This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attached
copy is furnished to the author for internal non-commercial research
and education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling or
licensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of the
article (e.g. in Word or Tex form) to their personal website or
institutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies are
encouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Fowling during the Gravettian: the avifauna of Pavlov I, the Czech Republic
Zbigniew M. Bochenski a,*, Teresa Tomek a, Jaros1aw Wilczynski a, Jiri Svoboda b, Krzysztof Wertz a,Piotr Wojtal a
a Institute of Systematics and Evolution of Animals, Polish Academy of Sciences. Sławkowska 17, 31-016 Krakow, PolandbDepartment of Palaeolithic and Palaeoethnology, Institute of Archaeology, ASCR, 69201 Dolnı Vestonice 25, Czech Republic
a r t i c l e i n f o
Article history:
Received 20 May 2009
Received in revised form
5 August 2009
Accepted 11 August 2009
Keywords:
Bird remains
Gravettian
Upper Palaeolithic
Hunting
Broad spectrum revolution
Corvus corax
Lagopus lagopus
Lagopus muta
Tetrao tetrix
a b s t r a c t
This is the first article that describes in detail the bird remains from the Gravettian mega-site Pavlov I in
the Czech Republic. More than 1000 bird bones represent at least 19 taxa, of which the most numerous
are tetraonids including black grouse (Tetrao tetrix), willow grouse (Lagopus lagopus) and ptarmigan
(Lagopus muta), and ravens (Corvus corax). The archaeological and biological contexts indicate that most
birds were hunted by people in the vicinity of the site, possibly with the help of knotted nets made from
plant fibres. We suggest that ravens were killed while feeding on carcasses and/or food remains that may
have been disposed of intentionally, and feathers may have been used for arrow fletching. Human-
modified bones indicate that birds were used not only for food but also as raw material for tools and
decoration. Although mammals were certainly more important in the subsistence of the Gravettian
people, it is clear that birds played a role in their culture. The location of Pavlov I enlarges the explanatory
scope of the Broad Spectrum Revolution hypothesis to include higher latitudes north of the
Mediterranean.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
The open-air loess site of Pavlov I, situated in the Pavlovian Hills
in the region of Moravia, the Czech Republic, is one of the largest
and most important Upper Palaeolithic sites in Europe. It is part of
a large complex of Gravettian sites that extend from the village of
Dolnı Vestonice in the northwest to Pavlov in the southeast. The
site is located on a gentle slope between 190 and 200 m a.s.l.,
several dozen meters above the Dyje river. The 14C dates indicate
a relatively short time-span of occupation between 27 and 25 ka BP,
i.e. in the Evolved Pavlovian stage, an equivalent of the final Inter-
pleniglacial or OIS 3 (Svoboda, 2005). The great advantage of the
site is that it was excavated between 1952 and 1971 by one person,
Bohuslav Klıma, using a single research strategy of investigating the
area in squares measuring 1 or 4 m2.
An analysis of the environmental, archaeozoological, anthro-
pological, and archaeological records has been underway since the
1990s (Svoboda, 1994, 1997, 2005; Trinkaus et al., 2009). Both
pollen and charcoal analyses from the Gravettian cultural layers
show that the landscape was partly covered by woodland areas,
forming a parkland (arboreal pollen usually exceeds 50%), with
conifers such as pine, spruce and larch dominating, but accompa-
nied by deciduous trees such as willow and alder along the
riverbed. The sporadic occurrence of species such as oak, beach, and
yew is also indicated (Damblon, 1997; Mason et al., 1994; Opravil,
1994; Rybnıckova and Rybnıcek, 1991; Svobodova, 1991a,b). Large
meadow-steppe mixed with conifer patches, and subalpine grass
communities on the rocky summits were also present. Studies of
the vertebrate fauna confirm the environmental reconstructions
based on the plant remains (Musil, 1955, 1959, 2005; Nadachowski,
2005).
Pavlov I has been interpreted as a typical example of a large
settlement occupied for a long period, and used repeatedly by
hunters-gatherers, where the settlement levels accumulated as
a thick complex of layers, creating local palimpsests (Novak, 2005:
70). An abundance of artefacts indicate a very intensive occupation
of the site. For instance, the site yielded more than 200,000 stone
artefacts (Verpoorte, 2005), 1000 ceramic fragments (Soffer and
* Corresponding author.
E-mail addresses: [email protected] (Z.M. Bochenski), tomek@
isez.pan.krakow.pl (T. Tomek), [email protected] (J. Wilczynski),
[email protected] (J. Svoboda), [email protected] (K. Wertz), wojtal@
isez.pan.krakow.pl (P. Wojtal).
Contents lists available at ScienceDirect
Journal of Archaeological Science
journal homepage: ht tp: / /www.elsevier .com/locate/ jas
0305-4403/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jas.2009.08.002
Journal of Archaeological Science 36 (2009) 2655–2665

Author's personal copy
Vandiver, 2005) and more than 600 bone, antler and ivory tools
(Bruhl, 2005). Current fieldwork focuses on the smaller ‘‘satellite’’
camps in the vicinity, which are characterised by having a more
restricted fauna (Pavlov VI; Svoboda et al., 2009).
The site of Pavlov I also yielded a large number of animal
remains (Musil, 1955, 1959, 1994, 1997). At present, more precise
data on faunal remains are available only for the material excavated
between 1952 and 1956 in the southeastern part of Pavlov I. This
area covers about a half the sitewhere some 46,000mammal bones
were found. The most numerous remains belonged to game
species, including hare (Lepus sp.), reindeer (Rangifer tarandus),
mammoth (Mammuthus primigenius), and horse (Equus sp.). A
characteristic feature of this archaeological site is the large number
of carnivores. These comprise nearly 40% of all mammal remains –
the most numerous being wolf (Canis lupus), red fox (Vulpes vulpes)
and Arctic fox (Alopex lagopus). Other less commonmammals found
at the site include the wolverine (Gulo gulo), cave lion (Panthera
spelaea), cave bear (Ursus spelaeus) and brown bear (Ursus arctos)
(Musil, 2005). The southeastern part of Pavlov I also yielded about
two dozen remains of small mammals including rodents (Nada-
chowski, 2005). The presence of all parts of their skeletons indi-
cated that most of the hunted herbivores and carnivores were
brought to the site and dismembered there. However, the over-
representation of foot bones of both cave lion and cave bear indi-
cates that their skins rather than whole carcasses were brought to
the site.
Mammal bones modified by people make up less that 2% of all
the remains. Of the 16,000 bones studied from the 1954 to 1956
excavations, 299 fragments had some traces of human activity such
as cut and burn marks. Human related marks were found on bones
representing all size categories: relatively small species (hare and
fox), medium size species (wolf and reindeer) and large mammals
(horse andmammoth). It is noteworthy that the marks observed on
the bones indicate that the Gravettian people hunted large and
medium-sizemammals for their skin andmeat (Wojtal et al., 2005).
Zooarchaeological studies in Pavlov I have so far focused on
mammals, with birds generally being treated superficially. Musil
(1958,1959) states that birds constitute some 8–9% of the total faunal
assemblage, and include raven (Corvus corax), black grouse (Lyrurus
tetrix), and partridge (Alectoris sp.) (see also Musil, 1994, 1997, 2003,
2005;Wojtal et al., 2005).Our aimhere is tofill the gap in information
about the birds and provide insights into Gravettian use of birds.
2. Material and methods
This study includes all avian remains found in the entire area of
Pavlov I during the excavations undertaken between 1952 and 1971.
Althoughwedonotknowwhether thematerialwas sieved, the large
number of very small objects recovered (e.g. isolated teeth of small
mammals, some fifteen hundred phalanges of hare and fox) indicate
that the excavations were carried out carefully and that the remains
available for study are representative. Due to the lack of precise data
we cannot exclude the possibility that the bird remains come from
two or more facies. Therefore they are interpreted here as a single
assemblage. The material belongs to the Moravian Museum (Brno,
Czech Republic) and is housed in Budisov castle.
The bird bones were identified by Teresa Tomek and Krzysztof
Wertz, based on comparisons with modern skeletons in the
collection of the Institute of Systematics and Evolution of Animals,
Polish Academy of Sciences, and with the help of available manuals
(Erbersdobler, 1968; Kraft, 1972; Tomek and Bochenski, 2000). For
practical reasons, in the case of body part representation we
combined the few taxa of the Tetraonidae family (black grouse
Tetrao tetrix, willow grouse Lagopus lagopus, ptarmigan Lagopus
muta, Lagopus sp. and Tetraonidae indet.) into one group
‘‘tetraonids’’, this being justified on morphological and ecological
grounds (Cramp and Simmons, 1980; Erbersdobler, 1968; Kraft,
1972). However, we analysed the remains of the raven (Corvus
corax) separately from other species of the Corvid family present at
Pavlov I, i.e. magpie (Pica pica), jay (Garrulus glandarius), chough
(Pyrrhocorax pyrrhocorax), and jackdaw (Corvus monedula) because
ravens were easily distinguished by their large size. We examined
the bone assemblage for element frequency, cut marks, traces of
burning, perforation of the distal humerus due to the overextension
of the elbow joint (Laroulandie, 2005b; Laroulandie et al., 2008),
predator activity marks including perforations made with beaks or
claws (Bochenski et al., 2009), root etching and weathering
(Bochenski and Tomek, 1997).
The results are presented in three different ways – as Number of
Identified Specimens (NISP), Minimum Number of Individuals
(MNI), and Minimum Number of Elements (MNE). The MNE was
calculated for each major type of bone only, i.e., omitting small
elements such as phalanges or vertebra. The MNE is the sum of
complete bones (left and right) and proximal (left and right) or
distal (left and right) parts – whichever is more numerous (Lyman,
1994, 2008).
The ratio of wing-to-leg elements was calculated as the number
of wing fragments (humerusþ ulnaþ carpometacarpus) divided by
the sum of wing and leg fragments (femurþ tibiotarsusþ tarsome-
tatarsus), and then expressed as a percentage (Ericson, 1987;
Livingston, 1989). The calculations were based on NISP and MNE.
A Chi-square test was used to evaluate the significance of differ-
ences in the proportion of wing and leg elements.
3. Results
The avian remains from Pavlov I include 1045 fragments rep-
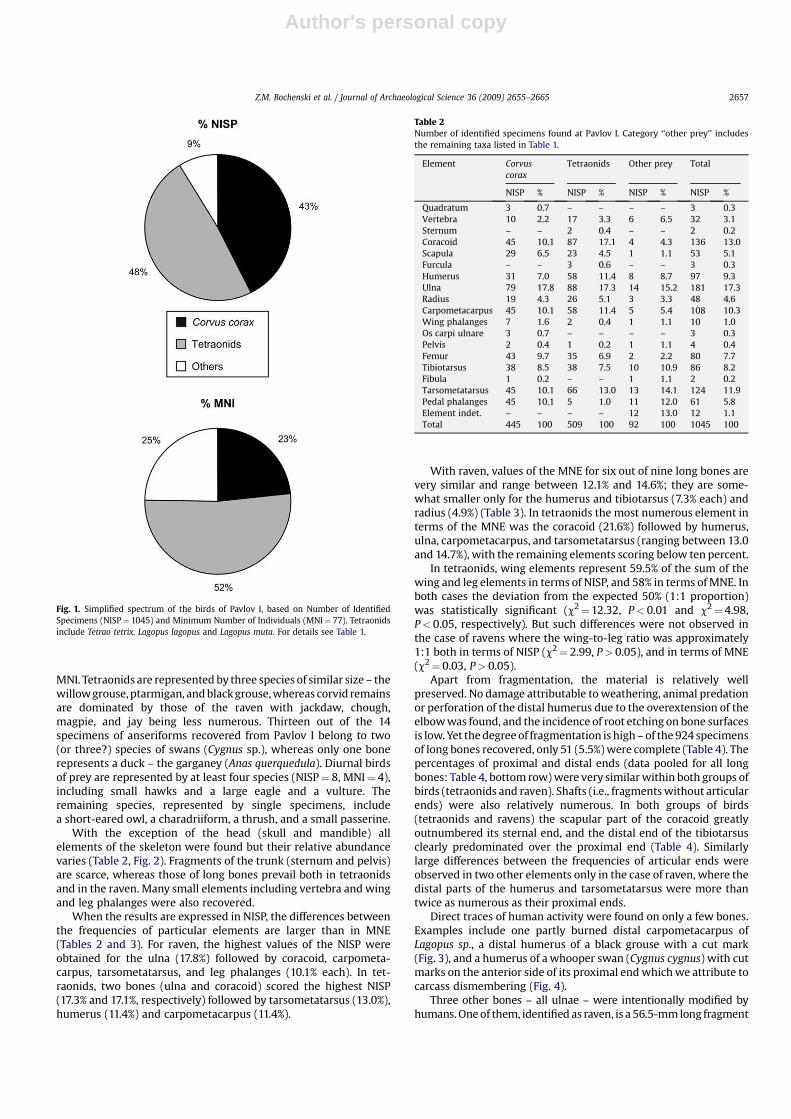
resenting at least 19 taxa (Table 1, Fig. 1). Members of two families –
Tetraonidae and Corvidae – clearly predominate, comprising
respectively 48.8 and 45.8% of the total NISP, or 52.0 and 29.9% of the
Table 1
Avian species found at Pavlov I.
Taxon NISP MNI
N % N %
1 Cygnus cygnus Whooper swan 2 0.2 1 1.3
Cygnus cygnus/Cygnus olor Whooper/Mute swan 5 0.5 1 1.3
2 Cygnus columbianus Bewick’s swan 6 0.6 2 2.6
3 Anas querquedula Garganey 1 0.1 1 1.3
4 Asio flammeus Short-eared owl 1 0.1 1 1.3
5 Haliaetus albicilla White-tailed eagle 2 0.2 1 1.3
6 Gyps fulvus Griffon vulture 2 0.2 1 1.3
Accipitridae indet. 1 0.1 – 0.0
7 Falco cf. tinnunculus cf. Kestrel 1 0.1 1 1.3
8 Falco cf. peregrinus cf. Peregrine 2 0.2 1 1.3
9 Lagopus lagopus Willow grouse 312 29.9 31 40.3
10 Lagopus muta Ptarmigan 21 2.0 4 5.2
Lagopus sp. 58 5.6 – 0.0
11 Tetrao tetrix Black grouse 62 5.9 5 6.5
Tetraonidae indet. (Tetrao tetrix/Lagopus sp.) 56 5.4 – 0.0
12 Perdix perdix Partridge 1 0.1 1 1.3
13 Charadriiformes indet. (size Larus) 1 0.1 1 1.3
14 Turdus sp. Thrush indet. 1 0.1 1 1.3
15 Passeriformes indet. (size Emberiza) Perching birds 1 0.1 1 1.3
16 cf. Pica pica cf. Magpie 1 0.1 – 0.0
Pica pica/Garrulus glandarius Magpie/Jay 1 0.1 1 1.3
17 Pyrrhocorax pyrrhocorax Chough 7 0.7 1 1.3
Pyrrhocorax/Corvus monedula Chough/Jackdow 9 0.9 1 1.3
18 Corvus monedula Jackdow 12 1.1 2 2.6
small Corvidae 3 0.3 – 0.0
19 Corvus corax Raven 445 42.6 18 23.4
Total identifiable 1014 97.0 77 100
Aves indet. 31 3.0
Total 1045 77 100
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–26652656
Author's personal copy
MNI. Tetraonids are represented by three species of similar size – the
willowgrouse,ptarmigan, andblackgrouse,whereas corvid remains
are dominated by those of the raven with jackdaw, chough,
magpie, and jay being less numerous. Thirteen out of the 14
specimens of anseriforms recovered from Pavlov I belong to two
(or three?) species of swans (Cygnus sp.), whereas only one bone
represents a duck – the garganey (Anas querquedula). Diurnal birds
of prey are represented by at least four species (NISP¼ 8, MNI¼ 4),
including small hawks and a large eagle and a vulture. The
remaining species, represented by single specimens, include
a short-eared owl, a charadriiform, a thrush, and a small passerine.
With the exception of the head (skull and mandible) all
elements of the skeleton were found but their relative abundance
varies (Table 2, Fig. 2). Fragments of the trunk (sternum and pelvis)
are scarce, whereas those of long bones prevail both in tetraonids
and in the raven. Many small elements including vertebra and wing
and leg phalanges were also recovered.
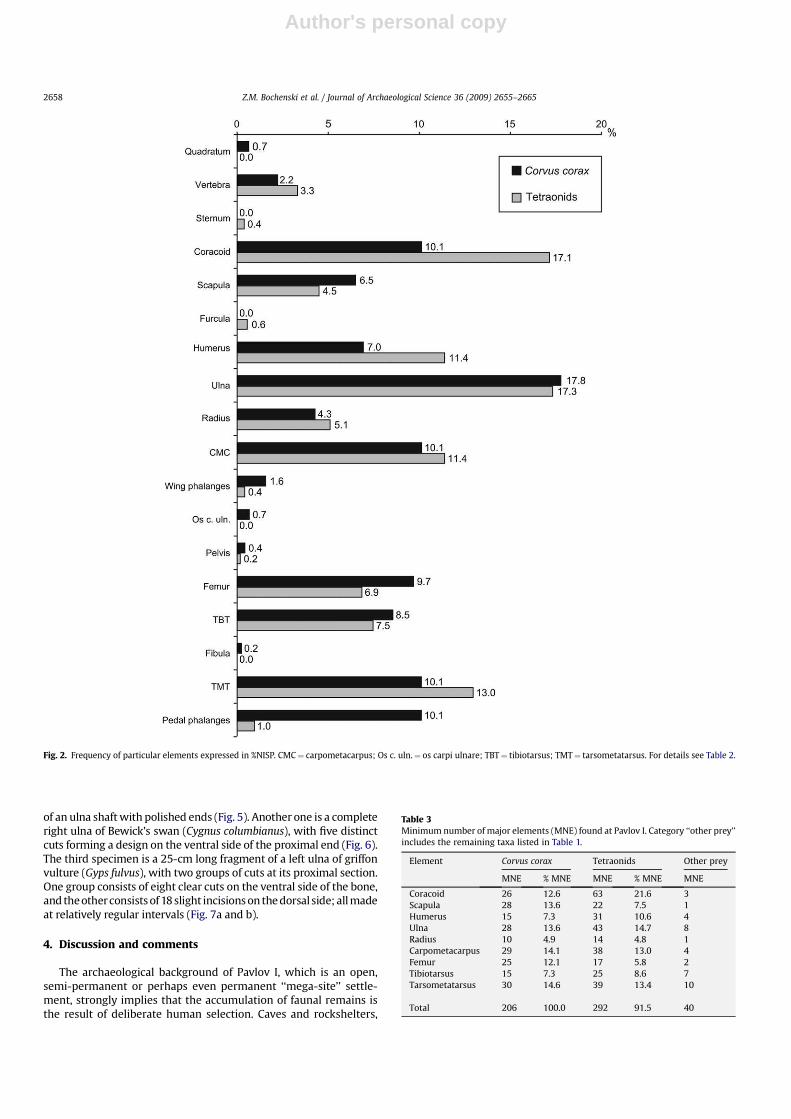
When the results are expressed in NISP, the differences between
the frequencies of particular elements are larger than in MNE
(Tables 2 and 3). For raven, the highest values of the NISP were
obtained for the ulna (17.8%) followed by coracoid, carpometa-
carpus, tarsometatarsus, and leg phalanges (10.1% each). In tet-
raonids, two bones (ulna and coracoid) scored the highest NISP
(17.3% and 17.1%, respectively) followed by tarsometatarsus (13.0%),
humerus (11.4%) and carpometacarpus (11.4%).
With raven, values of the MNE for six out of nine long bones are
very similar and range between 12.1% and 14.6%; they are some-
what smaller only for the humerus and tibiotarsus (7.3% each) and
radius (4.9%) (Table 3). In tetraonids the most numerous element in
terms of the MNE was the coracoid (21.6%) followed by humerus,
ulna, carpometacarpus, and tarsometatarsus (ranging between 13.0
and 14.7%), with the remaining elements scoring below ten percent.
In tetraonids, wing elements represent 59.5% of the sum of the
wing and leg elements in terms of NISP, and 58% in terms ofMNE. In
both cases the deviation from the expected 50% (1:1 proportion)
was statistically significant (c2¼12.32, P< 0.01 and c2¼ 4.98,
P< 0.05, respectively). But such differences were not observed in
the case of ravens where the wing-to-leg ratio was approximately
1:1 both in terms of NISP (c2¼ 2.99, P> 0.05), and in terms of MNE
(c2¼ 0.03, P> 0.05).
Apart from fragmentation, the material is relatively well
preserved. No damage attributable toweathering, animal predation
or perforation of the distal humerus due to the overextension of the
elbowwas found, and the incidence of root etching on bone surfaces
is low. Yet thedegree of fragmentation is high–of the924 specimens
of long bones recovered, only 51 (5.5%)were complete (Table 4). The
percentages of proximal and distal ends (data pooled for all long
bones: Table 4, bottom row)were very similarwithin both groups of
birds (tetraonids and raven). Shafts (i.e., fragmentswithout articular
ends) were also relatively numerous. In both groups of birds
(tetraonids and ravens) the scapular part of the coracoid greatly
outnumbered its sternal end, and the distal end of the tibiotarsus
clearly predominated over the proximal end (Table 4). Similarly
large differences between the frequencies of articular ends were
observed in two other elements only in the case of raven, where the
distal parts of the humerus and tarsometatarsus were more than
twice as numerous as their proximal ends.
Direct traces of human activity were found on only a few bones.
Examples include one partly burned distal carpometacarpus of
Lagopus sp., a distal humerus of a black grouse with a cut mark
(Fig. 3), and a humerus of awhooper swan (Cygnus cygnus) with cut
marks on the anterior side of its proximal endwhichwe attribute to
carcass dismembering (Fig. 4).
Three other bones – all ulnae – were intentionally modified by
humans.Oneof them, identified as raven, is a 56.5-mmlong fragment
Fig. 1. Simplified spectrum of the birds of Pavlov I, based on Number of Identified
Specimens (NISP¼ 1045) and Minimum Number of Individuals (MNI¼ 77). Tetraonids
include Tetrao tetrix, Lagopus lagopus and Lagopus muta. For details see Table 1.
Table 2
Number of identified specimens found at Pavlov I. Category ‘‘other prey’’ includes
the remaining taxa listed in Table 1.
Element Corvus
corax
Tetraonids Other prey Total
NISP % NISP % NISP % NISP %
Quadratum 3 0.7 – – – – 3 0.3
Vertebra 10 2.2 17 3.3 6 6.5 32 3.1
Sternum – – 2 0.4 – – 2 0.2
Coracoid 45 10.1 87 17.1 4 4.3 136 13.0
Scapula 29 6.5 23 4.5 1 1.1 53 5.1
Furcula – – 3 0.6 – – 3 0.3
Humerus 31 7.0 58 11.4 8 8.7 97 9.3
Ulna 79 17.8 88 17.3 14 15.2 181 17.3
Radius 19 4.3 26 5.1 3 3.3 48 4.6
Carpometacarpus 45 10.1 58 11.4 5 5.4 108 10.3
Wing phalanges 7 1.6 2 0.4 1 1.1 10 1.0
Os carpi ulnare 3 0.7 – – – – 3 0.3
Pelvis 2 0.4 1 0.2 1 1.1 4 0.4
Femur 43 9.7 35 6.9 2 2.2 80 7.7
Tibiotarsus 38 8.5 38 7.5 10 10.9 86 8.2
Fibula 1 0.2 – – 1 1.1 2 0.2
Tarsometatarsus 45 10.1 66 13.0 13 14.1 124 11.9
Pedal phalanges 45 10.1 5 1.0 11 12.0 61 5.8
Element indet. – – – – 12 13.0 12 1.1
Total 445 100 509 100 92 100 1045 100
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–2665 2657
Author's personal copy
of an ulna shaftwith polished ends (Fig. 5). Another one is a complete
right ulna of Bewick’s swan (Cygnus columbianus), with five distinct
cuts forming a design on the ventral side of the proximal end (Fig. 6).
The third specimen is a 25-cm long fragment of a left ulna of griffon
vulture (Gyps fulvus), with two groups of cuts at its proximal section.
One group consists of eight clear cuts on the ventral side of the bone,
and theotherconsistsof18slight incisionson thedorsal side; allmade
at relatively regular intervals (Fig. 7a and b).
4. Discussion and comments
The archaeological background of Pavlov I, which is an open,
semi-permanent or perhaps even permanent ‘‘mega-site’’ settle-
ment, strongly implies that the accumulation of faunal remains is
the result of deliberate human selection. Caves and rockshelters,
Fig. 2. Frequency of particular elements expressed in %NISP. CMC¼ carpometacarpus; Os c. uln.¼ os carpi ulnare; TBT¼ tibiotarsus; TMT¼ tarsometatarsus. For details see Table 2.
Table 3
Minimum number of major elements (MNE) found at Pavlov I. Category ‘‘other prey’’
includes the remaining taxa listed in Table 1.
Element Corvus corax Tetraonids Other prey
MNE % MNE MNE % MNE MNE
Coracoid 26 12.6 63 21.6 3
Scapula 28 13.6 22 7.5 1
Humerus 15 7.3 31 10.6 4
Ulna 28 13.6 43 14.7 8
Radius 10 4.9 14 4.8 1
Carpometacarpus 29 14.1 38 13.0 4
Femur 25 12.1 17 5.8 2
Tibiotarsus 15 7.3 25 8.6 7
Tarsometatarsus 30 14.6 39 13.4 10
Total 206 100.0 292 91.5 40
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–26652658
Author's personal copy
Fig.3.Distal
humerusoftheblack
grouse
(Tetraotetrix)
with
acu
tmark
on
the
poste
riorsid
e.
Table 4
Fragmentation of long bones found at Pavlov I. Category ‘‘other prey’’ includes the remaining taxa listed in Table 1.
Element Corvus corax Tetraonids Other prey Total
N Whole
bone (%)
Proximal
part (%)
Distal part
(%)
Shaft
(%)
N Whole
bone (%)
Proximal
part (%)
Distal part
(%)
Shaft
(%)
N Whole
bone (%)
Proximal
part (%)
Distal part
(%)
Shaft
(%)
N Whole
bone (%)
Proximal
part (%)
Distal part
(%)
Shaft
(%)
Coracoid 45 13.3 51.1 22.2 13.3 87 5.7 66.7 12.6 14.9 4 50.0 25.0 25.0 – 136 9.6 60.3 16.2 14.0
Scapula 29 – 96.6 – 3.4 23 – 100 – – 1 – 100 – – 53 – 98.1 0.0 1.9
Humerus 31 – 19.4 48.4 32.3 58 – 37.9 53.4 8.6 8 – 50.0 37.5 12.5 97 – 33.0 50.5 16.5
Ulna 79 – 19.0 35.4 45.6 88 2.3 47.7 39.8 10.2 14 14.3 21.4 42.9 21.4 181 2.2 33.1 38.1 26.5
Radius 19 – 26.3 42.1 31.6 26 – 53.8 34.6 11.5 3 – – 33.3 66.7 48 – 39.6 37.5 22.9
Carpometacarpus 45 11.1 40.0 24.4 24.4 58 19.0 46.6 25.9 8.6 5 20.0 60.0 20.0 – 108 15.7 44.4 25.0 14.8
Femur 43 7.0 32.6 41.9 18.6 35 2.9 31.4 45.7 20.0 2 – 50.0 50.0 – 80 5.0 32.5 43.8 18.8
Tibiotarsus 38 – 10.5 39.5 50.0 38 – 15.8 65.8 18.4 10 – 10.0 70.0 20.0 86 – 12.8 54.7 32.6
Tarsometatarsus 45 8.9 20.0 57.8 13.3 66 13.6 28.8 45.5 12.1 13 – 23.1 76.9 – 124 10.5 25.0 53.2 11.3
Element indet. – – – – – – – – – – 11 – – – 100 11 – – 0.0 100
Total NISP 374 4.8 32.6 35.0 27.5 479 5.8 46.3 35.9 11.9 71 7.0 23.9 42.3 26.8 924 5.5 39.1 36.0 19.4
Fig.4.Proxim
alhumerusofthewhoopersw
an(Cygn
uscygn
us)
with
cutmark
sonthe
anterio
rsid
e.
Z.M
.Boch
enski
etal./JournalofArch
aeo
logica
lScien
ce36(2009)2655–2665
2659

Author's personal copy
especially those occupied seasonally by people, are more likely to
have mixed deposits of human debris, non-human prey remains,
and bones derived from animals dying naturally. We would expect
to find numerous remains of rodents and small birds in Pavlov I if
the site had been inhabited also by owls, but this is not the case.
Their absence cannot be attributed to poor recovery because many
very small phalanges of hare and fox are present. Theoretically, the
relatively large birds recovered from the deposits (tetraonids and
ravens) could be food remains of a large predator such as an eagle
or eagle owl, but it is very unlikely that a raptor would chose to live
within a large human settlement and even less likely that it would
not be quickly disturbed or killed by the human inhabitants.
Moreover, we did not notice characteristic perforations of certain
bones which large raptors often make with their beaks and claws
(Bochenski et al., 2009). Mammalian predators can also be ruled out
as accumulating agents because they would destroy most of the
fragile bird bones or at least leave gnaw marks (Andrews, 1990;
Lyman, 1994). Water transport is not an option either because the
site is situated high above the river. However, the species compo-
sition of the birds recovered from the site and the presence of the
aforementioned bones modified by humans, confirm the conclu-
sion reached on the basis of mammalian fauna that the remains
were accumulated by humans (Svoboda, 2005).
The relatively lownumberof identifiedbird species (19) certainly
does not reflect the real diversity of avifauna in the habitat. More-
over, the relative abundance of particular taxa in the assemblage
differs considerable from any known modern avifauna. The over-
representation of one ormore groups of birds (at Pavlov I, tetraonids
and ravens made up more than 90% of the total NISP) is a typical
feature of an assemblage deposited by a selective predator – in this
case humans. In Europe there are many examples of assemblages
attributable to human origin in which one or more bird taxa
predominate. These usually include galliforms (Alectoris, Lagopus,
Tetrao), ducks, bustards, or snowy owls, depending on the locally
available prey (Cassoli and Tagliacozzo, 1997; Koumouzelis et al.,
2001; Laroulandie, 1998, 2003, 2005a; Tagliacozzo and Gala, 2002;
Tyrberg, 1998). Nearby Late Pleistocene sites that contain large
accumulations of tetraonids include Mamutowa Cave, Deszczowa
Cave, Ob1azowa Cave, Krucza Ska1a and Komarowa Cave where
tetraonids account for 40–70%of all avian remains (Bochenski,1981;
Bochenski and Tomek, 2004; Cyrek et al., 2000; Tomek and
Bochenski, 2005; Tomek et al., 2003).
According to Mourer-Chauvire (1983), humeri and femora are
the most numerous elements in human-derived assemblages of
bird bones, whereas in avian derived assemblages tarsometatarsi
and carpometacarpi prevail in middle size prey (e.g. Lagopus), and
tarsometatarsi and coracoids in large prey (e.g. Tetrao). However, as
shown by Bochenski (2005), the problem is more complicated and
the predominance of the humerus is not specific only to materials
deposited by humans. Avian remains from Pavlov I are puzzling
because while on the one hand various elements are well repre-
sented which makes it difficult to apply the criterion of human
versus non-human origin of thematerial, on the other hand, femora
of ravens and tetraonids are somewhat less numerous than several
of the other elements which does not agree with the assumption
that the assemblage includes human food remains. Yet the
contextual data indicate that the birds – in any case tetraonids –
were probably eaten by people. A similar situation where in an
obviously human-derived assemblage femora were less numerous
Fig. 5. Ulna shaft of the raven (Corvus corax), with polished ends. The fragment is
65.5 mm long.
Fig. 6. Complete right ulna of the bewick’s swan (Cygnus columbianus), with five
distinct cuts forming a design on the ventral side of the proximal end.
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–26652660

Author's personal copy
than other elements was also observed in other archaeological sites
(Guminski, 2005; Steadman et al., 2002), which indicates that the
criterion must be used cautiously. A possible explanation for this
phenomenon has been suggested by Higgins (1999) who claims
that femora are most subjected to destruction in natural as well as
cultural depositions. This is on the one hand because femora
contain large quantities of marrow desirable for human
consumption (Higgins, 1999: 1453–4), and on the other hand, the
bones are more fragile due to their relatively thin cortical walls and
low compactness. However, we are unsure whether the latter
explanation is valid also for galliforms whose leg bones seem to be
particularly sturdy.
Another method often used to distinguish human food remains
from birds which have been subject only to natural decomposition
factors is the ratio of the wing-to-leg elements (Ericson, 1987).
According to Ericson (1987) in assemblages accumulated by people,
bones from the meat-rich leg bones prevail. Although the theory
has many supporters, it has also been partly criticized (Bochenski,
2005; Livingston, 1989; Serjeantson et al., 1993). Recent studies
have shown that the ratio may also be useful in distinguishing
pellet and uneaten food remains of owls and diurnal birds of prey.
Wing bones greatly predominate in uneaten food remains of many
diurnal birds of prey (Bochenski et al., 1999; Bochenski and Torn-
berg, 2003; Bramwell et al., 1987; Laroulandie, 2000, 2002; Mli-
kovsky, 1996) whereas their occurrence in owl and diurnal raptor
pellets is much lower or they are absent (Bochenski et al., 1993,
1998; Bochenski and Nekrasov, 2001; Bochenski and Tomek, 1994).
However, it may occasionally happen that leg elements prevail in
assemblages deposited by raptors as in the case of pellet material
derived from the imperial eagle (Bochenski et al., 1999; Bochenski,
2005) or that wing bones predominate in assemblages deposited by
humans (Guminski, 2005; Livingston, 1989; Serjeantson et al.,
1993). Our results, showing predominance of wing elements in
tetraonids and a nearly equal ratio of wing and leg bones in ravens,
confirm the conclusion reached by Serjeantson et al. (1993) that
anatomical distribution alone cannot provide an answer to the
question of who accumulated the bones – humans or animals.
Although it is an important factor in taphonomic analysis, other
data including the archaeological context may have priority. It is
also true that it is better to use a combination of many criteria to
successfully attribute analysed material to a particular predator or
a group of predators (Bochenski, 2005).
The overrepresentation of wing elements in archaeological sites
is a well-known phenomenon which cannot be explained by one
universal theory (Bovy, 2002; Guminski, 2005). Various factors
including cultural and behavioural as well as post-depositional
modifications play a role in producing this pattern, and it is difficult
to pinpoint the main reason in the case of tetraonid remains at
Pavlov I. While it is very likely that tetraonids and possibly also
ravens were eaten by the Gravettian people, the birds were used
also for other purposes. The few artefacts made from avian bone at
the site are direct evidence that birds were not just a source of food.
Wing bones, especially those of large taxa, were often selected for
tool-making in prehistory (d’Errico et al., 2003; Gal, 2005; Lar-
oulandie, 2003; Wijngaarden-Bakker, 1997), which was also the
case at Pavlov I where all three bones decorated by humans
belonged to large birds. It is also possible that wing feathers were
used as decoration or for utilitarian purposes. However, direct
evidence for the use of feathers in archaeological sites is hard to
obtain and therefore it often remains only a possible option (Bovy,
2002; Mulkeen and O’Connor, 1997; Reichstein and Pieper, 1986;
Reisch, 1976).
The small number of bones with human-induced modifications
is not unusual in materials attributable to human origin. Assem-
blages are known with high frequencies of cut marks and/or traces
of burning (Cassoli and Tagliacozzo, 1997; Laroulandie, 2005a;
Tagliacozzo and Gala, 2002; Reisch, 1976), but in other assemblages
such evidence is scarce even when the sites are unequivocally
cultural in origin (Alhaique et al., 2004; Steadman et al., 2002). In
comparison to large mammals, birds are relatively easy to
Fig. 7. A 25-cm long fragment of a left ulna of the griffon vulture (Gyps fulvus), with two groups of cuts at its proximal section. One group consists of 18 light cuts on the dorsal side
of the bone (a), and the other consists of eight clear cuts on the ventral side (b).
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–2665 2661

Author's personal copy
disarticulate and one can do it with one’s bare hands without using
any tools (Steadman et al., 2002). Such a procedure may result in
characteristic perforation of the distal humerus and damage to the
proximal ulna due to overextension of the elbow (Laroulandie,
2005b; Laroulandie et al., 2008) but no such damage was noted at
Pavlov I. One would expect to recover similar number of scapulae
and coracoids because the two bones are firmly articulated and
tend to stay together when bird carcasses are pulled apart by hand.
Yet, the assemblage shows a paucity of scapulae relative to cora-
coids whichmay be due to the fact that the thin blade of scapulae is
easily broken and the remaining articular part is small and less
likely to be recovered.
Skeletal representation for themain prey species, tetraonids and
ravens, indicates the presence of nearly all major and many minor
anatomical elements which suggests that whole carcasses were
brought to the site and subsequently processed there. The only
missing part of the body is the head (beak, brain case and
mandible). However, the presence of three quadrates of the raven
suggests that the birds were not decapitated outside the site. The
absence of heads in the material may be due to their fragility and
tendency to disintegrate in pre- and post-depositional processes
such as human trampling and diagenesis. The scarcity or lack of
elements of the head and neck has also been observed at other
human-derived sites (Steadman et al., 2002), indirectly supporting
our suggestions.
The high degree of fragmentation of the bones and over-
representation of shafts without articular ends may reflect the
consumption of the ends of the bones (Laroulandie, 2005b;
Steadman et al., 2002) or post-depositional damage resulting from
the unique osteological characters of avian bones (Higgins, 1999). It
is likely that somedamage at Pavlov I is due to tramplingbecause the
site was occupied intensively. The overrepresentation of some
portions of certain bonesmay be best explained by the properties of
thebones. For instance, the sternal endof the coracoid is thin and the
proximal end of the tibiotarsus also consists of some thin crests – in
both cases those parts of the bones may be easily damaged. As
a consequence, scapular parts of coracoids clearly predominate in
many cultural and non-cultural assemblages (Bochenski, 2005;
Bochenski et al., 1993, 1997, 1998; Steadman et al., 2002).
Members of the Tetraonidae family are ground-dwelling, weak
flyers, and relatively heavy with bodies weighing between 0.4 and
1.4 kg. This makes them vulnerable to predation. They have always
been a common source of food for various predators including
birds, mammals, and people (Boev, 1997; Laroulandie, 2005a;
Stewart, 2007; Tyrberg, 1995) and it seems that Pavlov I is no
exception. Although direct traces attributable to food processing
are very scarce on bones from the site, they include one partly
burned wing bone of Lagopus which may indicate cooking for
consumption.
The large number of ravens at Pavlov I is also interesting. Their
body weight is comparable to that of tetraonids (0.99–1.56 kg) but
unlike some galliforms they fly very well and their flight feathers
are stiff and strong (Cramp and Perrins, 1994). The habitat around
the site (rocks and trees not far from the dwellings) was certainly
suitable for them to nest in, but they tend to keep their territories to
themselves and, depending on the season of the year, one can see
solitary birds, couples or small family flocks only. Therefore the
Gravettian people must have either hunted ravens over larger
territories outside the immediate vicinity of their site or, which
seemsmore likely, they took advantage of the fact that ravens often
feed on carrion. It is tempting to speculate that ravenswere killed at
the nearby mammoth bone accumulation of Dolnı Vestonice
I. Large carcasses of mammoths and their bones left by people must
have attracted many carrion-eating animals. Of course, remains of
smaller game that are abundant at Pavlov I also could have
attracted ravens. It is impossible to determine whether the people
lured ravens, and if so, whether they did it intentionally by
exposing carcasses or merely took advantage of the fact that the
birds were already nearby. It is noteworthy that no other Upper
Palaeolithic site contains somany remains of ravens (Stewart, 2007;
Tyrberg, 1998). Pavlov I is unique in this respect. Unfortunately
there is no direct evidencewhy the Gravettian people specialized in
killing ravens. They may have hunted the birds because they were
easy prey close to their dwellings, because ravens stole their food,
or perhaps because they needed the birds for raw materials (bones
or feathers). The lack of cut marks and traces of burning on raven
remains may suggest that the birds were used for purposes other
than food. The fragment of a raven ulna with polished edges (Fig. 5)
supports this hypothesis. It may have been made into a tube, either
for use as a drinking ‘‘straw’’, blowing or sniffing, or with both ends
plugged, as a container for some substance such as ochre (Wijn-
gaarden-Bakker, 1997). Ravens have symbolic meaning in many
cultures throughout the world (Goodwin, 1976) and therefore it is
possible that they played a similar role at Pavlov I during the Upper
Palaeolithic. This would agree with the archaeological context that
suggests symbolic activities and social rituals performed at the
Moravian mega-sites of Pavlov I and Dolnı Vestonice I (Musil, 1994;
Svoboda, 2005).
The remaining smaller species of corvids make up about 3.2% of
all bird remains (NISP¼ 33, MNI¼ 5) and are representedmainly by
jackdaws that are currently considered synanthropes, living near
human settlements, but they also tend to nest among rocks and in
the hollows of older trees (Cramp and Perrins, 1994). It is likely that
this group of birds also nested in the vicinity of Pavlov I. Jackdaws,
like ravens, could have been hunted but because their remains are
scarce, it is difficult to draw further conclusions.
An interesting feature of the analysed material is the relatively
large number (NISP¼ 13, MNI¼ 4) of swan remains (Cygnus sp.),
especially in comparison to the number of the remaining anseri-
forms (only one bone of a duck was found). The presence of swans
indicates a largewater body in the vicinity of the site which implies
other waterfowl must have been present as well. Yet, it seems that
of all anseriforms only the swans were of particular interest to the
Gravettian people, which can be deduced not only from their
relative abundance but also from the fact that one of the three
decorated bones belonged to a swan (Fig. 6). It is difficult to know
why, but large size is a possible explanation. It has already been
noted in many archaeological sites that large species were
preferred to smaller ones (Guminski, 2005; Laroulandie, 2003).
Another interesting feature of the material is the relatively large
variety of birds of prey. We include in this category all meat-eating
species – accipitrids and owls. Although their remains are scarce, at
least five different species are represented. The griffon vulture ulna
with cut marks (Fig. 7a and b) – one of the only four artefacts made
from bird bones at the site – suggests that raptors may have had
some special significance to the Gravettian people. This would
agree with the result of other studies on the mammalian remains
that show the use of carnivores by the Gravettian people in Central
Europe (Fladerer, 2001; Lipecki and Wojtal, 1998; Munzel, 2004;
West, 1996; Wojtal, 2000; Wojtal et al., 2005). The regularity of the
cut marks on the vulture ulna suggests decoration. Ravens are in
some ways similar to birds of prey because they also eat meat and
they are capable of actively killing smaller prey (Cramp and Perrins,
1994). Therefore, the Gravettian huntersmay have treated ravens as
birds of prey and attributed special meaning to them too, which
could explain the large number of raven remains at the site.
Small animals made an increasingly important contribution to
human diet in the Mediterranean Basin from the early Middle
Palaeolithic onward, especially just before the Neolithic –
a hypothesis called ‘‘Broad Spectrum Revolution’’ (Davis, 2005;
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–26652662

Author's personal copy
Flannery, 1969; Munro, 2004; Stiner, 2001; Stiner et al., 2000).
While small-game assemblages of the Middle Palaeolithic were
dominated by slow-moving tortoises, more agile small vertebrates
including lagomorphs and birds were common prey beginning
from the Upper Palaeolithic. The integration of relatively fast-
moving (and therefore more difficult to catch) small animals into
the diet is interpreted as a necessary consequence of overhunting
slow-moving and slow-reproducing resources such as tortoises.
The bird-rich material from Pavlov I supports the hypothesis. A
unique feature of Pavlov I is the species composition: besides gal-
liforms which were typically hunted throughout Europe, ravens
played an important role. Most of the data in support of the
hypothesis of the Broad Spectrum Revolution derive from sites
around the Mediterranean (Davis, 2005; Stiner, 2001). The location
of Pavlov I enlarges the geographical zone of the hypothesis to
include higher latitudes.
Although we are unsure how the birds were procured at Pavlov
I, their high numbers indicate that the Gravettian people caught
them efficiently. Fully ossified bones indicate that the birds were
adult when killed, thus the people did not rely on collecting young
birds from their nests – a relatively easy task. Hunting so many
birds without a trap or a good weaponwould be a time-consuming
and ineffective procedure. One possible clue to themethods used to
procure birds comes in the form of negative clay impressions of
fibre-based constructions (Adovasio et al., 1996, 1997, 2005). Ado-
vasio and colleagues have proved that the sites of Pavlov I and Dolnı
Vestonice I and II contain the world’s oldest evidence for the use of
plant fibres in the manufacture of textiles, basketry, and netting.
Knotted nets may have served as highly efficient animal traps,
supported indirectly by our results. Another possibility is that bows
were used to hunt birds and medium-size mammals such as lep-
orids and foxes. It is tempting to speculate that the stiff and strong
flight feathers of ravens were a source of arrow fletching. Because
hard evidence of bows and arrows before the Mesolithic is lacking
(Cattelain, 2006; Rosendahl et al., 2006), only indirect pathways
remain to address the question in the Upper Palaeolithic. The
anatomical analysis of ligament attachments and enthesopathies
on the upper limb bones (Churchill, 1994; Villotte, 2008) cannot be
used as an unequivocal and convincing argument. What remains is
an analysis of the potential lithic projectiles, with emphasis on their
form, size, and the typical impact scars on their tips (Knecht, 1997;
Plisson and Geneste, 1989; Villa et al., 2009). Given their extremely
small size, the geometric microliths of Pavlov are better candidates
for arrowpoints than some of the other European specimens, which
are generally interpreted as spearpoints. Yaroshevich (personal
communication) who studied impact scars on the Pavlov microliths
concluded that some of them may have been used as projectiles,
most probably as arrows.
5. Conclusions
This is one of the very few studies of avian remains from the
Gravettian, and certainly the most abundant in bird bones. The
archaeological context with extremely numerous artefacts and tens
of thousands of mammal remains indicate a very intensive occu-
pation of Pavlov I. The skewed taxonomical profile of the avian
assemblage (i.e. more than 90% of the NISP being either grouse or
raven), and bird bones modified by humans are clear suggestions
that birds also played a role in the culture and subsistence of the
Gravettian people.
A unique feature of Pavlov I is the relative abundance of ravens
whose bones had some utilitarian purposes (an ulna made into
a tube), and whose feathers may have been used for decoration or
arrow fletching. It is likely that ravens were killed while feeding on
carcasses or food remains that may have been disposed
intentionally.
Anatomical distribution of particular bones and their fragments
in an assemblage should be used cautiously when studying the
taphonomy of a site. Relying exclusively on one taphonomic feature
such as the wing-to-leg ratio or the predominance of femora may
be misleading. It is advisable to use as wide a range of factors as
possible, including the archaeological and biological contexts.
The numerous bird remains (tetraonids and possible also other
taxa) from Pavlov I that contributed to the diet of its human
inhabitants can be interpreted as supporting the so-called Broad
Spectrum Revolution hypothesis, according to which people in
Europe and the Near East started to hunt swift, small animals from
the Upper Palaeolithic onwards. Until now, most of the data in
support of the theory derive from sites around the Mediterranean.
Pavlov I, which is situated far to the north, enlarges the
geographical zone of the hypothesis. The site also provides a clue as
to how the swift, fast-moving small-game animals were hunted.
Knotted nets made from plant fibres, whose world’s oldest
evidence was found at Pavlov I, may have been used as efficient
traps for catching birds and small mammals.
Acknowledgments
We thank Martin Oliva (Anthropos Institute, Moravian Museum,
Brno) and Moravske Zemske Museum for allowing this study of
bird bones from the Pavlov I site stored in Budisov castle. We are
very grateful to Erika Gal, Veronique Laroulandie, Cecile Mourer-
Chauvire and Tommy Tyrberg for fruitful discussions on various
aspects of this paper and supplying us with relevant literature, and
to Gary Haynes and Simon Davis for helpful comments and cor-
recting the English language of the manuscript. Five anonymous
reviewers provided useful suggestions how to improve the original
manuscript. Our research was partly supported by the Polish
Ministry of Science and Higher Education (grants No. 2 P04C 08130
for years 2006–2009 awarded to P. Wojtal and No. 0903/P01/2006/
31 for years 2007–2009 awarded to J. Wilczynski).
References
Adovasio, J.M., Soffer, O., Klıma, B., 1996. Upper palaeolithic fibre technology:interlaced woven finds from Pavlov I, Czech Republic, c. 26,000 years ago.Antiquity 70, 526–534.
Adovasio, J.M., Hyland, D.C., Soffer, O., 1997. Textiles and cordage: a preliminaryassessment. In: Svoboda, J. (Ed.), Pavlov I – Northwest. The Upper PaleolithicBurial and Settlement Context. Dolnı Vestonice Studies, vol. 4, pp. 432–443.
Adovasio, J.M., Soffer, O., Hyland, D.C., 2005. Textiles and cordage. In: Svoboda, J.(Ed.), Pavlov I Southeast. A Window into the Gravettian Lifestyles. Dolnı Ves-tonice Studies, vol. 14, pp. 432–443.
Alhaique, F., Bisconti, M., Castiglioni, E., Cilli, C., Fasani, L., Giacobini, G., Grifoni, R.,Guerreschi, A., Iacopini, A., Malerba, G., Peretto, C., Recchi, A., Rocci Ris, A.,Ronchitelli, A., Rottoli, M., Thun Hohenstein, U., Tozzi, C., Visentini, P.,Wilkens, B., 2004. Animal resources and subsistence strategies. CollegiumAntropologicum 28, 23–40.
Andrews, P., 1990. Owls, Caves and Fossils. Predation, Preservation and Accumula-tion of Small Mammal Bones in Caves, with an Analysis of the Pleistocene CaveFaunas from Westbury-Sub-Mendib, Somerset. UK. Natural History MuseumPublications, London.
Bochenski, Z., 1981. Fossil remains of birds from Mamutowa Cave. Folia Quaternaria54, 1–24 (in Polish with English summary).
Bochenski, Z.M., 2005. Owls, diurnal raptors and humans: signatures on avianbones. In: O’Connor, T. (Ed.), Biosphere to Lithosphere. New Studies in Verte-brate Taphonomy. Oxbow Books, Oxford, pp. 31–45.
Bochenski, Z.M., Boev, Z., Mitev, I., Tomek, T., 1993. Patterns of bird bone frag-mentation in pellets of the Tawny Owl (Strix aluco) and the Eagle Owl (Bubobubo) and their taphonomic implications. Acta Zoologica Cracoviensia 36 (2),313–328.
Bochenski, Z.M., Huhtala, K., Jussila, P., Pulliainen, E., Tornberg, R., Tunkkari, P.S.,1998. Damage to bird bones in pellets of Gyrfalcon Falco rusticolus. Journal ofArchaeological Science 25, 425–433.
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–2665 2663

Author's personal copy
Bochenski, Z.M., Huhtala, K., Sulkava, S., Tornberg, R., 1999. Fragmentation andpreservation of bird bones in food remains of the Golden Eagle Aquila chrys-aetos. Archaeofauna 8, 31–39.
Bochenski, Z.M., Korovin, V.A., Nekrasov, A.E., Tomek, T., 1997. Fragmentation of birdbones in food remains of Imperial Eagles Aquila heliaca. International Journal ofOsteoarchaeology 7 (2), 165–171.
Bochenski, Z.M., Nekrasov, A.E., 2001. The taphonomy of Sub-Atlantic bird remainsfrom Bazhukovo III, Ural mountains, Russia. Acta Zoologica Cracoviensia 44 (2),93–106.
Bochenski, Z.M., Tomek, T., 1994. Pattern of bird bone fragmentation in pellets of thelong-eared owl Asio otus and its taphonomic implications. Acta Zoologica Cra-coviensia 37 (1), 177–190.
Bochenski, Z.M., Tomek, T., 1997. Preservation of bird bones: erosion versus diges-tion by owls. International Journal of Osteoarchaeology 7 (4), 372–387.
Bochenski, Z., Tomek, T., 2004. Bird remains from rock-shelter in the Krucza Ska1a(Central Poland). Acta Zoologica Cracoviensia 47 (1–2), 27–47.
Bochenski, Z.M., Tomek, T., Tornberg, R., Wertz, K., 2009. Distinguishing nonhumanpredation on birds: pattern of damage done by the white-tailed eagle Haliaetusalbicilla, with comments on the punctures made by the golden eagle Aquilachrysaetos. Journal of Archaeological Science 36, 122–129. doi:10.1016/j.jas.2008.07.018.
Bochenski, Z.M., Tornberg, R., 2003. Fragmentation and preservation of bird bonesin uneaten food remains of the Gyrfalcon Falco rusticolus. Journal of Archaeo-logical Science 30, 1665–1671.
Boev, Z., 1997. Wild galliform and gruiform birds (Avis, Galliformes and Gruiformes)in the archaeological record of Bulgaria. International Journal of Osteo-archaeology 7, 430–439.
Bovy, K.M., 2002. Differential avian skeletal part distribution: explaining theabundance of wings. Journal of Archaeological Science 29, 965–978.
Bramwell, D., Yalden, W., Yalden, P.E., 1987. Black grouse as the prey of the goldeneagle at an archaeological site. Journal of Archaeological Science 14, 195–200.
Bruhl, E., 2005. Bone, antler, and ivory tools. In: Svoboda, J. (Ed.), Pavlov I Southeast.A Window into the Gravettian Lifestyles. Dolnı Vestonice Studies, 14, pp.252–293.
Cassoli, P.F., Tagliacozzo, A., 1997. Butchering and cooking of birds in the Palaeolithicsite of Grotta Romanelli (Italy). International Journal of Osteoarchaeology 7,303–320.
Cattelain, P., 2006. Apparition et evolution de larc et des pointes de fleches dans laprehistoire europeene (Paleo-, Meso-, Neolithique). In: Bellintani, P. (Ed.), Cat-ene operative dellarco preistorico. Giunta della Provincia Autonoma di Trento,Trento, pp. 45–66.
Cramp, S., Perrins, C.M., 1994. The Birds of the Western Palearctic, vol. VIII. OxfordUniversity Press, Oxford, New York.
Cramp, S., Simmons, K.E.L., 1980. The Birds of the Western Palearctic, vol. II. OxfordUniversity Press, Oxford, London, New York.
Churchill, S.E., 1994. Human Upper Body Evolution in the Eurasian Later Pleistocene.PhD thesis, University of New Mexico, Albuquerque.
Cyrek, K., Nadachowski, A., Madeyska, T., Bochenski, Z., Tomek, T., Wojtal, P.,Miekina, B., Lipecki, G., Garapich, A., Rzebik-Kowalska, B., Stworzewicz, E.,Wolsan, M., Godawa, J., Ko�sciow, R., Fostowicz-Frelik, q., Szyndlar, Z., 2000.Excavation in the Deszczowa Cave (Kroczyckie Rocks, Czestochowa Upland,Central Poland). Folia Quaternaria 71, 5–84.
Damblon, F., 1997. Anthracology and past vegetation reconstruction. In: Svoboda, J.(Ed.), Pavlov I – Northwest. Dolnı Vestonice Studies, 4, pp. 437–442.
Davis, S.J.M., 2005. Why domestic food animals? Some zoo-archaeological evidencefrom the Levant. Journal of Archaeological Science 32, 1408–1416.
d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Tillier, A.-M., Soressi, M.,Bresson, F., Maureille, B., Nowell, A., Lakarra, J., Backwell, L., Julien, M., 2003.Archaeological evidence for the emergence of language, symbolism, and music– an alternative multidisciplinary perspective. Journal of World Prehistory 17(1), 1–70.
Erbersdobler, K., 1968. Vergleichend morphologische Untersuchungen an Einzel-knochen des postcranialen Skeletts in Mitteleuropa vorkommender mittel-grober Huhnervogel. Ludwig-Maximilians-Universitat Munchen.
Ericson, P.G.P., 1987. Interpretations of archaeological bird remains: a taphonomicapproach. Journal of Archaeological Science 14, 65–75.
Fladerer, F.A., 2001. Die Faunareste vom jungpalaolithischen Lagerplatz Krems-Wachtberg, Ausgrabung 1930. Jagdwild und Tierkorpernutzung an der Donauvor 27.000 Jahren, 39. Mitteilungen der Prahistorischen Kommision, Osterrei-chische Akademie der Wissenschaften, Wien. Philosophish-historiche Klasse.
Flannery, K.V., 1969. Origins and ecological effects of early domestication in Iran andthe near East. In: Ucko, P.J., Dimbleby, G.W. (Eds.), The Domestication andExploitation of Plants and Animals. Duckworth, London, pp. 73–100.
Gal, E., 2005. New data to the bird bone artefacts from Hungary and Romania. In:Luik, H., Choyke, A.M., Batey, C.E., L}ougas, L. (Eds.), From Hooves to Horns, fromMollusc to Mammoth. Manufacture and Use of Bone Artefacts from PrehistoricTimes to the Present. Proceedings of the 4th Meeting of the ICAZ Worked BoneResearch Group at Tallinn, 26th–31st of August 2003, 15. Muinasaja teadus,Tallinn, pp. 325–338.
Goodwin, D., 1976. Crows of the World. Cornell University Press, Ithaca, New York.Guminski, W., 2005. Stone age hunters of Dudka and Szczepanki, Masurian Lake-
land, NE Poland. Acta Archaeologica 76, 111–143.Higgins, J., 1999. Tunel: a case study of avian zooarchaeology and taphonomy.
Journal of Archaeological Science 26, 1449–1457.Knecht, H., 1997. Projectile Technology. Plenum, New York.
Koumouzelis, M., Ginter, B., Koz1owski, J.K., Pawlikowski, M., Bar-Yosef, O.,Albert, R.M., Litynska-Zajac, M., Stworzewicz, E., Wojtal, P., Lipecki, G., Tomek, T.,Bochenski, Z.M., Pazdur, A., 2001. The early upper Palaeolithic in Greece: theexcavations in Klisoura cave. Journal of Archaeological Science 28, 515–539.
Kraft, E., 1972. Vergleichend morphologische Untersuchungen an Einzelknochennord- und mitteleuropaischer kleinerer Huhnervogel. Ludwig-Maximilians-Universitat Munchen.
Laroulandie, V., 1998. Etudes archeozoologique et taphonomique des Lagopedes dessaules de la grotte magdalenienne des Eglises (Ariege). Anthropozoologica 28,45–54.
Laroulandie, V., 2000. Taphonomie et archeozoologie des oiseaux en grotte:applications aux sites Paleolithiques du Bois-Ragot (Vienne), de Combe Sau-niere (Dordogne) et de la Vache (Ariege). These d’Universite, Universite deBordeaux I.
Laroulandie, V., 2002. Damage to pigeon long bones in pellets of the eagle owl Bubobubo and food remains of peregrine falcon Falco peregrinus: zooarchaeologicalimplications. In: Bochenski, Z.M., Bochenski, Z., Stewart, J.R. (Eds.), Proceedingsof the 4th Meeting of the ICAZ Bird Working Group, Krakow, Poland, 11–15September, 2001. Acta Zoologica Cracoviensia 45 (special issue), 331–339.
Laroulandie, V., 2003. Exploitation des Oiseaux au Magdalenien en France: Etat deslieux. In: Costamagno, S., Laroulandie, V. (Eds.), Mode de vie au Magdalenien:Apports de l’archeozoologie/Zooarchaeological insights into Magdalenian life-ways. Actes du colloque 6.4 du XIVe Congres du l’UISPP, Liege, Belgique, 2–8septembre 2001. BAR n� 1144. Royaume-Uni, Oxford.
Laroulandie, V., 2005a. Bird exploitation pattern: the case of Ptarmigan Lagopus sp.in the upper Magdalenian site of La Vache (Ariege, France). DocumentaArchaeobiologiae. In: Grupe, G., Peters, J. (Eds.), Feathers, Grit and Symbolism.Birds and Humans in the Ancient Old and New Worlds, Proceedings of the 5thMeeting of the ICAZ Bird Working Group, Munich, 26–28 July 2004, 3, pp.165–178.
Laroulandie, V., 2005b. Anthropogenic versus non-anthropogenic bird boneassemblages: new criteria for their distinction. In: O’Connor, T. (Ed.), Biosphereto Lithosphere. New Studies in Vertebrate Taphonomy. Oxbow Books, Oxford,pp. 25–30.
Laroulandie, V., Costamagno, S., Cochard, D., Mallye, J.-B., Beauval, C., Castel, J.-C.,Ferrie, J.-G., Gourichon, L., Rendu, W., 2008. Quand desarticuler laisse destraces: le cas de l’hyperextension du coude. Annales de Paleontologie 94,287–302.
Lipecki, G., Wojtal, P., 1998. Mammal remains. In: Kozlowski, J.K. (Ed.), Complex ofUpper Palaeolihic Sites near Moravany, Western Slovakia, vol. 2. Institute ofArchaeology, Jagellonian University, Cracow/Archaeological Institute, SlovakAcademy of Sciences, Nitra, pp. 103–126. Moravany-Lopata (Excavations1993–1996).
Livingston, S.D., 1989. The taphonomic interpretation of avian skeletal partfrequencies. Journal of Archaeological Science 16, 537–547.
Lyman, R.L., 1994. Vertebrate Taphonomy. Cambridge Manuals in Archaeology.Cambridge University Press, Cambridge.
Lyman, R.L., 2008. Quantitative Paleozoology. Cambridge Manuals in Archaeology.Cambridge University Press, Cambridge.
Mason, S., Hather, J., Hillman, G., 1994. Preliminary investigation of the plant macro-remains from Dolnı Vestonice II and its implications for the role of plant foodsin Palaeolithic and Mesolithic Europe. Antiquity 68, 48–57.
Mlikovsky, J., 1996. New data on the food of the White-tailed Sea Eagle (Haliaetusalbicilla) in the Svjatoj Nos wetlands, Lake Baikal. Buteo 8, 115–118.
Mourer-Chauvire, C., 1983. Les oiseaux dans les habitats paleolithiques: gibier deshommes ou proies des rapaces? In: Grigson, C., Clutton-Brock, J. (Eds.), Animaland Archaeology: 2. Shell Middens, Fishes and Birds. British ArchaeologicalReports, International Series, 183, pp. 111–124.
Mulkeen, S., O’Connor, T.P.O., 1997. Raptors in towns: towards an ecological model.International Journal of Osteoarchaeology 7, 440–449.
Munro, N.D., 2004. Zooarchaeological measures of hunting pressure and occupationintensity in the Natufian. Current Anthropology 45 (s4), s5–s34.
Munzel, S.C., 2004. Subsistence patterns in the Gravettian of the Ach valley,a former tributary of the Danube in the Swabian Jura. In: Svoboda, J., Sedlack-ova, L. (Eds.), The Gravettian Along the Danube. Dolnı Vestonice Studies, 11, pp.71–85.
Musil, R., 1955. Osteologick�y material z paleolitickeho sidliste v Pavlove. (Dasosteologische Material aus der palaolithischen Siedlungsstatte in Pollau). ActaAcademiae Scientarum �Cechoslovenicae, Basis Brunensis 27/318 (6), 279–320.
Musil, R., 1958. Funde von Wirbeltieren in Mahren (Tschechoslowakei) aus demletzten Wurm-Stadial. Eiszeitalter und Gegenwart 9, 61–68.
Musil, R., 1959. Osteologick�y material z paleolitickeho sidliste v Pavlove. �Cast II. (Dasosteologische Material aus der palaolithischen Siedlungsstatte in Pavlov. II Teil).Anthropozoikum 8, 83–106.
Musil, R., 1994. The fauna. Hunting game of the culture layer of Pavlov. In:Svoboda, J. (Ed.), Pavlov I. Excavations 1952–53. Eraul 66/Dolnı VestoniceStudies, 2, pp. 181–209.
Musil, R., 1997. Hunting analysis. In: Svoboda, J. (Ed.), Pavlov IdNorthwest. DolnıVestonice Studies, 4, pp. 443–468.
Musil, R., 2003. The middle and upper palaeolithic game suit in central andsoutheastern Europe. In: Andel, T.H.van, Davies, W. (Eds.), Neanderthals andModern Humans in the European Landscape During the Last Glaciation. Cam-bridge, pp. 167–190.
Musil, R., 2005. Animal prey. In: Svoboda, J. (Ed.), Pavlov I Southeast. AWindow intothe Gravettian Lifestyles. Dolnı Vestonice Studies, 14, pp. 190–228.
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–26652664

Author's personal copy
Nadachowski, A., 2005. Small vertebrates and environmental reconstruction. In:Svoboda, J. (Ed.), Pavlov I Southeast. A Window into the Gravettian Lifestyles.Dolnı Vestonice Studies, 14, pp. 187–189.
Novak, M., 2005. Pavlov I – Southeast. Review of spatial distributions. In: Svoboda, J.(Ed.), Pavlov I Southeast. A Window into the Gravettian Lifestyles. Dolnı Ves-tonice Studies, 14, pp. 53–71.
Opravil, E., 1994. The vegetation. In: Svoboda, J. (Ed.), Pavlov I. Excavations 1952–53.Eraul 66/Dolnı Vestonice Studies, 2, pp. 175–180.
Plisson, H., Geneste, J.M., 1989. Analyse technologique des pointes a cran solutreensdu Placard (Charente), du Fourneau du Diable, du Pech de la Boissiere et deCombe Sauniere (Dordogne). Paleo 1, 65–106.
Reichstein, H., Pieper, H., 1986. Untersuchungen an Skelettresten von Vogeln ausHaithabu (Ausgrabung 1966-1969). In: Berichte uber die Ausgrabungen inHaithabu Bericht, 22. Karl Wachholz Verlag, Neumunster.
Reisch, L., 1976. Beobachtungen an Vogelknochen aus dem Spatpleistozan der Hohlevon Kephalari (Argolis, Griechenland). Archaologisches Korrespondenzblatt 6,261–265.
Rosendahl, G., Beinhauer, K.-W., Loscher, M., Kreipl, K., Walter, R., Rosendahl, W.,2006. Le plus vieil arc du monde? Une piece interessante en provenance deMannheim, Allemagne. L’Anthropologie 110, 371–382.
Rybnıckova, E., Rybnıcek, K., 1991. The environment of the Pavlovian – Palae-oecological results from Bulhary, South Moravia. In: Edler-Kovacs, J. (Ed.),Palaeovegetational Development in Europe, Pan-European PalaeobotanicalConference Vienna, pp. 73–79.
Serjeantson, D., Irving, B., Hamilton-Dyer, S., 1993. Bird taphonomy from the insideout: the evidence of gull predation on the manx shearwater Puffinus puffinus.Archaeofauna 2, 191–204.
Soffer, O., Vandiver, P., 2005. Ceramic fragments. In: Svoboda, J. (Ed.), Pavlov ISoutheast. AWindow into the Gravettian Lifestyles. Dolnı Vestonice Studies, 14,pp. 415–431.
Steadman, D.W., Plourde, A., Burley, D., 2002. Prehistoric butchery and consumptionof birds in the Kingdom of Tonga, South Pacific. Journal of ArchaeologicalScience 29, 571–584.
Stewart, J.R., 2007. An Evolutionary Study of Some Archaeologically SignificantAvian Taxa in the Quaternary of the Western Palaearctic. In: BAR InternationalSeries, 1653.
Stiner, M.C., 2001. Thirty years on the ‘‘Broad Spectrum Revolution’’ and paleolithicdemography. Proceedings of the National Academy of Sciences 98 (13),6993–6996.
Stiner, M.C., Munro, N.D., Surovell, T.A., 2000. The tortoise and the hare: small-gameuse, the broad spectrum revolution, and Paleolithic demography. CurrentAnthropology 41, 39–73.
Svoboda, J. (Ed.), 1994, Pavlov I. Excavations 1952–53. Eraul 66/Dolnı VestoniceStudies, 2 Liege.
Svoboda, J. (Ed.), 1997. Pavlov I – Northwest. Dolnı Vestonice Studies, 4 Brno.Svoboda, J., 2005. Pavlov I – Southeast. Location, Stratigraphy, Microstratigraphies,
and Features. In: Svoboda, J. (Ed.), Pavlov I Southeast. A Window into theGravettian Lifestyles. Dolnı Vestonice Studies, 14, pp. 25–52.
Svoboda, J., Kralık, M., �Culıkova, V., Hladilova, S., Novak, M., N�yvltova Fisakova, M.,N�yvlt, D., Zelinkova, M., 2009. Pavlov VI: an upper paleolithic living unit.Antiquity 83, 282–295.
Svobodova, H., 1991a. The pollen analysis of Dolnı Vestonice II, section No 1. In:Svoboda, J. (Ed.), Dolnı Vestonice II – Western Slope, 54, pp. 75–88. Eraul.
Svobodova, H., 1991b. Pollen analysis of the upper paleolithic triple burial at DolnıVestonice. Archeologicke rozhledy 43, 505–510.
Tagliacozzo, A., Gala, M., 2002. Exploitation of Anseriformes at two Upper Palae-olithic sites in Southern Italy: Grotta Romanelli (Lecce, Apulia) and Grotta delSantuario della Madonna a Praia a Mare (Cosenza, Calabria). In: Bochenski, Z.M.,Bochenski, Z., Stewart, J.R. (Eds.), Proceedings of the 4th Meeting of the ICAZBird Working Group, Krakow, Poland, 11–15 September, 2001. Acta ZoologicaCracoviensia 45 (special issue), 117–131.
Tomek, T., Bochenski, Z.M., 2000. The Comparative Osteology of European Corvids(Aves: Corvidae), with a Key to the Identification of their Skeletal Remains.Publications of the Institute of Systematics and Evolution of Animals, Krakow.
Tomek, T., Bochenski, Z., 2005. Weichselian and Holocene bird remains fromKomarowa Cave, Central Poland. Acta Zoologica Cracoviensia 48A (1–2),43–65.
Tomek, T., Bochenski, Z., Bochenski, Z.M., 2003. Birds (Aves). In: Valde-Novak, P.,Nadachowski, A., Madeyska, T. (Eds.), Oblazowa Cave: Human Activity, Stra-tigraphy and Palaeoenvironment. Institute of Archaeology and Ethnology PolishAcademy of Sciences, Krakow, pp. 102–113.
Trinkaus, A.E., Svoboda, J.A., Wojtal, P., N�yvltova Fisakova, M., Wilczynski, J., 2009.Human remains from the Moravian Gravettian: morphology and taphonomy ofadditional elements from dolnı Vestonice II and Pavlov I. International Journalof Osteoarchaeology. doi:10.1002/oa.1088.
Tyrberg, T., 1995. Palaeobiogeography of the genus Lagopus in the West Palearctic.Courier Forschungsinstitut Senckenberg 181, 275–291.
Tyrberg, T., 1998. Pleistocene Birds of the Palearctic: a Catalogue. In: Publication ofthe Nuttall Ornithological Club, 27 Cambridge, Massatchusetts, and updates athttp://web.telia.com/wu11502098/pleistocene.pdf (2008-02-24).
Verpoorte, A., 2005. Lithic assemblage of Pavlov I South-Central (1954, 1956, 1963,1964). In: Svoboda, J. (Ed.), Pavlov I Southeast. A Window into the GravettianLifestyles. Dolnı Vestonice Studies, 14, pp. 75–111.
Villa, P., Boscato, P., Ranaldo, F., Ronchitelli, A., 2009. Stone tools for the hunt: pointswith impact scars from a Middle Paleolithic site in southern Italy. Journal ofArchaeological Science 36, 850–859.
Villotte, S., 2008. Enthesopathies et Activites des hommes prehistoriques. Thesepresente a l’Universite de Bordeaux 1 pour obtenir le grade de Docteur.Bordeaux.
West, D.L., 1996. Krakow-Spadzista, unit E and unit F. Faunal remains. In:Koz1owski, J.K. (Ed.), The Upper Palaeolithic site Krakow-Spadzista. Units D, Eand F (Excavations1986–1989). Folia Quaternaria 67, 21–34.
Wijngaarden-Bakker, L.H., 1997. The selection of bird bones for artefact productionat Dutch Neolithic sites. International Journal of Osteoarchaeology 7, 339–345.
Wojtal, P., 2000. Taphonomy. In: Cyrek, K., Nadachowski, A., Madeyska, T.,Bochenski, Z., Tomek, T., Wojtal, P., Miekina, B., Lipecki, G., Garapich, A., Rzebik-Kowalska, B., Stworzewicz, E., Wolsan, M., Godawa, J., Ko�sciow, R., Fostowicz-Frelik, q., Szyndlar, S. (Eds.), Excavation in the Deszczowa Cave (KroczyckieRocks, Czestochowa Upland, central Poland). Folia Quaternaria 71, 5–84.
Wojtal, P., Sedlackova, L., Wilczynski, J., 2005. Human activities on the faunalmaterial. In: Svoboda, J. (Ed.), Pavlov I Southeast. AWindow into the GravettianLifestyles. Dolnı Vestonice Studies, 14, pp. 229–231.
Z.M. Bochenski et al. / Journal of Archaeological Science 36 (2009) 2655–2665 2665
Related Documents











