Functional and metabolic characterization of hemocytes of the green mussel, Perna viridis: in vitro impacts of temperature Ludovic Donaghy, Aswani K. Volety * Department of Marine and Ecological Sciences, Coastal Watershed Institute, College of Arts and Science, Florida Gulf Coast University, 10501 FGCU Boulevard South, Fort Myers, FL 33965, USA article info Article history: Received 29 April 2011 Received in revised form 5 July 2011 Accepted 10 July 2011 Available online 20 July 2011 Keywords: Green mussel Perna viridis Hemocytes Temperature Florida abstract The green mussel, Perna viridis, is a bivalve mollusk native to Asia and was recently introduced to Florida, USA. Since its first observation in 1999 in Tampa Bay, Florida, green mussel population has expanded considerably, to reach the Atlantic coast of Florida, Georgia and South Carolina. Most of currently available studies about the ecology and biology of green mussels were performed in the Indian and Pacific oceans. Very recently, it has been suggested that due to a weak low temperature resistance, green mussels might have already reached the Northern edge of their distribution in the USA. However, there is currently an obvious lack of data about the adaptation capacities of Perna viridis to environmental conditions in Florida, especially at the physiological and cellular levels. In the present work, we deter- mined and characterized the populations of circulating hemocytes, and the cellular components of hemolymph involved in various physiological functions, including immunity. Two main populations were characterized, hyalinocytes and granulocytes. Granulocytes accounted for 60% of circulating cells, and displayed higher phagocytic capacities, lysosomal content and basal oxidative metabolism than hyali- nocytes. Hemocyte parameters were not influenced by the size of green mussels. In addition, hemocytes were subjected to acute temperature challenges (10, 20 and 30 C) and their immune-related functions and metabolism analyzed. Our results showed that 10 C represent a stressful condition for the Floridian green mussels, as depicted by a low phagocytosis capacity and an increase of oxidative metabolism. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction The green mussel Perna viridis is a bivalve mollusk native from the Indian coast and throughout the Indo-Pacific and Asia-Pacific. In the early to mid-1990s, populations were discovered along the coasts of Venezuela, Trinidad, and Jamaica [1,2]. In 1999, green mussels genetically related to the ones previously described in the Caribbean were observed in Tampa Bay, Florida, USA [3]. This was the first reported occurrence of this Indo-Pacific marine bivalve in North America. Green mussel population has then expanded considerably, most likely through local reproduction and settlement, along the coastal regions of southwestern Florida. Recruits of Perna viridis can now also be found on the Atlantic coast of Florida, Georgia and South Carolina [1,2,4]. Green mussels are known biofoulers of boats, navi- gational aids but also of submerged power plants and other indus- tries infrastructures, potentially clogging water intakes and outflow pipes [1]. Furthermore, this non-native species seems to have the potential to displace local native species such as the Eastern oyster, Crassostrea virginica [5]. The long-term ecological and economic impact of the green mussel introduction to Florida might then be considerable but cannot be predicted at this time. Due to their origin, the population dynamics [6e8] as well as the reproduction and growth [9e11] of green mussels were, until now, mostly studied in Asia, where P. viridis are widely used for the biomonitoring of coastal environments. Indeed, many studies measured the tissue levels of contaminants such as heavy metals, PAHs, PCBs or pesticides in P. viridis [12e15]. Similarly, the toxico- logical impacts of such contaminations on the green mussels were investigated, once again mostly in Asia [16e21]. The populations of green mussels from Florida were, until now, almost exclusively studied for their geographic distribution patterns, in order to understand their origin and to predict the population dynamics [1e4]. Very recently, Urian et al. [22] investigated on the effects of seawater temperature on the survival of Floridian P. viridis, and concluded that due to a weak low temperature resistance, green mussels might have already reached the Northern edge of their distribution in USA. Indeed, temperature is a predominant factor determining the geographic distribution of marine invertebrates * Corresponding author. Tel.: þ1 239 590 7216; fax: þ1 239 590 7200. E-mail address: [email protected] (A.K. Volety). Contents lists available at ScienceDirect Fish & Shellfish Immunology journal homepage: www.elsevier.com/locate/fsi 1050-4648/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.fsi.2011.07.018 Fish & Shellfish Immunology 31 (2011) 808e814

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Fish & Shellfish Immunology 31 (2011) 808e814
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate/ fs i
Functional and metabolic characterization of hemocytes of the green mussel,Perna viridis: in vitro impacts of temperature
Ludovic Donaghy, Aswani K. Volety*
Department of Marine and Ecological Sciences, Coastal Watershed Institute, College of Arts and Science, Florida Gulf Coast University,10501 FGCU Boulevard South, Fort Myers, FL 33965, USA
a r t i c l e i n f o
Article history:Received 29 April 2011Received in revised form5 July 2011Accepted 10 July 2011Available online 20 July 2011
Keywords:Green musselPerna viridisHemocytesTemperatureFlorida
* Corresponding author. Tel.: þ1 239 590 7216; faxE-mail address: [email protected] (A.K. Volety).
1050-4648/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.fsi.2011.07.018
a b s t r a c t
The green mussel, Perna viridis, is a bivalve mollusk native to Asia and was recently introduced to Florida,USA. Since its first observation in 1999 in Tampa Bay, Florida, green mussel population has expandedconsiderably, to reach the Atlantic coast of Florida, Georgia and South Carolina. Most of currentlyavailable studies about the ecology and biology of green mussels were performed in the Indian andPacific oceans. Very recently, it has been suggested that due to a weak low temperature resistance, greenmussels might have already reached the Northern edge of their distribution in the USA. However, there iscurrently an obvious lack of data about the adaptation capacities of Perna viridis to environmentalconditions in Florida, especially at the physiological and cellular levels. In the present work, we deter-mined and characterized the populations of circulating hemocytes, and the cellular components ofhemolymph involved in various physiological functions, including immunity. Twomain populations werecharacterized, hyalinocytes and granulocytes. Granulocytes accounted for 60% of circulating cells, anddisplayed higher phagocytic capacities, lysosomal content and basal oxidative metabolism than hyali-nocytes. Hemocyte parameters were not influenced by the size of green mussels. In addition, hemocyteswere subjected to acute temperature challenges (10, 20 and 30 �C) and their immune-related functionsand metabolism analyzed. Our results showed that 10 �C represent a stressful condition for the Floridiangreen mussels, as depicted by a low phagocytosis capacity and an increase of oxidative metabolism.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
The greenmussel Perna viridis is a bivalvemollusknative from theIndian coast and throughout the Indo-Pacific and Asia-Pacific. In theearly to mid-1990s, populations were discovered along the coasts ofVenezuela, Trinidad, and Jamaica [1,2]. In 1999, green musselsgenetically related to the ones previously described in the Caribbeanwere observed in Tampa Bay, Florida, USA [3]. This was the firstreported occurrence of this Indo-Pacific marine bivalve in NorthAmerica. Green mussel population has then expanded considerably,most likely through local reproduction and settlement, along thecoastal regions of southwestern Florida. Recruits of Perna viridis cannowalso be found on the Atlantic coast of Florida, Georgia and SouthCarolina [1,2,4]. Green mussels are known biofoulers of boats, navi-gational aids but also of submerged power plants and other indus-tries infrastructures, potentially clogging water intakes and outflowpipes [1]. Furthermore, this non-native species seems to have the
: þ1 239 590 7200.
All rights reserved.
potential to displace local native species such as the Eastern oyster,Crassostrea virginica [5]. The long-term ecological and economicimpact of the green mussel introduction to Florida might then beconsiderable but cannot be predicted at this time.
Due to their origin, the population dynamics [6e8] as well as thereproduction and growth [9e11] of green mussels were, until now,mostly studied in Asia, where P. viridis are widely used for thebiomonitoring of coastal environments. Indeed, many studiesmeasured the tissue levels of contaminants such as heavy metals,PAHs, PCBs or pesticides in P. viridis [12e15]. Similarly, the toxico-logical impacts of such contaminations on the green mussels wereinvestigated, once again mostly in Asia [16e21]. The populations ofgreen mussels from Florida were, until now, almost exclusivelystudied for their geographic distribution patterns, in order tounderstand their origin and to predict the population dynamics[1e4]. Very recently, Urian et al. [22] investigated on the effects ofseawater temperature on the survival of Floridian P. viridis, andconcluded that due to a weak low temperature resistance, greenmussels might have already reached the Northern edge of theirdistribution in USA. Indeed, temperature is a predominant factordetermining the geographic distribution of marine invertebrates

L. Donaghy, A.K. Volety / Fish & Shellfish Immunology 31 (2011) 808e814 809
[23]. For instance, cold temperatures have been shown to increasemortality of the tropical species Perna perna [24]. But even nonlethal temperatures can induce a stress that may reduce long-termsurvival, growth and/or reproductive success [22,23]. There iscurrently an obvious lack of data about the adaptation capacities ofP. viridis in Floridian environmental conditions, at the physiologicaland cellular levels.
Hemocytes are cells circulating in hemolymphatic systemand infiltrating in tissues of bivalve mollusks [25e28]. Bivalvehomeostasis relies, at least partially, on hemocytes through theirinvolvement in various physiological functions [29] includingnutrient digestion, transportation, and distribution [30,31], shelland tissue repair [32,33], detoxification processes [34e36]and cellular immune defense [37e41]. Although various hemocytesub-populations have been described in marine bivalves [28], twomain types are generally accepted: granulocytes, containing manyintra-cytoplasmic granules, and hyalinocytes, with no granules[25,27,28]. Environmental factors including temperature, salinity,nutrients and toxicants affect marine bivalves. Hemocytes weresuggested to be sensitive to variations of these factors, reflecting thephysiological status of bivalves and their adaptation to the envi-ronment [29,41]. The characterization of hemocytes under natural,ambient condition, as well as their response to environmentalvariations is then essential to further understand cell-mediatedresponses of bivalves to anthropogenic and pathological stresses,as well as their adaptation capacities compared with other species.
The present study aimed at better understanding the Floridiangreen mussels’ adaptation capacities to changing environmentalconditions, such as temperature, through (i) the characterization ofthe biology and types of circulating hemocytes and (ii) the study ofthe in vitro effects of temperature variations on the hemocytes’functional and metabolic activities.
2. Material and methods
2.1. Animals
Adult green mussels were collected from Estero Bay, Florida,USA (26�220N 81�510W; depth from 1.5 to 2 m), where ambientsalinity was 31.6 and seawater temperature 22 �C. For character-ization of hemocytes, mussels were field collected and processedimmediately. The size of mussels was determined as the length ofthe shell. The size of P. viridis individuals ranged from86 to 138mm,with a mean value of 109.75 � 7.5 mm. Mussels used in tempera-ture experiments (Section 2.4) were placed in tanks of fully aeratedartificial seawater (salinity 30) and acclimated for at least 1 weekprior to experiments. Mussels were fed daily with Shellfish Dietalgae mix (Reed Mariculture Inc.).
2.2. Hemolymph collection
Using a 3 mL syringe fitted with a 25-G needle, approximately1.5e2.0 mL of hemolymph was collected from each mussel. Theneedle was inserted between shell valves into the posterioradductor muscle. Collected hemolymph was microscopicallyexamined for purity, filtered through 41 mmnylon filters (accordingflow cytometer manufacturer recommendations) and kept on ice tominimize cell clumping. All subsequent analyses were performedon individual samples.
2.3. Flow cytometry analyses
Flow cytometry analyses were performed on a Cytomics FC500flow cytometer (Beckman Coulter), equipped with a 488 nm Argonlaser.
2.3.1. Hemocyte concentration, cytomorphology and mortalityDetermination of hemocyte concentration, cytomorphology and
mortality was performed using a double staining procedureincluding SYBR Green I (1/1000 of stock solution) (Invitrogen) andPropidium iodide (PI; 10 mg mL�1) (Sigma) [42]. Hemocytemorphology was based upon relative flowecytometric parameters,Forward Scatter (FSC) and Side Scatter (SSC). FSC and SSCcommonly measure particle size and internal complexity, respec-tively. Membranes of viable cells do not allow PI to penetrate;whereas, altered membranes are permeable to PI. Dead cells arecharacterized by loss of membrane integrity and are, therefore,double stained by SYBR Green I and PI. Concentration is reported asthe number of cells mL�1 of hemolymph; morphological parame-ters are expressed in arbitrary units (A.U.), and hemocyte mortalityis the percentage of double stained cells.
2.3.2. Lysosome quantificationThe presence and relative amount of lysosomes in hemocytes
was determined using LysoTracker Red (Molecular Probes, Invi-trogen), a membrane permeable, fluorescent probe (emissionmaximum at w 590 nm) that accumulates within lysosomalcompartments. Hemolymph was diluted in artificial filteredseawater (AFSW) containing LysoTracker Red (final concentration1 mM). Mixed solutions were incubated 60 min in the dark, at roomtemperature. Relative intracellular lysosomal quantity is expressedas the level of red fluorescence (FL3 detector of the flow cytometer)in arbitrary units (A.U.).
2.3.3. Phagocytic capacitiesEvaluation of phagocytic capacities was based on the ingestion
of fluorescent latex microbeads (2.0 mm, Polysciences Inc.) byhemocytes. Incubation of hemolymph with microbeads was per-formed at 20 �C for 120 min. Conditions of incubation for in vitroexperiment are described in Section 2.4. Phagocytic capacities werethen defined as (i) the percentage of cells that had ingested three ormore microbeads [42,43], and (ii) the average number ofmicrobeads per phagocytic hemocytes [44].
2.3.4. Intracellular oxidative metabolismDetermination of intracellular oxidative metabolism was per-
formed using 2070-dichlorofluorescein diacetate (final concentra-tion 10 mM) (DCFH-DA; Molecular Probes, Invitrogen), a membranepermeable, non-fluorescent probe. Inside hemocytes, the -DAradical is first hydrolyzed by esterase enzymes. Intracellularhydrogen peroxide (H2O2), as well as superoxide ion (O2�
� [45,46];),then oxidizes DCFH to the fluorescent DCF molecule. Oxidation ofDCFH can also be mediated by nitrite radicals (NO2 or N2O3) [47]and various oxidase and peroxidase enzymes [48]. DCF greenfluorescence is proportional to the intracellular oxidative metabo-lism of hemocytes. Relative intracellular oxidative metabolism istherefore expressed as the level of green fluorescence (FL1detector) in arbitrary units (A.U.). Incubation of hemolymph withDCFH-DA was performed at 20 �C for 30 min. Conditions of incu-bation for in vitro experiment are described in Section 2.4.
2.3.5. Mitochondrial membrane potentialEstimation of mitochondrial membrane potential (MMP) was
performed using JC10 (final concentration 5 mM) (Enzo LifeSciences), a membrane permeable fluorescent probe. JC-10 accu-mulates and aggregates in mitochondria, selectively generating anorange emission profile (l 590 nm). As membrane potentialdecreases, JC-10 monomers are generated, resulting in a shift togreen fluorescence emission (l 525 nm). Relative MMP intensity isexpressed as the ratio between the levels of orange fluorescence(FL2 detector) and green fluorescence (FL1 detector).
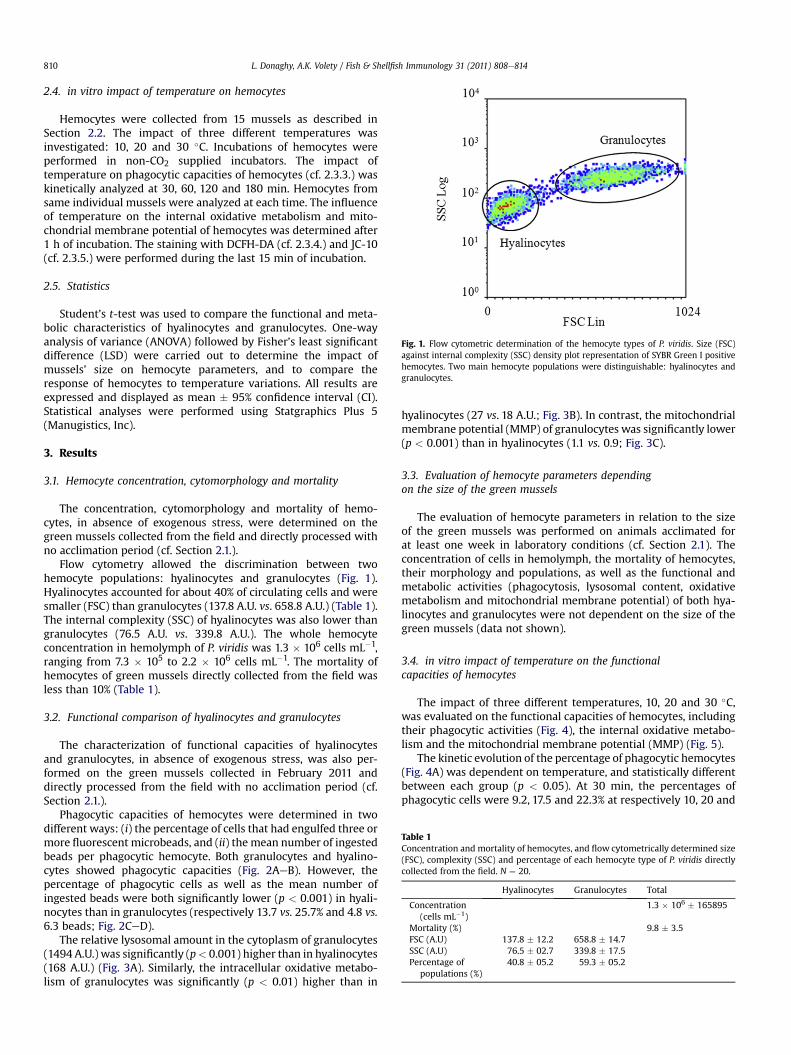
Fig. 1. Flow cytometric determination of the hemocyte types of P. viridis. Size (FSC)against internal complexity (SSC) density plot representation of SYBR Green I positivehemocytes. Two main hemocyte populations were distinguishable: hyalinocytes andgranulocytes.
Table 1Concentration and mortality of hemocytes, and flow cytometrically determined size(FSC), complexity (SSC) and percentage of each hemocyte type of P. viridis directlycollected from the field. N ¼ 20.
Hyalinocytes Granulocytes Total
Concentration(cells mL�1)
1.3 � 106 � 165895
Mortality (%) 9.8 � 3.5FSC (A.U) 137.8 � 12.2 658.8 � 14.7SSC (A.U) 76.5 � 02.7 339.8 � 17.5Percentage of
populations (%)40.8 � 05.2 59.3 � 05.2
L. Donaghy, A.K. Volety / Fish & Shellfish Immunology 31 (2011) 808e814810
2.4. in vitro impact of temperature on hemocytes
Hemocytes were collected from 15 mussels as described inSection 2.2. The impact of three different temperatures wasinvestigated: 10, 20 and 30 �C. Incubations of hemocytes wereperformed in non-CO2 supplied incubators. The impact oftemperature on phagocytic capacities of hemocytes (cf. 2.3.3.) waskinetically analyzed at 30, 60, 120 and 180 min. Hemocytes fromsame individual mussels were analyzed at each time. The influenceof temperature on the internal oxidative metabolism and mito-chondrial membrane potential of hemocytes was determined after1 h of incubation. The staining with DCFH-DA (cf. 2.3.4.) and JC-10(cf. 2.3.5.) were performed during the last 15 min of incubation.
2.5. Statistics
Student’s t-test was used to compare the functional and meta-bolic characteristics of hyalinocytes and granulocytes. One-wayanalysis of variance (ANOVA) followed by Fisher’s least significantdifference (LSD) were carried out to determine the impact ofmussels’ size on hemocyte parameters, and to compare theresponse of hemocytes to temperature variations. All results areexpressed and displayed as mean � 95% confidence interval (CI).Statistical analyses were performed using Statgraphics Plus 5(Manugistics, Inc).
3. Results
3.1. Hemocyte concentration, cytomorphology and mortality
The concentration, cytomorphology and mortality of hemo-cytes, in absence of exogenous stress, were determined on thegreen mussels collected from the field and directly processed withno acclimation period (cf. Section 2.1.).
Flow cytometry allowed the discrimination between twohemocyte populations: hyalinocytes and granulocytes (Fig. 1).Hyalinocytes accounted for about 40% of circulating cells and weresmaller (FSC) than granulocytes (137.8 A.U. vs. 658.8 A.U.) (Table 1).The internal complexity (SSC) of hyalinocytes was also lower thangranulocytes (76.5 A.U. vs. 339.8 A.U.). The whole hemocyteconcentration in hemolymph of P. viridis was 1.3 � 106 cells mL�1,ranging from 7.3 � 105 to 2.2 � 106 cells mL�1. The mortality ofhemocytes of green mussels directly collected from the field wasless than 10% (Table 1).
3.2. Functional comparison of hyalinocytes and granulocytes
The characterization of functional capacities of hyalinocytesand granulocytes, in absence of exogenous stress, was also per-formed on the green mussels collected in February 2011 anddirectly processed from the field with no acclimation period (cf.Section 2.1.).
Phagocytic capacities of hemocytes were determined in twodifferent ways: (i) the percentage of cells that had engulfed three ormore fluorescent microbeads, and (ii) the mean number of ingestedbeads per phagocytic hemocyte. Both granulocytes and hyalino-cytes showed phagocytic capacities (Fig. 2AeB). However, thepercentage of phagocytic cells as well as the mean number ofingested beads were both significantly lower (p < 0.001) in hyali-nocytes than in granulocytes (respectively 13.7 vs. 25.7% and 4.8 vs.6.3 beads; Fig. 2CeD).
The relative lysosomal amount in the cytoplasm of granulocytes(1494 A.U.) was significantly (p< 0.001) higher than in hyalinocytes(168 A.U.) (Fig. 3A). Similarly, the intracellular oxidative metabo-lism of granulocytes was significantly (p < 0.01) higher than in
hyalinocytes (27 vs. 18 A.U.; Fig. 3B). In contrast, the mitochondrialmembrane potential (MMP) of granulocytes was significantly lower(p < 0.001) than in hyalinocytes (1.1 vs. 0.9; Fig. 3C).
3.3. Evaluation of hemocyte parameters dependingon the size of the green mussels
The evaluation of hemocyte parameters in relation to the sizeof the green mussels was performed on animals acclimated forat least one week in laboratory conditions (cf. Section 2.1). Theconcentration of cells in hemolymph, the mortality of hemocytes,their morphology and populations, as well as the functional andmetabolic activities (phagocytosis, lysosomal content, oxidativemetabolism and mitochondrial membrane potential) of both hya-linocytes and granulocytes were not dependent on the size of thegreen mussels (data not shown).
3.4. in vitro impact of temperature on the functionalcapacities of hemocytes
The impact of three different temperatures, 10, 20 and 30 �C,was evaluated on the functional capacities of hemocytes, includingtheir phagocytic activities (Fig. 4), the internal oxidative metabo-lism and the mitochondrial membrane potential (MMP) (Fig. 5).
The kinetic evolution of the percentage of phagocytic hemocytes(Fig. 4A) was dependent on temperature, and statistically differentbetween each group (p < 0.05). At 30 min, the percentages ofphagocytic cells were 9.2, 17.5 and 22.3% at respectively 10, 20 and

Fig. 2. Flow cytometric comparison of phagocytic capacities of P. viridis hyalinocytes and granulocytes. (AeB) Size (FSC) against internal complexity (SSC) density plot of hemocytesincubated with fluorescent microbeads (B). Size against fluorescence intensity (FL1) density plot of the same hemocyte populations associated or not with fluorescent microbeads(A). Hemocyte types were designated as hyalinocytes (Hy), granulocytes (Gr) and aggregated cells (Ag). The histogram (upper right) represents the intensity of fluorescencedepending on the quantity of ingested beads. Representative results of 20 individuals. (CeD) Percentage of phagocytic hyalinocytes and granulocytes (C) and average number ofingested beads in phagocytic hemocytes (D). An asterisk indicates a significant (t-test, p < 0.001) difference between groups.
L. Donaghy, A.K. Volety / Fish & Shellfish Immunology 31 (2011) 808e814 811
30 �C. At all temperatures, the percentage of phagocytic cellsincreased more quickly during the first 2 h, reaching values of 18.1,41.7 and 48.2%. After 2 h, the percentage of phagocytic cellsincreased more slowly, reaching 21.6, 46 and 53.8% after 180 minincubation.
The quantity of ingested beads did not evolve in the sameway asthe percentage of phagocytic hemocytes (Fig. 4B). At 10 �C, theaverage number of beads inside hemocytes remained constant,around 4 beads/cell, from 30 to 180 min. At 20 �C, ingested beadsincreased to 5.2 and 6.1 beads/cell at 120 and 180 min (p < 0.05),respectively. Themean quantity of beads in hemocytes incubated at30 �C increased as early as 60 min (p< 0.05), to reach 8.4 beads/cellat 180 min.
The impact of temperature on the intracellular oxidativemetabolism differed between hyalinocytes and granulocytes(Fig. 5A). In hyalinocytes, the oxidativemetabolism remained equal,whatever the incubation temperature. Contrastingly, the oxidativemetabolism in granulocytes was highest at 10 �C (65 A.U.) and
Fig. 3. Comparison of (A) lysosomal content expressed as fluorescence intensity (FL3), (B)mitochondrial membrane potential in P. viridis hyalinocytes and granulocytes. Asterisks ind
decreased to 56 A.U. and 38 A.U. at 20 �C and 30 �C, respectively.The values obtained at 20 and 30 �C were significantly differentfrom the other groups (p < 0.001).
The MMPwas dependant on the incubation temperature and itsvariations were similar between hyalinocytes and granulocytes(Fig. 5B). Although it was significantly different between hyalino-cytes and granulocytes (p < 0.001), the MMP was the highest inhemocytes incubated at 10 �C. The MMP then decreased when cellswere maintained at higher temperatures.
4. Discussion
In the present study, the use of flow cytometry allowed thedistinction between two main hemocyte populations: hyalinocytesand granulocytes. In spite of controversies about the classificationof hemocytes, flow cytometry and microscopy studies alwaysreported these two major cell types in mussels such as Mytilusedulis [49,50] and Mytilus galloprovincialis [51e53] as well as in
intracellular oxidative metabolism expressed as fluorescence intensity (FL1) and (C)icate a significant (t-test) difference between groups; *: p < 0.01; **: p < 0.001.
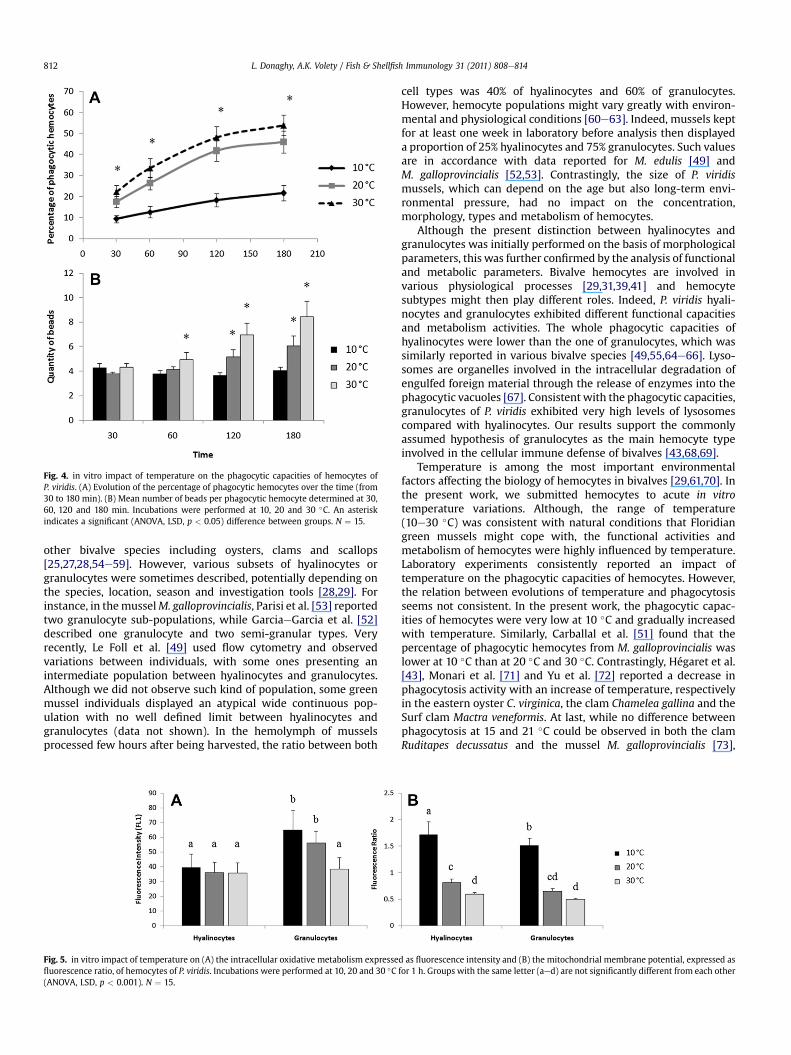
Fig. 4. in vitro impact of temperature on the phagocytic capacities of hemocytes ofP. viridis. (A) Evolution of the percentage of phagocytic hemocytes over the time (from30 to 180 min). (B) Mean number of beads per phagocytic hemocyte determined at 30,60, 120 and 180 min. Incubations were performed at 10, 20 and 30 �C. An asteriskindicates a significant (ANOVA, LSD, p < 0.05) difference between groups. N ¼ 15.
L. Donaghy, A.K. Volety / Fish & Shellfish Immunology 31 (2011) 808e814812
other bivalve species including oysters, clams and scallops[25,27,28,54e59]. However, various subsets of hyalinocytes orgranulocytes were sometimes described, potentially depending onthe species, location, season and investigation tools [28,29]. Forinstance, in themusselM. galloprovincialis, Parisi et al. [53] reportedtwo granulocyte sub-populations, while GarciaeGarcia et al. [52]described one granulocyte and two semi-granular types. Veryrecently, Le Foll et al. [49] used flow cytometry and observedvariations between individuals, with some ones presenting anintermediate population between hyalinocytes and granulocytes.Although we did not observe such kind of population, some greenmussel individuals displayed an atypical wide continuous pop-ulation with no well defined limit between hyalinocytes andgranulocytes (data not shown). In the hemolymph of musselsprocessed few hours after being harvested, the ratio between both
Fig. 5. in vitro impact of temperature on (A) the intracellular oxidative metabolism expressefluorescence ratio, of hemocytes of P. viridis. Incubations were performed at 10, 20 and 30 �C(ANOVA, LSD, p < 0.001). N ¼ 15.
cell types was 40% of hyalinocytes and 60% of granulocytes.However, hemocyte populations might vary greatly with environ-mental and physiological conditions [60e63]. Indeed, mussels keptfor at least one week in laboratory before analysis then displayeda proportion of 25% hyalinocytes and 75% granulocytes. Such valuesare in accordance with data reported for M. edulis [49] andM. galloprovincialis [52,53]. Contrastingly, the size of P. viridismussels, which can depend on the age but also long-term envi-ronmental pressure, had no impact on the concentration,morphology, types and metabolism of hemocytes.
Although the present distinction between hyalinocytes andgranulocytes was initially performed on the basis of morphologicalparameters, this was further confirmed by the analysis of functionaland metabolic parameters. Bivalve hemocytes are involved invarious physiological processes [29,31,39,41] and hemocytesubtypes might then play different roles. Indeed, P. viridis hyali-nocytes and granulocytes exhibited different functional capacitiesand metabolism activities. The whole phagocytic capacities ofhyalinocytes were lower than the one of granulocytes, which wassimilarly reported in various bivalve species [49,55,64e66]. Lyso-somes are organelles involved in the intracellular degradation ofengulfed foreign material through the release of enzymes into thephagocytic vacuoles [67]. Consistent with the phagocytic capacities,granulocytes of P. viridis exhibited very high levels of lysosomescompared with hyalinocytes. Our results support the commonlyassumed hypothesis of granulocytes as the main hemocyte typeinvolved in the cellular immune defense of bivalves [43,68,69].
Temperature is among the most important environmentalfactors affecting the biology of hemocytes in bivalves [29,61,70]. Inthe present work, we submitted hemocytes to acute in vitrotemperature variations. Although, the range of temperature(10e30 �C) was consistent with natural conditions that Floridiangreen mussels might cope with, the functional activities andmetabolism of hemocytes were highly influenced by temperature.Laboratory experiments consistently reported an impact oftemperature on the phagocytic capacities of hemocytes. However,the relation between evolutions of temperature and phagocytosisseems not consistent. In the present work, the phagocytic capac-ities of hemocytes were very low at 10 �C and gradually increasedwith temperature. Similarly, Carballal et al. [51] found that thepercentage of phagocytic hemocytes from M. galloprovincialis waslower at 10 �C than at 20 �C and 30 �C. Contrastingly, Hégaret et al.[43], Monari et al. [71] and Yu et al. [72] reported a decrease inphagocytosis activity with an increase of temperature, respectivelyin the eastern oyster C. virginica, the clam Chamelea gallina and theSurf clam Mactra veneformis. At last, while no difference betweenphagocytosis at 15 and 21 �C could be observed in both the clamRuditapes decussatus and the mussel M. galloprovincialis [73],
d as fluorescence intensity and (B) the mitochondrial membrane potential, expressed asfor 1 h. Groups with the same letter (aed) are not significantly different from each other

L. Donaghy, A.K. Volety / Fish & Shellfish Immunology 31 (2011) 808e814 813
Perrigault et al. [70] reported a higher phagocytic activity at 21 �Cthan at 13 and 27 �C in the hard clam Mercenaria mercenaria. Then,as suggested by Monari et al. [71], temperatures above or belowa certain threshold may result in stressful conditions for hemo-cytes, so that they are less responsive. Such a threshold mightdepend on both the natural field environment and the geneticbackground of the species itself.
During cellular respiration, dioxygen (O2) is reduced into water(H2O), and the mitochondrial respiratory chain transfers protonsacross the inner mitochondrial membrane, producing an electro-chemical gradient. Such proton motive force is a physico-chemicalparameter consisting of two components: a voltage gradient, themitochondrial membrane potential (MMP), and a pH gradient [74].It has been reported that respiration rates of ectothermal animals[75,76] increase at higher temperature. In the present study, wecontrastingly observed a decrease of MMP from 10 to 30 �C. Usingthe same dye (JC-10) with flow cytometry analysis, similar patternof response was observed in hemocytes of the Pacific oysterCrassostrea gigas: MMP remained constant at 10 and 15 �C, thendecreased at 20 and 25 �C (Soudant, personal communication).First, the MMP is only one of the components involved in mito-chondrial respiration, and the present results do not allowcomplete understanding of mitochondrial changes occurringduring the increase of temperature. Second, the MMP is used toproduce ATP, the energy transfer nucleotide. High temperaturesmight stimulate the green mussel’s metabolism, increasing theneed for ATP which might therefore result in a decrease of theMMP. During cellular respiration, an estimated 1 or 2% of oxygenconsumed is not completely reduced to water but instead partiallyreduced to superoxide anion (O2�
�) which can be converted tohydrogen peroxide (H2O2) and the highly reactive hydroxyl radical(OH�) [77,78]. These reactive oxygen species (ROS) participate to thewhole oxidative metabolism, and most studies investigating theeffect of temperature on hemocytes reported an increase ofROS production (unstimulated and stimulated) with increasingtemperature [43,79,80]. Increase of ROS production is thought to berelated to increase in respiration at high temperature, as statedabove. In contrast, results from our study showed that unstimu-lated oxidative metabolism was highest at 10 �C and decreased at20 and 30 �C. Perrigault et al. [70] reported similar observationwith hemocytes of Mercenaria mercenaria maintained for 2 and 4months at 13, 21 and 27 �C. In these cases, low temperatureexposures might represent stressful conditions for the studiedspecies, while the higher temperatures remained under the stressthreshold, as suggested for phagocytic activity. However, due to thecurrent lack of fundamental knowledge about the tight relationsbetween oxidative metabolism, mitochondrial activity and envi-ronmental conditions in bivalves, present results do not allow us todefinitely conclude this relationship.
The present study characterized, for the first time, the two mainpopulations of circulating hemocytes from the green mussel,P. viridis. Such knowledge was crucial to determining the impact oftemperature variation on the metabolism and immune-relatedfunctions of hemocytes. In accordance with the very recent workof Urian et al. [22], our results showed that relatively low temper-ature (10 �C) represents a stressful condition for the Floridian greenmussels, as depicted by the low phagocytic capacity and theincrease of oxidative metabolism. Then, as suggested by Urian et al.[22], in the current environmental conditions, green mussels mighthave already reached the Northern edge of their distribution in theUSA. However such statement should further be argued by thestudy of the impact of temperature and other environmentalparameters on both the hemocyte activities and physiology ofgreen mussels, including filtration, reproduction or growth ofindividuals.
Acknowledgments
The authors thank the staff of the Vester Marine and Environ-mental Science Research Field Station. Sincere thanks are especiallydue to Lesli Haynes for help in animal maintenance, and KatieMcFarland and BobWasno for field collection of greenmussels. Thecontents of this scientific publication were developed undera Congressionally-directed grant (P116Z090117) from the U.S.Department of Education. However, the contents do not necessarilyrepresent the policy of the U.S. Department of Education, and youshould not assume endorsement by the Federal Government.
References
[1] Benson AJ, Marelli DC, Frischer ME, Danforth JM, Williams JD. Establishment ofthe green mussel, Perna viridis (Linnaeus 1758) (Mollusca:Mytilidae) on thewest coast of Florida. J Shellfish Res 2001;20:21e9.
[2] BakerP, Fajans JS, ArnoldWS, IngraoDA,Marelli DC, BakerSM.Rangeanddispersalof a tropical marine invader, the Asian green mussel, Perna viridis, in subtropicalwaters of the southeastern United States. J Shellfish Res 2007;26:345e55.
[3] Ingrao DA, Mikkelsen PM, Hicks DW. Another introduced marine mollusk inthe Gulf of Mexico: the Indo-Pacific green mussel, Perna viridis, in Tampa Bay.Florida J Shellfish Res 2001;20:13e9.
[4] Power AJ, Walker RL, Payne K, Hurley D. First occurrence of the nonindigenousgreen mussel, Perna viridis (Linnaeus, 1758) in coastal Georgia, United States.J Shellfish Res 2004;23:741e4.
[5] Kravitz AR, Campanella R. The invasive green mussel and its potential impacton the State’s oyster industry. Louisiana Oyster Industry Convention. NewOrleans; 2004.
[6] Khan MAA, Assim ZB, Ismail N. Population dynamics of the green-lippedmussel, Perna viridis from the Offshore waters of Naf River coast,Bangladesh. Chiang Mai J Sci 2010;37:344e54.
[7] Prakoon W, Tunkijjanukij S, Nguyen TTT, Na-Nakorn U. Spatial and temporalgenetic variation of green mussel, Perna viridis in the Gulf of Thailand andimplication for aquaculture. Mar Biotech 2010;12:506e15.
[8] Stafford H, Willan RC, Neil KM. The invasive Asian green mussel, Perna viridis(Linnaeus, 1758) (Bivalvia: Mytilidae), breeds in Trinity Inlet, tropical northernAustralia. Molluscan Res 2007;27:105e9.
[9] Acosta V, Glem ME, Natera Y, Urbano T, Himmelman JH, Rey-Mendez M, et al.Differential Growth of the Mussels Perna perna and Perna viridis (Bivalvia:Mytilidae) in Suspended Culture in the Golfo de Cariaco, Venezuela. J WorldAquacult Soc 2009;40:226e35.
[10] Bigatti G, Miloslavich P, Penchaszadeh PE. Sexual differentiation and size atfirst maturity of the invasive mussel Perna viridis (Linnaeus, 1758) (Mollusca:Mytilidae) at La Restinga Lagoon (Margarita Island, Venezuela). Am MalacolBull 2005;20:65e9.
[11] Rajagopal S, Venugopalan VP, Nair KVK, van der Velde G, Jenner HA, denHartog C. Reproduction, growth rate and culture potential of the greenmussel, Perna viridis (L.) in Edaiyur backwaters, east coast of India. Aquacul-ture 1998;162:187e202.
[12] Choi HG, Moon HB, Choi M, Yu J, Kim SS. Mussel watch program for organiccontaminants along the Korean coast, 2001e2007. Environ Monit Assess2010;169:473e85.
[13] Fang JKH, Wu RSS, Zheng GJ, Lam PKS, Shin PKS. Seasonality of bio-accumulation of trace organics and lysosomal integrity in green-lipped musselPerna viridis. Sci Total Environ 2010;408:1458e65.
[14] Rojas N, Lemus M, Rojas L, Martinez G, Ramos Y, Chung KS. Mercury levels inPerna viridis from the north coast of Sucre State, Venezuela. Cienc Mar 2009;35:91e9.
[15] Shahbazi A, Zakaria MP, Yap CK, Tan SG, Surif S, Mohamed CAR, et al. Use ofdifferent tissues of perna viridis as Biomonitors of Polycyclic AromaticHydrocarbons (PAHs) in the coastal waters of Peninsular Malaysia. EnvironForensics 2010;11:248e63.
[16] Aanand S, Purushothaman CS, Pal AK, Rajendran KV. Toxicological studies onthe effect of copper, lead and zinc on selected enzymes in the adductor muscleand intestinal diverticula of the green mussel Perna viridis. Indian J Mar Sci2010;39:299e302.
[17] Long AM, Li CD, Chen SY, Yan W, Dang AC, Cheng YY, et al. Short-term metalaccumulation and MTLP induction in the digestive glands of Perna virdisexposed to Zn and Cd. J Environ Sci-China 2010;22:975e81.
[18] Nusetti O, Tovar M, Zapata-Vivenes E. Pyruvate kinase, PhosphoenolpyruvateCarboxykinase, Cytochrome C oxidase and Catalase activities in cadmiumexposed perna viridis subjected to Anoxic and Aerobic conditions. J ShellfishRes 2010;29:203e8.
[19] Verlecar XN, Jena KB, Chainy GBN. Modulation of antioxidant defences indigestive gland of Perna viridis (L.), on mercury exposures. Chemosphere2008;71:1977e85.
[20] Wu HF, Wang WX. NMR-based metabolomic studies on the toxicologicaleffects of cadmium and copper on green mussels Perna viridis. Aquat Toxicol2010;100:339e45.

L. Donaghy, A.K. Volety / Fish & Shellfish Immunology 31 (2011) 808e814814
[21] Wu HF, Wang WX. Tissue-Specific toxicological effects of cadmium in greenmussels (Perna viridis): Nuclear Magnetic Resonance-based Metabolomicsstudy. Environ Toxicol Chem 2011;30:806e12.
[22] Urian AG, Hatle JD, Gilg MR. Thermal Constraints for range Expansion of theinvasive green mussel, Perna viridis, in the southeastern United States. J ExpZool A Ecol Genet Physiol 2011;315A:12e21.
[23] Delgado M, Camacho AP. Influence of temperature on gonadal development ofRuditapes philippinarum (Adams and Reeve, 1850) with special reference toingested food and energy balance. Aquaculture 2007;264:398e407.
[24] Hicks DW, McMahon RF. Temperature acclimation of upper and lowerthermal limits and freeze resistance in the nonindigenous brown mussel,Perna perna (L.), from the Gulf of Mexico. Marine Biol 2002;140:1167e79.
[25] Auffret M. Bivalve hemocyte morphology. Am Fish Soc Spec Pub 1988;18:169e77.
[26] Cheng TC. Functional morphology and biochemistry of molluscan phagocytes.Ann New York Acad Sci 1975;266:343e79.
[27] Cheng TC. Bivalves. In: Ratcliffe NARA, editor. Invertebrate blood cells. Lon-don, UK: Academic Press; 1981. p. 233e300.
[28] Hine PM. The inter-relationships of bivalve haemocytes. Fish ShellfishImmunol 1999;9:367e85.
[29] Donaghy L, Lambert C, Choi KS, Soudant P. Hemocytes of the carpet shell clam(Ruditapes decussatus) and the Manila clam (Ruditapes philippinarum): currentknowledge and future prospects. Aquaculture 2009;297:10e24.
[30] Beninger PG, Le Pennec G, Le Pennec M. Demonstration of nutrient pathwayfrom the digestive system to oocytes in the gonad intestinal loop of thescallop Pecten maximus L. Biol Bull 2003;205:83e92.
[31] Cheng TC. Hemocytes: forms and functions. In: Kennedy VS, Eble AF,Newell RIE, editors. The eastern oyster Crassostrea virginica. College Park, MD,USA: Maryland Sea Grant Book; 1996. p. 299e333.
[32] Mount AS, Wheeler AP, Paradkar RP, Snider D. Hemocyte-mediated shellmineralization in the eastern oyster. Science 2004;304:297e300.
[33] Sparks AK, Morado JF. Inflammation and wound repair in bivalve molluscs. In:WS F, editor. Disease processes in marine bivalve molluscs. Bethesda, MD,USA: American Fisheries Society; 1988. p. 139e52.
[34] Fisher WS. Antimicrobial activity of copper and zinc accumulated in easternoyster amebocytes. J Shellfish Res 2004;23:321e51.
[35] Fisher WS. Relationship of amebocytes and terrestrial elements to adult shelldeposition in eastern oysters. J Shellfish Res 2004;23:353e67.
[36] Matozzo V, Ballarin L, Pampanin DM, Marin MG. Effects of copper andcadmium exposure on functional responses of hemocytes in the clam, Tapesphilippinarum. Arch Environ Contam Toxicol 2001;41:163e70.
[37] Canesi L, Gallo G, Gavioli M, Pruzzo C. Bacteria-hemocyte interactions andphagocytosis in marine bivalves. Microsc Res Tech 2002;57:469e76.
[38] Chagot D, Comps M, Boulo V, Ruano F, Grizel H. Histological study of a cellularreaction in Ruditapes decussatus infected by a protozoan. Aquaculture 1987;67:260e1.
[39] Cheng TC. Cellular defense mechanisms in oysters. In: Fingerman NNR, editor.Recent advances in marine biotechnology. Immunobiology and pathology.Enfield, NH, USA: Science Publishers; 2000. p. 43e83.
[40] Chu FLE. Humoral defense factors in marine bivalves. Am Fish Soc Spec Pub;1988:18.
[41] Chu FLE. Defense mechanisms of marine bivalves. In: Fingerman NNR, editor.Recent advances in marine biotechnology. Immunobiology and pathology.Enfield, NH, USA: Science Publishers; 2000. p. 1e42.
[42] Delaporte M, Soudant P, Moal J, Lambert C, Quere C, Miner P, et al. Effect ofa mono-specific algal diet on immune functions in two bivalve species -Crassostrea gigas and Ruditapes philippinarum. J Exp Biol 2003;206:3053e64.
[43] Hégaret H, Wikfors GH, Soudant P. Flow-cytometric analysis of haemocytesfrom eastern oysters, Crassostrea virginica, subjected to a sudden temperatureelevation II. Haemocyte functions: aggregation, viability, phagocytosis, andrespiratory burst. J Exp Mar Biol Ecol; 2003:249e65.
[44] Donaghy L, Hong H-K, Lambert C, Park H-S, Shim WJ, Choi K-S. First charac-terisation of the populations and immune-related activities of hemocytesfrom two edible gastropod species, the disk abalone, Haliotis discus discus andthe spiny top shell, Turbo cornutus. Fish Shellfish Immunol 2010;28:87e97.
[45] Curtin JF, Donovan M, Cotter TG. Regulation and measurement of oxidativestress in apoptosis. J Immunol Methods 2002;265:49e72.
[46] Rao KMK, Padmanabhan J, Kilby DL, Cohen HJ, Currie MS, Weinberg JB. Flowcytometric analysis of Nitric-Oxide production in Human Neutrophils usingdichlorofluorescein diacetate in the presence of A Calmodulin Inhibitor. J LeukBiol 1992;51:496e500.
[47] Wardman P. Fluorescent and luminescent probes for measurement ofoxidative and nitrosative species in cells and tissues: progress, pitfalls, andprospects. Free Rad Biol Med 2007;43:995e1022.
[48] Gomes A, Fernandes E, Lima J. Fluorescence probes used for detection ofreactive oxygen species. J Biochem Biophys Methods 2005;65:45e80.
[49] Le Foll F, Rioult D, Boussa S, Pasquier J, Dagher Z, Leboulenger F. Character-isation of Mytilus edulis hemocyte subpopulations by single cell time-lapsemotility imaging. Fish Shellfish Immunol 2010;28:372e86.
[50] Pipe RK. Hydrolytic enzymes associated with the Antigranulocytes hemocytesof the marine mussel Mytilus edulis. Histochem J 1990;22:595e603.
[51] Carballal MJ, Lopez MC, Azevedo C, Villalba A. Hemolymph cell types of themussel Mytilus galloprovincialis. Dis Aquat Org 1997;29:127e35.
[52] Garcia-Garcia E, Prado-Alvarez M, Novoa B, Figueras A, Rosales C. Immuneresponses of mussel hemocyte subpopulations are differentially regulated by
enzymes of the PI3-K, PKC, and ERK kinase families. Dev Comp Immunol2008;32:637e53.
[53] Parisi MG, Li H, Jouvet LBP, Dyrynda EA, Parrinello N, Cammarata M, et al.Differential involvement of mussel hemocyte sub-populations in the clear-ance of bacteria. Fish Shellfish Immunol 2008;25:834e40.
[54] Cima F, Matozzo V, Marin MG, Ballarin L. Haemocytes of the clam Tapesphilippinarum (Adams & Reeve, 1850): morphofunctional characterisation.Fish Shellfish Immunol 2000;10:677e93.
[55] Donaghy L, Kim B-K, Hong H-K, Park H-S, Choi K- S. Flow cytometry studies onthe populations and immune parameters of the hemocytes of the Suminoeoyster, Crassostrea ariakensis. Fish Shellfish Immunol 2009;27:296e301.
[56] Hégaret H, Wikfors GH, Soudant P. Flow-cytometric analysis of haemocytesfrom eastern oysters, Crassostrea virginica, subjected to a sudden temperatureelevation I. Haemocyte Types morphology. J Exp Mar Biol Ecol; 2003:237e48.
[57] Lopez C, Carballal MJ, Azevedo C, Villalba A. Morphological characterization ofthe hemocytes of the clam, Ruditapes decussatus (Mollusca: Bivalvia). J InvertPathol 1997;69:51e7.
[58] Xing J, Zhan W. Characterisation of monoclonal antibodies to haemocyte typesof scallop (Chlamys farreri). Fish Shellfish Immunol; 2005:17e25.
[59] Zhang W, Wu X, Wang M. Morphological, structural, and functional charac-terization of the haemocytes of the scallop. Argopecten Irradians Aquacult;2006:19e32.
[60] da Silva PM, Comesana P, Fuentes J, Villalba A. Variability of haemocyte andhaemolymphparameters in Europeanflat oysterOstrea edulis families obtainedfrom brood stocks of different geographical origins and relation with infectionby the protozoan Bonamia ostreae. Fish Shellfish Immunol 2008;24:551e63.
[61] Flye-Sainte-Marie J, Soudant P, Lambert C, Le Goic N, Goncalvez M,Travers MA, et al. Variability of the hemocyte parameters of Ruditapes phil-ippinarum in the field during an annual cycle. J Exp Mar Biol Ecol 2009;377:1e11.
[62] Lambert C, Soudant P, Degremont L, Delaporte M, Moal J, Boudry P, et al.Hemocyte characteristics in families of oysters, Crassostrea gigas, selected fordifferential survival during summer and reared in three sites. Aquaculture2007;270:276e88.
[63] Soudant P, Paillard C, Choquet G, Lambert C, Reid HI, Marhic A, et al. Impact ofseason and rearing site on the physiological and immunological parameters ofthe Manila clam Venerupis (¼Tapes, ¼Ruditapes) philippinarum. Aquaculture2004;229:401e18.
[64] Goedken M, De Guise S. Flow cytometry as a tool to quantify oyster defencemechanisms. Fish Shellfish Immunol 2004;16:539e52.
[65] Lopez C, Carballal MJ, Azevedo C, Villalba A. Differential phagocytic ability ofthe circulating haemocyte types of the carpet shell clam Ruditapes decussatus(Mollusca: Bivalvia). Dis Aquat Org 1997;30:209e15.
[66] Tripp MR. Phagocytosis by hemocytes of the hard clam, Mercenaria merce-naria. J Invert Pathol 1992;59:222e7.
[67] Luzio JP, Pryor PR, Bright NA. Lysosomes: fusion and function. Nat Rev MolCell Biol 2007;8:622e32.
[68] Terahara K, Takahashi KG, Nakamura A, Osada M, Yoda M, Hiroi T, et al.Differences in integrin-dependent phagocytosis among three hemocytesubpopulations of the Pacific oyster “Crassostrea gigas”. Dev Comp Immunol2006;30:667e83.
[69] Carballal MJ, Lopez C, Azevedo C, Villalba A. in vitro study of phagocytic abilityof Mytilus galloprovincialis Lmk haemocytes. Fish Shellfish Immunol 1997;7:403e16.
[70] Perrigault M, Dahl SF, Espinosa EP, Gambino L, Allam B. Effects of temperatureon hard clam (Mercenaria mercenaria) immunity and QPX (Quahog ParasiteUnknown) disease development: II. Defense parameters. J Invert Pathol 2011;106:322e32.
[71] Monari M, Matozzo V, Foschi J, Cattani O, Serrazanetti GP, Marin MG. Effects ofhigh temperatures on functional responses of haemocytes in the clam Cha-melea gallina. Fish Shellfish Immunol 2007;22:98e114.
[72] Yu JH, Song JH, Choi MC, Park SW. Effects of water temperature change onimmune function in surf clams, Mactra veneriformis (Bivalvia: Mactridae).J Invert Pathol 2009;102:30e5.
[73] Ordas MC, Novoa B, Figueras A. Phagocytosis inhibition of clam and musselhaemocytes by Perkinsus atlanticus secretion products. Fish Shellfish Immunol1999;9:491e503.
[74] Solaini G, Sgarbi G, Lenaz G, Baracca A. Evaluating mitochondrial membranepotential in cells. Biosci Rep 2007;27:11e21.
[75] Abele D, Heise K, Portner HO, Puntarulo S. Temperature-dependence ofmitochondrial function and production of reactive oxygen species in theintertidal mud clam Mya arenaria. J Exp Biol 2002;205:1831e41.
[76] Sommer AM, Portner HO. Metabolic cold adaptation in the lugworm Arenicolamarina: comparison of a North Sea and a White Sea population. Mar EcolProgr Ser 2002;240:171e82.
[77] Murphy MP. How mitochondria produce reactive oxygen species. Biochem J2009;417:1e13.
[78] Poyton RO, Ball KA, Castello PR. Mitochondrial generation of free radicals andhypoxic signaling. Trends Endocrinol Metab 2009;20:332e40.
[79] Chen MY, Yang HS, Delaporte M, Zhao SJ. Immune condition of Chlamys farreriin response to acute temperature challenge. Aquaculture 2007;271:479e87.
[80] Cheng W, Hsiao IS, Hsu CH, Chen JC. Change in water temperature on theimmune response of Taiwan abalone Haliotis diversicolor supertexta and itssusceptibility to Vibrio parahaemolyticus. Fish Shellfish Immunol 2004;17:235e43.
Related Documents











