cells were treated with the indicatedagonists and antagonists for the time denoted by the black bar. Datawere acquired at 3.2 s/scan in C–H. F, representative trace of Ca2�
responses in isolated mGlu5-expressing nuclei. Unless otherwise indi-cated, DHPG was used at 100 �M, LY393053 and LY367366 at 40 �M,quisqualate (Quis) and glutamate at 10 �M, and MPEP at 1 �M. In allcases, n � 10 from three to five independent experiments.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium30478
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
These data suggest that there may be distinct mechanisms ofregulating nuclear Ca2� within different types of neurons. In-deed, the present study extends this concept further by dem-onstrating unique distributions of nuclear receptors coupled toIP3 metabolism that may generate such a signal in situ.
What are the functional consequences of nuclear receptoractivation? Changes in nuclear Ca2� play an integral role incellular functions such as protein import, apoptosis, and genetranscription (49). Of note here, varying levels of nuclear Ca2�
can activate different transcription factors. For example, thetranscription factors NF-�B and c-Jun N-terminal kinase areselectively activated by large transient Ca2� rises, whereasnuclear factor of activated T cells is activated by low, sustainedCa2� plateaus (50). Thus the amplitude and duration of cal-cium signals may lead to differential activation of various tran-scriptional regulators, co-activators, and/or modifying enzymes(51). One prominent nuclear target may be calcium/calmodu-lin-activated kinase IV. This kinase is known to phosphorylateCREB within the nucleus (52). Very recently this kinase and/orits other nuclear isoforms have been implicated in long termmemory consolidation. Specifically, Limback-Stokin et al. (53)demonstrated in vivo that nuclear calcium signaling pathways,not cytoplasmic, were responsible for converting short termmemory into long term memory. Conceivably, mGlu5-mediatedchanges in nuclear Ca2� play a critical role in this fundamentalproperty as well.
Although rises in nucleoplasmic Ca2� may affect transcrip-tion, recent studies have shown that depletion of luminal Ca2�
displaces a mass, possibly representing cargo, within the nu-clear pore complex itself. Luminal Ca2� loss is mediated by theactivation of the IP3 or the ryanodine receptor, because specificinhibitors of these calcium channels block changes in the con-formation of the complex (54). Thus another potential conse-quence of the activation of nuclear mGlu5 may be the indirectmodulation of nuclear pore complexes. Taken together thesedata suggest that alterations in the amplitude, frequency, andduration of nuclear Ca2� may have profound effects upon manyprocesses, including transport and transcription.
These data help establish the role of nuclear mGlu5 recep-tors in a native, physiological setting. Moreover, contrary to theidea that intracellular mGlu5 receptors are nonfunctional andmerely constitute an internal reserve of receptors waiting to go
FIG. 10. Activation of nuclear mGlu5 receptors leads to CREBphosphorylation. Top, exposure of dissociated striatal cultures and/orpurified striatal nuclei (bottom) to 10 �M quisqualate (Quis) increasedCREB phosphorylation within minutes. No CREB activation is seen inthe presence of MPEP.
FIG. 11. Proposed model of nuclear and plasma membranemGlu5 receptors. Proposed topology of nuclear mGlu5 receptors to-gether with relevant transporter proteins. PSD, post-synaptic density.
FIG. 9. Glutamate generates a fasternuclear response than DHPG in het-erologous cells. Representative traces ofcytoplasmic (light gray line) or nuclear(dark gray line) Ca2� responses in mGlu5-stable HEK cells (21) represented as thefractional change in fluorescence relativeto the basal level. Cells were treated withDHPG (A) or glutamate (Glu, B) for thetime denoted by the black bar andscanned at 0.25 s/scan. C, independentexperiments showing paired sets of cyto-plasmic (Cyto, circle) and nuclear (Nuc,square) data. p � 0.02 for DHPG; p �0.004 for glutamate. D, compiled datafrom paired sets of cytoplasmic (C, lightgray bar) and nuclear (N, dark gray bar)maximal responses (�F/Fo, %) for eachdrug, n � 40 cells. *, p � 0.003 for DHPG;p � 0.002 for glutamate.
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium 30479
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from

to the cell surface, the present data argue strongly for a dy-namic intracellular role in signal transduction. Given that theER and nuclear lumen serve as unique intracellular stores ofCa2�, intracellular receptors such as mGlu5 may play a pivotalrole in generating and shaping intracellular Ca2� signals.
Acknowledgments—We thank Seta Dikranian and Steve Harmon fortechnical assistance, Michael Morgan for MATLAB® analyses, andDr. J. Wang (University of Missouri, Kansas City) for teaching us thestriatal culture system. We also thank Dr. J. Rothstein (The JohnsHopkins University), Dr. J. P. Pin (Institute of Functional Genomics,CNRS, France), and Drs. Rachel Wong and Steve Mennerick for insight-ful comments and critical reading of the manuscript.
REFERENCES
1. Lee, D. K., Lanca, A. J., Cheng, R., Nguyen, T., Ji, X. D., Gobeil, F., Jr.,Chemtob, S., George, S. R., and O’Dowd, B. F. (2004) J. Biol. Chem. 279,7901–7908
2. Chen, R., Mukhin, Y. V., Garnovskaya, M. N., Thielen, T. E., Iijima, Y., Huang,C., Raymond, J. R., Ullian, M. E., and Paul, R. V. (2000) Am. J. Physiol. 279,F440–F448
3. Moughal, N. A., Waters, C., Sambi, B., Pyne, S., and Pyne, N. J. (2004) Cell.Signal. 16, 127–136
4. Bhattacharya, M., Peri, K., Ribeiro-da-Silva, A., Almazan, G., Shichi, H., Hou,X., Varma, D. R., and Chemtob, S. (1999) J. Biol. Chem. 274, 15719–15724
5. Bkaily, G., Choufani, S., Hassan, G., El-Bizri, N., Jacques, D., and D’Orleans-Juste, P. (2000) J. Cardiovasc. Pharmacol. 36, S414–S417
6. Boivin, B., Chevalier, D., Villeneuve, L. R., Rousseau, E., and Allen, B. G.(2003) J. Biol. Chem. 278, 29153–29163
7. O’Malley, K. L., Jong, Y. J., Gonchar, Y., Burkhalter, A., and Romano, C.(2003) J. Biol. Chem. 278, 28210–28219
8. Hermans, E., and Challiss, R. A. (2001) Biochem. J. 359, 465–4849. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y., Nakahara, K., Watanabe, D.,
Yamaguchi, S., Kawabata, S., and Okada, M. (1998) Brain Res. Rev. 26,230–235
10. Flint, A. C., Dammerman, R. S., and Kriegstein, A. R. (1999) Proc. Natl. Acad.Sci. U. S. A. 96, 12144–12149
11. Kettunen, P., Krieger, P., Hess, D., and El Manira, A. (2002) J. Neurosci. 22,1868–1873
12. Prothero, L. S., Richards, C. D., and Mathie, A. (1998) Br. J. Pharmacol. 125,1551–1561
13. Marino, M. J., Awad-Granko, H., Ciombor, K. J., and Conn, P. J. (2002)Neuropharmacology 43, 147–159
14. Paquet, M., and Smith, Y. (2003) J. Neurosci. 23, 7659–766915. Mao, L., and Wang, J. Q. (2001) Mol. Brain Res. 86, 125–13716. Mao, L., and Wang, J. Q. (2003) J. Neurochem. 84, 233–24317. Danbolt, N. C. (2001) Prog. Neurobiol. 65, 1–10518. Kanai, Y., and Endou, H. (2001) Curr. Drug Metab. 2, 339–35419. Patel, S. A., Warren, B. A., Rhoderick, J. F., and Bridges, R. J. (2004) Neuro-
pharmacology 46, 273–28420. Romano, C., Miller, J. K., Hyrc, K., Dikranian, S., Mennerick, S., Takeuchi, Y.,
Goldberg, M. P., and O’Malley, K. L. (2001) Mol. Pharmacol. 59, 46–5321. Romano, C., van den Pol, A. N., and O’Malley, K. L. (1996) J. Comp. Neurol.
367, 403–41222. Ferraguti, F., Conquet, F., Corti, C., Grandes, P., Kuhn, R., and Knopfel, T.
(1998) J. Comp. Neurol. 400, 391–40723. Chase, L. A., Roon, R. J., Wellman, L., Beitz, A. J., and Koerner, J. F. (2001)
Neuroscience 106, 287–30124. Kniazeff, J., Bessis, A. S., Maurel, D., Ansanay, H., Prezeau, L., and Pin, J. P.
(2004) Nat. Struct. Mol. Biol. 11, 706–71325. Kingston, A. E., Griffey, K., Johnson, M. P., Chamberlain, M. J., Kelly, G.,
Tomlinson, R., Wright, R. A., Johnson, B. G., Schoepp, D. D., Harris, J. R.,Clark, B. P., Baker, R. S., and Tizzano, J. T. (2002) Neurosci. Lett. 330,127–130
26. Lopez-Bendito, G., Shigemoto, R., Fairen, A., and Lujan, R. (2002) Cereb.Cortex 12, 625–638
27. Di Giorgi Gerevini, V. D., Caruso, A., Cappuccio, I., Ricci Vitiani, L., Romeo, S.,Della Rocca, C., Gradini, R., Melchiorri, D., and Nicoletti, F. (2004) Dev.Brain. Res. 150, 17–22
28. Ott, C. M., and Lingappa, V. R. (2002) J. Cell Sci. 115, 2003–200929. Arriza, J. L., Fairman, W. A., Wadiche, J. I., Murdoch, G. H., Kavanaugh,
M. P., and Amara, S. G. (1994) J. Neurosci. 14, 5559–556930. Kanai, Y., and Hediger, M. A. (2004) Pflugers Arch. 447, 469–47931. Gonzalez, M. I., Kazanietz, M. G., and Robinson, M. B. (2002) Mol. Pharmacol.
62, 901–91032. Fournier, K. M., Gonzalez, M. I., and Robinson, M. B. (2004) J. Biol. Chem.
279, 34505–3451333. Bannai, S. (1984) Biochim. Biophys. Acta 779, 289–30634. Baker, D. A., Xi, Z. X., Shen, H., Swanson, C. J., and Kalivas, P. W. (2002)
J. Neurosci. 22, 9134–914135. Cavelier, P., Hamann, M., Rossi, D., Mobbs, P., and Attwell, D. (2005) Prog.
Biophys. Mol. Biol. 87, 3–1636. Gassmann, M., Haller, C., Stoll, Y., Aziz, S., Biermann, B., Mosbacher, J.,
Kaupmann, K., and Bettler, B. (2005) Mol. Pharmacol. 68, 137–14437. Goudet, C., Kniazeff, J., Hlavackova, V., Malhaire, F., Maurel, D., Acher, F.,
Blahos, J., Prezeau, L., and Pin, J. P. (2005) J. Biol. Chem. 280,24380–24385
38. Kanner, B. I. (1983) Biochim. Biophys. Acta 726, 293–31639. Funicello, M., Conti, P., De Amici, M., De Micheli, C., Mennini, T., and Gobbi,
M. (2004) Mol. Pharmacol. 66, 522–52940. Scholze, P., Norregaard, L., Singer, E. A., Freissmuth, M., Gether, U., and
Sitte, H. H. (2002) J. Biol. Chem. 277, 21505–2151341. Ralphe, J. C., Segar, J. L., Schutte, B. C., and Scholz, T. D. (2004) J. Mol. Cell.
Cardiol. 37, 33–4142. Kharazia, V. N., and Weinberg, R. J. (1994) J. Neurosci. 14, 6021–603243. Nicotera, P., Orrenius, S., Nilsson, T., and Berggren, P. O. (1990) Proc. Natl.
Acad. Sci. U. S. A. 87, 6858–686244. Gerasimenko, O. V., Gerasimenko, J. V., Tepikin, A. V., and Petersen, O. H.
(1995) Cell 80, 439–44445. Stehno-Bittel, L., Perez-Terzic, C., and Clapham, D. E. (1995) Science 270,
1835–183846. Humbert, J. P., Matter, N., Artault, J. C., Koppler, P., and Malviya, A. N.
(1996) J. Biol. Chem. 271, 478–48547. Gerasimenko, J. V., Maruyama, Y., Yano, K., Dolman, N. J., Tepikin, A. V.,
Petersen, O. H., and Gerasimenko, O. V. (2003) J. Cell Biol. 163, 271–28248. Marchenko, S. M., Yarotskyy, V. V., Kovalenko, T. N., Kostyuk, P. G., and
Thomas, R. C. (2005) J. Physiol. (Lond.) 565, 897–91049. Berridge, M. J. (2001) Novartis Found. Symp. 239, 52–6450. Dolmetsch, R. E., Lewis, R. S., Goodnow, C. C., and Healy, J. I. (1997) Nature
386, 855–85851. Dolmetsch, R. (2003) Sci. STKE 2003, PE452. Soderling, T. R. (1999) Trends Biochem. Sci. 24, 232–23653. Limback-Stokin, K., Korzus, E., Nagaoka-Yasuda. R., and Mayford, M. (2004)
J. Neurosci. 24, 10858–1086754. Erickson, E. S., Mooren, O. L., Moore-Nichols, D., and Dunn, R. C. (2004)
Biophys. Chem. 112, 1–7
Intracellular mGlu5 Receptors Mobilize Nuclear Calcium30480
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from

O'MalleyYuh-Jiin I. Jong, Vikas Kumar, Ann E. Kingston, Carmelo Romano and Karen L.
LIGANDCultured Striatal Neurons: ROLE OF TRANSPORTERS IN DELIVERING
Functional Metabotropic Glutamate Receptors on Nuclei from Brain and Primary
doi: 10.1074/jbc.M501775200 originally published online June 15, 20052005, 280:30469-30480.J. Biol. Chem.
10.1074/jbc.M501775200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/280/34/30469.full.html#ref-list-1
This article cites 53 references, 24 of which can be accessed free at
by guest on June 19, 2016http://w
ww
.jbc.org/D
ownloaded from
Related Documents











