1 This is an author-created version of the following manuscript: 1 2 Zimmer, H., Auld, T., Hughes, L., Offord, C. and Baker, P. (2015) Fuel flammability and 3 fire responses of juvenile canopy species in a temperate rainforest ecosystem. 4 International Journal of Wildland Fire 24, 349-360. 5 6 The published version is available online at http://www.publish.csiro.au/paper/WF14054.htm 7 8 9 Fuel flammability and fire responses of juvenile canopy species in a 10 temperate rainforest ecosystem 11 12 Running head: Flammable fuels and fire responses in rainforest 13 14 Heidi C. Zimmer A,E , Tony D. Auld B , Lesley Hughes C , Catherine A. Offord D and Patrick J. 15 Baker A 16 17 A Department of Forest and Ecosystem Science, University of Melbourne, 500 Yarra Blvd, 18 Richmond, Victoria 3121, Australia 19 B Office of Environment and Heritage NSW, P.O. Box 1967, Hurstville, NSW 2220, Australia 20 C Department of Biological Sciences, Macquarie University, North Ryde, NSW 2109, 21 Australia 22 D The Royal Botanic Gardens and Domain Trust, The Australian Botanic Garden, Mount 23 Annan, NSW 2567, Australia 24 E Corresponding author: [email protected] 25 26 27 28 Additional keywords: Australia, Blue Mountains, bushfire, Doryphora sassafras, 29 experimental burn, litter, resprouting, Syzygium smithii, Wollemia nobilis. 30 31 32 33 34

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
This is an author-created version of the following manuscript: 1
2
Zimmer, H., Auld, T., Hughes, L., Offord, C. and Baker, P. (2015) Fuel flammability and 3
fire responses of juvenile canopy species in a temperate rainforest ecosystem. 4
International Journal of Wildland Fire 24, 349-360. 5
6
The published version is available online at http://www.publish.csiro.au/paper/WF14054.htm 7
8
9
Fuel flammability and fire responses of juvenile canopy species in a 10
temperate rainforest ecosystem 11
12
Running head: Flammable fuels and fire responses in rainforest 13
14
Heidi C. ZimmerA,E, Tony D. AuldB, Lesley HughesC, Catherine A. OffordD and Patrick J. 15
BakerA 16
17 ADepartment of Forest and Ecosystem Science, University of Melbourne, 500 Yarra Blvd, 18
Richmond, Victoria 3121, Australia 19 BOffice of Environment and Heritage NSW, P.O. Box 1967, Hurstville, NSW 2220, Australia 20 CDepartment of Biological Sciences, Macquarie University, North Ryde, NSW 2109, 21
Australia 22 DThe Royal Botanic Gardens and Domain Trust, The Australian Botanic Garden, Mount 23
Annan, NSW 2567, Australia 24 E Corresponding author: [email protected] 25
26
27
28
Additional keywords: Australia, Blue Mountains, bushfire, Doryphora sassafras, 29
experimental burn, litter, resprouting, Syzygium smithii, Wollemia nobilis. 30
31 32
33
34

2
Abstract 35 36
Historically, rainforests have been considered vulnerable to fire. Recent research, however, 37
has shown that many rainforest species can survive fire by resisting burning and by 38
resprouting and seeding post-fire. We investigated the response of a warm temperate 39
rainforest community to fire by burning juveniles of the dominant canopy tree species 40
(Doryphora sassafras, Syzygium smithii and Wollemia nobilis) and examining litter 41
flammability in a controlled environment. The three species resprouted after the experimental 42
burn, predominately from buds on the stem that were below the soil surface. Higher fire 43
temperatures resulted in reduced overall plant height and resprouting from buds lower on the 44
stem. Increasing proportions of W. nobilis litter generated fires with higher intensities and 45
fuel consumption compared with rainforest angiosperm litter. Moreover, fuel moisture 46
content decreased with increasing W. nobilis litter proportions. Higher litter flammability 47
may result in increased likelihood of fire ignition and fire severity near W. nobilis trees, 48
which would negatively impact the juveniles of all three rainforest species. Alternatively, 49
lower-severity fires (e.g. in rainforest angiosperm litter) may favour W. nobilis juveniles over 50
its rainforest angiosperm competitors 51
52
53

3
Introduction 54
55
The distribution of many of the world’s vegetation communities is influenced by specific fire 56
regimes (Bond and Keeley 2005; Bond et al. 2005), and their interactions with vegetation 57
composition and structure (D’Antonio and Vitousek 1992; Bond and Midgley 1995; Brooks 58
et al. 2004). Fire adaptation varies among plant species with key functional traits conferring 59
survival in fire-controlled ecosystems including the ability to resprout and fire-triggered 60
seedling recruitment (Bond and Keeley 2005). Vegetation communities and their associated 61
fuel characteristics vary widely, and interspecific variation in flammability and contribution 62
to fuel load is substantial (Philpot 1977; van Wilgen et al. 1990; Fonda 2001). 63
64
Until recently, it was generally accepted that rainforests are highly sensitive to fire (e.g. 65
Jackson 1968; Ash 1988; Bowman 2000 cf. Baker et al. 2012). Rainforest composition and 66
structure varies with climate and edaphics (Webb 1968). In areas of lower productivity, or 67
high fire activity, rainforests are limited to areas topographically protected from fire, such as 68
gullies and valleys, or areas adjacent to water bodies, barren or rocky areas (Webb 1968; 69
Bond and van Wilgen 1996, Bowman 2000; Bond et al. 2005; Wood and Bowman 2011). 70
Rainforest species typically require longer to mature compared with shrub and woodland 71
species (Noble and Slatyer 1980) and are tolerant of competition and shade during 72
establishment (Noble and Slatyer 1980). They do not have recruitment strategies reliant on 73
fire, such as serotiny (Lamont et al. 1991), or soil-stored seeds with germination promoted by 74
heat and/or smoke (Keeley and Fotheringham 2000). Several studies, however, describe 75
responses of rainforest species to fire, such as post-fire resprouting (Ashton and Frankenberg 76
1976; Hill and Read 1984; Bowman 1991; Melick and Ashton 1991; Benson and McDougall 77
1998; Williams 2000; Prior et al. 2007; Clarke et al. 2009; Pappas 2010; Williams et al. 78
2012) and germination in fire-generated gaps (Hill and Read 1984; Williams 2000; Jennings 79
and Neyland 2012). The fire-resistant properties of rainforest have also been highlighted 80
(Pappas 2010, Baker et al. 2012; Clarke et al. 2014). Fire resistance is attributed to the moist, 81
shaded, fire-inhibiting microclimates that rainforests promote (Keith 2012) and their position 82
lower in the landscape (i.e. fires burning downhill are generally of lower intensity; Noble et 83
al. 1980). Part of the problem in understanding the relationship between rainforest and fire 84
has been language (Hill 2000). The descriptor ‘fire-sensitive’ is an oversimplification of 85
species responses to complex fire regimes. Fire regimes are a product of frequency, intensity 86
and season (Gill 1975), and individual fires vary at the microsite scale (Bradstock and Auld 87

4
1995; Scarff and Westoby 2006). Rather than being fire-sensitive (i.e. killed by fire), many 88
rainforest species have strategies to cope with certain fire regimes. 89
90
Fire survival and post-fire resprouting have been described for several Australian rainforest 91
species (e.g. Nothofagus cunninghamii, Syzigium smithii, Tristaniopsis laurina; Stocker 92
1981; Hill and Read 1984; Melick and Ashton 1991; Williams 2000; Pappas 2010; Williams 93
et al. 2012). In general, resprouting can be triggered by damage from fire, but also from 94
flooding, herbivory, landslides, logging and windstorms (Del Tredici 2001; Bond and 95
Midgeley 2003), although resprouting can also occur in undamaged plants (Johnstone and 96
Lacey 1983; Midgley and Cowling 1993; Wooley et al. 2008). To resprout, plants require 97
stored resources and undamaged meristems (Bond and van Wilgen 1996; Vesk and Westoby 98
2004b, Clark et al. 2013). 99
100
In trees, resprouting typically takes one of two forms: basal (including underground 101
lignotubers and lower stem collar sprouts; Del Tredici 2001) or epicormic (Burrows 2008). 102
After a severe crown fire, basal sprouting is more common than epicormic sprouting 103
(Burrows 2008). Epicormic resprouting requires protection of vascular cambium and 104
meristems from high temperatures (Burrows 2008, Clarke et al. 2013). Consequently, 105
resprouting occurs higher on the stem when disturbance is less severe (Bellingham and 106
Sparrow 2000). Resprouting ability can also vary with life history stage. Some species can 107
resprout as seedlings and saplings, but not as mature trees (Del Tredici 2001), such as 108
savannah trees that outgrow the risk of mortality from grass fires (Williams et al. 1999). In 109
other species, resprouting ability increases with size (Hodgkinson 1998; Bond and Midgley 110
2001), due to increased resources (Bellingham and Sparrow 2000). Bark thickness, which 111
protects meristems and vascular cambium, typically increases with size and age (Pinard and 112
Huffmann 1997; Baker and Bunyavejchewin 2002; Lawes et al. 2011). However, many 113
rainforest species are thin barked (Uhl and Kauffman 1990; Hoffman et al. 2003), and bark 114
thickness is not always a good indicator of fire response (Lawes et al. 2014). 115
116
Surface and near-surface fuels, typically dead leaf material, are fundamental for fire spread in 117
most Australian vegetation types (Catchpole 2002). Although little work has been done on 118
rainforest fuels in Australia (but see Hill 1982; Dickinson and Kirkpatrick 1985), a review of 119
fire in tropical rainforests showed fires frequently occur in surface fuels, particularly litter 120
(Cochrane 2003). These conclusions are supported by a litter addition experiment in the 121

5
Amazon that found fire spread and intensity were determined by fine (leaf litter) fuels (Balch 122
et al. 2008). Fuel loads in rainforest are, however, typically less than those of sclerophyll 123
forest (Barker 1992; Leonard et al. 2014). 124
125
The Sydney Basin region of south-eastern Australia is prone to experiencing large fires 126
(Cunningham 1984; Gill and Moore 1996; Bradstock 2010), yet patches of warm temperate 127
rainforest are common (Keith 2004). The sandstone landscapes in the Sydney Basin are 128
characterised by broad expanses of predominantly dry sclerophyll vegetation communities on 129
plateaus and ridges, with warm temperate rainforest in protected southerly aspects and deep 130
gullies (Keith 2004). Important changes in vegetation, topography and fuel occur over 131
relatively fine scales (tens to hundreds of metres; Hammill and Bradstock 2006). While 132
weather is the most important determinant of fire behaviour, differences in fuel (e.g. 133
structure, composition, moisture) constitute a key source of small-scale variability in fire 134
behaviour (Chafer et al. 2004; Hammill and Bradstock 2006; Bradstock et al. 2009; 135
Bradstock et al. 2010). The impact of fire on individual plants is dependent upon the heat and 136
duration of the fire, which itself is controlled by the amount of fine fuel consumed (Bradstock 137
and Auld 1995; Scarff and Westoby 2006). We aimed to assess the effect of fire temperature 138
on species from a Sydney Basin warm temperate rainforest community that includes the high 139
conservation value, rare species Wollemia nobilis (Benson and Allen 2007). We also aimed to 140
assess the role of litter composition on surface fire behaviour. This community was selected 141
in particular because the relationship between W. nobilis and fire was unknown (NSW 142
Department of Conservation 2006) but important for conservation management. We asked: 143
1. How does fire temperature influence the response of juvenile rainforest trees post-144
fire? 145
2. How does rainforest litter composition influence fire behaviour (i.e. fuel consumption, 146
flame height, temperature)? 147
3. What are the implications of local-scale heterogeneity in litter flammability and 148
interspecific variation in resprouting ability for forest dynamics? 149
150
Materials and methods 151
152
Effect of fire on juvenile rainforest trees 153
154

6
Twenty-eight individuals each of Doryphora sassafras Endl. (Arthrospermaceae), Syzygium 155
smithii Neid. (Myrtaceae) and Wollemia nobilis Jones et al. (Araucariaceae) were used to 156
examine the impact of fire on plant survival and resprouting. The D. sassafras and S. smithii 157
were grown from seed and were 6 months old. Wollemia nobilis were grown from cuttings 158
because of the extremely low availability of seed, due to the status of W. nobilis as a 159
threatened species with <100 mature (seed-producing) trees in the wild. The W. nobilis 160
juveniles had been propagated one year previously. Plants of all species were grown in 160 161
mm diameter pots and were on average 182 mm tall (± 7 mm [1 SD]) and had multiple 162
branches along their stems. All plants were well watered. 163
164
To conduct burning trials, 1.2 m tall windbreaks were established around a 2 m × 2 m gravel 165
area at the University of Melbourne’s Burnley Campus (37.823 °S, 144.998 °E). Within the 166
windbreaks, a circular area of 1 m2 was established with moistened filtered sand as a base 167
material. Plants were burned in groups of three, one individual of each species. Once placed 168
in the fire area, plants were partially covered with sand such that the base of each plant was 169
protected to a height of 40 mm along the stem. The relative position of the species within the 170
enclosure was randomised for each fire. Thermocouples (Pyrosales, Australia) were placed at 171
(1) the base of each plant and (2) beside the main stem at 50 mm above where it protruded 172
from the sand. Dry straw with a moisture content of 3–4% was used as fuel. Five different 173
fuel loads were used: 200 g, 400 g, 600 g, 800 g and 1000 g (equivalent to 2–10 t/ha). Fuel 174
was loosely and evenly spread over the circular (1 m2) area. 175
176
Fires were replicated five times for each fuel load, each fire with one plant of each species 177
(i.e. three plants). Fires were lit from one edge and allowed to burn across the area until they 178
naturally extinguished. Temperature data from the thermocouples were recorded at 5-second 179
intervals using DeltaLog 9 (DeltaOhm, Netherlands). Flame height was estimated visually 180
using a metal stake marked at 10 cm intervals at the centre of the fire. Weather conditions 181
during the burns, conducted in spring, were fine and calm: winds were at 5–12 km/h, mean 182
temperature was 18°C and mean humidity was 51%. Three individuals each of D. sassafras 183
and S. smithii, and two W. nobilis, were kept unburnt as controls; these plants were 184
transported to the burn site then returned to the nursery. These plants were not buried. After 185
the experimental fires, plants were retrieved and placed in the nursery where they were 186
watered daily. The first sprouts appeared four weeks after the experimental burns. At nine 187

7
weeks we assessed whether plants had resprouted, and if they had we measured (a) post-fire 188
live plant height (the total height of post-fire plant growth); (b) the height on the stem at 189
which plants resprouted; (c) resprout length (the length of new post-fire sprouts); and (d) total 190
number of resprouting buds per plant. Plant height and resprout length are indicative of plant 191
resource storage and the ability to mobilise resources after damage, while the height of 192
resprouting buds and the total number of buds are indicative of plant morphology (i.e. 193
meristem locations) and damage to and protection of buds during fire. 194
195
Effect of rainforest litter composition on fire characteristics 196
The vegetation of the field location of W. nobilis is warm temperate rainforest dominated by 197
Ceratopetalum apetalum, D. sassafras and S. smithii with an understorey of ferns and vines 198
(Benson and Allen 2007). At Site 1 (refer Benson and Allen 2007 for the specific vegetation 199
characteristics of the W. nobilis sites), 26 litter samples were collected from the areas 200
surrounding W. nobilis individuals. These 26 samples varied in the relative proportion of W. 201
nobilis litter and rainforest angiosperm litter, with seven samples of pure W. nobilis branches 202
from the base of the W. nobilis trunks, 13 of mixed litter (W. nobilis branches and rainforest 203
litter) collected from 5 m and 10 m away from the W. nobilis trunks, and six rainforest 204
angiosperm litter samples taken >20 m away from the W. nobilis grove. All samples were 205
collected using a 25 cm × 25 cm quadrat. Samples were placed in sealed plastic bags and 206
transported to Macquarie University in Sydney. The wet weights of all samples were 207
measured, then the samples were dried at 70°C until constant mass. The difference between 208
wet and dry weight was used to calculate fuel moisture content. The number and volume of 209
litter samples collected was limited by both conservation considerations (minimising the level 210
of disturbance near W. nobilis) and the difficulties of site access. 211
212
For the experimental burning of litter, a circular plot, 0.25 m2 in area, with a rim 60 mm high 213
and raised 50 mm from the ground was constructed after Plucinski (2003) and built from 214
perforated steel. A solid steel plate, the same area as the plot and with five small holes to 215
expose thermocouples was also constructed. The solid plate was then placed within the plot 216
to cover the perforated steel base and capture ash while allowing for the exposure of the 217
thermocouples. The five thermocouples were equidistant from the centre of the plate. The 218
maximum temperature that could be recorded by the thermocouples was 500° C. A cotton 219
ball soaked with 1 ml of methylated spirits, placed in the centre of the arena, was the 220
standardised ignition source. Experimental burns were conducted in a large shed in order to 221

8
protect the burns from wind and provide standardised light, wind and temperature conditions. 222
Experimental burns were carried out on oven-dry litter subsamples of 100 g from angiosperm 223
rainforest (n = 6), W. nobilis (n = 8), and mixed rainforest and W. nobilis (n = 13; ranging 224
from 5–93% W. nobilis). This allowed direct comparison of flammability among litter types. 225
During each burn we measured (a) maximum flame height (an indicator of fire intensity in 226
homogenous fuels; Keeley 2009; Sullivan et al. 2012), visually recorded every 30 seconds for 227
the duration of the burn using a metal bar with 5 cm increments placed at the back of the 228
plate; (b) maximum temperature, recorded every second using the thermocouples and data 229
logger and (c) mass of litter consumed by fire, equal to the difference in weight of each 230
sample before and after burning. 231
232
Statistical analysis 233
234
For the experimental burning of juvenile rainforest trees we used maximum temperature 235
(measured at the plant stem) as our primary explanatory variable. Maximum temperature was 236
selected because of its objectivity and because it was correlated both with duration of burn 237
above 60°C (the temperature likely to cause leaf damage; Helgerson 1990; Cunningham and 238
Read 2006; van Wagner 1973; Figure 1) and flame height (Figure S1). To assess the 239
influence of fire on the juvenile trees we related maximum fire temperature to post-fire live 240
plant height, and the height, length and number of resprouting stems. To assess the influence 241
of pre-fire plant characteristics on fire response, we related pre-fire stem diameter to these 242
same post-fire plant characteristics. Initially, we fitted linear models to the data. If a linear 243
model did not provide a good fit then models of increasing complexity were fitted, based on 244
examination of residual plots. In some cases it was necessary to fit different functional forms 245
for the different species. Akaike’s Information Criterion for small samples (AICc; Hurvich 246
and Tsai 1989) was used to select the best model. If the linear model was non-significant, and 247
there was no pattern in the residuals, then the (non-significant) parameter estimates from the 248
linear model were reported. 249
250
For the experimental burning of litter we determined the per cent weight of litter consumed 251
during the fire. The relationships between litter composition and flame height, per cent 252
weight of litter consumed and fuel moisture content were examined using linear regression. 253
All statistical analyses were conducted in R Version 3.03 (R Core Team 2013). 254

9
255
Results 256
257
Effect of fire on juvenile rainforest trees 258
The maximum temperatures of the experimental burns, measured at 50 mm on the plant stem, 259
ranged from 32°C to 606°C. The duration of burning at temperatures likely to damage living 260
tissues (>60°C; Helgerson 1990; Cunningham and Read 2006; van Wagner 1973) ranged 261
from 0 to 245 seconds. All plants exposed to experimental burns resprouted (Figure 2), 262
except for one individual W. nobilis plant. The W. nobilis that did not resprout was burnt at a 263
maximum temperature of 508°C. (The highest temperature experienced by any W. nobilis 264
was 538 °C.) The majority of resprouts were from buds below the sand. 265
266
Post-fire live plant height, measured from where stem was covered with sand during 267
experimental burn to the highest living part of the plant, decreased with increasing maximum 268
fire temperature in all three species (Figure 3a; W. nobilis R2 = 0.48; D. sassafras R2 = 0.14, 269
P = 0.04; S. smithii R2 = 0.22, P = 0.01). Mean post-fire live plant height was similar among 270
species: 23 (± 16) mm, S. smithii; 8 (± 16) mm, D. sassafras; 32 (± 43) mm, W. nobilis. The 271
relationship between W. nobilis post-fire live plant height and maximum temperature 272
represented a different functional form (exponential) to D. sassafras and S. smithii (linear), 273
influenced by several W. nobilis with taller post-fire heights, resulting from lower maximum 274
temperature burns. The relationship between post-fire live height and maximum temperature 275
was similar for D. sassafras and S. smithii, and this was reflected in their parameter estimates 276
and overlapping confidence intervals (Table 1). 277
278
The height on the stem where the highest post-fire resprouting bud occurred was also 279
negatively correlated with maximum temperature in S. smithii and W. nobilis (Figure 3b; W. 280
nobilis R2 = 0.36; S. smithii R2 = 0.35, P = 0.001), but not for D. sassafras (P = 0.67). 281
Relatively low maximum fire temperatures allowed two W. nobilis individuals to sprout from 282
high on the stem (≥ 90 mm), contributing to the relationship between W. nobilis post-fire 283
maximum sprout height and temperature taking a different functional form (exponential) to S. 284
smithii (linear). These W. nobilis were very lightly burnt. It must be noted that the mean 285
maximum post-fire bud height in W. nobilis at lower temperatures (46 mm (±46) at <200 °C) 286
was only 27 mm more than that of S. smithii (19 (±11) mm)). 287

10
288
The maximum length of (post-fire) resprouts on W. nobilis was also higher where maximum 289
temperature was lower (Figure 3c; W. nobilis, R2 = 0.28). There was no correlation between 290
maximum temperature and post-fire resprout length for D. sassafras (P = 0.29). Syzygium 291
smithii showed a weak (P = 0.20) trend for increasing post-fire resprout length with 292
maximum temperature. The longest five post-fire resprouts belonged to W. nobilis. Four of 293
these five plants with the longest resprouts had buds that were obviously present before, and 294
had survived, the fire. 295
296
The total number of buds that resprouted post-fire was weakly negatively correlated with 297
maximum temperature in S. smithii and W. nobilis (Figure 3d; S. smithii R2 = 0.09, P = 0.08; 298
W. nobilis R2 = 0.10, P = 0.06). However, this was not the case for D. sassafras (P = 0.94). 299
300
Post-fire live plant height and pre-fire stem diameter were not related in W. nobilis (P = 301
0.44). In contrast, there was a weak trend for increasing post-fire plant height with pre-fire 302
stem diameter with D. sassafras (R2 = 0.11, P = 0.06) and S. smithii (Figure 4a; Table 2; R2 = 303
0.12, P = 0.05). In contrast, the total number of post-fire resprouting buds increased with pre-304
fire stem diameter in W. nobilis (R2 = 0.26, P = 0.005) and to a lesser extent in D. sassafras 305
(R2 = 0.07, P = 0.09), but not in S. smithii (Figure 4b; P = 0.65). This relationship in W. 306
nobilis was particularly influenced by one plant (with the largest diameter, 18 mm). This 307
plant also influenced the relationship between W. nobilis post-fire maximum post-fire 308
resprout length and pre-fire stem diameter (P = 0.03). There were no relationships between 309
pre-fire stem diameter and post-fire maximum resprout length in the other species (D. 310
sassafras, P = 0.58; S. smithii, P = 0.08). No species showed significant relationships 311
between pre-fire diameter and post-fire maximum sprout height (D. sassafras, P = 0.65; S. 312
smithii, P = 0.30; W. nobilis, P = 0.85) 313
314
The control plants also sprouted, in line with the timing of the study in the spring (i.e. 315
growing season). Sprouts occurred along the length of the stem in S. smithii, the mean 316
number of sprouts was 7 (±3) and the mean maximum length of post-fire resprouts was 30 317
(±9) mm. Sprouts also occurred along the length of the stem in D. sassafras, the mean 318
number sprouts was 6 (±3) and the maximum resprout length was 35 mm, other plants had 319
maximums of 1 mm. One of the W. nobilis control plants grew three new sprouts from its top; 320
maximum new resprout length was 125 mm. 321

11
322
Effect of rainforest litter composition on fire characteristics 323
Per cent weight of litter consumed during fire increased with the proportion of W. nobilis 324
litter in samples (Figure 5a; Table 3; R2 = 0.87, P < 0.001). Flame heights (up to a maximum 325
of 80 cm) also increased with increasing proportions of W. nobilis litter (R2 = 0.80, P < 326
0.001, Figure 5b). Logger data showed that 16 fires (of 26) were still increasing in 327
temperature when they reached the maximum measurable temperature (500° C). Notably, all 328
except one of the 16 fires that reached temperatures ≥500°C included some W. nobilis litter, 329
while the maximum temperature of pure angiosperm litter fires was 392° C; Figure S2). Fuel 330
moisture content (before oven drying) showed increasing proportions of W. nobilis litter was 331
also associated with decreasing fuel moisture (Figure 5c; R2 = 0.21, P = 0.002). 332
333
Discussion 334
335
Seedlings and juveniles of shade-tolerant rainforest trees are vulnerable to surface fires 336
because of their small size, thin bark and the long time they spend in the understorey. In our 337
study, immature individuals of all three rainforest species resprouted after experimental fires, 338
indicating that these species can survive, and recover from fire. In all species, the majority of 339
resprouting occurred from the base of the plant. Resprouting occurred higher on the stem, 340
post-fire live plant height was taller and resprout length was longer in W. nobilis after lower 341
temperature fires. However, per cent of litter consumed, flame height and temperature 342
increased, and fuel moisture content decreased, with increasing proportions of W. nobilis 343
litter. As a consequence, the probability of ignition, given an ignition source, may be 344
increased in W. nobilis litter relative to rainforest angiosperm litter. Increased fire 345
temperature and fuel consumption may also influence fire spread, via pre-heating of fuels. 346
347
Resprouting in immature D. sassafras, S. smithii and W. nobilis occurred after exposure to a 348
range of fire temperatures, from 32 to 606°C. Chandler (1983) describes fires with surface 349
temperatures from 500 to 750°C as having severe effects. The maximum temperature in our 350
experiment was somewhat lower than that reported by Hobbs and Atkins (1988), who 351
described maximum surface and litter fire temperatures up to 800°C in southwestern 352
Australian woodland fires. Alternately, the maximum temperatures in our study are 353
comparable to the maximum temperature (650°C) recorded in Pinus pinaster litter fires 354

12
(Valette et al. 1994). The majority of resprouts, in all species in our study, occurred on the 355
lower stem below where they were covered by sand during the experimental burn. The degree 356
of soil heating during fire decreases rapidly with increasing soil depth (Auld and Bradstock 357
1996; Vesk et al. 2004). It appears that the sand insulated dormant buds from high 358
temperatures, countering the lack of protection by bark in these small, thin-barked rainforest 359
species. The insulating properties of soil account for observations that basal resprouting is 360
more common after severe fires, compared with elevated, epicormic sprouting (Burrows 361
2008). Resprouting could also be a response to the effects of burial in sand (abrasion and 362
low-level heat transfer); effects of burial cannot be separated from effects of burning here. 363
Further, it must be acknowledged that plant responses to fire vary not only in response to fire 364
characteristics, but also according to their physiological status (e.g. water stress), which is 365
influenced by climate, which in turn influences the likelihood of fire occurrence. The plants 366
in this study were well watered and the conditions under which the burns occurred were mild, 367
hence the post-fire responses of these species to intense fire in dry conditions in the wild may 368
be weaker than those observed here. 369
370
Post-fire resprouting of mature S. smithii has been demonstrated in previous studies (Ashton 371
and Frankenberg 1976, Melick and Ashton 1991), as has the capacity of mature D. sassafras 372
to resprout from root suckers and the stem base (Campbell and Clarke 2006). Syzygium 373
smithii and D. sassafras can also resprout in the absence of disturbance (Johnston and Lacey 374
1983). The presence of resprouting in rainforest species is unsurprising, as vegetative 375
recruitment is beneficial for recovery from injury, including herbivory, and can provide intra-376
specific competitive advantage (Poorter et al. 2010; Tanenzap et al. 2011). This is 377
particularly important to shade-tolerant juveniles with long residence times in the 378
understorey. The capacity D. sassafras, S. smithii and W. nobilis juveniles to resprout after 379
fire was unknown prior to our study. Our study highlights the capacity of D. sassafras and S. 380
smithii species to resprout at the juvenile stage, and after high temperature fire, provided 381
some buds are insulated by the soil. Both D. sassafras and S. smithii experienced topkill (i.e. 382
stem mortality, Hoffman and Solbrig 2003) in our experimental burns, such that resprouting 383
was basal, with a maximum height of resprouting buds of 18 and 30 mm (for D. sassafras 384
and S. smithii, respectively). Nevertheless, post-fire resprouts may be more competitive than 385
newly germinated seedlings, through their access to a well-developed root system and 386
resources not available to seedlings (Vesk and Westoby 2004a; Dietze and Clark 2008). 387
388

13
The ability of W. nobilis to produce multiple stems is well documented (Hill 1997; Burrows 389
et al. 2003). Moreover, the presence of fire scars on mature, live trees in the wild population 390
(Offord et al. 1999; New South Wales Department of Conservation 2006; Benson and Allen 391
2007) suggests that adult plants can survive fire. Our study shows that immature W. nobilis, 392
like other rainforest taxa, have the potential to survive fire by resprouting. While the capacity 393
to resprout is common in angiosperms, it is much less common in gymnosperms, which lack 394
cotyledonary buds and do not develop root collars with the potential to sprout (Del Tredici 395
2001). However, resprouting after injury in the rainforest conifer family Araucariaceae is 396
relatively common (Burrows 1987; Burrows et al. 2003; Alabrace and Dillenberg 2012; also 397
in Southern Hemisphere conifer family Podocarpaceae, Ladd and Enright 2010). Juvenile W. 398
nobilis plants resprouted from higher on the stem, grew taller and had greater maximum 399
resprout length, compared to other rainforest species juveniles, suggesting greater protection 400
of dormant meristems from the lethal heat effects of fire (e.g. through deeper burial of 401
meristem in the stem, or greater insulation of meristem by bark). Increased resprouting height 402
and vigour in juveniles, as demonstrated here, may give W. nobilis a competitive advantage 403
over other rainforest taxa after low severity fires. However, this result may also be attributed 404
to the clonal (cf. seed) production of W. nobilis individuals used in this study. Future studies 405
should be completed on seed-grown W. nobilis when availability of such plants increases. 406
407
Wollemia nobilis litter generated fires that were more intense and consumed more fuel, 408
compared with the litter of the surrounding rainforest. Litter ventilation is a fundamental 409
driver of increased flammability, and well-ventilated litter beds burn more rapidly (Scarff et 410
al. 2006). Large leaves are more likely to create well-ventilated litter beds (Scarff et al. 411
2006), and less dense fuels can sustain burning at higher moisture content compared to dense 412
fuels (Ganteaume et al. 2009). Although W. nobilis does not have large leaves, it abscises 413
entire branches, rather than leaves (Burrows et al. 2006). Wollemia nobilis branches are 414
unbranched, and have broad-based leaves that remain attached to the branch when it is 415
abscised (Burrows et al. 2006), creating more woody, structured and ventilated litter. 416
Moreover, W. nobilis litter decays slowly (Banks 2002), resulting in increased particle sizes, 417
and therefore ventilation. Hence, W. nobilis litter is more likely to sustain burning than 418
rainforest angiosperm litter because of its structure and ventilation. Wollemia nobilis litter is 419
also more likely to ignite than rainforest angiosperm litter because it has lower fuel moisture 420
content (Sullivan et al. 2012). Wollemia nobilis contains a range of volatile compounds (as 421
determined by steam extraction; Brophy et al. 2000). Organic compounds not extractable by 422

14
steam, but potentially contributing to flammability, remain unknown. Nevertheless, the 423
overall leaf oil content of W. nobilis (0.5% fresh weight; Brophy et al. 2000) is small 424
compared with famously flammable Eucalyptus species (up to 3.5% fresh weight; Barker and 425
Smith 1902). Increased flammability can also drive fire propagation, as high burning 426
temperatures can pre-heat surrounding fuels and potentially raise them ignition temperature. 427
428
Fires in pure rainforest litter displayed low flame height (mean flame height was 30 cm 429
versus 71 cm in pure W. nobilis litter), fuel consumption (36% versus 95% in pure W. nobilis 430
litter), and relatively high fuel moisture content (mean 90% versus 31% in pure W. nobilis 431
litter). In general, rainforest experiences less fire than dry and/or sclerophyll forest because 432
its fuel moisture content is higher, and the moisture content required for ignition (of 433
rainforest fuels) is lower (Dickinson and Kirkpatrick 1985). Indeed, extended drought or 434
severe fire weather are required for rainforest to burn (forest fire is unlikely to be sustained if 435
moisture >22%, Sullivan et al. 2012; see also Hill 1982; Pyrke and Marsden-Smedley 2005; 436
Hammill and Bradstock 2009). In our study, the mean fuel moisture content of the pure 437
rainforest angiosperm litter was 81% (±31%). Litter was dried to constant weight before 438
flammability experiments were performed, allowing direct comparison of flammability 439
among litter types. While an even drying of litter in the field is improbable, it is likely that 440
comparison of wet litter flammability would only accentuate the differences reported. 441
Wollemia nobilis litter is so well ventilated that it will always be relatively dry, and therefore 442
flammable, compared with angiosperm litter, which is more closely packed and lays flat on 443
the forest floor (Catchpole 2002; Scarff et al. 2006). 444
445
While rainforest is less flammable than sclerophyll forest and woodland (Pyrke and Marsden 446
Smedley 2005), rainforest edges are more likely to burn than rainforest. Pappas (2010), 447
working in cool temperate rainforest in the Central Highlands of Victoria, found that the 448
mean distance from eucalypt/rainforest ecotone to burnt rainforest patches was only 2.5 m, 449
demonstrating higher probabilities of burning at rainforest edges than in the rainforest 450
interior. Wollemia nobilis occurs at the margins of (and intergrades into) rainforest and dry 451
sclerophyll forest on steep slopes and on ledges above the canyon floor (Department of 452
Conservation 2006; Benson and Allen 2007). Surrounding ridge tops above these locations 453
consist of dry sclerophyll forest and woodlands. The habitat of W. nobilis may therefore be 454
relatively more exposed to fire, in particular fire ignited by burning material falling down 455
slope (Banks 2002), and more likely to burn, compared to surrounding rainforest. This may 456

15
explain observed fire scars on W. nobilis (Offord et al. 1999; New South Wales Department 457
of Conservation 2006; Benson and Allen 2007), not evident on other co-occurring rainforest 458
species. 459
460
What are the implications of heterogeneity in litter flammability within rainforest, combined 461
with variation in resprouting ability among co-occurring tree species? Fire temperatures, 462
flame heights and fuel consumption are likely to be higher with increasing proportions of W. 463
nobilis litter in the ground fuel layer, potentially increasing the probability of ignition and fire 464
spread. Fire may reduce inter- and intra-specific competition with mature W. nobilis, 465
assuming the fire resistance and resilience is relatively higher in mature W. nobilis, compared 466
with its competitors. Few seedlings of any species occur in pure W. nobilis litter (H. Zimmer, 467
unpublished data). While this may be attributed to the relatively small area of pure W. nobilis 468
litter in the rainforest, it may also be attributed to the inhospitable environment created by W. 469
nobilis (e.g., phytotoxicity, Seal et al. 2010; increased fire). Juveniles of W. nobilis have 470
enhanced capacity for survival and growth after low severity fire, compared with co-471
occurring rainforest taxa. As such, low severity fires may provide some limited competitive 472
advantages for immature W. nobilis plants, compared with the immature rainforest 473
angiosperms. 474
475
Conclusion 476
The ability to survive and resprout after injury is critical for species with long residence times 477
in the understorey (Poorter et al. 2010; Tanenzap et al. 2011; e.g. W. nobilis; Zimmer et al. 478
2013). We have shown that juveniles of three rainforests taxa, D. sassafras, S. smithii and W. 479
nobilis, can survive fire when buds in the stem are insulated from lethal temperatures by soil. 480
Notably, several W. nobilis individuals resprouted from higher on the stem, and grew taller 481
with longer resprouts, after lower temperature fire. On the other hand, an increasing 482
proportion of W. nobilis litter resulted in increased flame height, maximum temperature and 483
fuel consumption and lower fuel moisture content. These factors, attributed to the structure 484
and ventilation of W. nobilis litter, may contribute to increased ignitability and fire spread 485
where the proportion of W. nobilis litter is higher (i.e. in the vicinity of mature W. nobilis). 486
The likelihood of exposure of W. nobilis litter to an ignition source may also be increased, 487
relative to the rainforest angiosperm litter, due to the position of the W. nobilis at the edge of 488
the rainforest, adjacent to the more fire-prone dry sclerophyll woodland (NSW Department of 489
Conservation 2006; Benson and Allen 2007). While high severity fire (i.e. in W. nobilis litter) 490

16
is likely to limit resprouting to belowground buds in juveniles all three rainforest trees, W. 491
nobilis juveniles may have some competitive advantage, via resprouting from higher on the 492
stem, after low severity fire (i.e. in angiosperm litter). 493
494
Acknowledgements 495
We thank Tracey Adams for performing the litter burning experiments. We are also grateful 496
to Chris Pavich, Steve Clark, Jaime Plaza, Geoff Burrows and Patricia Meagher for their help 497
and guidance in the field during litter collection. We thank Scott Nichols for assistance with 498
the designing the plant burning experimental set up, and Scott Nichols and Linda Parker for 499
assistance in undertaking the plant burning experiment. We thank Steve Clark for sharing his 500
insights into the relationship between Wollemi pines and fire. Comments from the editor and 501
two anonymous referees greatly improved this manuscript. Heidi Zimmer’s doctoral research 502
is funded by an Australian Postgraduate Award and the Wollemi Pine Recovery Team, which 503
includes the New South Wales Office of Environment and Heritage, New South Wales 504
National Parks and Wildlife Service and the Royal Botanic Gardens and Domain Trust. 505
506

17
References 507
Alabarce F, Dillenburg L (2012) Resprouting ability and intensity after damage in seedlings 508 of the large-seeded species Araucaria angustifolia. Brazilian Journal of Plant Physiology 509 24, 131–140. 510
Ash J (1988) The location and stability of rainforest boundaries in north-eastern Queensland, 511 Australia. Journal of Biogeography 15, 619–630. 512
Ashton DH, Frankenberg J (1976) Ecological studies of Acmena smithii (Poir.) Merrill & 513 Perry with special reference to Wilson's Promontory. Australian Journal of Botany 24, 514 453–487. 515
Auld TD, Bradstock RA (1996) Soil temperatures after the passage of a fire: Do they 516 influence the germination of buried seeds? Austral Ecology 21, 106–109. 517
Baker PJ, Simkin R, Pappas N, McLeod A, McKenzie M (2012) Fire on the mountain: A 518 multi-scale, multi- proxy assessment of the resilience of cool temperate rainforest to fire 519 in Victoria’s Central Highlands. Pp 375-391 in SG Haberle and B David (eds), Peopled 520 landscapes: Archaeological and Biogeographic Approaches to Landscapes. ANU E-press, 521 Canberra, Australia. 522
Baker PJ, Bunyavejchewin S (2006) Bark thickness and the influence of forest fire on tree 523 population structure in a seasonal evergreen tropical forest. Natural History Bulletin of 524 the Siam Society 54, 215–225. 525
Balch JK, Nepstad DC, Brando PM, Curran LM, Portela O, de Carvalho O Jr, Levebvre P 526 (2008) Negative fire feedback in a transitional forest of southeastern Amazonia. Global 527 Change Biology 14, 2276–2287. 528
Banks J (2002) Wollemi pine: tree find of the 20th century. Pp 85-89 in J, Dargavel, D 529 Gaughwin and B Libbis (eds), Australia’s Ever-changing Forests V, Proc Fifth National 530 Conference on Australian Forest History. ANU Centre for Resource and Environmental 531 Studies, Canberra, Australia. 532
Barker P (1992) Fire and the stability of a Victorian cool temperate rainforest. Pp 27–131 in 533 AP Gell, D Mercer (eds)Victoria’s Rainforests : Perspectives on Definition, 534 Classification and Management. Dept. of Geography and Environmental Science, 535 Monash University, Melbourne, Australia. 536
Barker R, Smith H (1902) A research on the eucalypts: especially in regard to their essential 537 oils. Series: Technical education series no 24. Technological Museum, Sydney Australia. 538
Bellingham PJ, Sparrow AD (2000) Resprouting as a life history strategy in woody plant 539 communities. Oikos 89, 409–416. 540
Benson DH, McDougall L (1997) Ecology of Sydney Plant Species: Myrtaceae. 541 Cunninghamia 5, 809–987. 542
Benson J, Allen C (2007) Vegetation associated with Wollemia nobilis (Araucariaceae). 543 Cunninghamia 10, 255–262. 544

18
Bond W, van Wilgen BW (1996) Fire and Plants. Springer. 545
Bond WJ, Keeley JE (2005) Fire as a global ‘herbivore’: the ecology and evolution of 546 flammable ecosystems. Trends in Ecology & Evolution 20, 387–394. 547
Bond WJ, Midgley JJ (1995) Kill thy neighbour: an individualistic argument for the evolution 548 of flammability. Oikos 73, 79–85. 549
Bond WJ, Midgley JJ (2001) Ecology of sprouting in woody plants: the persistence niche. 550 Trends in Ecology & Evolution 16, 45–51. 551
Bond WJ, Midgley JJ (2003) The evolutionary ecology of sprouting in woody plants. 552 International Journal of Plant Sciences 164, S103–S114 553
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a 554 world without fire. New Phytologist 165, 525–538. 555
Bowman DJMS (1991) Recovery of some northern Australian monsoon tree species 556 following fire. Proceedings of the Royal Society of Queensland 101, 21–25. 557
Bowman DJMS (2000) Australian Rainforests. Cambridge University Press, UK. 558
Bradstock R, Auld T (1995) Soil temperatures during experimental bushfires in relation to 559 fire intensity: consequences for legume germination and fire management in south-560 eastern Australia. Journal of Applied Ecology 32, 76–84. 561
Bradstock RA, Hammill KA, Collins L, Price O (2009) Effects of weather, fuel and terrain on 562 fire severity in topographically diverse landscapes of south-eastern Australia. Landscape 563 Ecology 25, 607–619. 564
Bradstock RA, Cohn JS, Gill AM, Bedward M, Lucas C (2009) Prediction of the probability 565 of large fires in the Sydney region of south-eastern Australia using fire weather. 566 International Journal of Wildland Fire 18, 932–943. 567
Bradstock RA (2010) A biogeographic model of fire regimes in Australia: current and future 568 implications. Global Ecology and Biogeography 19, 145–158. 569
Brooks M, D’Antonio CM, Richardson D, Grace J, Keeley JE, Di Tomaso J, Hobbs RJ, 570 Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. BioScience 54, 571 677–688. 572
Brophy JJ, Goldsack RJ, Wu MZ, Fookes CJ, Forster PI (2000) The steam volatile oil of 573 Wollemia novilis and its comparison with other members of the Araucariaceae (Agathis 574 and Araucaria). Biochemical Systematics and Ecology 28, 563–578. 575
Burrows G (1987) Leaf axil anatomy of the Araucariaceae. Australian Journal of Botany 35, 576 631–640. 577
Burrows GE (2008) Syncarpia and Tristaniopsis (Myrtaceae) possess specialised fire-578 resistant epicormic structures. Australian Journal of Botany 56, 254–264. 579
Burrows GE, Meagher PF, Heady RD (2006) An anatomical assessment of branch abscission 580

19
and branch-base hydraulic architecture in the endangered Wollemia nobilis. Annals of 581 Botany 99, 609–623. 582
Burrows GE, Offord CA, Meagher PF, Ashton K (2003) Axillary meristems and the 583 development of epicormic buds in Wollemi pine (Wollemia nobilis). Annals of Botany 584 92, 835–844. 585
Campbell ML, Clarke PJ (2006) Response of montane wet sclerophyll forest understorey 586 species to fire: evidence from high and low intensity fires. Proceedings of the Linnean 587 Society of New South Wales 127, 63–74. 588
Catchpole W (2002) Fire properties and burn patterns in heterogenous landscapes. Pp 49-76 589 in RA Bradstock, J Williams, AM Gill (eds) Flammable Australia’. Cambridge 590 University Press, Cambridge, UK. 591
Chafer CJ, Noonan M, Macnaught E (2004) The post-fire measurement of fire severity and 592 intensity in the Christmas 2001 Sydney wildfires. International Journal of Wildland Fire 593 13, 227–240. 594
Chandler C (1983) Forest Fire Management. Fire in Forestry; Vol 2. Wiley, USA. 595
Clarke PJ, Knox KJE, Campbell M, Copeland L (2009) Post-fire recovery of woody plants in 596 the New England Tableland Bioregion. Cunninghamia 11, 221–239. 597
Clarke PJ, Lawes MJ, Midgley JJ, Lamont BB, Ojeda F, Burrows GE, Enright NJ, Knox KJE 598 (2013) Resprouting as a key functional trait: how buds, protection and resources drive 599 persistence after fire. New Phytologist 197, 19–35. 600
Clarke PJ, Knox KLE, Bradstock RA, Munoz-Robles C, Kumar L (2014) Vegetation, terrain 601 and fire history shape the impact of extreme weather on fire severity and ecosystem 602 response. Journal of Vegetation Science, in press. 603
Cochrane MA (2003) Fire science for rainforests. Nature 421(6926), 913–919. 604
Cunningham CJ (1984) Recurring natural fires hazards: a case study of the Blue Mountains, 605 New South Wales, Australia. Applied Geography 4, 5–27. 606
Cunningham SC, Read J (2006) Foliar temperature tolerance of temperate and tropical 607 evergreen rain forest trees of Australia. Tree Physiology 26, 1435–1443. 608
Del Tredici P (2001) Sprouting in temperate trees: a morphological and ecological review. 609 The Botanical Review 67, 121–140. 610
Dickinson K, Kirkpatrick J (1985) The flammability and energy content of some important 611 plant species and fuel components in the forests of southeastern Tasmania. Journal of 612 Biogeography 12, 121–134. 613
Dietze MC, Clark JS (2008) Changing the gap dynamics paradigm: vegetative regeneration 614 control on forest response to disturbance. Ecological Monographs 78, 331–347. 615
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire 616 cycle, and global change. Annual Review of Ecology and Systematics 23, 63–87. 617

20
Fonda RW (2001) Burning characteristics of needles from eight pine species. Forest Science 618 47, 390–396. 619
Ganteaume A, Lampin-Maillet C, Guijarro M, Hernando C, Jappiot M, Fonturbel T, Pérez-620 Gorostiaga P, Vega JA (2009) Spot fires: fuel bed flammability and capability of 621 firebrands to ignite fuel beds. International Journal of Wildland Fire 18, 951–969. 622
Gill AM (1975) Fire in the Australian flora: A review. Australian Forestry 38, 4–25. 623
Gill AM, Moore P (1996) Regional and historical fire weather patterns pertinent to the 624 January1994 Sydney bushfires. Proceedings of the Linnaean Society of NSW 116, 27–36. 625
Hammill KA, Bradstock RA (2006) Remote sensing of fire severity in the Blue Mountains: 626 influence of vegetation type and inferring fire intensity. International Journal of 627 Wildland Fire 15, 213–226. 628
Hammill KA, Bradstock RA (2009) Spatial patterns of fire behaviour in relation to weather, 629 terrain and vegetation. Proceedings of the Royal Society of Queensland 115, 129–135. 630
Helgerson, OT (1989) Heat damage in tree seedlings and its prevention. New Forests 3, 333–631 358. 632
Hill KD (1997) Architecture of the Wollemi Pine (Wollemia nobilis, Araucariaceae), a unique 633 combination of model and reiteration. Australian Journal of Botany 45, 817–826. 634
Hill RS (1982) Rainforest fire in western Tasmania. Australian Journal of Botany 30, 583–635 589. 636
Hill RS (2000) Attempting to define the impossible: a commentary on ‘Australian 637 Rainforests: islands of green in a land of fire’. Australian Geographical Studies 38, 320–638 326. 639
Hill RS, Read J (1984) Post-fire regeneration of rainforest and mixed forest in western 640 Tasmania. Australian Journal of Botany 32, 481–493. 641
Hobbs RJ, Atkins L (1988) Spatial variability of experimental fires in south‐west Western 642 Australia. Austral Ecology 13, 295–299. 643
Hodgkinson KC (1998) Sprouting success of shrubs after fire: height-dependent relationships 644 for different strategies. Oecologia 115, 64–72. 645
Hoffmann WA, Orthen B, Nascimento PKVD (2003) Comparative fire ecology of tropical 646 savanna and forest trees. Functional Ecology 17, 720–726. 647
Hoffmann WA, Solbrig OT (2003) The role of topkill in the differential response of savanna 648 woody species to fire. Forest Ecology and Management 180, 273–286. 649
Hurvich CM, Tsai CL (1989) Regression and time series model selection in small samples. 650 Biometricka 76, 297–307. 651
Jackson WD (1968) Fire, air, water and earth - an elemental ecology of Tasmania. 652 Proceedings of the Ecological Society of Australia 3, 9–16. 653

21
Jennings B (2014) Seedling regeneration of celery-top pine. Tasforests 19, 1–16. 654
Johnston R, Lacey C (1983) Multi-stemmed trees in rainforest. Australian Journal of Botany 655 31, 189–195. 656
Keeley JE (2009) Fire intensity, fire severity and burn severity: a brief review and suggested 657 usage. International Journal of Wildland Fire 18,11–126. 658
Keeley JE, Fotheringham CJ (2000) Role of fire in regeneration from seed. Pp 311-310 in M 659 Fenner (ed) Seeds: The ecology of regeneration in plant communities. CABI, UK. 660
Keith DA (2012) Functional traits: their roles in understanding and predicting biotic 661 responses to fire regimes from individuals to landscapes. Pp 97–126 in RA Bradstock, 662 AM Gill, RJ Williams. Flammable Australia. Cambridge University Press, Cambridge, 663 UK. 664
Keith DA (2004) Ocean Shores to Desert Dunes: Native vegetation of NSW and ACT. Office 665 of Environment and Heritage NSW, Australia. 666
Ladd PG, Enright NJ (2010) Ecology of fire-tolerant Podocarps in temperate Australian 667 forests. Smithsonian Contributions to Botany 95, 141–156. 668
Lamont BB, Le Maitre DC, Cowling RM, Enright NJ (1991) Canopy seed storage in woody 669 plants. The Botanical Review 57, 277-317. 670
Lawes MJ, Richards A, Dathe J, Midgley JJ (2011) Bark thickness determines fire resistance 671 of selected tree species from fire-prone tropical savanna in north Australia. Plant Ecology 672 212, 2057–2069. 673
Lawes MJ, Richardson SJ, Clarke PJ, Midgley JJ, McGlone MS, Bellingham PJ (2014) Bark 674 thickness does not explain the susceptibility of Australian and New Zealand temperate 675 rainforests to anthropogenic fire. Journal of Biogeography, in press. 676
Leonard SWJ, Bennett AF, Clarke MF (2014) Determinants of the occurrence of unburnt 677 forest patches: Potential biotic refuges within a large, intense wildfire in south-eastern 678 Australia. Forest Ecology and Management 314, 83–93. 679
Melick DR, Ashton DH (1991) The Effects of natural disturbances on warm temperate rain-680 forests in south-eastern Australia. Australian Journal of Botany 39, 1–30. 681
Midgley JJ, Cowling RM (1993) Regeneration patterns in Cape subtropical transitional 682 thicket; where are all the seedlings? South African Journal of Botany 59, 496–499. 683
NSW Department of Environment and Conservation (2006) Wollemi Pine (Wollemia nobilis) 684 Recovery Plan. NSW Department of Environment and Conservation, Hurstville NSW. 685
Noble IR, Slatyer R (1980) The use of vital attributes to predict successional changes in plant 686 communities subject to recurrent disturbances. Plant Ecology 43, 5–21. 687
Noble IR, Bary GAV, Gill AM (1980) McArthurs fire danger meters expressed as equations. 688 Australian Journal of Ecology 5, 201–203. 689

22
Offord C, Porter C, Meagher P, Errington G (1999) Sexual reproduction and early plant 690 growth of the Wollemi pine (Wollemia nobilis), a rare and threatened Australian conifer. 691 Annals of Botany 84, 1–9. 692
Pappas N (2010) The impacts of the 2009 bushfires on cool temperate rainforest in the 693 Central Highlands of Victoria. Unpublished BSc (Honours) thesis, Monash University. 694
Philpot C (1977) Vegetative features as determinants of fire frequency and intensity. 695 Proceedings of the Symposium on the Environmental Consequences of Fire and Fuel 696 Management in Mediterranean Ecosystems. USDA Forest Service Technical Report, 697 WO-3, Washington, DC. 698
Pinard MA, Huffman J (1997) Fire resistance and bark properties of trees in a seasonally dry 699 forest in eastern Bolivia. Journal of Tropical Ecology 13, 727–740. 700
Plucinski MP (2003) The investigation of factors governing ignition and development of fires 701 in heathland vegetation. PhD Thesis. University of New South Wales. 702
Poorter L, Kitajima K, Mercado P, Chubiña J, Melgar I, Prins HH (2010) Resprouting as a 703 persistence strategy of tropical forest trees: relations with carbohydrate storage and shade 704 tolerance. Ecology 91, 2613–2627. 705
Prior LD, Bowman DMJS, Brook BW (2007) Growth and survival of two north Australian 706 relictual tree species, Allosyncarpia ternata (Myrtaceae) and Callitris intratropica 707 (Cupressaceae). Ecological Research 22, 228–236. 708
Pyrke AF, Marsden-Smedley JB (2005) Fire-attributes categories, fire sensitivity, and 709 flammability of Tasmanian vegetation communities. Tasforests 16, 35–46. 710
R Core Team (2013) R: A language and environment for statistical computing. R Foundation 711 for Statistical Computing, Vienna, Austria. URL. http://www.R-project.org/. 712
Seal AN, Pratley JE, Haig TJ, An M, Wu H (2010) Plants with phytotoxic potential: Wollemi 713 pine (Wollemia nobilis). Agriculture, Ecosystems and Environment 135, 52–57. 714
Scarff FR, Westoby M (2006) Leaf litter flammability in some semi-arid Australian 715 woodlands. Functional Ecology 20, 745–752. 716
Sullivan AL, WL McCaw, Cruz M, Matthews S, Ellis PF (2012) Fuel, fire weather and fire 717 behaviour in Australian ecosystems. Pp 51-78 in Bradstock RA, Gill AM, William RJ 718 (eds) Flammable Australia: Fire regimes, biodiversity and ecosystems in a changing 719 world. CSIRO, Australia. 720
Stocker GC (1981) Regeneration of a North Queensland rain forest following felling and 721 burning. Biotropica 13, 86–92. 722
Tanentzap AJ, Mountford EP, Cooke AS, Coomes DA (2011) The more stems the merrier: 723 advantages of multi-stemmed architecture for the demography of understorey trees in a 724 temperate broadleaf woodland. Journal of Ecology 100, 171–183. 725
Uhl JB, Kauffman C (1990) Deforestation, fire susceptibility, and potential tree responses to 726 fire in the Eastern Amazon Ecology 71, 437-449 727

23
Valette JC, Gomendy V, Marechal J, Houssard C, Gillon D (1994) Heat-transfer in the soil 728 during very low-intensity experimental fires-the role of duff and soil-moisture content. 729 International Journal of Wildland Fire 4, 225–237. 730
van Wagner CE (1973) Height of crown scorch in forest fires. Canadian Journal of Forest 731 Research 3, 373–378. 732
van Wilgen BW, Higgins KB, Bellstedt DU (1990) The role of vegetation structure and fuel 733 chemistry in excluding fire from forest patches in the fire-prone fynbos shrublands of 734 South Africa. Journal of Ecology 78,210–222. 735
Vesk PA, Westoby M (2004a) Sprouting by plants: the effects of modular organisation. 736 Functional Ecology 18, 939-945. 737
Vesk PA, Westoby M (2004b) Funding the bud bank: a review of the costs of buds. Oikos 738 106, 200–208. 739
Vesk PA, Warton DI, Westoby M (2004) Sprouting by semi-arid plants: testing a dichotomy 740 and predictive traits. Oikos 107, 72–89. 741
Webb LJ (1968) Environmental relationships of the structural types of Australian rain forest 742 vegetation. Ecology 49, 296–311. 743
Williams PR (2000) Fire-stimulated rainforest seedling recruitment and vegetative 744 regeneration in a densely grassed wet sclerophyll forest of north-eastern Australia. 745 Australian Journal of Botany 48, 651–658. 746
Williams PR, Parsons M, Jensen R, Tran C (2012) Mechanisms of rainforest persistence and 747 recruitment in frequently burnt wet tropical eucalypt forests. Austral Ecology 37, 268–748 275. 749
Williams RJ, Cook GD, Gill AM, Moore P (1999) Fire regime, fire intensity and tree survival 750 in a tropical savanna in northern Australia. Austral Ecology 24, 50–59. 751
Wood SW, Bowman DMJS (2011) Alternative stable states and the role of fire–vegetation–752 soil feedbacks in the temperate wilderness of southwest Tasmania. Landscape Ecology 753 27, 13–28. 754
Wooley L, Henkel T, Sillett SC (2008) Reiteration in the monodominant tropical tree 755 Dicymbe corymbosa (Caesalpiniaceae) and its potential adaptive significance. Biotropica 756 40, 32–43. 757
Zimmer HC, Auld TA, Benson J, Baker PJ (2013) Recruitment bottlenecks in a rare 758 Australian conifer: Wollemia nobilis. Biodiversity and Conservation 23, 203–215. 759
760
761
762
24
Figure captions 763
764
765 766
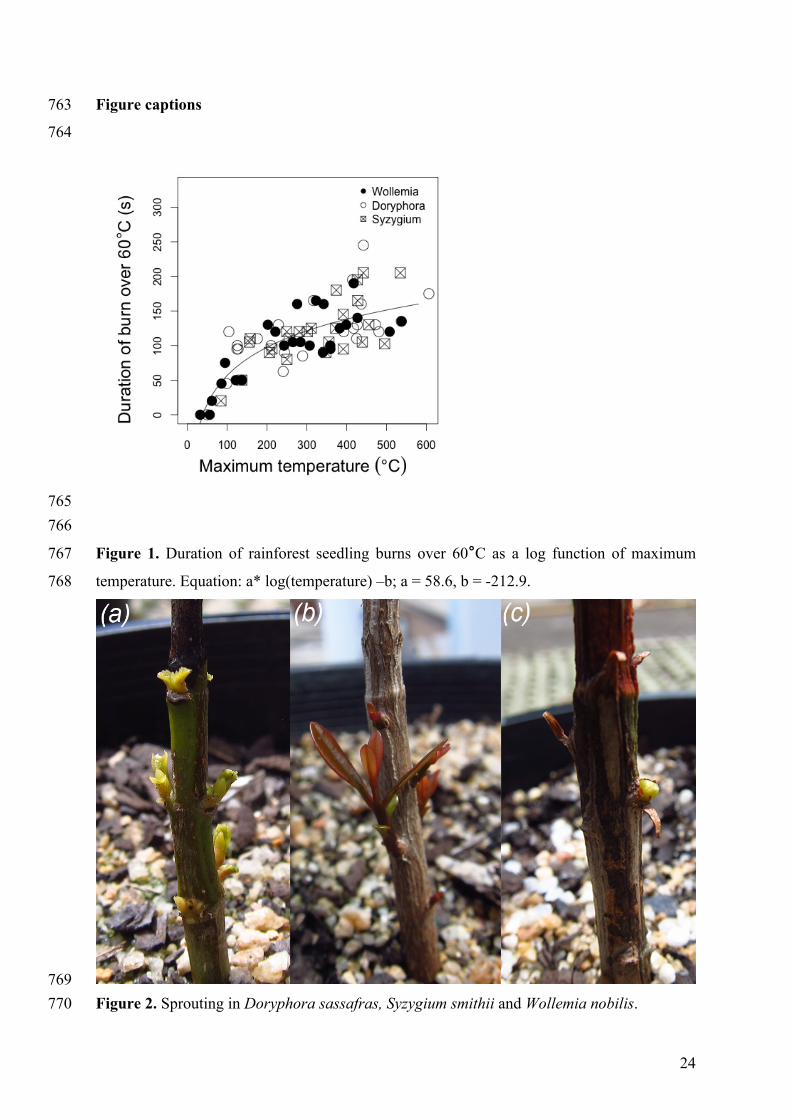
Figure 1. Duration of rainforest seedling burns over 60°C as a log function of maximum 767
temperature. Equation: a* log(temperature) –b; a = 58.6, b = -212.9. 768
769 Figure 2. Sprouting in Doryphora sassafras, Syzygium smithii and Wollemia nobilis. 770
25
771
772 773
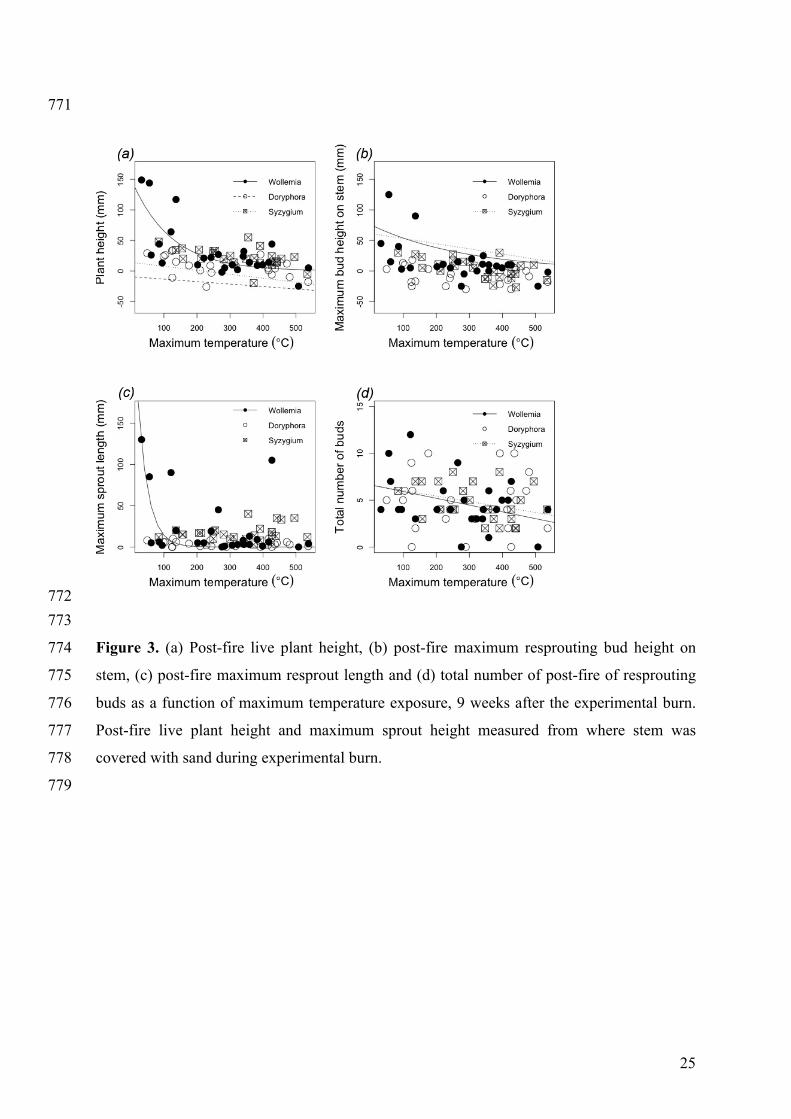
Figure 3. (a) Post-fire live plant height, (b) post-fire maximum resprouting bud height on 774
stem, (c) post-fire maximum resprout length and (d) total number of post-fire of resprouting 775
buds as a function of maximum temperature exposure, 9 weeks after the experimental burn. 776
Post-fire live plant height and maximum sprout height measured from where stem was 777
covered with sand during experimental burn. 778
779

26
780 781
Figure 4. (a) Post-fire live plant height and (b) total number of re-sprouting buds as a 782
function of (pre-fire) stem diameter. Post-fire live plant height measured from where stem 783
was covered with sand during experimental burn. 784
785 786

27
Figure 5. Characteristics of litter fire (a) post-burn percent weight (b) flame height and (c) 787
fuel moisture content as a function of litter composition. 788
789
790
791
792
793
794
795
796
797
798 799
28
800
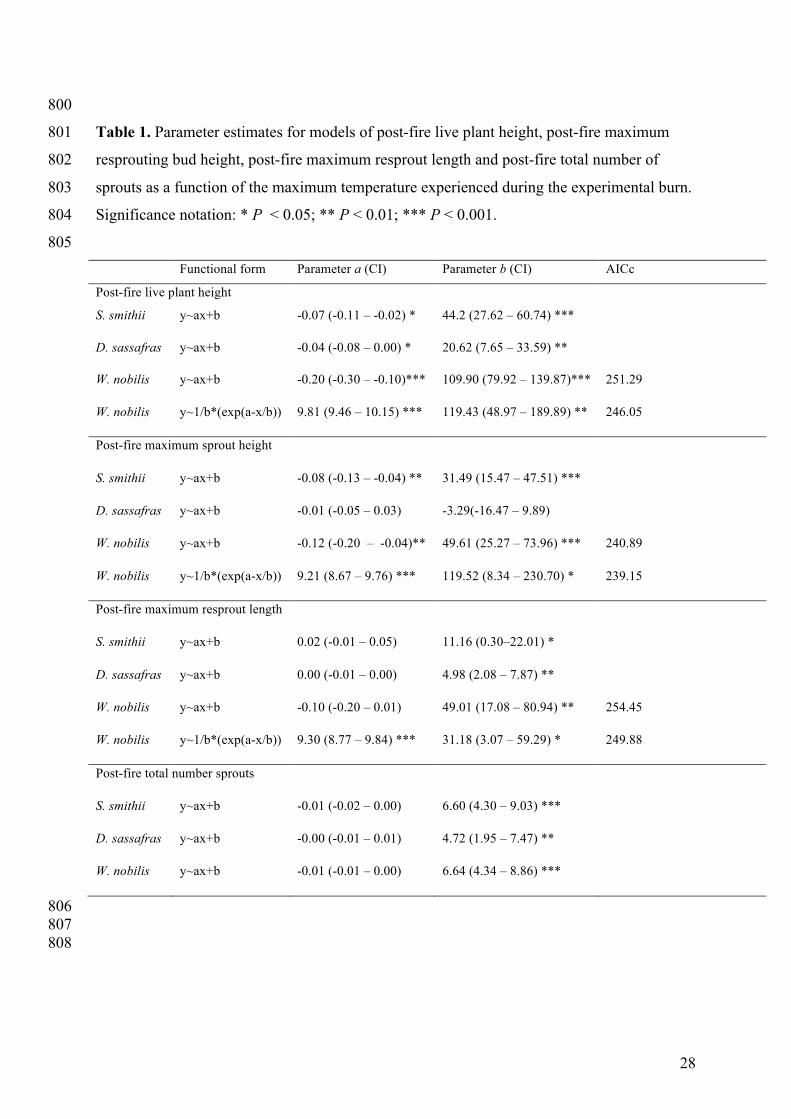
Table 1. Parameter estimates for models of post-fire live plant height, post-fire maximum 801
resprouting bud height, post-fire maximum resprout length and post-fire total number of 802
sprouts as a function of the maximum temperature experienced during the experimental burn. 803
Significance notation: * P < 0.05; ** P < 0.01; *** P < 0.001. 804
805 Functional form Parameter a (CI) Parameter b (CI) AICc
Post-fire live plant height
S. smithii y~ax+b -0.07 (-0.11 – -0.02) * 44.2 (27.62 – 60.74) ***
D. sassafras y~ax+b -0.04 (-0.08 – 0.00) * 20.62 (7.65 – 33.59) **
W. nobilis y~ax+b -0.20 (-0.30 – -0.10)*** 109.90 (79.92 – 139.87)*** 251.29
W. nobilis y~1/b*(exp(a-x/b)) 9.81 (9.46 – 10.15) *** 119.43 (48.97 – 189.89) ** 246.05
Post-fire maximum sprout height
S. smithii y~ax+b -0.08 (-0.13 – -0.04) ** 31.49 (15.47 – 47.51) ***
D. sassafras y~ax+b -0.01 (-0.05 – 0.03) -3.29(-16.47 – 9.89)
W. nobilis y~ax+b -0.12 (-0.20 – -0.04)** 49.61 (25.27 – 73.96) *** 240.89
W. nobilis y~1/b*(exp(a-x/b)) 9.21 (8.67 – 9.76) *** 119.52 (8.34 – 230.70) * 239.15
Post-fire maximum resprout length
S. smithii y~ax+b 0.02 (-0.01 – 0.05) 11.16 (0.30–22.01) *
D. sassafras y~ax+b 0.00 (-0.01 – 0.00) 4.98 (2.08 – 7.87) **
W. nobilis y~ax+b -0.10 (-0.20 – 0.01) 49.01 (17.08 – 80.94) ** 254.45
W. nobilis y~1/b*(exp(a-x/b)) 9.30 (8.77 – 9.84) *** 31.18 (3.07 – 59.29) * 249.88
Post-fire total number sprouts
S. smithii y~ax+b -0.01 (-0.02 – 0.00) 6.60 (4.30 – 9.03) ***
D. sassafras y~ax+b -0.00 (-0.01 – 0.01) 4.72 (1.95 – 7.47) **
W. nobilis y~ax+b -0.01 (-0.01 – 0.00) 6.64 (4.34 – 8.86) ***
806 807 808

29
Table 2. Parameter estimates for models of post-fire live plant height, post-fire maximum 809
resprouting bud height, post-fire maximum resprout length and post-fire total number of 810
resprouting buds as a function of (pre-fire) plant diameter. Significance notation: * P < 0.05; 811
** P < 0.01; *** P < 0.001. 812
813
Functional form Parameter a (CI) Parameter b (CI)
Post-fire live plant height
S. smithii y~ax+b 8.00 (-0.15 – 16.16) -7.67 (-39.65 – 24.32)
D. sassafras y~ax+b -16.79 (-43.49 – 9.91) 7.94 (-0.23 – 16.11)
W. nobilis y~ax+b 2.50 (-4.12 – 9.13) 10.92 (-47.07 – 68.91)
Post-fire maximum sprout height
S. smithii y~ax+b 4.70 (-4.45 – 13.86) -13.81 (-49.73 – 22.11)
D. sassafras y~ax+b 1.84 (-6.34 – 10.04) -11.64 (-38.41 – 15.13)
W. nobilis y~ax+b -0.59 (-5.51 – 4.31) 44.68 (1.65 – 87.72) *
Post-fire maximum resprout length
S. smithii y~ax+b 4.24 (0.63 – 9.10) 1.28 (-17.80 – 20.36)
D. sassafras y~ax+b 0.50 (-1.33 – 2.33) 2.06 (-3.93 – 8.04)
W. nobilis y~ax+b 5.84 (0.64 – 11.05)* -25.78 (-71.35 – 19.79)
Post-fire total number sprouts
S. smithii y~ax+b 0.25 (-0.86 – 1.36) 3.76 (-0.60 – 8.12)
D. sassafras y~ax+b 1.35 (-0.26 – 2.95) 0.52 (-4.73 – 5.77)
W. nobilis y~ax+b 0.54 (0.17 – 0.90) ** 0.21 (-2.98 – 3.40)
814 815 816
30
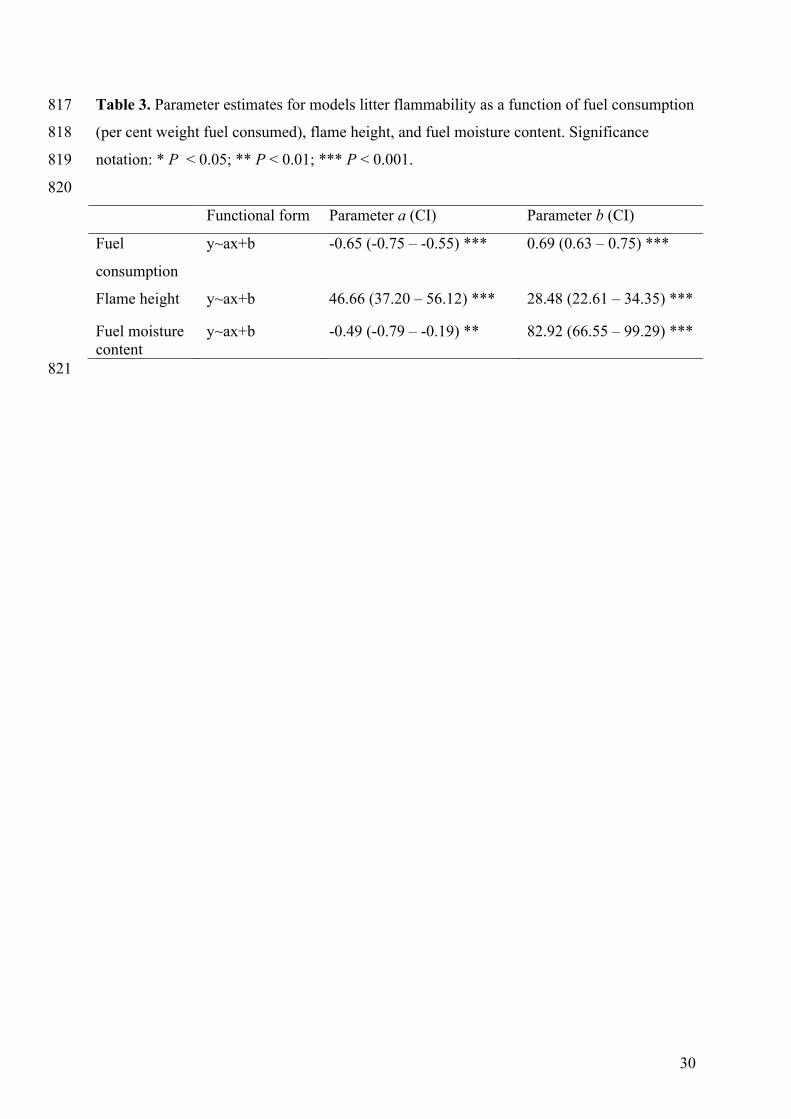
Table 3. Parameter estimates for models litter flammability as a function of fuel consumption 817
(per cent weight fuel consumed), flame height, and fuel moisture content. Significance 818
notation: * P < 0.05; ** P < 0.01; *** P < 0.001. 819
820
Functional form Parameter a (CI) Parameter b (CI)
Fuel
consumption
y~ax+b -0.65 (-0.75 – -0.55) *** 0.69 (0.63 – 0.75) ***
Flame height y~ax+b 46.66 (37.20 – 56.12) *** 28.48 (22.61 – 34.35) ***
Fuel moisture content
y~ax+b -0.49 (-0.79 – -0.19) ** 82.92 (66.55 – 99.29) ***
821
Related Documents











