Molecular Ecology (2001) 10 , 2003 – 2012 © 2001 Blackwell Science Ltd Blackwell Science, Ltd Frequent cytoplasmic exchanges between oak species that are not closely related: Quercus suber and Q. ilex in Morocco N. BELAHBIB,*† M.-H. PEMONGE,† A. OUASSOU,‡ H. SBAY,§ A. KREMER† and R. J. PETIT† * Université Ibn Tofail, Faculté des Sciences, Département de Biologie, BP 133, 14000 Kénitra, Morocco, † INRA, Laboratoire de génétique et d’amélioration des arbres forestiers, BP 45, F-33611 Gazinet Cedex, France, ‡ Institut Agronomique et Vétérinaire Hassan II, BP 6301 — Instituts, M-10101 Rabat, Morocco, § Centre National de la Recherche Forestière, Charia Omar Ibn Khattab, BP 763, M-10050 Rabat-Agdal, Morocco Abstract Chloroplast (cp) and mitochondrial (mt) DNA variation were studied in 97 populations of cork oak ( Quercus suber ) in Morocco; in 31 of these populations, holm oak ( Quercus ilex ), a clearly distinct species, also occurred and was compared with Q. suber . Three cpDNA and one mtDNA primer pairs were used in the survey, each in combination with one restriction enzyme. Six haplotypes belonging to two very divergent lineages were detected; one lin- eage predominates in each species, and is probably ancestral, as inferred from comparisons with other oak species. In the mixed-species populations, cytoplasmic genomes were fre- quently shared across species, as indicated by an introgression ratio of 0.63. This index is a new measure of the propensity of species to share locally genetic markers, varying from zero (complete differentiation) to one (no differentiation). By contrast, more closely related deciduous oak species ( Q. robur , Q. petraea and Q. pubescens ) have introgression ratios vary- ing from 0.82 to 0.97. The introgression events appear to have been more frequent in the direction Q. ilex (female) × Q. suber (male), a finding which seems attributable to the flower- ing phenology of these two species. This asymmetry may have favoured immigration of Q. suber beyond its main range, in regions already colonized by Q. ilex . There, rare hybrid- ization and further introgression through long distance pollen flow have established populations that are morphologically indistinguishable from Q. suber but that have cytoplasmic genomes originating from the local Q. ilex populations. Keywords : cpDNA, geographical structure, hybridization, introgression, mtDNA, PCR–RFLP Received 9 December 2000; revision received 22 March 2001; accepted 3 April 2001 Introduction Since the first review of the topic by Rieseberg & Soltis (1991), who demonstrated the high frequency of cytoplasmic exchanges across plant species boundaries, the sharing of chloroplast (cp) DNA or mitochondrial (mt) DNA genomes between different plant taxa has remained a common observation. To account for these exchanges, the most frequently cited mechanism is introgression, rather than convergence or incomplete lineage sorting, reflecting the high potential for interspecific gene flow in plants (Anderson 1953). Among closely related and at least partly sympatric plant species, the existence of cytoplasmic exchanges could well turn out to be the rule, rather than the exception, although to prove this would require a full and unbiased evaluation of the available evidence. Examples of cyto- plasmic exchanges at higher taxonomic levels, between more distantly related congeneric species (for instance classified in different sections of a genus), or even between distinct genera, have also been reported, and in many cases these were not anticipated. As a consequence, it is difficult to ‘demarcate a zone of safety’, where cytoplasmic exchanges become impossible (Rieseberg & Soltis 1991). Because cytoplasmic exchanges involving more diver- gent species may be less frequent than those involving closely related taxa, the evidence for introgression should be considered carefully. If introgression is confirmed, one may expect that the donor species will be identified more easily than when the species are closely related. Further- more, the underlying processes could differ from those identified in previous studies involving less distantly Correspondence: R. J. Petit. Fax: + 33 557979088; E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Ecology (2001)
10
, 2003–2012
© 2001 Blackwell Science Ltd
Blackwell Science, Ltd
Frequent cytoplasmic exchanges between oak species that are not closely related:
Quercus suber
and
Q. ilex
in Morocco
N . BELAHBIB, *† M. -H. PEMONGE,† A. OUASSOU,‡ H. SBAY,§ A. KREMER† and R . J . PETIT†*
Université Ibn Tofail, Faculté des Sciences, Département de Biologie, BP 133, 14000 Kénitra, Morocco,
†
INRA, Laboratoire de génétique et d’amélioration des arbres forestiers, BP 45, F-33611 Gazinet Cedex, France,
‡
Institut Agronomique et Vétérinaire Hassan II, BP 6301
—
Instituts, M-10101 Rabat, Morocco,
§
Centre National de la Recherche Forestière, Charia Omar Ibn Khattab, BP 763, M-10050 Rabat-Agdal, Morocco
Abstract
Chloroplast (cp) and mitochondrial (mt) DNA variation were studied in 97 populations ofcork oak (
Quercus suber
) in Morocco; in 31 of these populations, holm oak (
Quercus ilex
), aclearly distinct species, also occurred and was compared with
Q. suber
. Three cpDNA andone mtDNA primer pairs were used in the survey, each in combination with one restrictionenzyme. Six haplotypes belonging to two very divergent lineages were detected; one lin-eage predominates in each species, and is probably ancestral, as inferred from comparisonswith other oak species. In the mixed-species populations, cytoplasmic genomes were fre-quently shared across species, as indicated by an introgression ratio of 0.63. This index is anew measure of the propensity of species to share locally genetic markers, varying fromzero (complete differentiation) to one (no differentiation). By contrast, more closely relateddeciduous oak species (
Q. robur
,
Q. petraea
and
Q. pubescens
) have introgression ratios vary-ing from 0.82 to 0.97. The introgression events appear to have been more frequent in thedirection
Q. ilex
(female)
×
Q. suber
(male), a finding which seems attributable to the flower-ing phenology of these two species. This asymmetry may have favoured immigration of
Q. suber
beyond its main range, in regions already colonized by
Q. ilex
. There, rare hybrid-ization and further introgression through long distance pollen flow have establishedpopulations that are morphologically indistinguishable from
Q. suber
but that havecytoplasmic genomes originating from the local
Q. ilex
populations.
Keywords
: cpDNA, geographical structure, hybridization, introgression, mtDNA, PCR–RFLP
Received 9 December 2000; revision received 22 March 2001; accepted 3 April 2001
Introduction
Since the first review of the topic by Rieseberg & Soltis(1991), who demonstrated the high frequency of cytoplasmicexchanges across plant species boundaries, the sharing ofchloroplast (cp) DNA or mitochondrial (mt) DNA genomesbetween different plant taxa has remained a commonobservation. To account for these exchanges, the mostfrequently cited mechanism is introgression, rather thanconvergence or incomplete lineage sorting, reflecting thehigh potential for interspecific gene flow in plants (Anderson1953). Among closely related and at least partly sympatricplant species, the existence of cytoplasmic exchanges couldwell turn out to be the rule, rather than the exception,
although to prove this would require a full and unbiasedevaluation of the available evidence. Examples of cyto-plasmic exchanges at higher taxonomic levels, betweenmore distantly related congeneric species (for instanceclassified in different sections of a genus), or even betweendistinct genera, have also been reported, and in manycases these were not anticipated. As a consequence, it isdifficult to ‘demarcate a zone of safety’, where cytoplasmicexchanges become impossible (Rieseberg & Soltis 1991).
Because cytoplasmic exchanges involving more diver-gent species may be less frequent than those involvingclosely related taxa, the evidence for introgression shouldbe considered carefully. If introgression is confirmed, onemay expect that the donor species will be identified moreeasily than when the species are closely related. Further-more, the underlying processes could differ from thoseidentified in previous studies involving less distantly
Correspondence: R. J. Petit. Fax: + 33 557979088; E-mail:[email protected]
MEC1330.fm Page 2003 Monday, July 23, 2001 5:32 PM

2004
N . B E L A H B I B
E T A L .
© 2001 Blackwell Science Ltd,
Molecular Ecology
, 10, 2003–2012
related species. For instance, will these exchanges beasymmetric, as often reported at lower taxonomic levels?Will they involve pollen movements during range expan-sions (pollen swamping, whereby a native nuclear genomeis replaced by an alien one without affecting the (mater-nal) cytoplasmic genetic structure, that remains static;Potts & Reid 1988; Petit
et al
. 1997)? Or will they implyinstead the complete displacement of the native cytoplasmby an alien one in the absence of significant nuclear geneflow (Rieseberg & Soltis 1991; Rieseberg 1995)?
To answer these questions, the genus
Quercus
, which iswidely distributed over the northern hemisphere, and whichincludes over 500 species, is particularly well-suited. In theEuropean and American white oaks, cytoplasmic exchangescan be so frequent that the origin of the cpDNA or mtDNAgenomes shared cannot be identified unambiguously(Whittemore & Schaal 1991; Petit
et al
. 1993a, 2001a,b;Dumolin-Lapègue
et al
. 1997a, 1999). Quantitative analysesof cytoplasmic exchanges across a range of closely relatedoak species in Europe has revealed that the levels of intro-gression, although always of strong amplitude, could varysignificantly across species pairs (Dumolin-Lapègue
et al
.1999; Petit
et al
. 2001a). Simultaneously, phylogeneticstudies in the genus
Quercus
, using both nuclear and cyto-plasmic markers, have started to clarify its taxonomic struc-ture (Manos
et al
. 1999), providing some support for furtheranalyses of the amplitude of interspecific gene flow acrossincreasingly divergent taxa. Chloroplast exchanges havebeen identified at a variety of levels, including across sep-arate sections of the subgenus
Quercus
(Manos
et al
. 1999).According to the latest taxonomic interpretations, the
subgenus
Quercus
is divided into four sections, two strictlyAmerican (sections
Lobatae
and
Protobalanus
), one foundboth in America and in Eurasia (section
Quercus
sensustricto), and one strictly Eurasian (section
Cerris
) (Manos
et al
. 1999). Cork oak (
Q. suber
L.) and holm oak (
Q. ilex
Lam.) are two Mediterranean species of oak that belong tothe latter section. However, this section includes two welldifferentiated lineages, using both cpDNA and ribosomalDNA (Manos
et al
. 1999). These two groups of species hadbeen given various taxonomic status in the past, up to thesubgenus level (Schwarz 1937, 1964). One of the two groupsincludes
Q. ilex
and related evergreen species (such as
Q. coccifera
), and the other includes the deciduous species
Q. cerris
, as well as the evergreen
Q. suber
. Further work isneeded to decide whether the divergence between the
Q. ilex
and
Q. suber
groups is at the sectional level ornot. At any rate, they do represent clear distinct lineageswithin the
Quercus
genus, whether cpDNA, ribosomalDNA or morphological evidence is used.
Q. ilex
and
Q. suber
have a fairly similar geographicaldistribution throughout southwestern Europe and northAfrica;
Q. suber
is less abundant than
Q. ilex
, but theunusual feature of its bark, used to produce the familiar
cork, explains the great economic role of this partly domest-icated tree species. In Morocco, cork oak covers 320 200 ha,approximately 16% of the world’s surface for this species,and about 7% of the Moroccan forest. With 137 000 ha, i.e.half its original size, the Maâmora forest, northeast ofRabat, is still by far the largest cork oak forest in the world(M’Hirit 1996). In Morocco,
Q. suber
is facing, as in thesouthern part of its European range, an unprecedenteddecline due to several factors such as excessive pruningfor fodder, overexploitation of the bark and overgrazing,which prevents natural regeneration (Benzyane 1997). Corkoak genetic resources from Morocco are important not onlyfor the local economy but also internationally. Indeed,since global change may lead to warmer and drier climatein southern Europe, cork oak genetic resources from northAfrica may become of particular interest there, in the formof
ex situ
plantations (Varela & Eriksson 1995).In his study of cork oak in Morocco, Sauvage (1961)
summarizes the information available at that time on thebiogeography of
Q. suber
in this country. The most likelyhypothesis is that
Q. suber
originated from the Iberianpeninsula, and colonized Morocco from Gibraltar and thenthe Rif region no earlier than at the end of the Miocene. Forthe Postglacial, palynological data are now available fromnorth Africa, and a distinction is usually made between thedifferent pollen types of oaks (e.g. Bernard & Reille 1987;Carrión
et al
. 2000). These results indicate that both
Q. ilex
and
Q. suber
have been present in Morocco since at least theearly Postglacial (Reille
et al
. 1996). Between 14 000
bp
and12 000
bp
, in the Middle Atlas, oaks (
Q.
cf.
ilex
) seemed toextend in a landscape essentially dominated by the steppe(Lamb
et al
. 1989). On the other hand, it is clear from otherstudies that at higher altitude (in the Rif, in particular),deciduous oak forests (
Q. faginea
s. l.) were largely domin-ant and that the present distribution of
Q. suber
and
Q. ilex
had been facilitated by man at their expense, especiallyaround 1000
bp
, at the time of the invasion by the Arabs(Reille 1977; Reille
et al
. 1996). The high tolerance to graz-ing in the case of
Q. ilex
, and the protection against forestfires conferred by the cork in the case of
Q. suber
(Carrión
et al
. 2000), may explain their development at that time. Inaddition, in Morocco, and especially in the Maâmora forest,
Q. suber
produces sweet acorns that constitute an appreci-ated source of food for human populations, a fact that mayalso account for its extension. In the High Atlas, southeastof Marrakech, where isolated patch of
Q. suber
(mixed withthe more abundant
Q. ilex
) disconnected from the mainrange can still be found at high altitude (1600–2009 m),pollen attributed to this species has been present since atleast the Boreal (but never in high percentages), whendeciduous oaks were dominating (Bernard & Reille 1987).
In Morocco,
Q. suber
is now mostly restricted to the north-east, whereas
Q. ilex
, the most abundant tree species (23.5%of the forests), grows further south at high elevation in the
MEC1330.fm Page 2004 Monday, July 23, 2001 5:32 PM

I N T R O G R E S S I O N A M O N G O A K S P E C I E S I N M O R O C C O
2005
© 2001 Blackwell Science Ltd,
Molecular Ecology
, 10, 2003–2012
Atlas mountains (Benabid 1985). The overlap betweenthe range of the two species in Morocco is indicated inFig. 1.
Examples of hybrids between the two species are listedin Camus (1936–39). Although usually considered to berare, they are mentioned in Algeria, France, Italy, Portugaland Spain (Camus 1936–39; Franco 1990 in Gil Sánchez
et al
. 1996). Reports of other hybrids with
Q. suber
involvedeciduous species such as
Q. afares
Pomel in Algeria(Camus 1936–39) and
Q. cerris
L. in Italy (Bellarosa
et al
.1996) (all belonging to section
Cerris
) but also
Q. faginea
s.l. in Spain (Gil Sánchez
et al
. 1996), which belongs to thesection
Quercus
s.s. (Manos
et al
. 1999). Here, we will exam-ine the level of cpDNA variation within
Q. suber
, and wewill quantify the level of introgression with
Q. ilex
using aspecifically developed introgression index that allowscomparisons with other cp/mtDNA (= cyDNA) studiesinvolving different pairs of oak species.
Materials and methods
Plant material
The material used in the study was collected from 97populations distributed in seven geographically contrastingregions of Morocco (Fig. 1). The
Quercus suber
populationswere pure or mixed with
Quercus ilex
(31 cases out of 97).First (in 1996), 24 populations were sampled, includingseven where both species were present (267 trees, mean of8.6 trees per population and per species). Second (from
1997 to 2000), 73 other populations were added (508 trees,mean of 5.8 trees per population). Increasing the number ofpopulations appeared more interesting than retaining highsample sizes within population (Pons & Petit 1995). Intotal, 775 trees were sampled throughout Morocco: 610
Q. suber
trees and 165
Q. ilex
trees; all individuals sampledhad typical bark characters.
DNA isolation, amplification and digestion
Total DNA was isolated from leaves. The procedure de-scribed in Dumolin
et al
. (1995) was followed, except that 1
m
1,4-Dithiothreitol was used in place of
β
-mercaptoethanol.The polymerase chain reaction (PCR) method was asdetailed in (Demesure
et al
. 1995). We used five pairs ofchloroplast primers (DT, TF, CD, AS and VL) and two pairsof mitochondrial primers (
nad
7/1–2 and
nad
7/2–3) for thefirst 24 populations; for the 73 other populations, thechloroplast primer pairs DT and TF and one mitochondrialprimer pair (
nad
7/2–3) were retained, together with a newcpDNA primer pair (SR) that revealed additional variation(see Table 1 for details and sources of primers). PCRproducts were digested with five units of either
Hin
fI or
Taq
I, and electrophoresis in polyacrylamide gels was asdescribed in El Mousadik & Petit (1996).
Relationships between haplotypes
The computer program
tcs
(Clement
et al
. 2000) was usedto depict relationships among the haplotypes.
tcs
implements
Fig. 1 Distribution of Quercus suber L. and Quercus ilex L. in Morocco, and location of the 97 populations sampled.
MEC1330.fm Page 2005 Monday, July 23, 2001 5:32 PM

2006
N . B E L A H B I B
E T A L .
© 2001 Blackwell Science Ltd,
Molecular Ecology
, 10, 2003–2012
the estimation of gene genealogies from DNA sequences asdescribed by Templeton
et al
. (1992). This cladogram estima-tion method is also known as statistical parsimony. Becausethere were no more than four length variants for any givenpolymorphic restriction fragment in our study, it waspossible to code the haplotypes found using arbitrarilythe bases A, T, C or G. To indicate the abundance ofeach haplotype, circles were made proportional totheir frequencies.
Genetic diversity analysis
For the 73
Q. suber
populations and the 24
Q. ilex
popula-tions studied with DT, TF, SR and
nad
7/2–3, cpDNA hap-lotype frequencies were computed for each population. Theaverage intrapopulation diversity (
h
S
), the total diversitywithin species (
h
T
), and the coefficient of differentiation(
G
ST
= 1 –
h
S
/
h
T
) and their standard deviations were estim-ated following Pons & Petit (1995). Taking advantage of thenumerous mixed-species populations where both
Q. suber
and
Q. ilex
had been sampled, an introgression index wasobtained as follows. First, inter- and intraspecific iden-tities were measured on the basis of the 31 mixed-speciespopulations: for species 1 (respectively 2) in population
k
,the estimator of the intraspecific identity is given by:
(and , respectively), where
i
indexes the
haplotypes (alleles), whereas the estimator of the inter-specific identity is given by:
(after Dumolin-Lapègue
et al.
1999).
We define here the introgression ratio
IG
as the meanacross all pairs of the interspecific identities, divided by themean intraspecific identities:
.
This ratio will tend to be one when genetic variation isspecies-independent (i.e. when identity between specieswithin population is equivalent to identities within specieswithin population, that is, for all populations
k
:
J
12
k
≈
J
1
k
≈
J
2
k
), and zero when the species are fully differentiated,implying absolutely no variant shared across species (all
J
12
k
= 0, regardless of the values taken by
J
1
k
and
J
2
k
). It ispossible to compute the value that this index would takeif haplotypes were shared between species but have inde-pendent geographical distributions; in that case
IG
wouldnevertheless be higher than zero because locally the twospecies may share the same haplotype(s) by chance. Tocheck that there is some concordant geographical associ-ation across species, we computed an expected value forthis index called
IG
e
. It is the ratio of the overall identitybetween species, based on the mean frequencies in eachspecies (
J
12T
), divided by the mean intraspecific identitieswithin population, with both the numerator and the denom-inator being computed throughout the complete range,and not only in areas of sympatry, as in IG above. Being aratio, IG can be compared across studies, contrary to theidentities themselves.
Results
In the first screening of 24 populations, up to 62 restrictionfragments were obtained with the six cpDNA PCR frag-ments digested with HinfI and TaqI, and six with the twomtDNA PCR fragments. We could identify four point muta-tions and 10 insertion/deletions (indels): 12 cpDNA andtwo mtDNA restriction fragments were polymorphic butthey allowed the detection of only two highly divergenthaplotypes. One was referred to as haplotype I since it wasfound in all but two Quercus ilex individuals, whereas theother one (named haplotype S) was found in a majority ofQ. suber trees but was nearly absent in Q. ilex. Comparisonswith other oak species indicated that Q. cerris is character-ized by haplotypes closely related to S, and that both haplo-types I and S differ significantly from all types found in
Table 1 Description of the eight pairs of chloroplast and mitochondrial primers used in this study
Abbr. Primer 1 Primer 2 Reference* Ta °C Restr. Enz. Survey†
AS psaA [PS I (P 700 apoprotein A1)] trnS [tRNA-Ser(GGA)] 1 58.0 HinfI 1DT trnD [tRNA-Asp(GUC)] trnT [tRNA-Thr(GGU)] 1 54.5 TaqI 3TF trnT [tRNA-Thr(UGU)] trnF [tRNA-Phe(GAA)] 2 57.5 HinfI 3CD trnC [tRNA-Cys(GCA)] trnD [tRNA-Aps(GUC)] 1 58.0 TaqI 1VL trnV [tRNA-Val(UAC)3′exon] rbcL [RuBis CO large subunit] 3 57.5 HinfI 1SR trnS [tRNA-Ser(GCU)] trnR [tRNA-Arg(UCU)] 4 50.0 HinfI 2
nad7/1–2 nad7 exon 1 nad7 exon 2 3 47.0 HinfI 1nad7/2–3 nad7 exon 2 nad7 exon 3 3 57.0 HinfI 3
*1: Demesure et al. 1995; 2: Taberlet et al. (1991); 3: Dumolin-Lapègue et al. (1997b); 4: D. Grivet, unpublished.†1: initial survey (24 populations), 2: second survey (73 populations), 3: complete survey (97 populations).
J1k =
n1k x1ki2
i∑
1–
n1k 1–------------------------------------- J2k
J12k x1kix2kii
∑=
IGJ12k
k∑
J1k J2k+( )/2k∑
--------------------------------------------=
MEC1330.fm Page 2006 Monday, July 23, 2001 5:32 PM

I N T R O G R E S S I O N A M O N G O A K S P E C I E S I N M O R O C C O 2007
© 2001 Blackwell Science Ltd, Molecular Ecology, 10, 2003–2012
oaks of the section Quercus s.s., including Q. canariensisand Q. faginea from Morocco (Petit et al. 2001b).
The combination SR-HinfI gave additional information,as six haplotypes were found instead of two. The haplo-types S-1 and S-2 differ by only one mutation from eachother, and the four haplotypes I are also closely related(Fig. 2 and Table 2). In Q. suber, all six haplotypes werefound. The haplotypes S-1 and I-2 are the most frequentones. Haplotype S-2 is present in the north of the CentralPlateau and in the Maâmora forest, between Rabat andKénitra, I-1 and I-3 are limited to the centre of the CentralPlateau, and I-4 is found in two populations of the Rif. Fourof these haplotypes are shared across species, especially
I-2, which is the most frequent haplotype in Q. ilex, but isalso found frequently in Q. suber when both species growtogether or in close proximity, such as in the Bab Azharforest (Middle Atlas, east of Fes), in the six isolated popu-lations of the High Atlas, and in the south of the CentralPlateau. The exceptions are two populations from theMiddle Atlas (Jaâba and Ribaâ, south of Fes) growing inforests dominated by Q. ilex, that have the S-1 type (Fig. 3a).In Q. ilex, four of the six haplotypes were found (S-1, I-1,I-2 and I-4); most individuals present haplotypes of lineageI, except in five isolated cases (in two forests from theAtlantic plain and in three from western Rif) (Fig. 3b).
The level of differentiation GST among the 73 MoroccanQ. suber populations analysed during the second surveywas 0.84, significantly higher than the value found inQ. ilex (0.33) (Table 3). The introgression index IG was 0.63,compared to values ranging between 0.82 and 0.97 in otheroak species pairs studied by Dumolin-Lapègue et al. (1999)(Table 4). Assuming that the distribution of haplotypes inthe two oak species is independent, we would expect anintrogression ratio IGe of 0.33 instead, a value almost twiceas low as that which was actually found (0.63). The differ-ence is even stronger for the three pairs of oaks studied sofar (Table 4).
Discussion
Cytoplasmic exchanges in plants have been described in anunexpectedly high number of cases at different taxonomiclevels (Rieseberg & Soltis 1991). In a genus like Quercus,there are a variety of such levels to explore, and our studyextends the quantitative description of cytoplasmic intro-gression to the case of oak species that are not closelyrelated, in contrast with previous population studies in thisgenus (Dumolin-Lapègue et al. 1999; Petit et al. 2001a). InMorocco, neither Q. ilex nor Q. suber, which belong to thesection Cerris (Manos et al. 1999), are sharing haplotypeswith oaks of the section Quercus s.s. (such as Q. canariensis,Q. faginea or Q. pyrenaica; see Petit et al. 2001b). On theother hand, the analysis of both cpDNA and mtDNAdiversity of Q. suber in Morocco revealed a clear-cut
S-1 S-2
I-2
I-1
I-3I-1
Fig. 2 Relationships among the six haplotypes detected. Boxeswere used when it could not be inferred to which haplotype theother lineage was most closely related. The sizes of the circles areproportional to the frequency of the corresponding haplotypes.
Table 2 Description of the six haplotypes identified
Haplotype DTTaq-3* TFHinf-1 TFHinf-3 SRHinf-2 SRHinf-4a SRHinf-4b† SRHinf-5 nad7Hinf-1
S-1 1 2 1 2 9 1 3 2S-2 1 2 1 2 9 2 3 2I-1 2 1 2 2 1 ? 1 1I-2 2 1 2 2 1 ? 2 1I-3 2 1 2 2 1 ? 4 1I-4 2 1 2 1 1 ? 2 1
*Length variants for each fragment are coded according the their mobility in the gel (1–4), whereas 9 indicate a point mutation.†This length variation occurs within one of the two fragments produced by a point mutation (cf. SRHinf-4a).
MEC1330.fm Page 2007 Monday, July 23, 2001 5:32 PM

2008 N . B E L A H B I B E T A L .
© 2001 Blackwell Science Ltd, Molecular Ecology, 10, 2003–2012
cytoplasmic exchange with Q. ilex. Usually, only cpDNAis used to characterize these interspecific cytoplasmicexchanges. Since both genomes are maternally inherited inoaks (Dumolin et al. 1995), their persistent association wasexpected, but confirms the findings of Dumolin-Lapègueet al. (1998) on the strict cotransmission of both cytoplasmicgenomes across generations in oaks. Furthermore, the fact
that several independent mutations from both cyto-plasmic genomes yielded the same information rulesout the possibility of convergence to account for theinterspecific cytoplasmic sharing.
In principle, joint sharing of haplotypes due to incom-plete lineage sorting could also account for the cytoplasmicexchanges. Many plant species are thought to be initiallyparaphyletic shortly after speciation (Rieseberg & Brouillet1994). Depending on the species’ effective populationsizes, this situation could last for a long time (Neigel &Avise 1986). Two arguments militate against this hypo-thesis in our example: the likely ancestry of the divergencebetween the two taxa, which is very high, and the similar-ity of the geographical distribution of the haplotypeswhere Q. ilex and Q. suber are in sympatry in Morocco.
Actually, previous observations based on isozymemarkers had confirmed that isolated cases of hybrid-ization between Q. ilex and Q. suber species could occur(Elena-Rosselló et al. 1992). Nevertheless, as stressed in
Table 3 Results of the analysis of diversity
Quercus suber Quercus ilex
Number of populations 73 24Mean sample size/population 5.26 5.17Number of haplotypes 6 4
hS 0.06 (0.02) 0.15 (0.05)hT 0.38 (0.06) 0.22 (0.08)GST 0.84 (0.05) 0.33 (0.11)
Fig. 3 Distribution of the six haplotypesfound in this study: (a) Quercus suber, and(b) Quercus ilex. The symbols are proportionalto the number of individuals analysed. Onlythe results of the complete screening areillustrated.
MEC1330.fm Page 2008 Monday, July 23, 2001 5:32 PM
I N T R O G R E S S I O N A M O N G O A K S P E C I E S I N M O R O C C O 2009
© 2001 Blackwell Science Ltd, Molecular Ecology, 10, 2003–2012
Varela & Valdiviesso (1996), the probability of hybridiza-tion is likely to be low given the flowering phenology ofthe two species in mixed stands. Indeed, Q. ilex usuallyends flowering some weeks before Q. suber begins.Because these trees are generally protandrous, hybridswith Q. ilex as the mother tree should be less rare thanthe reciprocals.
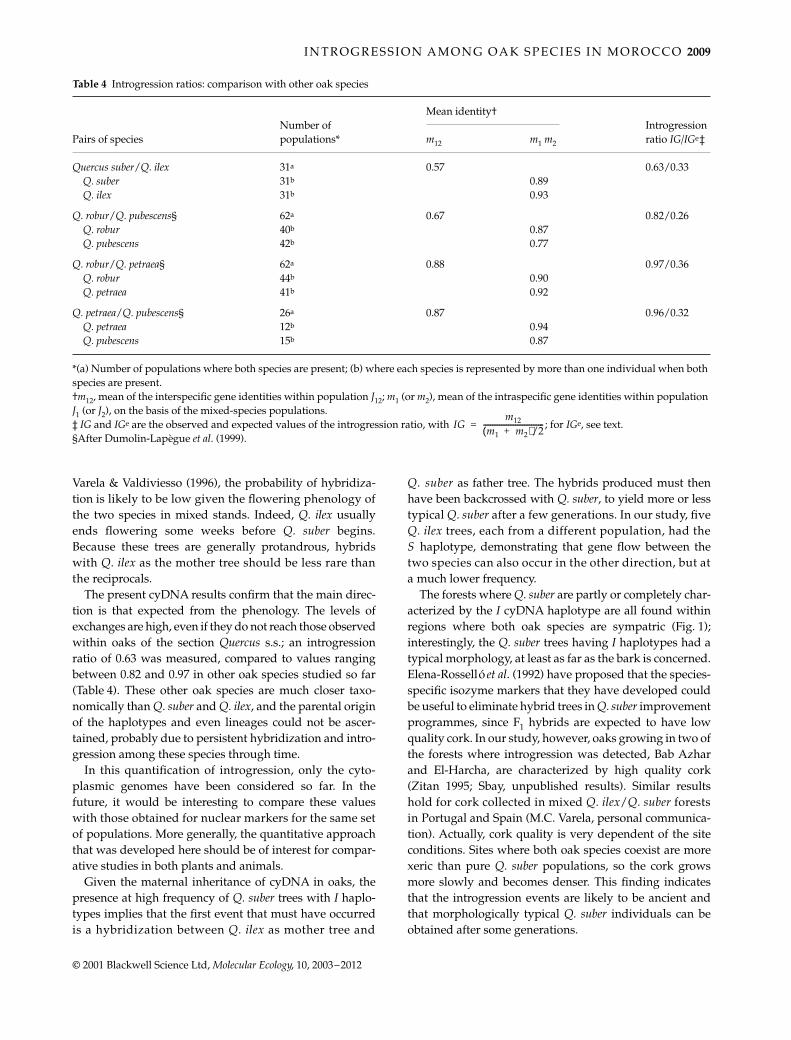
The present cyDNA results confirm that the main direc-tion is that expected from the phenology. The levels ofexchanges are high, even if they do not reach those observedwithin oaks of the section Quercus s.s.; an introgressionratio of 0.63 was measured, compared to values rangingbetween 0.82 and 0.97 in other oak species studied so far(Table 4). These other oak species are much closer taxo-nomically than Q. suber and Q. ilex, and the parental originof the haplotypes and even lineages could not be ascer-tained, probably due to persistent hybridization and intro-gression among these species through time.
In this quantification of introgression, only the cyto-plasmic genomes have been considered so far. In thefuture, it would be interesting to compare these valueswith those obtained for nuclear markers for the same setof populations. More generally, the quantitative approachthat was developed here should be of interest for compar-ative studies in both plants and animals.
Given the maternal inheritance of cyDNA in oaks, thepresence at high frequency of Q. suber trees with I haplo-types implies that the first event that must have occurredis a hybridization between Q. ilex as mother tree and
Q. suber as father tree. The hybrids produced must thenhave been backcrossed with Q. suber, to yield more or lesstypical Q. suber after a few generations. In our study, fiveQ. ilex trees, each from a different population, had theS haplotype, demonstrating that gene flow between thetwo species can also occur in the other direction, but ata much lower frequency.
The forests where Q. suber are partly or completely char-acterized by the I cyDNA haplotype are all found withinregions where both oak species are sympatric (Fig. 1);interestingly, the Q. suber trees having I haplotypes had atypical morphology, at least as far as the bark is concerned.Elena-Rosselló et al. (1992) have proposed that the species-specific isozyme markers that they have developed couldbe useful to eliminate hybrid trees in Q. suber improvementprogrammes, since F1 hybrids are expected to have lowquality cork. In our study, however, oaks growing in two ofthe forests where introgression was detected, Bab Azharand El-Harcha, are characterized by high quality cork(Zitan 1995; Sbay, unpublished results). Similar resultshold for cork collected in mixed Q. ilex/Q. suber forestsin Portugal and Spain (M.C. Varela, personal communica-tion). Actually, cork quality is very dependent of the siteconditions. Sites where both oak species coexist are morexeric than pure Q. suber populations, so the cork growsmore slowly and becomes denser. This finding indicatesthat the introgression events are likely to be ancient andthat morphologically typical Q. suber individuals can beobtained after some generations.
Table 4 Introgression ratios: comparison with other oak species
Number of populations*
Mean identity†Introgression ratio IG/IGe‡Pairs of species m12 m1 m2
Quercus suber/Q. ilex 31a 0.57 0.63/0.33Q. suber 31b 0.89Q. ilex 31b 0.93
Q. robur/Q. pubescens§ 62a 0.67 0.82/0.26Q. robur 40b 0.87Q. pubescens 42b 0.77
Q. robur/Q. petraea§ 62a 0.88 0.97/0.36Q. robur 44b 0.90Q. petraea 41b 0.92
Q. petraea/Q. pubescens§ 26a 0.87 0.96/0.32Q. petraea 12b 0.94Q. pubescens 15b 0.87
*(a) Number of populations where both species are present; (b) where each species is represented by more than one individual when both species are present.†m12, mean of the interspecific gene identities within population J12; m1 (or m2), mean of the intraspecific gene identities within population J1 (or J2), on the basis of the mixed-species populations.‡ IG and IGe are the observed and expected values of the introgression ratio, with ; for IGe, see text.§After Dumolin-Lapègue et al. (1999).
IGm12
m1 m2+( )/2----------------------------------=
MEC1330.fm Page 2009 Monday, July 23, 2001 5:32 PM

2010 N . B E L A H B I B E T A L .
© 2001 Blackwell Science Ltd, Molecular Ecology, 10, 2003–2012
The fact that all Q. suber trees from the isolated BabAzhar forest (74 trees in seven different sites), fromsouthern Central Plateau forests (25 trees from five dif-ferent sites) and from the High Atlas (38 trees in six differ-ent sites) had without exception the cytoplasm of type I isremarkable. Given the isolated location of these forests, farfrom the main range of Q. suber, and the distribution ofsampling sites, these results suggest that the populationsare virtually fixed for the I cytoplasm. Although some ofthese populations include only a few dozen of trees (inthe High Atlas), others are very large (about 15 000 ha inthe case of Bab Azhar). To account for this completecytoplasmic substitution, two contrasting hypotheses canbe suggested.
The first and more classical one is the so-called ‘cyto-plasmic capture’ (Rieseberg & Soltis 1991; Rieseberg 1995).In this scenario, Q. suber having typical S haplotypesand initially present in these forests, were in contactwith Q. ilex trees with I haplotypes. Progressively, thesecork oaks were replaced by other cork oaks having an Ihaplotype acquired by hybridization of a Q. ilex mothertree with a Q. suber acting as father, followed by back-crossing with Q. suber. In the forests from Central Plateauand from the Rif region, where Q. suber is characterized byboth haplotypes I and S, this process would be incomplete,whereas in Bab Azhar and in the five other forests ofsouthern Central Plateau and the High Atlas a full replace-ment of the S haplotype by the I haplotype would haveoccurred. This suggests some kind of selective sweeps infavour of the I haplotype, and/or chance effects that ledto the elimination of the S haplotype through drift.
Second, considering the limited extent of gene flowthrough acorns but the much more prevalent pollen flow inoaks (Petit et al. 1993b; Ennos 1994), as well as the isolationof the Bab Azhar forest or the High Atlas populations fromthe remaining of the species’ range, a mechanism similarto that proposed by Petit et al. (1997) for pedunculate andsessile oaks in France can be proposed. In this hypothesis(so-called ‘nuclear capture’ or ‘pollen swamping’), cork oakwould have colonized these forests by means of pollen,through hybridization and backcrossing with the alreadyestablished Q. ilex trees. Hence, the species would haveextended its range in the south and in the east of the coun-try without having to rely on long distance seed dispersalevents. Because colonization by seed is not required forQ. suber in this scenario, there is no need for a mechanismto eliminate the S haplotype. This makes the hypothesisquite parsimonious. The high level of genetic differenti-ation found for the 73 Moroccan populations, based on thedistribution of the six cpDNA haplotypes (GST = 0.84),contrasts clearly with the low level of differentiationfor nuclear (i.e. biparentally inherited) markers: FST variesfrom 0.03 to 0.11 and 0.16, in three studies involving differentloci and different regions (Elena-Rosselló & Cabrera 1996;
Toumi & Lumaret 1998; Jiménez et al. 1999), suggesting amuch more extensive pollen than seed flow. The low level ofcyDNA differentiation among Q. ilex populations (GST = 0.33)should be re-evaluated, given the low levels of cyDNAdiversity for this species in Morocco and the more limitedsample size; the occasional but incomplete cases of intro-gression with Q. suber (observed in five populations) mayalso contribute to produce this unusually low estimate (cf.Petit et al. 2001b for comparisons with other oak species).
Given that the Q. suber populations characterized byhaplotypes I are found in very distant regions, it wouldseem that these introgression events have occurred severaltimes independently. The similar geographical distributionof two rare haplotypes (I-1 and I-4) in both oak species rein-forces this opinion (compare Fig. 3a,b). Additionally, thesharing of the same cytoplasm by the two species in theHigh Atlas region supports the idea that these extremelysmall stands [sometimes involving only a few trees,Sauvage (1961); and personal observations] have not beenartificially introduced. Indeed, if seeds of Q. suber collectedin the main range of the species had been used to establishthese stands during historical times, we would insteadexpect to find at least some trees characterized by the moreabundant S haplotype. Furthermore, these conclusions aresupported by the finding of a continuous presence, sincethe Boreal, of Q. suber pollen types at low densities in fossilsediments of the High Atlas (Bernard & Reille 1987). Toresolve this issue more convincingly, it would be neces-sary to have additional variation within Q. ilex. That thesesmall and extremely isolated cork oak stands could be ofnative origin is an important issue, because they are clearlythreatened, given their very reduced size. Actually, duringsampling in the High Atlas, we were unable to find asingle Q. suber tree in two of the stations quoted by Sauvagein 1961; obviously, Q. suber does not exist there anymore.
In the Middle Atlas, we have another situation: in twoforests dominated by Q. ilex, a few old individuals ofQ. suber were found (total population of less than 30 treesin each case). All Q. suber trees analysed have the S-1haplotype, implying that they have colonized this regionby seed, most probably through human seed transfer, giventheir small size and isolation from other Q. suber stands.
Given the strong divergence between Q. ilex andQ. suber, the finding of relatively high level of cyDNA intro-gression between these taxa was unexpected. Incompletelineage sorting and convergence were not likely to apply inthis case. The geographical pattern of cytoplasmic sharingand its directionality indicate that processes involved aresimilar to those detected among less divergent oak species.In plants, even when dealing with relatively divergent taxa,it is, therefore, conceivable that hybridization and intro-gression play some significant adaptative role and thatinterspecific gene flow is to some extent evolutionarilyadjusted rather than merely accidental.
MEC1330.fm Page 2010 Monday, July 23, 2001 5:32 PM

I N T R O G R E S S I O N A M O N G O A K S P E C I E S I N M O R O C C O 2011
© 2001 Blackwell Science Ltd, Molecular Ecology, 10, 2003–2012
Acknowledgements
This study has benefited from a grant from a Projet de Rechercheet d’Aide au Dévelopement nos 98–11, from INRA. We thank NajatAlfioun for her initial contribution in the project, and Mrs. Tahri,Lamnaouar and El Arabi who participated in the collection ofoak material in the Central Plateau, in Bab Azhar and in the Rif.We also thank El Alaoui Hajjam for his help in the collection ofoaks from the High Atlas, the Middle Atlas and the Rif, and threeanonymous reviewers for their helpful comments on a previousversion of the manuscript.
References
Anderson E (1953) Introgressive hybridization. Biological Review,28, 280–307.
Bellarosa B, Schirone B, Maggini F, Fineschi S (1996) Inter- andintraspecific variation in three Mediterranean oaks (Q. cerris,Q. suber, Q. crenata). In: Inter- and Intraspecific Variation in EuropeanOaks: Evolutionary Implications and Practical Consequences (edsKremer A, Muhs H-J), pp. 239–276. Office for Official Publica-tions of the European Communities, Luxembourg.
Benabid A (1985) Les écosystèmes forestiers, préforestiers etprésteppiques du Maroc: diversité, répartition, biogéographieet problèmes posés par leur aménagement. Forêt Méditerranéenne,7, 53–64.
Benzyane M (1997) Improvement strategy of forest tree speciesin Morocco: the case of cork oak. In: European Forest GeneticRessources Programme (EUFORGEN) — Quercus Suber Network(eds Turok J, Varela MC, Hansen C), pp. 60–67. Compilers Reportof the third and fourth meeting, 1996/Italy, 1997/Spain.
Bernard J, Reille M (1987) Nouvelles analyses polliniques dansl’Atlas de Marrakech, Maroc. Pollen et Spores, 39, 225–240.
Camus A (1936–39) Les Chênes. Monographie du genre Quercus. PaulLechevallier, Paris.
Carrión JS, Parra I, Navarro C, Munuera M (2000) Past distributionand ecology of the cork oak (Quercus suber) in the Iberian penin-sula: a pollen-analytical approach. Diversity and Distributions, 6,29–44.
Clement M, Posada D, Crandall K (2000) TCS: a computer programto estimate gene genealogies. Molecular Ecology, 9, 1657–1660.
Demesure B, Sodzi N, Petit RJ (1995) A set of universal primersfor amplification of polymorphic non-coding regions of mito-chondrial and chloroplast DNA in plants. Molecular Ecology, 4,129–131.
Dumolin S, Demesure B, Petit RJ (1995) Inheritance of chloroplastand mitochondrial genomes in pedunculate oak investigatedwith an efficient PCR method. Theoretical and Applied Genetics,91, 1253–1256.
Dumolin-Lapègue S, Demesure B, Le Corre V, Fineschi S, Petit RJ(1997a) Phylogeographic structure of white oaks throughoutthe European continent. Genetics, 146, 1475–1487.
Dumolin-Lapègue S, Kremer A, Petit RJ (1999) Are chloroplastand mitochondrial DNA variation species independent in oaks?Evolution, 53, 1406–1413.
Dumolin-Lapègue S, Pemonge M-H, Petit RJ (1997b) An enlargedset of consensus primers for the study of organelle DNA inplants. Molecular Ecology, 6, 393–397.
Dumolin-Lapègue S, Pemonge M-H, Petit RJ (1998) Associationbetween chloroplast and mitochondrial ligneages in oaks.Molecular Biology and Evolution, 15, 1321–1331.
El Mousadik A, Petit RJ (1996) Chloroplast DNA phylogeographyof the argan tree of Morocco. Molecular Ecology, 5, 547–555.
Elena-Rosselló JA, Cabrera E (1996) Isozyme variation in naturalpopulations of cork-oak (Quercus suber L.). Population structure,diversity, differentiation and gene flow. Silvae Genetica, 45, 229–235.
Elena-Rosselló JA, Lumaret R, Cabrera E, Michaud H (1992)Evidence for hybridization between sympatric holm-oak andcork-oak in Spain based on diagnostic enzyme markers.Vegetatio, 99–100, 115–118.
Ennos RA (1994) Estimating the relative rates of pollen and seedmigration among plant populations. Heredity, 72, 250–259.
Gil Sánchez L, Jiménez Sancho MP, Díaz-Fernández PM (1996)Quercus complex in Spain: an overview of its present state. In:Inter- and Intraspecific Variation in European Oaks: EvolutionaryImplications and Practical Consequences (eds Kremer A, Muhs H-J),pp. 295–318. Office for Official Publications of the EuropeanCommunities, Luxembourg.
Jiménez P, Agúndez D, Alía R, Gil L (1999) Genetic variation incentral and marginal populations of Quercus suber L. SilvaeGenetica, 48, 278–284.
Lamb HF, Eicher U, Switsur VR (1989) An 18 000 years record ofvegetation, lake-level and climatic change from Tigalmamine,Middle Atlas, Morocco. Journal of Biogeography, 16, 65–74.
M’Hirit O (1996) Le chêne liège au Maroc. Inventaire, caractérisationet conservation des ressources génétiques. AEFCS, Rabat-Chellah,Maroc.
Manos PS, Doyle JJ, Nixon KC (1999) Phylogeny, biogeography,and processes of molecular differentiation in Quercus subgenusQuercus (Fagaceae). Molecular Phylogenetics and Evolution, 12,333–349.
Neigel JE, Avise JC (1986) Phylogenetic relationships of mitochon-drial DNA under various demographic models of speciation.In: Evolutionary Processes and Theory (eds Karlin S, Nevo E),pp. 515–534. Academic Press, New York.
Petit RJ, Csaikl U, Bordacs S et al. (2001b) Chloroplast DNA varia-tion in European white oaks: phylogeography and patterns ofdiversity based on data from over 2, 600 populations. ForestEcology and Management, in press.
Petit RJ, Kremer A, Wagner DB (1993a) Geographic structure ofchloroplast DNA polymorphisms in European oaks. Theoreticaland Applied Genetics, 87, 122–128.
Petit RJ, Kremer A, Wagner DB (1993b) Finite island model fororganelle and nuclear genes in plants. Heredity, 71, 630–641.
Petit RJ, Latouche-Hallé C, Pemonge M-H, Kremer A (2001a)Chloroplast DNA variation of oaks in France and the influenceof forest fragmentation on genetic diversity. Forest Ecology andManagement, in press.
Petit RJ, Pineau E, Demesure B et al. (1997) Chloroplast DNA foot-prints of postglacial recolonization by oaks. Proceedings of theNational Academy of Sciences of the USA, 94, 9996–10001.
Pons O, Petit RJ (1995) Estimation, variance and optimal samplingof gene diversity. I. Haploid locus. Theoretical and AppliedGenetics, 90, 462–470.
Potts BM, Reid JB (1988) Hybridization as a dispersal mechanism.Evolution, 42, 1245–1255.
Reille M (1977) Contribution pollenanalytique à l’histoire Holocènede la végétation des montagnes du Rif (Maroc septentrional).Xe Congrès INQUA, Birmingham, AFEQ, 50 (Suppl.), 53–76.
Reille M, Andrieu V, de Beaulieu J-L (1996) Les grands traits del’histoire de la végétation des montagnes méditerranéennesoccidentales. Ecologie, 27, 153–169.
MEC1330.fm Page 2011 Monday, July 23, 2001 5:32 PM

2012 N . B E L A H B I B E T A L .
© 2001 Blackwell Science Ltd, Molecular Ecology, 10, 2003–2012
Rieseberg LH (1995) The role of hybridization in evolution: oldwine in new skins. American Journal of Botany, 82, 944–953.
Rieseberg LH, Brouillet L (1994) Are many plant species para-phyletic? Taxon, 43, 21–32.
Rieseberg LH, Soltis DE (1991) Phylogenetic consequences of cyto-plasmic gene flow in plants. Evolutionary Trends in Plants, 5, 65–84.
Sauvage C (1961) Recherches géobotaniques sur les subéraiesmarocaines. Série botanique, 21–22. Institut Cherifien, Rabat.
Schwarz O (1937) Monographie der Eichen Europas und des Mit-telmeergebietes. I. Textband. Dahlem bei Berlin, 200 p.
Schwarz O (1964) Quercus. In: Flora Europaea, Vol. I. (ed. Tuttin et al.),pp. 61–64. Cambridge University Press, Cambridge.
Taberlet P, Gielly L, Pautou G, Bouvet J (1991) Universal primersfor amplification of three non-coding regions of chloroplastDNA. Plant Molecular Biology, 17, 1105–1109.
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysisof phenotypic associations with haplotypes inferred fromrestriction endonuclease mapping and DNA sequence data. III.Cladogram estimation. Genetics, 132, 619–633.
Toumi L, Lumaret R (1998) Allozyme variation on cork oak(Quercus suber L.): the role of phylogeography and geneticintrogression by other Mediterranean oak species and humanactivities. Theoretical and Applied Genetics, 97, 647–656.
Varela MC, Eriksson G (1995) Mutipurpose gene conservationin Quercus suber — a Portuguese example. Silvae Genetica, 44,28–37.
Varela MC, Valdiviesso T (1996) Phenological phases of Quercussuber L. flowering. Forest Genetics, 3, 93–102.
Whittemore AT, Schaal BA (1991) Interspecific gene flow in oaks.Proceedings of the National Academy Sciences of the USA, 88, 2540–2544.
Zitan L (1995) Analyse des caractères morphologiques pour l’évaluationde la variabilité génétique du chêne liège (Quercus suber L.). Mémoirede 3ème cycle agronomie, IAV Hassan II, Rabat.
This research forms part of Nadia Belahbib’s state thesis on corkoak genetic resources in Morocco, and was conducted at the ForestGenetics and Tree Improvement Laboratory of INRA. AbdellahOuassou and Hassan Sbay share an interest in the conservation offorest genetic resources in Morocco. In France, Antoine Kremer hasdeveloped a long-term research programme on genetic variationin forest trees, which largely focuses on oaks. Rémy Petit has aspecial interest in cytoplasmic markers and their use to unravelancient processes, and in their application to conservation.
MEC1330.fm Page 2012 Monday, July 23, 2001 5:32 PM
Related Documents











