ELSEVIER Brain Research 707 (1996) 213-220 BRAIN RESEARCH Research report Formation and dissolution of spinules and changes in nematosome size require optic nerve integrity in black bass ( Micropterus salmoides) retina Joaquln De Juan *, Magdalena Garcla, Nicolfis Cuenca Departamento de Histologia e lnstituto de Neurociencias, Uniuersidad de Alicante, Apdo. Correos 374, Alicante 03080, Spain Acxrepted 28 September 1995 Abstract Teleost retinas adapted to light show numerous spinules invaginated in the cone pedicles and small nematosomes in the distal horizontal cells. Darkness induces the dissolution of spinules and the presence of large and numerous nematosomes. The aim of this work is to study the influence of optic nerve integrity on spinule formation/dissolution and changes in nematosome size during light or dark adaptation of black bass (Micropterus salmoides) retinas. Eyes from fish, dark- or light-adapted, were removed and the eyecups placed in oxygenated Ringer's solution and immediately exposed to light or dark, respectively, for 1 h. The number of spinules per pedicle and the nematosome diameter were measured on electron micrographs. Isolation of eyecups in the dark, impaired both spinule formation and nematosome size reduction when they were superfused in light. In the same way, isolation of eyecups in the light, impaired both spinule dissolution and nematosome size increase when they were superfused in dark. No significant differences in spinule number and nematosome size, following dopamine superfusion, were found in comparison to retinas supeffused with Ringer's solution only. Our results suggest: (1) optic nerve integrity is necessary to yield spinule formation/disruption and changes in nematosome size during light or dark adaptation. (2) dopamine does not appear to be the primary agent responsible for spinule formation. Keywords: Cone pedicle; Horizontal cell; Synaptic plasticity; Dopamine; Nucleolus-like body; Light- and dark-adaptation; Efferent fiber 1. Introduction In teleost retinas, cone pedicles are invaginated by the fingerlike protrusions from lateral processes of cone hori- zontal cell dendrites termed spinules [32]. These spinules, which have characteristic densities beneath the membrane, are dynamic structures, forming during light adaptation and disappearing during dark adaptation [5,9- 11,17,19,25,32,35]. The combination of intracellular recording and quantitative ultrastructural analysis has sug- gested that spinules may be the sites of feedback transmis- sion from cone horizontal cells (HCs) to pedicles of cones [9-11,17,19,25,32,35]. Processes that might contribute to the formation of the spinules are cytoplasmic inclusions, resembling nucleoli and termed nematosomes, located in the external HCs of white perch (Roccus americana) retinas [5]. Nematosomes are larger and more numerous in dark-adapted retinas than in light-adapted ones. We found * Corresponding author. Departamento de Histologla, Facultad de Medicina, Apdo. Correos 374, Alicante 03080, Spain. Fax: (34) (6) 565-8511. 0006-8993/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 0006-8993(95)01259-1 that there exists an inverse relationship between number and size of nematosomes, and spinule number, suggesting that electron-dense material observed in spinules may orig- inate in the nematosomes [5]. What are the mechanisms regulating the spinules and nematosomes? It has been reported [12,20,21,34] that dopamine (DA) influences spinule formation. However, whereas the cited authors consider DA as an essential prerequisite for spinule formation, we have shown that this neuromodulator is not the key factor in spinule production [4]. In teleost fish, DA is released by the interplexiform cells [14,16,18,31]. These cells provide an intraretinal cen- trifugal pathway from the inner to the outer plexiform layer, where they make abundant synapses on cone-related HCs. The interplexiform cells receive all their input in the inner plexiform layer from amacrine cells and centrifugal fibers [15,37]. In a number of non-mammalian species, DA release seems to be influenced by centrifugal fibers project into the retina [24,26]. In fish retina, centrifugal fibers originate in the olfactory bulb and contain gonadotropin hormone- releasing hormone (GnRH)-like and FMRFamide-like pep- tides [23,28]. Contacts between centrifugal fibers and

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

E L S E V I E R Brain Research 707 (1996) 213-220
BRAIN RESEARCH
R e s e a r c h repor t
Formation and dissolution of spinules and changes in nematosome size require optic nerve integrity in black bass ( Micropterus salmoides) retina
J o a q u l n De Juan *, M a g d a l e n a Garc la , Nicolf is C u e n c a
Departamento de Histologia e lnstituto de Neurociencias, Uniuersidad de Alicante, Apdo. Correos 374, Alicante 03080, Spain
Acxrepted 28 September 1995
Abstract
Teleost retinas adapted to light show numerous spinules invaginated in the cone pedicles and small nematosomes in the distal horizontal cells. Darkness induces the dissolution of spinules and the presence of large and numerous nematosomes. The aim of this work is to study the influence of optic nerve integrity on spinule formation/dissolution and changes in nematosome size during light or dark adaptation of black bass (Micropterus salmoides) retinas. Eyes from fish, dark- or light-adapted, were removed and the eyecups placed in oxygenated Ringer's solution and immediately exposed to light or dark, respectively, for 1 h. The number of spinules per pedicle and the nematosome diameter were measured on electron micrographs. Isolation of eyecups in the dark, impaired both spinule formation and nematosome size reduction when they were superfused in light. In the same way, isolation of eyecups in the light, impaired both spinule dissolution and nematosome size increase when they were superfused in dark. No significant differences in spinule number and nematosome size, following dopamine superfusion, were found in comparison to retinas supeffused with Ringer's solution only. Our results suggest: (1) optic nerve integrity is necessary to yield spinule formation/disruption and changes in nematosome size during light or dark adaptation. (2) dopamine does not appear to be the primary agent responsible for spinule formation.
Keywords: Cone pedicle; Horizontal cell; Synaptic plasticity; Dopamine; Nucleolus-like body; Light- and dark-adaptation; Efferent fiber
1. Introduction
In teleost retinas, cone pedicles are invaginated by the fingerlike protrusions from lateral processes of cone hori- zontal cell dendrites termed spinules [32]. These spinules, which have characteristic densities beneath the membrane, are dynamic structures, forming during light adaptation and disappearing during dark adaptation [5 ,9- 11,17,19,25,32,35]. The combination of intracellular recording and quantitative ultrastructural analysis has sug- gested that spinules may be the sites of feedback transmis- sion from cone horizontal cells (HCs) to pedicles of cones [9-11,17,19,25,32,35]. Processes that might contribute to the formation of the spinules are cytoplasmic inclusions, resembling nucleoli and termed nematosomes, located in the external HCs of white perch (Roccus americana) retinas [5]. Nematosomes are larger and more numerous in dark-adapted retinas than in light-adapted ones. We found
* Corresponding author. Departamento de Histologla, Facultad de Medicina, Apdo. Correos 374, Alicante 03080, Spain. Fax: (34) (6) 565-8511.
0006-8993/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 0006-8993(95)01259-1
that there exists an inverse relationship between number and size of nematosomes, and spinule number, suggesting that electron-dense material observed in spinules may orig- inate in the nematosomes [5].
What are the mechanisms regulating the spinules and nematosomes? It has been reported [12,20,21,34] that dopamine (DA) influences spinule formation. However, whereas the cited authors consider DA as an essential prerequisite for spinule formation, we have shown that this neuromodulator is not the key factor in spinule production [4]. In teleost fish, DA is released by the interplexiform cells [14,16,18,31]. These cells provide an intraretinal cen- trifugal pathway from the inner to the outer plexiform layer, where they make abundant synapses on cone-related HCs. The interplexiform cells receive all their input in the inner plexiform layer from amacrine cells and centrifugal fibers [15,37].
In a number of non-mammalian species, DA release seems to be influenced by centrifugal fibers project into the retina [24,26]. In fish retina, centrifugal fibers originate in the olfactory bulb and contain gonadotropin hormone- releasing hormone (GnRH)-like and FMRFamide-like pep- tides [23,28]. Contacts between centrifugal fibers and

214 J.D. Juan et al. / Brain Research 707 (1996) 213-220
dopaminergic interplexiform cells were observed by elec- tron microscopy to be conventional synaptic junctions [37]. Also, it has been observed [30] that GnRH closely mim- icked the effects of DA on HCs, indicating that GnRH acts by stimulating the release of DA from interplexiform cells. FMRFamide suppresses the effects of GnRH on HCs in some experiments [30]. Finally, it has been reported that GnRH induces light-adaptive spinule formation in dark- adapted retinas [3].
This work has three aims: (1) to determine whether in isolated eyecups, formation/dissolution of horizontal cell spinules, and changes in nematosome size during light or dark adaptation are possible, (2) to determine whether DA is involved in the control of spinules and nematosomes in isolated eyecups, "and (3) to determine whether in these experiments, as in previous ones [5], there exists a inverse co-variation between the number of spinules and nemato- some size.
By means of an ultrastructural and morphometric study, we showed that spinule formation/dissolution and changes in nematosomes size were impaired by isolation of the eyecups. In contrast, when DA was added to them, there were not effects.
• 2 hours 60 minutes
' l© b Protocol II
10 minutes
. @ I
Fig. 1. Protocols used for ultrastructural study of the fish retinas. Left side represents the initial state of illumination: light- (white fish) or dark- (black fish) adaptation for 2 hours. In Protocols I, II, V and VI, the eyes were removed (scissors) and theirs eyecups superfused in light- (white retina) or dark- (black retina) conditions for 60 min. In Protocols III and IV, after the 2-hours light- and dark-adaptation, the animals were dark- (black fish in the middle) and light- (white fish in the middle) adapted for 10 min respectively, with the eyes then being removed (scissors) and theirs eyecups superfused in light- (white retina) or dark- (black retina) conditions for 60 min.
2. Materials and methods
Black bass (Micropterus salmoides) 12-15 cm in length were kept in an aquarium under standard conditions and light-dark cycles of 12 h. For this work, six experimental protocols were performed (Fig. 1). Groups of two black bass were used in each protocol. The animals were decapi- tated and the eyes enucleated, hemisected, and the poste- rior part (the eyecups) immersed in Ringer's solution. Standard Ringer's solution consisted of (mM): NaCI 145.0, KCI 2.5, MgSO 4 0.1, CaCI 2 0.7, NaHCO 3 25.0, glucose 20.0. This solution was continuously bubbled with a mix- ture of 97% 02 and 3% CO 2, maintained at 20°C and pH 7.3. The superfusion medium that flowed over the eyecups was delivered to the chamber via fine polyethyline tubing and was drained out by tissue paper placed on the edge of the chamber. Surgery and preparation of dark-adapted fish were performed under dim red light.
In order to determine the effect of DA on spinule formation and changes in nematosome size, eyecups from two additional fish, for each protocol, were superfused with 50 /xM DA in the Ringer's solution. Dopamine solutions, freshly prepared, were protected from oxidation by adding 0.1 mM ascorbic acid to the superfusion medium. The DA concentration used was in accordance with con- centration reported to be electrophysiologically active [29,36] and capable to produced spinules [34].
After completion of the protocols, the retinas were dissected and divided in four pieces and fixed overnight at 4°C with 1% paraformaldehyde, 1.6% glutaraldehyde, 0.15 mM CaC12 in 0.1 M phosphate buffer (pH 7.4). After
fixation, four blocks of tissue were taken from each of these pieces, rinsed in 5% sucrose phosphate buffer and postfixed with 2% OsO 4 for 1 h. The retinas were dehy- drated in ethanol and embedded in Epon-812. The blocks were vertically sectioned in an LKB-III ultramicrotome. Sections of 1 /~m thick were stained by the Rosenquist's [27] method or with Toluidine blue. Semi-thin sections were observed in a light microscope to study changes in the length of cones as an internal control to ensure that DA, and light and dark adaptation have the effects on the retina expected from previous experiments. Five vertical sections, 50-100 nm thick, were obtained from each block and stained with uranyl acetate and lead citrate. Electron micrographs of cone pedicles and the adjacent HCs were printed at final magnification of x 15,000 for cone pedi- cles and × 30,000 for nematosomes. Using a homemade computer program, the number of spinules was counted in fifty sections of cone pedicles per retina and nematosome diameter was calculated for fifty sections of nematosomes. In order to obtain a representative sample across the retina, the sections number of pedicles and nematosomes exam- ined was distributed among the four blocks and the struc- tures randomly selected from different sections. Statistical differences were checked by the Student's t-test.
3. Results
In prolonged dark adapted retinas (Protocol VI), the Black bass cone myoides (Fig. 2a) are widely elongated (46 ___ 5 ~m, S.E.) from the outer limitant membrane

J.D. Juan et al. / Brain Research 707 (1996) 213-220 215
OLM
k
OLM
Fig. 2. Light micrographs of 1 p,m semithin retinal section from dark- (a) and light- (b) adapted retinas. In dark-adapted retinas, cone myoids are elongated and the ellipsoids ( * ) are far to the outer limiting membrane (OLM). In contrast, in light adapted retinas the ellipsoids are close to the OLM. Bar: 20 #m.
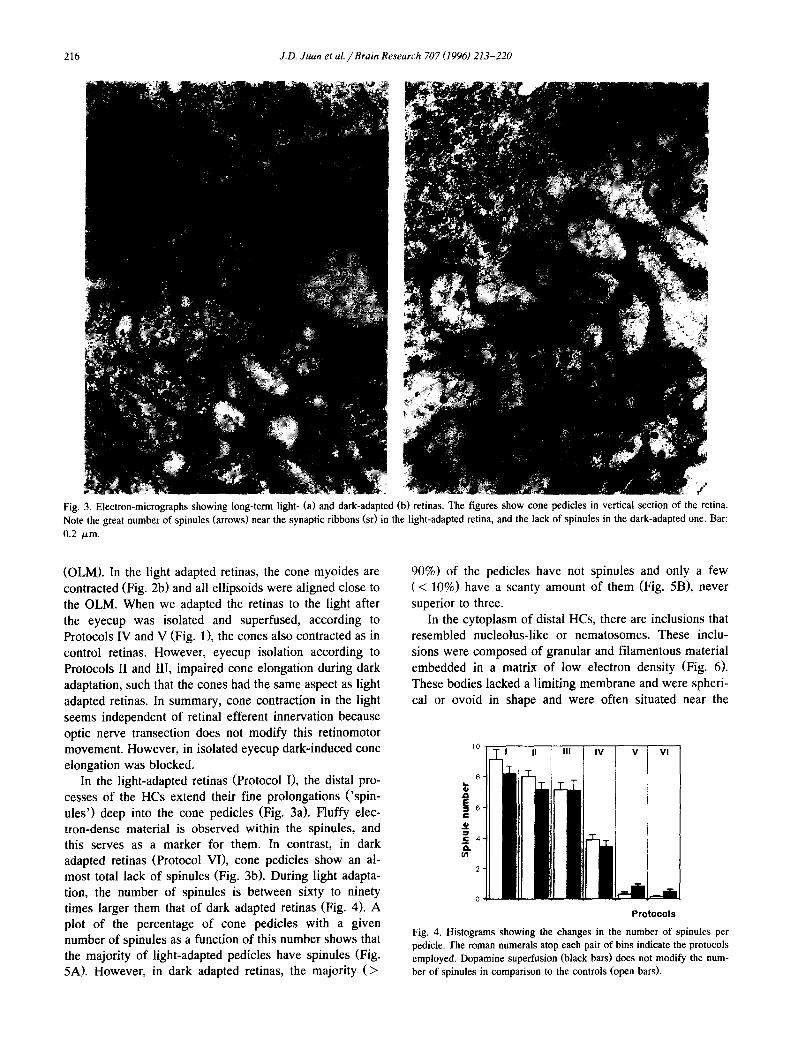
216 J.D. Juan et al. // Brain Research 707 (1996) 213-220
Fig. 3. Electron-micrographs showing long-term light- (a) and dark-adapted (b) retinas. The figures show cone pedicles in vertical section of the retina. Note the great number of spinules (arrows) near the synaptic ribbons (st) in the light-adapted retina, and the lack of spinules in the dark-adapted one. Bar: 0.2 /.tm.
(OLM). In the light adapted retinas, the cone myoides are contracted (Fig. 2b) and all ellipsoids were aligned close to the OLM. When we adapted the retinas to the light after the eyecup was isolated and superfused, according to Protocols IV and V (Fig. 1), the cones also contracted as in control retinas. However, eyecup isolation according to Protocols II and Ill, impaired cone elongation during dark adaptation, such that the cones had the same aspect as light adapted retinas. In summary, cone contraction in the light seems independent of retinal efferent innervation because optic nerve transection does not modify this retinomotor movement. However, in isolated eyecup dark-induced cone elongation was blocked.
In the light-adapted retinas (Protocol I), the distal pro- cesses of the HCs extend their fine prolongations ('spin- ules') deep into the cone pedicles (Fig. 3a). Fluffy elec- tron-dense material is observed within the spinules, and this serves as a marker for them. In contrast, in dark adapted retinas (Protocol VI), cone pedicles show an al- most total lack of spinules (Fig. 3b). During light adapta- tion, the number of spinules is between sixty to ninety times larger them that of dark adapted retinas (Fig. 4). A plot of the percentage of cone pedicles with a given number of spinules as a function of this number shows that the majority of light-adapted pedicles have spinules (Fig. 5A). However, in dark adapted retinas, the majority (>
90%) of the pedicles have not spinules and only a few (< 10%) have a scanty amount of them (Fig. 5B), never superior to three.
In the cytoplasm of distal HCs, there are inclusions that resembled nucleolus-like or nematosomes. These inclu- sions were composed of granular and filamentous material embedded in a matrix of low electron density (Fig. 6). These bodies lacked a limiting membrane and were spheri- cal or ovoid in shape and were often situated near the
1o ~ II I III iv V Vl
e I
Protocols
Fig. 4. Histograms showing the changes in the number of spinules per pedicle. The roman numerals atop each pair of bins indicate the protocols employed. Dopamine superfusion (black bars) does not modify the num- ber of spinules in comparison to the controls (open bars).
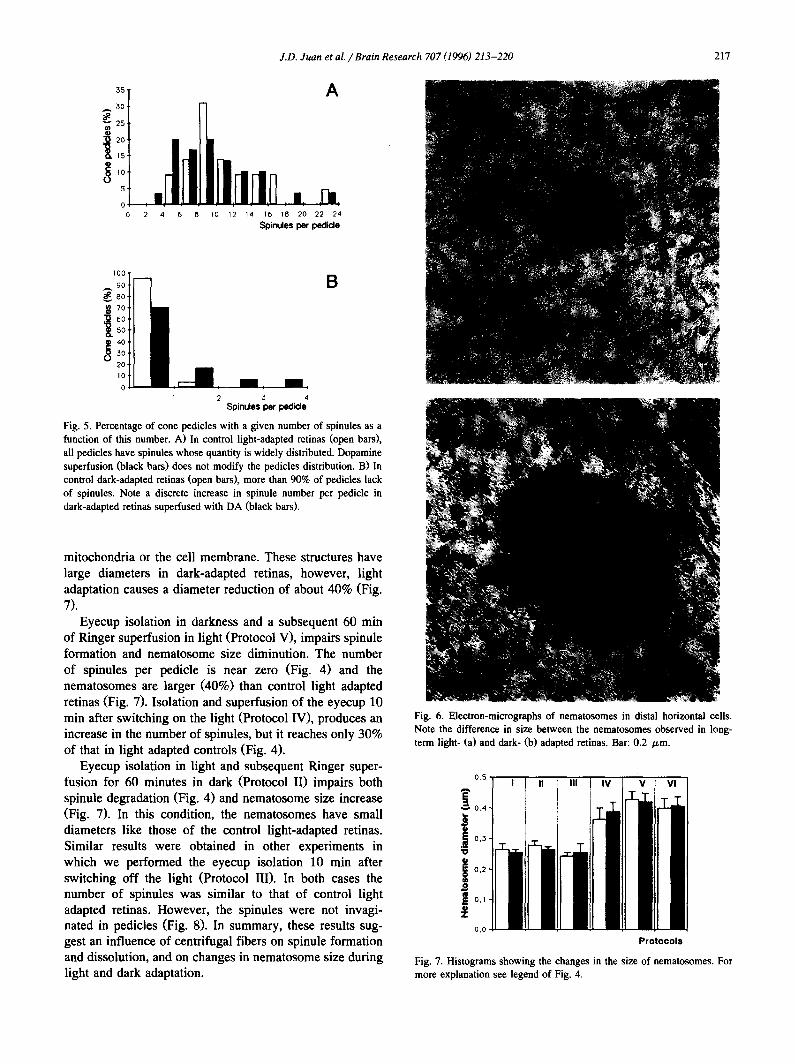
J.D. Juan et aL / Brain Research 707 (1996) 213-220 217
3O 7
20
1S
5
0 0 2 4 6 8 IO 12 14 16 18 20 22 24
S p i n u l e s pe r p e d i d e
IOO
9o
8o
70 L 30 20 IO 0
1 2 5 4 S p i n u l e s pe r p e d i d e
Fig. 5. Percentage of cone pedicles with a given number of spinules as a function of this number. A) In control light-adapted retinas (open bars), all pedicles have spinules whose quantity is widely distributed. Dopamine superfusion (black bars) does not modify the pedicles distribution. B) In control dark-adapted retinas (open bars), more than 90% of pedicles lack of spinules. Note a discrete increase in spinule number per pedicle in dark-adapted retinas superfused with DA (black bars).
mitochondria or the cell membrane. These structures have large diameters in dark-adapted retinas, however, light adaptation causes a diameter reduction of about 40% (Fig. 7).
Eyecup isolation in darkness and a subsequent 60 min of Ringer superfusion in light (Protocol V), impairs spinule formation and nematosome size diminution. The number of spinules per pedicle is near zero (Fig. 4) and the nematosomes are larger (40%) than control light adapted retinas (Fig. 7). Isolation and superfusion of the eyecup 10 min after switching on the light (Protocol IV), produces an increase in the number of spinules, but it reaches only 30% of that in light adapted controls (Fig. 4).
Eyecup isolation in light and subsequent Ringer super- fusion for 60 minutes in dark (Protocol II) impairs both spinule degradation (Fig. 4) and nematosome size increase (Fig. 7). In this condition, the nematosomes have small diameters like those of the control light-adapted retinas. Similar results were obtained in other experiments in which we performed the eyecup isolation 10 rain after switching off the light (Protocol III). In both cases the number of spinules was similar to that of control light adapted retinas. However, the spinules were not invagi- nated in pedicles (Fig. 8). In summary, these results sug- gest an influence of centrifugal fibers on spinule formation and dissolution, and on changes in nematosome size during light and dark adaptation.
Fig. 6. Electron-micrographs of nematosomes in distal horizontal cells. Note the difference in size between the nematosomes observed in long- term light- (a) and dark- (b) adapted retinas. Bar: 0.2 /zm.
0 . 5 ~
! ill Eli Hi i o. ll ill ill ill ill ill il
I°"tl II II I11 Ill Itl I 0,0 ' '
Protocols
Fig. 7. Histograms showing the changes in the size of nematosomes. For more explanation see legend of Fig. 4.

218 J.D. Juan et al. / Brain Research 707 (1996) 213-220
In order to determine the possible role of dopamine (DA) in spinule formation, six additional experiments were performed. In these experiments, DA was added to the Ringer's solution in each of the protocols described in Fig. 1. The results obtained showed that DA perfusion does not modify the number of spinules (Fig. 4) and nematosome size (Fig. 7) in comparison to the controls. These data suggest that DA is not a major factor in spinule formation.
4. Discussion
In Micropterus salmoides retinas, spinules form when light dawns and disappear almost completely during dark adaptation. Similar to others species [32,33], the time course of total formation or disappearance is near 60 min (data not shown). However, in Micropterus salmoides, the number of spinules per pedicle in light adapted retinas is greater than in other teleosts investigated so far, being double that in Carassius auratus, Tinca tinca and Cypri- nus carpio [33]. In dark adapted retinas, this number is close to zero in Micropterus salmoides, whereas in the other investigated teleosts ranged near one to two spinules per pedicle [32].
Retinas isolated from long-term dark adapted Mi- cropterus salmoides (> 2 h), and afterwards incubated at light during 60 min (Protocol V), produced cone contrac- tion but no spinule formation. This indicated that spinule
outgrowth needs, simultaneously, retinal illumination and optic nerve integrity. This suggested a central influence, probably via centrifugal fibres, on spinule formation. How- ever, cone contraction was absolutely independent of cen- tral influence as is the case with other species [8].
It was reported that spinule formation and cone contrac- tion are independent processes that can be triggered by only a brief (1-2 min) exposure to light [7,33,35]. In one experiment (Protocol IV), we analyzed the influence of a short light exposition (10 min) in spinule formation. In this experiment, long-time dark adapted fishes (> 2 h) were exposed to light for 10 min before eyecups isolation. In this case, the number of spinules did not reach the values of light-adapted retinas, rising to only 30% of these values. This suggests that although spinule formation has an en- dogenous control [13] and can be started by only a brief exposure to light, optic nerve integrity is necessary for its total development. However, cones were contracted as expected.
In a similar way, the dissolution of spinules also needs optic nerve integrity. In fact, in long-term light adapted retinas, which were isolated and superfused in dark for 1 h (Protocol II), the number of spinule did not decrease. (However, the spinules did not invaginate into the cone pedicles.) The significance of this non dissolution of spin- ules in deafferentized retinas remains to be established. As it occurs in other teleosts [8], in our experiments using Protocol II, cone elongation is not reached.
Fig. 8. Electron-micrographs of a vertical section from a dark-adapted retina studied according to Protocol II. Note the great number of spinules (arrows) not invaginated into cone pedicles, which remain near the pedicle base. Similar results were obtained using Protocol III. Bar: 0.4/zm.

J.D. Juan et al. / Brain Research 707 (1996) 213-220 219
This absence of elongation could, in principle, have been attributed to cone contraction due to a high concen- tration of extracellular Ca ++ (to 10 -3 M or upper) as described by Dearry and Burnside [6]. However, our re- sults discard this explanation, because in control eyecups, previously dark adapted and super/used in darkness with the same concentrations of extracellular Ca 2+, no cone contracted, even though 5 mM taurine was added to the superfusate as indicated by Dearry and Burnside [7].
Several observations strongly suggest that DA synthesis and/or release by interplexiform cells is controlled by centrifugal fibers. For instance, in fish retinas, suppression of interplexiform cells by 6-OHDA [2] or the optic nerve transection [22] leads to an increase in connexon density in gap junctions of HCs. Dopamine is known to reduce the connexon density in the gap junctions of HCs [1]. Finally, exogenous GnRH, which is the likely neurotransmitter of the centrifugal fibers, acts by stimulating the release of DA from interplexiform cells and reproducing the effects of DA on HCs [30].
Our results are in agreement with an influence of centrifugal fibers on spinule formation and dissolution. However, these results do not support that DA acts by triggering spinule formation. These results are in contradic- tion with the interpretation of other authors [12,20,21,34], who consider DA as an essential prerequisite for spinule formation. In all our experiments, DA superfusion did not modify the number of spinules and this is in agreement with previous results obtained by us [4]. In addition, our unpublished data suggest that dopamine is not the only factor responsible for the formation of spinules and it must act in conjunction with an other neuroactive substance. Despite intraocular injection of DA, a fish adapted to the darkness can produce a normal number of spinules (unpub- lished data). Furthermore, in retinas with intact optic nerve treated with 6-hydroxydopamine (6-OHDA), intraocular injection of DA restores only partially (40%) the number of spinules. We propose that putative non-dopaminergic factor is located in the interplexiform cell (IPC) and that this factor's liberation is eliminated when the IPC is destroyed by 6-OHDA or in the absence of efferent fiber stimulation in optic nerve section.
Why are there differences between our results and that of other authors? The manner of obtaining histological sections cannot account for these differences, since the use of tangential or transverse section does not seem to cause differences when spinules are counted [31]. We also do not believe that the problem stems from too little DA reaching the HCs in our experiments. The doses of DA used by us (50 /xM) were adjusted at the concentration required to produce a response between maximal and half maximal from the dose-response data of Kirsch et al. [20]. This dose is also in agreement with values employed in electro- physyological studies [29,37] and by Weiler et al. [34] in the study of effects of DA in formation of spinules. Furthermore, our study of the changes in the length of
cones served as an internal control to ensure that DA had the effects on the retina expected from previous experi- ments. On the other hand, the differences between our results and that of other authors could be due to the species of fish used, as we comment at the beginning of this discussion.
The large size of nematosomes when spinules are scanty or non existent (in dark-adapted retinas and in retinas with impaired spinule formation) and vice versa, suggests a close relationship between nematosomes and the electron- dense material located beneath the spinule plasmalemma. More information about the significance of nematosomes can be found in our previous work [5].
In conclusion, our results are in agreement with a central control of spinule formation through centrifugal fibers, probably by means of GnRH as a suggested Behrens et al. [3]. However, we think that DA release from inter- plexiform cells is not the only factor responsible for starting spinule formation. Similarly, spinule disruption is also centrally controlled but we know nothing about this mechanism. Finally, our data suggest, as in a previous work [5], that spinule electron-dense material originates in nematosomes.
Acknowledgements
We wish to thank J. Stobbs for improving the English, Drs. John E. Dowling and N.M. Grzywacz for critically reading the manuscript, and the Conselleria de Medi Am- bient of the Generalitat Valenciana for helping us to obtain the fish. This research was supported by DGICYT Grant PM92-0113 and GV-2521-94 to J.D.J.
References
[1] Baldridge, W.H., Ball, A. and Miller, R.G., Dopaminergic regulation of horizontal cell gap junction particle density in goldfish retina, J. Comp. Neurol., 265 (1987) 428-436.
[2] Baldridge, W.H. Ball, A.K. and Miller, R.G., Gap junction particle density of horizontal cells in goldfish retinas lesioned with 6-OHDA, J. Comp. Neurol., 287 (1989) 238-246.
[3] Behrens, U.D., Douglas, R.H. and Wagner, H.J., Gonadotropin-re- leasing hormone, a neuropeptide of efferent projections to the teleost retina induces light-adaptive spinule formation on horizontal cell dendrites in dark-adapted preparations kept in vitro, Neurosci. Lett. 164 (1993) 59-62.
[4] De Juan, J. Ifiiguez, C. and Dowling, J.E., Influence of dopamine on spinules, synaptic ribbons and nematosomes in white perch retina, Invest. Ophtalmol. Vis. Sci. Suppl., 31 (1990) 333.
[5] De Juan, J., Ifiiguez, C. and Dowling, J.E., Nematosomes in external horizontal cells of white perch (Roccus americana) retina: changes during dark and light adaptation, Brain Res., 546 (1991) 176-180.
[6] Dearry, A. and Burnside, B., Effects of extracellular Ca+ + , K + , and Na+ on cone and RPE retinomotor movements in isolated teleost retinas, J. Gen. Physiol., 83 (1984) 589-611.
[7] Dearry, A. and Burnside, B., Dopaminergic regulation of cone retinomotor movement in isolated teleost retinas. I. Induction of

220 J.D. Juan et aL / Brain Research 707 (1996) 213-220
cone contraction is mediated by D2 receptors, J. Neurochem., 46 (1986) 1006-1021.
[8] Dearry, A. and Barlow, R.B., Circadian rhythms in the green sunfish retina, J. Gen. Physiol., 89 (1987) 745-770.
[9] Djamgoz, M.B.A., Downing, J.E.G., Kirsch, M., Prince, D.J, and Wagner, H.J., Plasticity of cone horizontal cell function in cyprinid fish retina: effects of background illumination of moderate intensity, J. Neurocytol., 17 (1988) 701-710.
[10] Djamgoz, M.B.A, Downing, J.E.G., Prince, D.J. and Wagner, H.J., Physiological and ultrastructural evidence for light-dependent plas- ticity of cone photoreceptor horizontal cell feedback interactions in the isolated retina of cyprinid fish, J. Physiol., 362 (1985) 20.
[11] Djamgoz, M.B.A., Downing, J.E.G., and Wagner, H.J., The cellular origin of an unusual type of S-potential: an intraceUular horseradish peroxidase study in a cyprinid retina, J. Neurocytol., 14 (1985) 469-486.
[12] Djamgoz, M.B.A. Kirsch, M., Wagner, H.J., Haloperidol suppresses light-induced spiriule formation and biphasic responses of horizontal cells in fish (roach) retina, Neurosci. Lett., 107 (1989) 200-204.
[13] Douglas, R.H. and Wagner, H.J., Endogenous control of spinule formation in horizontal cells of the teleost retina, Cell Tissue Res., 229 (1983) 443-449.
[14] Dowling, J.E., Retinal neuromodulation: the role of dopamine, Vis. Neurosci., 7 (1991) 87-97.
[15] Dowling, J.E. and Ehinger, B., Synaptic organization of the amine- containing interplexiform cells of the goldfish and Cebus monkey retinas, Science, 188 (1975) 270-273.
[16] Dowling, J.E. and Ehinger, B., The interplexiform cell system. 1. Synapses of the dopaminergic neurons of the goldfish retina, Proc. Roy. Soc. B. (Lond.), 201 (1978) 7-26.
[17] Downing, J.E.G. and Djamgoz, M.B.A., Quantitative analysis of cone photoreceptor-horizontal cell connectivity patterns in the retina of a cyprinid fish: electron microscopy of functionally identified and HRP-labelled horizontal cells, J. Comp. Neurol., 289 (1989) 537- 553.
[18] Ehinger, B., Falck, B. and Laties, A.M., Adrenergic neurons in teleost retina, Z. Zellforsch. Mikrosk. Anat., 97 (1969) 285-297.
[19] Kirsch, M., Djamgoz, M.B.A., and Wagner, H.J., Correlation of spinule dynamics and plasticity of the horizontal cell spectral re- sponse in cyprinid fish retina: quantitative analysis, Cell Tissue Res., 260 (1990) 123-130.
[20] Kirsch, M., Wagner, H.J., Djamgoz, M.B.A., Dopamine and plastic- ity of horizontal cell function in the teleost retina: regulation of a spectral mechanism through Dl-receptors, Brain Res., 31 (1988) 401-412.
[21] Kohler, K. and Weiler, R., Dopaminergic modulation of transient neurite outgrowth from horizontal cells of the fish retina is not mediated by cAMP, Eur. J. Neurosci., 2 (1990) 788-794.
[22] Kurz-Isler, G., Voigt, T., and Wolburg, H., Modulation of connexon densities in gap junctions of horizontal cell perikarya and axon terminals in fish retina: effects of light/dark cycles, interruption of the optic nerve and application of dopamine, Cell Tissue Res., 268 (1992) 267-275.
[23] Miinz, H., Class, B., Stumpf, W.E., Jennes, L., Centrifugal innerva- tion of the retina by luteinizing hormone releasing hormone (LHRH-immunoreactive telencephalic neurons in teleostean fishes), Cell Tissue Res., 222 (1982) 313-323.
[24] Ramrn y Cajal, S., Textura del Sistema Nervioso del Hombre y de los Vertebrados. Tomo 11. Capitulo XXX. Continuacirn del Estudio de la Retina, Imprenta y Libreria de Nicol:~s Moya, Madrid, 1904, pp 610-645.
[25] Raynauld, J.P., Laviolette, J.R. and Wagner, H.J., Goldfish retina: a correlate between cone activity and morphology of the horizontal cell in cone pedicles, Science, 204 (1979) 1436-1438.
[26] Reperant, J., Miceli, D., Vesselkin, N.P. and Molorchnikoff, S., The centrifugal visual system of vertebrates: a century-old search re- viewed, Int. Rev. Cytol., 118 (1989) 115-171.
[27] Rosenquist, T.H., Slavin, B.G. and Bernick, S., The pearson silver- gelatin method for light microscopy of 0.5-2 /xm plastic sections, Stam Technol., 46 (1971) 253-257.
[28] Stell, W.K., Walker, S.E., Chohan, K.S. and Ball, A.K., The gold- fish nervous terminalis: a luteinizing hormone-releasing hormone and moluscan cardioexcitatory peptide immunoreactive olfactoreti- nal pathway, Proc. Natl. Acad. Sci. USA, 81 (1984) 940-944.
[29] Tornquist, K., Yang, X.-L. and Dowling, J.E., Modulation of cone horizontal cell activity in the teleost fish retina. IlL Effects of prolonged darkness and dopamine on electrical coupling between horizontal cells, J. Neurosci., 8 (1988) 2279-2288.
[30] Umino, O. and Dowling, J.E., Dopamine release from interplexiform cells in the retina: effects of GnRH, FMRFamide, bicuculline, and enkephalin on horizontal cell activity, J. Neurosci., 11 (1991) 3034-3046.
[31] Umino, O., Lee, Y., and Dowling, J.E., Effects of light stimuli on the release of dopamine from interplexiform cells in the white perch retina, Vis. Neurosci., 7 (1991) 451-458.
[32] Wagner, H.J., Light-dependent plasticity of the morphology of hori- zontal cell terminals in cone pedicles of fish retinas, J. Neurocytol., 9 (1980) 573-590.
[33] Wagner, H.J. and Douglas, R.H., Morphologic changes in teleost primary and secondary retinal cells following brief exposure to light, Invest. Ophthalmol. Vis. Sci., 24 (1983) 24-29.
[34] Weiler, R., Kohler, K., Kirsch, M., and Wagner, H.J., Glutamate and dopamine modulate synaptic plasticity in horizontal cell dendrites of fish retina, Neurosci. Lett., 87 (1988) 205-209.
[35] Weiler, R. and Wagner, H.J., Light-dependent change of cone horizontal cell interactions in carp retina, Brain Res. 298 (1984) 1-9.
[36] Yang, X.-L., Tornquist, K.,. and Dowling, J.E., Modulation of cone horizontal cell activity in the teleost fish retina. III. Effects of prolonged darkness and dopamine on electrical coupling between horizontal cells, J. Neurosci., 8 (1988) 2279-2288.
[37] Zucker, C.L. and Dowling, J.E., Centrifugal fibres synapse on dopaminergic interplexiform cells in the teleost retina, Nature, 300 (1987) 166-168.
Related Documents











