ORIGINAL ARTICLE Forced aggregation and defined factors allow highly uniform-sized embryoid bodies and functional cardiomyocytes from human embryonic and induced pluripotent stem cells Martin Pesl • Ivana Acimovic • Jan Pribyl • Renata Hezova • Aleksandra Vilotic • Jeremy Fauconnier • Jan Vrbsky • Peter Kruzliak • Petr Skladal • Tomas Kara • Vladimir Rotrekl • Alain Lacampagne • Petr Dvorak • Albano C. Meli Received: 25 May 2013 / Accepted: 25 October 2013 / Published online: 21 November 2013 Ó Springer Japan 2013 Abstract In vitro human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs) can differentiate into functional cardiomyocytes (CMs). Pro- tocols for cardiac differentiation of hESCs and hiPSCs include formation of the three-dimensional cell aggregates called embryoid bodies (EBs). The traditional suspension method for EB formation from clumps of cells results in an EB population heterogeneous in size and shape. In this study we show that forced aggregation of a defined number of single cells on AggreWell plates gives a high number of homogeneous EBs that can be efficiently differentiated into functional CMs by application of defined growth factors in the media. For cardiac differentiation, we used three hESC lines and one hiPSC line. Our contracting EBs and the resulting CMs express cardiac markers, namely myosin heavy chain a and b, cardiac ryanodine receptor/calcium release channel, and cardiac troponin T, shown by real- time polymerase chain reaction and immunocytochemistry. Using Ca 2? imaging and atomic force microscopy, we demonstrate the functionality of RyR2 to release Ca 2? from the sarcoplasmic reticulum as well as reliability in contractile and beating properties of hESC-EBs and hiPSC- EBs upon the stimulation or inhibition of the b-adrenergic pathway. Keywords Human pluripotent stem cell Embryoid body Differentiation Cardiomyocyte Calcium Introduction Human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs) have the ability to grow indefinitely and differentiate in all somatic cell types. Both of these cell types can be differentiated into functional cardiomyocytes (CMs) in vitro. Differentiation of hESCs and hiPSCs across the cardiac lineage can be achieved using different methods: coculture with mouse visceral endoderm-like cells (END-2 cells), formation of three- dimensional aggregates called embryoid bodies (blasto- cyst-like structures [1, 2]), or culturing as a monolayer on Matrigel (BD Biosciences, San Jose, CA, USA) in medium supplemented with defined factors. The efficiency of these methods depends on the developmental potential of each cell line, which may vary between lines [3–5]. While the differentiation of CMs in embryoid bodies (EBs) has been reported as reliable, resulting in up to 60 % contracting M. Pesl and I. Acimovic contributed equally to this work. Electronic supplementary material The online version of this article (doi:10.1007/s00380-013-0436-9) contains supplementary material, which is available to authorized users. M. Pesl I. Acimovic A. Vilotic J. Vrbsky V. Rotrekl P. Dvorak A. C. Meli Department of Biology, Faculty of Medicine, Masaryk University, Brno, Czech Republic M. Pesl J. Vrbsky P. Kruzliak T. Kara P. Dvorak ICRC, St Anne’s University Hospital, Brno, Czech Republic J. Pribyl R. Hezova P. Skladal CEITEC, Masaryk University, Brno, Czech Republic J. Fauconnier A. Lacampagne A. C. Meli (&) INSERM U1046, University of Montpellier I, University of Montpellier II, 371 Avenue du Doyen G. Giraud, CHU Arnaud de Villeneuve, Building INSERM Crastes de Paulet, 34295 Montpellier, France e-mail: [email protected] P. Skladal Department of Biochemistry, Faculty of Science, Brno, Czech Republic 123 Heart Vessels (2014) 29:834–846 DOI 10.1007/s00380-013-0436-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Forced aggregation and defined factors allow highly uniform-sizedembryoid bodies and functional cardiomyocytes from humanembryonic and induced pluripotent stem cells
Martin Pesl • Ivana Acimovic • Jan Pribyl • Renata Hezova • Aleksandra Vilotic •
Jeremy Fauconnier • Jan Vrbsky • Peter Kruzliak • Petr Skladal •
Tomas Kara • Vladimir Rotrekl • Alain Lacampagne • Petr Dvorak •
Albano C. Meli
Received: 25 May 2013 / Accepted: 25 October 2013 / Published online: 21 November 2013
� Springer Japan 2013
Abstract In vitro human embryonic stem cells (hESCs)
and human induced pluripotent stem cells (hiPSCs) can
differentiate into functional cardiomyocytes (CMs). Pro-
tocols for cardiac differentiation of hESCs and hiPSCs
include formation of the three-dimensional cell aggregates
called embryoid bodies (EBs). The traditional suspension
method for EB formation from clumps of cells results in an
EB population heterogeneous in size and shape. In this
study we show that forced aggregation of a defined number
of single cells on AggreWell plates gives a high number of
homogeneous EBs that can be efficiently differentiated into
functional CMs by application of defined growth factors in
the media. For cardiac differentiation, we used three hESC
lines and one hiPSC line. Our contracting EBs and the
resulting CMs express cardiac markers, namely myosin
heavy chain a and b, cardiac ryanodine receptor/calcium
release channel, and cardiac troponin T, shown by real-
time polymerase chain reaction and immunocytochemistry.
Using Ca2? imaging and atomic force microscopy, we
demonstrate the functionality of RyR2 to release Ca2?
from the sarcoplasmic reticulum as well as reliability in
contractile and beating properties of hESC-EBs and hiPSC-
EBs upon the stimulation or inhibition of the b-adrenergic
pathway.
Keywords Human pluripotent stem cell � Embryoid
body � Differentiation � Cardiomyocyte � Calcium
Introduction
Human embryonic stem cells (hESCs) and human induced
pluripotent stem cells (hiPSCs) have the ability to grow
indefinitely and differentiate in all somatic cell types. Both
of these cell types can be differentiated into functional
cardiomyocytes (CMs) in vitro. Differentiation of hESCs
and hiPSCs across the cardiac lineage can be achieved
using different methods: coculture with mouse visceral
endoderm-like cells (END-2 cells), formation of three-
dimensional aggregates called embryoid bodies (blasto-
cyst-like structures [1, 2]), or culturing as a monolayer on
Matrigel (BD Biosciences, San Jose, CA, USA) in medium
supplemented with defined factors. The efficiency of these
methods depends on the developmental potential of each
cell line, which may vary between lines [3–5]. While the
differentiation of CMs in embryoid bodies (EBs) has been
reported as reliable, resulting in up to 60 % contracting
M. Pesl and I. Acimovic contributed equally to this work.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00380-013-0436-9) contains supplementarymaterial, which is available to authorized users.
M. Pesl � I. Acimovic � A. Vilotic � J. Vrbsky � V. Rotrekl �P. Dvorak � A. C. Meli
Department of Biology, Faculty of Medicine, Masaryk
University, Brno, Czech Republic
M. Pesl � J. Vrbsky � P. Kruzliak � T. Kara � P. Dvorak
ICRC, St Anne’s University Hospital, Brno, Czech Republic
J. Pribyl � R. Hezova � P. Skladal
CEITEC, Masaryk University, Brno, Czech Republic
J. Fauconnier � A. Lacampagne � A. C. Meli (&)
INSERM U1046, University of Montpellier I, University
of Montpellier II, 371 Avenue du Doyen G. Giraud,
CHU Arnaud de Villeneuve, Building INSERM Crastes de
Paulet, 34295 Montpellier, France
e-mail: [email protected]
P. Skladal
Department of Biochemistry, Faculty of Science, Brno,
Czech Republic
123
Heart Vessels (2014) 29:834–846
DOI 10.1007/s00380-013-0436-9

EBs [4, 6, 7], the heterogeneity in size and shape of the
EBs may eventually contribute to inconsistency in effi-
ciency of cardiac differentiation.
By using well-characterized CCTL12, CCTL13, and
CCTL14 hESC lines [8, 9] as well as clone 4 hiPSC line
[10], we have established a method to form highly homo-
geneous EBs containing equal numbers of stem cells that
can be differentiated to CMs. This was achieved through
forced aggregation in AggreWell plates followed by three
specific steps of induction [11]. Importantly, while the first
contracting EBs appear between days 14 and 20 of dif-
ferentiation, we observed interline variability in differen-
tiating efficiency between lines. Quantitative reverse
transcription–polymerase chain reaction (qRT-PCR) and
immunocytochemistry indicate that the contracting hESC-
EBs and hiPSC-EBs of ±200 lm in diameter express
cardiac markers including atrial and ventricular myosin
heavy chain a (MYH6) and b (MYH7), and cardiac ryan-
odine receptor/calcium (Ca2?)-release channel (RYR2),
yielding a population of up to 95 % of cardiac troponin T
(cTnT)-positive cells. Using a Ca2? imaging technique and
atomic force microscopy (AFM), we observed spontaneous
Ca2? transients and demonstrated the functionality of
RyR2 in releasing Ca2? from the sarcoplasmic reticulum
(SR). We also observed reliability in contractile and beat-
ing properties of EBs from both hESCs and hiPSCs in
response to pharmacologic modulators of the b-adrenergic
pathway (i.e., isoproterenol and metoprolol). We showed
that caffeine, a RyR2 agonist, increases contraction force
(CF) in correlation with the depletion of Ca2? from the SR
and through RyR2.
Taken together, these data indicate that using both hESC
and hiPSC lines, AggreWell plates are suitable for for-
mation of highly uniform EBs with similar starting cell
numbers, which can be differentiated to contracting EBs
containing functional CMs.
Materials and methods
Maintenance of pluripotent stem cells
The CCTL12, CCTL13, and CCTL14 hESC lines [8, 9] as
well as the clone 4 hiPSC line [10] used in this study were
maintained in an undifferentiated state as colonies on the
mitomycin C (10 lg/ml; Molekula, Gillingham, UK)
inactivated mouse embryonic fibroblast (MEF) feeder
(seeded at 5 9 105 MEFs per 60-mm diameter Petri dish)
in hESC medium consisting of Dulbecco’s modified
Eagle’s medium (DMEM)/F-12 (Gibco, Grand Island, NY,
USA), 15 % KnockOut Serum Replacement (KO-SR;
Gibco), 1 % L-glutamine (Gibco), 1 % nonessential amino
acids (PAA, Pasching, Austria), 0.5 % penicillin–
streptomycin (PAA), 0.1 mM b-mercaptoethanol (Sigma,
St Louis, MO, USA) and 4 ng/ml human fibroblast growth
factor 2 (FGF2; R&D Systems, Minneapolis, MN, USA).
All cells were passaged with 1 mg/ml collagenase type IV
(Gibco). Passages were: CCTL12 (33–65), CCTL13
(34–60), CCTL14 (36–50), hiPSC clone 4 (63–96).
Formation of embryoid bodies
Prior to formation of EBs, cells treated with collagenase type
IV were plated on Matrigel-coated plates and cultured in
MEF-conditioned medium (MEF-CM) supplemented with
10 ng/ml human FGF2 and 1 % L-glutamine. The next two
passages of cells were done using 19 TrypLE Express
(Gibco). Incubation of the cells with 19 TrypLE was for 2
min at 37 �C, followed by centrifugation of cells at 170g for 4
min (Eppendorf centrifuge 5804R), and cells were grown as
monolayers on Matrigel. When cells reached 100 % con-
fluent monolayer, they were treated with 19 Accutase
(Sigma) for 4 min at 37 �C, and centrifuged at 350g for 7 min.
EB formation was induced by seeding of ±2.5 9 106 cells in
2 ml of EB medium consisting of KO-DMEM (Invitrogen,
Carlsbad, CA, USA), 10 % heat-inactivated fetal bovine
serum (Invitrogen), 1 % L-glutamine, 1 % nonessential
amino acids, 1 % penicillin–streptomycin, and 0.1 mM b-
mercaptoethanol per well on an AggreWell 400 plate
(Stemcell Technologies, Grenoble, France). EB medium was
supplemented with 25 lg/ml Y-27632 (Sigma), and forced
aggregation of cells was achieved by centrifugation of the
plate at 100g for 3 min at room temperature (23 �C). The
plate was then held under hypoxic conditions (90 % N2, 5 %
O2, and 5 % CO2) at 37 �C.
Cardiac differentiation
Twenty-four hours after seeding of the cells, formed EBs
were transferred to low-adhesive 60-mm diameter Petri
dishes (ratio 1:2) in Induction 1 medium composed of EB
medium supplemented with 10 ng/ml bone morphogenetic
protein 4 (BMP4; R&D), 5 ng/ml FGF2, and 6 ng/ml activin
A (R&D). After 4 days, medium was replaced with Induction
2 medium composed of EB medium supplemented with 10
ng/ml vascular endothelial growth factor (VEGF; R&D) and
10 lM inhibitor of Wnt response 1 (IWR-1; Sigma) for 3
days, followed by Induction 3 medium (EB medium with 10
ng/ml VEGF and 5 ng/ml FGF2). Induction 3 medium was
changed every fourth day until day 28 of differentiation, after
which EBs were kept only in EB medium without growth
factors. Dishes were held in the hypoxic conditions (5 % O2
and 5 % CO2) at 37 �C for the first 12 days of the differen-
tiation and then transferred to normoxic conditions (20 % O2,
75 % N2, 5 % CO2). First beating EBs appeared between days
14 and 20 of differentiation.
Heart Vessels (2014) 29:834–846 835
123

Quantitative RT-PCR
One-month-old beating CCTL12 hESC-EBs and undiffer-
entiated CCTL12 hESCs on Matrigel were used for RNA
extraction. Total RNA was isolated from the cells using a
PureLink RNA Mini Kit (Ambion, Carlsbad, CA, USA)
followed by cDNA synthesis using Revert Aid H Minus
First Strand cDNA Synthesis Kit (Fermentas, Gummers-
bach, Germany) according to the manufacturer’s protocols.
Real-time PCR was performed using the Applied Biosys-
tems 7500 Sequence Detection System. The 20-ll PCR
reaction mixture consisted of 10 ll TaqMan Gene Expres-
sion Master Mix (Applied Biosystems, Foster City, CA,
USA), 8 ll nuclease-free water, 1 ll primer and probe mix
of the TaqMan Gene Expression assay (MYH6, MYH7 and
RYR2; Applied Biosystems), and 1 ll cDNA. Reactions
were running on 96-well optical plates (Applied Biosys-
tems) at 50 �C for 2 min, followed by 95 �C for 10 min, and
40 cycles at 95 �C for 15 s and 60 �C for 1 min. Expression
levels were normalized to the housekeeping gene glycer-
aldehyde-3-phosphate dehydrogenase (GAPDH).
Immunocytochemistry
To dissociate cells from EBs, 10 beating EBs were col-
lected in 1 ml culture medium and washed three times in 10
ml Ca2?-free solution (120 mM NaCl, 5.4 mM KCl, 5 mM
MgSO4, 5 mM sodium pyruvate, 20 mM glucose, 20 mM
taurine, and 10 mM HEPES; pH 6.9). EBs were then
incubated in Ca2?-free solution for 20 min before centri-
fugation at 15g for 3 min. After removing the supernatant,
EBs were incubated in 1 ml digestion solution (Ca2?-free
solution supplemented with 0.8 mg/ml of collagenase from
Clostridium histolyticum (Sigma) and 0.04 mg/ml protease
from Streptomyces griseus (Sigma)) prewarmed to 37 �C,
for 30 min at 37 �C, with periodic shaking. Dissociated
cells were centrifuged at 170g for 3 min and the cell pellet
was resuspended in 1 ml KB solution (85 mM KCl, 30 mM
K2HPO4, 1 mM EGTA, 2 mM ATP-Na2, 5 mM sodium
pyruvate, 5 mM creatine, 20 mM taurine, and 20 mM
glucose; pH 7.3) prewarmed to 37 �C and incubated for 30
min at 37 �C, with periodic gentle shaking. To stain dis-
sociated CMs, cells were plated onto gelatin-coated 12-mm
diameter glass coverslips in EB medium. The fixation of
the samples was done with 4 % paraformaldehyde for 1 h
on ice and washed with 19 phosphate-buffered saline
(PBS). A blocking solution 1 (1 % bovine serum albumin
(BSA), 0.1 % Triton in 19 PBS) was used for 15 min at
room temperature and then washed out with 19 PBS. A
blocking solution 2 (1 % BSA, 0.03 % Tween, 0.01 %
NaN3 in 19 PBS) was used for 1 h at room temperature
and then washed out with 19 PBS. Incubation with pri-
mary antibody anti-human cTnT (1:500, Cell Signaling,
Danvers, MA, USA) took place overnight at 4 �C. Incu-
bation with donkey anti-rabbit Alexa 594 secondary anti-
body (1:500, Invitrogen) was for 1 h at room temperature.
Samples were then mounted in Mowiol–diamidino-2
phenylindole (DAPI; Sigma) and were observed under a
living-cell confocal Carl Zeiss LSM 700 microscope
(Oberkochen, Germany). Images were analyzed using
ImageJ, version 4.6 (rsweb.nih.gov/ij/). For counting of
cTnT-positive cells, images with DAPI and cTnT signals
were overlapped. The total number of nuclei and cTnT-
negative cells were counted manually and were used to
calculate the percentage of cTnT-positive cells. cTnT-
positive cells were scored in each experiment on at least
three slides. The average percentage of positive cells and
standard deviation were calculated from a minimum of 500
cells per experiment.
For immunostaining of cut sections of EBs, beating EBs
were embedded in 1 % low melting agarose (Promega,
Madison, WI, USA) in 19 PBS and stored at -80 �C until
sectioning. Prior cryosectioning (Leica CM 1850; Leica,
Wetzlar, Germany), EBs in low melting agarose were
frozen in Jung Tissue Freezing Medium (Leica). Cryosec-
tions (5 lm) were fixed in 4 % paraformaldehyde for 1 h on
ice and washed with 19 PBS. A blocking solution 1 (1 %
BSA, 0.1 % Triton in 19 PBS) was used for 15 min at
room temperature and then washed out three times for 5
min with 19 PBS. A blocking solution 2 (1 % BSA, 0.03 %
Tween in 19 PBS) was used for 1 h at room temperature
and then washed out with 0.05 % Tween in 19 PBS.
Samples were incubated with primary antibody anti-human
cTnT (1:200, Santa Cruz Biotechnologies, Santa Cruz, CA,
USA) for 1.5 h at room temperature, washed three times for
5 min with 0.05 % Tween in 19 PBS, and incubated with
donkey anti-goat Alexa 488 secondary antibody (1:400,
Invitrogen) for 1 h at room temperature. After incubation
with secondary antibody, samples were incubated for 5 min
with DAPI (1:1000 in 19 PBS; Sigma), washed three times
for 5 min with 19 PBS, and mounted in Mowiol (Sigma).
All solutions were administered in drops. Pictures of the
samples were taken using a living-cell confocal Carl Zeiss
LSM 700 microscope.
Measurements of cytosolic Ca2? variation
Cells were incubated in a Tyrode’s solution (135 mM
NaCl, 10 mM HEPES, 5.4 mM KCl, 0.9 mM MgCl2; pH
7.4) containing 1 mM CaCl2 and loaded with 3–5 lM
Fluo4-AM (Invitrogen) for 20 min. Cells were then placed
in an experimental chamber on the stage of an inverted
microscope. Ca2? images were recorded with a laser
scanning confocal microscope (Zeiss LSM Exciter, 409
water immersion objective, in x–y mode, 1 image/0.495 s).
To enable comparisons between cells, changes in the
836 Heart Vessels (2014) 29:834–846
123

Fluo-4 fluorescence signal (DF) were divided by basal
fluorescence (F0) using ImageJ. Ca2? transient amplitudes
were analyzed by Prism (version 6.0; GraphPad, San
Diego, CA, USA).
Atomic force microscopy
Beating EBs were plated on a 60-mm diameter Petri dish in
the presence of Tyrode’s solution. Prior to the experiments,
1.8 mM CaCl2 and 10 mM glucose were added freshly to
the solution maintained at 37 �C. To avoid the cantilever
thermic deflection, the parts of AFM located in liquid were
heated up to 37 �C and maintained for 15 min. Landing
procedure was finalized with the deflection (DFL) param-
eter changed to value of 2 nA, which was proportional to the
force value of 25 nN (force acting on the tip surface). In all
AFM experiments, we used a silicon nitride triangular-lever
probe SNL-10 (Bruker AFM Probes, Camarillo, CA, USA)
with a silicon cantilever of very low spring constant (0.06
N/m). Data acquisition was performed using an AFM
microscope (NTgra Vita) equipped with scanning by probe-
measuring head (NT-MDT, Zelenograd, Russia). Temper-
ature inside the measuring chamber was driven by Petri dish
thermostatic module SU045NTF (NT-MDT). Nova soft-
ware version 1.0.26.1297 (NT-MDT) was used to drive the
AFM microscope, capture the data, and partially evaluate
the measured data.
According to Hook’s law, the cantilever deflection
(bending) is proportional to the force acting between the
EB and cantilever:
F ¼ �k � Dh ð1Þ
where F is the acting force, k is stiffness of the cantilever
material, and Dh is the change of cantilever position caused
by the force. The recorded DFL–height calibration curve
was fitted using the Boltzmann function allowing simple
calculation of Dh (height):
DFL ¼ A1 � A2
1þ eðh�h0=dhÞ þ A2 ð2Þ
where A1 (initial value), A2 (final value), h0 (center), and dh
are the parameters of regression function, height h is the
function argument, and DFL is the dependent variable. The
data (DFL vs time curves) recorded by AFM microscope
were analyzed by Microcal Origin software, version 8.07
(OriginLab, Northampton, CA, USA), using an internal
peak-fitting module, whereby the proper peaks in the
recorded curves can be marked by setting peak orientation,
width, height, and minimum height as the peak parameters.
Position of peak maxima in time was exported and used for
further analysis of beat frequency and force (using Eqs. 1
and 2, and cantilever spring constant k equal to 0.06 N/m).
Young’s modulus describing stiffness of the cells in cluster
was calculated from the measured force-distance (FD)
curves (DFL vs height). DFL was further transformed to
the force values and fitted with the Hertzian model,
described by the equation:
F ¼ 4Ka2
2ð1� vÞp tan Hð3Þ
where F is the force acting on cell surface, K is Young’s
modulus, a is the depth of indentation, m is Poisson’s ratio
(equal to 0.5 for incompressible materials), and H is the
angle between tip walls and EB surface.
Statistical analysis
Statistical comparisons were performed with the nonpara-
metric Wilcoxon signed-rank test or the Mann–Whitney test
when specified, using Prism (version 6.0; GraphPad). Data
are expressed as mean ± standard error of the mean (SEM),
and differences were considered significant at P\0.05. To
compare percentage of contracting EBs in all lines, one-way
analysis of variance (ANOVA) followed by post hoc Bon-
ferroni analysis with a two-sided method was used.
Results
Efficiency in generating contracting EBs from hESCs
and hiPSCs
EBs were derived from CCTL12, CCTL13, and CCTL14
hESC lines [8, 9] and clone 4 hiPSC line [10] using stem
cells grown on Matrigel-coated plates. EBs were formed
with the same initial amount of stem cells per EB (2000
cells) by centrifugation of AggreWell 400 plates. Forced
aggregation by a mild centrifugation step (100g, 3 min at
room temperature) using AggreWell 400 plates and 24 h of
hypoxic incubation allows highly uniformly sized and
shaped EBs of ±200 lm in diameter (Fig. 1a).
To quantify the efficiency in obtaining beating EBs from
the different stem cell lines, we adapted a protocol of
cardiac differentiation with three steps of induction under
hypoxia using defined cytokines, as previously published
(Fig. 1b) [11]. EBs from all tested cell lines started to beat
from days 14 to 20 of differentiation.
To evaluate the efficiency of our method using Aggre-
Well 400 plates, we counted manually the number of
beating EBs over the total number of EBs per plate and per
line, and at least three times. From days 20 to 30 of dif-
ferentiation, CCTL12 hESC, CCTL13 hESC, and clone 4
hiPSC lines exhibit between 33 and 46 % of beating EBs
on average (CCTL13 hESC mean 34 ± 6 %, n = 6 versus
clone 4 hiPSC mean 44 ± 5 %, n = 6; not significant;
Heart Vessels (2014) 29:834–846 837
123

Fig. 1c). CCTL14 hESC line delivers fewer beating EBs on
average, with high variability, and significantly different to
the CCTL12 hESC line (CCTL12 mean 46 ± 6 %, n = 3
versus CCTL14 mean 10 ± 9 %, n = 3; P\0.05; Fig. 1c),
whereas EBs that were not treated with growth factors
produced \1 % (data not shown). Thus, while interline
variability is observed, CCTL12 hESC and clone 4 hiPSC
lines display the highest rate of contracting EBs.
Fig. 1 a Formed embryoid bodies (EBs; from CCTL13 hESC line) after
24 h on AggreWell 400 plate and after transfer to low-adhesive 60-mm
diameter Petri dish. b Schematic outlines of cardiac differentiation
protocol. c Percentage of beating EBs over the total number of EBs formed
from days 20 to 30 of differentiation. For each line displayed, 3–6 dishes
were used for counting. Data presented as mean ± SEM. *P\0.05. BMP4,
bone morphogenetic protein 4; FGF2, fibroblast growth factor 2; VEGF,
vascular endothelial growth factor; IWR1, inhibitor of Wnt response 1
838 Heart Vessels (2014) 29:834–846
123

We then tried to form EBs using V96 microwell plates,
which are used to generate beating EBs from many lines
without interline variability, as previously published [4].
When using V96 microwell plates with similar conditions of
media and cell density, EBs do not form (data not shown).
Interestingly, we noticed that the 24-h postformation
density of plated EBs per Petri dish modulates the ratio of
‘‘attached’’ EBs over floating EBs. Thus, when ±500 EBs
are placed in a low-adhesive 60-mm diameter Petri dish,
more than 90 % of them result in floating EBs eventually.
A density of ±1000 EBs per low-adhesive 60-mm Petri
dish results in more than 50 % of ‘‘attached’’ EBs over the
total number of EBs (data not shown). These observations
indicate the importance of EB density, in agreement with
observations made for murine ESCs [12].
To assess relative expression of cardiac markers in
1-month-old EBs, we performed qRT-PCR to detect specific
atrial myosin heavy-chain a (MYH6) and ventricular myosin
heavy-chain b (MYH7). We also evaluated the expression of
cardiac ryanodine receptor (RYR2). The comparison of the
relative expression of these three genes between CCTL12
hESC-EBs and the corresponding undifferentiated stem cells
indicates that all genes are significantly upregulated (Table 1;
Fig. 2a). Correspondingly, CCTL12 hESC-EBs at day 30 of
differentiation exhibit significantly higher levels of MYH6,
MYH7, and RYR2 mRNA with a 98,661-fold, 507-fold, and
38-fold increase, respectively.
In addition, we immunostained enzymatically dissoci-
ated cells from the contracting EBs by using an antibody
against the sarcomeric cTnT and detected its expression
(Fig. 2b). Dissociated cells from beating CCTL13 hESC-
EBs are cTnT-positive, exhibiting a typical striated pattern
[13, 14]. However, we observed that these cells exhibit
heterogeneous shapes (e.g., rectangular, triangular, or
round), likely due to the EB spheroid spatial organization
as previously described [15] (Fig. 2c). We then counted the
number of cTnT-positive cells over the total number of
cells and based on the number of DAPI-stained nuclear
DNA, and found a cardiac efficiency of 95.2 ± 0.9 % (n =
3) for the CCTL13 hESC line. To confirm that most of cells
contained in beating EBs are CMs, we also immunostained
cryosections of beating EBs for cTnT. Cut sections of
beating EBs appear to contain mainly cTnT-positive cells,
confirming the percentage of cTnT-positive cells obtained
from dissociated EBs (Fig. 2d). Taken together, these
results indicate that our beating EBs are mostly composed
of CMs expressing typical cardiac markers.
Spontaneous intracellular Ca2? transients
in hESC-CMs and hiPSC-CMs are RyR2 dependent
To monitor the spontaneous intracellular Ca2? transients
and assess the functionality of the SR in hESC-CMs and
hiPSC-CMs, we performed laser scanning confocal
microscopy on enzymatically dissociated CMs. Cells were
first loaded with a fluorescent Ca2? indicator (Fluo-4AM)
to monitor these transients. Under control conditions, with
1 mM CaCl2 in the external compartment and without any
stimulation, clusters of enzymatically dissociated hESC-
CMs exhibit heterogeneous and spontaneous rhythmic
Ca2? transients (Fig. 3a, b). In several clusters, diffusion of
Ca2? transients can be seen between CMs composing a
cluster, suggesting the existence of cardiac electrical cou-
pling. When performed on enzymatically dissociated single
hESC-CMs, spontaneous Ca2? transients occur more fre-
quently on average as shown in Fig. 3d (mean frequency of
clusters of 0.15 ± 0.01/s, n = 10, versus mean frequency of
single CM of 0.38 ± 0.03/s, n = 3, P \ 0.01). A similar
phenomenon was observed using clusters of hiPSC-CMs
and single hiPSC-CMs (mean frequency of clusters of 0.13
± 0.02/s, n = 7, versus mean frequency of single CM of
0.30 ± 0.04/s, n = 6, P\0.05; Fig. 3c, d). Overall, there is
no difference in frequencies of spontaneous Ca2? transients
between hESC-CMs and hiPSC-CMs, indicating a similar
maturity for both cell types to release Ca2? spontaneously
from the SR stores.
To assess the SR Ca2? loading, we used caffeine as a RyR2
agonist. Exposure to 10 mM caffeine induces a quick eleva-
tion of Ca2? transient immediately followed by absence of
Ca2? events (Fig. 3a, b) in both types of CMs. These results
suggest that the recorded spontaneous Ca2? events are mainly
initiated by RyR2 dependent and not by inositol triphosphate
receptor (IP3R) activation that could, however, be second-
arily activated. These data also suggest that hESC-CMs and
hiPSC-CMs contain a high amount of Ca2? in the intracel-
lular SR stores that is totally depleted upon caffeine-induced
RyR2 activation, consistent with previous reports [16, 17].
hESC- and hiPSC-derived CMs exhibit consistent
and similar contractile and beating properties
We then focused on the functional maturity of our beating
EBs by measuring their mechanical properties rather than
their electrophysiologic properties, which do not imply that
the resulting CMs actually contract properly. We set up an
AFM-based method to monitor the contracting force (CF)
and beat rate (BR) properties among the homogeneous
beating EBs from hESC and hiPSC lines. Thus, by gently
touching the whole beating EB with the AFM cantilever, we
were able to measure CF and BR in physiologic solution
(Tyrode solution with 1.8 mM CaCl2 at 37 �C) (Fig. 4a).
Under basal conditions, hESC-EBs and hiPSC-EBs contract
similarly on average (31 ± 7 vs 39 ± 9 nN), both with sub-
stantial reliability (Fig. 4b). hESC-EBs exhibit a BR (i.e.,
beats per unit of time) with an average of 51 ± 5 beats/min,
which is significantly slower than the mean BR of hiPSC-EBs
Heart Vessels (2014) 29:834–846 839
123

Table 1 mRNA levels of MYH6, MYH7, and RYR2 in CCTL12 hESC-derived contracting EBs (EBc) and undifferentiated CCTL12 hESCs (SC)
Gene EBc (n = 4)b SC (n = 4)b Fold change P valuea
MYH6 4.128 ± 1.375 4.184e-005 ± 1.044e-005 98661 0.0286
MYH7 0.004092 ± 0.001419 8.057e-006 ± 6.900e-006 507 0.0286
RYR2 0.02429 ± 0.008714 0.0006365 ± 0.0002177 38 0.0286
a According to Mann–Whitney testb Expression levels (mean ± SEM) were normalized to glyceraldehyde-3-phosphate dehydrogenase expression
Fig. 2 a Gene expression in contracting CCTL12 hESC-EBs (EBc) and
undifferentiated CCTL12 hESCs (SC). MYH6 (n = 4), MYH7 (n = 4),
and RYR2 (n = 4) are overexpressed in EBc in comparison with SC.
Expression levels were normalized to glyceraldehyde-3-phosphate
dehydrogenase expression. *P \ 0.05, according to Mann–Whitney
test. b Representative immunocytochemistry of cardiac troponin T
(cTnT) in CCTL13 hESC-derived cardiomyocytes (CMs) from enzy-
matically dissociated beating EBs. The cTnT is stained in red while the
nuclear DNA is stained in blue (diamidino-2 phenylindole (DAPI)). The
scale bar represents 100 lm. c CCTL13 hESC-derived CMs from
enzymatically dissociated beating EBs visualized under higher mag-
nification. The cTnT is stained in red while the nuclear DNA is stained
in blue (DAPI). The scale bar represents 20 lm. d Cryosection of
CCTL13 hESC beating EBs stained for cTnT (green) and nuclear DNA
(blue). Both panels show the immunostaining performed on cryosec-
tions of CCTL13 hESC beating EBs. For immunostaining experiments,
negative controls were used with significantly lower signal. The scale
bar represents 50 lm
840 Heart Vessels (2014) 29:834–846
123
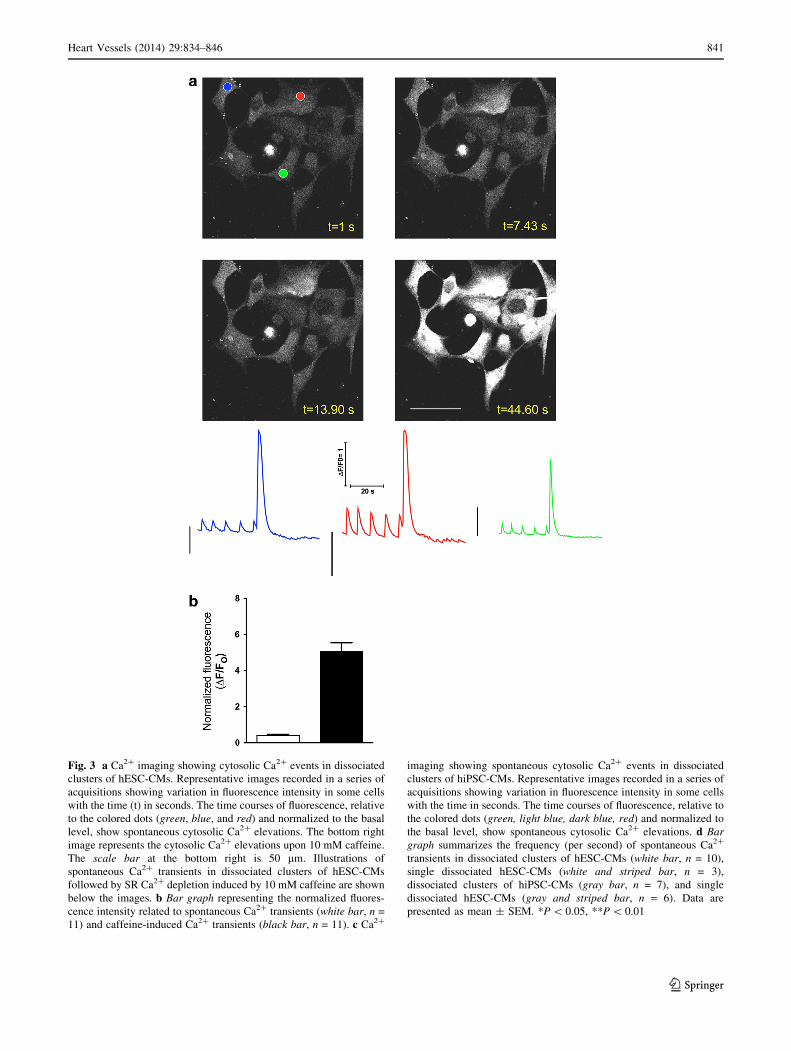
Fig. 3 a Ca2? imaging showing cytosolic Ca2? events in dissociated
clusters of hESC-CMs. Representative images recorded in a series of
acquisitions showing variation in fluorescence intensity in some cells
with the time (t) in seconds. The time courses of fluorescence, relative
to the colored dots (green, blue, and red) and normalized to the basal
level, show spontaneous cytosolic Ca2? elevations. The bottom right
image represents the cytosolic Ca2? elevations upon 10 mM caffeine.
The scale bar at the bottom right is 50 lm. Illustrations of
spontaneous Ca2? transients in dissociated clusters of hESC-CMs
followed by SR Ca2? depletion induced by 10 mM caffeine are shown
below the images. b Bar graph representing the normalized fluores-
cence intensity related to spontaneous Ca2? transients (white bar, n =
11) and caffeine-induced Ca2? transients (black bar, n = 11). c Ca2?
imaging showing spontaneous cytosolic Ca2? events in dissociated
clusters of hiPSC-CMs. Representative images recorded in a series of
acquisitions showing variation in fluorescence intensity in some cells
with the time in seconds. The time courses of fluorescence, relative to
the colored dots (green, light blue, dark blue, red) and normalized to
the basal level, show spontaneous cytosolic Ca2? elevations. d Bar
graph summarizes the frequency (per second) of spontaneous Ca2?
transients in dissociated clusters of hESC-CMs (white bar, n = 10),
single dissociated hESC-CMs (white and striped bar, n = 3),
dissociated clusters of hiPSC-CMs (gray bar, n = 7), and single
dissociated hESC-CMs (gray and striped bar, n = 6). Data are
presented as mean ± SEM. *P \ 0.05, **P \ 0.01
Heart Vessels (2014) 29:834–846 841
123

(74 ± 7 beats/min, Fig. 4c). Thus, these data indicate
consistent CF and BR and reliability in the recording
parameters, and difference in mean BR between hESC-EBs
and hiPSC-EBs (Fig. 4b, c and Supplementary Movie 1).
Furthermore, we observed a 10–20 % change in both CF
and BR values when the temperature was 37 ± 0.5 �C
while a drop in frequency of more than 50 % was observed
when the temperature was 32 �C (data not shown). This
variation is likely due to the high thermal sensitivity of the
resulting CMs forming the contracting EBs, which is
known to slow down contractions and the intracellular
Ca2? release [18–20].
Because the ‘‘fight-or-flight’’ response is an important
mechanism occurring in the heart under stress conditions,
we tested the b-adrenergic pathway in the beating EBs.
Thus, addition of metoprolol (specific b1-receptor inhibi-
tor) significantly reduces both CF and BR in comparison
with basal conditions (BC) in hiPSC-EBs (mean CF 13 ± 3
nN and mean BR 23 ± 6 beats/min, Fig. 4b, c). Similarly,
metoprolol induces a trend toward a decrease in both CF
and BR in hESC-EBs in comparison with basal conditions
(Fig. 4b, c), with no difference when compared with
hiPSC-EBs. Addition of isoproterenol (b-receptor agonist)
20 min after metoprolol induces an increase of both
Fig. 3 continued
842 Heart Vessels (2014) 29:834–846
123
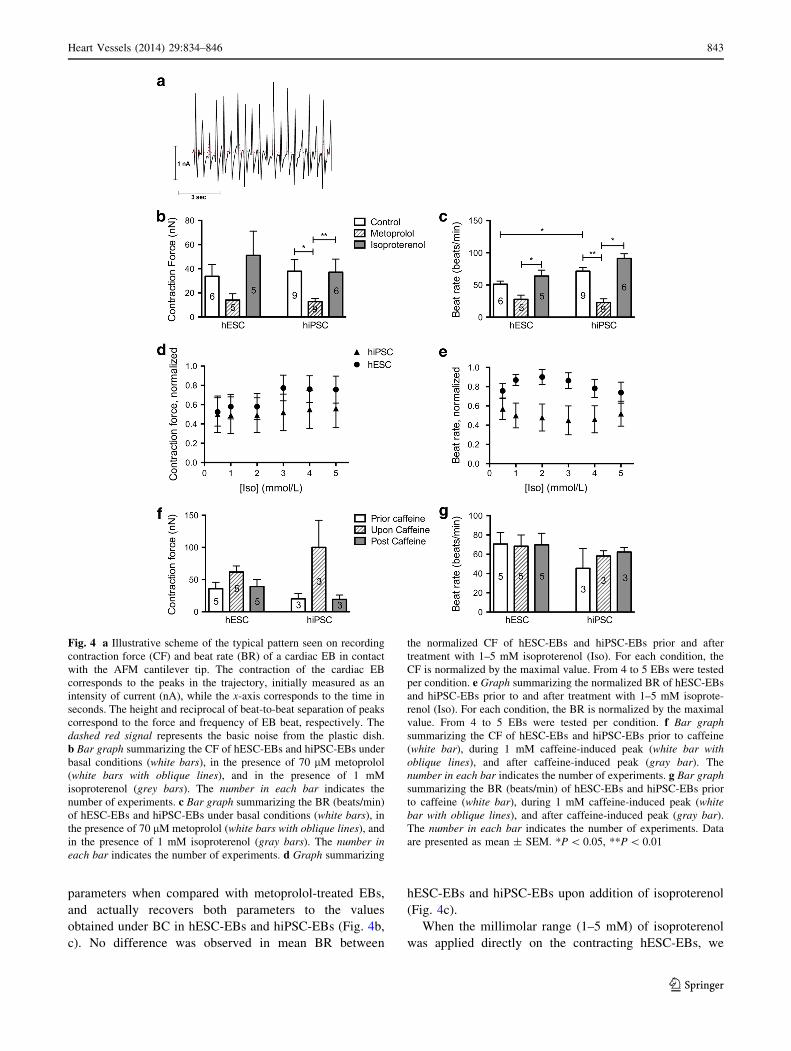
parameters when compared with metoprolol-treated EBs,
and actually recovers both parameters to the values
obtained under BC in hESC-EBs and hiPSC-EBs (Fig. 4b,
c). No difference was observed in mean BR between
hESC-EBs and hiPSC-EBs upon addition of isoproterenol
(Fig. 4c).
When the millimolar range (1–5 mM) of isoproterenol
was applied directly on the contracting hESC-EBs, we
Fig. 4 a Illustrative scheme of the typical pattern seen on recording
contraction force (CF) and beat rate (BR) of a cardiac EB in contact
with the AFM cantilever tip. The contraction of the cardiac EB
corresponds to the peaks in the trajectory, initially measured as an
intensity of current (nA), while the x-axis corresponds to the time in
seconds. The height and reciprocal of beat-to-beat separation of peaks
correspond to the force and frequency of EB beat, respectively. The
dashed red signal represents the basic noise from the plastic dish.
b Bar graph summarizing the CF of hESC-EBs and hiPSC-EBs under
basal conditions (white bars), in the presence of 70 lM metoprolol
(white bars with oblique lines), and in the presence of 1 mM
isoproterenol (grey bars). The number in each bar indicates the
number of experiments. c Bar graph summarizing the BR (beats/min)
of hESC-EBs and hiPSC-EBs under basal conditions (white bars), in
the presence of 70 lM metoprolol (white bars with oblique lines), and
in the presence of 1 mM isoproterenol (gray bars). The number in
each bar indicates the number of experiments. d Graph summarizing
the normalized CF of hESC-EBs and hiPSC-EBs prior and after
treatment with 1–5 mM isoproterenol (Iso). For each condition, the
CF is normalized by the maximal value. From 4 to 5 EBs were tested
per condition. e Graph summarizing the normalized BR of hESC-EBs
and hiPSC-EBs prior to and after treatment with 1–5 mM isoprote-
renol (Iso). For each condition, the BR is normalized by the maximal
value. From 4 to 5 EBs were tested per condition. f Bar graph
summarizing the CF of hESC-EBs and hiPSC-EBs prior to caffeine
(white bar), during 1 mM caffeine-induced peak (white bar with
oblique lines), and after caffeine-induced peak (gray bar). The
number in each bar indicates the number of experiments. g Bar graph
summarizing the BR (beats/min) of hESC-EBs and hiPSC-EBs prior
to caffeine (white bar), during 1 mM caffeine-induced peak (white
bar with oblique lines), and after caffeine-induced peak (gray bar).
The number in each bar indicates the number of experiments. Data
are presented as mean ± SEM. *P \ 0.05, **P \ 0.01
Heart Vessels (2014) 29:834–846 843
123

observed a slight increase, although nonsignificant, in both
normalized mean CF and BR when compared with basal
conditions, while a similar effect of isoproterenol was
observed in CF of hiPSC-EBs but with no significant
change in BR (Fig. 4d, e).
Taken together, these results indicate a functional b-
adrenergic pathway in both contracting hESC-EBs and
hiPSC-EBs, with no difference between the two types of
stem cell clusters.
We then sought to evaluate the effect of caffeine on the
contractile properties of the EBs, in relation with the SR
Ca2? depletion effect observed in dissociated CMs (Fig. 3).
It is believed that caffeine potentiates CF in skeletal and
cardiac muscle by penetrating the cell membrane and
potentiating Ca2? release from the SR, which contributes
to its stimulating and ionotropic effect [21–23]. When
hESC-EBs and hiPSC-EBs are treated with 1 mM caffeine,
a quick and pronounced increase in the CF immediately
occurs, followed by a progressive decrease of the CF (e.g.,
mean CF 20 ± 8 nN prior to caffeine, 100 ± 42 nN under
caffeine, 19 ± 7 nN post caffeine for hiPSC-EBs, Fig. 4f).
Interestingly, in this particular case application of 1 mM
caffeine does not change the BR (e.g., mean BR 45 ± 20
nN prior to caffeine, 58 ± 5 nN under caffeine, 62 ± 5 nN
post caffeine for hiPSC-EBs, Fig. 4g), suggesting that the
molecular mechanisms underlying the repeatable beats are
not affected by the caffeine-induced RyR2 activation.
These data indicate that caffeine quickly causes an increase
of the contraction of the CMs, due to the release of SR
Ca2? through RyR2, followed by a progressive decrease in
contraction, due to the depletion of Ca2? in the SR stores as
well as the Ca2? efflux through the Na?/Ca2? exchanger at
the sarcolemma. Thus, these data are then in agreement
with the SR Ca2? depletion and the absence of Ca2? events
after caffeine application observed by fluorescent micros-
copy in dissociated CMs (Fig. 3).
Discussion
In the this study, we present a forced aggregation method
using AggreWell 400 plates as a relatively homogeneous
starting point for stem cells. Using the same number of
cells offers the advantage to form uniformly sized and
shaped EBs. These EBs can be differentiated to beating
EBs upon application of defined factors with up to 50 %
efficiency from both hESCs and hiPSCs. Similarly to pre-
vious studies, we observed significant variability between
stem cell lines (hESC and iPSC) on differentiation in CMs
[24]. Our results indicate that the CMs composing the
beating EBs express specific cardiac markers, and we
found that in fact approximately 95 % of total number of
cells is positive for cTnT. Using fluorescent confocal
microscopy, we observed that enzymatically dissociated
CMs release Ca2? spontaneously from the intracellular SR
and mainly via the cardiac RyR2. Our results also dem-
onstrate that the resulting homogenous EBs, from both
hESC and hiPSC, can be used to investigate the contracting
force and the beating rate upon pharmacologic treatment in
a reliable manner. Thus, our results indicate that these EBs
can serve as suitable models for drug development and
testing because of their homogeneity.
We tested for the first time CCTL12, CCTL13, and
CCTL14 hESC lines [8, 9] for their cardiac differentiation
capacity. Our results indicate a substantial variability in
these lines, with the best efficiency of CM differentiation
for CCTL12 hESC line over CCTL13 and CCTL14 hESC
lines, estimated by the number of beating EBs following
treatment with specific induction media as previously
published [11]. Furthermore, the clone 4 hiPSC line used in
this study exhibits an efficiency similar to that of CCTL12
hESC, confirming its suitability for cardiac differentiation
as previously published [10].
Some groups revealed that the differentiation efficiency
is dependent of the EB size and the extracellular environ-
ment [25, 26]. Another group reported that some hESC
lines have a higher rate of cardiac differentiation in small
EBs [27]. Thus, it is reasonable to assume that a lower
amount of cells (i.e., \2000 cells per EB) might give a
higher percentage of CMs. In agreement with the afore-
mentioned published results, our preliminary and unpub-
lished data indicate that the smallest EBs developed using
the suspension method have a higher probability to beat in
comparison with the largest ones. While the suspension
method is cheaper than the AggreWell method, the
resulting EBs exhibit a high heterogeneity in shape and
size, which limits standardized protocols [28].
We also observed that the density of freshly formed EBs
is an important factor that leads to a different ratio of
‘‘attached’’ EBs versus floating EBs in low-adhesive Petri
dishes. Although counting of beating ‘‘attached’’ EBs is
easier to perform for practical reasons, they quickly lose
their three-dimensional shape. In the same way, a study has
reported that high density of EBs facilitates the cardiac
differentiation as well as the stimulation of BMP signaling
in murine ESCs [12].
It must be noted that among the different published
methods to generate human CMs through EB formation,
the composition of the media strongly varies. While the
universal system from Zambidis’ group uses, among oth-
ers, poly(vinyl alcohol), insulin and RPMI, and fetal bovine
serum (FBS) in the late phase of induction [4], in this study
we used, among others, FBS in media at early stage, activin
A, and VEGF as previously published [11]. Of note, our
EB formation medium contained FBS instead of KO-SR,
which was recently shown to improve cardiac
844 Heart Vessels (2014) 29:834–846
123

differentiation and contractility of CMs [29]. Endogenous
Wnt/b-catenin signaling is required for cardiac differenti-
ation of hESCs at an early stage of differentiation protocol
directed by BMP4 and activin A, while late inhibition of
Wnt/b-catenin pathway enhanced cardiogenesis. Thus,
while Keller’s group uses the Dickkopf 1 (Dkk1) protein as
a Wnt-signaling inhibitor through its specific affinity for
LRP6 [11], we used IWR-1, which acts as an inhibitor of
Wnt response 1. Moreover, it has been shown that the late
inhibition of Wnt signaling improves the efficiency of
BMP4 signaling-dependent cardiac differentiation from
hESCs and hiPSCs [30].
In this study, we demonstrated that the intracellular
Ca2? release through RyR2 channels appears functional in
both hESC and hiPSC lines, although further studies are
needed to fully characterize the Ca2? handling at the SR
level in these cells. For instance, other groups have shown
that the intracellular Ca2? homeostasis is immature in
hiPSC-CMs in comparison with hESC-CMs [17]. Further
evaluation of the contribution of both RyR2 and IP3R in
the calcium signaling of hESCs and hiPSCs is important,
since it is known that IP3R are highly expressed in
development and heart failure [31, 32]. Using AFM, we
verified that the inhibition of the b-adrenergic receptors
decreases the CF and BR in both hESC-CMs and hiPSC-
CMs composing the contracting EBs. The fact that
metoprolol inhibits both parameters without any prior b-
adrenergic stimulation suggests a high basal adrenergic
tonus in our hESC-EBs and hiPSC-EBs. Such a tonus
could also explain the mild effect of isoproterenol in our
experiments, in addition of the penetration rate of the
drugs into the multilayer EBs. This aspect needs further
exploration. Furthermore, we observed that caffeine
induces a trend toward increased CF but not BR in both
hESC-EBs and hiPSC-EBs, followed by a decrease. This
effect is likely due to the quick increase of cytosolic Ca2?
and by the final depletion of Ca2? from the SR stores [33,
34]. Overall, our AFM data match with those of the Ca2?
imaging upon caffeine treatment. However, it should be
considered that the effect of caffeine on EBs overall must
be relative to the accessibility of the contracting CMs
within the EB structure, also composed of extracellular
matrix.
Recently, Liu et al. [15] used a similar method to
investigate the mechanobiological properties of hESC-CMs
and hiPSC-CMs. Our results demonstrate that the EBs
formed using AggreWell 400 plates generate consistent
contractile and electrical properties with or without phar-
macologic stimulation. Of note, our results showed one-
order higher force of the EBs when compared with the
results from Liu et al. [15]. The same group has published
typical noise levels during force measurements of around
20 pN. This value is unusually low, as similar values are
usually obtained for interaction of single biomolecules.
Such a force for movement (noise connected with move-
ment) of a cell cluster is unexpected. The difference in
measurements can be due to the size of the EBs as well as
their final amount of CMs. Our data also indicate that the
beating EBs are highly sensitive to temperature variation,
suggesting that a constant temperature is a major factor in
maintaining reliability in recordings. This observation may
be related to alternative measurements that calibrate body
temperature as 1 �C per 10 % rise in heart beat (pulse-wave
frequency).
Finally, from a clinical point of view, the present study
offers a method to generate a high quantity of functional
CMs that could be potentially used for clinical application
and drug screening. Our data thus indicate the high
reproducibility of experiments on the resulting homoge-
neous EBs.
Acknowledgments We would like to thank Dr Livia Eiselleova,
Stanislava Koskova, Professor Ales Hampl, Dana Stritecka, and Eva
Peslova for their assistance, as well as Professor Majlinda Lako for
kindly providing the clone 4 hiPSC. This work was supported by
grants from the Ministry of Education, Youth, and Sports of the
Czech Republic (CZ.1.07/2.3.00/20.0011 and MSM0021622430),
project FNUSA-ICRC (no. CZ.1.05/1.1.00/02.0123) from the Euro-
pean Regional Development Fund, SoMoPro—Marie Curie
Actions—South Moravian Region, and by the European Society of
Cardiology (ESC) to Albano C. Meli. The research leading to these
results obtained a financial contribution from the European Com-
munity within the Seventh Framework Program (FP/2007-2013)
under Grant Agreement No. 229603. This work was supported by
CEITEC—Central European Institute of Technology (CZ.1.05/1.1.00/
02.0068) from the European Regional Development Fund. Albano C.
Meli was supported by a French Muscular Dystrophy Association
Research Grant (AFM). Ivana Acimovic was supported by a PLU-
RICELL grant (CZ.1.07/2.3.00/20.0011).
Conflict of interest There is no conflict of interest.
References
1. Doetschman TC, Eistetter H, Katz M, Schmidt W, Kemler R
(1985) The in vitro development of blastocyst-derived embryonic
stem cell lines: formation of visceral yolk sac, blood islands and
myocardium. J Embryol Exp Morphol 87:27–45
2. Doi K, Itoh H, Nakagawa O, Igaki T, Yamashita J, Chun TH,
Inoue M, Masatsugu K, Nakao K (1997) Expression of natriuretic
peptide system during embryonic stem cell vasculogenesis. Heart
Vessels Suppl 12:18–22
3. Burridge PW, Anderson D, Priddle H, Barbadillo Munoz MD,
Chamberlain S, Allegrucci C, Young LE, Denning C (2007)
Improved human embryonic stem cell embryoid body homoge-
neity and cardiomyocyte differentiation from a novel V-96 plate
aggregation system highlights interline variability. Stem Cells
25:929–938
4. Burridge PW, Thompson S, Millrod MA, Weinberg S, Yuan X,
Peters A, Mahairaki V, Koliatsos VE, Tung L, Zambidis ET
Heart Vessels (2014) 29:834–846 845
123

(2011) A universal system for highly efficient cardiac differen-
tiation of human induced pluripotent stem cells that eliminates
interline variability. PLoS One 6:e18293
5. Kattman SJ, Witty AD, Gagliardi M, Dubois NC, Niapour M,
Hotta A, Ellis J, Keller G (2011) Stage-specific optimization of
activin/nodal and BMP signaling promotes cardiac differentiation
of mouse and human pluripotent stem cell lines. Cell Stem Cell
8:228–240
6. Xu XQ, Graichen R, Soo SY, Balakrishnan T, Rahmat SN, Sieh
S, Tham SC, Freund C, Moore J, Mummery C, Colman A,
Zweigerdt R, Davidson BP (2008) Chemically defined medium
supporting cardiomyocyte differentiation of human embryonic
stem cells. Differentiation 76:958–970
7. Yang L, Soonpaa MH, Adler ED, Roepke TK, Kattman SJ,
Kennedy M, Henckaerts E, Bonham K, Abbott GW, Linden RM,
Field LJ, Keller GM (2008) Human cardiovascular progenitor
cells develop from a KDR ? embryonic-stem-cell-derived pop-
ulation. Nature 453:524–528
8. Dvorak P, Dvorakova D, Koskova S, Vodinska M, Najvirtova M,
Krekac D, Hampl A (2005) Expression and potential role of
fibroblast growth factor 2 and its receptors in human embryonic
stem cells. Stem Cells 23:1200–1211
9. Eiselleova L, Peterkova I, Neradil J, Slaninova I, Hampl A,
Dvorak P (2008) Comparative study of mouse and human feeder
cells for human embryonic stem cells. Int J Dev Biol 52:353–363
10. Armstrong L, Tilgner K, Saretzki G, Atkinson SP, Stojkovic M,
Moreno R, Przyborski S, Lako M (2010) Human induced plu-
ripotent stem cell lines show stress defense mechanisms and
mitochondrial regulation similar to those of human embryonic
stem cells. Stem Cells 28:661–673
11. Dubois NC, Craft AM, Sharma P, Elliott DA, Stanley EG,
Elefanty AG, Gramolini A, Keller G (2011) SIRPA is a specific
cell-surface marker for isolating cardiomyocytes derived from
human pluripotent stem cells. Nat Biotechnol 29:1011–1018
12. Lee MY, Cagavi Bozkulak E, Schliffke S, Amos PJ, Ren Y, Ge
X, Ehrlich BE, Qyang Y (2011) High density cultures of
embryoid bodies enhanced cardiac differentiation of murine
embryonic stem cells. Biochem Biophys Res Commun 416:51–57
13. Fujita E, Nakanishi T, Nishizawa T, Hagiwara N, Matsuoka R
(2013) Mutations in the cardiac troponin T gene show various
prognoses in Japanese patients with hypertrophic cardiomyopa-
thy. Heart Vessels. doi:10.1007/s00380-013-0332-3
14. Zwi-Dantsis L, Huber I, Habib M, Winterstern A, Gepstein A,
Arbel G, Gepstein L (2013) Derivation and cardiomyocyte dif-
ferentiation of induced pluripotent stem cells from heart failure
patients. Eur Heart J 34:1575–1586
15. Liu J, Sun N, Bruce MA, Wu JC, Butte MJ (2012) Atomic force
mechanobiology of pluripotent stem cell-derived cardiomyo-
cytes. PLoS One 7:e37559
16. Itzhaki I, Rapoport S, Huber I, Mizrahi I, Zwi-Dantsis L, Arbel G,
Schiller J, Gepstein L (2011) Calcium handling in human induced
pluripotent stem cell derived cardiomyocytes. PLoS One 6:e18037
17. Lee YK, Ng KM, Lai WH, Chan YC, Lau YM, Lian Q, Tse HF,
Siu CW (2011) Calcium homeostasis in human induced pluripo-
tent stem cell-derived cardiomyocytes. Stem Cell Rev 7:976–986
18. Liu B, Wohlfart B, Johansson BW (1990) Effects of low tem-
perature on contraction in papillary muscles from rabbit, rat, and
hedgehog. Cryobiology 27:539–546
19. Fu Y, Zhang GQ, Hao XM, Wu CH, Chai Z, Wang SQ (2005)
Temperature dependence and thermodynamic properties of Ca2?
sparks in rat cardiomyocytes. Biophys J 89:2533–2541
20. Wang SQ, Huang YH, Liu KS, Zhou ZQ (1997) Dependence of
myocardial hypothermia tolerance on sources of activator cal-
cium. Cryobiology 35:193–200
21. Degubareff T, Sleator W Jr (1965) Effects of caffeine on mam-
malian atrial muscle, and its interaction with adenosine and cal-
cium. J Pharmacol Exp Ther 148:202–214
22. Axelsson J, Thesleff S (1958) Activation of the contractile
mechanism in striated muscle. Acta Physiol Scand 44:55–66
23. Weber A, Herz R (1968) The relationship between caffeine
contracture of intact muscle and the effect of caffeine on retic-
ulum. J Gen Physiol 52:750–759
24. Bellin M, Marchetto MC, Gage FH, Mummery CL (2012)
Induced pluripotent stem cells: the new patient? Nat Rev Mol
Cell Biol 13:713–726
25. Bratt-Leal AM, Carpenedo RL, McDevitt TC (2009) Engineering
the embryoid body microenvironment to direct embryonic stem
cell differentiation. Biotechnol Prog 25:43–51
26. Mohr JC, Zhang J, Azarin SM, Soerens AG, de Pablo JJ,
Thomson JA, Lyons GE, Palecek SP, Kamp TJ (2010) The mi-
crowell control of embryoid body size in order to regulate cardiac
differentiation of human embryonic stem cells. Biomaterials
31:1885–1893
27. Bauwens CL, Peerani R, Niebruegge S, Woodhouse KA,
Kumacheva E, Husain M, Zandstra PW (2008) Control of human
embryonic stem cell colony and aggregate size heterogeneity
influences differentiation trajectories. Stem Cells 26:2300–2310
28. Antonchuk J (2013) Formation of embryoid bodies from human
pluripotent stem cells using AggreWell plates. Methods Mol Biol
946:523–533
29. Preda MB, Burlacu A, Simionescu M (2013) Defined-size
embryoid bodies formed in the presence of serum replacement
increases the efficiency of the cardiac differentiation of mouse
embryonic stem cells. Tissue Cell 45:54–60
30. Ren Y, Lee MY, Schliffke S, Paavola J, Amos PJ, Ge X, Ye M,
Zhu S, Senyei G, Lum L, Ehrlich BE, Qyang Y (2011) Small
molecule Wnt inhibitors enhance the efficiency of BMP-4-
directed cardiac differentiation of human pluripotent stem cells.
J Mol Cell Cardiol 51:280–287
31. Gutstein DE, Marks AR (1997) Role of inositol 1,4,5-trisphos-
phate receptors in regulating apoptotic signaling and heart failure.
Heart Vessels Suppl 12:53–57
32. Satin J, Itzhaki I, Rapoport S, Schroder EA, Izu L, Arbel G,
Beyar R, Balke CW, Schiller J, Gepstein L (2008) Calcium
handling in human embryonic stem cell-derived cardiomyocytes.
Stem Cells 26:1961–1972
33. Shibata S, Hollander PB (1967) Effects of caffeine on the con-
tractility and membrane potentials of rat atrium. Experientia
23:559
34. Shinohara T, Kim D, Joung B, Maruyama M, Vembaiyan K,
Back TG, Wayne Chen SR, Chen PS, Lin SF (2013) Carvedilol
analog modulates both basal and stimulated sinoatrial node
automaticity. Heart Vessels doi. doi:10.1007/s00380-013-0378-2
846 Heart Vessels (2014) 29:834–846
123
Related Documents











