Proc. Nati Acad. Sci. USA Vol. 79, pp. 3408-3412, June 1982 Biochemistry Functional exchange of components between light-activated photoreceptor phosphodiesterase and hormone-activated adenylate cyclase systems (rhodopsin/GTP-binding protein/phosphodiesterase inhibitor/cerebral cortex synaptic membranes/frog erythrocyte ghosts) MARK W. BITENSKY*, MARCIA A. WHEELER*, MARK M. RASENICK, AKIO YAMAZAKI*, PETER J. STEIN, KAREN R. HALLIDAY, AND GEORGE L. WHEELER* Yale University School of Medicine, Department of Pathology, New Haven, Connecticut 06510 Communicated by Lewis Thomas, February 11, 1982 ABSTRACT Previous studies have noted profound similarities between the regulation of light-activated 3',5'-cyclic nucleotide phosphodiesterase (3',5'-cyclic-nucleotide 5'-nucleotidohydro- lase, EC 3.1.4.17) in retinal rods and hormone-activated ade- nylate cyclase [ATP pyrophosphate-lyase (cyclizing), EC 4.6.1.1] in a variety of tissues. We report here the functional exchange of components isolated from the photoreceptor system, which dis- played predicted functional characteristics when incubated with recipient adenylate cyclase systems from rat cerebral cortical and hypothalamic synaptic membranes and frog erythrocyte ghosts. We demonstrate functional exchange of photoreceptor compo- nents at each ofthree loci: the hormone receptor, the GTP-binding protein (GBP), and the catalytic moiety of adenylate cyclase. Il- luminated (but not unilluminated) rhodopsin was found to mimic the hormone-receptor complex, causing GTP-dependent activa- tion of adenylate cyclase. The photoreceptor GBP complexed with guanosine 5'-[8,yJimidotriphosphate (p[NH]ppG) produced a marked activation of recipient adenylate cyclase systems. Much smaller activation was observed when GBP was not complexed with p[NH]ppG. A heat-stable photoreceptor phosphodiesterase inhibitor reduced both basal and Mn2+-activated adenylate cy- clase activities and this inhibition was reversed by photoreceptor GBP-p[NH]ppG. These data demonstrate a remarkable functional compatibility between subunits of both systems and furthermore imply that specialized peptide domains responsible for protein- protein interactions are highly conserved. In 1977 we reported detailed similarities between the activation mechanisms for the light-activated photoreceptor 3',5'-cycic- GMP phosphodiesterase (PDEase; 3',5'-cyclic-nucleotide 5'- nucleotidohydrolase, EC 3.1.4.17) and the hormone-activated adenylate cyclase [ATP pyrophosphate-lyase (cyclizing), EC 4.6.1.1] (1). In retinal rods, the binding of GTP to a specific protein was found to generate a nucleotide-protein complex that activates photoreceptor PDEase. This mechanism is virtually indistinguishable from that demonstrated for the activation of adenylate cyclase by a variety of hormones including epineph- rine and glucagon (2). Many additional similarities between the two systems have been extensively reviewed (3-6) and include the following: (i) an ordered, two-part switching mechanism wherein presentation of a specific signal (light or a hormone) must precede formation of the GTP-protein complex that acti- vates the catalytic moiety; (ii) in both systems, activation is ter- minated by hydrolysis of bound GTP and the hydrolysis-resist- ant analogue, guanosine 5'-[,B, y]imidotriphosphate (p[NH]ppG), produces persistent activation; (iii) the concentrations of gua- nine nucleotides that produce half-maximal activation are com- parable (<1 tLM); (iv) both systems can also be activated by either fluoride or limited trypsin proteolysis. In view of the many profound similarities between the two activation mechanisms it was reasonable to inquire whether one could regulate adenylate cyclase by the addition of correspond- ing components from the photoreceptor system. In this report, we describe interchangeability at each of three discrete loci in the activation sequence: (i) the signal receptor, (ii) the GTP- binding protein (GBP), and (iii) the catalytic moiety. At each of these loci, we have found functional interchangeability of reg- ulatory components from either mammalian or amphibian ade- nylate cyclase systems. These findings demonstrate a striking functional compatibility between the systems and further sug- gest an extraordinary degree of structural conservation sur- rounding the evolutionary development of these regulatory components. MATERIALS AND METHODS Cerebral Cortical and Hypothalamic Synaptic Membranes. Synaptic membrane-enriched fractions were prepared as de- scribed (7) and stored under liquid N2. After thawing, mem- branes (1 ml) were suspended in S ml of ice-cold 10 mM Hepes, pH 7.5/5 mM MgSO41 mM dithiothreitol, centrifuged (12,000 x g, 10 min), and resuspended in 1.5 ml of the same buffer. Adenylate cyclase was assayed as described (7) except that mem- branes were added in the Hepes/MgSO4dithiothreitol buffer and the reaction volume was 100 1.l. Hypothalamic membranes were prepared and their adenylate cyclase was assayed as de- scribed (8). Amphibian Erythrocyte Membranes. Heparinized (0.06 mg/ ml) frog blood was collected by cardiac puncture and the buffy coat was removed by washing (three times) in citrated Tris-buff- ered saline. Erythrocytes were lysed (30 min, 0QC) in 10 mM TrisEICl/1 mM MgSO4, pH 8.0 (1 ml of packed cells per 100 ml of buffer). The membranes were pelleted (12,000 X g, 10 min), washed twice in 75 mM Tris.HCI/25 mM MgSO4, pH 8.0, and suspended in that buffer. The levels of GBP were reduced in erythrocyte membranes as follows (9): Membranes (1.5 mg of protein per ml) were in- cubated (45 min, 0C) with 15 ml of 1 mM EDTA/0.1 mM di- thiothrmtol and then centrifuged (10,000 x g, 15 min). This cycle was repeated. Membranes were resuspended in 500 uLd of 75 mM Tris HCI/25 mM Mg0O4, pH 8.0. Adenylate cyclase in erythrocyte membranes was measured for 15 min at 300C in Abbreviations: p[NH]ppG, guanosine 5'-[f3, y]imidotriphosphate; PDEase, phosphodiesterase; GBP, GTP-binding protein; ROS, rod outer segment. * Present address: Division of Life Sciences, Los Alamos National Lab- oratory, Los Alamos, NM 87545. 3408 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertise- ment" in accordance with 18 U. S. C. §1734 solely to indicate this fact.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Proc. Nati Acad. Sci. USAVol. 79, pp. 3408-3412, June 1982Biochemistry
Functional exchange of components between light-activatedphotoreceptor phosphodiesterase and hormone-activatedadenylate cyclase systems
(rhodopsin/GTP-binding protein/phosphodiesterase inhibitor/cerebral cortex synaptic membranes/frog erythrocyte ghosts)
MARK W. BITENSKY*, MARCIA A. WHEELER*, MARK M. RASENICK, AKIO YAMAZAKI*, PETER J. STEIN,KAREN R. HALLIDAY, AND GEORGE L. WHEELER*Yale University School of Medicine, Department of Pathology, New Haven, Connecticut 06510
Communicated by Lewis Thomas, February 11, 1982
ABSTRACT Previous studies have noted profound similaritiesbetween the regulation of light-activated 3',5'-cyclic nucleotidephosphodiesterase (3',5'-cyclic-nucleotide 5'-nucleotidohydro-lase, EC 3.1.4.17) in retinal rods and hormone-activated ade-nylate cyclase [ATP pyrophosphate-lyase (cyclizing), EC 4.6.1.1]in a variety of tissues. We report here the functional exchange ofcomponents isolated from the photoreceptor system, which dis-played predicted functional characteristics when incubated withrecipient adenylate cyclase systems from rat cerebral cortical andhypothalamic synaptic membranes and frog erythrocyte ghosts.We demonstrate functional exchange of photoreceptor compo-nents at each ofthree loci: the hormone receptor, the GTP-bindingprotein (GBP), and the catalytic moiety of adenylate cyclase. Il-luminated (but not unilluminated) rhodopsin was found to mimicthe hormone-receptor complex, causing GTP-dependent activa-tion ofadenylate cyclase. The photoreceptor GBP complexed withguanosine 5'-[8,yJimidotriphosphate (p[NH]ppG) produced amarked activation of recipient adenylate cyclase systems. Muchsmaller activation was observed when GBP was not complexedwith p[NH]ppG. A heat-stable photoreceptor phosphodiesteraseinhibitor reduced both basal and Mn2+-activated adenylate cy-clase activities and this inhibition was reversed by photoreceptorGBP-p[NH]ppG. These data demonstrate a remarkable functionalcompatibility between subunits of both systems and furthermoreimply that specialized peptide domains responsible for protein-protein interactions are highly conserved.
In 1977 we reported detailed similarities between the activationmechanisms for the light-activated photoreceptor 3',5'-cycic-GMP phosphodiesterase (PDEase; 3',5'-cyclic-nucleotide 5'-nucleotidohydrolase, EC 3.1.4.17) and the hormone-activatedadenylate cyclase [ATP pyrophosphate-lyase (cyclizing), EC4.6.1.1] (1). In retinal rods, the binding of GTP to a specificprotein was found to generate a nucleotide-protein complex thatactivates photoreceptor PDEase. This mechanism is virtuallyindistinguishable from that demonstrated for the activation ofadenylate cyclase by a variety of hormones including epineph-rine and glucagon (2). Many additional similarities between thetwo systems have been extensively reviewed (3-6) and includethe following: (i) an ordered, two-part switching mechanismwherein presentation of a specific signal (light or a hormone)must precede formation of the GTP-protein complex that acti-vates the catalytic moiety; (ii) in both systems, activation is ter-minated by hydrolysis ofbound GTP and the hydrolysis-resist-ant analogue, guanosine 5'-[,B, y]imidotriphosphate (p[NH]ppG),produces persistent activation; (iii) the concentrations of gua-nine nucleotides that produce half-maximal activation are com-
parable (<1 tLM); (iv) both systems can also be activated byeither fluoride or limited trypsin proteolysis.
In view of the many profound similarities between the twoactivation mechanisms it was reasonable to inquire whether onecould regulate adenylate cyclase by the addition ofcorrespond-ing components from the photoreceptor system. In this report,we describe interchangeability at each of three discrete loci inthe activation sequence: (i) the signal receptor, (ii) the GTP-binding protein (GBP), and (iii) the catalytic moiety. At each ofthese loci, we have found functional interchangeability of reg-ulatory components from either mammalian or amphibian ade-nylate cyclase systems. These findings demonstrate a strikingfunctional compatibility between the systems and further sug-gest an extraordinary degree of structural conservation sur-rounding the evolutionary development of these regulatorycomponents.
MATERIALS AND METHODSCerebral Cortical and Hypothalamic Synaptic Membranes.
Synaptic membrane-enriched fractions were prepared as de-scribed (7) and stored under liquid N2. After thawing, mem-branes (1 ml) were suspended inS ml ofice-cold 10mM Hepes,pH 7.5/5mM MgSO41 mM dithiothreitol, centrifuged (12,000x g, 10 min), and resuspended in 1.5 ml of the same buffer.Adenylate cyclase was assayed as described (7) except that mem-branes were added in the Hepes/MgSO4dithiothreitol bufferand the reaction volume was 100 1.l. Hypothalamic membraneswere prepared and their adenylate cyclase was assayed as de-scribed (8).
Amphibian Erythrocyte Membranes. Heparinized (0.06mg/ml) frog blood was collected by cardiac puncture and the buffycoat was removed by washing (three times) in citrated Tris-buff-ered saline. Erythrocytes were lysed (30 min, 0QC) in 10 mMTrisEICl/1 mM MgSO4, pH 8.0 (1 ml of packed cells per 100ml of buffer). The membranes were pelleted (12,000 X g, 10min), washed twice in 75mM Tris.HCI/25mM MgSO4, pH 8.0,and suspended in that buffer.The levels of GBP were reduced in erythrocyte membranes
as follows (9): Membranes (1.5 mg of protein per ml) were in-cubated (45 min, 0C) with 15 ml of 1 mM EDTA/0.1 mM di-thiothrmtol and then centrifuged (10,000 x g, 15 min). Thiscycle was repeated. Membranes were resuspended in 500 uLdof 75 mM Tris HCI/25 mM Mg0O4, pH 8.0. Adenylate cyclasein erythrocyte membranes was measured for 15 min at 300C in
Abbreviations: p[NH]ppG, guanosine 5'-[f3, y]imidotriphosphate;PDEase, phosphodiesterase; GBP, GTP-binding protein; ROS, rodouter segment.* Present address: Division of Life Sciences, Los Alamos National Lab-oratory, Los Alamos, NM 87545.
3408
The publication costs ofthis article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U. S. C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 79 (1982) 3409
a reaction mix containing 50 mM Tris maleate at pH 8.0, 1 mMdithiothreitol, 0.62 mM EDTA, 0.8 mM 3-isobutyl-1-meth-ylxanthine, 6 mM MgSO4, 0.2 mM ATP, pyruvate kinase (0.05mg/ml), 60mM phosphoenolpyruvate, and myokinase (0.1 mg/ml).
Assay for cAMP Produced. cAMP was assayed by a modi-fication of the method of Brown et al. (10) using rabbit musclecAMP-binding protein. Values (pmol of cAMP/min per mg ofprotein) were corrected by subtraction of a boiled membraneblank. Recovery of cAMP added to a boiled membrane blankwas greater than 98%. All rates were linear for more than 15min and the ATP concentration did not decline measurably formore than 15 min as measured with luciferase (11).
Preparation of Photoreceptor GBP Fraction. GBP was pre-pared from Bufo marinus rod outer segment (ROS) membranesthat had been washed (four times) with 150mM Tris.HCl/5mMdithiothreitol, pH 7.3 (1 ml per 25 retinas), and (five times) with5 mM Tris.HCl/5 mM dithiothreitol, pH 7.3. All washes were
at 20,000 x g for 15 min. The GBP fraction was then preparedby washing the ROS with 100 ,uM GTP/20 mM Tris HCI/5mMdithiothreitol, pH 7.3. The supernatant from this wash is GBPand contains three polypeptides (39, 37, and approximately 10kilodaltons by NaDodSO4/polyacrylamide gel electrophoresis)known to copurify with light-activated ROS GTPase activity(12). The 39-kilodalton polypeptide is known to contain theGTP-binding site (13). In both GBP and the GBP-p[NH]ppGcomplex fraction (see below), these peptides account for >99%of total protein when analyzed densitometrically on Coomassieblue-stained NaDodSO4 gels.GBP-p[NH]ppG complex was prepared by further incubat-
ing the GTP-washed ROS membranes (see above) with 10 ,.Mp[NH]ppG/20 mM Tris.HCl/5 mM dithiothreitol, pH 7.3, for12-14 hours followed by centrifugation (as above). The super-natant containing GBP-p[NH]ppG was subjected to filtrationthrough an Amicon B15 concentrator (with 10 mM Tris HCl,pH 7.5/1 mM dithiothreitol) to remove unbound p[NH]ppG.
Preparation of Urea-Washed Rhodopsin-Containing Pho-toreceptor Membranes. Dark ROS membranes from Rana ca-
tesbiana were extracted with EDTA as previously described(14). After extraction, the pellet was resuspended in 30 vol of10 mM Tris.HCl/1 mM EDTA/1 mM dithiothreitol/5 M urea,
pH 7.5. The suspension was passed through a 21-gauge needlethree times, kept on ice for 60 min, and centrifuged (81,000x g, 4°C, 60 min). The resulting pellet was washed (three times)in 50 vol of 200 mM Tris HCl, pH 7.5/20 mM MgSO4/1 mMdithiothreitol and resuspended in the same buffer. Such "urea-washed" ROS membranes contain no PDEase or GTPaseactivity.
Preparation of Photoreceptor Inhibitor Fraction. Lyophi-lized EDTA extracts ofdark ROS from R. catesbiana were com-
bined with the urea-treated membranes and centrifuged(48,000 X g, 40C, 10 min). The resulting supernatant was heated(800C, 3 min) and centrifuged (as above). This heated inhibitoryfactor does not contain PDEase activity, GTPase activity, or
rhodopsin, but fully retains its ability to inhibit photoreceptorPDEase (15).
Other. Additional enzyme assays were cAMP PDEase (16),GTPase (17), luciferase (11), and ATPase (11). Proteins were
determined with the Bio-Rad assay (18).
EXPERIMENTAL
Substitution for the Hormone-Receptor Complex by Illu-minated Rhodopsin. The counterpart of hormone binding toreceptor in the cyclase system is illumination of rhodopsin inthe rod PDEase system. Table 1 shows the effects of highly
Table 1. Activation of adenylate cyclase by illuminatedrhodopsin and p[NH]ppG
Adenylate cyclase,pmol/min per mgUnilluminated Illuminated
Additions No rhodopsin rhodopsin rhodopsinRat cerebral cortex synaptic membranes
None 26 ± 3.1 14 ± 0.8 40 ± 0.5p[NH]ppG
(20 AM) 196 ± 8 252 ± 11 512 ± 20
Frog erythrocyte membranesNone 4.2 ± 0.41 4.0 ± 0.26 3.8 ± 0.1p[NH]ppG
(0.5 AM) 4.0 ± 0.19 4.9 ± 0.11 6.6 ± 0.1
Illuminated bovine rhodopsin was prepared from unilluminated rho-dopsin by full bleaching at ice temperature. All components, includingthe cyclase reaction mixture, were incubated for 25 min at 300C inabsolute darkness. For the rat cerebral cortex membranes ATP was 0.5mM, and the data are averages of duplicate values from one of fourrepresentative experiments. For the frog erythrocyte membranes ATPwas 0.1 mM, and the data shown are the means of three experimentsdone in triplicate. With both membranes, the ratio of rhodopsin-con-taining membranes to recipient cyclase membranes was 1:4 by totalprotein. Adenylate cyclase activities are expressed as pmol of cAMPformed per min per mg of recipient membrane protein (±SEM).
purified illuminated and unilluminated bovine rhodopsin (19)on mammalian and amphibian cyclase systems. With the mam-malian system, we note that the presence of p[NH]ppG is nec-essary for activation of cyclase by added rhodopsin, and acti-vation of cyclase with illuminated rhodopsin was 160% abovethat activity observed with p[NH]ppG alone. With unillumi-nated rhodopsin, we found little (20%) cyclase activation abovethat observed with p[NH]ppG alone. With the amphibian sys-tem, we again note preferential p[NH]ppG-dependent activa-tion of cyclase with illuminated rhodopsin. Activation of cyclaseby both highly purified bovine rhodopsin and by rhodopsin inurea-washed amphibian disc membranes was maximal at a rho-dopsin concentration of 1 ,M.
Illuminated rhodopsin (in urea-washed, frog disc mem-branes) activated hypothalamic cyclase above the levels pro-duced by p[NH]ppG alone at all p[NH]ppG concentrations ex-amined between 0.05 and 5 ,M. Maximal activation of cyclaseby rhodopsin (130%) was observed at 0.2 ,uM p[NH]ppG. At2 AM p[NH]ppG, illuminated rhodopsin activated frog eryth-rocyte cyclase by 10.7 pmol/min per mg of protein, which was75% of the activation produced by isoproterenol (14.2 pmol/min per mg of protein). Maximal activation of cortical or hy-pothalamic cyclase by admixture of rhodopsin vesicles wasachieved after a preincubation period of 25 min. Although de-tergent was unnecessary for cyclase activation by rhodopsin-containing membranes, 0.6% Brij-35 was found to enhance thisp[NH]ppG-dependent activation of cyclase by 20%. Neitherretinal nor phosphatidylcholine, alone or combined in vesicles,showed any effect on recipient cyclase systems at concentrationsequal to or exceeding those present in the rhodopsin prepara-tions. Finally, adenylate cyclase activity was undetectable in theurea-washed (amphibian) and reconstituted (bovine) rhodopsinpreparations.
Functional Interaction of Photoreceptor GBP with Recipi-ent Adenylate Cyclase Systems. We have examined the inter-action of photoreceptor GBP with a frog erythrocyte cyclasepreparation (Table 2). We note that the rod GBP-p[NH]ppGcomplex at 5 pug/ml is more effective as an activator of cyclasethan GBP at 50 ,ug/ml alone or in the presence of unbound
Biochemistry: Bitensky et al.
3410 Biochemistry: Bitensky et al
Table 2. Activation of frog erythrocyte adenylate cyclase by photoreceptor GBP alone or complexedwith p[NHlppG
Adenylate cyclase, pmol/min per mg
No GBP GBP GBP-p[NH]ppGAgonist GBP* (50 ,g/mi) (5,ug/ml) (5 Mg/ml)
None 7.8 ± 0.5 9.3 ± 0.2 8 ± 0.3 118 ± 3Isoproterenol(1UM) 14.3 ± 1.3 23 ± 1 11 ± 0.5 149 ± 4Isoproterenol (1 M)+ p[NH ppG (1 AM 85 ± 3 150 ± 4 73 ± 1 213 ± 6
p[NH]ppG(1 PM) 51.1 ± 2 60 ± 0.8t 59 ± 1.3 160 ± 3
Frog erythrocyte membranes were preincubated (10 min at 00C, then 10 min at 3000) with either GBP(50 pg/ml or 5 ug/ml) or the GBP-p[NH]ppG complex (5 pg/mi) (final concentrations). After the prein-cubation step, the cyclase reaction mixtures (containing 0.2 mM ATP and the above agonists) were addedto the preincubated membrane/GBP mixtures, and incubation was continued for an additional 15 minat 3000. Final concentration of the erythrocyte membranes was 1.5 mg of protein per ml. Results aremeans (±SEM) of duplicate experiments each done in triplicate.* In all cases, use of boiled GBP p[NH]ppG at 5 pg/ml in the preincubation step gave cyclase activitiesindistinguishable from those given in this column (buffer alone.)
t When the p[NH]ppG (1 pM) was added in the preincubation step the cyclase activity increased to 72 pmol/min per mg of protein.
p[NH]ppG. At a concentration of 5 ,g/ml, GBP did not acti-vate erythrocyte cyclase under any conditions examined (Table2). Neither GBP nor GBP-p[NH]ppG had cyclase activity them-selves, and when heated (90°C, 3 min) they did not activateadenylate cyclase.
Incubation of erythrocyte membranes with photoreceptorGBP alone or in combination with either isoproterenol or un-bound p[NH]ppG produced only minimal cyclase activation.However, when GBP was incubated with both p[NH]ppG andisoproterenol there was a striking activation (85 to 150 pmol/min per mg of protein, Table 2) of the recipient erythrocytecyclase. These data suggest that the erythrocyte cyclase hor-mone-receptor complex can enhance binding ofp[NH]ppG tophotoreceptor GBP. Similarily, additional activation by hor-mone in the presence of GBP-p[NH]ppG (149 to 213 pmol/min per mg of protein) indicates that, in the preparation used,not all GBP was previously complexed with p[NH]ppG. Thesedata also show that the recipient erythrocyte cyclase system it-
Table 3. Effect of photoreceptor GBP-p[NH]ppG complex onadenylate cyclase activity in EDTA/dithiothreitol-treatedfrog erythrocyte membranes
Adenylate cyclase,pmol/min per mg
EDTA/EDTA/ dithiothreitol-
dithiothreitol- treatedUntreated treated membranes +
Agonist membranes membranes GBP-p[NH]ppGNone 7 ± 0.5 14 ± 1 20 ± 1p[NH]ppG(1 M) 28±0.6 19±0.8 27±1.3
Isoproterenol(10PM) 4± 1.3 16±0.4 34± 1.3
Isoproterenol+ p[NH~ppG 52 ± 1 22 ± 1.5 55 ± 1*
EDTA/dithiothreitol-treated membranes (GBP-depleted) were pre-pared from erythrocyte membranes (1.8 mg/ml) and the treated mem-branes (1.5 mg/ml, final protein concentration) were incubated withGBP-p[NH]ppG (6 pug/ml) for 10 min at 00C and then 10 min at 300C.Reaction mix containing 0.2 mM ATP and the above agonists wasadded and incubation was continued (15 min, 3000). Reaction wasstopped andcAMP content was determined. Values are means (±SEM)of triplicate determinations from one of three similar experiments.*Value of EDTA-treated membranes and heated (900C, 3 min)GBP-p[NH]ppG complex was 22 pmol/min per mg of protein.
self is GBP limited because addition of the photoreceptorGBP-p[NH]ppG complex markedly enhanced cyclase activity.
Parallel experiments were carried out with an EDTA-washedGBP-depleted recipient erythrocyte cyclase system (Table 3).This treatment markedly diminished response to p[NH]ppGand isoproterenol. Although cyclase activity was diminished byEDTA washing, added rod GBP-p[NH]ppG fully restored cy-clase activity to the level observed in the unwashed system.
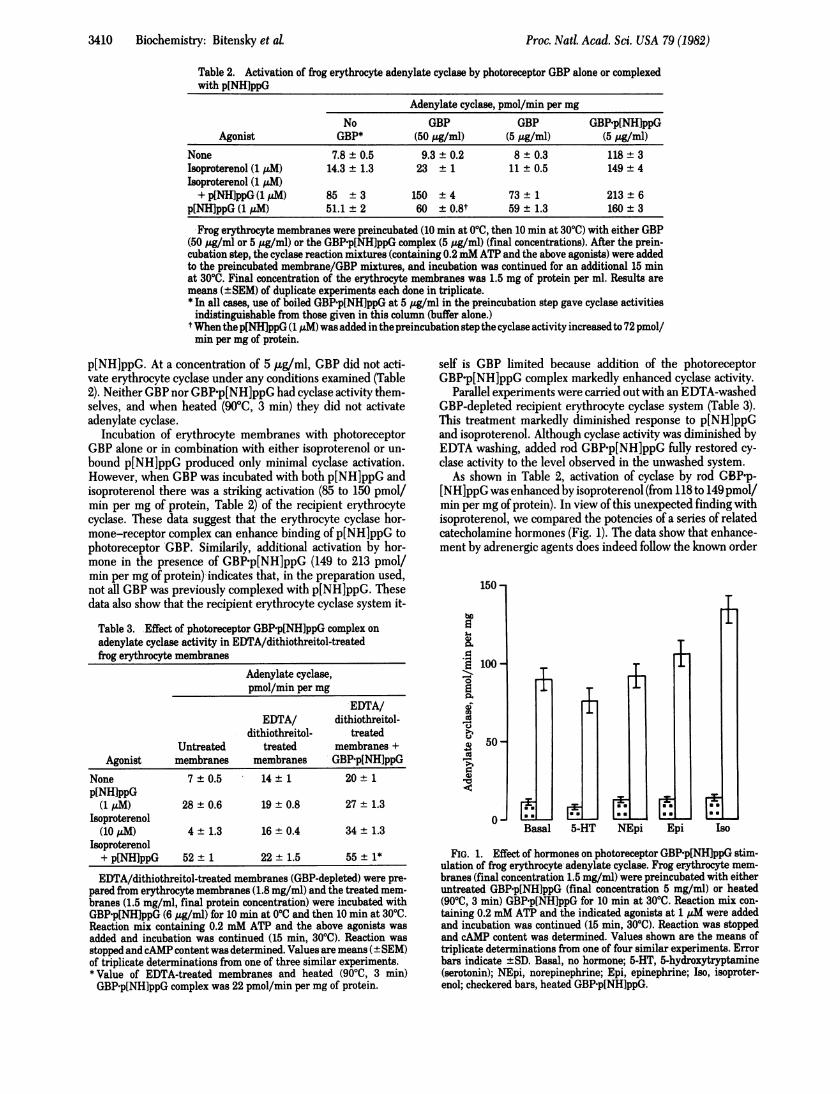
As shown in Table 2, activation of cyclase by rod GBP-p-[NH]ppG was enhanced by isoproterenol (from 118 to 149pmoVmin per mg of protein). In view of this unexpected finding withisoproterenol, we compared the potencies of a series of relatedcatecholamine hormones (Fig. 1). The data show that enhance-ment by adrenergic agents does indeed follow the known order
150-
100-to
Ei0
cd
50-¢U
'U4G)
0- BaaBasal
ni
5-HT
t
NEpi Epi Iso
FIG. 1. Effect of hormones on photoreceptor GBP'p[NH]ppG stim-ulation of frog erythrocyte adenylate cyclase. Frog erythrocyte mem-branes (final concentration 1.5 mg/ml) were preincubated with eitheruntreated GBP-p[NH]ppG (final concentration 5 mg/ml) or heated(9000, 3 min) GBP-p[NH]ppG for 10 min at 3000. Reaction mix con-taining 0.2 mM ATP and the indicated agonists at 1 ,uM were addedand incubation was continued (15 min, 3000). Reaction was stoppedand cAMP content was determined. Values shown are the means oftriplicate determinations from one of four similar experiments. Errorbars indicate ±SD. Basal, no hormone; 5-HT, 5-hydroxytryptamine(serotonin); NEpi, norepinephrine; Epi, epinephrine; Iso, isoproter-enol; checkered bars, heated GBP-p[NH]ppG.
Proc. Natl.-Acad- Sci. USA 79 (1982)
11

Proc. Natl. Acad. Sci. USA 79 (1982) 3411
of potency (20) shown by these agents in the frog erythrocyteadenylate cyclase system (isoproterenol > epinephrine > nor-epinephrine > 5-hydroxytryptamine). The data in Fig. 1 alsoshow that heating (90TC, 5 min) of the added GBP-p[NH]ppGdestroyed its capacity to activate cyclase with each of the hor-mones tested.
In studies with rat cerebral cortex synaptic membranes, wefound that heating (30 min, 420C) reduced p[NH]ppG-activatedcyclase by 85% while reducing Mn2+-activated cyclase by only40% (21). Activation by Mn2+ is independent of GBP and thusserves as an index of cyclase catalytic function. In heated prep-arations there was a 177% activation of adenylate cyclase (from31 to 86 pmol/min per mg of protein) by the photoreceptorGBPp[NH]ppG. This was comparable to the activation (165%)of this cyclase preparation that we observed with a GBP-p-[NH]ppG complex obtained from rat cerebral cortex synapticmembranes (21).
Functional Interaction of a Photoreceptor PDEase Inhibi-tory Factor with Recipient Adenylate Cyclase. The photore-ceptor system contains a heat-stable, trypsin-labile inhibitor oflight-activated PDEase (22). The effect of this inhibitor is re-versed by the action ofthe photoreceptor GBPp[NH]ppG com-plex (14, 23). We examined the effect of this photoreceptor-derived inhibitor on cerebral cortex synaptic membrane ade-nylate cyclase (Table 4). In these experiments we use both Mn2+and F- as activators of cyclase. Mn2+ stimulation is often usedas a measure of cyclase catalytic activity (independent of GBPregulation) (21, 24), whereas activation by F- requires the pres-ence of the cyclase-associated GBP (7, 24, 25).
The data in Table 4 reflect the known properties exhibitedby the inhibitor in the photoreceptor system and the knownproperties exhibited by Mn2+ and F- in the cyclase system. Thepresence of inhibitor does not block activation of cyclase bymodulators that depend upon the GBP (i.e., p[NH]ppG andF-). This finding reflects the known capacity of the rodGBP-p[NH]ppG complex to reverse the effects of the photo-receptor inhibitor on rod PDEase. The inhibitor does, however,markedly reduce both basal cyclase activity and Mn2+-stimu-lated cyclase activity.
Similar results were found with frog erythrocyte cyclase.Here inhibitor (0.2 mg/ml) reduced basal cyclase activity by82% and Mn2+-activated cyclase activity by 37%. In contrast,
Table 4. Effect of photoreceptor PDEase inhibitor on cerebralcortex synaptic membrane adenylate cyclase
Adenylate cyclase,pmol/min per mg
No PDEase PDEase Inhibition,Agonist inhibitor inhibitor %
None 43 ± 2 15 ± 0.7 65p[NH]ppG (5 ,uM) 206 ± 7 213 ± 13 0NaF (lOmM) 144 ± 6 126 ± 4 12Mn2+ (5mM) 246 ± 4 114 ± 9 54Mn2+ (2 mM) 178 ± 6 29 + 5 84
Synaptic membranes (2.1 mg/ml final protein concentration) wereincubated with PDEase inhibitor (200 ,ug/ml) and the above agonists(10 min at 0°C, then 10 min at 300C). Reaction mix containing 0.5 mMATP was added and incubation was continued (10 min at 3000). Thecyclase reaction was stopped by heating (900C, 3 min); the sampleswere then centrifuged (10,000 x g, 10 min) and an aliquot of the su-pernatant was treated with trypsin (75 ,ug/ml, 10 min, 370C), followedby trypsin inhibitor (375 tzg/ml). The samples were heated (900C, 3min) and aliquots were assayed for cAMP content. The above trypsintreatment prevented the PDEase inhibitor from interfering with thecAMP binding assay. Values shown are the means (±SEM) of tripli-cate determinations from one of four similar experiments.
the photoreceptor inhibitor did not diminish activation oferyth-rocyte cyclase by p[NH]ppG.
Using rat cerebral cortex synaptic membranes as a recipientcyclase system, we found that the PDEase inhibitor could re-verse Mn2+ activation by 54%, from 492 to 228 pmol/min permg of protein. This inhibition itself was then reversed by 91%(from 228 to 448 pmol/min per mg ofprotein) by addition ofthephotoreceptor GBP-p[NH]ppG complex.
The photoreceptor PDEase inhibitor did not inhibit frogerythrocyte membrane ATPase or firefly luciferase, nor did itaffect the ATP concentration during the cyclase assay. The ac-tivity of synaptic membrane PDEase (cAMP or cGMP) was notinfluenced by photoreceptor inhibitor.
DISCUSSIONIn these experiments, which explore the possibility of substi-tuting components between rod PDEase and hormone-acti-vated cyclase, a major concern was to separate specific fromnonspecific effects. The components ofthe cyclase system, withthe exception ofGBP (26), have not yet yielded to purification.Furthermore, the cyclase system is known to be a multi-subunitenzyme complex in which effects have been demonstrated witha wide range of modulators. Thus, the demonstration of eitherincreases or decreases in cyclase activity assumes regulatorysignificance only when such changes are accompanied by evi-dence ofbiochemical specificity. The results observed here, bythe simple mixing and preincubation of photoreceptor com-ponents with recipient adenylate cyclase membranes, displayedpredicted specific properties of the exchanged components.
Substitution of illuminated rhodopsin for the hormone-receptor complex of recipient cyclase systems displayed the fol-lowing functional specificity: (i) Both light and p[NH]ppG arerequired for rhodopsin activation of cyclase. (ii) Neither phos-phatidylcholine nor retinal could be responsible for the ob-served cyclase activation because in the absence ofopsin, thesecompounds were without effect. (iii) Illuminated rhodopsin in-teracts with the GBPs of the recipient cyclase system becausethe cyclase activation by rhodopsin depends upon the presenceof p[NH]ppG. The mechanisms that allow interaction betweenthe rhodopsin-containing membranes and the recipient cyclasesystem, and the molecular arrangements within the active com-plex, are unknown.
Photoreceptor GBP-p[NH]ppG displayed the following func-tional specificity in recipient cyclase systems: (i) In experimentsthat selectively compromise the GBP ofthe recipient adenylatecyclase system (EDTA washing in frog erythrocytes and heatingin cortical synaptosomes) the recipient adenylate cyclase activ-ity was enhanced by addition of the rod GBP-p[NH]ppG com-plex and much less by GBP alone (Table 3). These data indicatethat the amount of GBP in the recipient system was limiting.(ii) The combination of isoproterenol, uncomplexed photore-ceptor GBP, and free p[NH]ppG activated recipient erythro-cyte cyclase better than did any two ofthese agents alone. Thesedata suggest that the erythrocyte hormone-receptor complexcan interact with photoreceptor GBP and enhance formation ofadditional GBP-p[NH]ppG complex (Table 2). (iii) The addi-tional activation of frog erythrocyte membrane cyclase by rodGBP-p[NH]ppG due to the presence of catecholamines followsthe known order of potency for these adrenergic agents (Fig.1). These data indicate that occupation of the 3-adrenergic re-ceptor by agonist can facilitate functional intercalation of ex-ogenous GBP-p[NH]ppG. (iv) When the rod GBP-p[NH]ppGwas heated (90TC, 3 min) it failed to activate both adenylatecyclase and rod PDEase.
Inhibition of cyclase by rod PDEase inhibitor displays the
Biochemistry: Bitensky et al.

3412 Biochemistry: Bitensky et aL
following functional specificity: (i) Rod PDEase inhibitor inhib-ited both Mn2+-stimulated and basal cyclase activities but didnot significantly reduce F- or p[NH]ppG-dependent cyclaseactivities (Table 4). This is in keeping with the finding that, inthe photoreceptor system, the GBP-p[NH]ppG overcomes theeffect of the inhibitor and also with the fact that F- activationof cyclase depends on the presence of a GBP (7, 24, 25). (ii)Addition of rod GBP-p[NH]ppG substantially reversed the in-hibition ofMn2+-activated cyclase by the rod PDEase inhibitor.
As a result of the photoreceptor inhibitor-recipient cyclaseinteractions we speculate that a homologous inhibitor proteinmay also function in the physiological (hormone-dependent)regulation of adenylate cyclase. While Mn2+ (at both 2 and 5mM) appears to overcome any endogenous inhibitor that func-tions as a component of the recipient cyclase systems studiedhere, Mn2+ activation of such cyclase systems was significantlydiminished by the inhibitor of rod PDEase.
Evidence for the existence ofspecialized articulating domainsresponsible for the protein-protein interactions of the cyclasesystem has recently been reported in experiments with S49lymphoma variants (27). These cell lines possess GBPs that aredefective in their ability to interact with either the hormonereceptor or the catalytic moiety. Such variants, however, retainverifiable functional expression through the other unalteredsite.The functional exchange of photoreceptor PDEase compo-
nents for components of the cyclase system reported herestrongly supports the idea that the analogous proteins containcertain specific domains that are structurally similar. For ex-ample, although rhodopsin and the epinephrine receptor areobviously adapted for very different signal recognition, theynevertheless interactwith very similar GBPs. Moreover, where-as phosphodiesterase and adenylate cyclase catalyze opposingreactions, they are both activated by very similar GBPGTPcomplexes.The existence of shared structural domains within corre-
sponding proteins of the two systems may have important ge-netic implications. It is interesting to speculate that the genesfor rhodopsin and the epinephrine receptor may exhibit do-mains of highly conserved nucleotide sequences that corre-spond to those peptide domains responsible for receptor-GBPinteractions as well as unshared sequences corresponding tothose domains responsible for specialized photon capture orhormone reception. Moreover, the finding of such assembliesofshared and unshared nucleotide sequences may provide cluesto the evolutionary mechanisms that underlie the formation ofnew genes. Such evolutionary mechanisms could facilitate thegeneration ofnew combinations of existing functional domains,perhaps separated by intervening sequences (28-30).
We gratefully acknowledge the technical support ofRosa Carney andsecretarial support of Bibiane Roberts. Purified bovine rhodopsin wasa generous gift of Dr. Wayne Hubbell. This work was supported at YaleUniversity under National Institutes of Health Grant R01-AM 20179.
1. Wheeler, G. L. & Bitensky, M. W. (1977) Proc. Natl. Acad. Sci.USA 74, 4238-4242.
2. Rodbell, M., Lin, M. C., Salomon, Y., Londos, C., Haywood, J.P., Martin, B. R., Rondell, M. & Berman, M. (1975) Adv. CyclicNucleotide Res. 5, 3-29.
3. Bitensky, M. W., Wheeler, G. L., Aloni, B., Vetury, S. & Ma-tuo, Y. (1978) Adv. Cyclic Nucleotide Res. 9, 553-572.
4. Shinozawa, T., Sen, I., Wheeler, G. L. & Bitensky, M. W. (1979)J. Supramol Struct. 10, 185-190.
5. Pober, J. A. & Bitensky, M. W. (1979) Advances in Cyclic Nu-cleotide Research, eds. Greengard, P. & Robison, G. A. (Raven,New York), Vol. 11, pp. 265-301.
6. Bitensky, M. W., Wheeler, G. L., Yamazaki, A., Rasenick, M.M. & Stein, P. J. (1981) in Molecular Mechanisms ofPhotorecep-tor Transduction, Current Topics in Membranes and Transport,eds. Bronner, F. & Kleinzeller, A. (Academic, New York), Vol.15, pp. 237-271.
7. Rasenick, M. M. & Bitensky, M. W. (1980) Proc. Natl. Acad. Sci.USA 77, 4628-4632.
8. Wheeler, M. A., Tishler, M. & Bitensky, M. W. (1982) BrainRes. 231, 387-398.
9. Pfeuffer, T. (1977) J. Biol. Chem. 252, 7224-7234.10. Brown, B., Albano, J., Ekins, R. & Scherzis, A. (1971) Biochem.
J. 121, 561-562.11. Kimmich, G., Randles, J. & Brand, J. (1975) Anal. Biochem. 69,
187-206.12. Kuhn, H. (1980) Nature (London) 283, 589.13. Uchida, S., Wheeler, G. L., Yamazaki, A. & Bitensky, M. W.
(1981) J. Cyclic Nucleotide Res. 7, 95-104.14. Yamazaki, A., Sen, I., Bitensky, M. W., Casnellie, J. E. &
Greengard, P. (1980)J. Biol Chem. 255, 11619-11624.15. Yamazaki, A., Bartucca, F., Ting, A. & Bitensky, M. W. (1982)
Proc. Natl. Acad. Sci. USA 79, 3702-3706.16. Miki, N., Keirns, J. J., Marcus, F. R., Freeman, J. & Bitensky,
M. W. (1973) Proc. Natl Acad. Sci. USA 70, 3820-3824.17. Neufeld, A. H. & Levy, H. M. (1969) J. Biol Chem. 244,
6493-6497.18. Bradford, M. (1976) AnaL Biochem. 72, 248-254.19. Hong, K. & Hubbell, W. L. (1972) Proc. Natl Acad. Sci. USA 69,
2617-2621.20. Mukherjee, C., Caron, M. G., Coverstone, M. & Lefkowitz, R.
J. (1975)J. Biol Chem. 250, 4869-4876.21. Rasenick, M. M., Stein, P. J. & Bitensky, M. W. (1981) Nature
(London) 294, 560-562.22. Dumler, I. L. & Etingoff, R. N. (1976) Biochim. Biophys. Acta
429, 474-484.23. Hurley, J. B. (1980) Biochem. Biophys. Res. Commun. 92,
505-510.24. Ross, E. & Gilman, A. G. (1980) Annu. Rev. Biochem. 49,
533-564.25. Downs, R. W., Spiegel, A. M., Singer, M., Reen, S. & Aurbach,
G. D. (1980) J. Biol Chem. 255, 949-954.26. Northup, J. K., Sternweis, P. C., Smigel, M. D., Schleifer, L.
S., Ross, E. M. & Gilman, A. G. (1980) Proc. Natl Acad. Sci. USA77, 6516-6520.
27. Bourne, H. R., Kaslow, D., Kaslow, H. R., Salomon, M. P. &Licko, V. (1981) Mol Pharmacol 20, 435-441.
28. Darnell, J. E., Jr. (1978) Science 202, 1257-1260.29. Crick, F. (1979) Science 204, 264-271.30. Rogers, J., Early, P., Carter, C., Calame, K., Bond, M., Hood,
L. & Wall, R. (1980) Cell 20, 303-312.
Proc. Natl. Acad. Sci. USA 79 (1982)
Related Documents











