Journal of General Virology (1996), 77, 1211 1221. Printed in Great Britain 1211 Full protection against African horsesickness (AHS) in horses induced by baculovirus-derived AHS virus serotype 4 VP2, VP5 and VP7 Jorge L. Martinez-Torrecuadrada, 1 Maria Diaz-Laviada, 2 Polly Roy, 3.4 Carmen S~inchez, 2 Carmen Vela, 1 Jos6 Manuel S~inchez-Vizcaino 2 and J. Ignacio CasaP* 11NGENASA, Hnos Garc[a Noblejas 41, 2, 28037 Madrid, 2 Centro de hTvestigaci6n en Sanidad Animal-INIA, 28130 Valdeohnos, Madrid, Spain, 3 Laboratory of Molecular Biophysics, Institute of Virology and Environmental Microbiology, University of Oxford, MansfieM Road, Oxford OX1 3SR, UK and 4 University of Alabama, Birm#Tgham, AL 35294, USA African horsesickness virus serotype 4 (AHSV-4) outer capsid protein VP2, or VP2 and VP5 plus inner capsid protein VP7, derived from single or dual recombinant baculovirus expression vectors were used in different combinations to immunize horses. When the proteins were purified by affinity chromatography, the com- bination of all three proteins induced low levels of neutralizing antibodies and conferred protection against virulent virus challenge. However, purified VP2 or VP2 and VP5 in the absence of VP7 failed to induce neutralizing antibodies and protection. Immunization with non-purified proteins enhanced the titres of neutralizing antibodies. Again, the combination of the three proteins was able to confer total protection to immunized horses, which showed absence of viraemia. The antigenicity of recombinant VP2 was analysed with a collection of 30 MAbs. Both purified and unpurified recombinant VP2 proteins showed different antigenic patterns in comparison to that of VP2 on virions. An immunization experiment with four more horses con- firmed these results. The vaccine described here would not only prevent the disease, but would drastically reduce the propagation of the virus by vectors. Introduction African horsesickness (AHS) is a virus disease that causes morbidity in equids and a high mortality in horses (often exceeding 90%). The disease is caused by an arthropod-borne virus (African horsesickness virus; AHSV) within the genus Orbivirus of the family Reoviridae (Holmes, 1991). AHSV is transmitted to susceptible animals by biting midges (Culicoides spp.), which become infected by feeding on blood from sick animals containing high concentrations of infectious virus. The virus is mainly confined to sub-Saharan Africa, although severe epizootics have occurred on occasions in other parts of the world, including northern Africa, the Middle East and southern Europe (Lubroth, 1988; Rodrfguez et al., 1992; Mellor, 1993). As in other gnat-transmitted orbiviruses, AHSV contains seven structural proteins (VP1-VP7), which are organized into two concentric protein capsids (Oellermann et al., 1970), and a genome of 10 dsRNA segments (Bremer, 1976). The outer capsid consists of two major protein species, VP2 and VP5, of which VP2 * Author for correspondence. Fax + 34 1 4087598. (with a molecular mass of 124 kDa) is the major serotype- specific antigen (Bremer et al., 1990) and the main target for the neutralizing response of the host (Ranz et al., 1992; Burrage et al., 1993; Martinez-Torrecuadrada et al., 1994; Martinez-Torrecuadrada & Casal, 1995). The function of VP5 in protection remains unclear. The inner capsid is formed by two major proteins, VP3 and VP7, enclosing three other minor proteins, VP1, VP4 and VP6, that are closely associated with the virus genome (for review see Roy et al., 1994a). Like other virus diseases, vaccination and preventive measures are essential to protect horses against AHSV and to control the spread of the virus. Currently, both attenuated live virus vaccines and inactivated virus vaccines for AHSV are available commercially (Erasmus, 1978; Dubourget et al., 1992; House et al., 1994). Inactivated AHSV vaccine is generally preferred for two main reasons: (i) risks associated with the live vaccine can be avoided; and (it) infected animals can be distinguished from vaccinated animals (Laviada et at., 1995). However, like insufficient attenuation, incomplete inactivation of the virus is a threat to animal health. Recent developments in the use of baculovirus as a gene expression system have provided a powerful strategy for the development of a non-infectious, safe virus vaccine. 0001-3789 © 1996 SGM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of General Virology (1996), 77, 1211 1221. Printed in Great Britain 1211
Full protection against African horsesickness (AHS) in horses induced by baculovirus-derived AHS virus serotype 4 VP2, VP5 and VP7
Jorge L. Martinez-Torrecuadrada, 1 Maria Diaz-Laviada, 2 Polly Roy, 3.4 Carmen S~inchez, 2 Carmen Vela, 1 Jos6 Manuel S~inchez-Vizcaino 2 and J. Ignacio CasaP*
11NGENASA, Hnos Garc[a Noblejas 41, 2, 28037 Madrid, 2 Centro de hTvestigaci6n en Sanidad Animal-INIA, 28130 Valdeohnos, Madrid, Spain, 3 Laboratory o f Molecular Biophysics, Institute of Virology and Environmental Microbiology, University of Oxford, MansfieM Road, Oxford OX1 3SR, UK and 4 University o f Alabama, Birm#Tgham, A L 35294, USA
African horsesickness virus serotype 4 (AHSV-4) outer capsid protein VP2, or VP2 and VP5 plus inner capsid protein VP7, derived from single or dual recombinant baculovirus expression vectors were used in different combinations to immunize horses. When the proteins were purified by affinity chromatography, the com- bination of all three proteins induced low levels of neutralizing antibodies and conferred protection against virulent virus challenge. However, purified VP2 or VP2 and VP5 in the absence of VP7 failed to induce neutralizing antibodies and protection. Immunization with non-purified proteins enhanced the titres of
neutralizing antibodies. Again, the combination of the three proteins was able to confer total protection to immunized horses, which showed absence of viraemia. The antigenicity of recombinant VP2 was analysed with a collection of 30 MAbs. Both purified and unpurified recombinant VP2 proteins showed different antigenic patterns in comparison to that of VP2 on virions. An immunization experiment with four more horses con- firmed these results. The vaccine described here would not only prevent the disease, but would drastically reduce the propagation of the virus by vectors.
Introduction
African horsesickness (AHS) is a virus disease that causes morbidity in equids and a high mortality in horses (often exceeding 90%). The disease is caused by an arthropod-borne virus (African horsesickness virus; AHSV) within the genus Orbivirus of the family Reoviridae (Holmes, 1991). AHSV is transmitted to susceptible animals by biting midges (Culicoides spp.), which become infected by feeding on blood from sick animals containing high concentrations of infectious virus. The virus is mainly confined to sub-Saharan Africa, although severe epizootics have occurred on occasions in other parts of the world, including northern Africa, the Middle East and southern Europe (Lubroth, 1988; Rodrfguez et al., 1992; Mellor, 1993).
As in other gnat-transmitted orbiviruses, AHSV contains seven structural proteins (VP1-VP7), which are organized into two concentric protein capsids (Oellermann et al., 1970), and a genome of 10 dsRNA segments (Bremer, 1976). The outer capsid consists of two major protein species, VP2 and VP5, of which VP2
* Author for correspondence. Fax + 34 1 4087598.
(with a molecular mass of 124 kDa) is the major serotype- specific antigen (Bremer et al., 1990) and the main target for the neutralizing response of the host (Ranz et al., 1992; Burrage et al., 1993; Martinez-Torrecuadrada et al., 1994; Martinez-Torrecuadrada & Casal, 1995). The function of VP5 in protection remains unclear. The inner capsid is formed by two major proteins, VP3 and VP7, enclosing three other minor proteins, VP1, VP4 and VP6, that are closely associated with the virus genome (for review see Roy et al., 1994a).
Like other virus diseases, vaccination and preventive measures are essential to protect horses against AHSV and to control the spread of the virus. Currently, both attenuated live virus vaccines and inactivated virus vaccines for AHSV are available commercially (Erasmus, 1978; Dubourget et al., 1992; House et al., 1994). Inactivated AHSV vaccine is generally preferred for two main reasons: (i) risks associated with the live vaccine can be avoided; and (it) infected animals can be distinguished from vaccinated animals (Laviada et at., 1995). However, like insufficient attenuation, incomplete inactivation of the virus is a threat to animal health. Recent developments in the use of baculovirus as a gene expression system have provided a powerful strategy for the development of a non-infectious, safe virus vaccine.
0001-3789 © 1996 SGM

1212 J. L. Martinez-Torrecuadrada and others
It has been reported that baculovirus-expressed outer capsid protein VP2, or VP2 plus VP5, of bluetongue virus (BTV), the prototype of the genus Orbivirus, is able to induce protective immunity against bluetongue disease in sheep (Roy et al., 1990). Such protection lasted for a long period ( > 15 months) when baculovirus-synthesized virus-like particles (VLPs) consisting of VP2, VP3, VP5 and VP7 were tested (French et al., 1990; Roy et al., 1992, 1994b).
We recently reported the construction of recombinant Autographa californica multinucleocapsid nuclear poly- hedrosis viruses that expressed AHSV-4 VP2 alone or together with VP5 (Martinez-Torrecuadrada et al., 1994) and VP7 (Chuma et al., 1992). Subsequently, we have demonstrated that antisera raised against purified recom- binant VP2 in rabbits were able to neutralize AHSV-4 in vitro (M artinez-Torrecuadrada & Casal, 1995). However, in spite of its relevance to vaccine design, systematic antigenic analysis of baculovirus-expressed VP2 derived from either AHSV or BTV has not beeninvestigated to date. Synthesis of a recombinant protein in a hetero- logous system may lead to inappropriate folding, which may affect the proper presentation of the antigens. Therefore, an additional objective of this study was to examine the antigenicity of recombinant VP2 and the effect of expression in insect cells on the biological activity of the protein. Previous vaccination studies with BTV VP2 involved only infected cell extracts containing VP2 protein. For safety and from a regulatory point of view, purified antigen, free of cellular contaminants, would be preferable. Therefore, for this study purified proteins as well as infected cell extracts were used in the immunization protocols to investigate the potential for inducing complete protection in horses against AHSV. Since AHSV VLPs are not yet available, we analysed the available recombinant proteins for their potential as candidate vaccines. In this report, we describe the evaluation of the protective properties of several combi- nations of recombinant AHSV-4 VP2, VP5 and VP7 in horses, the natural host of AHSV.
Methods Cells and viruses. AHSV-4 was grown and assayed in confluent
monolayers of Vero cells (ATCC CCL81) in Dulbecco's modified Eagle's medium containing 5 % fetal calf serum (FCS) and antibiotics at 37 °C. Purified AHSV particles were obtained according to a procedure described previously (Mertens et al., 1987). AHSV-4/89 isolate that had been isolated from infected cells (Marfinez- Torrecuadrada et al., 1994) was used for challenge. After three passages on Vero cell monolayers, the virus was titrated by plaque assay as described previously (OeUermann, 1970) and stored at - 8 0 °C.
The Spodopterafrugiperda cell clone Sf9 (ATCC CRLIT11) was used to propagate recombinant baculoviruses. Sf9 cells were grown and maintained in suspension or monolayer cultures using TNM-FH medium supplemented with 5% FCS plus antibiotics at 27 °C.
Recombinant baculoviruses AcAHSV-4.2, which expresses the VP2 protein of AHSV-4 (Martinez-Torrecuadrada et al., 1994), AcAHSV- 4.2/5, which synthesizes AHSV-4 VP2 and VP5 simultaneously (Martinez-Torrecuadrada et al., 1994) and AcAHSV-4.7, expressing AHSV-4 VP7 (Chuma et al., 1992), were used for antigen preparation.
Antisera and MAbs. Horse sera from different sources, either from AHSV-4-infected horses or from horses vaccinated with live attenuated monovalent vaccine specific for AHSV-4, were used as positive controls.
Thirty VP2-specific MAbs defining nine different antigenic sites ~Ranz et al., 1992) were kindly provided by Dr A. Sanz (INGENASA, Madrid, Spain). They were used to characterize the antigenic structure of baculovirus-expressed VP2.
Animals. Fifteen mixed breed horses of various ages and sizes, seronegative for AHSV, were used for the vaccination experiments. Two weeks before the experiment, the horses were housed at bio- containment level 3 in the Centro de Investigaci6n en Sanidad Animal (INIA, Valdeolmos, Madrid. Spain).
Preparation o f recombinant antigens for horse inoculation. Sf9 cells were infected at a multiplicity of 2 5 p.f.u./cell with each recombinant baculovirus. Cells were harvested at 72 h post-infection by centri- fugation at 200 g for l0 min. washed once with PBS and resuspended in PBS at a density of 3-7 z 10 v cells per ml. A portion of each sample was analysed by SDS PAGE and Coomassie blue staining to estimate the amounts of AHSV-4 proteins present. Each sample was then divided into aliquots and stored at - 7 0 °C until the day of immunization.
For preparation of purified baculovirus-expressed proteins, infected cells were harvested at 72 h, washed with PBS and lysed by osmotic shock with 25 mM-NaHCQ pH 8'3. Nuclei and cellular debris were then removed by centrifugation at 12 000 g. VP2 and VP5 purifications were achieved by affinity chromatography using IgGs purified from a polyclonal anti-AHSV-4 horse serum and specific MAbs as described previously (Martinez-Torrecuadrada et al., 1994). VP7 was purified by banding on a 30 % to 80 % (w/v; 10 ml) continuous sucrose gradient (Chuma et al., 1992). Each purified sample was subsequently stored at - 2 0 °C until use in the immunization experiments.
Horse immunization and challenge. Three groups of horses were used for the immunization experiments. They were immunized in the presence of adjuvants by the intramuscular route with one dose of AHSV proteins as indicated in Table 1. A booster was given 30 days after the first immunization. Alumina (Alhydrogel; Superfos) plus 500 lag of Quil A (Superfos) per horse were used as adjuvants. Adjuvants such as Freund's adjuvant and other oil emulsion adjuvants were not used as horses are generally hypersensitive to oil emulsions. Control animals received a placebo of saline plus adjuvant~ Blood samples for serological assays were taken before the start of the experiments and at approximately weekly intervals until the challenge.
For challenge, horses received 106 TCIDs0 of AHSV-4/89 in 1 ml of saline, intravenously in the jugular vein, 21-22 days after the second dose of vaccine. Clinical manifestations of the disease including rectal temperatures were observed from 1 to 21 days post-challenge. Blood was collected daily from each horse until the animals died or were sacrificed, in order to determine the virus and antibody titres.
ELISA. For detection of specific antibodies against the whole virion or VP7, an indirect ELISA was carried out similar to that previously described (Laviada etal . , 1992a). Briefly, polystyrene plates (Nunc) were coated with 0.5 lag of CsCI gradient-purified AHSV-4 or 1 lag of purified VP7 in 50 m~a-carbonate buffer pH 9.6 overnight at 4 oC. Washes between steps were performed with 0.05% Tween-20 in distilled water. Horse sera were serially diluted in blocking buffer (0.05 % Tween-20, 10 % FCS in PBS) and incubated for 1 h at 37 °C.

Protection against A HS V in horses 1213
Table 1. AHSV-4 recombinant antigens used in vaccination experiments
Antigen Group Horse (dose ~tg) Form Protection
I 1 VP2 (100) Purified*
II
III
2 VP2 (100) Purified VP5 (50)
3 VP2 (100) Purified + vP5 (50) VP7 (100)
4 Saline
5 VP2 (100) + VP7 (100)
6 VP2 (100) VP7 (100)
7 VP2 (100) vP5 (50) VP7 (100)
8 VP2 (~ 100) VP7 (~ 100)
9 VP2 (~ 100) + VP5 (~ 100) VP7 (~ 100)
Saline
VP2 (,.~ 100) VP5 (~ 100) VP7 (~ 100)
Purified
Purified
Purified
7 x 107 AcAHSV-4.2-infected cells 5 x 106 AcAHSV-4.7-infected cells
6 x 107 AcAHSV-4.2/5-infected cells
5 x 106 AcAHSV-4.7-infected cells
10
1 t 6 x 107 AcAHSV-4.2/5-infected cells + 12 + 13 5 x 10 ~ AcAHSV-4.7-infected cells + 14 +
15 Saline
* VP2 and VP5 were purified by affinity chromatography; VP7 was purified by sucrose gradient centrifugation.
Peroxidase-labelled anti-horse IgG (Sigma) at optimum dilution in blocking buffer was used as conjugate. Colour was developed by adding ABTS [2,2'-azinobis(3-ethylbenzthiazolinesulphonic acid); Sigma] as substrate. The reaction was stopped with 1% SDS and the absorbance was measured at 405 nm using an ELISA reader. Titres were determined as the reciprocal of the highest dilution giving three times the blank (pre-immunization sera) A40 ~.
Detection of VP2- and VP5-specific antibodies was performed by a double-antibody sandwich ELISA (DAS-ELISA). Basically, microtitre plates were coated with 1 lag/well of purified anti-VP2 MAb (8DF 1) or purified anti-VP5 MAb (10AE12) (Ranz et al., 1992) in PBS by overnight incubation at 4 °C. After five washes, plates were incubated with AcAHSV-4.2/5-infected cell extracts diluted 1/100 in PBS for 1 h at 37 °C, washed again and incubated with serial dilutions of horse sera for 1 h at 37 °C. Bound antibodies were detected as described above.
To test the antigenic structure of recombinant VP2 an indirect ELISA was designed. Plates were coated with three types of antigens, 0.5 lag/well of CsCl-purified AHSV-4 virus, 0.2 lag/well of purified recombinant VP2 or 100 lal/well of AcAHSV-4.2-infected cell extracts lysed by osmotic shock (diluted 1:100 in carbonate buffer). VP2- specific MAbs and peroxidase-labelled rabbit anti-mouse IgG (Sigma) were added as first and conjugate antibodies, respectively.
Virus neutralization test and viraemia assays. To determine the ability of the horse sera to neutralize the virus #l vitro, a Veto cell monolayer protection assay was used as described previously (Martinez- Torrecuadrada et al., 1994).
To detect red blood cell-associated virus, horse erythrocytes were washed three times in PBS at 4 °C and resuspended in PBS containing 30 lag/ml of gentamicin for storage at 4 °C until testing. Immediately
before using, red blood cells were lysed by osmotic shock in distilled water for 30 s. The ionic strength of the lysed cell sample was subsequently restored by diluting 10 times with PBS and cells were sonicated for 30 s to ensure complete disruption. Each lysed sample was seeded onto monolayers of Vero cells. The monolayers were observed 5-7 days post-infection for CPE. When needed, three consecutive passages were made in Veto cells. Viraemia titres of each viraemic blood sample were determined by plaque assay on Vero cell monolayers as described previously (Oellermann, 1970). CPE was confirmed by an AHSV-specific DAS-ELISA (Laviada et al., 1992b).
To confirm the viraemia, suckling BALB/c mice between 1 and 3 days of age were inoculated intracranially with 30 ~tl per mouse of lysed erythrocytes diluted 1/10 in saline. Mice were observed for the development of neurological disorders for 15 days. Brains from mice which showed such clinical symptoms or died were examined for virus replication by inoculating Vero cells.
Resul t s Antigenic characterization of baculovirus-expressed AHSV-4 VP2
To determine the antigenic properties of the baculovirus- expressed protein compared to the authentic VP2 protein present in the virion, we analysed the antibody-binding ability of VP2 obtained by three different methods: (i) non-purified recombinant VP2, (ii) purified recombinant VP2 and (iii) native AHSV-4 VP2 in the form of intact
1214 J. L. Mart[nez-Torrecuadrada and others
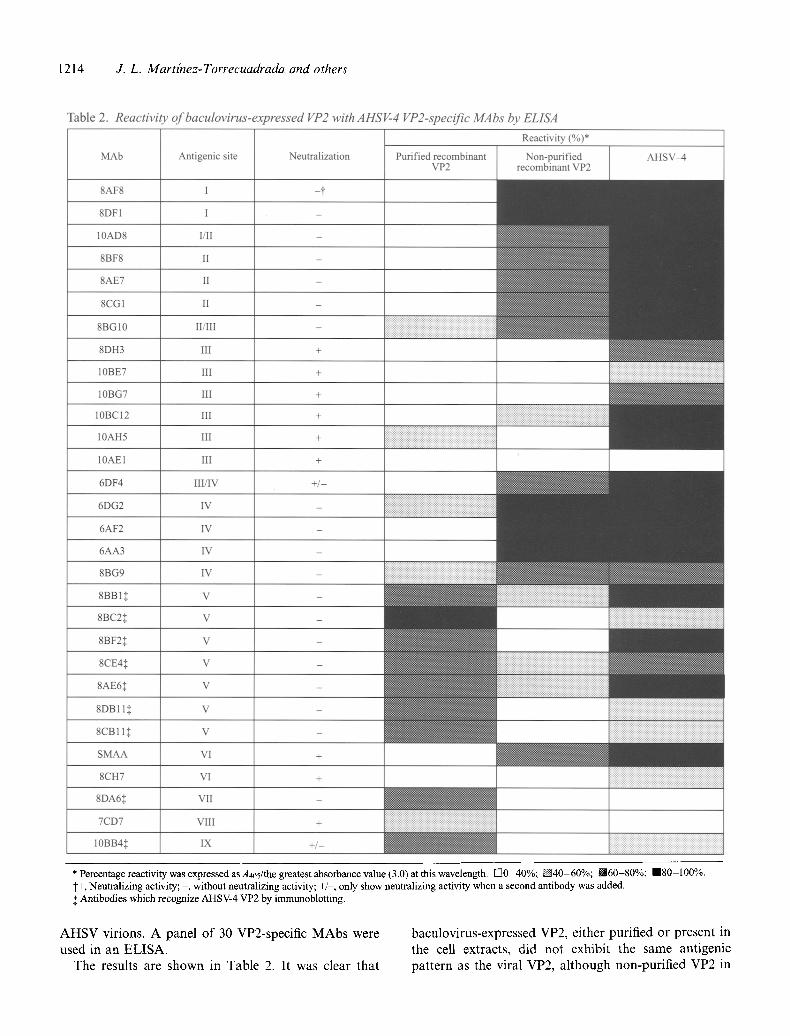
Table 2. Reactivitv of baculovirus-expressed VP2 with AHSV-4 VP2-specific MAbs by ELISA
MAb
8AF8
8DF 1
10AD 8
8BF8
8AE7
8CG 1
8BG10
8DH3
Antigenic site
I/II
II/III
III
Neutralization
10BE7 III +
10BG7 III +
10BC12 III +
10AH5
IOAE1
6DF4
6DG2
III
III
II1/IV
IV
IV
IV
IV
V
V
g m
V
V
V
6AF2
6AA3
8BG9
8BBI~
8BC2:~
8BF2~
8CE43~
8AE6:~
8DBll~
8CB11:~ V
SMAA VI +
8CH7 VI +
8DA6~: VII
+/
7CD7 VIII +
10BB4:~ IX +/_
Reactivity (%)*
Purified recombinant Non-purified AHSV-4 VP2 recombinant VP2
iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii!iiiiiiiiiiiiiiiiii i iiii i iiiiiiiii
i i! ! .~iiiiiiiiiiiiiii~!iliiiiiiiiii!iiiiiiiiiiiiiiiiiiiii~i~!~:~iiiiiiiiiii .......................
* Percentage reactivity was expressed as A4•5/the greatest abs•rbance va•ue (3.•) at this wave•ength. I-]0 40%; D40-60%; 160-80%; m80-100%. t+, Neutralizing activity; - , without neutralizing activity; +/-, only show neutralizing activity when a second antibody was added. ~: Antibodies which recognize AHSV-4 VP2 by immunoblotting.
AHSV virions. A panel of 30 VP2-specific MAbs were used in an ELISA.
The results are shown in Table 2. It was clear that
baculovirus-expressed VP2, either purified or present in the cell extracts, did not exhibit the same antigenic pattern as the viral VP2, although non-purified VP2 in
Protection against A H S V in horses 1215
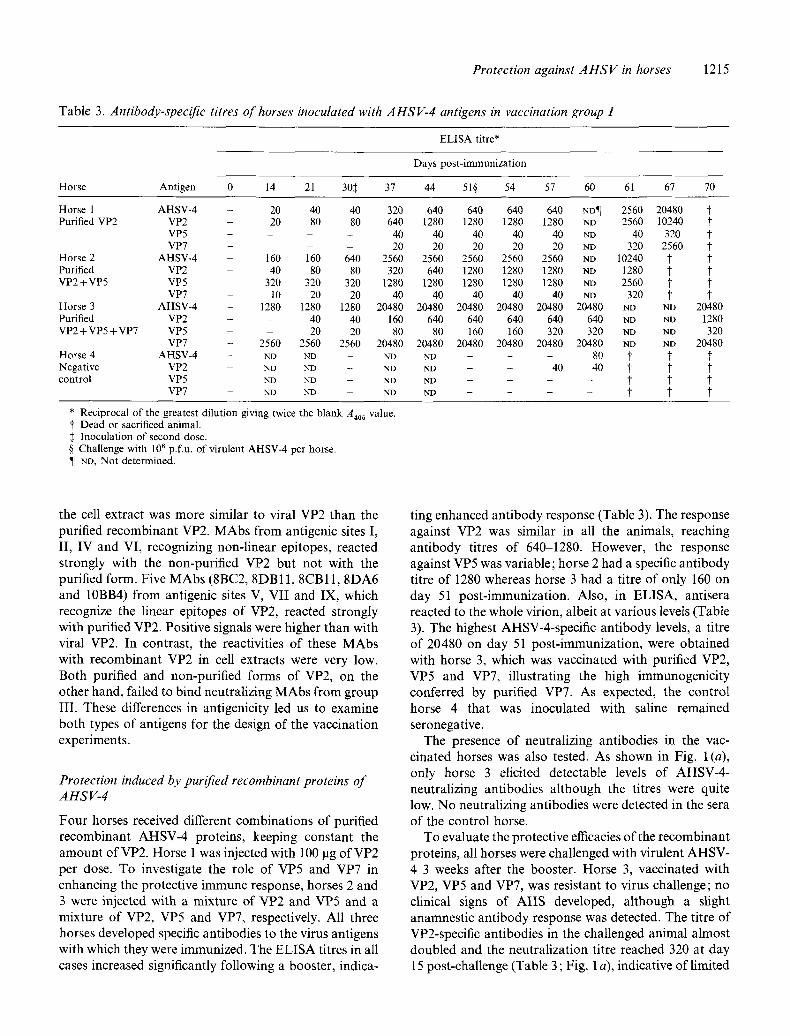
Table 3. Antibody-specific titres of horses inoculated with AHSV-4 antigens in vaccination group I
E L I S A titre*
D a y s p o s t - i m m u n i z a t i o n
H o r s e A n t i g e n 0 14 21 302 37 44 51§ 54 57 60 61 67 70
H o r s e 1 A H S V - 4 - 20 40 40 320 640 640 640 640 Nr~¶ 2560 20480 t Pur i f ied V P 2 V P 2 - 20 80 80 640 1280 1280 1280 1280 ND 2560 10240 t
VP5 . . . . 40 40 40 40 40 ND 40 320 t V P 7 - - - 20 20 20 20 20 ND 320 2560 ~"
H o r s e 2 A H S V - 4 - 160 160 640 2560 2560 2560 2560 2560 ND 10240 t f Pur i f ied VP2 - 40 80 80 320 640 1280 1280 1280 ND 1280 "~ t V P 2 + V P 5 VP5 - 320 320 320 1280 1280 1280 1280 1280 ND 2560 "j" ~"
VP7 - 10 20 20 40 40 40 40 40 ND 320 t ~" H o r s e 3 A H S V - 4 - 1280 1280 1280 20480 20480 20480 20480 20480 20480 NO ND 20480 Pur i f ied V P 2 - - 40 40 160 640 640 640 640 640 ND ND 1280 V P 2 + VP5 + VP7 VP5 - - 20 20 80 80 160 160 320 320 ND ND 320
V P 7 - 2560 2560 2560 20480 20480 20480 20480 20480 20480 NO ND 20480 H o r s e 4 A H S V - 4 - ND ND ND N D - - - - - 80 ~" "~ N e g a t i v e V P 2 - NO NO -- NO NO -- -- 40 40 t t t c o n t r o l VP5 ND NO -- NO ND -- -- "~ "~ t
V P 7 - ND ND ND ND . . . . t t t
* R e c i p r o c a l o f the g rea te s t d i l u t i on g iv ing twice the b l a n k A405 value . f D e a d o r sacr i f iced an ima l . :~ I n o c u l a t i o n o f s e c o n d dose . § C h a l l e n g e w i th 106 p.f .u , o f v i ru l en t A H S V - 4 pe r horse . ¶ ND, N o t de t e rmined .
the cell extract was more similar to viral VP2 than the purified recombinant VP2. MAbs from antigenic sites I, II, IV and VI, recognizing non-linear epitopes, reacted strongly with the non-purified VP2 but not with the purified form. Five MAbs (8BC2, 8DB11, 8CB11, 8DA6 and 10BB4) from antigenic sites V, VII and IX, which recognize the linear epitopes of VP2, reacted strongly with purified VP2. Positive signals were higher than with viral VP2. In contrast, the reactivities of these MAbs with recombinant VP2 in cell extracts were very low. Both purified and non-purified forms of VP2, on the other hand, failed to bind neutralizing MAbs from group III. These differences in antigenicity led us to examine both types of antigens for the design of the vaccination experiments.
Protection induced by purified recombinant proteins of AHS V-4
Four horses received different combinations of purified recombinant AHSV-4 proteins, keeping constant the amount of VP2. Horse 1 was injected with 100 lag of VP2 per dose. To investigate the role of VP5 and VP7 in enhancing the protective immune response, horses 2 and 3 were injected with a mixture of VP2 and VP5 and a mixture of VP2, VP5 and VP7, respectively. All three horses developed specific antibodies to the virus antigens with which they were immunized. The ELISA titres in all cases increased significantly following a booster, indica-
ting enhanced antibody response (Table 3). The response against VP2 was similar in all the animals, reaching antibody titres of 640-1280. However, the response against VP5 was variable; horse 2 had a specific antibody titre of 1280 whereas horse 3 had a titre of only 160 on day 51 post-immunization. Also, in ELISA, antisera reacted to the whole virion, albeit at various levels (Table 3). The highest AHSV-4-specific antibody levels, a titre of 20480 on day 51 post-immunization, were obtained with horse 3, which was vaccinated with purified VP2, VP5 and VP7, illustrating the high immunogenicity conferred by purified VP7. As expected, the control horse 4 that was inoculated with saline remained seronegative.
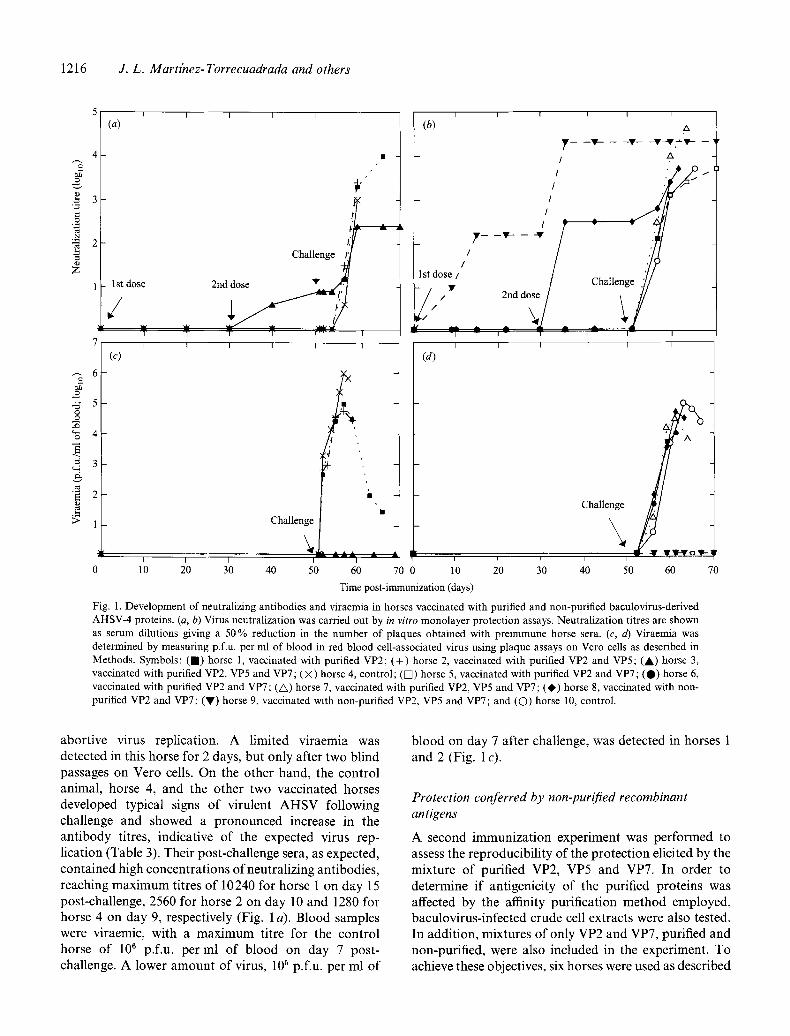
The presence of neutralizing antibodies in the vac- cinated horses was also tested. As shown in Fig. 1 (a), only horse 3 elicited detectable levels of AHSV-4- neutralizing antibodies although the titres were quite low. No neutralizing antibodies were detected in the sera of the control horse.
To evaluate the protective efficacies of the recombinant proteins, all horses were challenged with virulent AHSV- 4 3 weeks after the booster. Horse 3, vaccinated with VP2, VP5 and VP7, was resistant to virus challenge; no clinical signs of AHS developed, although a slight anamnestic antibody response was detected. The titre of VP2-specific antibodies in the challenged animal almost doubled and the neutralization titre reached 320 at day 15 post-challenge (Table 3 ; Fig. 1 a), indicative of limited
1216 J. L. Mart[nez-Torrecuadrada and others
@
z
O Q
I I I I I I I I I I I I
(a) (b) LX
7 - -Iv-- - - Y - - v - - v ~ - - -
4
• / ' / /
: / /~ - - - - V - - - - -~
2 Challenge
/ d ~ st dose / Challe
1 1st dose 2n / / ~ 2nd dose
- ~ T Y T r . . . . . . . i ~ i i A _
I I I I I I I I I [ I
(c) (d) 6
5 m -
4 ¢ , i , , "" !
3
2 i - m Challenge
1 Challenge _ \
0 110 210 310 410 510 6; 70 0 10 20 30 40 50 60 70
Time post-immunization (days)
Fig. 1. Development of neutralizing antibodies and viraemia in horses vaccinated with purified and non-purified baculovirus-derived AHSV-4 proteins. (a, b) Virus neutralization was carried out by in vitro monolayer protection assays. Neutralization titres are shown as serum dilutions giving a 50 % reduction in the number of plaques obtained with preimmune horse sera. (c, d) Viraemia was determined by measuring p.f.u, per ml of blood in red blood cell-associated virus using plaque assays on Vero cells as described in Methods. Symbols: (m) horse 1, vaccinated with purified VP2; (+) horse 2, vaccinated with purified VP2 and VP5; (A) horse 3, vaccinated with purified VP2, VP5 and VP7; (×) horse 4, control; (D) horse 5, vaccinated with purified VP2 and VP7; (O) horse 6, vaccinated with purified VP2 and VP7; (A) horse 7, vaccinated with purified VP2, VP5 and VP7; (0) horse 8, vaccinated with non- purified VP2 and VP7; (V) horse 9, vaccinated with non-purified VP2, VP5 and VP7; and (O) horse 10, control.
abortive virus replication. A limited viraemia was detected in this horse for 2 days, but only after two blind passages on Vero cells. On the other hand, the control animal, horse 4, and the other two vaccinated horses developed typical signs of virulent AHSV following challenge and showed a pronounced increase in the antibody titres, indicative of the expected virus rep- lication (Table 3). Their post-challenge sera, as expected, contained high concentrations of neutralizing antibodies, reaching maximum titres of 10240 for horse 1 on day 15 post-challenge, 2560 for horse 2 on day 10 and 1280 for horse 4 on day 9, respectively (Fig. 1 a). Blood samples were viraemic, with a maximum titre for the control horse of 106 p.f.u, per ml of blood on day 7 post- challenge. A lower amount of virus, 105 p.f.u, per ml of
blood on day 7 after challenge, was detected in horses 1 and 2 (Fig. 1 c).
Protection conferred by non-purified recombinant antigens
A second immunization experiment was performed to assess the reproducibility of the protection elicited by the mixture of purified VP2, VP5 and VP7. In order to determine if antigenicity of the purified proteins was affected by the affinity purification method employed, baculovirus-infected crude cell extracts were also tested. In addition, mixtures of only VP2 and VP7, purified and non-purified, were also included in the experiment. To achieve these objectives, six horses were used as described
Protection against AHSV in horses 1217
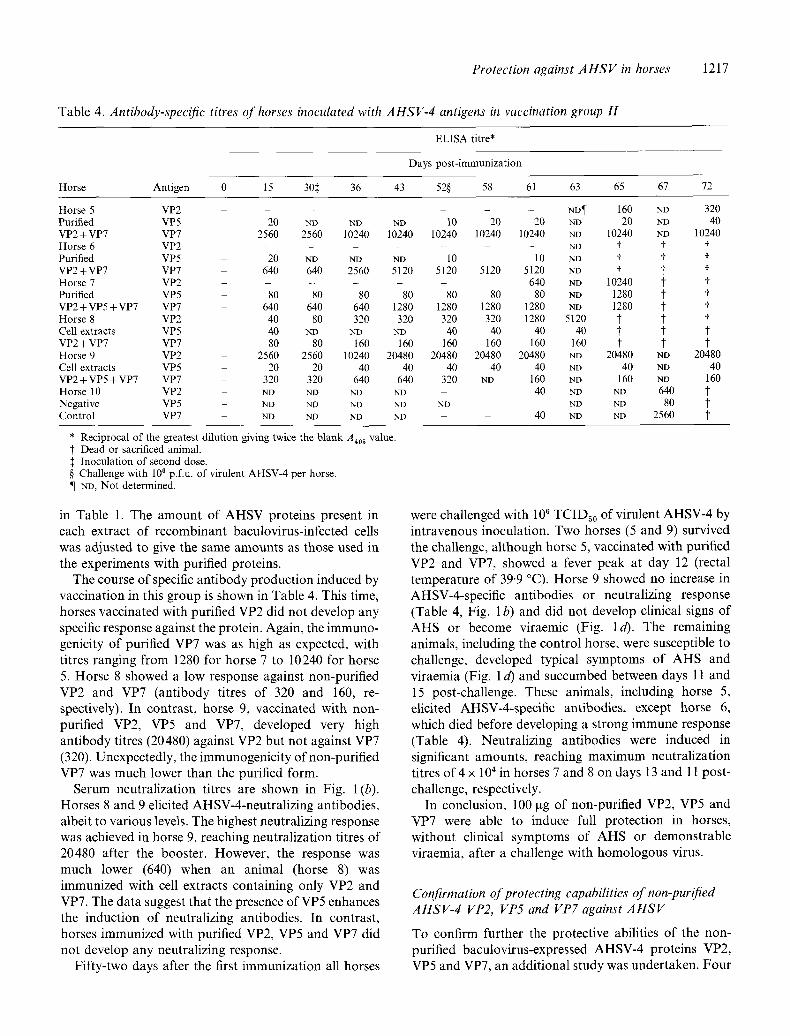
Table 4. Antibody-specific titres of horses inoculated with AHSV-4 antigens in vaccination group H
ELISA titre*
Days post-immunization
Horse Antigen 0 15 30:~ 36 43 52§ 58 61 63 65 67 72
Horse 5 VP2 . . . . . . N D ¶ 160 ND 320 Purified VP5 20 ND ND ND 10 20 20 ND 20 ND 40 VP2+VP7 VP7 2560 2560 10240 10240 10240 10240 10240 ND 10240 ND 10240 H o r s e 6 V P 2 . . . . . . ND "~ "~
Purified VP5 20 ND ND ND 10 10 ND ~" t t VP2+VP7 VP7 640 640 2560 5120 5120 5120 5120 ND t t t Horse 7 VP2 - - - 640 NO 10240 ~" t Purified VP5 80 80 80 80 80 80 80 ND 1280 t t VP2 +VP5+VP7 VP7 640 640 640 1280 1280 1280 1280 ND 1280 t t Horse 8 VP2 40 80 320 320 320 320 1280 5120 ~" t t Cell extracts VP5 - 40 ND ND ND 40 40 40 40 t" t t" VP2+VP7 VP7 - 80 80 160 160 160 160 160 160 t t t Horse 9 VP2 2560 2560 10240 20480 20480 20480 20480 ND 20480 ND 20480 Cell extracts VP5 - 20 20 40 40 40 40 40 ND 40 ND 40 VP2 + VP5 + VP7 VP7 320 320 640 640 320 ND 160 ND 160 ND 160 Horse 10 VP2 ND NP ND ND 40 ND ND 640 t Negative V P 5 - ND ND ND ND ND -- ND ND 8 0 "~
Control VP7 ND ND ND NO 40 ND NO 2560 t
* Reciprocal of the greatest dilution giving twice the blank A40 ~ value. t Dead or sacrificed animal. :~ Inoculation of second dose. § Challenge with 106 p.f.u, of virulent AHSV-4 per horse. ¶ ND, Not determined.
in Table 1. The amoun t o f A H S V proteins present in each extract o f recombinant baculovirus-infected cells was adjusted to give the same amounts as those used in the experiments with purified proteins.
The course o f specific an t ibody produc t ion induced by vaccinat ion in this group is shown in Table 4. This time, horses vaccinated with purified VP2 did not develop any specific response against the protein. Again, the immuno- genicity o f purified VP7 was as high as expected, with titres ranging f rom 1280 for horse 7 to 10240 for horse 5. Horse 8 showed a low response against non-purif ied VP2 and VP7 (ant ibody titres o f 320 and 160, re- spectively). In contrast , horse 9, vaccinated with non- purified VP2, VP5 and VP7, developed very high ant ibody titres (20480) against VP2 but not against VP7 (320). Unexpectedly, the immunogenic i ty o f non-purified VP7 was much lower than the purified form.
Serum neutral izat ion titres are shown in Fig. 1 (b). Horses 8 and 9 elicited AHSV-4-neutral iz ing antibodies, albeit to various levels. The highest neutralizing response was achieved in horse 9, reaching neutral ization titres o f 20480 after the booster. However , the response was much lower (640) when an animal (horse 8) was immunized with cell extracts containing only VP2 and VP7. The data suggest that the presence o f VP5 enhances the induct ion o f neutralizing antibodies. In contrast , horses immunized with purified VP2, VP5 and VP7 did not develop any neutralizing response.
Fif ty- two days after the first immunizat ion all horses
were challenged with 106 TCIDs0 of virulent AHSV-4 by intravenous inoculation. Two horses (5 and 9) survived the challenge, a l though horse 5, vaccinated with purified VP2 and VP7, showed a fever peak at day 12 (rectal temperature o f 39-9 °C). Horse 9 showed no increase in AHSV-4-specific antibodies or neutralizing response (Table 4, Fig. 1 b) and did no t develop clinical signs o f A H S or become viraemic (Fig. l d). The remaining animals, including the control horse, were susceptible to challenge, developed typical symptoms of A H S and viraemia (Fig. 1 d) and succumbed between days l 1 and 15 post-challenge. These animals, including horse 5, elicited AHSV-4-specific antibodies, except horse 6, which died before developing a s trong immune response (Table 4). Neutral izing antibodies were induced in significant amounts , reaching max imum neutral ization titres o f 4 x 10 ~ in horses 7 and 8 on days 13 and 1l post- challenge, respectively.
In conclusion, 100 gg o f non-purif ied VP2, VP5 and VP7 were able to induce full protect ion in horses, wi thout clinical symptoms of A H S or demonstrable viraemia, after a challenge with homologous virus.
Confirmation of protecting capabilities of non-purified AHSV-4 VP2, VP5 and VP7 against AHSV
To confirm further the protective abilities o f the non- purified baculovirus-expressed AHSV-4 proteins VP2, VP5 and VP7, an addit ional study was undertaken. F o u r
1218 J. L. Martinez-Torrecuadrada and others
%
<
2
0
Z
I I I I I I
(a)
2nd dose
- - - - - - - ~ . . . . I . . . . 4 - - - - ~ l . . . .
Z Z Z
II /
"L "F
I i i y I
X A I I
I / I!
Challenge I/L l/
U
(b) I I I I [ I
1st d o s <
/ 2nd dose
i
I I
0 10 2o 3'o 4o Time post-immunization (days)
/
/ !
I
I
I
Challenge I I
-N- --N I
50 60 70
Fig. 2. Time-course of neutralizing and VP2-, VP5- and VP7-specific antibodies in horses vacci- nated with insect cell extracts containing AHSV- 4 VP2, VP5 and VP7. The results are given for four vaccinated horses and one non-vaccinated control. (a) ELISA for detection of AHSV-4- specific antibodies using baculovirus-derived VP2 (.) , VP5 (A) and VP7 ()<) as antigens. Detection of anti-VP7 antibodies was carried out by indirect ELISA, whereas anti-VP2 and anti-VP5 anti- bodies were detected by DAS-ELISA (see Methods). Solid lines represent data from vacci- nated horses and dotted lines from the control horse. Titres were given as the arithmetic mean _+so of log10 titres. (b) Neutralizing antibodies developed by vaccinated (B) and non-vaccinated (X) horse pre- and post-challenge were deter- mined by bt vitro monolayer protection. Neutral- ization titres are given as serum dilutions giving a 50 % reduction in the number of plaques obtained with preimmune horse sera.
horses were injected intramuscularly with two doses of Sf9 cells infected with recombinant viruses (6 x 107 cells for AcAHSV-4.2/5 and 5 x l0 s cells for AcAHSV-4.7; Table 1). A fifth horse received only saline as control.
In all vaccinated animals, anti-AHSV-4 and specific antibody titres were detected. As in horse 9 of the previous experiment, a very high level of VP2-specific antibodies was elicited in all vaccinated horses (Fig. 2 a), reaching titres of 10 240-20 480 after the booster immuni- zation. Also, the response against non-purified VP7 was lower than that induced by non-purified VP2, with an average titre of 640. In addition, the level of virus- neutralizing antibodies to AHSV-4 increased signi- ficantly (average neutralization titres > 104) following the booster injection (Fig. 2b), which was in agreement with previous results obtained with horse 9. The control
horse remained seronegative until after the challenge seroconversion.
Experimental exposure to AHSV-4 led to infection of the non-vaccinated horse, which showed typical clinical signs of AHS such as supraorbital oedema, congestion of oral and conjunctival mucosa, cardiac symptoms, res- piratory distress and prostration. In contrast, the vaccinated animals were completely protected from virulent virus challenge, remaining healthy without any clinical signs or detectable viraemia. The levels of specific and neutralizing antibodies were also maintained despite the severity of the challenge inoculation, indicating lack of virus replication in all the vaccinated horses. As expected, the post-challenge blood of the control horse was viraemic from day 6 (data not shown) and the serum was seropositive to AHSV-4 from day 7, which indicated

Protection against A H S V in horses 1219
virus replication (Fig. 2a). It is clear from this study that the recombinant vaccine efficiently protect~d horses against infection with the homologous virulent AHSV.
Discussion
Outer shell proteins VP2 and VP5 from orbiviruses have been shown to play a major role in virus neutralization (Inumaru & Roy, 1989). The use of baculovirus-derived outer capsid proteins from BTV in the development of subunit vaccines is extensively documented (Roy et al., 1990). They have been used individually or co-expressed with inner core proteins VP3 and VP7 to form VLPs (Roy et al., 1992). The efficacy of these VLPs in terms of induction of neutralizing antibodies and protection seems superior to that of the individual proteins. As little as 10 lag of VLPs fully protected sheep against virulent BTV even when challenged 15 months post-vaccination. Since AHSV-4 VLPs are not currently available, we decided to test the immunogenicity of the three available recombinant proteins in various combinations in the natural host, and to evaluate their protection efficacies against virulent AHSV-4.
In this report, we have studied for the first time the antigenicity of the recombinant baculovirus-expressed VP2, both in purified and unpurified form. Such information is relevant for the design of a candidate vaccine. A collection of 30 VP2-specific MAbs allowed us a precise antigenic comparison between the recom- binant VP2 and the authentic AHSV VP2. Our data clearly indicate that the antigenic properties of recom- binant VP2 protein, both in purified form and in recombinant virus-infected cell extract, differ signi- ficantly from that of virus VP2. Although the neutralizing MAbs did not recognize the unpurified VP2 in the infected cell extracts, the protein was recognized by most of the MAbs specific for non-linear epitopes. These results suggest that recombinant VP2, when expressed individually, does not possess the conformation necess- ary to expose the immunodominant neutralization epitopes. Purified VP2, on the other hand, was exclusively recognized by MAbs against linear epitopes (sites V, VII and IX). It is probable that the purification process altered the conformation of the recombinant VP2 in such a way that only the linear epitopes were recognized. This is probably due to the use of chaotropic agents for elution of VP2 from the affinity column.
Subsequently, the protective capability of recombinant VP2 in combination with VP5 and VP7 was determined. Combinations of VP5 and VP7 alone were not tested for two major reasons: (i) antibodies raised against BTV VP5 do not neutralize BTV infectivity (Yang et al., 1992); and (ii) previous immunization of rabbits with AHSV VP7 alone showed a total absence of neutralizing
antibodies in serum (J. L. Martinez-Torrecuadrada & J. I. Casal, unpublished results). The immunization experi- ments were performed on three groups of horses. The first group of horses were vaccinated only with purified proteins. In these conditions, VP2 alone or together with VP5 was unable to protect the horses. Only a mixture of the three proteins VP2, VP5 and VP7 induced low levels of neutralizing antibodies, but sufficient to confer protection. The slight increase in neutralizing antibodies after challenge is indicative of an anamnestic response. Our results are unlike those of previous studies on BTV (Roy et al., 1990), where VP2 alone fully protected sheep against the disease. Furthermore, the presence of purified VP5 did not result in an enhancement of protection in these conditions. Several factors might contribute to these differences. These include the use of purified proteins instead of cell extracts and a different antigenic structure of VP2 and VP5 in AHSV with respect to BTV.
A second group of horses was immunized with recombinant virus-infected cell extracts and purified proteins stored at - 2 0 °C for 7 months. The use of cellular extracts improved the development of neutra- lizing antibodies by three orders of magnitude. There- fore, in cell extracts and especially when co-expressed with VP5, the recombinant VP2 preserved the natural conformation better than the purified protein. It is possible that the interaction between VP2 and VP5 preserves the correct conformation of the VP2 molecule and consequently induces a more effective subset of neutralizing antibodies. Also, VP5 and probably VP7 may contain important T cell epitopes, which could enhance the immune response to the virus. In some cases, horses developed significant titres of neutralizing anti- bodies without protection, as in the case of the horse vaccinated with cell extracts containing VP2 and VP7. On the other hand, horse 5, which survived the challenge, had no neutralizing antibodies. This result implies that there is not a direct correlation between neutralization in vitro and protection in vivo, and perhaps other immune mechanisms (e.g. cell-mediated immunity) are involved in the process of protection. Similar evidence was found in BTV infection (for a review see Osburn, 1992), where sheep could resist challenge with active virus in the absence of neutralizing antibody.
The superior immunogenicity of purified VP7 is most likely explained by the fact that VP7 is over-expressed at very high levels in the infected cells, inducing disc-shaped crystals (Chuma et al., 1992). Crystalline intracellular VP7 might be less immunogenic that the purified VP7, which has been solubilized and has probably renatured enough to recover its major conformational immuno- dominant epitope.
Several conclusions were obtained from this exper- iment: (i) non-purified VP2 is much more immunogenic

1220 J. L. Mart[nez-Torrecuadrada and others
when it is used together with VP5 and VP7, otherwise immunogenicity is similar to that of the purified form; (ii) VP7 seems more immunogenic when it is purified; and (iii) the affinity column-purified forms of VP2 and VP5 are not only poor immunogens, but they are also quite unstable. Therefore, either the infected cell extracts or recombinant proteins purified by alternative tech- niques will be more efficacious as an AHSV subunit vaccine. To confirm these data, an additional vaccination study was undertaken involving infected cell extracts containing VP2, VP5 and VP7 in combination. The reproducibility of the results was high, similar neutrali- zation titres as before were obtained and, as expected, all the horses were fully protected against the challenge.
Two important aspects should be considered in the control and eradication of AHSV in a non-endemic area. One is protection against the disease. The other is to prevent the spread of the virus by its vectors, in this case Culicoides (gnat). In BTV, 104 infectious particles per ml of blood are necessary to recover at least one virus per biting (Mellor, 1990). Assuming similar conditions for AHSV, the vaccine described here would not only protect the animals, but would drastically reduce the possibility that a vector became infected and spread the disease after biting a vaccinated horse. In our studies, viraemia in vaccinated horses was not detectable by conventional methods.
In summary, the crude cell extracts containing the three structural proteins VP2, VP5 and VP7 was sufficient to obtain a complete protective immune response. Although further experiments need to be performed to estimate the duration of the immunity induced by these antigens, the data clearly indicate the effectiveness of this candidate vaccine, which also has the potential to be produced cost-effectively.
J, L. M.T. is a recipient of a fellowship of the Spanish Ministry of Education and Science. We thank Dr A. Sanz for supplying the MAbs used in this study. This work was partially supported by INIA grant SC93-158.
References
BREMER, C. W. (1976). A gel electrophoretic study of the protein and nucleic acid components of African horsesickness virus. Onder- stepoort Journal of Veterinary Research 43, 193-200.
BREMER, C.W., HUISMANS, H. & VAN DIJK, A.A. (1990). Charac- terization and cloning of the African horsesickness virus genome. Journal of General Virology 71, 793-799.
BURRAGE, T. G., TREVEJO, R., STONE-MARSCHAT, M. & LAEGREID, W. W. (1993). Neutralizing epitopes of African horsesickness virus serotype 4 are located on VP2. Virology 196, 79%803.
CHUMA, T,, LE BLOIS, H., SANCHEZ-VIZCAiNO, J. M., D/AZ-LAVIADA, M. & ROY, P. (1992). Expression of the major core antigen VP7 of African horsesickness virus by a recombinant baculovirus and its use as a group-specific diagnostic reagent. Journal of General Virology 73, 925-931.
DUBOURGET, P. H., PREAUD, J. M., DETRAZ, N., LACOSTE, F., FABRY, A.C., ERASMUS, B. & LOMBARD, M. (1992). Development, pro-
duction and quality control of an industrial inactivated vaccine against AHSV type IV. In Second b,ternational Symposium on Bluetongue, African Horse Sickness and Related Orbiviruses, pp. 874-886. Edited by T. E. Walton and B. I. Osburn. Boca Raton: CRC Press.
ERASMUS, B. T. (1978). A new approach to polyvalent immunization against African horsesickness. In Proceeding of the Fourth lu- ternational Conference on Equine Infectious Diseases, pp. 401-403. Edited by J.T. Bryans and H. Gerber. Princeton: Veterinary Publications.
FRENCH, T.J., MARSHALL, J. J. A. & ROY, P. (1990). Assembly of double-shelled, virus-like particles of bluetongue virus by the simultaneous expression of four structural proteins. Journal of Virology 64, 5695-5700.
HOLMES, I .H. (1991). Family Reoviridae. In Classification and No- menclature of Viruses. Fifth Report of the International Committee on Taxonomy of Viruses, pp. 186-199. Edited by R. I. B. Francki, C. M. Fauquet, D.L. Knudson and F. Brown. Wein and New York: Springer-Verlag.
HOUSE, J. A., LOMBARD, M., DUBOURGET, P. HOUSE, C. & MEBUS, C. A. (1994). Further studies on the efficacy of an inactivated African horse sickness serotype 4 vaccine. Vaccine 12, 142 144.
INUMARU, S. & ROY, P. (1989). Development of bluetongue virus vaccine using a baculovirus expression vector. In Biotechnology for Livestock and Health, pp. 347-357. New York: Plenum Press.
LAVIADA, M.D., ROY, P. & S.~NCHEZ-VIzCAINO, J .M. (1992a). Adaptation and evaluation of an indirect ELISA and imnmno- blotting test for African horsesickness antibody detection. In Second International Symposium on Bhwtongue, African Horse Sickness and Related Orbiviruses, pp. 646-650. Edited by T. E. Walton and B. I. Osburn. Boca Raton: CRC Press.
LAVIADA, M.D., BABIN, M., DOMINGUEZ, J. & SANCHEZ-VIZCAiNO, J. M. (1992b). Detection of African horsesickness virus in infected spleens by a sandwich ELISA using two monoclonal antibodies specific for VP7. Journal of Virological Methods 38, 229-242.
LAVIADA, M. D., RoY, P., SANCHEZ-VIZCAfNO, J .M. & CASAL J. I. (1995). The use of African horsesickness virus NS3 protein, expressed in bacteria, as a marker to differentiate infected from vaccinated horses. Virus Research 38, 205-218.
LUBROTH, J. (1988). African horsesickness and the epizootic in Spain 1987. Equine Practice 10, 26-33.
MARTiNEZ-TORRECUADRADA, J. L., IWATA, H., VENTEO, A., CASAL, J. I. & Roy, P. (1994). Expression and characterization of the two outer capsid proteins of African horsesickness virus : the role of VP2 in virus neutralization. Virology 202, 348-359.
MARTiNEZ-TORRECUADRADA, J. L. & CASAL, J. L (1995). Identification of a linear neutralization domain in the protein VP2 of African horsesickness virus. Virology 210, 391-399.
MELLOR, P. S. (1990). The replication of bluetongue virus in Culicoides vector. Current Topics in Microbiology and Immunology 162, 143-161,
MELLOR, P. (1993). African horsesickness transmission and epidemi- ology. Veterinary Research 24, 199 212.
MERTENS, P. P. C., BURROUGHS, J. N. & ANDERSON, J. (1987). Purifi- cation and properties of virus particles, infections subviral particles and cores of bluetongue virus serotypes 1 and 4. Virology 157, 375-386.
OELLERMANN, R. A. (1970). Plaque formation by African horsesickness virus and characterization of its RNA. Onderstepoort Journal of Veterinary Research 37, 137 144.
OELLERMANN, R.A., ELS, H.J. ~¢ ERASMUS, B.J. (1970). Charac- terization of African horsesickness virus. Archiv fiir die gesamte F\irusforsehung 29, 163-174.
OSBURN, B.I. (1992). Immune responses to orbiviruses. In Second btternational Symposium on Bluetongue, African Horse Sickness and Related Orbiviruses, pp. 511-524. Edited by T. E. Walton and B. I. Osburn. Boca Raton: CRC Press.
RANZ, A. I., MIGUET, J. G., ANAYA, C., VENTEO, A., CORTI~S, E., VELA, C. & SANZ, A. (1992). Diagnostic methods for African horsesickness virus using monoclonal antibodies to structural and nonstructural proteins. Veterinary Microbiology 33, 143-153.

Protection against A H S V in horses 1221
RODRfGUEZ, M., HOOGHUIS, H. & CASTAI~O, M. (1992). African horsesickness in Spain. Veterinary Microbiology 33, 129-142.
ROY, P , URAKAWA, T., VAN DIJK, A.A. & ERASMUS, B.J. (1990). Recombinant virus vaccine for bluetongue disease in sheep. Journal of Virology 64, 1998 2003.
RoY, P., FRENCH, T. J. & ERASMUS, B. J. (1992). Protective efficacy of virus-like particles for bluetongue disease. Vaccine 10, 28-32.
Roy, P., MERTENS, P. P. C. & CASAL, J. 1. (1994a). African horse- sickness virus structure. Comparative Immunology and Microbiology of lnfectious Diseases 17, 243-273.
RoY, P., BISHOP, D. H. L., LEBLOIS, H. & ERASMUS, B.J. (1994b). Long-lasting protection of sheep against bluetongue challenge after vaccination with virus-like particles: evidence for homologous and partial heterologous protection. Vaccine 12, 805-811.
YANG, Y.-Y., JOHNSON, T. M., MECHAM, J. O., TAM, J. P. & LI, J. K.- K. (1992). Epitopic mapping of linear and conformation-dependent antigenic determinants on gp5 of five US bluetongue viruses. Virology 188, 530-536.
(Received 12 December 1995; Accepted 12 Februa O, 1996)
Related Documents











