Journal of Animal Ecology 2007 76, 801–813 © 2007 The Authors. Journal compilation © 2007 British Ecological Society Blackwell Publishing Ltd Foraging behaviour of a frugivorous bat helps bridge landscape connectivity and ecological processes in a fragmented rainforest MICKAËL HENRY, JEAN-MARC PONS* and JEAN-FRANÇOIS COSSON† Département Ecologie et Gestion de la Biodiversité, MNHN/CNRS, UMR 5176, 4, av. du petit château, F-91800 Brunoy, France; * Origine, structure et évolution de la biodiversité (UMR 5202), Département Systématique et Evolution, Muséum National d’Histoire Naturelle, Case Postale 51, 55 rue Buffon, F-75005 Paris, France; and † INRA-EFPA, UMR Centre de Biologie et de Gestion des Populations (CBGP), Campus International de Baillarguet, CS 30016, F-34988 Montferrier/Lez cedex, France Summary 1. Landscape connectivity may greatly influence the distribution of animals when it alters their movements and their ability to reach food patches. Depending on their foraging behaviour, organisms may or may not adapt to anthropogenic changes in landscape connectivity and may eventually undergo local extinctions. 2. Recent studies underlined the need to use indicators of functional landscape connectivity based on the behaviour and movement abilities of studied animals to better link landscape structure to ecological processes in disturbed and fragmented areas. 3. The objectives of this study were: to elaborate an index of functional connectivity for Rhinophylla pumilio, a Neotropical understorey frugivorous bat; to use this index to investigate the possible mechanisms controlling its distribution and sustainability in a fragmented landscape; and to test whether this index could be applied to other species of the same guild. 4. We pursued a 10-year bat mist-net survey, coupled to local estimates of food availability, in a mature forest of French Guiana that was recently fragmented by the completion of a reservoir lake. The 18 sampling sites range from undisturbed continuous forest sites to small remote forest fragments. A connectivity value, based on radio-tracking surveys, was attributed to each site. Connectivity measures mean forest cover within neighbouring landscape units, weighted by the probability that bats would use them, as estimated by frequency distribution of flight distance data. 5. The abundance of R. pumilio was positively correlated with landscape connectivity and not correlated with local food availability. Its foraging strategy has evolved in response to the highly scattered distribution of its fruit resource. In spite of its high mobility, R. pumilio apparently failed to exploit a food resource that is distributed patchily over a low-connective habitat because its foraging movements are not well adapted to habitat disruptions. 6. The connectivity index contributed to explain general tendencies of abundance variations in other understorey frugivorous bats, although the spatial scale we examined was probably too small for these species. We make recommendations to adapt a functional connectivity index to species whose large-scale movements are difficult to survey. Key-words: forest fragmentation, functional connectivity, landscape ecology, phyllostomid frugivorous bats, structural connectivity. Journal of Animal Ecology (2007) 76, 801–813 doi: 10.1111/j.1365-2656.2007.01258.x Correspondence: M. Henry, Centro de Investigaciones en Ecosistemas, Universidad National Autónoma de México, Apartado Postal 27–3 (Xangari), Morelia, Michoacán, Mexico. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Animal Ecology
2007
76
, 801–813
© 2007 The Authors.Journal compilation© 2007 British Ecological Society
Blackwell Publishing Ltd
Foraging behaviour of a frugivorous bat helps bridge landscape connectivity and ecological processes in a fragmented rainforest
MICKAËL HENRY, JEAN-MARC PONS* and JEAN-FRANÇOIS COSSON†
Département Ecologie et Gestion de la Biodiversité, MNHN/CNRS, UMR 5176, 4, av. du petit château, F-91800 Brunoy, France;
*
Origine, structure et évolution de la biodiversité (UMR 5202), Département Systématique et Evolution, Muséum National d’Histoire Naturelle, Case Postale 51, 55 rue Buffon, F-75005 Paris, France; and
†
INRA-EFPA, UMR Centre de Biologie et de Gestion des Populations (CBGP), Campus International de Baillarguet, CS 30016, F-34988 Montferrier/Lez cedex, France
Summary
1.
Landscape connectivity may greatly influence the distribution of animals when italters their movements and their ability to reach food patches. Depending on theirforaging behaviour, organisms may or may not adapt to anthropogenic changes inlandscape connectivity and may eventually undergo local extinctions.
2.
Recent studies underlined the need to use indicators of functional landscapeconnectivity based on the behaviour and movement abilities of studied animals to betterlink landscape structure to ecological processes in disturbed and fragmented areas.
3.
The objectives of this study were: to elaborate an index of functional connectivity for
Rhinophylla pumilio
, a Neotropical understorey frugivorous bat; to use this index toinvestigate the possible mechanisms controlling its distribution and sustainability in afragmented landscape; and to test whether this index could be applied to other speciesof the same guild.
4.
We pursued a 10-year bat mist-net survey, coupled to local estimates of foodavailability, in a mature forest of French Guiana that was recently fragmented by thecompletion of a reservoir lake. The 18 sampling sites range from undisturbed continuousforest sites to small remote forest fragments. A connectivity value, based on radio-trackingsurveys, was attributed to each site. Connectivity measures mean forest cover withinneighbouring landscape units, weighted by the probability that bats would use them, asestimated by frequency distribution of flight distance data.
5.
The abundance of
R. pumilio
was positively correlated with landscape connectivityand not correlated with local food availability. Its foraging strategy has evolved inresponse to the highly scattered distribution of its fruit resource. In spite of its highmobility,
R. pumilio
apparently failed to exploit a food resource that is distributedpatchily over a low-connective habitat because its foraging movements are not welladapted to habitat disruptions.
6.
The connectivity index contributed to explain general tendencies of abundancevariations in other understorey frugivorous bats, although the spatial scale we examinedwas probably too small for these species. We make recommendations to adapt afunctional connectivity index to species whose large-scale movements are difficult tosurvey.
Key-words
: forest fragmentation, functional connectivity, landscape ecology,phyllostomid frugivorous bats, structural connectivity.
Journal of Animal Ecology
(2007)
76
, 801–813 doi: 10.1111/j.1365-2656.2007.01258.x
Correspondence: M. Henry, Centro de Investigaciones en Ecosistemas, Universidad National Autónoma de México, ApartadoPostal 27–3 (Xangari), Morelia, Michoacán, Mexico. E-mail: [email protected]

802
M. Henry, J.-M. Pons & J.-F. Cosson
© 2007 The Authors.Journal compilation© 2007 British Ecological Society,
Journal of Animal Ecology
,
76
, 801–813
Introduction
One of the most crucial concerns of tropical ecologistsis to assess the impact of habitat loss and fragmentationon species diversity, as well as the underlying ecologicalprocesses leading to local species extinctions (Stouffer& Bierregaard 1995a,b; Laurance & Bierregaard 1997;Gascon
et al
. 1999; Laurance
et al
. 2002; Ewers &Didham 2006). Habitat fragmentation may affectanimal species’ densities through a variety of direct andindirect mechanisms, such as pure habitat loss, increas-ing edge effect and associated microclimatic changeson small fragments, isolation of habitat fragments,or greater extinction risks of small populations(Saunders, Hobbs & Margules 1991; Andrén 1994). Tosatisfy the needs of both theoretical and appliedconservation biology, various hypotheses have beenproposed to explain the density and distribution ofanimal species within fragmented landscapes. Earlyhypotheses were based on the equilibrium theory ofisland biogeography (MacArthur & Wilson 1967),stating that as in true islands, the occurrence of speciesin habitat fragments is a function of fragment area andisolation distance from the mainland (Simberloff 1988).
However, the theory of island biogeography has itsown limits, and the ‘island metaphor’, consisting intreating habitat fragments as islands embedded in aninhospitable matrix, gives imperfect insights about thedistribution of organisms in fragmented areas. Bordershape of both fragments and mainland may alsogreatly influence colonization and dispersal rates,independently from fragment size or isolation distance(Taylor 1987a,b). Moreover, the occurrence of somespecies in fragments may depend more on dispersalmovements from other fragments rather than from themainland (Fahrig & Merriam 1994), making the notionof isolation distance difficult to apply. Conversely,some mainland areas bordering the inhospitablematrix are likely to undergo noticeable faunal changesas well, due to habitat disruptions, edge effect, or invad-ing species. Thus, whenever possible, it is important toconsider ecological processes along a continuousgradient of landscape disturbance rather than in a patch-based approach. Finally, the assumption that the matrixsurrounding fragments is inhospitable regarding thefocal species is often violated (Norton, Hannon &Schmiegelow 2000; Ricketts 2001; Brotons, Mönkkönen& Martin 2003; Kupfer, Malanson & Franklin 2006).For instance, forest birds may find valuable foodresources in pastures or logged areas surrounding theirnatural habitat, leading to greater population densitiesthan expected in fragmented areas (Brotons, Herrando& Martin 2004; Brotons
et al
. 2005). Therefore, it is ofspecial interest to control for possible mitigating effectsrelated to matrix quality.
The recent discipline of landscape ecology, usingGIS-based analyses of landscape structure, offerssolutions to side-step these limitations. Landscapeanalyses can provide integrative measurements of
spatial heterogeneity, combining many environmentalvariables and their spatial interactions. Among thedescriptors of landscape structure, landscape connec-tivity may provide valuable insights for understandingecological processes related to habitat fragmentation.Landscape connectivity describes the degree to whichthe landscape facilitates or impedes movements ofanimals among resource patches (Taylor
et al
. 1993)and may be used to predict the distribution of theirmovements and activity in fragmented areas. Landscapeconnectivity has been successfully linked to ecologicalprocesses in an increasing number of studies (With,Gardner & Turner 1997), including analyses of dispersaldistances and genetic relatedness among animalindividuals or populations (Manel
et al
. 2003; Coulon
et al
. 2004).More recently, Bélisle (2005) reviewed the notions of
structural and functional connectivity. On one hand,landscape parameters that measure ‘the degree towhich some landscape elements of interest are contiguousor physically linked to one another’ refer to structuralconnectivity (With
et al
. 1997; Tischendorf & Fahrig2000). On the other hand, landscape parameters thattake into account dispersal or movement abilities ofthe studied organisms – i.e. that measure landscapeconnectivity with respect to how organisms mayperceive landscape – refer to functional connectivity.Species-specific studies would gain in efficiency andapplicability if behaviour-based functional connectivitywas favoured. Without any tight link to behaviour, it isnecessary to consider in connectivity analyses severalto many landscape descriptors, as well as their respectiveinteractions. Furthermore, several trials may be necessaryto determine which spatial scale is the most relevant forthe target species. Overall, batteries of nonindependentanalyses repeated at different spatial scales may berequired. This increases risks of misinterpretations dueto type I statistical errors (null hypothesis rejected whileit is true), or even to type II errors (null hypothesisaccepted while it is false) if probability values arecorrected for too many tests.
The objectives of this study were: (1) to develop anindex of functional landscape connectivity for aNeotropical understorey frugivorous bat
Rhinophyllapumilio
Peters (Phyllostomidae), whose foragingpattern has been well described (Henry & Kalko 2007);(2) to test whether functional connectivity, jointly withresource availability, may contribute to explain thedistribution and sustainability of these bats within afragmented rainforest; and (3) to assess whether thefunctional connectivity index may be also applied tothe other species of the guild of understorey frugivorousbats, including the genera
Carollia
and
Sturnira
(Phyllostomidae). These understorey frugivorous batsact as important seed dispersers in tropical forests (Henry& Jouard 2007) and are sensitive to modifications oflandscape structure (Estrada, Coates-Estrada &Merrit 1993; Brosset
et al
. 1996; Kalko 1998; Schulze,Seavy & Whitacre 2000; Estrada & Coates-Estrada

803
Frugivorous bats and landscape connectivity
© 2007 The Authors.Journal compilation© 2007 British Ecological Society,
Journal of Animal Ecology
,
76
, 801–813
2001, 2002; Faria 2006). Previous studies (Gorresen &Willig 2004) succeeded to link the abundance of frugiv-orous bats to various indicators of structural landscapeconnectivity (e.g. forest cover, densities of fragments).In this study, we further introduce keystone behaviouralcomponents to develop an indicator of functionallandscape connectivity.
Understorey frugivorous bats feed on well-scatteredfood items, mainly small fruits produced in smallamounts and for extended periods of time (‘steady-state’ fruit crops; Kalko 1998; Thies & Kalko 2004) byunderstorey plants. This forces bats to be constantlyengaged in flights devoted to food search, and to rarelycommute over long distances (Fleming, Heithaus &Sawyer 1977; Heithaus & Fleming 1978; Thies, Kalko& Schnitzler 2006; Henry & Kalko 2007). This foragingbehaviour appears incompatible with the patchydistribution of resources in fragmented forests. Thus,we hypothesized that the distribution of understoreyfruit bats was more dependent on the degree oflandscape connectivity (ensuring spatial continuityof resource distribution) than on local conditions ofresource availability.
For that purpose, we pursued a long-term bat survey,coupled with local estimates of resource availability,that was initiated 10 years ago in Saint-Eugène, FrenchGuiana (Granjon
et al
. 1996; Cosson, Pons & Masson1999a; Cosson
et al
. 1999b; Pons & Cosson 2002). Inthis study area, forest fragments are land bridge islandsrecently isolated from the continuous forest by areservoir lake. Therefore, one can consider that thelandscape is composed of two elements only – themature forest and an aquatic matrix devoid of fruitresources that could modify the local conditions ofresource availability and spatial continuity (Leigh
et al
.2002). After validating a connectivity modelling in thissimplified system, one may transpose it into morecomplex landscape mosaics with matrix habitats ofvarious quality.
Methods
Surveys were undertaken at the Saint-Eugène studyarea (4
°
51
′
N, 53
°
04
′
W), northern French Guiana. Themature forest surrounding Saint-Eugène was fragmentedby the creation of the Petit-Saut hydroelectric dambuilt on the Sinnamary River, 60 km downstream fromthe study area in early 1994 (Cosson
et al
. 1999a). Thesubsequent flooding transformed 465 km
2
of continuousforest into a reservoir lake covered by 100 km
2
of tinyforested islands, mostly < 10 ha in area (Claessens
et al
. 2002). Total annual rainfall averages 3250 mm,with a main dry season from August to November anda shorter, less marked one in early March.
R. pumilio
is the most abundant understorey fru-givorous bat in this area, the other species from the guildbeing
Carollia brevicauda
(Schintz),
C. perspicillata
(Linnaeus) and
Sturnira tildae
(E. Geoffroy) (Cosson
et al
. 1999a). All bat surveys were conducted during 1–1·5-month sessions during main dry seasons of years1995–97 and 2002–04. These two periods corresponded,respectively, to second to fourth and ninth to 11th yearsfollowing fragmentation. They were termed periods of‘recent’ and ‘older’ fragmentation, respectively.
We selected four mainland sites and 14 islands (size0·8–7·5 ha, Fig. 1) surveyed at least twice in 1995–97(Cosson
et al
. 1999a; Cosson & Pons, unpublished data),and repeated the same sampling methods in 2002–04.The 18 capture sites are located within a 4
×
4-km areaencompassing a portion of the flooded lake and theadjacent mainland. A pre-fragmentation survey in1993–94 ensured that fruit bat abundances andcommunities were homogeneous across the study areabefore fragmentation (Cosson
et al
. 1999a). Withineach site, bats were captured simultaneously at two tofive capture stations located at least 50 m apart and atleast 20 m away from the shoreline. Each capturestation consisted of a group of three mist nets (12
×
2·5 m,mesh 38 mm) set at ground level in T pattern whenpossible, or in line otherwise. To avoid biases due totrap-shy behaviour of bats, each site was sampled asingle night per field session, but four to six times out ofthe six field sessions. From one field session to the next,we tried to set stations at the same place, but this wasoften difficult due to tree falls and vegetation regrowthon trails that remained unattended between fieldsessions. The spatial extent occupied by capturestations at the mainland sites roughly equalled that inislands (except for island 22, which was too small toaccommodate more than two capture stations).
Fig. 1. Study area map showing the location of the fragmentswhere the radio-tracking survey was undertaken, as well as the18 capture sites: four mainland sites (letters) and 14 forestedfragments (numbers).

804
M. Henry, J.-M. Pons & J.-F. Cosson
© 2007 The Authors.Journal compilation© 2007 British Ecological Society,
Journal of Animal Ecology
,
76
, 801–813
During capture nights, nets were opened from duskto dawn (18.30–06.30 h) and were continuouslychecked during the first and the last 2 h of the night,and every 2 h otherwise. Capture interruptions due toheavy rain were rare and short because we workedduring dry seasons. No netting was done during the 6-day periods encompassing the full moon owing topossible slowdown in bat captures due to lunar phobia(Lang
et al
. 2006). Captured bats were kept in clothbags before being identified to species following a keyderived from Charles-Dominique, Brosset & Jouard(2001) and Simmons & Voss (1998), and released oncapture sites.
In mature forests of French Guiana, the differentunderstorey fruit bat species consume the same fruitresources but in different proportions. To quantifyfood available to understorey fruit bats, we focused onseveral epiphytes (Cyclanthaceae and
Philodendron
spp., Araceae) and on
Piper
(Piperaceae) plantsthat are known to constitute keystone resources for
R. pumilio
and
Carollia
spp., respectively (Fleming1982, 1985; Cosson 1994; Cockle 1997; Charles-Dominique & Cockle 2001; Thies & Kalko 2004;Delaval, Henry & Charles-Dominique 2005).
Sturnira
spp. consumes mostly
Solanum
fruits within forestedges and second growth in lowland disturbed rainforestsof Guiana and Amazon (Marinho-Filho 1991; Cosson1994; Lobova & Mori 2004). These plants are scarceand probably restricted to large tree fall gaps withinundisturbed rainforests. They were so rarely encoun-tered in our study sites that we could not assess theirdensity. However,
S. tildae
mostly feeds on epiphytesand
Piper
sp. within undisturbed rainforests (Cosson1994; Cockle 1997; Charles-Dominique & Cockle2001; Delaval
et al
. 2005), making our food availabilitysurvey appropriate for it as well.
Plant resources were censused within four to five200-m
2
plots (5
×
40 m) per site, uniformly distributedalong capture stations. Botanical surveys occurred inNovember 1999 and 2004 and were associated withrecent and older fragmentation periods, respectively.
Only shrubby
Piper
individuals > 50 cm tall werecounted, i.e. the estimated minimum size required forfruit production. To better estimate potential fruitproduction on each
Piper
individual, we counted thenumber of terminal branches that could potentiallybear fruits, and the number of flowering and fruitingspikes.
Piper
spikes ripen in the evening and are gener-ally removed by bats within the following hours or elsefall off in the morning (Thies & Kalko 2004), whichrendered it difficult to introduce removal rate as anadditional variable in our analyses.
Epiphyte infructescences constitute the main diet of
R. pumilio
and are frequently consumed by
Carollia
and
Sturnira
species in French Guiana. Followingdiet descriptions provided by Cockle (1997), we
concentrated our interest on
Asplundia heteranthera
,
Evodianthus funifer
and
Thoracocarpus bissectus
(Cyclanthaceae), and several
Philodendron
species(Araceae), including
P. billietae
,
P. grandifolium
,
P.insigne
,
P. linnaei
,
P. pedatum
,
P. squamiferum
and
P.
subgenus
Pteromischum
(
P. duckei
,
P. guianense
,
P.placidum
). Most of these are epiphytes whose adventi-tious roots develop on trunks, at understorey tosubcanopy levels (1–8 m above ground). Therefore, wecould visually census adult individuals with reasonableaccuracy, but the presence of fruits could not bedocumented as in
Piper
.
To describe the local level of forest connectivity at eachcapture site, we used two landscape descriptors derivedfrom Hewison
et al
. (2001) and Coulon
et al
. (2004), alandscape connectivity index and a habitat remotenessindex. The connectivity index measures the extent offorest cover within a certain radius around capturesites, while the remoteness index is an indicator ofisolation measuring the potential difficulty for bats toreach this site from the nearest area of continuousforest. These calculations were based on a SPOTsatellite image (resolution 20 m) of the study area takenin 1996, transformed into a binary map (water vs. foresthabitat) of 250
×
250 pixels and exported as a binarytext matrix using the software ImageJ 1·33u (NationalInstitutes of Health, USA; URL: http://rsb.info.nih.gov/ij). Connectivity and remoteness values werecalculated in three steps (Fig. 2). First, we assigned to
Fig. 2. Treatment of the map for calculation of the landscapeconnectivity index (CI) and remoteness index. Step 1: initialbinary map where each pixel (or landscape unit) can take onlytwo values, 0 for ‘water’ and 200 for ‘forest’. Step 2, weassigned to each landscape unit the mean value of allneighbouring units located within a 400-m radius. Step 3 is asimilar process, but values of neighbouring units in step 2 areweighted by a coefficient depending on their respective distanceto the considered landscape unit (see Results, Fig. 3) to transformthe step 2 matrix into the CI matrix. Upper graph shows the CIprofile of all landscape units transected by the segment [AB]bridging island 22 to the nearest landscape unit of maximumCI. Lower graph is the same, but showing the reverse values 1/(CI + 1) that are summed to produce the final remoteness index.

805
Frugivorous bats and landscape connectivity
© 2007 The Authors.Journal compilation© 2007 British Ecological Society,
Journal of Animal Ecology
,
76
, 801–813
each landscape unit, i.e. each map pixel correspondingto a 20
×
20 m plot, an arbitrary suitability value equal-ling 0 or 200, for water and forested units, respectively.Second, we computed for each landscape unit the meansuitability value of all neighbouring landscape unitswithin a given radius. A mean value of 200 indicatesthat the considered landscape unit is completelysurrounded by forested units within the chosen radius,while a value of 100 indicated that only half of thelandscape units are forested. These mean values denotethe proportion of forest cover within the chosen radiusbut do not take into account the size of each forestisland. To overcome this limitation, we assigned higherweighting coefficients to closer landscape units, andlower weighting coefficients to farther ones, for thecalculation of mean suitability values (see below). Theresulting forest connectivity index ranges from 0 (noforested habitat within the chosen radius) to 200 (onlyforested habitat) and decreases sharply when forestedhabitat becomes scarce in the immediate vicinity of theconsidered landscape unit.
Once the connectivity matrix was computed, weassigned to each capture site the connectivity index(CI) of their central landscape unit. The remotenessindex (RI) was calculated as the sum of the inverse [1/(CI + 1)] of all landscape units a bat has to cross whenreaching the capture site by flying in a straight line fromthe nearest landscape unit of maximum connectivityindex (CI = 200). Thus, RI equals
c.
0 when the capturesite has a CI = 200, and otherwise increases sharplywhen isolated by large numbers of landscape units withCI = 0.
Following this method, two important parametersare needed for CI and RI calculations, namely (1) thelength of the radius enclosing the so-called neighbouringlandscape units, and (2) the mathematical functiondetermining the distance-dependent weighting coeffi-cients assigned to these neighbouring units. To defineweighting coefficients, we sought a mathematicalfunction describing the frequency distribution ofminimal distances bats cover when they search forfood. Search flights devoted to food localization are animportant component of foraging activity of understoreyfruit bats (Fleming
et al
. 1977; Henry & Kalko 2007),and their lengths give an indication of the extent ofhabitat bats can investigate in a single flight to findfruiting plants. The longest distances bats cover in asingle flight set the radius length, i.e. the distance afterwhich weighting coefficients would become zero.Detailed flight distance data of
R. pumilio
wereobtained by radio-tracking surveys.
R
.
P U M I L I O
To assess flight distances, we radio-tracked five individ-uals. The radio-tracking survey was carried out inDecember 1995 on a small 7·5-ha fragment (one maleand one female) and in November 1999 on a larger 28-ha fragment (two males and one female) (Fig. 1). The
latter, rather flat, fragment was crossed by numeroustrails forming a 100-m spacing grid, which greatlyfacilitated radio-tracking. Bats were mist-netted onthese fragments and fitted with 0·70-g radio-transmitters(Biotrack, Dorset, UK) representing < 7·5% of theirbody mass. Transmitters were attached to the back ofbats using surgical SkinBound® (Smith and NephewInc., Mississauga, Ontario, Canada) after a smallamount of dorsal fur was trimmed. Bats were releasedat the capture site within 30 min after capture. Notracking data were taken on the capture night to avoidbiases resulting from stress response to manipulation.Bats were tracked afterward for 3–4 nights (19.00–06.00 h) by two observers in radio-contact and eachequipped with a CE-12 receiver (Custom Electronics,Urbana, IL, USA) and a four-element Yagi antenna.Radio-tracking nights involved determining wheneverpossible the hanging locations of bats by triangulation.Bats were considered to be hanging when the signalintensity was judged constant in direction and intensityfor at least 1 min. Triangulation data were computedand analysed with the software Tracker 1·1 (CamponotusAB, Solna, Sweden 1994) after invalid bat hanginglocations were discarded, i.e. points > 400 m away fromobserver positions (maximal estimated detection range).Flight distances are defined by the linear distancebetween each pair of successive hanging locationsvisited by bats. As discussed in Henry & Kalko (2007),the particular diet of
R. pumilio
in our study area,mostly epiphyte fruits, makes search flights easy tomonitor. As epiphyte infructescences are too large tobe removed and transported in feeding roosts, batsconsume them
in situ
(Cockle 1997). Therefore, a partof the hanging locations reported by radio-tracking on
R. pumilio
may correspond to the location of consumedfruiting plants.
To improve the reliability of the mathematical mod-elling of the flight distance frequency distribution, wesupplemented our data set with similar data collectedon six nonreproductive individuals of
R. pumilio
usingthe same methodology (two males and four females;Henry 2005; Henry & Kalko 2007) in the undisturbedforest of the Nouragues natural reserve (110 km south-east from Saint-Eugène; Charles-Dominique 2001).Flight distance data were compared among study areasusing a nested
to measure to what extent datasets should be considered separately to produce twodistinct series of connectivity index.
The purpose of our study was to discriminate betweenthe respective contributions of landscape connectivity,food resource availability and age of fragmentation inexplaining the local abundance (capture rate) of
R.pumilio
, and to determine if the same connectivity indexcould be applied in a similar way to the other understoreyfrugivorous bats. Owing to low capture rates, data onthe other understorey frugivorous bats were pooled

806
M. Henry, J.-M. Pons & J.-F. Cosson
© 2007 The Authors.Journal compilation© 2007 British Ecological Society,
Journal of Animal Ecology
,
76
, 801–813
together and referred to as ‘shrub-frugivorous bats’to mark the contrast with
R. pumilio
that is ratherspecialized on epiphytes.
Raw data of capture rate (number of captures perunit of capture effort) were strongly asymmetrical dueto low capture rate in most of forest fragments. Instead,we used as indicators of bat abundance the residualvalues of the linear regression of capture numbersagainst capture effort (expressed in ‘station-night’, i.e.three mist-nets open during one night). As residualscan be either positive or negative, they tend to have asymmetrical distribution. This procedure re-establishednormality of capture data when both capture numbersand capture efforts were transformed using log(value + 1).We produced two indicators of bat abundance.Abundance 1 refers to residual values where theregression line of capture numbers against effort isforced to pass through the origin of axes (intercept = 0)as what we would logically expect. In abundance 2, theregression line had no constraints, which resulted inslightly negative intercepts, steeper slopes, and offereddata sets with greater variance. In all cases, the log-logregression of capture numbers against effort wassignificant (
n
= 36 and
P
< 0·001,
R
2
= 0·29–0·67).In a first step, we used generalized linear models
(GLMs) to determine if explicative variables (connec-tivity index, remoteness index, numbers of epiphytesand of
Piper
branches and spikes per census plot) werelinked to each other, and particularly whether foodavailability varied along gradients of forest connectivityand from one study period to the next. Then, we averagedresource availability values of the various census plotswithin each capture site, and introduced in models thebat abundance values as dependent variables. We useda forward selection process and report the amount ofvariability introduced variables explained at each step.Statistical analyses were performed on Systat 9·0.
Results
The six field sessions totalled a capture effort of 255station-nights (or 765 net-nights) and yielded 267understorey fruit bats (Table 1) belonging to our fourtarget species, namely the epiphyte-specialist R. pumilio(59·2% of all captures), and the three shrub-frugivores:C. brevicauda (18·7%), C. perspicillata (10·9%) and S.tildae (11·2%). Only 6·5% of the shrub-frugivores werecaptured in fragments (of a total of 109) against 31·6%of R. pumilio individuals (out of 158). Therefore, thedecrease in abundance in fragments compared withthe mainland was significantly more pronouncedfor the former species than for the latter ones (χ2 =24·366, d.f. = 1, P < 0·001). While shrub-frugivoreswere detected in five of the 14 (35·7%) fragments duringthe recent fragmentation period, their capture ratesremained zero within fragments during the olderfragmentation period. No intersite movements were
recorded in the course of our bat surveys (which werenot designed for that purpose). Conversely, two malesR. pumilio were recaptured at the same capture site atwhich they were initially banded as long as 7–9 yearsbefore.
Radio-tracking surveys conducted on R. pumilio werenot equally successful on the two fragments. The threeindividuals from the large 28-ha fragment could beconveniently tracked all night (contact time > 95%)because they apparently never left the fragment forforaging. A total of 93 flight distances between successivehanging locations were obtained along tracking sessionsof 3–5 nights per individual. On the other hand, the twoindividuals from the small fragment 20 regularlycommuted between adjacent fragments (16, 19 and 22see Fig. 1), leading to a fairly low contact time andhighly disrupted, intractable, tracking data sets due tothe difficulty of navigating by night among unstablesnags. The female roosted once on island 20 and twiceon island 19. Therefore, analyses were restricted toindividuals from the large 28-ha fragment.
Like the six individuals tracked in Nouragues(Henry & Kalko 2007), R. pumilio individuals in Saint-Eugène had rather small home ranges and displayedshort flight distances between successive hanginglocations (90% of flight distances < 200 m). Meanflight distances were slightly longer in Saint-Eugènethan in Nouragues (116 ± 72 m, n = 93, and 102 ± 75 m,n = 231, respectively). However, a nested performedon the square-root transformed flight distances revealedthat this difference could be attributed to a significantinterindividual variability (F7,315 = 2·317, P = 0·026),rather than an effect of the study area (F1,315 = 0·248,P = 0·248). Therefore, we felt comfortable usingtracking data from Nouragues together with thosefrom Saint-Eugène to improve our modelling of flightdistances of R. pumilio.
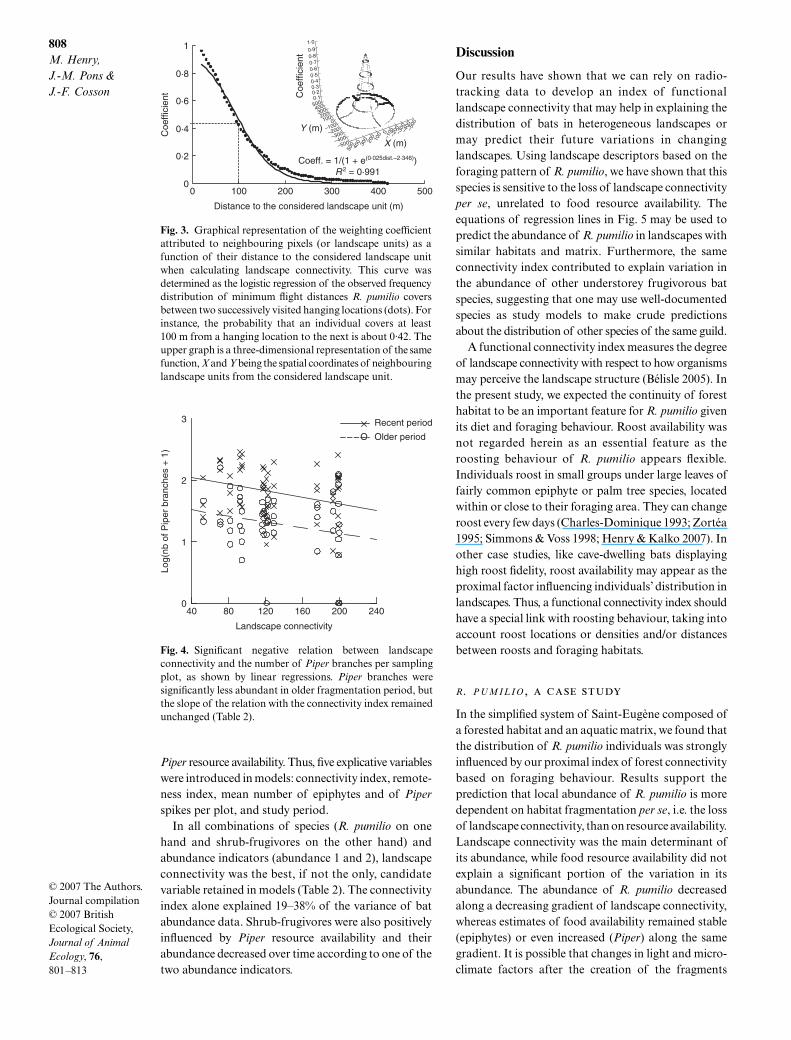
To model frequency distribution of minimal flightdistances, we first pooled all of the 324 flight distancevalues and calculated for each 5-m distance classranging from 15 to 415 m the proportion of flightsobserved (Fig. 3). Then, we applied a logistic regressionon these values as a function of distance, whichdescribes the probability that a given flight will at leastencompass a certain distance. Owing to the absence oflong commuting flights, flight distances displayed ahomogeneous unimodal distribution, and the logisticregression explained a high proportion of variability(R2 > 0·99). The logistic regression was then used todetermine the distance-dependent weighting coefficientsrequired for the calculation of the forest connectivityindex. According to this function, the weighting coef-ficient becomes nearly zero at a distance of 400 m. Inother words, for each landscape unit, the connectivityindex is calculated over a 400 m radius circular area

807Frugivorous bats and landscape connectivity
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
(50·3 ha). The resulting connectivity values for the 18capture sites ranged from 53·2 (the smallest island) to200 (one of the mainland sites) and averaged 119·7 ±45·8 (Table 1). The remoteness index was minimum(< 0·01) for the latter mainland site, peaked at 9·3–9·5for two remote islands, and averaged 2·0 ± 3·2.
A total of 886 Piper individuals was censused on onehundred and fifteen 200-m2 plots (14 sites per period).Epiphyte estimates are available for only 78 of these 115plots (six and 14 sites in recent and older fragmentationperiods, respectively) and totalled 1085 individuals.Piper productivity within plots (number of fruiting orflowering spikes per terminal branch) did not differbetween mainland and islands (Mann–WhitneyU = 1466, d.f. = 1, P = 0·211, n = 109). Assuming thatthe number of terminal branches indicates the potential
fruiting rate (correlation between number of branchesand number of fruiting spikes: Pearson r = 0·81,P < 0·001), we found that Piper resource availabilitywas negatively and significantly influenced by the forestconnectivity (F1,111 = 7·25, P = 0·008), and also under-went a significant 67% decrease from recent to olderfragmentation periods (F1,111 = 4·12, P = 0·045, Fig. 4).Epiphyte density did not vary with landscape connec-tivity or between fragmentation periods (F1,74 = 0·60,P = 0·441, and F1,74 = 1·22, P = 0·272, respectively).
Variation in bat abundance was analysed using GLM,after resource availability values were averaged toproduce a single value per capture site. As the numbersof terminal Piper branches and the numbers of floweringand fruiting spikes were tightly correlated (see above),we only retained the latter variable as an indicator of
Table 1. Detailed bat capture data (Effort: capture effort in numbers of station-nights; Rhp: number of captured R. pumilio, Shf:number of captured shrub-frugivorous bats), descriptors of landscape connectivity (connectivity and remoteness indexes) andmean values of food resource availability (number of Piper branches, Piper spikes and epiphytes per survey plot) within each siteand each fragmentation period (recent 2–4 years post-reservoir construction vs. older 9–11 years)
Period Site Effort Rhp Shf P. branches P. spikes EpiphytesConnectivityindex
Remotenessindex
Recent 5 13 2 0 146·25 14·00 – 71·46 1·97fragmentation 6 12 0 0 160·75 27·50 19·21 94·67 1·17
8 6 3 1 43·00 10·00 16·56 121·43 0·409 4 2 0 – – – 68·65 0·43
12 3 1 0 175·25 52·75 – 92·78 0·9716 7 7 1 81·00 17·50 12·75 129·20 0·3417 6 8 0 73·75 20·50 15·68 122·69 0·5119 7 3 0 98·00 7·50 – 117·13 0·4720 8 0 1 83·50 14·00 – 118·57 0·7721 6 1 0 – – – 87·98 9·5022 4 2 0 57·33 15·00 – 53·24 0·6324 6 1 3 – – – 91·63 9·2834 4 1 0 – – – 135·20 1·7853 6 3 1 68·67 4·67 – 81·98 7·43Cp 11 22 38 97·57 31·57 20·97 199·17 0·03Oi 21 16 9 48·50 8·75 – 200·00 0·00Ro 16 28 1 87·00 19·25 – 176·19 0·20Vi 5 1 0 41·75 3·75 10·40 193·48 0·09
Older 5 8 0 0 54·00 6·75 17·75 71·46 1·97fragmentation 6 6 0 0 14·25 1·75 19·50 94·67 1·17
8 3 3 0 14·00 0·75 10·75 121·43 0·409 6 1 0 – – – 68·65 0·43
12 3 0 0 46·50 2·75 11·25 92·78 0·9716 3 2 0 25·50 6·75 12·25 129·2 0·3417 3 0 0 31·50 3·25 11·50 122·69 0·5119 6 4 0 25·50 4·00 14·50 117·13 0·4720 8 3 0 23·25 1·50 20·75 118·57 0·7721 8 0 0 – – – 87·98 9·5022 4 0 0 32·50 7·00 12·00 53·24 0·6324 3 0 0 – – – 91·63 9·2834 3 2 0 – – – 135·2 1·7853 3 1 0 30·50 1·50 9·75 81·98 7·43Cp 17 31 50 47·13 10·50 17·13 199·17 0·03Oi 13 3 2 7·75 4·00 6·50 200·00 0·00Ro 9 3 0 26·00 4·50 13·25 176·19 0·20Vi 4 4 2 34·50 2·75 21·75 193·48 0·09
808M. Henry, J.-M. Pons & J.-F. Cosson
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
Piper resource availability. Thus, five explicative variableswere introduced in models: connectivity index, remote-ness index, mean number of epiphytes and of Piperspikes per plot, and study period.
In all combinations of species (R. pumilio on onehand and shrub-frugivores on the other hand) andabundance indicators (abundance 1 and 2), landscapeconnectivity was the best, if not the only, candidatevariable retained in models (Table 2). The connectivityindex alone explained 19–38% of the variance of batabundance data. Shrub-frugivores were also positivelyinfluenced by Piper resource availability and theirabundance decreased over time according to one of thetwo abundance indicators.
Discussion
Our results have shown that we can rely on radio-tracking data to develop an index of functionallandscape connectivity that may help in explaining thedistribution of bats in heterogeneous landscapes ormay predict their future variations in changinglandscapes. Using landscape descriptors based on theforaging pattern of R. pumilio, we have shown that thisspecies is sensitive to the loss of landscape connectivityper se, unrelated to food resource availability. Theequations of regression lines in Fig. 5 may be used topredict the abundance of R. pumilio in landscapes withsimilar habitats and matrix. Furthermore, the sameconnectivity index contributed to explain variation inthe abundance of other understorey frugivorous batspecies, suggesting that one may use well-documentedspecies as study models to make crude predictionsabout the distribution of other species of the same guild.
A functional connectivity index measures the degreeof landscape connectivity with respect to how organismsmay perceive the landscape structure (Bélisle 2005). Inthe present study, we expected the continuity of foresthabitat to be an important feature for R. pumilio givenits diet and foraging behaviour. Roost availability wasnot regarded herein as an essential feature as theroosting behaviour of R. pumilio appears flexible.Individuals roost in small groups under large leaves offairly common epiphyte or palm tree species, locatedwithin or close to their foraging area. They can changeroost every few days (Charles-Dominique 1993; Zortéa1995; Simmons & Voss 1998; Henry & Kalko 2007). Inother case studies, like cave-dwelling bats displayinghigh roost fidelity, roost availability may appear as theproximal factor influencing individuals’ distribution inlandscapes. Thus, a functional connectivity index shouldhave a special link with roosting behaviour, taking intoaccount roost locations or densities and/or distancesbetween roosts and foraging habitats.
R. P U M I L I O ,
In the simplified system of Saint-Eugène composed ofa forested habitat and an aquatic matrix, we found thatthe distribution of R. pumilio individuals was stronglyinfluenced by our proximal index of forest connectivitybased on foraging behaviour. Results support theprediction that local abundance of R. pumilio is moredependent on habitat fragmentation per se, i.e. the lossof landscape connectivity, than on resource availability.Landscape connectivity was the main determinant ofits abundance, while food resource availability did notexplain a significant portion of the variation in itsabundance. The abundance of R. pumilio decreasedalong a decreasing gradient of landscape connectivity,whereas estimates of food availability remained stable(epiphytes) or even increased (Piper) along the samegradient. It is possible that changes in light and micro-climate factors after the creation of the fragments
Fig. 3. Graphical representation of the weighting coefficientattributed to neighbouring pixels (or landscape units) as afunction of their distance to the considered landscape unitwhen calculating landscape connectivity. This curve wasdetermined as the logistic regression of the observed frequencydistribution of minimum flight distances R. pumilio coversbetween two successively visited hanging locations (dots). Forinstance, the probability that an individual covers at least100 m from a hanging location to the next is about 0·42. Theupper graph is a three-dimensional representation of the samefunction, X and Y being the spatial coordinates of neighbouringlandscape units from the considered landscape unit.
Fig. 4. Significant negative relation between landscapeconnectivity and the number of Piper branches per samplingplot, as shown by linear regressions. Piper branches weresignificantly less abundant in older fragmentation period, butthe slope of the relation with the connectivity index remainedunchanged (Table 2).

809Frugivorous bats and landscape connectivity
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
locally increased the density of light-demandingpioneer Piper. Newly created edges are characterizedby greater light penetration due to increased treemortality and foliage drop induced by the physiologicalstress of moisture and temperature changes (Lovejoyet al. 1986; Kapos 1989; Malcolm 1994; Ferreira &Laurance 1997; Laurance et al. 1998, 2002) and itappears that Piper flourishes under these conditions.
The sensitivity of R. pumilio to the loss of landscapeconnectivity, despite the maintenance of food resourceavailability, could stem from the incompatibility oftheir foraging strategy with the obligation to crossexpanses of matrix devoid of food sources. The foragingstrategy, i.e. the manner in which bats move acrosslandscapes to search for and exploit food resources, canbe roughly decomposed into two components: searchflights and commuting flights (Fleming et al. 1977;Heithaus & Fleming 1978; Henry 2005; Thies et al.2006). While search flights are devoted to finding fooditems within foraging areas, commuting flights refer tolonger straightforward flights conducted by batsamong several foraging areas. To find their widely scat-tered food items, understorey fruit bats mostly rely on
search flights and less frequently on longer commutingflights (Fleming et al. 1977; Heithaus & Fleming 1978;Bonaccorso & Gush 1987; Henry 2005; Thies et al.2006). As a consequence, they might be reluctant to, orfail to, efficiently exploit patchily distributed foodresources that impose frequent commuting flights overan unexploited matrix.
The foraging strategy of R. pumilio could even beconsidered as an extreme search strategy because thesebats use almost exclusively search flights and thereforeexploit a single small foraging area (Henry & Kalko2007). Most of the surveyed fragments (0·8–7·5 ha) aresmaller in size than their foraging area (3·5–14·1 ha;Henry & Kalko 2007). This might force individuals tosplit their foraging area into smaller ones distributedover two or several contiguous fragments, resulting inregular disruptions of search flights and thus lowertheir foraging efficiency.
Nevertheless, our data suggest that although R.pumilio was strongly affected by the loss of landscapeconnectivity, it could subsist at low densities in poorlyconnected habitats away from the mainland and couldbe a resident in the fragmented area. As a pre-requisite,we got direct evidence from telemetry that individualscan cross narrow habitat disruptions between fragmentswhen these fragments are smaller in size than theirhome range. Most importantly, habitat remoteness,which was directly calculated from the proximalconnectivity index, did not influence the abundanceof R. pumilio. This indicates that the presence ofR. pumilio in rather remote sites does not depend on theproximity of mainland, and suggests that local repro-ductive recruitment may occur in fragmented areas.Accordingly, on many occasions we found juvenilesand reproductive (gestating or lactating) females in smallfragments. Reproductive activity does not seem pre-cluded by the loss of landscape connectivity for R. pumilio.
R. P U M I L I O
R. pumilio was a successful model to build a connectivityindex applicable to other species of the same guild,
Table 2. Sources of variation in the abundance of the epiphyte-specialist R. pumilio and of the shrub-frugivorous C. brevicauda,C. perspicillata and S. tildae within the different capture sites. Results are outputs of a forward multiple regression (GLM) wherethe introduced explicative factors were: landscape structure (landscape connectivity and remoteness), resource availability (meannumber of epiphytes and of piper spikes per plot), and study period (recent vs. older fragmentation period). Factors contributingto explain a significant part of the total variation of bat abundance are given by order of appearance in the model, with their effectsign and their respective contribution to total R2. Abundance 1 and 2 refers to the two indicators of bat abundance (see Methods)
Factors
Abundance 1 Abundance 2
F-ratio P Sign R2 F-ratio P Sign R2
Rhinophylla pumilio1. Connectivity index F1,34 = 18·6 < 0·001 (+) 0·35 F1,34 = 10·8 0·002 (+) 0·24
Shrub-frugivorous bats1. Connectivity index F1,24 = 14·7 0·001 (+) 0·38 F1,34 = 8·07 0·008 (+) 0·192. Piper spikes F1,24 = 9·26 0·008 (+) 0·123. Period F1,24 = 4·57 0·048 (–) 0·11
Fig. 5. Effect of landscape connectivity on abundance index 2 of understorey fruit batsduring the recent and older fragmentation periods. All periods combined, equations oflinear regressions are [y = 0·0039x – 0·4623] and [y = 0·0034x – 0·4046] for R. pumilioand shrub-frugivorous bats, respectively.

810M. Henry, J.-M. Pons & J.-F. Cosson
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
namely Carollia and Sturnira species, for which searchflights are also thought to be an important componentof foraging behaviour. However, the equation linkingconnectivity index to abundance (Fig. 5) should beregarded as a very rough predictor because it is basedon scarce capture data combining several species. Morecapture data would be welcomed to refine the model,but this seems hardly feasible in this context whereshrub-frugivorous bats have apparently desertedfragments during the transition from recent to olderfragmentation periods. This general decline inabundance, also depicted by GLM analyses (Table 2)may be related to the pervasive decrease of Piper as afood resource over the study area (Fig. 4). A progressiveclosure of canopy foliage within edges could haveresulted in the disappearance of many light-demandingPiper plants during the 5 years separating the two plantsurveys. Many stems of dead young Piper were found inthe later period.
We suggest that enlarging the spatial scale of thestudy would provide better insights about the relationbetween landscape connectivity and the abundance ofCarollia and Sturnira species. At least, Carollia perspicillataexploits larger home ranges than R. pumilio and mayregularly perform longer commuting flights amongseveral (two to three) foraging areas located 500–1500 mapart (Heithaus & Fleming 1978). Thus, C. perspicillata islikely to perceive landscape connectivity in a differentway, or at a different spatial scale. Accordingly, Gorresen& Willig (2004) found significant effects of landscapestructure on the abundance of C. perspicillata withinfocus windows of 1–5 km radii, which is 2·5–12 timesthe spatial scale used for R. pumilio. We can imaginethat our study transposed to C. perspicillata would yield amuch extended curve of flight frequency distribution(Fig. 3) with high weighting coefficients attributed tolandscape units situated within a much larger radius.This would produce connectivity values much smallerthan for R. pumilio in the fragmented area, and alsowith much less intersite variability. In other words,C. perspiscillata is likely to perceive and respond tofragmentation at an earlier degree of structuralconnectivity loss compared with R. pumilio. This couldalso explain why Carollia species eventually disappearedfrom fragments at Saint-Eugène while R. pumilio did not.
Collecting data on the foraging pattern of bats poses atechnical challenge due to their high mobility. Detaileddata on flight distance collected herein could even beseen as an exception within bat studies, given the unu-sually short displacements displayed by R. pumilio(mostly < 200 m at a time) compared with other well-studied species. Available tracking data commonlystate home ranges encompassing several tens toseveral hundreds of hectares in a few days, makingflight distances difficult to monitor. Nevertheless, anincreasing number of well-documented studies with
standardized tracking protocols are now available,concerning either shrub-frugivorous bats (Thies et al.2006) or insectivorous bats (Weinbeer & Kalko 2004;Meyer, Weinbeer & Kalko 2005). Although thesestudies may not document flight distance betweensuccessive hanging locations with as much accuracy asin the small home-ranged R. pumilio, they can at leastdocument flight distance observed at fixed time intervals(e.g. every 3 min, or every 10 min).
In studies where the positioning of bats is irregular intime, we propose to rely on density kernels generated byanalyses of home range or foraging area (e.g. Weinbeer& Kalko 2004; Henry & Kalko 2007). Methods ofprobabilistic kernels produce concentric densitycontours delineating areas of a given probability ofpresence. By plotting the inverse probability of presence(1 – P) on the vertical axis, against the radius of thecorresponding area on the horizontal axis, one mayobtain curves similar in shape to the frequency distri-bution of maximal flight distance (Fig. 3).
Finally, other methods than telemetry could providedata on flight distance frequency distribution, butthese could only be operational at greatest temporaland spatial scales. Capture–recapture data, when summedover many individuals in a well-sampled study area,could be treated as flight distances. Genetic analysesperformed on pollen or seeds collected on capturedindividuals can give data on the location of visitedplants, providing plants are already indexed in thevicinity. Finally, homing experiments measuring thetime it takes individuals to return to their roost as afunction of translocation distance, have been used invarious taxa as a means to assess landscape connectivity(see the review by Bélisle 2005, p. 1993). Homing experi-ments could be conducted on cave-dwelling bats.
Our study contrasts with the other bat surveys conductedin forest fragments surrounded by abundant second-growth vegetation characterized by very high densitiesof the bat plants Piper, Solanum or Vismia (Estradaet al. 1993; Brosset et al. 1996; Kalko 1998; Schulzeet al. 2000; Estrada & Coates-Estrada 2001, 2002;Gorresen & Willig 2004). In these studies, Carollia andSturnira species remained abundant in forest fragments,second growth and pastures where they are effectiveseed dispersers (Galindo-González, Guevara & Sosa2000). Therefore, bat surveys in Saint-Eugène indi-rectly underline the important role of second growthvegetation in shaping fruit bat communities in frag-mented forests. A matrix colonized by second growthvegetation may provide bats with compensatory foodsupplies that favour the spatial continuity of resourcedistribution (matrix compensation and supplementa-tion hypotheses; Dunning, Danielson & Pulliam 1992;Norton et al. 2000). In further studies, indicators oflandscape connectivity should be adapted to morecomplex environments including various habitat types.

811Frugivorous bats and landscape connectivity
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
In the modelling calculation, the different habitat typesshould be assigned to different suitability values rang-ing between 0 and 200, depending for instance on theirrelative contribution to the diet of bats. Such modellingconnectivity in more realistic landscape mosaics isurgently needed. Among others, it would help under-standing how bat diversity patterns may be influencedby the spatial configuration of the various neighbour-ing habitat types, and how communities are likely to bemodified in changing landscapes.
Indicators of landscape connectivity may alsocontribute to an understanding of ecological processesat different levels of analysis. One may, for instance,expect intraspecific variation in the perception oflandscape connectivity. Breeding females have to facecritical energetic, temporal and behavioural constraints,and are usually forced to reduce the spatial extent oftheir foraging movements (e.g. Racey & Swift 1985;Kurta et al. 1989; Charles-Dominique 1991; Henryet al. 2002). In particular, lactating R. pumilio reducetheir foraging area and flight distances by 42% and25%, respectively (Henry & Kalko 2007), and are thenlikely to perceive landscape connectivity in a differentway. This raises questions about spatial variation ofpopulation dynamics in fragmented areas. At theinterspecific level, landscape connectivity may helpinvestigate or predict possible effects of competitionrelaxation in areas where perceived landscape connec-tivity varies greatly among competitors. Conversely,studies of the mutualistic interactions between plantsand seed-dispersing bats could benefit from a betterknowledge of what landscape connectivity meansfrom dispersers’ eyes.
Acknowledgements
We are grateful to G. Dubost and C. Erard who initi-ated this project. We thank R. Barbault, P. Charles-Dominique, T.H. Fleming, P.M. Forget, E.K.V. Kalko,L. Granjon, J.L. Martin, N.G. Yoccoz and H.J. Youngfor providing valuable criticism and suggestions. Manythanks also go to all the people who participated orhelped in any aspect of the fieldwork, especially P.Cerdan (Laboratoire Hydreco, barrage de Petit-Saut),S. Poirot, A.S. Hennion, A. Lyet, R. Kirsch and D.Pons. The fieldwork was supported by the ‘Electricitéde France’ (Convention Muséum/EDF CQZH 1294)and the Laboratoire d’Ecologie Générale de Brunoy(MNHN-CNRS, UMR 5176, France). M.H. receiveda Ph.D. grant from La Fondation des Treilles.
References
Andrén, H. (1994) Effects of habitat fragmentation on birdsand mammals in landscapes with different proportions ofsuitable habitat: a review. Oikos, 71, 355–366.
Bélisle, M. (2005) Measuring landscape connectivity: thechallenge of behavioral landscape ecology. Ecology, 86,1988–1995.
Bonaccorso, F.J. & Gush, T.J. (1987) Feeding behaviour and
foraging strategies of captive phyllostomid fruit bats:an experimental study. Journal of Animal Ecology, 59,907–920.
Brosset, A., Charles-Dominique, P., Cockle, A., Cosson, J.F.& Masson, D. (1996) Bat communities and deforestationin French Guiana. Canadian Journal of Zoology, 74,1974–1982.
Brotons, L., Mönkkönen, M. & Martin, J.L. (2003) Arefragments islands? Landscape context and density-arearelationships in boreal forest birds. American Naturalist,162, 343–357.
Brotons, L., Herrando, S. & Martin, J.L. (2004) Birdassemblages in forest fragments within Mediterraneanmosaics created by wild fires. Landscape Ecology, 19, 663–675.
Brotons, L., Wolff, A., Paulus, G. & Martin, J.L. (2005) Effectof adjacent agricultural habitat on the distribution ofpasserines in natural grasslands. Biological Conservation,124, 407–414.
Charles-Dominique, P. (1991) Feeding strategy and activitybudget of the frugivorous bat Carollia perspicillata(Chiroptera: Phyllostomidae) in French Guiana. Journal ofTropical Ecology, 7, 243–256.
Charles-Dominique, P. (1993) Tent-use by the bat Rhinophyllapumilio (Phyllostomidae: Carolliinae) in French Guiana.Biotropica, 25, 111–116.
Charles-Dominique, P. (2001) The field station. Nouragues:Dynamics and Plant–Animal Interactions in a NeotropicalRainforest (eds F. Bongers, P. Charles-Dominique, P.M.Forget & M. Théry), pp. 1–7. Kluwer Academic Publishers,Dordrecht.
Charles-Dominique, P. & Cockle, A. (2001) Frugivory anddispersal by bats. Nouragues: Dynamics and Plant–AnimalInteractions in a Neotropical Rainforest (eds F. Bongers, P.Charles-Dominique, P.M. Forget & M. Théry), pp. 207–216. Kluwer Academic Publishers, Dordrecht.
Charles-Dominique, P., Brosset, A. & Jouard, S. (2001) AtlasDes Chauves-Souris de Guyane. Patrimoines naturels, no.49. Muséum National d’Histoire Naturelle, Paris.
Claessens, O., Granjon, L. & de Massary, J.C.& Ringuet, S.(2002) La station de recherche de Saint-Eugène: situation,environnement et présentation générale. Revue d’Ecologie(Terre et Vie), 57, 21–39.
Cockle, A. (1997) Modalités de dissémination et d’établissementde lianes de sous-bois (Cyclanthaceae et Philodendron) EnForêt Guyanaise. PhD Thesis, Université Paris VI, France.
Cosson, J.F. (1994) Dynamique de population et dispersion dela chauve-souris frugivore Carollia perspicillata en GuyaneFrançaise. PhD Thesis, Université Paris XI, France.
Cosson, J.F., Pons, J.M. & Masson, D. (1999a) Effects offorest fragmentation on frugivorous and nectarivorousbats in French Guiana. Journal of Tropical Ecology, 15,515–534.
Cosson, J.F., Ringuet, S., De Massary, J.C., Claessens, O.,Dalecky, A., Villiers, J.F., Granjon, L. & Pons, J.M. (1999b)Ecological changes in recent land-bridge islands in FrenchGuiana, with emphasis on vertebrate communities.Biological Conservation, 91, 213–222.
Coulon, A., Cosson, J.F., Angibault, J.M., Cargnelutti, B.,Galan, M., Morellet, N., Petit, E., Aulagnier, S. & Hewison,A.J.M. (2004) Landscape connectivity influences gene flowin a roe deer population inhabiting a fragmented landscape:an individual-based approach. Molecular Ecology, 13,2841–2850.
Delaval, M., Henry, M. & Charles-Dominique, P. (2005)Interspecific competition and niche partitioning: exampleof a Neotropical rainforest bat community. Revue d’Ecologie(Terre et Vie), 60, 149–166.
Dunning, J.B., Danielson, B.J. & Pulliam, H.R. (1992)Ecological processes that affect populations in complexlandscapes. Oikos, 65, 169–175.

812M. Henry, J.-M. Pons & J.-F. Cosson
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
Estrada, A. & Coates-Estrada, R. (2001) Bat species richnessin live fences and in corridors of residual rain forestvegetation at Los Tuxtlas, Mexico. Ecography, 24, 94–102.
Estrada, A. & Coates-Estrada, R. (2002) Bats in continuousforest, forest fragments and in an agricultural mosaichabitat-island at Los Tuxtlas, Mexico. Biological Conservation,103, 237–245.
Estrada, A., Coates-Estrada, R. & Merrit, D. Jr (1993) Batspecies richness and abundance in tropical rain forestfragments and in agricultural habitats at Los Tuxtlas,Mexico. Ecography, 16, 309–318.
Ewers, R.M. & Didham, R.K. (2006) Confounding factors inthe detection of species responses to habitat fragmentation.Biological Reviews, 81, 117–142.
Fahrig, L. & Merriam, G. (1994) Conservation of fragmentedpopulations. Conservation Biology, 8, 50–59.
Faria, D. (2006) Phyllostomid bats of a fragmented landscapein the north-eastern Atlantic forest, Brazil. Journal of TropicalEcology, 22, 531–542.
Ferreira, L.V. & Laurance, W.F. (1997) Effects of forestfragmentation on mortality and damage of selected trees incentral Amazonia. Conservation Biology, 11, 797–801.
Fleming, T.H. (1982) Foraging strategies in plant-visitingbats. Ecology of Bats (ed. T.H. Kunz), pp. 287–386. PlenumPress, New York.
Fleming, T.H. (1985) Coexistence of five sympatric Piper(Piperaceae) species in a tropical dry forest. Ecology, 66,688–700.
Fleming, T.H., Heithaus, E.R. & Sawyer, W.B. (1977) Anexperimental analysis of the food location behavior offrugivorous bats. Ecology, 58, 619–627.
Galindo-González, J., Guevara, S. & Sosa, V.J. (2000) Bat-and bird-generated seed rains at isolated trees in pastures ina tropical rainforest. Conservation Biology, 14, 1693–1703.
Gascon, C., Lovejoy, T.E., Bierregaard, R.O. Jr, Malcolm,J.R., Stouffer, P.C., Vasconcelos, H., Laurance, W.F.,Zimmerman, B., Tocher, M. & Borges, S. (1999) Matrixhabitat and species persistence in tropical forest remnants.Biological Conservation, 91, 223–229.
Gorresen, P.M. & Willig, M.R. (2004) Landscape responses ofbats to habitat fragmentation in Atlantic forest of Paraguay.Journal of Mammalogy, 85, 688–697.
Granjon, L., Cosson, J.F., Judas, J. & Ringuet, S. (1996)Influence of tropical rainforest fragmentation on mammalcommunities in French Guiana: early trends. Acta Oecologia,17, 673–684.
Heithaus, E.R. & Fleming, T.H. (1978) Foraging movementsof a frugivorous bat, Carollia perspicillata (Phyllostomatidae).Ecological Monographs, 48, 127–143.
Henry, M. (2005) Le déclin des populations de chauves-sourisfrugivores en forêt néotropicale fragmentée – conséquencessur la dispersion des graines. PhD Thesis, Université ParisVI, France.
Henry, M. & Jouard, S. (2007) Effect of bat exclusion on pat-terns of seed rain in tropical rainforest in French Guiana.Biotropica, doi: 10.1111/j.1744-7429.2007.00286.x, in press.
Henry, M. & Kalko, E.K.V. (2007) Foraging strategy andbreeding constraints of Rhinophylla pumilio (Phyllostomidae)in the Amazon lowlands. Journal of Mammalogy, 88, 81–93.
Henry, M., Thomas, D.W., Vaudry, R. & Carrier, M. (2002)Foraging distances and home range of pregnant and lactatinglittle brown bats (Myotis lucifugus). Journal of Mammalogy,83, 767–774.
Hewison, A.J.M., Vincent, J.P., Joachim, J., Angibault, J.M.,Cargnelutti, B. & Cibien, C. (2001) The effects of woodlandfragmentation and human activity on roe deer distributionin agricultural landscapes. Canadian Journal of Zoology,79, 679–689.
Kalko, E.K.V. (1998) Organisation and diversity of tropicalbat communities through space and time. Zoology, 101,281–297.
Kapos, V. (1989) Effects of isolation on the water status offorest patches in the Brazilian Amazon. Journal of TropicalEcology, 5, 173–185.
Kupfer, J.A., Malanson, G.P. & Franklin, S.B. (2006) Notseeing the ocean for the islands: the mediating influence ofmatrix-based processes on forest fragmentation effects.Global Ecology and Biogeography, 15, 8–20.
Kurta, A., Bell, G.P., Nagy, K.A. & Kunz, T.H. (1989)Energetics of pregnancy and lactation in free-ranging littlebrown bat (Myotis lucifugus). Physiological Zoology, 62,804–818.
Lang, A.B., Kalko, E.K.V., Romer, H., Bockholdt, C. &Dechmann, D.K.N. (2006) Activity levels of bats andkatydids in relation to the lunar cycle. Oecologia, 146, 659–666.
Laurance, W.F. & Bierregaard, R.O. Jr (1997) Tropical ForestRemnants: Ecology, Management, and Conservation ofFragmented Communities. University of Chicago Press.
Laurance, W.F., Ferreira, L.V., Rankin-de Merona, J.M. &Laurance, S.G. (1998) Rain forest fragmentation and thedynamics of Amazonian tree communities. Ecology, 79,2032–2040.
Laurance, W.F., Lovejoy, T.E., Vasconcelos, H.L., Bruna,E.M., Didham, R.K., Stouffer, P.C., Gascon, C., Bierre-gaard, R.O. Jr, Laurance, S.G. & Sampiao, E. (2002) Eco-system decay of Amazonian forest fragments: a 22-yearinvestigation. Conservation Biology, 16, 605–618.
Leigh, E.G. Jr, Cosson, J.F., Pons, J.M. & Forget, P.M. (2002)En quoi l’étude des îlots forestiers permet-elle de mieuxconnaître le fonctionnement de la forêt tropicale? Revued’Ecologie (Terre et Vie), 57, 181–194.
Lobova, T.A. & Mori, S.A. (2004) Epizoochorous dispersalby bats in French Guiana. Journal of Tropical Ecology, 20,581–582.
Lovejoy, T.E., Bierregaard, R.O. Jr, Rylands, A.B., Malcolm,J.R., Quintela, C.E., Harper, L.H., Brown, K.S. Jr, Powel,A.H., Powell, G.V.N., Schubart, H.O. & Hays, M.B. (1986)Edge and other effects of isolation on Amazon forestfragments. Conservation Biology: the Science of Scarcityand Diversity (ed. M.E. Soulé), pp. 257–285. Sinauer,Sunderland, MA.
MacArthur, R.H. & Wilson, E.O. (1967) The Theory of IslandBiogeography. Princeton University Press, Princeton,NJ.
Malcolm, J.R. (1994) Edge effects in central Amazonianforest fragments. Ecology, 75, 2438–2445.
Manel, S., Schwartz, M.K., Luikart, G. & Taberlet, P.(2003) Landscape genetics: combining landscape ecology andpopulation genetics. Trends in Ecology and Evolution, 18,189–197.
Marinho-Filho, J.S. (1991) The coexistence of two frugivorousbat species and the phenology of their food plants in Brazil.Journal of Tropical Ecology, 7, 59–67.
Meyer, C.F.J., Weinbeer, M. & Kalko, E.K.V. (2005) Homerange size and spacing patterns of Macrophyllum macro-phyllum (Phyllostomidae) foraging over water. Journal ofMammalogy, 86, 587–598.
Norton, M.R., Hannon, S.J. & Schmiegelow, F.K.A. (2000)Fragments are not islands: patch vs landscape perspectiveson songbird presence and abundance in a harvested borealforest. Ecography, 23, 209–223.
Pons, J.M. & Cosson, J.F. (2002) Effect of forest fragmentationon animalivorous bats in French Guiana. Revue d’Ecologie(Terre et Vie), 57, 117–130.
Racey, P.A. & Swift, S.M. (1985) Feeding ecology of Pipistrelluspipistrellus (Chiroptera: Vespertilionidae) during pregnancyand lactation. I. Foraging behaviour. Journal of AnimalEcology, 54, 205–215.
Ricketts, T.H. (2001) The matrix matters: effective isolationin fragmented landscapes. American Naturalist, 158, 87–99.

813Frugivorous bats and landscape connectivity
© 2007 The Authors.Journal compilation© 2007 British Ecological Society, Journal of Animal Ecology, 76, 801–813
Saunders, D.A., Hobbs, R.J. & Margules, C.R. (1991) Biologicalconsequences of ecosystem fragmentation: a review.Conservation Biology, 5, 18–32.
Schulze, M.D., Seavy, N.E. & Whitacre, F.F. (2000) Acomparison of the Phyllostomid bat assemblages inundisturbed neotopical forest and in forest fragments of aslash-and-burn farming mosaic in Petén, Guatemala.Biotropica, 32, 174–184.
Simberloff, D. (1988) The contribution of population andcommunity biology to conservation science. Annual Reviewof Ecology and Systematic, 19, 473–471.
Simmons, N.B. & Voss, R. (1998) The mammals of Paracou,French Guyana: a neotropical lowland rainforest fauna.Part I. Bats. Bulletin of the American Museum of NaturalHistory, 237, 1–219.
Stouffer, P.C. & Bierregaard, R.O. Jr (1995a) Use of Amazo-nian forest fragments by understorey insectivorous birds.Ecology, 76, 2429–2445.
Stouffer, P.C. & Bierregaard, R.O. Jr (1995b) Effects of forestfragmentation on understorey hummingbirds in AmazonianBrazil. Conservation Biology, 9, 1085–1094.
Taylor, R.J. (1987a) The geometry of colonization: 1. Islands.Oikos, 48, 225–231.
Taylor, R.J. (1987b) The geometry of colonization: 2. Peninsulas.Oikos, 48, 232–237.
Taylor, P.D., Fahrig, L., Henein, K. & Merriam, G. (1993)Connectivity is a vital element of landscape structure.Oïkos, 68, 571–573.
Thies, W. & Kalko, E.K.V. (2004) Phenology of neotropicalpepper plants (Piperaceae) and their association with theirmain dispersers, two short-tailed fruit bats, Carollia perspicillataand C. castanea (Phyllostomidae). Oikos, 104, 362–376.
Thies, W., Kalko, E.K.V. & Schnitzler, H.-U. (2006) Influenceof environment and resource availability on activitypatterns of Carollia castanea (Phyllostomidae) in Panama.Journal of Mammalogy, 87, 331–338.
Tischendorf, L. & Fahrig, L. (2000) On the usage and meas-urement of landscape connectivity. Oikos, 90, 7–19.
Weinbeer, M. & Kalko, E.K.V. (2004) Morphologicalcharacteristics predict alternate foraging strategy andmicrohabitat selection in the orange-bellied bat, Lampronycterisbrachiotis. Journal of Mammalogy, 85, 1116–1123.
With, K.A., Gardner, R.H. & Turner, M.G. (1997) Landscapeconnectivity and population distributions in heterogeneousenvironments. Oikos, 78, 151–169.
Zortéa, M. (1995) Observations on tent-using in the carollinebat Rhinophylla pumilio in southeastern Brazil. ChiropteraNeotropical, 1, 85–88.
Received 28 December 2006; accepted 11 April 2007
Related Documents











