
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript


Culture Media for Food Microbiology

Vol. 14 (1978) edited by M.J. Bull (lst reprint 1983) Vol. 15 (1979) edited by M.J. Bull Vol. 16 (1982) edited by M.J. Bull Vol. 17 (1983) edited by M.E. Bushell Vol. 18 (1983) Microbial Polysaccharides, edited by M.E. Bushell Vol. 19 (1984) Modern Applications of Traditional Biotechnologies, edited by
M.E Bushell Vol. 20 (1984) Innovations in Biotechnologie, edited by E.H. Houwink and
R.R. van der Meer Vol. 21 (1989) Statistical Aspects of the Microbiological Analysis of Foods, by B. Jarvis Vol, 22 (1986) Moulds and Filamentous Fungi in Technical Microbiology, by
O. Fassatiov~ Vol. 23 (1986) Micro-organisms in the Production of Food, edited by M.R. Adams Vol. 24 (1986) Biotechnology of Animo Acid Production; edited by K. Aida, I. Chibata,
K. Nakayama, K. Takinama and H. Yamada Vol. 25 (1988) Computers in Fermentation Technology, edited by M.E. Bushell Vol. 26 (1989) Rapid Methods in Food Microbiology, edited by M.R. Adams and
C.F.A. Hope Vol. 27 (1989) Bioactive Metabolites from Microorganisms, edited by M.E. Bushell and
U. Gr~fe Vol. 28 (1993) Micromycetes in Foodstuffs and Feedstuffs; edited by Z. Jesenskfi Vol. 29 (1994) Aspergillus: 50 years on; edited by S.D. Martinelli and J.R. Kinghorn Vol. 30 (1994) Bioactive Secondary Metabolites of Microorganisms, edited by
V. Betina Vol. 31 (1995) Techniques in Applied Microbiology, edited by B. Sikyta Vol. 32 (1995) Biotransformations: Microbial Degradation of Health Risk Compounds,
edited by V.P. Singh Vol. 33 (1995) Microbial Pentose Utilization. Current Applications in Biotechnology,
by A. Singh and P. Mishra

Culture Media for Food Microbiology
Edited by
Janet E.L. Corry Department of Clinical Veterinary Science, University of Bristol, Langford, Avon, BS18 7DU, U.K.
G.D.W. Curtis Bacteriology Department, John Radcliffe Hospital, Oxford OX3 9D U, U.K.
Rosamund M. Baird Summer/ands House, Summer/ands, Yeovil, Somerset BA21 3AL, U.K.
progress in industrial microbiology
E L S E V I E R
A m s t e r d a m - L a u s a n n e - N e w Y o r k - O x f o r d - S h a n n o n - S i n g a p o r e - T o k y o

ELSEVIER SCIENCE B.V. Sara Burgerhartstraat 25 P.O. Box 211, 1000 AE Amsterdam, The Netherlands
First printing 1995 Second impression 1999
ISBN 0-444-81498-1 (Vol. 34) ISBN 0-444-41668-8 (Series)
�9 1995 Elsevier Science B.V. All rights reserved
No part of this publication may be reproduced, stored in a retrieval system or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior written permission of the publisher, Elsevier Science B.V., Copyright & Permissions Department, P.O. Box 521, 1000 AM Amsterdam, The Netherlands.
Special regulations for readers in the U.S.A. - This publication has been registered with the Copyright Clearance Center Inc. (CCC), 222 Rosewood Drive, Danvers, MA 01923. Information can be obtained from the CCC about conditions under which photocopies of parts of this publica- tion may be made in the U.S.A. All other copyright questions, including photocopying outside of the USA, should be referred to the copyright owner, Elsevier Science B.V., unless otherwise specified.
No responsibility is assumed by the publisher for any injury and/or damage to persons or property as a matter of products liability, negligence or otherwise, or from any use or operation of any methods, products, instructions or ideas contained in the material herein.
This book is printed on acid-free paper.
Printed in The Netherlands

Acknowledgements
This book is the result of the work of many microbiologists who have taken part in the six meetings of the Working Party on Culture Media since its inception in 1978. The International Committee on Food Microbiology and Hygiene of the International Union of Microbiological Societies thanks all those who participated in the meetings and in particular those who prepared monographs for discussion: R.M. Baird, V. Bartl, L.R. Beuchat, R.M. Blood, E. de Boer, F.J. Bolton, R.E. Brackett, G.D.W. Curtis, A.R. Datta, L. Dominguez Rodriguez, E. Elliot, J.M. Farber, G.A. Gardner, W.H. Holzapfel, R. Holbrook, R.V. Lachica, J.V. Lee, W.H. Lee, S.J. Lewis, B.M. Mackey, G.C. Mead, S.C. Morgan Jones, D.A.A. Mossel, P. van Netten, I. Perales, T. Petersen, D.J. Pusch, B. Ralovich, G. Reuter, M. van Schothorst, D.A.L. Seiler, N.P. Skovgaard, B. Swaminathan, G. Wauters and G. Weenk. H. SCgaard and M. Jakobsen, as successive Editors-in-Chief of the International Journal of Food Microbiology, have been unfailingly helpful and patient in the publication of proceedings in that journal.
This work has been supported by the following companies whose generosity is gratefully acknowledged: BDH, Becton Dickinson, Difco, Elsevier, Gibco, Lab M, Merck and Oxoid.

This Page Intentionally Left Blank

Introduction
The roots of this book are an idea of the past president of the International Committee for Food Microbiology and Hygiene (ICFMH), Professor David Mos- sel, who formed the Working Party on Quality Assurance of Culture Media in 1978. This group convened its first meeting in Mallorca, Spain in 1979. In those days little attention was paid by many food microbiologists to the possibility that the media they used might not always function optimally. This applied especially to selective media used to isolate pathogens. The Mallorca meeting, funded by the Merck Society for Arts and Science, brought together 35 microbiologists from 13 countries. Topics covered included quality assurance tests used for raw materials for media, microbiological methods of monitoring complete media and particular problems encountered with media developed for specific groups of micro- organisms. Areas identified as requiring particular investigation were: standard methods and choice of reference strains for use in media monitoring; guidance concerning the effect of substrate (e.g. type of food examined); the effects of sublethal damage and competitive flora; information concerning the shelf-life of dehydrated media, rehydrated media and poured plates.
Inhibitors such as bile salts and brilliant green were identified as unsatisfactory because they were poorly defined and methods were needed for monitoring them. The proceedings were published in a book (Corry, 1982).
The sequel to the Mallorca meeting was held in Dallas, USA, in 1981, funded by Oxoid Ltd. Most contributions were concerned with selective media developed for specific groups of food-related pathogens but it was at this time that the possibility of producing a 'pharmacopoeia' of culture media was first discussed in depth following a paper presented by Dr Vladimir Bartl from Prague. The proceedings were published in a special issue of the Archiv f/Jr Lebensmittel- hygiene (Corry and Baird, 1982).
The third meeting of the Working Party, held in London in January 1984 and funded by Difco Inc., was the first at which all three editors of this pharmacopoeia were present. The proceedings appeared in a special issue of the new ICFMH journal, the International Journal of Food Microbiology (Baird et al., 1985) which set the scene for (i) the format of the information to be included in the mono- graphs for each medium (supplied, appropriately, by Rosamund Baird, whose first degree is in pharmacy); (ii) the protocol for microbiological testing of culture

viii Introduction
media, devised by a committee of the Working Party chaired by Gordon Curtis and (iii) the media for which monographs were to be written.
There followed two years of hard work editing the pharmacopoeia and organis- ing the fourth meeting of the Working Party. This was held in association with the IUMS 14th International Congress of Microbiology in Manchester, England in 1986 and funded by a consortium of manufacturers and Elsevier Science B.V. It was a highly productive meeting lasting two very full days and considered draft monographs for 42 different media. As a result of this meeting a complete issue of the International Journal of Food Microbiology was published containing all the monographs as well as standard methods of testing the selectivity and productivity of solid and liquid media and a list of standard strains to be used for testing media performance (Baird et al., 1987).
A subsequent meeting in Budapest in 1988 added more monographs, many of which were for Listeria monocytogenes media (Baird et al., 1989). This bacterium was by then attracting Widespread attention as a 'new' food-borne pathogen.
There followed a period of consolidation while food microbiologists from all parts of the world were requested to monitor their media using the test strains and methods prescribed. In 1992 a four day meeting was held in Heidelberg, funded by Becton Dickinson, at which all the monographs were reviewed, some were added and some deleted. The results of medium monitoring using standard strains as well as 'in house' strains of test organisms were presented and analysed by approximately 30 participants, and as a result, the numbers of test organisms recommended for monitoring each medium were reduced and the total number of strains recommended for use were rationalised. The new monographs as well as a series of reviews of media for different groups of food micro-organisms were published in the International Journal of Food Microbiology. All current mono- graphs, together with these reviews are collected in this new volume.
We hope that microbiologists specialising in food and related areas, particularly those who are members of or who aspire to join a laboratory accreditation scheme, will find this book useful.
We have tried to include all the media most commonly used in food microbiol- ogy. Inclusion of a medium, however, implies no endorsement of its superiority over other media, and likewise, there will be good media that are absent from our book. Topics that still need to be addressed include the standardisation of undefined ingredients such as blood, plasma, bile and brilliant green, procedures for resuscitation of sublethally-damaged organisms and the effect of the type of food on the optimal method of examination.
We rely upon readers to use the methods and test organisms suggested and to inform the editors of any errors or ambiguities found. Please send us any com- ments or suggestions you may have concerning improvements, deletions or addi- tions that can be made in future editions.
Janet E.L. Corry, G.D.W. Curtis and Rosamund M. Baird

Introduction ix
References
Baird, R.M., Barnes, E.M., Corry, J.E.L., Curtis, G.D.W. and Mackey, B.M. (editors, 1985) Proceedings of the third international symposium on quality assurance and quality control of culture media. Int. J. Food Microbiol. 2, 1-138.
Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (editors, 1987) Pharmacopoeia of culture media for food microbiology. Int. J. Food Microbiol. 5, 187-300.
Baird, R.M, Corry, J.E.L., Curtis, G.D.W., Mossel, D.A.A. and Skovgaard, N.P. (editors, 1988) Pharmacopoeia of culture media for food microbiology- additional monographs. Int. J. Food Microbiol. 9, 85-144.
Corry, J.E.L. (editor, 1982) Quality assurance and quality control of culture media. G.I.T. Verlag, Darmstadt.
Corry, J.E.L. and Baird, R.M. (editors, 1982) Proceedings of the second international symposium on quality assurance of microbiological culture media. Arch. Lebensmittelhyg. 33, 137-175..

This Page Intentionally Left Blank

Contents
Acknowledgements/v
Introduction/vii
Part 1 Reviews of media
Chapter 1 Microbiological assessment of culture media - comparison and statistical evaluation of methods (G Weenk)/1
Chapter 2 Principles involved in the detection and enumeration of clostridia in foods (G C Mead)/25
Chapter 3 Media for the detection and enumeration of Bacillus cereus in foods (P van Netten and J M Kramer)/35
Chapter 4 Culture media for enterococci and group D streptococci (G Reuter)/51
Chapter 5 Culture media and methods for the isolation of Listeria monocytogenes (G D W Curtis and W H Lee)/63
Chapter 6 Media used in the detection and enumeration of Staphylococcus aureus (Rosamund M Baird and W H Lee)/77
Chapter 7 Culture media for non-sporulating Gram-positive food spoilage bacteria (W H Holzapfel)/89
Chapter 8 Media for Aeromonas spp., Plesiomonas shigelloides and Pseudomonas spp. from food and environment (C Jeppesen)/111
Chapter 9 Culture media for the isolation of campylobacters (Janet E L Corry, D E Post, P Colin and M J Laisney)/129
Chapter 10 Media for coliforms, Escherichia coli and 'total' Enterobacteriaceae (Ruth M Blood and G D W Curtis)/163
Chapter 11 Media for salmonella (M Busse)/187 Chapter 12 Culture media for the isolation and enumeration of pathogenic Vibrio
species in foods and environmental samples (T J Donovan and P van Netten)/203
Chapter 13 Isolation of Yersinia enterocolitica from foods (E de Boer)/219 Chapter 14 Media for detecting and enumerating yeasts and moulds (L R
Beuchat)/229
Part 2 Pharmacopoeia of culture media
Notes on the use of the monographs/243 Summary of organisms and recommended media/247

xii Contents
Monographs
All Purpose Tween (APT) agar- modified (for H202 detection)/251 Aspergillus flavus and parasiticus agar (AFPA)/254 Baird-Parker agar/257 Baird-Parker liquid (LBP) medium/261 Bile Oxalate Sorbose (BOS) broth/263 Bismuth sulphite agar/266 Briggs agar/269 Brilliant Green Bile (BGB) broth/272 Cefsulodin Irgasan Novobiocin (CIN) agar/275 Cellobiose Polymyxin B Colistin (CPC) agar/278 Cephaloridin Fucidin Cetrimide (CFC) agar/281 Charcoal Cefoperazone Deoxycholate (CCD) agar - modified/284 Charcoal Cefoperazone Deoxycholate (CCD) broth/287 Citrate Azide Tween Carbonate (CATC) agar/290 Cresol red Thallium Acetate Sucrose (CTAS) agar/293 Diagnostic Salmonella Selective Semisolid Medium (DIASALM)/296 Dichloran Glycerol (DG18) agar/300 Dichloran Rose Bengal Chloramphenicol (DRBC) agar/303 Differential Reinforced Clostridial agar- Modified (mDRCM)/306 Dominguez Rodriguez LSAMm agar/309 Enterobacteriaceae Enrichment (EE) broth/314 FDA Listeria enrichment broth/317 Fraser broth - modified/320 Giolitti and Cantoni Broth (GCB)/323 Hektoen Enteric (HE) agar/326 Irgasan Ticarcillin Chlorate (ITC) broth/329 Iron sulphite agar/332 Kanamycin Aesculin Azide (KAA) agar/335 KRANEP agar/337 Lactobacillus Sorbic acid (LaS) agar (syn. sorbic acid agar base)/340 Lauryl tryptose broth/343 Levinthal broth with acriflavine and nalidixic acid/345 Lithium chloride Ceftazidime Tween 80 (LCT) agar/347 Lithium chloride Phenylethanol Moxalactam (LPM) agar/350 L-S Differential (LSD) agar/354 Lysine Iron Cystine Neutral Red (LICNR) broth/357 M 17 agar/360 de Man, Rogosa and Sharpe (MRS) agar/362 de Man, Rogosa and Sharpe agar with Sorbic acid (MRS-S) agar/364 Mannitol Egg Yolk Polymyxin (MEYP) agar/367 Mannitol Lysine Crystal violet Brilliant green (MLCB) agar/369 M-Enterococcus (ME) agar/372 Muller Kauffman tetrathionate broth/374

Contents xiii
Oleandomycin Polymyxin Sulphadiazine Perfringens Agar (OPSPA)/378 Oxford agar/381 Oxford agar- Modified (MC)/385 Oxytetracycline Glucose Yeast extract (OGY) agar//388 Phenol red brilliant green agar (syn. modified brilliant green agar)/391 Polymyxin Acriflavine Lithium Chloride Ceftazidime Aesculin Mannitol (PALCAM) agar/395 Polymyxin Acriflavine Lithium Chloride Ceftazidime Aesculin Mannitol Egg Yolk (L-PALCAMY) broth/399 Polymyxin Pyruvate Egg Yolk Mannitol Bromothymol blue Agar (PEMBA)/402 Preston campylobacter selective agar/405 Preston enrichment broth/408 Rambach agar/410 Rapid Perfringens Medium (RPM)/413 Rappaport-Vassiliadis (RV) broth/416 Rappaport (SR) medium - semisolid modification/419 Rappaport-Vassiliadis (MSRV) Medium - Semisolid modification/422 Rogosa Agar/425 Rogosa agar- modified (pH 6.2)/428 Rose Bengal Chloramphenicol (RBC) agar/431 Salmonella Shigella Deoxycholate Calcium (SSDC) agar//434 Selenite cystine broth/437 Skirrow Campylobacter selective agar/440 Streptomycin Thallous Acetate Actidione (STAA) agar/442 Sulphite Cycloserine Azide (SCA) agar/445 Thallous Acetate Tetrazolium Glucose (T1TG) agar/448 Thiosulphate Citrate Bile-salt Sucrose (TCBS) agar/450 Tryptone Bile Agar (TBA)/453 Tryptone Soya Broth with 10% NaC1 & 1% Sodium Pyruvate (PTSBS)/456 Tryptose Sulphite Cycloserine (TSC) agar (without egg yolk)/458 University of Vermont (UVM) broths I & 11/461 Violet Red Bile Glucose (VRBG) agar//464 Violet Red Bile (VRB) agar (syn. violet red bile lactose agar)/467 Xylose Lysine Deoxycholate (XLD) agar/470
Appendix I
Testing methods for use in quality assurance of culture media/473
Appendix II
Test strains/479
Index/485

This Page Intentionally Left Blank

Part 1
Reviews of Media

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Chapter 1
Microbiological assessment of culture media" comparison and statistical evaluation of methods
G.H. Weenk Nutricia Research, P.O. Box 1, 2700 MA Zoetermeer, The Netherlands
In this review methods for the quality control of media were compared, taking the following questions as guidelines. (i) Which methods are easy to use and give reliable results? (ii) Which experimental design should be used in order to obtain reliable data with a minimal input of resources (staff and materials)? These questions can be answered satisfactorily using statistical methods. This review shows that solid media can be assessed with acceptable accuracy using well established methods like the spread plate technique. In order to assure a minimum of statistical error, at least two plates with an average count of 100 colonies per plate seems to be the best design. This also applies to the ecometric streaking technique, a good alternative to the more quantitative methods. For an accurate assessment of liquid media, large numbers of tubes need to be tested. This is very expensive in terms of laboratory resources and therefore unlikely to be used routinely. Therefore it is proposed to use the serial dilution technique, in which the broths are tested in triplicate (Richard, 1982).
The recommendations in this review can be used together with the methods recommended in this volume to assist laboratories setting up QC tests for culture media.
Introduction
The International Committee for Food Microbiology and Hygiene Working Party for Culture Media (ICFMH, WPCM) proposed a standardized scheme for the quality control (QC) of microbiological media (Baird et al., 1985), which was elaborated in a number of monographs (Baird et al., 1987; Baird et al., 1989 reviged and expanded in this volume).

In addition the scheme proposed criteria and techniques to test medium quality in order to:
�9 assess and compare the quality of commercially available dehydrated formulae or ready-to-use plates or tubes or their ingredients (Labots and Galesloot, 1960; Mossel et al., 1974, 1979);
�9 check the quality of purchased batches of commercially available media, before use (Mossel et al., 1980);
�9 check on medium preparation procedures (Mossel, 1970; Corry et al., 1986).
The testing methods used for QC purposes have also been applied in the development of new media or to enable food microbiologists to chose the most suitable medium for a defined purpose (Moussa et al., 1973; Morris et al., 1982; Terplan et al., 1982; Thomson, 1984; Mossel, 1985a; Peterz et al., 1985; Seiler, 1985; Mossel, 1986, 1989).
Several methods have been proposed with which the quality of media can be checked (Corry, 1982; Corry et al., 1982; Mossel, 1982; Reusse, 1982; Richard, 1982; Mossel et al., 1983; Baird et al., 1987). However, there is no guidance concerning the scope and field of application for each method. In this paper the available methods are reviewed. The methods are compared using statistics and a scope and field of application is presented. Statistics are also used to determine the most cost effective experimental design of culture medium QC in order to obtain results as accurate as possible and with a minimal input of labour and materials.
QC methods for solid media
The media which are to be checked for their performance are inoculated with overnight cultures of pure, well defined and appropriate test strains, preferably obtained from type culture collections. The use of mixed cultures and stressed strains has also been proposed for this purpose and a number of studies on medium performance have been carried out with naturally contaminated samples (Baird and van Doorne, 1982; Morris et al., 1982; Tzannetis et al., 1982; Peterz et al., 1985; Baird et al., 1987; Chain and Fung, 1991). However, such inocula are difficult to standardize and are usually specific to the laboratory or the foods and media which are to be investigated. Therefore these mixed cultures have, up to now, not been included in the list of test strains given in the ICFMH-monographs (Baird et al., 1987, 1989; this volume). The European Community Bureau of Reference has taken an initiative to develop and provide laboratories with standardized reference strains and mixed cultures for the QC of microbiological methods and media (Beckers et al., 1987; In 't Veld, 1991; In 't Veld et al., 1991). This work is carried out by the Dutch Institute of Health and Environmental

Hygiene and has already resulted in a number of reference samples, which are now being tested on a pilot scale. In the near future these samples should become available as certified samples and will certainly contribute significantly to perfecting the methods for medium QC.
The majority of the techniques used in the QC procedures for solid media rely on colony counting. The most widely used techniques are: the pour plate technique (Thomson, 1984); the spread plate technique (Peterz et al., 1985); the modified Miles-Misra technique (Miles et al., 1938; Corry, 1982) and the spiral plate technique (Gilchrist et al., 1973).
They are all routinely used in food microbiology laboratories and described in nationally and internationally recognized methods (Thomas and Thomas, 1975; Jarvis et al., 1977; ISO, 1987, 1988).
Colony counts are used to calculate the Absolute Growth Index (AGI) and Relative Growth Index (RGI), with which the selectivity and productivity of a medium are determined (Baird et al., 1987; van Netten et al., 1991). As plates are inoculated with overnight cultures, more than one plate is needed in order to obtain a reliable count of 30-300 colonies per plate. With the Miles-Misra technique several dilutions are inoculated onto one plate and with the spiral plater a wide range of counts can be assessed by using one plate, so both these methods use relatively few plates. They also are less labour intensive than the traditional spread plate technique. Of the four frequently used techniques, the spiral plater will have the lowest labour costs (Jarvis et al., 1977; Kramer et al., 1979). However, the high capital cost of spiral plater may outweigh savings in labour costs. Colony counting can be done automatically and thus speed up the collection of data and decrease the workload of the laboratory (Chain and Fung, 1991).
The colony morphology and the diagnostic system, summarized with the term 'electivity' (Baird et al., 1985, 1987), should also be checked in order to get a complete picture of the performance of a medium (van Netten et al., 1991). These cannot be checked properly using the pour plate or microcolony techniques such as the agar droplet technique (Richard et al., 1983; Adams, 1989).
T A B L E 1
S t r e n g t h - w e a k n e s s analys is o f co lony c o u n t i n g m e t h o d s u s e d for t he qua l i ty c o n t r o l o f sol id m e d i a
( a d a p t e d f r o m v a n N e t t e n et al. 1991)
T e c h n i q u e N u m b e r of Vis ib i l i ty of Cos t o f O v e r a l l
p l a t e s n e e d e d a C o l o n i a l D i a g n o s t i c L a b o u r ~ E q u i p m e n t ~ r e su l t
m o r p h o l o g y b sy s t em b
P o u r p l a t e . . . . + 3 -
S p r e a d p l a t e - + + - + 1 +
M i l e s - M i s r a + + + - + 3 +
Sp i r a l p l a t e + + + + - 3 +
a + _ lOW; -- -- h igh .
b + = good ; - = poo r .

Fig. 1. The ecometric streaking template.
Table 1 summarizes the weaknesses and strengths of the four most frequently used colony count techniques. This qualitative comparison shows a preference for using the spiral plate or Miles-Misra technique. The pour plate technique is not favoured, because it does not allow a proper analysis of the elective properties of a solid medium.
The ecometric streaking method was developed by Mossel and co-workers as a semi-quantitative alternative to the techniques described above (Mossel et al., 1983). It is the only technique which was specially designed to check the quality of solid media. Overnight cultures of test strains are inoculated onto the agar in a defined standardized way, as is shown in Fig. 1. One loopful of inoculum only is sequentially diluted from streak to streak. Growth on the plates is not recorded as a colony count, but as a score. Five streaks of growth in each quadrant score as one, growth on up to three streaks score 0.5. A maximum score of five is obtained when all streaks in the four quadrants show growth and the final streak in the centre of the plate is also colonized. The ecometric technique has successfully been applied by several authors to validate the quality and properties of solid media or to trace errors in medium preparation (Mossel et al., 1979, 1983; Corry et al., 1986; van Netten et al., 1991). The main advantages of the technique are that it is not labour intensive and the material costs are low since the overnight culture is diluted on one plate. However, the ecometric technique requires training in order to obtain reproducible results. As a dilution in colony numbers should be obtained from streak to streak, slight changes in the angle of the loop or the pressure applied to the loop during inoculation will lead to a disturbance of the desired
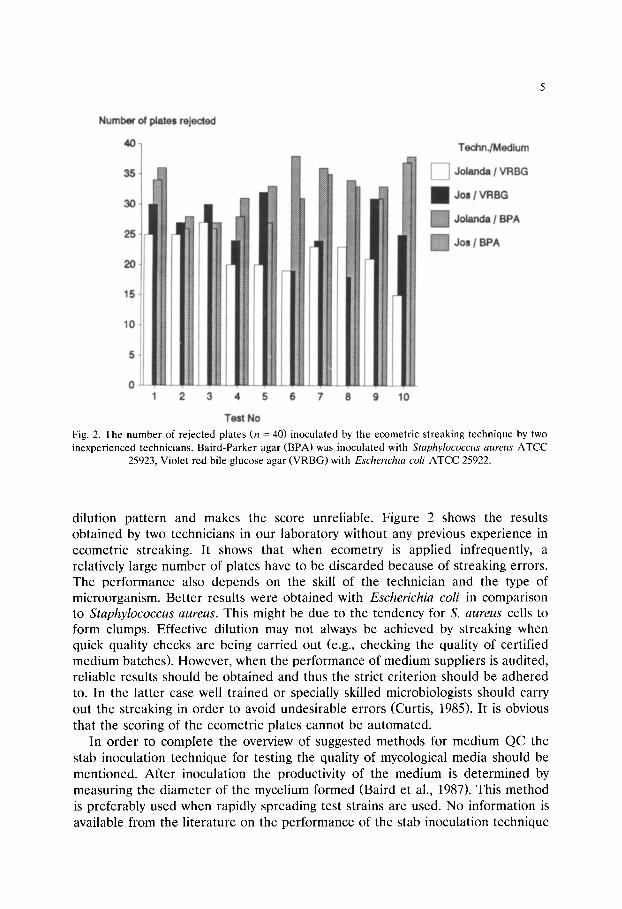
Fig. 2. The number of rejected plates (n = 40) inoculated by the ecometric streaking technique by two inexperienced technicians. Baird-Parker agar (BPA) was inoculated with Staphylococcus aureus ATCC
25923, Violet red bile glucose agar (VRBG) with Escherichia coli ATCC 25922.
dilution pattern and makes the score unreliable. Figure 2 shows the results obtained by two technicians in our laboratory without any previous experience in ecometric streaking. It shows that when ecometry is applied infrequently, a relatively large number of plates have to be discarded because of streaking errors. The performance also depends on the skill of the technician and the type of microorganism. Better results were obtained with Escher ich ia coli in comparison to S taphy lococcus aureus. This might be due to the tendency for S. aureus cells to form clumps. Effective dilution may not always be achieved by streaking when quick quality checks are being carried out (e.g., checking the quality of certified medium batches). However, when the performance of medium suppliers is audited, reliable results should be obtained and thus the strict criterion should be adhered to. In the latter case well trained or specially skilled microbiologists should carry out the streaking in order to avoid undesirable errors (Curtis, 1985). It is obvious that the scoring of the ecometric plates cannot be automated.
In order to complete the overview of suggested methods for medium QC the stab inoculation technique for testing the quality of mycological media should be mentioned. After inoculation the productivity of the medium is determined by measuring the diameter of the mycelium formed (Baird et al., 1987). This method is preferably used when rapidly spreading test strains are used. No information is available from the literature on the performance of the stab inoculation technique

TABLE 2
Sources of error in colony count procedures (from Jarvis 1989)
Source of error Includes errors due to
Sampling error Weighing Maceration Pipette volumes Diluent volumes Pipetting error Culture medium faults Incubation faults Non-randomness of propagules Counting errors 'Recording' errors
Dilution error
Plating error
Distribution error
Calculation error
(Curtis, 1985). As this method has reliable alternatives (Mossel et al., 1983; Thomson, 1984), it will not be discussed.
The statistics of methods used for the QC of solid media
In the QC of solid media the performance of the test medium is compared to that of a very nutritious reference medium (e.g., tryptone soy agar + 0.3% yeast extract). When both media are inoculated, errors can be made and thus should be allowed for when the medium quality is assessed. The errors associated with colony counting methods are extensively reviewed and further elaborated by Jarvis (1989) and are listed in Table 2. The contribution of each individual error source to the magnitude of the overall error may be method dependent. Therefore the methods used for the QC of solid media are divided into two groups: the quantitative methods (inoculation by spread plate, spiral plate and Miles-Misra technique) and the the semi-quantitative ecometric streaking technique. As they are based on different principles they will be treated separately.
Several authors have shown that there is no statistically significant difference in the performance of the spread plate, spiral plate and Miles-Misra technique (Badger and Pankhurst, 1960; Gilchrist et al., 1977; Jarvis et al., 1977; Kramer and Gilbert, 1978; Donegan et al., 1991). Therefore they will be regarded in the rest of the paper as one method and generally referred to as surface plating techniques.
The total error associated with colony counts obtained with surface plating techniques can be calculated from Table 2 according to the following formula given by Jarvis (1989):
% total error = + ( (A 2 + B2 + C 2)
where A = % sampling error; B = % distribution error; C = % dilution error.
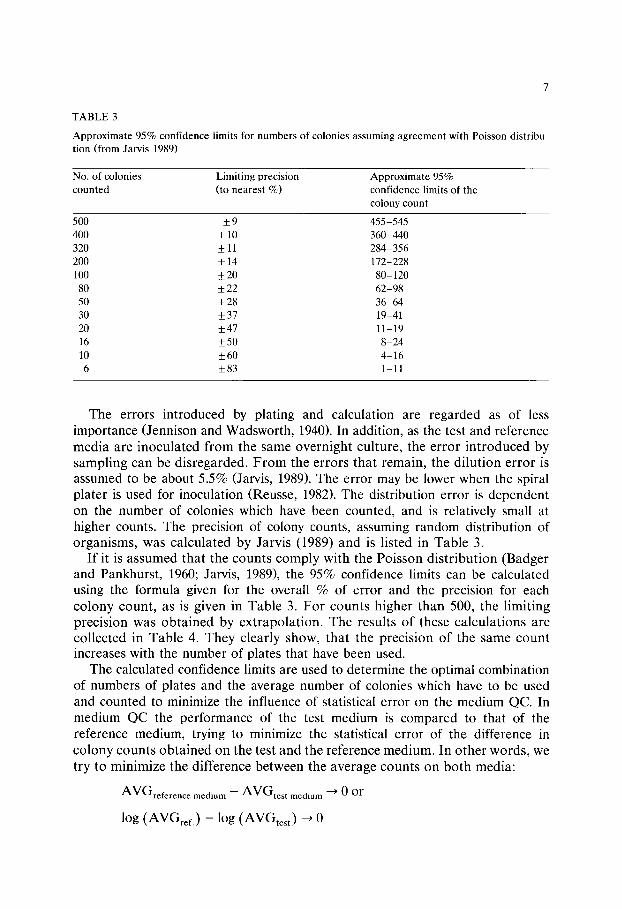
TABLE 3
Approximate 95% confidence limits for numbers of colonies assuming agreement with Poisson distribu- tion (from Jarvis 1989)
No. of colonies Limiting precision Approximate 95% counted (to nearest %) confidence limits of the
colony count
500 _+ 9 455-545 400 + 10 360-440 320 + 11 284-356 200 + 14 172-228 100 _+ 20 80-120
80 _+ 22 62-98 50 +_ 28 36-64 30 _+ 37 19-41 20 _+47 11-19 16 _+50 8-24 10 +_60 4-16 6 +_83 1-11
The errors introduced by plating and calculation are regarded as of less importance (Jennison and Wadsworth, 1940). In addition, as the test and reference media are inoculated from the same overnight culture, the error introduced by sampling can be disregarded. From the errors that remain, the dilution error is assumed to be about 5.5% (Jarvis, 1989). The error may be lower when the spiral plater is used for inoculation (Reusse, 1982). The distribution error is dependent on the number of colonies which have been counted, and is relatively small at higher counts. The precision of colony counts, assuming random distribution of organisms, was calculated by Jarvis (1989) and is listed in Table 3.
If it is assumed that the counts comply with the Poisson distribution (Badger and Pankhurst, 1960; Jarvis, 1989), the 95% confidence limits can be calculated using the formula given for the overall % of error and the precision for each colony count, as is given in Table 3. For counts higher than 500, the limiting precision was obtained by extrapolation. The results of these calculations are collected in Table 4. They clearly show, that the precision of the same count increases with the number of plates that have been used.
The calculated confidence limits are used to determine the optimal combination of numbers of plates and the average number of colonies which have to be used and counted to minimize the influence of statistical error on the medium QC. In medium QC the performance of the test medium is compared to that of the reference medium, trying to minimize the statistical error of the difference in colony counts obtained on the test and the reference medium. In other words, we try to minimize the difference between the average counts on both media:
AVGreference medium -- A V G t e s t medium ~ 0 or
l og ( A V G r e f . ) - l og ( A V G t e s t ) ~ 0
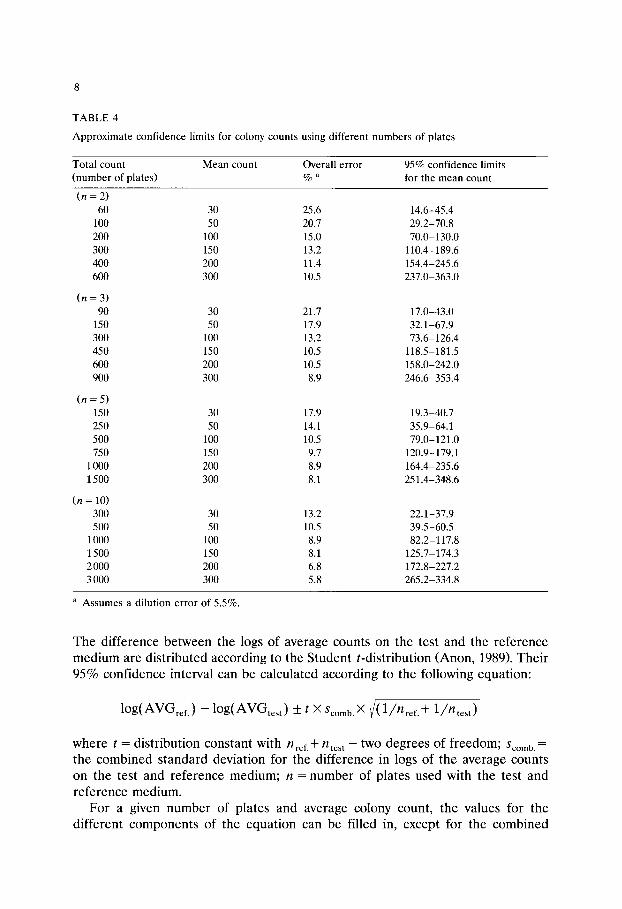
TABLE 4
Approximate confidence limits for colony counts using different numbers of plates
Total count Mean count Overall error 95% confidence limits (number of plates) % a for the mean count
( n = 2) 60 30 25.6 14.6-45.4
100 50 20.7 29.2-70.8 200 100 15.0 70.0-130.0 300 150 13.2 110.4-189.6 400 200 11.4 154.4-245.6 600 300 10.5 237.0-363.0
( n = 3) 90 30 21.7 17.0-43.0
150 50 17.9 32.1-67.9 300 100 13.2 73.6-126.4 450 150 10.5 118.5-181.5 600 200 10.5 158.0-242.0 900 300 8.9 246.6-353.4
( n = 5) 150 30 17.9 19.3-40.7 250 50 14.1 35.9-64.1 500 100 10.5 79.0-121.0 750 150 9.7 120.9-179.1
1000 200 8.9 164.4-235.6 1500 300 8.1 251.4-348.6
(n = 10) 300 30 13.2 22.1-37.9 500 50 10.5 39.5-60.5
1000 100 8.9 82.2-117.8 1500 150 8.1 125.7-174.3 2 000 200 6.8 172.8-227.2 3 000 300 5.8 265.2-334.8
a Assumes a dilution error of 5.5%.
The difference betWeen the logs of average counts on the test and the reference medium are distributed according to the Student t-distribution (Anon, 1989). Their 95% confidence interval can be calculated according to the following equation:
l o g ( A W G r e f . ) - l o g ( A V G t e s t ) + t X Scomb. X ~ ( 1 / n r e f . + 1 /n te s t )
where t = distribution constant with nref.-k- n t e s t - tWO degrees of freedom; Scomb . -
t h e combined standard deviation for the difference in logs of the average counts on the test and reference medium; n = number of plates used with the test and reference medium.
For a given number of plates and average colony count, the values for the different components of the equation can be filled in, except for the combined

standard deviation. For Scomb the following equation is valid:
Scomb.= ~(dfref. X s 2 -k- dftest X s 2 ) / ( d f r e f . + dftest )
When it is assumed that Sref. and Stest should in principle be equal, and for the sake of convenience the same numbers of plates are used for the test and the reference media, it can be derived that"
Scomb" = Sref.-- Stest = S
The s tandard deviation for the means can be derived from the 95% confidence intervals (CI) for the mean colony counts as they were calculated according to Jarvis (1989) and are collected in Table 4:
AVG _+ (overall error% • 2 • AVG) = (CI) or
(AVG + (CI)) - ( A V G - (CI)) = 4 • or
log(AVG + (CI)) - l o g ( A V G - (CI)) = 4 • s log units
So from the confidence intervals given in Table 4, the combined s tandard deviation can be calculated for each set of plates and average number of counted colonies, leading to the 95% confidence intervals for log(AVGref.)- log(AVGtest). These are shown in Fig. 3. As a reference, Fig. 3 also includes a set criterion for the productivity of a test medium: a medium is accepted when the (average) count on the test medium does not differ more than 0.7 log units from the growth on the reference medium (Baird et al., 1987).
Figure 3 clearly shows that the statistical error is minimized when the average count on the plate and the number of counted plates are increased. However, a mean count of more than 100 per plate does not significantly reduce statistical error when more than two plates are used. The use of more than three plates is not worthwhile as the statistical error is only reduced marginally. Therefore it is concluded that the use of three plates with an average count of about 100 colonies per plate is the opt imal combinat ion in respect to minimizing the statistical counting error in the QC of the media. For practical reasons it may however be decided to use two plates per medium. When an average count of about 100 CFU per plate is used, this seems to be acceptable. The maximum difference between test and reference medium introduced by statistical error, given by the sum of the 95% confidence intervals, still remains below the reference value of 0.7 log units.
The 95% confidence interval for the mean count defines the range of means which can be expected on average in 19 tests out of 20 when a mean is calculated from a set of colony counts. The interval is derived from the mean count calculated from the colony counts obtained from a number of samples. In other words the 95% confidence interval gives an estimate of the population within which the true mean count will lie. In one test out of 20 it is possible that the mean lies outside

10
Fig. 3. The 95% confidence interval (in log units) for log(AVGref.)-logAVG(tcst) in relation to the average plate count and the number of plates counted (n). As a reference, the criterion for the acceptance of non-selective media is included (log(AVGref)-log(AVGtest)~< 0.7 log units (Baird et al.,
1987).
this interval and might therefore result in the conclusion that the mean does not belong to this population. This is the so-called fault of the first order, and defines the chance that a mean may be unjustifiably discarded from the population. In addition, a second order fault defines the chance that a result is regarded as belonging to the population, although in fact it belongs to another one which partly overlaps. In order to calculate this chance, the following situation is taken as an example. The test medium and reference medium are checked for their productiv- ity, and are considered to perform equally well. So according to the criteria proposed by the ICFMH working party (Baird et al., 1987) their colony counts will lie within 0.7 log units from each other. In statistical terms the following chance should be determined:
P { 1 o g ( A g G r e f . ) - l o g ( A g G t e s t ) < 0.7}
under the condition that
ref . - ~ test :# 0
Where /Zref. and ~test are the population means.
11
PIF 2nd)In %
1 0 0 ""-.~ ...~ \
\ ',,
!x '\ ',\
2 O
",. \\ ~
0 . . . . I ' ' ; ' 1 " ' ' '
0 0,5 1 log ref- log test
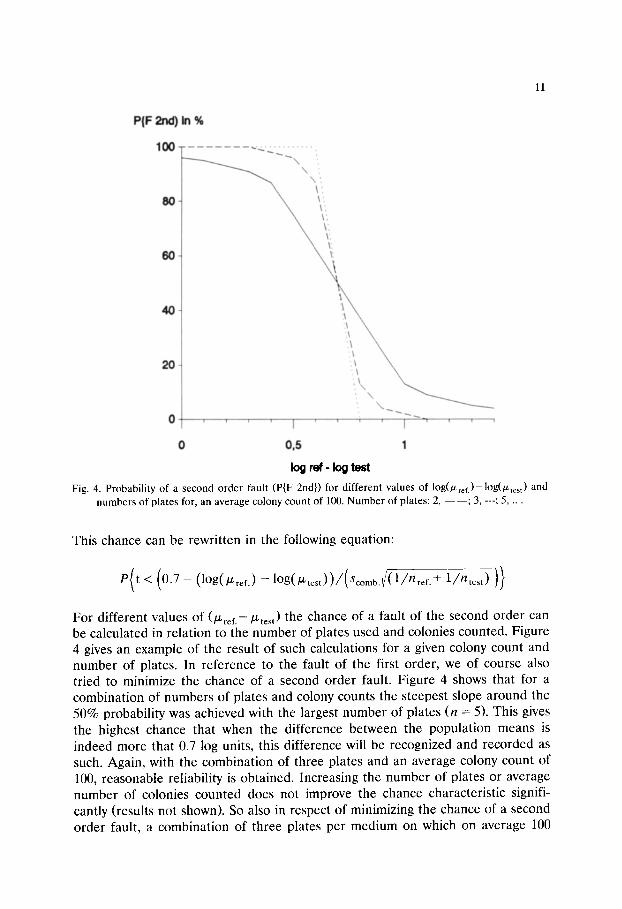
Fig. 4. Probability of a second order fault (P{F 2nd}) for different values of log(/Zref.)-log(~test) and numbers of plates for, an average colony count of 100. Number of plates: 2, - --- ; 3, ---; 5, ...
This chance can be rewritten in the following equation:
e ( t < ( 0 . 7 - (1og([/~ref.) --1og([Ztest))/(Scomb.~/(1/P'/ref. -]- 1/?/test ) )}
For different values of (~ref.--[Ltest) the chance of a fault of the second order can be calculated in relation to the number of plates used and colonies counted. Figure 4 gives an example of the result of such calculations for a given colony count and number of plates. In reference to the fault of the first order, we of course also tried to minimize the chance of a second order fault. Figure 4 shows that for a combination of numbers of plates and colony counts the steepest slope around the 50% probability was achieved with the largest number of plates (n = 5). This gives the highest chance that when the difference between the population means is indeed more that 0.7 log units, this difference will be recognized and recorded as such. Again, with the combination of three plates and an average colony count of 100, reasonable reliability is obtained. Increasing the number of plates or average number of colonies counted does not improve the chance characteristic signifi- cantly (results not shown). So also in respect of minimizing the chance of a second order fault, a combination of three plates per medium on which on average 100

12
Fig. 5. The relation between the colony count and the score on the ecogram. Each point represents the average score of five accepted plates. D, Escherichia coli on violet red bile agar, v, Staphylococcus
aureus on Baird-Parker agar.
colonies should be counted, is the best choice. When medium assessment is carried out in duplicate, it has to be accepted that the chance of mistakenly approving a medium batch, is significant.
The ecometric streaking technique is semi-quantitative and is based on an arbitrary scoring system (Mossel et al., 1980). The precision of the ecometric data is considerably lower than that of the surface plating techniques discussed above (Mossel et al., 1980). Figure 5 shows the relation between the ecometric score and the colony count as determined for S. aureus and E. coli grown on Baird-Parker agar and VRBG agar respectively. They confirm earlier observations that within the score of five a difference in colony counts of 1 log unit will not be observed (Mossel et al., 1980; van Netten et al., 1991). Mossel and co-workers (1980) calculated the confidence interval from the ecograms they obtained in several experiments and arrived at 0.6 scoring units for a score below 4.6. Compared to the confidence intervals calculated for the conventional surface inoculation techniques (Tables 3 and 4) this is relatively poor. However, the purpose of the medium QC determines whether this is important or not. The technique may have sufficient accuracy in medium development trials, where, in general, only large differences in medium performance are considered to be relevant.

13
TABLE 5
Criteria for the assessment of solid media for productivity and selectivity, using surface plating (SP) or the ecometric streaking technique (EC)
Medium Test strain Criterion SPa Criterion EC a
(Log(AVGref.) - Log(AVGtest)) AGI ref.-- AGI t e s t
Non-selective Wanted < 0.7 < 1 Selective Wanted < 1.0 < 1 Selective Not wanted > 5.0 4-5
a The test should be carried out in duplicate or triplicate and (for SP) with a count of at least 100 colonies per plate.
Several authors have proposed criteria and units of measure for medium assessment using the ecometric streaking technique (Mossel et al., 1983; Baird et al., 1987; van Netten et al., 1991). The statistics of the ecometric streaking technique, as shown in Fig. 5, allow these attempts to be harmonized. Figure 5 clearly shows that consistent scores are obtained at counts higher than 107 and below 102. At intermediate counts, the ecometric score jumps in steps of 0.5 units from 0 to 5. Assuming that overnight cultures contain 10 s to 109 microorganisms per ml and taking the criteria proposed for surface plating techniques (Baird et al., 1987), as a guide, criteria and units of measure can be derived. They are listed in Table 5 and agree very well with those proposed by van Netten and co-workers (1991). They use the Absolute Growth Index (AGI) as unit of measure.
It is not possible to calculate a statistically acceptable experimental design. However, in line with the statistics of the surface plating techniques, it is assumed that ecometric medium assessment should also be carried out in duplicate or triplicate in order to obtain reliable results.
Comparison of methods used for the QC of solid media
From the qualitative and statistical comparison of the methods which are frequently used for the quality control of media, a scope and field of application for each of the methods can be drawn up. In general, conventional surface plating techniques can be applied in every area of medium QC as they allow the accumulation of relatively precise quantitative (and qualitative) data. The ecomet- ric technique has its application in those areas where semi-quantitative data are sufficient, e.g., medium development where relatively large differences in medium performance are being investigated. However, the ecometric technique can also be the method of choice in the control of a number of good laboratory practices. It should however be stressed that for this purpose conventional surface plating techniques provide more accurate data. In a number of cases more than one method applies. Personal preference and cost will determine in these cases which

14
TABLE 6
Scope and field of application for methods for the quality control of solid microbiological media
Field of application Scope SPa EC b
Good laboratory practices Quality check of purchased batches of ready to use media (or their ingredients) before use + c + Check of medium preparation procedures + + Audit or compare the performance of _ medium suppliers + -
Medium development Relatively large differences are investigated + +
a SP = su r f ace p l a t e ( M i l e s - M i s r a + Spi ra l p l a t e + S p r e a d p l a t e ) t e c h n i q u e s .
b E C -- e c o m e t r i c t e c h n i q u e .
c + = m e t h o d r e c o m m e n d e d for this p u r p o s e ; - = m e t h o d is no t r e c o m m e n d e d for this p u r p o s e .
method a laboratory adopts. The scope and fields of application are summarized in Table 6.
QC methods for liquid media
Liquid media are used to promote (or inhibit) microbial growth, induce the production of visible metabolites or provoke specific reactions. Several methods for the QC of broths have been proposed and can be divided into tw6 groups (Baird et al., 1985, 1987; van Netten et al., 1991): methods which determine kinetic parameters and endpoint determinations.
The kinetic parameters relate to the growth curve: the lag phase a n d / o r the growth rate are used as criteria. It has been shown that they can be determined with great accuracy (Papadopoulou and Ioannidis, 1990). However, they have not been used frequently in medium QC and proper validated criteria are lacking (Curtis, 1985; van Netten et al., 1991). In addition they are relatively labour intensive and therefore seem to be more suitable for research purposes than for routine quality checks.
Endpoint determinations focus on the final result of growth: the increase in biomass. This can be determined indirectly by: measuring turbidity or changes in electrical properties, e.g., conductance, of the broth; estimating viable numbers within the broth; looking at the production of visible metabolites (e.g., gas) or reactive compounds (e.g., indole). Adding 15 g agar per litre to broth has also been suggested to determine its quality, using techn!ques developed for solid media (van Netten et al., 1991). As such an approach would limit the range of QC data (e.g., gas product ion would not be observed), the method is not recommended.
The most widely used method for the QC of liquid media is the serial dilution technique, as proposed by Richard (1982). This method is also used for assessment of recently developed semisolid media for salmonella detection (Baird et al., 1989).

TABLE 7
Strength-weakness analysis of methods used for the quality control of liquid media
15
Technique Accuracy a Visibility of Cost of diagnostic system a Labour b Equipment b
Overall result
Length of lag phase - + - - Growth rate + + - - Agar addition c + _ + + MPN + + + + Agar inoculation d + + _ +
2 m
0 2+ 4+ 2+
a 3r - - - good; - = poor. b . . [_ _ _ low; - = high. c Agar is added to the liquid medium. d Inoculation of agar after growth in the liquid medium.
A dilution series of an overnight culture is prepared in the test and reference highest titre at which microbia l activity is still appa ren t is scored. The serial dilu- tion technique is not as l abour intensive as the kinetic me thods and easy to dilution technique is not as labour intensive as the kinetic methods and easy to perform on a routine basis. Growth can either be determined by eye, by measure- ment of the optical density, by changes in conductance or by streaking onto agars (Mackey, 1985). In our hands determination of growth by eye gave, in general, as reliable results as using instrumentation (results not shown). Only opaque media, like Rappaport-Vassiliadis broth, gave problems and confirmation of growth by inoculation onto agar, as described by Stokes and Ridgeway (1979), became necessary. Mossel and co-workers determined the biomass at the end of growth by colony counting (Mossel et al., 1974). The same approach was taken by Carvalhal et al. (1991), who applied MPN statistics to microorganisms which were grown in liquid medium and subsequently transferred to agar. The method was shown to give as accurate results as the spread plate and drop plate methods. The use of solid media to determine the microbial yield at the end of growth also allows QC of broths in which mixed strains or naturally contaminated food samples are being used. It permits accurate quantification of growth yields after the test strains have been al lowed to express their physiological capacit ies in the broth. Table 7 gives an overview of the strengths and weaknesses of the methods .
The statistics of methods used for the QC of liquid media
Microbial growth in liquid media is generally considered to comply with the Poisson distribution (Cochran, 1950; Cowell and Morisetti, 1969; Jarvis, 1989). Before Poisson dis t r ibut ion can be applied the fol lowing condi t ions have to be met: the microorganism should be randomly distr ibuted in the broth (the microor- ganism is equally likely to be found in any par t of the b ro th and there is no

16
Fig. 6. The relation between the standard error of the log l0(MPN) (SE Log MPN) and the dilution factor and the number of replicate tubes used in the test (From Jarvis (1989)).
tendency to form pairs, groups or clusters of organisms or to repel one another); one or more microbes will result in good (turbid) growth in the medium.
In the QC of microbiological media these requirements are generally met, as pure overnight cultures are used. However, naturally contaminated samples have also been proposed for checking the performance of a medium (Mossel, 1985b). In such a case the distribution of microorganisms within the broth will very likely not conform with the Poisson distribution and therefore other distributions should be applied (Galesloot, 1986; Haas and Heller, 1988; Jarvis, 1989).
The accuracy of growth predictions based on a single inoculated tube is relatively poor (Halvorson and Ziegler, 1933). Therefore the 'Most Probable Number' (MPN) concept was developed, using dilution series and multiple tubes per dilution to predict the number of microorganisms (Cochran, 1950; Jarvis, 1989). The MPN became an important tool within food microbiology after ready to use tables became available (de Man, 1977, 1983). As it is not as accurate as colony counts (Thomas, 1955; McCarthy et al., 1958), attempts have been made to improve it (Aspinall and Kilsby, 1979; Reichart, 1991). With the aid of computer programs an optimal combination of dilutions, tubes per dilution and repeats can be defined in order to get an accurate MPN result with a minimal input of media, materials and labour (Hurley and Roscoe, 1983; Strijbosch, 1989). Figure 6 illustrates the relation between the standard error of the log(MPN) and the

17
number of tubes and the dilution factor used. It clearly shows that the accuracy of the estimate for the density of the microorganisms in the broth can be improved significantly when the dilution factor is decreased or the number of tubes per dilution are increased. Increasing the number of tubes to more than five tubes per dilution seems not to be worthwile.
The experimental design, defined by the dilution factor and the number of replicates, determines the accuracy of the quantitative data obtained from the QC of liquid media. Therefore the criteria for judgement of the quantitative perfor- mance of the test medium are also dependent on the experimental design. The following calculations will illustrate this, using the most probable number as estimate for the true density in the liquid medium.
The starting point for the statistical analysis is again the Null hypothesis:
H0: the productivity of the test and reference medium are the same. H~: the productivity of the test and reference medium are not the same.
The statistical significance of differences between the two M P N estimations can be calculated using the formula given by Jarvis (1989):
t - - ( log MPNref.-log MPNtest)/0.58V/((log(aref./nref. ) + (log(atest/ntest))
with t = Student t-test variable with (nref.+ ntest- 2) degrees of freedom; MPN = most probable number obtained with the test (MPi~ltest) and reference medium (MPNref.); a = dilution factor used in the tube series with test (atest) and reference medium (aref.); n = number of replicate tubes used in the tube series with test (ntest) and reference medium (nref.).
When a dilution factor lower than 10 is used, the formula should use 0.55 instead of 0.58. For each combination of dilution factors, tubes and the resulting t-value, the critical ratio of MPNref. and MPNtest can be calculated. At higher values of the critical ratio the Null hypothesis is rejected at a defined significance level (i.e., P = 0.05).
Table 8 illustrates the result of such calculations. For the sake of convenience the same dilution factor and number of tubes were used in the tube series with test and reference medium. Depending on the discriminating requirements of the test, an experimental design can be chosen. For instance, to be able to detect a difference between test and reference medium of 1 titre unit, five replicates per dilution and a dilution factor of 10 would be the best approach.
When the statistics of presence/absence tests are compared with the criteria proposed for medium assessment (Baird et al., 1987) it is obvious that large numbers of tubes have to be analysed in order to achieve an acceptable accuracy. This will impose a significant pressure on the resources of the lab and thus will discourage medium assessment studies. Therefore it seems to be more realistic to follow the serial dilution technique and to carry out the tests in triplicate (Richard, 1982). It has to be accepted that statistical error will significantly interfere with the results. Table 9 summarizes the criteria for the assessment of liquid media.
18
TABLE 8
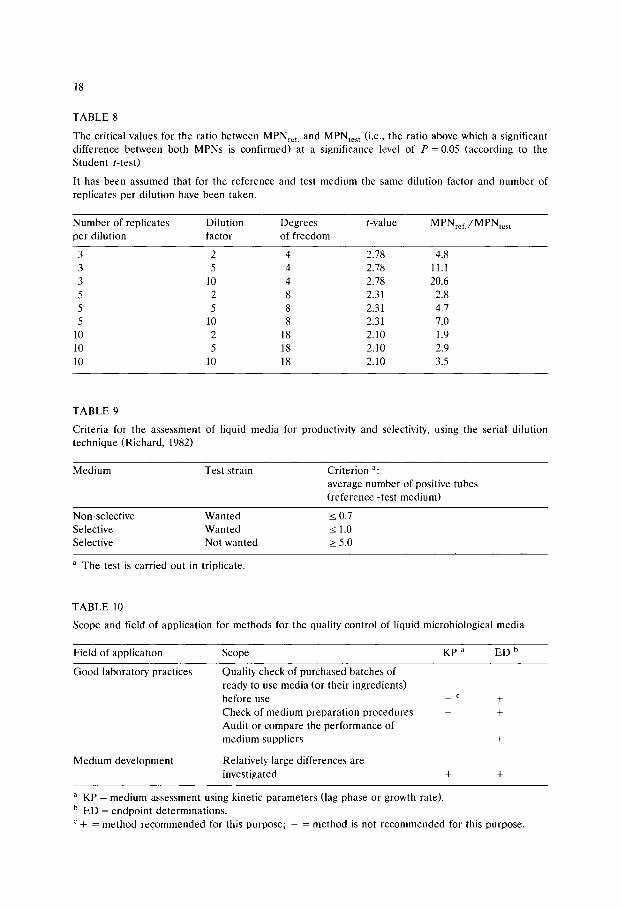
The critical values for the ratio between MPNref. and MPNtest (i.e., the ratio above which a significant difference between both MPNs is confirmed) at a significance level of P = 0.05 (according to the Student t-test)
It has been assumed that for the reference and test medium the same dilution factor and number of replicates per dilution have been taken.
Number of replicates Dilution Degrees t-value MPNrcf./MPNtest per dilution factor of freedom
3 2 4 2.78 4.8 3 5 4 2.78 11.1 3 10 4 2.78 20.6 5 2 8 2.31 2.8 5 5 8 2.31 4.7 5 10 8 2.31 7.0
10 2 18 2.10 1.9 10 5 18 2.10 2.9 10 10 18 2.10 3.5
TABLE 9
Criteria for the assessment of liquid media for productivity and selectivity, using the serial dilution technique (Richard, 1982)
Medium Test strain Criterion a: average number of positive tubes (reference-test medium)
Non-selective Wanted < 0.7 Selective Wanted < 1.0 Selective Not wanted > 5.0
a The test is carried out in triplicate.
TABLE l0
Scope and field of application for methods for the quality control of liquid microbiological media
Field of application Scope KP a ED b
Good laboratory practices Quality check of purchased batches of ready to use media (or their ingredients) before use _ c +
Check of medium preparation procedures - + Audit or compare the performance of medium suppliers - +
Medium development Relatively large differences are investigated + +
a K P = medium assessment using kinetic parameters (lag phase or growth rate). b ED = endpoint determinations. c + = method recommended for this purpose; - = method is not recommended for this purpose.

19
When the endpoint yield is determined by taking the number of positive tubes (p) out of n inoculated tubes (n 4: p), growth in the reference and test medium can be assessed and compared according to the Chi-squared test (Labots and Galesloot, 1960). The criteria for rejection or acceptance of medium batches, again depend on the number of tubes used. Where growth yields are expressed as CFUs, after subsequent inoculation onto agar (Mossel et al., 1974), the same statistics apply as shown for the QC of solid media.
Comparison of methods used for the QC of liquid media
With the introduction of growth determination equipment kinetic methods have become available for QC laboratories. Before they can be used properly the experimental design, and the criteria for rejection and release need to be defined. Currently, the methods of choice are therefore endpoint determinations. For this purpose the serial dilution technique is preferred. Good alternatives are growth yield determinations based on the total number of positive tubes or CFUs on agar inoculated from the test tubes. For all these methods, the experimental design and the resulting criteria are available. The proposed scope and fields of application are summarized in Table X. Endpoint determinations can be (routinely) used for all fields of application. Criteria for rejection and release of liquid media are not yet available for methods using the lag phase or growth rate as unit of measure. Therefore methods using kinetic parameters have not been recommended for use to check good laboratory practices.
Conclusions
In this overview methods for the quality control of media are compared, from the view point of a manager of a quality control laboratory. He or she will ask the following questions before undertaking extensive testing of media. Which methods are easy to use and give reliable results? Which experimental design should I use in order to obtain reliable data with a minimal input of resources (staff and materials)?
Using statistics as a yardstick, these questions can be answered satisfactorily. This overview shows that solid media can be assessed with acceptable accuracy using well established methods like the spread plate technique. In order to assure minimum statistical error at least two plates with an average count of 100 CFU per plate seems to be the best design. Although less accurate, the ecometric technique provides a good alternative to the more quantitative counting methods. In line with the experimental design proposed for the latter, at least two plates should be tested by ecometry to obtain a reliable result. For an accurate assessment of liquid media, large numbers of tubes need to be tested. This is expensive in laboratory resources. Therefore it is proposed to use the serial dilution technique, in which the broths are tested in triplicate, according to Richard (1982).

20
It is proposed to complete the criteria, as proposed by the ICFMH Working Party on the Quality Control of Microbiological Media, by adding the required experimental design (Tables 5 and 9). This will help QC laboratories to set up medium assessment as a quality control tool within the framework of Good Laboratory Practices and promote further standardization.
Acknowledgements
The author wishes to thank Prof. B. Jarvis, Dr F.P.G.M. la Fors, Dr P. van Netten and Dr L.W.G. Strijbosch for helpful discussions. The technical assistance of Jos Meeuwisse and Jolanda van den Brink is greatly appreciated.
References
Adams, M.R. (1989) Miscellaneous labour and materials saving methods. In: M.R. Adams and C.F.E. Hope (Eds.). Rapid Methods in Microbiology. Progress in Industrial Microbiology, 26, Elsevier, Amsterdam. pp. 239-254.
Anon. (1989) Introduction and statistics for microbiologists, course manual. Statistics for Industry, Knaresborough, UK.
Aspinall, L.J. and Kilsby, D.C. (1979) A microbiological quality control procedure based on tube counts. J. Appl. Bacteriol. 46, 325-330.
Badger, E.H.M. and Pankhurst, E.S. (1960) Experiments on the accuracy of surface drop bacterial counts. J. Appl. Bacteriol. 23, 28-36.
Baird, R.M. and van Doorne, H. (1982) Enrichment techniques for Staphylococcus aureus. In: J.E.L. Corry, R. Baird and G. Terplan (Eds.). Proceedings of the second Symposium of Quality Assurance and Quality Control of Microbiological Culture Media. February 27-28 1981, Dallas, USA. Archiv. Lebensmittelhyg. 33, 151-153.
Baird, R.M., Barnes, L.M., Corry, J.E.L., Curtis, G.D.W. and Mackey, B.M. (Eds.). (1985) Quality assurance and quality control of microbiological culture media. Int. J. Food Microbiol. 2, 1-136.
Baird, R.M., Corry, J.E.L. and Curtis G.D.W. (Eds.). (1987) Pharmacopoeia of culture media for food microbiology. Proceedings of the 4th International Symposium on Quality Assurance and Quality Control of Microbiological Culture Media. Manchester, 4-5 September. Int. J. Food Microbiol. 5, 187-299.
Baird, R.M., Corry, J.E.L., Curtis, G.D.W., Mossel D.A.A. and Skovgaard, N.P. (Eds.). (1989) Pharmacopoeia of culture media for food microbiology: additional monographs. Proceedings of the 5th International Symposium on Quality Assurance and Quality Control of Microbiological Culture Media. P6cs, Hungary, 23 August 1988. Int. J. Food Microbiol. 9, 85-144.
Beckers, H.J., Roberts, D., Pietsch, O., van Schothorst, M., Vassiliadis P. and Kampelmacher, E.H. (1987) Reference samples for checking the performance of Salmonella isolation methods. Int. J. Food Microbiol. 4, 51-57.
Carvalhal, M.L.C., Oliveira, M.S. and Alterthum, F. (1991) An economical and time saving alternative to the most-probable-number method for the enumeration of microorganisms. J. Microbiol. Meth- ods 14, 165-170.
Chain, V.S. and Fung, D.Y.C. (1991) Comparison of redigel, petrifilm, spiral plate system, isogrid and aerobic plate count for determining the numbers of aerobic bacteria in selected foods. J. Food Protect. 54, 208-211.

21
Cochran, W.G. (1950) Estimation of bacterial densities by means of the 'most probable number'. Biometrics 6, 1105-116.
Corry, J.E.L. (1982) Quality assessment of culture media by the Miles-Misra method. In: J.E.L. Corry (Ed.). Quality Assurance and Quality Control of Microbiological Culture Media. Proceedings of the Symposium held on 6-7 September 1979, Callas de Mallorca, Spain, G.I.T.-Verlag Ernst Giebeler, Darmstadt. pp. 21-38.
Corry, J.E.L., Baird, R. and Terplan, G. (Eds.). (1982) Proceedings of the second Symposium of Quality Assurance and Quality Control of Microbiological Culture Media. February 27-28 1981, Dallas, USA. Archiv. Lebensmittelhyg. 33, 137-175.
Corry, J.E.L., Leclerc, M., Mossel, D.A.A., Skovgaard, N., Terplan G. and van Netten, P. (1986) An investigation into the quality of media prepared and poured by an automatic system. Int. J. Food Microbiol. 3, 109-120.
Cowell, N.D. and Morisetti, M.D. (1969) Microbiological techniques: some statistical aspects. J. Sci. Food Agric. 20, 573-579.
Curtis, G.D.W. (1985) A review of methods for the quality control of culture media. Int. J. Food Microbiol. 2, 13-20
de Man, J.C. (1977) MPN tables for more than one test. Eur. J. Appl. Microbiol. 4, 307-316. de Man, J.C. (1983) MPN tables, corrected. Eur. J. Microbiol. Biotechnol. 17, 301-305. Donegan, K., Matyac, C., Seidler, R. and Portecous, A. (1991) Evaluation of methods for sampling,
recovery and enumeration of bacteria applied to the phylloplane. Appl. Environm. Microbiol. 57, 51-56.
Galesloot, T.E. (1986) Statistical remarks concerning the limiting dilution test used for the bacteriologi- cal testing of milk and milk products. Neth. Milk Dairy J. 40, 31-40.
Gilchrist, J.E., Campbell, J.E., Donelly, C.B., Peeler, J.T. and Delanay, J.M. (1973) Spiral plate method for bacterial determination. Appl. Microbiol. 25, 244-252.
Gilchrist, J.E., Donelly, C.B., Peeler, J.T. and Campbell, J.E. (1977) Collaborative study comparing the spiral plate and aerobic plate count methods. J. Assoc. Offic. Anal. Chem. 60, 807-812.
Haas, C.N. and Heller, B. (1988) Statistical approaches to monitoring. In: McFeters, A. (Ed.). Drinking Water Microbiology. Springer Verlag. New York. pp. 412-427.
Halvorson, H.O. and Ziegler, N.R. (1933) Application of statistics to problems in bacteriology. II. A consideration of the accuracy of dilution data obtained by using a single dilution. J. Bacteriol. 26, 331-339.
Hurley, M.A. and Roscoe, M.E. (1983) Automated statistical analysis of microbial enumeration by dilution series. J. Appl. Bacteriol. 55, 159-164.
In 't Veld, P.H. (1991) Towards certified reference materials for microbiology. De Ware(n) Chemicus. 21, 20-25.
In 't Veld, P.H., Soentoro, P.S., Delfgou-van Asch, E.H.M. and Notermans, S. (1991) Influence of reconstitution on isolation and enumeration of Listeria monocytogenes from milk powder used for reference samples. J. Food Protect. 54, 124-126.
ISO. (1987) Microbiology. General guidance for enumeration of Bacillus cereus: colony count technique at 30~ International Standard ISO 7932.
ISO. (1988) Meat and meat products. Enumeration of micro-organisms: colony count technique at 30~ (Reference method). International Standard ISO 2293.
Jarvis, B., Lach, V.H. and Wood, J.M. (1977) Evaluation of the spiral plate maker for the enumeration of micro-organisms in foods. J. Appl. Bacteriol. 43, 149-157.
Jarvis, B. (1989) Statistical aspects of the microbiological analysis of foods. Progress in Industrial Microbiology, 21. Elsevier Scientific Publishers B.V., Amsterdam.
Jennison, M.W. and Wadsworth, G.P. (1940) Evaluation of the errors involved in estimating bacterial numbers by the plating method. J. Bacteriol. 43, 149-157.
Kramer, J.M. and Gilbert, R.J. (1978) Enumeration of microorganisms in food: a comparative study of five methods. J. Hyg., Cambridge 61, 151-159.
Kramer, J.M., Kendall, M. and Gilbert, R.J. (1979) Evaluation of the spiral plate and the laser colony counting techniques for the enumeration of bacteria in foods. Eur. J. Appl. Microbiol. Biotechnol. 6, 289-299.

22
Labots, H. and Galesloot, Th.E. (1960) Een onderzoek naar de geschiktheid van het gedroogde briljantgroengallactose medium van oxo ltd voor het aantonen van coliachtige bacteriEn in melk. NIZO beproevingsrapport 25.
Mackey, B.M. (1985) Quality control monitoring of liquid selective enrichment media used for isolating salmonellae. Int. J. Food Microbiol. 2, 41-48.
McCarthy, J.A., Thomas, H.A. and Delaney, J.E. (1958) Evaluation of the reliability of coliform density tests. Am. J. Public Health. 48, 1628-1635.
Miles, A.A., Misra, S.S. and Irwin, J.O. (1938) The estimation of the bacteriocidal power of blood. J. Hyg. 38, 732-749.
Morris, G.K., Bopp, C.A., Patton, C.M. and Wells, J.G. (1982) Media for isolating Campylobacter. In: J.E.L. Corry, R. Baird. and G. Terplan (Eds.). Proceedings of the second Symposium of Quality Assurance and Quality Control of Microbiological Culture Media. February 27-28 1981, Dallas, USA. Archiv. Lebensmittelhyg. 33, 151-153.
Mossel, D.A.A., (1970) Microbiological culture media as ecosystems. In: J. van Bragt, D.A.A. Mossel, R.L.M. Pierik, and H. Veldstra (Eds.). Effects of sterilization on components in nutrient media. H. Veenman & Zonen N.V. Wageningen, pp. 15-40.
Mossel, D.A.A., Harrewijn, G.A. and Nesselrooy-van Zadelhoff, C.F.M. (1974) Standardisation of the selective inhibitory effect of surface active compounds used in media for the detection of Enterobac- teriaceae in foods and water. Health Lab. Sci. 11,260-267.
Mossel, D.A.A., van Rossem, F. and Rantama, A. (1979) Ecometric monitoring of agar immersion plating and contact (AIPC)-slides used in assuring the microbiological quality of perishable foods. Lab. Pract. 28, 470-475.
Mossel, D.A.A., van Rossem, F., Koopmans, M., Hendriks, M., Verouden M. and Eelderink, I. (1980) A comparison of the classical and the so-called ecometric technique for the quality control of selective culture media. J. Appl. Bacteriol. 49, 439-454.
Mossel, D.A.A. (1982) Ecological essentials of the use of selective culture media in public health microbiology. In: J.E.L. Corry (Ed.). Quality Assurance and Quality Control of Microbiological Culture Media. Proceedings of the Symposium held on 6-7 September 1979, Callas de Mallorca, Spain, G.I.T.-Verlag Ernst Giebeler, Darmstadt. pp. 11-19.
Mossel, D.A.A., Bonants-van Laarhoven, T.M.G., Lichtenberg-Merkus, A.M.T. and Werdler, M.E.B. (1983) Quality assurance of selective culture media for bacteria, moulds and yeasts: an attempt at standaardisation at the international level. J. Appl. Bacteriol. 54, 313-327.
Mossel, D.A.A. (1985a) Media for Enterobacteriaceae. Int. J. Food Microbiol. 2, 27-32. Mossel, D.A.A. (1985b) Introduction and prospective. Int. J. Food Microbiol. 2, 1-7. Mossel, D.A.A. (1986) Developing methodology for foodborne microorganisms-fundamentals of analyti-
cal techniques. In: M.D. Pierson, and N.J. Stern (Eds.). Foodborne Microorganisms and their Toxins: Developing Methodology. Marcel Dekker, New York. pp. 1-22.
Mossel, D.A.A. (1989) Listeria monocytogenes in foods. Isolation, characterization and control. Int. J. Food Microbiol. 8, 183-195.
Moussa, R.S., Keller, N., Curiat, G. and de Man, J.C. (1973) Comparison of five media for the isolation of coliform organisms from dehydrated and deep frozen foods. J. Appl. Microbiol. 36, 619-624.
Papadopoulou, C. and Ioannidis, K. (1990) Differentation of S. gallinarum and S. pullorum by means of growth-kinetics analysis. J. Microbiol. Methods 11,247-253.
Peterz, M., Wiberg, C. and Norberg, P. (1985) Comparison of media for isolation of Bacillus cereus from foods. J. Food Protect. 48, 969-970.
Reichart, O. (1991) Some remarks on the bias of the MPN method. Int. J. Food Microbiol. 13, 131-142. Reusse, U. (1982) The use of the stomacher and spiral plate methods in food microbiology. In: J.E.L.
Corry (Ed.). Quality Assurance and Quality Control of Microbiological Culture Media. Proceedings of the Symposium held on 6-7 September 1979, Callas de Mallorca, Spain, G.I.T.-Verlag Ernst Giebeler, Darmstadt. pp. 59-61.
Richard, N. (1982) Monitoring the quality of selective liquid media used in the official serial dilution technique for the bacteriological examination of food. In: J.E.L. Corry (Ed.). Quality Assurance and Quality Control of Microbiological Culture Media. Proceedings of the Symposium held on 6-7 September 1979, Callas de Mallorca, Spain, G.I.T.-Verlag Ernst Giebeler, Darmstadt. pp. 51-58.

23
Richard, J., Moller Andersen, H. and Gratadoux, J.J. (1983) Rapid method for selecting appropriate solid media for the enumeration of aerobic micro-organisms. J. Appl. Bacteriol. 54, 329-334.
Seiler, D.A.L. (1985) Monitoring mycological media. Int. J. Food Microbiol. 2, 123-131. Stokes, E.J. and Ridgway, G.L. (1979) Media-testing and other techniques. In: Clinical bacteriology. 5th
Edn. Edward Arnold, London. pp. 342-381. Strijbosch, L.W.G. (1989) Experimental design and statistical evaluation of limiting dilution assays. PhD
Thesis. University of Maastricht (The Netherlands). Terplan, G., Zaadhof, K.J. and Becker, H. (1982) Quality assurance of newer media for the enumera-
tion of Staphylococcus aureus in food. In: J.E.L. Corry, R. Baird. and Terplan, G. (Eds.). Proceedings of the Second Symposium of Quality Assurance and Quality Control of Microbiological Culture Media. February 27-28 1981, Dallas, USA. Archiv. Lebensmittelhyg. 33, 151-153.
Thomas, H.A. (1955) Statistical analysis of coliform data. Sewage Ind. Wastes. 27, 212-222. Thomas, S.B. and Thomas, B.F. (1975) The bacteriological grading of bulk collected milk. Dairy
Industries. July, 262-265. Thomson, G.F. (1984) Enumeration of yeasts and moulds: media trial. Food Microbiol. 1,223-227. Tzannetis, S.E., Papavassilliou, J. and Konidaris, N. (1982) The production of selective media for
Lancefield group D streptococci after damage due to heat and cold. In: J.E.L. Corry, R. Baird. and G. Terplan (Eds.). Proceedings of the second Symposium of Quality Assurance and Quality Control of Microbiological Culture Media. February 27-28 1981, Dallas, USA. Archiv. Lebensmittelhyg. 33, 151-153.
van Netten, P., Weenk, G. and van der Zee, H. (1991) Quality control of culture media. De Ware(n) Chemicus. 21, 26-60.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
25
Chapter 2
Principles involved in the detection and enumeration of clostridia in foods
G.C. Mead Department of Animal Health, Royal Veterinary College, Boltons Park, Hawkshead Road, Herts EN6 INB, UK
The clostridia are a group of anaerobic bacteria that vary considerably in their biochemical and physiological properties. Not surprisingly, attempts to develop a single isolation medium for all species that occur in foods have not been entirely successful, and the problem is compounded by the need to recover both vegetative cells and spores, some of the latter being unable to germinate without heat activation.
Most available isolation media, except some of those used in the dairy industry, include sulphite and an appropriate iron salt, so that blackening due to sulphite reduction can serve as a differential test for clostridia. The limitations of this test in solid agar media are discussed and some advantages described in relation to its use in liquid media for Most Probable Number determinations. A medium favoured for the purpose is the Differential Reinforced Clostridial Medium of Gibbs and Freame (1965).
An unresolved issue is whether or not special precautions are needed to exclude oxygen during food sample preparation and dilution, preparation of media, and in conditions used for anaerobic incuba- tion. Although such stringency may be required for maximum recovery of sub-lethally damaged cells or spores, practical constraints in food control laboratories necessitate use of relatively simple procedures for detecting clostridia routinely.
Introduction
T h e g e n u s Clostr idium c o m p r i s e s a l a rge a n d m e t a b o l i c a l l y d ive r se co l l ec t ion of
bac t e r i a , all of wh ich a re a n a e r o b i c s p o r e - f o r m e r s . B e r g e y ' s M a n u a l of S y s t e m a t i c
B a c t e r i o l o g y ( H o l t e t al., 1986) lists 83 spec ie s a n d o t h e r s have b e e n a d d e d in
r e c e n t years , bu t r e la t ive ly few a re l ikely to be e n c o u n t e r e d in foods . T h o s e t h a t
a re c o m m o n as food c o n t a m i n a n t s a r e ma in ly m e s o p h i l e s ; n e v e r t h e l e s s , s o m e
t h e r m o p h i l e s a r e i m p o r t a n t in r e l a t i o n to spo i l age of c a n n e d foods , a n d f r o m t ime
to t ime c los t r id ia l spo i l age of ch i l l - s t o r ed p r o d u c t s is a lso r e p o r t e d , e.g., milk,
p o t a t o e s , m e a t ( B h a d s a v l e et al., 1972; B r o c k l e h u r s t a n d L u n d , 1982; D a i n t y et al.,
1989).

26
According to Gibbs and Freame (1965), contamination of foods with clostridia is largely derived from soil and, of the species normally present, only Cl. perfringens is also common in faeces. Unless food is badly mishandled, the occurrence of clostridia is likely to be very low.
From the public health viewpoint, the two most important species are CL botulinum and Cl. perfringens, but some other species or strains are also known to be hazardous. The organisms in question include Cl. barati, Cl. butyricum, Cl. difficile, Cl. sordellii, Cl. sphenoides and Cl. spiroforme and all would be of possible concern in foods such as powdered formulations for infants (Mossel, 1989).
In many countries, 'sulphite-reducing clostridia' may be sought as index organ- isms for CL botulinum, as general hygiene indicators, or as a means of detecting faults in food processing. Such tests are not generally favoured in the United Kingdom, however, because of questions concerning their relevance and suitability, as well as the known limitations of existing media and methods (see below).
Factors affecting isolation
Vegetative cells of clostridia tend to be oxygen-sensitive and usually are de- stroyed by heat processing. Thus, some foods, e.g., dried milk products, will contain only spores. In raw meats, on the other hand, conditions are more favourable for survival of vegetative cells and these tend to outnumber any spores. It is clear, therefore, that laboratory isolation procedures should be equally suitable for both viable forms of clostridia.
In practice, there are inherent difficulties in meeting this requirement. For rapid spore germination, heat treatment is often necessary, but inactivates vegeta- tive cells. When beef was inoculated with spores of a heat-resistant strain of Cl. perfringens, Barnes et al. (1963) found that only about 3% germinated without prior heat shock over a range of storage temperatures. Furthermore, not only do strains of clostridia vary with respect to the heating regime needed for optimum germination, but spores in a single population germinate at different rates, suggesting a need for prolonged incubation of isolation media. In addition, some clostridia, e.g., Cl. botulinum type E, produce unusually heat-sensitive spores which may be inactivated by the degree of heat treatment used to detect spores in the laboratory.
Other factors have been discussed by Mossel and de Waart (1968) and involve the nature of the isolation medium, the varying properties of different clostridia and the type of food under examination. Clostridia differ widely in their energy requirements and some grow poorly, if at all, in media that do not contain a fermentable carbohydrate. However, where sugars are fermented, abundant gas is generally produced, causing considerable disruption of agar media. For this reason, liquid media may be preferred when inclusion of a fermentable carbohy- drate is deemed necessary. Because the clostridia are so diverse, it is difficult to find selective agents that are equally favourable for all relevant species whilst inhibiting other organisms, such as some coliform bacteria, that are likely to grow

27
under the test conditions. The few alternatives were considered by Gibbs and Freame (1965), and only polymyxin seems to be widely applicable for this purpose, although it is not entirely selective for clostridia.
The nature of the food being examined in any instance will determine whether damaged spores are likely to be present as a result of, e.g., heat processing, or whether organisms capable of interfering with clostridial growth and differentia- tion may cause a problem. Since clostridia are frequently present in only small numbers, there is also the possibility that food ingredients may interfere with the isolation medium at low dilutions of the test sample.
Types of isolation media
It was stated by Gibbs and Freame (1965) that "there is still no fully satisfactory medium for selecting the clostridia as a whole". More than 25 years later, this statement remains true and, during the intervening years, no major advances in media development have occurred. Those media that are presently available include both liquid and solid formulations, selective and non-selective, and virtu- ally all depend on sulphite reduction as the key differential criterion.
Sulphite reductase activity is a common property among clostridia that is found in only a few other bacteria capable of growth in clostridial media. Some clostridia, however, are particularly sensitive to sulphite, despite their ability to reduce it. Among the species known to include sensitive strains are Cl. butyricum, Cl. saccharobutyricum, Cl. septicum, Cl. sporogenes and Cl. tertium. For this reason, most isolation media contain no more than 0.05% sulphite (e.g., Gibbs and Freame, 1965; Mossel and de Waart, 1968; Weenk et al., 1991).
Reduction of sulphite and precipitation of the resultant sulphide as a black deposit involves an appropriate iron salt that yields iron sulphide. With solid media, the reaction is seen as a black halo around each colony. Inclusion of a fermentable carbohydrate in the medium can lead to a rapid fall in pH during bacterial growth and failure to precipitate the sulphide. This is generally avoided by restricting the sugar concentration to 0.1% and incorporating a buffer in the medium.
Sulphite-containing media are used in both solid and liquid form. A liquid medium that has been used widely and permits counts of clostridia by the Most Probable Number (MPN) method is the differential reinforced clostridial medium (DRCM) of Gibbs and Freame (1965). The basic medium contains no selective inhibitors and, when used in screw-capped bottles that leave only a small head- space, it can be incubated aerobically. Reducing conditions are maintained by inclusion of cysteine. The procedure recommended for detection and enumeration of clostridia in foods is given in Table 1. Following heat treatment of the food-sample suspension to eliminate vegetative cells and activate spores, D R C M is used to obtain a spore count. In parallel, the medium is used without any heat treatment. This yields a presumptive 'total' count of clostridia in the sample, i.e., vegetative cells plus (some) spores, but all bottles showing blackening are heat-

28
TABLE 1
Detection and enumeration of clostridia using the differential reinforced clostridial medium (DRCM) of Gibbs and Freame (1965)
Food homogenate (1 : 10) in 0.1% peptone diluent
I I Heat sample in bottles MPN in DRCM
(78~ for 30 min) ]
Black bottles give I
MPN in DRCM presumptive "total" count
I I Black bottles give Heat in situ
spore count (78~ for 30 min)
I Subculture into fresh DRCM
I Black bottles give
"total" count
For most purposes, spore counts are obtained after incubation at 37~ for 2 days, while 'total ' counts require 3 days at the same temperature. Different time-temperature regimes may be more appropriate in some instances. Gibbs and Freame (1965) make no specific recommendations in this respect. MPN, most probable number.
treated as before and subcultured into flesh DRCM to confirm the presence of clostridia. Thus, the overall method is relatively laborious for routine purposes, especially with the suggested use of five replicates per dilution. However, Gibbs and Freame (1965) indicate a number of advantages in the MPN method, including a greater sensitivity by comparison with solid media in detecting low numbers of clostridia, because of the larger inoculum. They also claim a higher recovery of the organisms from liquid rather than solid media and lower toxicity to clostridia of any selective agents used. Apart from the added advantage mentioned previously that a liquid medium is not disrupted by gas produced during sugar fermentation, the DRCM method also allows development of blackening to be followed without the need to disturb conditions inside an anaerobic jar.
Despite the advantages of DRCM, some limitations must be recognised. The MPN method is less accurate than direct counting and DRCM itself can support the growth of other organisms capable of producing blackening. These include strains of Bacteroides spp. and Enterobacteriaceae (Gibbs and Freame, 1965). Hence, pasteurisation of unheated cultures is required for confirmation. This assumes, of course, that any clostridia present in cultures showing blackening will have formed heat-resistant spores in the medium. In the case of Cl. perfringens, sporulation in laboratory media tends to be poor, and recovery of vegetative cells is likely to be low in DRCM because of failure to confirm on heating (Gibbs, 1971).
Recently, a modified DRCM has been evaluated for isolating clostridia from foods (Weenk et al., 1991). In this study, the medium (MDRCM)was used in a

29
solid form for deep agar cultures and incubated at 30~ for up to 3 days. Blackening by clostridia was facilitiated by using 0.1% ferric ammonium citrate and samples were pasteurised for at least one minute at 80~ before inoculation. The method gave good results, but the occurrence of Bacillus spp. that formed small sulphite-reducing colonies necessitated confirmatory tests for metronidazole sensitivity and inability to grow aerobically, as distinguishing features of clostridia. Attempts to prevent inhibition of clostridial blackening in MDRCM due to copious acid formation involved addition of N-2-hydroxyethylpiperazine-N'2-ethane- sulphonic acid and sodium bicarbonate to increase buffering, but no clear benefit was evident.
Use of lactate instead of glucose in isolation media to reduce growth of lactic acid bacteria was suggested by Hirsch and Grinsted (1954). In practice, Weenk et al. (1991) found 1.5% lactate to be inhibitory for some clostridia when incorpo- rated in MDRCM.
Liquid media used with the MPN method for counting clostridia in the dairy industry are sometimes based on milk, which supports growth of cheese spoilage species such as Cl. butyricum, Cl. sporogenes and Cl. tyrobutyricum. The milk used is that under test. After heating to kill vegetative cells, the milk surface in each tube is sealed with a paraffin plug, which is subsequently displaced by growth of gas-producing clostridia and thus is indicative of their presence. In a study of five MPN methods, Senyk et al. (1989) evaluated two milk-based media, one of which contained added glucose and lactate. These were compared with laboratory media containing lactate, either in liquid or semi-solid form, and again sealed with paraffin or agar. The fifth medium was essentially Reinforced Clostridial Medium with added lactate and an agar seal. All the test media were inoculated with spores of pure cultures of the relevant organisms and incubated at 32 or 37~ for up to 14 days. The milk media gave the lowest recoveries of clostridia and gas formation occurred more slowly in comparison with the other media. This may have been due to the absence of any added reducing agent in the milk substrate. Even with the most effective media, however, long periods of incubation were needed to detect small inocula (Table 2). The medium favoured by Senyk et al. (1989) on the basis of simplicity and sensitivity was the modified BB-lactate formulation. It contained (g/l): peptone, 15; beef extract, 10; yeast extract, 5; L-cysteine HC1, 0.5; sodium acetate, 5; agar, 2; sodium lactate (60%), 8.4 ml; pH 6.0. The tubes were sealed after inoculation with 2% agar.
As with liquid media used for MPN determinations, the available solid media for direct counting vary in both nutrient status and selectivity. For example, the iron sulphite agar of Attenborough and Scarr (1957) is a medium of low nutrient content that is suitable for detecting spores of thermophiles such as Desulfotomac- ulum (Clostridium) nigrificans. In this case, considerable selectivity is achieved by incubating the medium at 56~ For mesophilic species, one of the more selective solid media is the sulphite-polymyxin-sulphadiazine (SPS) agar of Angelotti et al. (1962). The medium, which contains tryptone and yeast extract, with no added carbohydrate, was designed for selective isolation of Cl. perfringens but was regarded by Gibbs and Freame (1965) as the best solid medium for clostridia as a
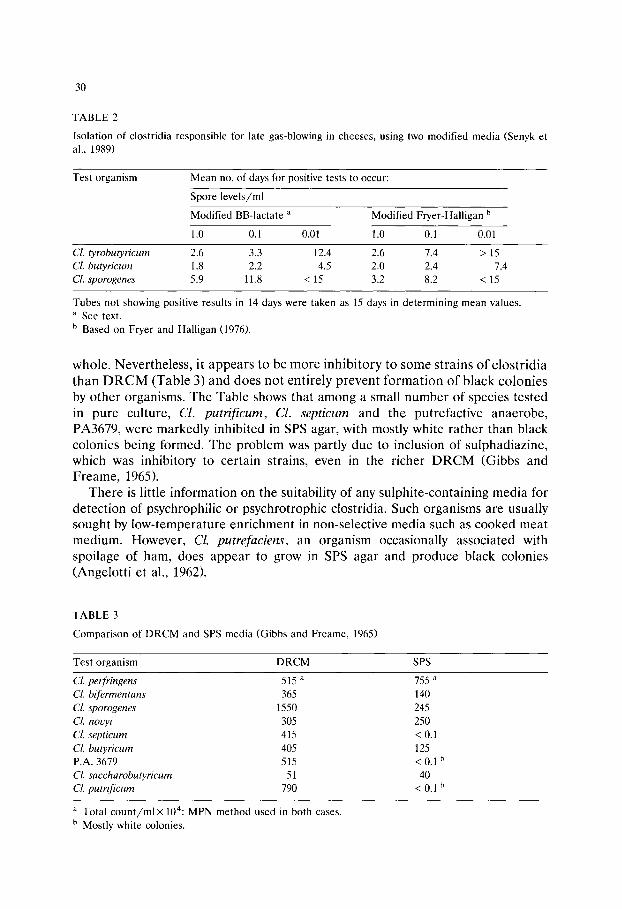
30
T A B L E 2
Isolation of clostridia responsible for late gas-blowing in cheeses, using two modified media (Senyk et al., 1989)
Test organism Mean no. of days for positive tests to occur:
Spore levels/ml
Modified BB-lactate ~ Modified Fryer-Halligan b
1.0 0.1 0.01 1.0 0.1 0.01
Cl. tyrobutyricum 2.6 3.3 12.4 2.6 7.4 > 15 Cl. butyricum 1.8 2.2 4.5 2.0 2.4 7.4 Cl. sporogenes 5.9 11.8 < 15 3.2 8.2 < 15
Tubes not showing positive results in 14 days were taken as 15 days in determining mean values. a See text. b Based on Fryer and Halligan (1976).
whole. Nevertheless, it appears to be more inhibitory to some strains of clostridia than D R C M (Table 3) and does not entirely prevent formation of black colonies by other organisms. The Table shows that among a small number of species tested in pure culture, Cl. putrificum, Cl. septicum and the putrefactive anaerobe, PA3679, were markedly inhibited in SPS agar, with mostly white rather than black colonies being formed. The problem was partly due to inclusion of sulphadiazine, which was inhibitory to certain strains, even in the richer DRCM (Gibbs and Freame, 1965).
There is little information on the suitability of any sulphite-containing media for detection of psychrophi|ic or psychrotrophic clostridia. Such organisms are usually sought by low-temperature enrichment in non-selective media such as cooked meat medium. However, Cl. putrefaciens, an organism occasionally associated with spoilage of ham, does appear to grow in SPS agar and produce black colonies (Angelotti et al., 1962).
T A B L E 3
Comparison of DRCM and SPS media (Gibbs and Freame, 1965)
Test organism D R C M SPS
Cl. perfringens 515 a 755 ~
Cl. bifermentans 365 140
Cl. sporogenes 1550 245
Cl. nouyi 305 250
Cl. septicum 415 < O. 1 Cl. butyricum 405 125 P.A. 3679 515 < 0.1 b
Cl. saccharobutyricum 51 40 Cl. putrificum 790 < 0.1 b
a Total count /mix 104: M P N method used in both cases. b Mostly white colonies.

31
The use of sulphite-containing media for isolation and enumeration of Cl. perfringens was reviewed by Mead (1985) and will not be discussed in detail here. Even with this single species it has not been possible to develop an entirely specific medium, but use of appropriate antibiotics gives greater scope for selectivity than is likely for clostridia as a whole. With some media, selectivity for Cl. perfringens can be further increased by incubation at 46~ again an approach that is not applicable to all clostridia found in foods, because of their varying temperature requirements.
Detection of Cl. botulinum in foods is more concerned with demonstrating the presence of neurotoxin than isolating the organism itself. Moreover, the species actually encompasses a small group of metabolically different organisms, all of which happen to produce a similar toxin. Because of this diversity, it is unlikely than any single detection method or protocol will be equally suitable for all types or for all kinds of food sample (Hobbs et al. 1982). In principle, the first stage of isolation invariably requires an enrichment step involving either incubation of the food sample or inoculation of a non-selective medium. Enrichment cultures are plated on suitable agar media, e.g., egg yolk agar and purified for toxin testing. There seems to be some doubt about the value of using selective sulphite-contain- ing media to enrich for Cl. botulinum (Hobbs et al., 1982).
Sample preparation and conditions of anaerobiosis
It can be argued that the efficacy of any isolation medium and procedure for anaerobic bacteria is critically dependent on the measures taken to exclude oxygen. These may cover conditions for sample preparation and dilution, pre-re- duction of diluents and isolation media, and methods used for anaerobic incuba- tion.
There appear to be two schools of thought on the need for stringent anaerobic techniques in the detection of clostridia in foods. The first is supported by the experience of Gibbs and Freame (1965) and leads to the conclusion that no special steps to exclude oxygen are necessary. In comparing different diluents, Gibbs and Freame (1965) found 0.1% peptone entirely suitable for counting purposes when log phase cultures of clostridia were examined. Moreover, there was no significant advantage in boiling the diluent to expel oxygen prior to use. When clostridial cultures were subjected to mechanical blending in the presence of air or shaken with glass beads in medium with a large headspace, no marked loss of viability occurred. This suggests that foods can be macerated in the usual way to obtain good sample dispersion, without any adverse effects on vegetative clostridia. In addition, Gibbs and Freame (1965) saw no necessity to seal the surface of DRCM after inoculation, although the medium was freshly steamed and cooled before being inoculated. Thus, the recommended DRCM technique is very suitable for routine food control laboratories which may not have facilities for anaerobic incubation of media nor the time for tedious manipulations.
The second type of approach to isolating clostridia is that of the anaerobe specialist, such as Barnes (1985), who advocates stringent anaerobic techniques for

32
any food in which anae robes may be present . For clostridia, the use of freshly deoxygena ted diluent, and one that contains an appropr ia te reducing agent, is seen as essential, as are p re - r educed agar plates for isolating strains f rom liquid cultures. This approach is similar to that of A n d e r s o n and Fung (1983) who make the fur ther point that wi thout s t r ingent exclusion of oxygen in isolating clostridia, the re may be significant loss of cells that have been subjected to stress. Sublethal damage of both vegetat ive cells and spores may well arise f rom various food
processing and handl ing t rea tments . Maximal recovery of damaged organisms will requi re not only pro tec t ion f rom oxygen but the use of special resusci ta t ion procedures , as yet u n d e t e r m i n e d for most clostridia or types of damage . In
practice, all me thods of isolating microorganisms represen t some compromise , regardless of the organisms being sought. The diverse proper t ies of clostridia, even those associated with foods, renders such a compromise inevitable if isolation me thods for clostridia as a whole are to be used at all in the food industry. Existing methods , as descr ibed in the p resen t paper , would appea r to provide a reasonab le compromise for most practical purposes . The paucity of new deve lopments in this a rea under l ine the inheren t difficulty in formulat ing isolation media for such a var ied group of organisms.
References
Anderson, K.L. and Fung, D.Y.C. (1983) Anaerobic methods, techniques and principles for food bacteriology: a review. J. Food Protect. 46, 811-822.
Angelotti, R., Hall, H.E., Foter, M.J. and Lewis, K.H. (1962) Quantitation of Clostridium perfringens in foods. Appl. Microbiol. 10, 193-199.
Attenborough, S.J. and Scarr, M.P. (1957) The use of the membrane filter in techniques for control of thermophilic spores in the sugar industry. J. Appl. Bacteriol. 20, 460-466.
Barnes, E.M. (1985) Isolation methods for anaerobes in foods. Int. J. Food, Microbiol. 2, 81-87. Barnes, E.M., Despaul, J.E. and Ingram, M. (1963) The behaviour of a food poisoning strain of
Clostridium welchii in beef. J. Appl. Bacteriol. 26, 415-427. Bhadsavle, C.H., Shehata, T.E. and Collins, E.B. (1972) Isolation and identification of psychophilic
species of Clostridium from milk. Appl. Microbiol. 24, 699-702. Brocklehurst, T.F. and Lund, B.M. (1982) Isolation and properties of psychrotrophic and psychrophilic
pectolytic strains of Clostridium. J. Appl. Bacteriol. 53, 355-361. Dainty, R.H., Edwards, R.A. and Hibbard, C.M. (1989) Spoilage of vacuum-packed beef by a
Clostridium sp. J. Sci. Food Agric. 49, 473-486. Fryer, T.F. and H alligan, A.C. (1976) The development of a method for detecting spores of Clostridium
tyrobutyricum in milk. N.Z.J. Dairy Sci. Technol. 11, 100-106. Gibbs, B.M. and Freame, B. (1965) Methods for the recovery of clostridia from foods. J. Appl.
Bacteriol. 28, 95-111. Gibbs, P.A. (1971) The incidence of clostridia in poultry carcasses and poultry processing plants. Br.
Poult. Sci. 12, 101-110. Hirsch, A. and Grinsted, E. (1954) Methods for the growth and enumeration of anaerobic spore-formers
from cheese, with observations on the effect of nisin. J. Dairy Res. 21, 101-110. Hobbs, G., Crowther, J.S., Neaves, P., Gibbs, P.A. and Jarvis, B. (1982) Detection and isolation of
Clostridium botulinum, In: Corry, J.E.L., Roberts, D. and Skinner, F.A. (Eds.). Isolation and identification methods for food poisoning organisms, Academic Press, London, pp. 151-164.
Holt, J.G., Sneath, P.H.A., Mair, N.S. and Sharpe, M.E. (1986) Bergey's Manual of Systematic Bacteriology, Vol. 2, Williams and Wilkins, Baltimore, pp. 1141-1200.

33
Mead, G.C. (1985) Selective and differential media for Clostridium perfringens. Int. J. Food Microbiol. 2, 89-98.
Mossel, D.A.A. (1989) Adequate protection of the public against food transmitted diseases of microbial aetiology. Achievements and challenges, half a century after the introduction of the Prescott- Meyer-Wilson Strategy. Int. J. Food Microbiol. 9, 271-294.
Mossel, D.A.A. and de Waart, J. (1968) The enumeration of clostridia in foods and feeds. Annal. Inst. Pasteur (Lille) 19, 13-27.
Senyk, G.F., Scheib, J.A., Brown, J.M. and Ledford, R.A. (1989) Evaluation of methods for determina- tion of spore-formers responsible for the late gas-blowing defect in cheese. J. Dairy Sci. 72, 360-366.
Weenk, G., Fitzmaurice, E. and Mossel, D.A.A. (1991) Selective enumeration of spores of Clostridium species in dried foods. J. Appl. Bacteriol. 70, 135-143.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
35
Chapter 3
Media for the detection and enumeration of Bacil lus cereus in foods
P e t e r v a n N e t t e n a a n d J o h n M . K r a m e r b
a Q Laboratories Ltd, Quayside, Navigation Way, Preston PR2 2 YP, UK h PHLS Food Hygiene Laboratory, Central Public Health Laboratory, London, UK.
Bacillus cereus is an established cause of food poisoning in addition to being a troublesome and persistent contaminant, responsible for a variety of spoilage defects in processed foods and dairy products. A range of diagnostic and selective media has been developed to facilitate the detection and enumeration of B. cereus in routine surveillance situations and food poisoning investigations. These media are reviewed with respect to the selective and diagnostic systems they employ, their ability to recover and differentiate the target organism, and their advantages and limitations in particular applications.
Introduction: the rationale for screening foods for B. cereus
Baci l lus cereus is now recognized throughout the world as a potential cause of food poisoning, and the majority of food microbiology laboratories include a regimen for the detection of this organism among their routine surveillance procedures. Before its pathogenicity was fully appreciated, B. cereus was well known as the cause of a common spoilage defect in milk referred to as 'broken' or 'bitty' cream (Stone and Rowlands, 1952).
B. cereus food poisoning arises from the ingestion of foods containing > 105 c fu /g of the organism, and two forms of the illness have been described: the 'diarrhoeal-syndrome' type, characterized by abdominal pain and diarrhoea of 8-16 h onset, and the acute-onset (1-5 h) 'emet ic-syndrome' type, in which the principal symptoms are nausea and vomiting (Kramer and Gilbert 1989). Both forms of illness are toxin-mediated. Foods that are common vehicles for B. cereus
food poisoning and those that are susceptible to B. cereus spoilage, share a number of intrinsic properties that favour growth of the organism and production of certain toxins and enzymes. These include a pH within the range of 5-9, a water

36
activity (a w) greater than 0.94 and, for mesophilic strains, a storage temperature between 10 and 45~
Two factors in the food production and distribution chain contribute most frequently to B. cereus food poisoning. Firstly, the temperature abuse of foods which are prepared several hours in advance of serving and held without adequate refrigeration, and secondly, the recontamination of processed foods either by the environment or by additional food ingredients (Kramer and Gilbert, 1989). Food additives, such as herbs, spices, seasonings and flavourings and pasteurized milk, have all been identified as a source of B. cereus (Mossel et al., 1967; van Netten et al., 1990). Foods subject to inadequate heat processing and post-process recontam- ination can become hazardous following extended storage at unsatisfactory tem- peratures, conditions which allow B. cereus to germinate and multiply. Moreover, psychrotrophic enterotoxigenic strains have been identified which may render food unsafe when held under refrigerated storage at temperatures of > 5~ (van Netten et al., 1990).
Although immunological and cell culture techniques have been developed to enable the direct detection of B. cereus diarrhoeal and emetic toxins in foods (Kramer and Gilbert, 1989), it is likely that most laboratories will continue to rely on conventional selective and diagnostic culture media for the routine isolation and enumeration of B. cereus in the foreseeable future.
B. cereus has a ubiquitous distribution. Thus surveillance procedures must be aimed at monitoring and controlling not only contamination of the end product, but also the ingredients and the process environment. However, this approach complicates the choice of isolation media to the extent that different selective systems are required to suppress the microbial populations characteristic of differ- ent foods and sampling sites.
Table 1 lists those foods that are most commonly contaminated with B. cereus
alongside related food products that have been implicated in B. cereus food poisoning. Apart from fresh vegetables, the microbial flora of these foods consists mainly of Gram-positive bacteria, such as Baci l lus species, staphylococci, micro- cocci, coryneform bacteria, enterococci, streptococci and lactobacilli. Gram-nega- tive organisms occur in addition in vegetables, spices and recontaminated cooked foods (Mossel et al., 1984). An effective B. cereus isolation medium must suppress competitive micro-organisms to the extent that the diagnostic system allows easy and reliable detection of typical strains of B. cereus.
Culture media available for the isolation and enumeration of B. cereus
The quantitative estimation of Baci l lus cereus in food and environmental samples is usually achieved by a standard plate counting method. However, a most probable number (MPN) technique employing trypticase soy-polymyxin B (100 I.U./ml) broth has been recommended for estimating B. cereus in (i) foods that are expected to contain < 10 cfu/g, or (ii) dehydrated starchy foods for which the plate count is inappropriate (Lancette and Harmon, 1980).

37
TABLE 1
Prevalence of Baci l lus cereus in routine food samples and food implicated in food poisoning
Foods commonly contaminated with Baci l lus cereus
Foods implicated in B. cereus food poisoning
Cereals and cereal products
Meat and meat products
Poultry and poultry products
Fish and fish products
Milk and dairy products
Egg and egg products
Vegetables and vegetable products
Desserts, pastries
Herbs, spices, seasonings
Fried rice Cooked rice dishes Cooked pasta dishes (including noodles, macaroni)
Meat loaf, forcemeat Boiled sausage Beef stew, veal broth
Turkey loaf, roast turkey Barbecued chicken
Boiled cod Lobster p~t6
Pasteurised milk, cream Infant formulae
Omelette
Alfalfa sprouts Vegetable stew Mashed potatoes Pea soup
Vanilla slices Vanilla pudding, sauce
Included in many food products associated with food poisoning
Modified from Kramer and Gilbert (1989).
A number of selective and non-selective agar media have been described for the isolation and enumeration of B. cereus in foods; these are listed Table 2. Tables 3 and 4 give details of the diagnostic systems and colony appearance on the media and Table 4 gives the recommended methods of incubation.
In the earliest studies on B. cereus food poisoning, Hauge (1955) used direct surface inoculation onto blood agar (BA). Gilbert and Taylor (1976) also recom- mended the use of horse blood agar for the enumeration of B. cereus in foods implicated in food poisoning, and the Nordic Committee on Food Analysis (Anon., 1982) recommended bovine blood agar for enumerating B. cereus in all types of food. This approach relies solely on the recognition of the characteristic colonial morphology of the organism on blood agar. However, this medium may conve- niently be rendered selective, and Kramer et al. (1982) described a method which involved surface spreading 0.1 ml polymyxin B (5000 I .U./ml) onto 20 ml Columbia base horse blood agar plates (CBA-P)when required. The polymyxin B solution is stable for 2 weeks at 4~
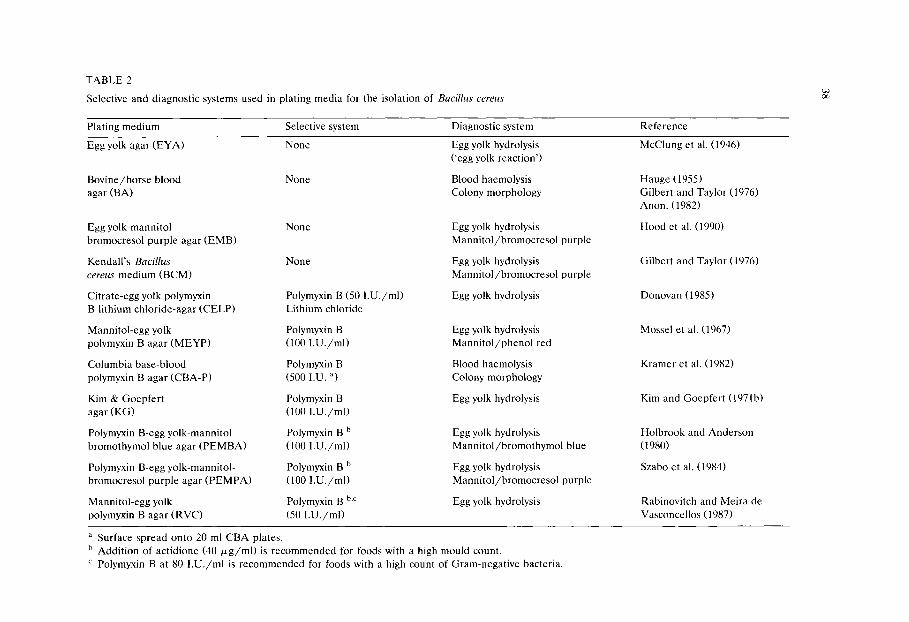
TABLE 2
Selective and diagnostic systems used in plating media for the isolation of Bacillus cereus
Plating medium Selective system Diagnostic system Reference
Egg yolk agar (EYA) None Egg yolk hydrolysis McClung et al. (1946) ('egg yolk reaction')
Bovine/horse blood None Blood haemolysis Hauge (1955) agar (BA) Colony morphology Gilbert and Taylor (1976)
Anon. (1982)
Egg yolk-mannitol bromocresol purple agar (EMB)
Kendall's Bacil lus
cereus medium (BCM)
Citrate-egg yolk polymyxin B-lithium chloride-agar (CELP)
Mannitol-egg yolk polymyxin B agar (MEYP)
Columbia base-blood polymyxin B agar (CBA-P)
Kim & Goepfert agar (KG)
Polymyxin B-egg yolk-mannitol bromothymol blue agar (PEMBA)
Polymyxin B-egg yolk-mannitol- bromocresol purple agar (PEMPA)
Mannitol-egg yolk polymyxin B agar (RVC)
None
None
Polymyxin B (50 I.U./ml) Lithium chloride
Polymyxin B (100 I.U./ml)
Polymyxin B (500 I.U. a)
Polymyxin B (100 I.U./ml)
Polymyxin B b (100 I.U./ml)
Polymyxin B b (100 I.U./ml)
Polymyxin B b,c (50 I.U./ml)
Egg yolk hydrolysis Mannitol/bromocresol purple
Egg yolk hydrolysis Mannitol/bromocresol purple
Egg yolk hydrolysis
Egg yolk hydrolysis Mannitol/phenol red
Blood haemolysis Colony morphology
Egg yolk hydrolysis
Egg yolk hydrolysis Mannitol/bromothymol blue
Egg yolk hydrolysis Mannitol/bromocresol purple
Egg yolk hydrolysis
Hood et al. (1990)
Gilbert and Taylor (1976)
Donovan (1985)
Mossel et al. (1967)
Kramer et al. (1982)
Kim and Goepfert (1971b)
Holbrook and Anderson (1980)
Szabo et al. (1984)
Rabinovitch and Meira de Vasconcellos (1987)
a Surface spread onto 20 ml CBA plates. b Addition of actidione (40 ~g/ml) is recommended for foods with a high mould count. c Polymyxin B at 80 I.U./ml is recommended for foods with a high count of Gram-negative bacteria.

39
Citrate-egg yolk-lithium chloride-polymyxin B (CELP) agar was the first selec- tive agar developed for the enumeration of B. cereus endospores and vegetative cells in milk and on dairy equipment (Donovan, 1958). The selective system consisted of polymyxin B (50 I.U./ml) and lithium chloride (0.5% w/v). The egg yolk precipitation test of McClung et al. (1946) was used for the presumptive identification of B. cereus and citrate was incorporated to enhance the egg yolk reaction. The recommended pH of CELP plates is 7.0 at which the medium has an opaque yellow appearance. The prepared medium should be stored refrigerated (Donovan 1958).
In mannitol-egg yolk-polymyxin B (MEYP) agar the single egg-yolk diagnostic system of McClung et al. (1946) was expanded to a double diagnostic system by the addition of a mannitol/phenol red combination. The selective system of Donovan's medium was modified by omitting lithium chloride and increasing the polymyxin B concentration to 100 I.U./ml (Mossel et al., 1967). The diagnos- tic and selective concept of MEYP has been adopted in all later formulae for B. cereus isolation media (Table 2). The appearance of MEYP agar is opaque light pink at pH 7.2. MEYP plates have a shelf-life of 4 days at 4~
Kim and Goepfert (1971a) observed that the mannitol/phenol red diagnostic system used in MEYP, after incubation at 32~ for 40 h, was not helpful in differentiating B. cereus from other organisms found in dried foods. Furthermore, the use of a fluorescein-labelled anti-exosporium antibody technique developed by this group as a rapid and less laborious serological confirmation of B. cereus, re- quired a culture medium that promoted free endospore production within 24 h, for which MEYP was found to be too nutrient-rich. Therefore, a new basal medium (KG) was formulated (Kim and Goepfert, 1971b) containing low levels of pep- tone (0.1% w/v) and no carbohydrate. The new formula included polymyxin B (100 I.U./ml) as the selective agent and egg yolk emulsion for differentiation, as in MEYP. Phenol red at 0.0025% w/v was added to facilitate the reading of the egg yolk reaction. At the recommended pH of 6.8, the appearance of KG agar is an opaque light pink. Plates can be stored for up to 7 days at 4~
Kendall's mannitol-egg yolk-bromocresol purple agar (BCM) was the first nutri- ent-limited agar medium to use an indicator other than phenol red. This medium was developed primarily to assist with the differentiation of B. cereus from other Baci l lus species, thus the formula does not include selective agents (Gilbert and Taylor, 1976). At the recommended pH of 7.0, BCM is an opaque purple in appearance. Plates have a shelf-life of approximately 7 days when stored at 4~
Poor differentiation of mannitol-utilizing organisms on MEYP after incubation at 32~ for 40 h, weak egg yolk reactions and poor differentiation of B. cereus
colonies on KG agar were the reasons for Holbrook and Anderson's (1980) de- velopment of a new B. cereus selective medium, polymyxin B-egg yolk-mannitol- bromothymol blue agar (PEMBA). The polymyxin and egg yolk concentrations used in PEMBA were the same as those in MEYP, but the double diagnos- tic system of MEYP was modified by replacing phenol red with bromothymol blue. The basal medium was also reformulated using (i) 0.1% w/v peptone, as in KG agar, (ii) 1% w/v sodium pyruvate, to improve the egg yolk reaction,

40
facilitate spore formation and reduce the spreading growth of some B. cereus
colonies and (iii) phosphates, to increase the buffeting capacity of the medium. The new formula was claimed to offer enhanced differentiation of B. cereus
from both mannitol-utilizing and non-mannitol-utilizing organisms. The addition of actidione (40 ktg/ml) was recommended for foods suspected of being contaminated with high levels of mould. PEMBA medium has a semi-opaque yellow-green appearance at pH 7.4. Plates stored at 4~ have a shelf-life of 4 days.
The not infrequent occurrence of atypical B. cereus colonies and inconsis- tent egg yolk reactions on PEMBA led Szabo and co-workers (1984) to de- velop polymyxin B-egg yolk-mannitol-bromocresol purple agar (PEMPA). The replacement of bromothymol blue by bromocresol purple, as in BCM, and a reduction in the incubation period to 18-24 h, was claimed to provide a more reliable diagnostic colony appearance and a consistent egg yolk reaction. At the recommended pH of 6.9, PEMPA medium is a semi-opaque purple in appearance. Information on appropriate storage conditions and the shelf-life of the medium was not provided in the original description, but it would be reasonable to assume that they are similar to those of MEYP and PEMBA.
A methylene blue-Mg2+/Zn2+-glucose-polymyxin agar (RVC) was developed by Rabinovitch and Meira de Vasconcellos (1987) as a less expensive alternative to MEYP and KG. The polymyxin B concentration in RVC agar is either 50 or 80 I.U./ml depending on the application, which is less than in other B. cereus selective media. In foods colonized with large numbers of Gram-negative bacteria, the higher concentration of polymyxin B is recommended. Moulds can be sup- pressed by the addition of actidione (40 ktg/ml). Polymyxin B may be replaced by polymyxin E (colistin) (Rabinovitch and Meira de Vasconcellos, 1987). The nutrient-limited composition of RVC basal medium promotes sporogenesis within 30-40 h. B. cereus can then be differentiated by microscopy from B. thuringiensis on the basis of the presence or absence of parasporal toxin crystals. Zn 2§ ions were incorporated in the basal medium to enhance the egg yolk reaction, medium has a sky-blue appearance at pH 7.3-7.4. Stored at 4~ under vacuum in polyethylene bags, RVC plates have a shelf-life of 3 weeks.
For the enumeration of both B. cereus and Clostridium per fr ingens in foods associated with food poisoning, the use of a non-selective blood-free egg yolk agar (BCP) containing inositol, mannitol and bromocresol purple was recently described (Hood et al., 1990). A similar technique that can be applied to routine food surveillance (D.A.A. Mossel and C.B. Struijk, personal communication) is the use of MEYP medium overlayered with tryptose-sulphite-cycloserine agar (Weenk et al., 1991).
Mode of action of the selective systems used in B. c e r e u s isolation media
The range of selective agents employed in B. cereus isolation media is limited to polymyxin B or E, lithium chloride and actidione (Table 2). The effects of sodium

41
chloride (> 5% w/v) and ethanol (8% v/v) have been studied, but these agents were found to inhibit B. cereus, albeit inconsistently (Mossel et al., 1967).
Polymyxin B is used in all the B. cereus selective agars. Polymyxins are bactericidal towards Gram-negative bacteria and interfere with the structure and function of their outer and cytoplasmic membranes. Polymyxin B suppresses the growth of Acinetobacter spp., Escherichia coli, Enterobacter spp., Klebsiella spp., Pseudomonas aeruginosa, Salmonella spp. and Shigella spp., but resistant strains of Pseudomonas spp., Proteus spp., (for example mannitol-negative, egg yolk-positive Proteus vulgaris) and mannitol-negative, egg yolk-positive Serratia marcescens have been reported. Polymyxin E (colistin) has an antibacterial spectrum similar to polymyxin B. Facultative and obligate aerobic Gram-positive bacteria, such as the mannitol-negative, egg yolk-positive S. aureus, and yeasts and moulds that are resist- ant to polymyxin B or E are also able to form colonies on B. cereus selective media.
The use of a lithium chloride (0.5% w/v)/polymyxin B combination has been described for the inhibition of lactic acid bacteria (MacLeod, 1954). However, certain strains of B. cereus have been identified as being susceptible to this selective system (Donovan, 1958; D.A.A. Mossel, personal communication). The addition of actidione to polymyxin-based agars such as PEMBA and RVC is recommended for the suppression of mould growth (Holbrook and Anderson, 1980; Rabinovitch and Meira de Vasconcellos, 1987).
Another factor that can contribute to selectivity is the composition of the basal medium itself. The nutritional conditions imposed by KG agar, for example, inhibit the lecithinase activity of B. polymyxa strains. These strains can form weakly egg yolk reactive colonies on MEYP.
Holbrook and Anderson (1980) noted that Bacillus species other than B. cereus were able to grow on PEMBA, including B. thuringiensis, B. cereus var. mycoides, B. licheniformis, B. subtilis, B. polymyxa, B. aleei and B. brevis. A similar observa- tion was reported by Szabo et al. (1984) for PEMPA. RVC medium prepared with 50 I.U./ml colistin suppressed the growth of Klebsiella pneumoniae and Pseu- domonas aeruginosa, but the concentration had to be increased to 80 I.U./ml in order to inhibit the growth of Salmonella and Arizona species. (Rabinovitch and Meira de Vasconcellos, 1987).
Mode of action of the diagnostic systems used in B. cereus isolation media
The principal diagnostic features of B. cereus exploited in the various B. cereus selective media are (i) the characteristic colonial appearance of the organism, (ii) the production of a lecithinase, responsible for the so-called egg yolk reaction and (iii) the inability of the organism to utilize mannitol (Tables 3 and 4). A second- ary diagnostic feature, the morphology of the endospore and sporangium, is also exploited by the nutrient-limited culture media: KG, BCM, PEMBA, PEMPA and RVC.
The characteristic egg yolk reaction of B. cereus involves an enzyme-mediated hydrolysis of lecithin or phosphatidylcholine into phosphoryl-choline and diglyc-

42
TABLE III
Diagnostic characteristics of Bacillus cereus isolation media
Diagnostic system Diagnostic characteristic
Bovine or horse blood
Egg yolk emulsion
Mannitol + phenol red
Mannitol + bromothymol blue
Mannitol + bromocresol purple
Nutrient limited basal medium
Haemolysis (a or/3)
Egg yolk precipitation (enhanced by Zn 2 + )
pH reduction associated with mannitol utilization causes pink ~ yellow colour change
pH reduction associated with mannitol utilization causes blue ~ green colour change
pH reduction associated with mannitol utilization causes purple ~ green colour change
Enhanced sporulation enables differentiation according to spore/sporangium morphology
eride. This results in the formation of a zone of precipitation surrounding the colonies of lecithinase-positive organisms. Typical strains of B. cereus produce a zone 3-5 mm wide on an optimally functioning selective agar (Baird et al., 1987). Other lecithinase-producing Bacillus species, aside from B. cereus, include B. anthracis, B. cereus var. mycoides, B. thuringiensis and B.laterosporus (Table 5).
Members of other bacterial genera that produce a lecithinase and are able to grow on B. cereus selective media, for example Staphylococcus aureus, Serratia marcescens and Proteus vulgaris, produce an egg yolk clearing reaction that is distinguishable from the egg yolk precipitation reaction associated with B. cereus.
The relative size of the egg yolk reaction zone is an important factor in the performance of B. cereus diagnostic and selective agars. Zones that are too large can coalesce and make the estimation of the number of presumptive B. cereus
colonies difficult (Harmon et al., 1984). This problem can be overcome either by using 14 cm diameter plates, or by reducing the countable colony range from the 30-300 per 9 cm plate normally recommended for quantitative analyses, to less than 30.
Reliance on the egg yolk reaction as the key diagnostic characteristic of B. cereus can result in weak or lecithinase-negative strains of this organism being overlooked. According to Holbrook and Anderson (1980), this is most likely to occur with KG agar and EYA and, to a lesser extent, with MEYP and PEMBA. Strains of B. cereus giving a weak egg yolk reaction have been implicated in outbreaks of food poisoning, and this was the principal reason for Gilbert and Taylor's (1976) recommendation that blood agar be used for primary isolation.

TABLE 4
Appearance of Baci l lus cereus colonies on various isolation media
43
Plating Colony appearance Reference medium
BA Gilbert and Taylor (1976) Anon. (1982) Kramer et al. (1982) Donovan (1958)
Raised grey-green, 3-8 mm matt or 'ground glass' surface. Edge crenate to fimbriate. Sur- rounded by zone of o~- or/3-haemolysis.
CELP Crenate to fimbriate. Surrounded by white zone of egg yolk hydrolysis.
MEYP Crenate to fimbriate, matt surface, 3-6 ram. Mossel et al. (1967) Surrounded by white zone of egg yolk hydrolysis against pink background.
KG Flat to slightly raised, crenate or irregular, dry matt surface, 2-5 ram. Surrounded by white zone of egg yolk hydrolysis against pink back- ground.
BCM Crenate to fimbriate raised colonies, mauve, Gilbert and Taylor (1976) 3-6 mm. Surrounded by violet zone of egg yolk Kramer et al. (1982) hydrolysis against purple background.
PEMBA Crenate to fimbriate raised peacock blue colonies, 3-5 mm. Surrounded by blue zone of egg yolk hydrolysis against yellow-green back- ground. Crenate to fimbriate raised mauve colonies, 2-5 mm. Surrounded by mauve-purple zone of egg yolk hydrolysis against purple background.
RVC Crenate to fimbriate raised colonies surrounded Rabinovitch and Meira de by pale blue zone of egg yolk hydrolysis against Vasconcellos (1987) celestial blue background.
Kim and Goepfert (1971b)
Holbrook and Anderson (1980)
PEMPA Szabo et al. (1984)
a Refer to Table 2 for key to abbreviations.
Such strains can still be recognised by their typical colonial morphology on this medium. Nonetheless, weak or negative lecithinase-producing isolates suspected as being strains of B. cereus require additional biochemical testing to confirm their identities.
Apart from KG and RVC agars, the differential manni tol /pH indicator systems of the other diagnostic/selective plating media give a presumptive identification of weak or negative egg yolk-reactive strains of B. cereus solely on the basis of their mannitol-negative colony appearance. For this system to function correctly, it is important that the pH is controlled both before and after autoclaving of the media. This is essential for the marginally buffered MEYP agar. It is the authors' experience that the usefulness of the manni tol /pH balance decreases with any variation from the recommended pH.
Inoculation and culture conditions
All the diagnostic/selective plating media for B. cereus are surface inoculated. This allows the formation of characteristic colonies of B. cereus that can be easily
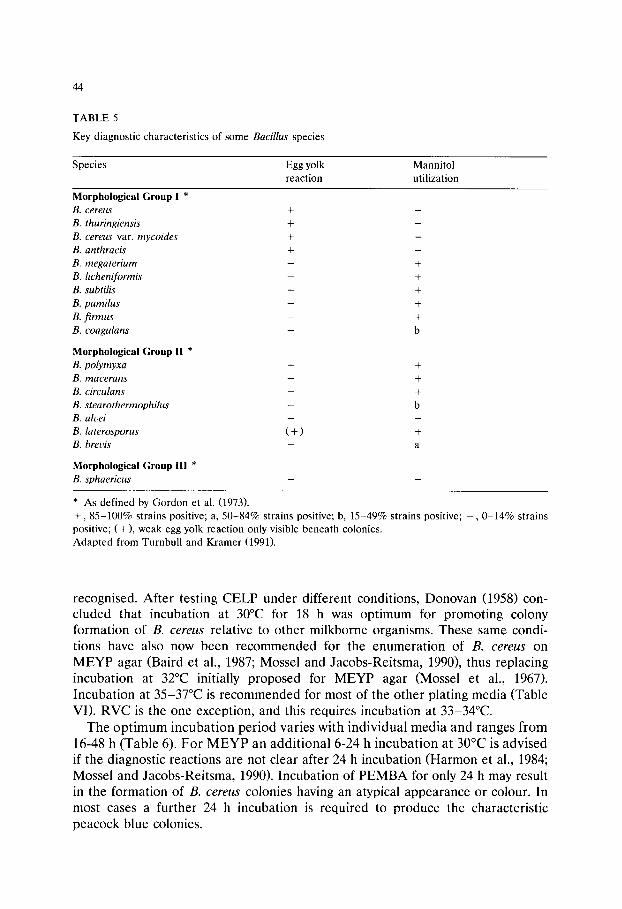
44
TABLE 5
Key diagnostic characteristics of some B a c i l l u s species
Species Egg yolk Mannitol reaction utilization
Morphological Group I * B. c e r e u s + -
B. t h u r i n g i e n s i s + -
B. c e r e u s var. r n y c o i d e s + -
B. a n t h r a c i s + -
B. m e g a t e r i u m - +
B. l i c h e n i f o r m i s - +
B. sub t i l i s - +
B. p u m i l u s - +
B. f i r m u s - +
B. c o a g u l a n s - b
Morphological Group II * B. p o l y m y x a - +
B. m a c e r a n s - +
B. c i r c u l a n s - +
B. s t e a r o t h e r m o p h i l u s - b
B. a l v e i - -
B. l a t e r o s p o r u s ( + ) +
B. b r e v i s - a
Morphological Group III * B. s p h a e r i c u s
* As defined by Gordon et al. (1973). + , 85-100% strains positive; a, 50-84% strains positive; b, 15-49% strains positive; - , 0-14% strains positive; (+) , weak egg yolk reaction only visible beneath colonies. Adapted from Turnbull and Kramer (1991).
recognised. After testing CELP under different conditions, Donovan (1958) con- cluded that incubation at 30~ for 18 h was optimum for promoting colony formation of B. cereus relative to other milkborne organisms. These same condi- tions have also now been recommended for the enumeration of B. cereus on MEYP agar (Baird et al., 1987; Mossel and Jacobs-Reitsma, 1990), thus replacing incubation at 32~ initially proposed for MEYP agar (Mossel et al., 1967). Incubation at 35-37~ is recommended for most of the other plating media (Table VI). RVC is the one exception, and this requires incubation at 33-34~
The opt imum incubation period varies with individual media and ranges from 16-48 h (Table 6). For MEYP an additional 6-24 h incubation at 30~ is advised if the diagnostic reactions are not clear after 24 h incubation (Harmon et al., 1984; Mossel and Jacobs-Reitsma, 1990). Incubation of PEMBA for only 24 h may result in the formation of B. cereus colonies having an atypical appearance or colour. In most cases a further 24 h incubation is required to produce the characteristic peacock blue colonies.

45
Enumeration of Baci l lus cereus in foods
The results of a collaborative study by Lancette and Harmon (1980) showed that the direct plating technique gave better repeatability and reproducibility than the MPN technique at both high and low levels of B. cereus contamination. For direct plating, blood agar is suitable for the isolation of large numbers of B. cereus in foods implicated in outbreaks (Gilbert and Taylor, 1976). However, the selective agars MEYP, PEMBA, KG and PEMBA are preferable for the enumeration of B. cereus in the presence of higher levels of other organisms found in food, environ- mental samples and faecal specimens. Blood agar base, MEYP and PEMBA are commercially available in dehydrated form or as pre-poured plates, and mono- graphs describing these appear in this volume. In North America and other ISO- affiliated (International Standards Organization) countries, MEYP is usually the medium of choice for the enumeration of B. c e r e u s in foods, and it is the medium recommended by the Association of Official Analytical Chemists (Lancette and Harmon, 1980). PEMBA is a widely used alternative, particularly in the UK. 1980). PEMBA is a widely used alternative, particularly in the UK.
Under standardized test conditions using 20 different strains of B. cereus
inoculated into cooked rice, Szabo and co-workers (1984) observed a difference in the recovery performance of selective agars and the non-inhibitory TSA (trypticase soy agar) as follows: TSA -- PEMPA > PEMBA, >> MEYP. This is in contrast to the findings of a similar study carried out in the PHLS Food Hygiene Laboratory ( J . M . Kramer, unpublished data) using 50 strains of different origin, in which no
TABLE 6
Recommended culture conditions for Bacillus cereus isolation media
Plating Incubation Reference medium a Temp. (~ Time (h)
BA 30 20-24 35-37 24-48
CBA-P 36 18-24
BCM 35-37 18
CELP 30 18
MEYP 32 18-40 30 24-30
KG 35-37 16-24
PEMBA 37 24-48
PEMPA 37 18-24
RVC 33-34 24-40
Anon. (1982) Gilbert and Taylor (1976)
Kramer et al. (1982)
Gilbert and Taylor (1976)
Donovan (1958)
Mossel et al. (1967) Baird et al. (1987)
Kim and Goepfert (1971b)
Holbrook an Anderson (1980)
Szabo et al. (1984)
Rabinovitch and Meira de Vasconcellos (1987)
a Refer to Table 2 for key to abbreviations.
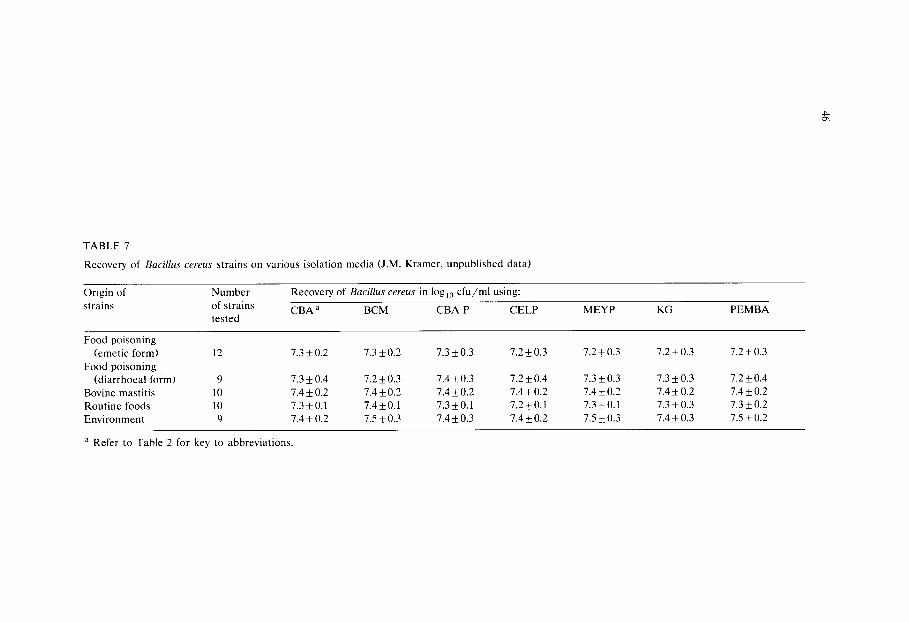
TABLE 7
Recovery of Baci l lus cereus strains on various isolation media (J.M. Kramer, unpublished data)
Origin of Number strains of strains
tested
Recovery of Baci l lus cereus in log10 c fu /ml using:
CBA a BCM CBA-P CELP MEYP KG PEMBA
Food poisoning (emetic form) 12
Food poisoning (diarrhoeal form) 9
Bovine mastitis 10 Routine foods 10 Environment 9
7.3 + 0.2 7.3 _+ 0.2 7.3 _+ 0.3 7.2 +_ 0.3 7.2 + 0.3 7.2 _+ 0.3 7.2 _+ 0.3
7.3 +_ 0.4 7.2 _+ 0.3 7.4 _+ 0.3 7.2 _+ 0.4 7.3 +_ 0.3 7.3 +_ 0.3 7.2 _+ 0.4 7.4 _+ 0.2 7.4 _+ 0.2 7.4 _+ 0.2 7.4 _+ 0.2 7.4 _+ 0.2 7.4 _+ 0.2 7.4 + 0.2 7.3 _+ 0.1 7.4 +_ 0.1 7.3 _+ 0.1 7.2 + 0.1 7.3 _+ 0.1 7.3 _+ 0.3 7.3 _+ 0.2 7.4 + 0.2 7.5 _+ 0.3 7.4 _+ 0.3 7.4 _+ 0.2 7.5 _+ 0.3 7.4 _+ 0.3 7.5 _+ 0.2
a Refer to Table 2 for key to abbreviations.

47
significant differences in recovery were observed between PEMBA, MEYP and other B. c e r e u s selective agars tested (Table 7).
In a comparison of selective media for the quantitative recovery of B. cereus
from naturally contaminated dried foods, Szabo et al. (1984) reported a rank order of: PEMPA > PEMBA > MEYP. The efficacy of PEMPA was attributed to a better suppression of background flora and the smaller colony and egg yolk reaction zone sizes obtained. However, in a similar study using dried foods and pasteurized milk samples containing 103-104 B. cereus per g or per ml, Holbrook and Anderson (1980) found no significant difference in the recovery of B. cereus
by McClung's egg yolk agar, MEYP, KG and PEMBA. Peterz et al., (1985) compared the recovery of B. cereus on BA, PEMBA and MEYP from pasteurized milk and cream, and dried products such as rice, potato powder, beans, peas, cocoa and soup powders, and also found no significant difference in the perfor- mance of these media.
As the selective systems used in these various B. cereus isolation media are similar, conflicting reports of their performance must be attributed either to the preparation of the media, the composition and microflora of the foods tested, or differences in the observation and interpretation of results. Clearly, the examina- tion of foods contaminated with low numbers of B. cereus in the presence of high numbers of mannitol-utilizing organisms would make the differentiation of B. cereus colonies on MEYP, KG, PEMBA and PEMPA difficult, and almost impos- sible on non-selective diagnostic media such as BA or BCM.
Finally it should be noted that although the egg yolk present in B. cereus
selective media confers some protection on sublethally injured strains (Mossel and van Netten, 1984), the use of a resuscitation procedure prior to selective plating may be required in order to achieve optimum recovery of viable but damaged cells of B. cereus.
Conclusion
A number of culture media specifically formulated for the detection and enumeration of B. cereus have been described. All have certain limitations on their use. Examples cited include the problems of detection of atypical strains of B. cereus , and poor suppression of the growth of background organisms. A knowledge of how isolation media function, their advantages and their drawbacks, is of primary importance in bacteriological investigations. It is hoped that the informa- tion contained in this review will assist the microbiologist in the selection of an isolation medium for B. cereus appropriate to the particular circumstances, whether in the context of routine surveillance and quality control or the investigation of specific contamination, spoilage, or public health problems attributed to this organism.

48
References
Anon. (1982) Nordic Committee on Food Analysis, 2nd Edn., Ord and Form AB, Uppsala. Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (Eds.) (1987) Pharmacopoeia of culture media for food
microbiology. Int. J. Food Microbiol. 5, 187-300. Donovan, K.O. (1958) A selective medium for Bacillus cereus in milk. J. Appl. Bacteriol. 21, 100-103. Gilbert, R.J. and Taylor, A.J. (1976) Bacillus cereus food poisoning. In: F.A. Skinner and J.G. Carr
(Eds.), Microbiology in Agriculture, Fisheries and Food, Society for Applied Bacteriology Sympo- sium Series No.4, Academic Press, London. pp. 197-213.
Gordon, R.E., Haynes, W.C. and Pang, C.H.N. (1973) The genus Bacillus. In: Agriculture Handbook No. 427, United States Department of Agriculture, Washington, DC. pp. 98-99.
Harmon, S.M., Kautter, D.A. and McClure, F.D. (1984) Comparison of selective plating media for enumeration of Bacillus cereus in foods. J. Food Protect. 47, 65-67.
Hauge, S. (1955) Food poisoning caused by aerobic spore-forming bacilli. J. Appl. Bacteriol. 18, 591-595.
Holbrook, R. and Anderson, J.M. (1980) An improved selective and diagnostic medium for the isolation and enumeration of Bacillus cereus in foods. Can. J. Microbiol. 26, 753-759.
Hood, A.M., Tuck, A. and Dane, C.R. (1990) A medium for the isolation, enumeration and rapid presumptive identification of injured Clostridium perfringens and Bacillus cereus. J. Appl. Bacteriol. 69, 359-372.
Kim, H.U. and Goepfert, J.M. (1971a) Occurrence of Bacillus cereus in selected dry food products. J. Milk Food Technol. 34, 12-15.
Kim, H.U. and Goepfert, J.M. (1971b) Enumeration and identification of Bacillus cereus in foods. Appl. Microbiol. 22, 581-587.
Kramer, J.M., Turnbull, P.C.B., Munshi, G. and Gilbert, R.J. (1982) Identification and characterisation of Bacillus cereus and other Bacillus species associated with foods and food poisoning. In: J.E.L. Corry, D. Roberts and F.A. Skinner (Eds.) Isolation and Identification Methods for Food Poisoning Organisms, Academic Press, London. pp. 261-286.
Kramer, J.M. and Gilbert R.J. (1989) Bacillus cereus and other Bacillus species. In: M.P. Doyle (Ed.), Foodborne bacterial pathogens, Marcel Dekker Inc., New York. pp. 21-70.
Lancette, G.A. and Harmon, S.M. (1980) Enumeration and confirmation of Bacillus cereus in foods: collaborative study. J. Assoc. Offic. Anal. Chem. 63, 581-586.
MacLeod, R.A. (1954) Dependence of the toxicity of cations for lactic acid bacteria on pH and incubation time. J. Bacteriol. 67, 23-26.
McClung, L.S., Heidenreich, P. and Toabe, R. (1946) A medium for the Nagler plate reaction for the identification of certain Clostridia. J. Bacteriol. 51, 751-752.
Mossel, D . A . A . , Koopman, M.J. and Jongerius, E. (1967) Enumeration of Bacillus cereus in foods. Appl. Microbiol. 15, 650-653.
Mossel, D . A . A . van der Zee, H., Corry, J.E.L. and van Netten, P. (1984) Microbiological quality control. In: S.M. Herschdorfer (Ed.), Quality control in the food industry, Vol. I, Academic Press, London, pp. 79-168.
Mossel, D.A.A. and Jacobs-Reitsma, W.F. (1990) Mikrobiologisch onderzoek van Levensmiddelen. Strategie, principes en methoden. P.C. Noordervliet B.V., Zeist.
Mossel, D.A.A. and van Netten, P. (1984) Harmful effects of selective media on stressed micro- organisms: nature and remedies. In: H.E. Andrew and A.D. Russell (Eds.) The revival of injured microbes, Academic Press, London. pp. 329-371.
Peterz, M., Wiberg, C. and Norberg, P. (1985) Comparison of media for the isolation of Bacillus cereus
from foods. J. Food Protect. 48, 969-970. Rabinovitch, L. and Meira de Vasconcellos, F.J. (1987) A new medium for the detection and
enumeration of Bacillus cereus in food. Rev. Microbiol., Sao Paulo. 18, 330-334. Stone, M.J. and Rowlands, A. (1952)"Broken" or "bitty" cream in raw and pasteurized milk. J. Dairy
Res. 19:51-62. Szabo, R.A., Todd, E.C.D and Rayman, M.K. (1984) Twenty-four hour isolation and confirmation of
Bacillus cereus in foods. J. Food Protect. 47, 856-860.

49
Turnbull, P.C.B. and Kramer, J.M. (1991) The genus Bacillus. In: A. Balows, W.J. Hausler, K.L. Herrmann, H.D. Isenberg and H.J. Shadomy (Eds.), Manual of Clinical Microbiology (5th Edn.), American Society for Microbiology, Washington, DC. pp. 296-303.
van Netten, P., van Moosdijk, A.M. and Mossel, D.A.A. (1990) Psychrotrophic strains of Bacillus cereus
producing enterotoxin. J. Appl. Bacteriol. 69, 73-79. Weenk, G., Fitzmaurice, E. and Mossel, D.A.A. (1991) Selective enumeration of spores of Clostridium
species in dried foods. J. Appl. Bacteriol. 70, 135-143.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
51
Chapter 4
Culture media for enterococci and group D-streptococci
G. Reuter Institute for Food Hygiene, Meat Hygiene and Technology, Veterinary Department,
Free University of Berlin, Brummerstrasse 10, D-IO00 Berlin, Germany
Lancefield group D-streptococci are contaminants of various food commodities, especially those of animal origin. They encompass the new genus Enterococcus comprising 13 known species and some species of streptococci which have their habitat in the intestine of animals, e.g. Streptococcus bouis, suis and equinus. The serologically based grouping may no longer constitute the best definition for streptococci from the food chain. Food hygiene monitoring systems using enterococci as indicators need reliable methods for selective cultivation and identification of marker strains. Up to now more than 100 modifications of selective media have been described for isolating streptococci or enterococci from various specimens. The selection of a medium requires either experience or consultation. It depends on the kind of specimen, the method of cultivation (plate count or membrane filter) and whether or not the habitat is heavily contaminated with other organisms. The choice of media is made more difficult as commercial versions of the same culture medium may vary in recipe a n d / o r performance from producer to producer. Therefore, reviewing the literature may help in the choice of medium and confirmation tests.
The selectivity and productivity of some commonly used or cited media are reported here, partly based on our own experience: citrate azide tween carbonate agar (CATC), kanamycin aesculin azide agar (KAA) and M-enterococcus agar (ME) including earlier results with aesculin bile azide agar (ABA), and thallous acetate tetrazolium glucose agar (T1TG). No medium was completely selective for all group D-streptococci or for all enterococci but some media were highly selective for a single Enterococcus species, e.g., for E. faecalis which serves as indicator of human pollution.
Confirmatory tests must be carried out when experience in the evaluation procedure is limited. Selective media for enterococci should be used only after or while checking in parallel their selectivity and productivity against appropriate test organisms.
Introduction
Enterococci may be considered an essential part of the autochthonous mi- croflora of humans and animals. Some host specificity exists, in humans Enterococ- cus faecalis and E. faecium are the most frequent species, while E. faecium is the predominant species in poultry and pigs. E. faecalis and faecium as well as E.

52
mundtii and casseliflavus can also be found in plants. This shows that there are some species which have their habitats in plants and insects. The ecology of enterococci needs further study especially on the basis of the new classification of the genus (Mundt, 1984).
With this wide distribution it is not surprising that enterococci occur in different food commodities, especially those of animal origin. Presence of E. faecalis and faecium is often used to indicate faecal contamination of food. That means that these species must be cultivated separately from the total count of a food specimen.
E. faecalis ssp. faecalis may be regarded as an indicator of contamination from human sources, whereas E. faecium and other species may indicate contamination from other sources.
Streptococcus boris of the group D-streptococci, which occurs in animal faeces, is not generally suitable as a hygienic indicator as it cannot survive very well in the environment. S. equinus has so far not been isolated from humans and is probably of no interest for food hygiene. Therefore, in the field of food microbiology culture media for representatives of the new genus Enterococcus especially those for E. faecalis and its subspecies are most often needed.
A wide variety of selective media for the enterococcus and /o r streptococcus group has been recommended and used in the past. More than 100 modifications have been described in the literature, summarized by Barnes (1976). Proposals for specific uses were made in the proceedings of a previous symposium (Reuter, 1985b). Some newer review data were given in the recent monograph of Devriese et al. (1991), but not all information related to food was included there. Taxonomic definitions must first be established to avoid confusion when using selective media or in interpreting results of counts of enterococci or streptococci. The last edition of Bergey's Manual (Mundt, 1984) does not contain all the new names of species which are accepted now (Devriese et al., 1991; Pompei et al., 1992).
Definitions to be considered
In the past, the ecological grouping seemed to be a useful means of separation, e.g., into the 'faecal', 'oral' and 'lactic' steptococci. However, some overlapping existed. Both oral and lactic streptococci could occasionally be isolated from human faecal samples, sometimes even in large numbers (Barnes, 1976).
Serological grouping was another well established way to divide the strepto- cocci. There seemed to be certain correlations between ecological and serological grouping, for example, the faecal streptococci commonly corresponded to the group D-streptococci, the mesophilic lactic streptococci to the group N. The oral streptococci, however, could not be defined serologically as a single group.
The new taxonomic separation into the genera 'Enterococcus' and 'Lactococcus' enables better identification of questionable isolates of streptococci: the outstand- ing problem is to separate the remaining D-positive strains from the enterococci easily. Streptococcus bocis, S. equinus and S. alactolyticus belong to the Lancefield

TABLE 1
Grouping of ' S t rep tococc i ' relevant to food
53
Serological: Group:
Taxonomical: I Genus : En terococcus
I Streptococcus Lac tococcus S treptococcus
Species: lyogroup I-VII (bacteriolytic activity)
I faecal is
spp. f a e c a l i s
spp. z y m o g e n e s
ssp. l i que fac iens
ma lodora tus
II f a e c i u m
durans
III cassel i f lavus* *)
IV a v i u m * * *)
p s e u d o a v i u m * * * * )
raf f inosus * * * * * ) solitarius
V h i r a e * * * * * * )
VI gal l inarum
VII mund t i i
O c e c o r u m * * * * )
boris (I + II) ( su is ) equinus * ) lactis + )
ssp. lactis
ssp. diacetilactis
ssp. cremoris
garviae + + )
p l a n t a r u m + + +
raff inolactis + + + + )
salivarius
ssp. thermophi lus
*) not isolated from humans **) epiphyte in plants (similar to faecium) * * *) serological group Q besides D * * * *) serological not D *****) less than 50% group D * * * * * *) similar to lyogroup II (durans) +) dairy lactococci ++) bovine sources (mastitis) + + + ) frozen peas + + + +) raw milk.
se ro logica l g r o u p D bu t a re no t en te rococc i . S o m e L e u c o n o s t o c and mos t P e d i o -
c o c c u s s t ra ins also n e e d to be d i f f e r e n t i a t e d f rom en t e rococc i ( F a c k l a m and
Coll ins , 1989). T h e r e f o r e , I have a t t e m p t e d h e r e to i l lus t ra te the g r o u p i n g of the
" s t r e p t o c o c c i in a b r o a d s e n s e " r e l evan t to food, in a c c o r d a n c e wi th the new
t a x o n o m y in T a b l e 1.
T h e c h a r a c t e r i s t i c s u se fu l fo r t he d i f f e r e n t i a t i o n o f t he e n t e r o c o c c i a r e g i v e n
in T a b l e s 2a a n d 2b. T h e d a t a w e r e c o l l e c t e d f r o m e x p e r i e n c e in o u r l a b o r a t o r y
a n d f r o m r e c e n t p a p e r s . T h e tes ts a re p r o p o s e d fo r r o u t i n e l a b o r a t o r y t e s t i n g
o f h u m a n a n d f o o d s p e c i m e n s . T h a t n o t all i so la t e s c o u l d be g r o u p e d m a y be seen

T A B L E 2a
Characteristics useful for differentiat ion of enterococci, according to: Hahn et al. (1970), Collins et al. (1984), Schleifer and Kilpper-B~ilz (1984), Reuter
(1985b), Facklam and Collins (1989), Nusser (1991), Devriese et al. (1991), Pompei et al. (1992)
Enterococcus Serogroup 1 2 3 4 5 6 7 8 9 10 11 12 spec ies / f requency ~ acid from:
subspecies (%) C A T C ~45 10 50 pt. arg. mm. /3-h. gel. mt.
13 14 14 16
lact. sorb. arab yp. mot. E
f aec ium D (68) pink + + ~ - + + - - + / - +
faecalis D (91 ) red + + - + + + - - + + +
ssp. faecalis
faecalis ssp. liquefaciens D(91) red + + - + + + - + + +
faecalis D(91) purple-red + + - + + + [ - ~ - / + - + + ssp. zymogenes I I
durans D (75) pink + + - hirae D (63) bright pink +
avium D (32) + + / - - Q
casseliflavus D (100) + / - + / - - gallinarum D (100) + +
maloduratus D (100) - + - mundt i i D (100) + +
pseudoa eiurn - + +
raffinosus D (40) + + solitarius D (100) + +
- + + ( + ) - _ + - _
- - + + - - + - - _
_ / + . . . . + + + +
- + / - + - _ + + + / - + +
- + + - + + + / - + -
. . . . + + + -- _
- / + + - + + + / - + +
_ _ _ + + - - _
_ _ _ + + + - -
- - + - - + + - - _
- I I
- V
- I V
+ I I I
+ V I
- I
- V I I
- I V
- I V
- I V
cecorum - + . . . . + / - + . . . . 0
% = frequency 1, growth on CATC; 2, growth at 45~ 3, growth at 10~ 4, growth at 50~ 5, growth in potassium tellurite 0.04%; 6, arginine dehydrolase; 7, growth in methylene blue milk; 8, /3-haemolytic; 9, gelatinase; 10, mannitol; 11, lactose; 12, sorbitol; 13, arabinose; 14, yellow pigment; 15, motility; 16,
lyo-group E I - V I I : marker reactions.

TABLE 2b
Lyogroups for separating species of Enterococcus according to Pompei et al. (1992)
55
Bacteriolytic activity a Activity inhibited b
E I strong E II not so strong
E III reduced E IV nearly none
E V strong E VI reduced
E VII reduced and weak
with suramin at pH 9.6
with sodium azide, 4% NaCl, at pH 9.6 with all specific conditions
only weak with suramine with bile salts, 6% NaC1 and suramine
a against Micrococcus luteus-cells. b experimental conditions: 0.02% sodium azide; 0.05% suramin; 0.04% bile salts; 4.0 and 6.0% NaC1;
pH 9.6
f rom a collection of atypical strains f rom C A T C - m e d i u m f rom Nusser (1991) (Table 3).
Requirements and composition of selective media for isolating group D-streptococci or enterococci
Selective agents Various selective agents may be used in these selective media: commonly
sodium azide, thallous acetate, and kanamycin or gentamicin. Other limiting ingredients or factors are crystal violet, Tween 80, carbonate and bile salts. Special growth conditions may be achieved by low pH (e.g., 6.0 or 6.2) or elevated incubation temperature (42 or 45~ Increased or decreased concentrations of selective agents will yield different degrees of selective productivity, therefore their correct use is of great importance. For instance, sodium azide is heat sensitive, and the solution should be added after the medium has been autoclaved. If commer- cially available media already contain this ingredient, its selective effect must be verified by testing the prepared medium. Many misleading results can be caused by neglecting this factor.
Indicators Indicator substances added to the media are useful for the recognition of
enterococci and for the rapid identification of single species on the basis of colony appearance. Tetrazolium, or in special cases tel|urite, reduction is commonly used. Recently the formation of an aesculin-ferrous complex has been applied as a criterion. However, some special facts must be considered. In a medium at pH 7.0, most enterococci will reduce tetrazolium to formazan, which becomes obvious as a bright red coloured colony; at pH 6.2 or even 6.0, this reaction is evident only for the species Enterococcus faecalis, while E. faecium or other streptococci show only
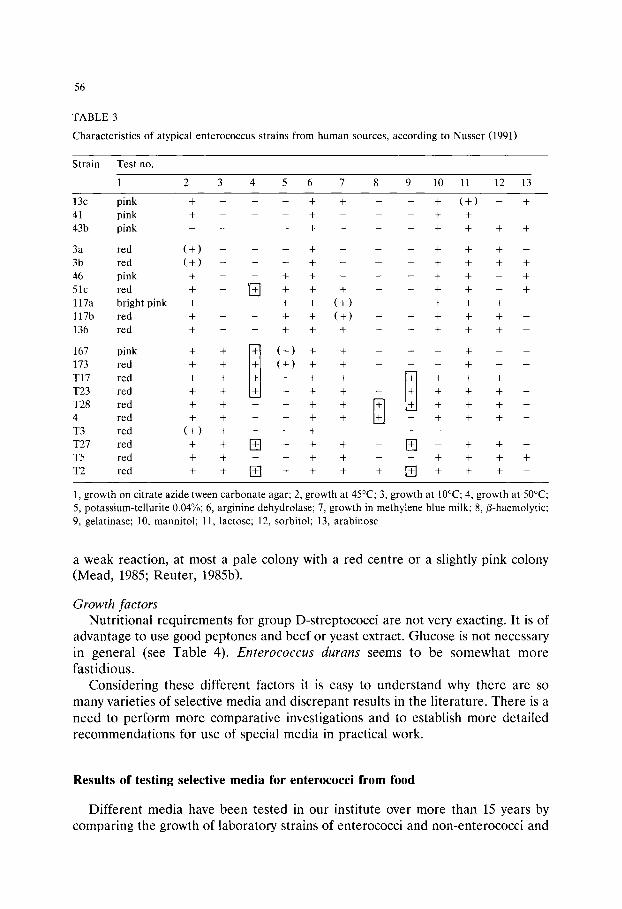
56
T A B L E 3
Characteristics of atypical enterococcus strains from human sources, according to Nusser ( 1 9 9 1 )
Strain Test no.
1 2 3 4 5 6 7 8 9 10 11 12 13
13c p i n k + - - - + + - - + ( + ) - +
41 p i n k + - - - + - - - + +
4 3 b p i n k . . . . + - - - + + + +
3a r e d ( + ) - - - + -
3b r e d ( + ) - - - + -
46 p i n k + - - + + -
51c r e d + - ~ + + +
1 1 7 a bright pink + - - + + ( + )
l 1 7 b r e d + - - + + ( + )
136 r e d + - - + + +
167 p i n k + + [ ' ~ ( + ) + +
173 red + + ~+~ ( + ) + +
T 1 7 r e d + + + + +
T 2 3 red + + + + +
T 2 8 red + + - + + +
4 r e d + + - + + +
T 3 r e d ( + ) + - - + -
T 2 7 red + + E ] + + +
T 5 red + + - + + +
T 2 red + + [+--7 + + +
m m
m J
_ m
m m
m
+ +
+ +
+ +
+ +
+ +
+ +
+ +
m _ m
-- M + + -- [++] + +
-- + +
-- ~ -- + _ _ + +
+ [ ~ + +
+ +
- +
- + + -
+ -
+ -
+
+
+
+
m
+ +
+ -
1, growth on citrate azide tween carbonate agar; 2, growth at 4 5 ~ 3, growth at IO~ 4, growth at 5 0 ~
5, p o t a s s i u m - t e l l u r i t e 0 . 0 4 % ; 6, arginine dehydrolase; 7, growth in methylene blue milk; 8,/3-haemolytic; 9, g e l a t i n a s e ; 10, m a n n i t o l ; 11, l a c t o s e ; 12, s o r b i t o l ; 13, arabinose
a weak reaction, at most a pale colony with a red centre or a slightly pink colony (Mead, 1985; Reuter, 1985b).
Growth factors Nutritional requirements for group D-streptococci are not very exacting. It is of
advantage to use good peptones and beef or yeast extract. Glucose is not necessary in general (see Table 4). Enterococcus durans seems to be somewhat more fastidious.
Considering these different factors it is easy to understand why there are so many varieties of selective media and discrepant results in the literature. There is a need to perform more comparative investigations and to establish more detailed recommendations for use of special media in practical work.
Results of testing selective media for enterococci from food
Different media have been tested in our institute over more than 15 years by comparing the growth of laboratory strains of enterococci and non-enterococci and

TABLE 4
Composition of culture media for enterococci (group D-streptococci)
57
Ingredients T1TG ME CATC g or ml/1
A B A K A A
aim of detection group D- group D- E. faecalis, group D- group D- streptococci streptococci and streptococci streptococci or E. faecalis E. faecium
Tryptone Peptone Meat extract Yeast extract
Glucose *) Aesculin Tween 80
NaCI
K2HPO 4 KH2PO 4
Ferric NH4-citrate Sodium citrate Sodium carbonate *)
Sodium azide *) Ox bile Kanamycin sulphate Thallous acetate *) TTC (1% sol.) *)
Agar * *)
Final pH
Incubation"
10.0 10.0
10.0
0.1 10.0
14.0
6.0
ld 37~ l d b . or 4h 37~ 1-2d 45~
15.0 15.0 17.0 20.0 5.0 3.0
5.0
2.0/5.0
4.0
5.0 5.0 5.0
1.0
5.0
15.0 2.0
1.0 1.0
5.0 5.0
0.5 0.5 1.0 1.0
0.4 0.4 0.25 0.15 10.0
0.02
10.0 10.0
10.0 15.0 13.5 10.0
7.2 7.0 7.1 7.0
l d 37~ l d 37~ 1-3d 37~ 18h 42~ + + or 1 d b. ld b. 3d 37~
Preparation: *) added separately according to some original descriptions; * *) concentration depends on quality of agar; d = day; h = hour; b. = on bench (22_+ 2~ T1TG, thallous acetate tetrazolium glucose agar (Barnes, 1956; Mead, 1985); ME, M enterococcus agar (Slanetz and Bartley, 1957); CATC, citrate azide tween carbonate agar (Reuter, 1968); ABA, aesculin bile azide agar (Isenberg et al., 1970); KAA, kanamycin aesculin azide agar (Mossel et al., 1978).
accompanying food microflora. Additionally, two or three of the media were used during routine investigations of food specimens, including meat and milk and similar products as well as faecal samples from humans and animals.
Results were first published for the following media (Reuter, 1978): ABA (aesculin bile azide agar); KAA (kanamycin aesculin azide agar); CATC (citrate azide tween carbonate agar); ME (M-enterococcus agar); T|TG (thallous acetate

58
tetrazolium glucose agar); ScS (streptococci selective agar); KA (crystal violet azide agar).
In a second paper (Reuter, 1985b) the use of ME-, KAA-agar and CATC- medium was reported again with comparative tests of selectivity, recovery rate and differentiating criteria. The testing was done qualitatively with the loop streaking technique and quantitatively with the modified drop-plating technique as shown in the paper on lactic acid bacteria (Reuter, 1985a).
These results and some details from other investigations within our institute (Schotte, 1986) are summarized in Tables 4 and 5, where the composition of the media and data on selectivity and productivity are presented. Monographs on CATC, KAA, ME and TITG media are included in this volume.
It can be concluded that a highly specific medium for enterococci still does not exist. Some media allowed growth of non-enterococci and others partially inhibited single strains of enterococci. In some cases growth of non-streptococci also occurred. That was the case with ABA, TITG and ScS media. Single strains of staphylococci, even Enterobacteriaceae (hafnia and arizona) and Pseudomon- adaceae were able to grow.
Kanamycin aesculin azide agar KAA medium allowed partial growth of mesophilic lactobacilli, some of which
split aesculin which should be the marker reaction for enterococci. This caused errors especially in specimens containing high numbers of lactobacilli. Incubation of KAA at 42~ did not prevent the growth of aesculin-splitting lactobacilli or other Gram-positive organisms, such as Corynebacteria. Moreover, all colonies on KAA appeared to be similar, i.e., small and greyish-white. Increasing the content of sodium azide would eliminate this problem of selectivity but would reduce the recovery rate of enterococci as well.
Citrate azide tween carbonate agar CATC-medium was sufficiently selective for the enterococci provided plates
were read at definite times (see Table 5). Very good growth and a brilliant for- mazan reaction was obtained with Enterococcus faecalis and its subspecies. Colonies of E. faecium appeared with a weaker formazan reaction. Single strains sometimes showed slightly reduced productivity and smaller colonies. They would be separated from E. faecalis very easily by colony appearance. Recovery of Streptococcus boris and S. equinus was reduced by more than one lOgl0 cycle. Oral streptococci showed weak colour formation and small colonies. Lactic streptococci and leuconostocs did not grow at all. If plates are read at a definite time i.e., after 1 day incubation at 37~ this medium may be considered as highly selective and very productive for E. faecalis and its subspecies and as an adequate medium for E. faecium.
M-Enterococcus medium ME-medium, in this case made up according to an ISO recommendation, did
not inhibit the lactococci at all and also failed with some lactobacillus strains. However, other non-streptococci were more or less eliminated. This kind of medium is widely used, because it is listed by nearly all producers of media. ME
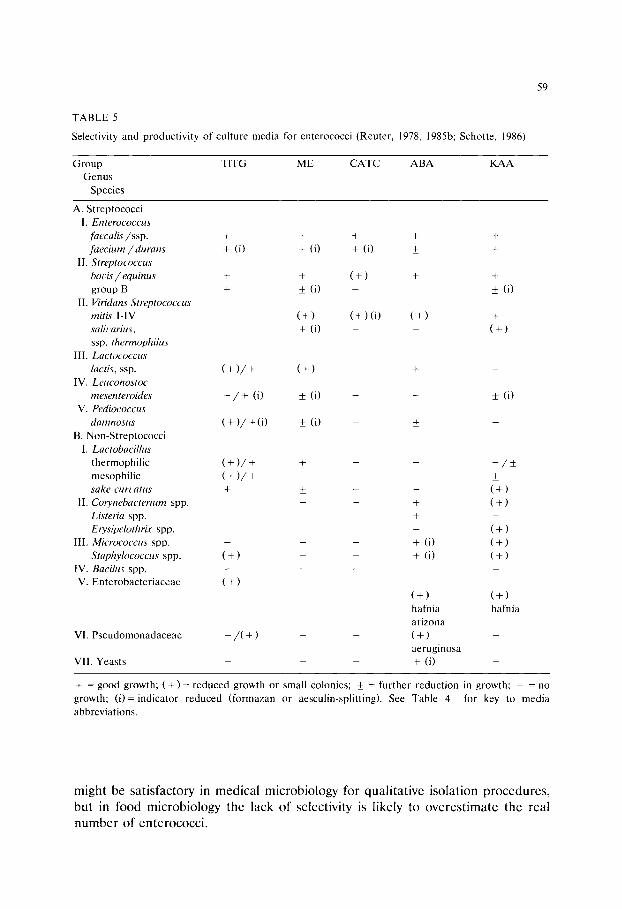
T A B L E 5
Selectivity and produc t iv i ty of cul ture media for enterococci (Reuter , 1978, 1985b; Schotte , 1986)
59
G r o u p Genus
Species
T1TG M E C A T C A B A K A A
A. St reptococci I. Enterococcus
faecalis/ssp. + + faecium/durans + (i) + (i)
II. Streptococcus boeis / equinus + + group B + + (i)
II. Viridans Streptococcus mitis I - IV ( + ) saliearius, + (i) ssp. thermophilus
III. Lactococcus lactis, ssp. ( + ) / + ( + )
IV. Leuconostoc mesenteroides - / + (i) ___ (i)
V. Pediococcus damnosus ( + ) / + (i) _+ ( i )
B. Non-S t rep tococc i
I. Lactobacillus the rmophi l i c ( + ) / + +
mesophi l ic ( + ) / + - sake-curvatus + +
II. Corynebacterium spp. Listeria spp. Erysipelothrix spp.
III. Micrococcus spp. - -
Staphylococcus spp. ( + ) -
IV. Bacilus spp. - - V. E n t e r o b a c t e r i a c e a e ( + ) -
VI. P s e u d o m o n a d a c e a e - / ( + )
VII . Yeas ts
+ + +
+ (i) + +
( + ) +
(+) (i) (+)
+
+
+ (i) + ( i )
(+) hafnia ar izona ( + )
ae rug inosa + (i)
+
• (i)
+
(+)
_ ( i )
- / + + (+) (+)
(+) (+) (+)
(+) hafnia
+ = good growth; ( + ) = reduced growth or small colonies; _+ - - f u r t h e r reduct ion in growth; - = no
growth; ( i ) = indicator r educed ( formazan or aesculin-spli t t ing). See Table 4 for key to media abbreviat ions.
might be satisfactory in medical microbiology for qualitative isolation procedures, but in food microbiology the lack of selectivity is likely to overestimate the real number of enterococci.

60
Thallous acetate tetrazolium glucose agar T1TG-medium was selective for group D-streptococci only if incubated at 42~
and read after not more than 18 h incubation. The pH of the medium should not exceed 6.2 (Mead, 1985). As shown previously (Reuter, 1968) continued incubation of this medium for a further 2 days at 20~ after 1 day at 30~ resulted in the appearance of streptococci, leuconostocs, pediococci and mesophilic lactobacilli. However, separation of these groups from the different colour reactions of the colonies was possible for the experienced worker. Less experienced workers would need to check identity using confirmatory tests.
Considerations for the correct use of the selective cultivation procedure
Sublethally damaged cells Resuscitation techniques are required in all cases where enterococci have been
subjected to any form of stress such as drying or freezing. Organisms may be incubated on or in a non-selective nutrient medium at 30-37~ (Barnes, 1976; Mossel and Corry, 1977) for 4-6 h, followed by overlaying with a selective medium. Alternatively, organisms on membrane filters may be incubated first on a non- selective and then on the selective medium (Barnes, 1976).
Storage of ready-to-use plates Following a short drying procedure, plates of selective media may be stored for
one week at 0-5~ in plastic bags. Storage at room temperature without drying is feasible for up to 2 weeks if plates are protected from light. If the selective agent is known to be unstable, media should be prepared just before use.
Confirmatory tests If new habitats or samples with unknown microflora composition are being
investigated confirmatory tests are required for colonies grown on selective media. First non-streptococci and then non-enterococci are eliminated. Micrococci and corynebacteria can be excluded by a positive catalase or benzidine-test on suspect colonies or by flooding the plates. Mesophilic lactic streptococci may be excluded by testing liquid subcultures for ability to grow at 45~ Further confirmation can be done by checking haemolytic or gelatinase-activities or mannitol fermentation and growth at 50~
If a high level of 'background flora' is expected, media with a high selectivity should be used.
References
Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (Eds.) 1987 Pharmacopoeia of Culture Media for Food Microbiology. Int. J. Food Microbiol. 5, 187-300.
Barnes, E.M. (1956) Methods for the isolation of faecal streptococci (Lancefield Group D) from bacon factories. J. Appl. Bacteriol. 19, 193-203.

61
Barnes, E.M. (1976) Methods for the isolation of faecal streptococci. Lab. Pract. 25, 145-147. Collins, M.D., Jones, D., Farrow, J.A.E., Kilpper-B~ilz, R. and Schleifer, K.H. (1984) Enterococcus
avium nom. rev., comb. nov.; E. casseliflavus nom. rev., comb. nov.; E. durans nora. rev., comb. nov; E. gallinarum comb. nov.; and E. malodoratus sp. nov. Int. J. Syst. Bacteriol. 34, 220-223.
Devriese, L.A., Collins, M.D. and Wirth, R. (1991) The genus Enterococcus. In: Balows, A., et al. (Eds.) The Prokaryotes, 2nd Edn., Vol. II, Springer-Verlag, Heidelberg, pp. 1465-1481.
Facklam, R.R. and Collins, D. (1989) Identification of Enterococcus species isolated from human infections by a conventional test scheme. J. Clin. Microbiol. 27, 731-734.
Hahn, G., Heeschen, W. and Tolle, W. (1970) Streptococcus: Struktur, Biochemie, Kultur und Klassifizierung. Kieler Milchwirtschaftl. Forschungsber., 22, 333-546.
Isenberg, H.D., Goldberg, J. and Sampson, J. (1970) Laboratory studies with a selective Enterococcus medium. Appl. Microbiol. 20, 433-436.
Mead, G.C. (1985) Isolation media for group D streptococci: comments. Int. J. Food Microbiol. 2, 115-117.
Mossel, D.A.A. and Corry, J.E.L. (1977) Detection and enumeration of sublethally injured pathogenic and index bacteria in foods and water processed for safety. Alimenta-Sonderausgabe pp. 19-34.
Mossel, D.A.A., Bijker, P.G.H. and Eelderink, I. (1978) Streptokokken der Lancefield-Gruppe D in Lebensmitteln und Trinkwasser: Ihre Bedeutung, Erfassung und Bek~impfung. Arch. Lebensmittel- hyg. 29, 121-127.
Mundt, J.O. (1984) Enterococci. In: Sneath, P.H.O., et al. (Eds.) Bergey's Manual of Systematic Bacteriology, Vol. 2, The Williams & Wilkins Co., Baltimore/London. pp. 1063-1066.
Nusser, E. (1991) Charakterisierung klinischer Isolate und einiger Kultursammlungs und Probiotika: Produktionsst~imme des Species E. faecium und faecalis durch Enzyme, Resistenz und Plasmide. Vet. Med. Diss., Free University Berlin.
Pompei, R., Thaller, M.C., Pittaluga, F., Flore, O. and Satta, G. (1992) Analysis of bacteriolytic activity patterns, a novel approval to the taxonomy of Enterococci. Int. J. System. Bacteriol., 42, 37-43.
Reuter, G. (1968) Erfahrungen mit N~ihrb6den fiir die selektive mikrobiologische Analyse von Fleis- cherzeugnissen. Arch. Lebensmittelhyg. 19, 53-57 and 84-89.
Reuter, G. (1978) Selektive Kultiviering von Enterokokken aus Lebensmitteln tierischer Herkunft. Arch. Lebensmittelhyg. 29, 128-131.
Reuter, G. (1985a) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68.
Reuter, G. (1985b) Selective media for group D-streptococci. Int. J. Food Microbiol. 2, 103-114. Schleifer, K.H. and Kilpper-B~ilz, R. (1984) Transfer of Streptococcus faecalis and Streptococcus
faecium to the Genus Enterococcus nom. rev. as Enterococcus faecalis comb. nov. and Enterococcus faecium comb. nov. Int. J. Syst. Bacteriol. 34, 31-34.
Schotte, M. (1986) Bewertung von 3 semi-quantitativen Prfifverfahren fiir die Qualit~itssicherung von N~ihrb6den in der Lebensmittelmikrobiologie mit Selektivmedien fiir Milchs~iurebakterien. Vet. Med. Diss., Free University Berlin.
Slanetz, L.W. and Bartley, C.H. (1957) Numbers of enterococci in water, sewage, and feces determined by the membrane filter technique with an improved medium. J. Bacteriol. 74, 591-595.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
63
Chapter 5 Culture media and methods for the isolation
of Listeria monocytogenes
G . D . W . Curt is a, W.H. Lee b
a Bacteriology Department John Radcliffe Hospital, Oxford OX3 9DU, UK b USDA-FSIS, Beltsville, MD 20705, USA
Abstract
The recovery of low numbers of Listeria monocytogenes from foods and environmental samples requires the use of enrichment cultures followed by selective plating and, where injured organisms are likely to be present, a pre-enrichment step. The development of selective and enrichment media for L. monocytogenes is traced and currently used media are discussed. Comparisons of media and methods for the culture of L. monocytogenes are reported but no single method can be recommended for all situations. Guidance is given on the choice of media and methods which is governed by the type of sample, number and nature of competing flora and cost.
During the past 10 years, large outbreaks of human listeriosis have been traced to contaminated coleslaw made from raw cabbage in Canada (Schlech et al., 1983), Mexican style soft cheese in the US (Linnan et al., 1988), soft cheese in Switzer- land (Bille, 1990) and liver pat6 in the UK (McLauchlin et al., 1991) and in Western Australia (Kittson, 1992). A very large human listeriosis outbreak oc- curred in France in 1992, the source of which has been identified as pork tongues in aspic (Goulet et al., 1993). Sporadic or single unrelated cases of human listeriosis are also common in many countries. Sporadic listeriosis in the US was associated with Listeria monocytogenes contaminated (opened) packages of ready- to-eat food from retail outlets in the domestic refrigerators of listeriosis patients by the US Centers for Disease Control (CDC)(Pinner et al., 1992). Schuchat et al.

64
(1992) showed that consumption of soft cheese and food from a delicatessen counter were each significantly associated with an increased risk of listeriosis and Schwartz et al. (1992)showed an association between consumption of uncooked hot dogs or undercooked chicken and sporadic cases of listeriosis. The CDC (Schuchat et al., 1992) estimated that there are about 1,850 cases of sporadic human listeriosis in the US with 425 deaths annually. The whole subject of L. monocytogenes as a food borne pathogen has been well reviewed by Farber and Peterkin (1991). Because human listeriosis is such a serious and often fatal disease, many countries have imposed a zero tolerance for L. monocytogenes in cooked ready-to-eat foods. Sampling of foods and the food plant environment for L. monocytogenes has become a major preoccupation worldwide. This in turn has lead to rapid improvements of L. monocytogenes recovery methods. Testing for L. monocytogenes from such diverse sources as farms, food plants, foods and kitchens requires different selective media and sampling procedures. This paper will at- tempt to review the current and some promising new recovery methods.
1. Listeria enrichment broths (LEB)
A liquid enrichment step is required for the recovery of low numbers ( < 20/g) of L. monocytogenes from foods and food plant environmental samples. For many years the only method available was the cold enrichment technique first advocated by Gray et al. (1948) in which sample material was inoculated into a nutrient broth without selective agents and held at 4~ for long periods. The method was primarily used for isolation from infected animal or human tissue but its usefulness in the examination of silage (Gray, 1960) and milk (Larsen, 1966) was also demonstrated. Its chief drawback was the need to make subcultures from the broth at intervals up to several months or even a year.
Early attempts at producing a selective broth which would allow the isolation of listeria within days of inoculation resulted in tryptose phosphate broth with polymyxin (Bojsen-Mr 1972) and Levinthal broth with trypaflavine and nalidixic acid (Ralovich et al., 1972). Watkins and Sleath (1981) combined a period of non-selective cold enrichment with subsequent selective enrichment in a broth containing potassium thiocyanate and nalidixic acid as selective agents. Although originally designed to recover listeria from sewage this system has been successfully used by Lewis and Corry (1991) for foods. Increased interest in L. monocytogenes as a food borne pathogen led to the development of many selective enrichment broths, the chief features of which are shown in Table 1.
With the exception of L-PALCAMY (van Netten et al., 1989) all these selective broths rely on nalidixic acid to inhibit Gram-negative organisms and acriflavine to provide additional selectivity especially with regard to Gram-positive organisms. L-PALCAMY incorporates only 5 mg/1 acriflavine and substitutes polymyxin, ceftazidime and lithium chloride for nalidixic acid. Lithium chloride is also used in Fraser's broth (Fraser and Sperber, 1988) whilst the FDA broth of Lovett et al. (1987) includes cycloheximide to inhibit yeasts and moulds.
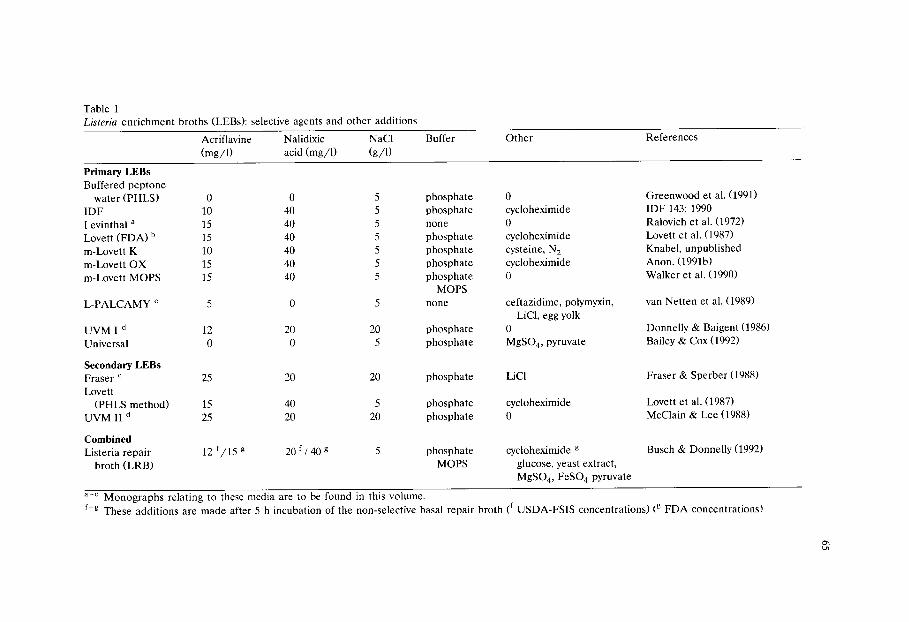
Table 1 Listeria enrichment broths (LEBs): selective agents and other additions
Acriflavine Nalidixic NaCl (mg/1) acid (mg/l) (g/ l)
Buffer Other References
Primary LEBs Buffered peptone
water (PHLS) 0 IDF 10 Levinthal a 15 Lovett (FDA) b 15 m-Lovett K 10 m-Lovett OX 15 m-Lovett MOPS 15
L-PALCAMY c 5
UVM I d 12 Universal 0
Secondary LEBs Fraser e 25 Lovett
(PHLS method) 15 UVM I Id 25
Combined Listeria repair
broth (LRB)
0 5 phosphate 40 5 phosphate 40 5 none 40 5 phosphate 40 5 phosphate 40 5 phosphate 40 5 phosphate
MOPS 0 5 none
20 20 phosphate 0 5 phosphate
0 cycloheximide 0 cycloheximide cysteine, N 2 cycloheximide 0
ceftazidime, polymyxin, LiCl, egg yolk
0 MgSO4, pyruvate
Greenwood et al. (1991) IDF 143:1990 Ralovich et al. (1972) Lovett et al. (1987) Knabel, unpublished Anon. (1991b) Walker et al. (1990)
van Netten et al. (1989)
Donnelly & Baigent (1986) Bailey & Cox (1992)
20 20 phosphate LiCl Fraser & Sperber (1988)
40 5 phosphate cycloheximide Lovett et al. (1987) 20 20 phosphate 0 McClain & Lee (1988)
12 f /15 g 20 f / 40 g 5 phosphate cycloheximide g /
MOPS glucose, yeast extract, MgSO4, FeSO 4 pyruvate
Busch & Donnelly (1992)
a - e Monographs relating to these media are to be found in this volume. f-g These additions are made after 5 h incubation of the non-selective basal repair broth (f USDA-FSIS concentrations) (g FDA concentrations)

66
The original formulation of Lovett et al. (1987) was designed for isolation of L. monocytogenes from milk and the University of Vermont (UVM) broths of Don- nelly and Baigent (1986) were developed by the USDA for use with meats (McClain and Lee, 1988). In view of the fact that the sample often constitutes 10% of the incubation mixture, it may prove useful to keep these applications in mind when choosing a selective broth. Samples likely to be heavily contaminated with interfering organisms may best be enriched in the more selective L-PALCAMY or even the thiocyanate nalidixic acid broth of Watkins and Sleath (1981). However, highly selective media should not be used unnecessarily as the concentrations of inhibitors may slow or prevent the growth of sublethally-damaged or of exception- ally sensitive strains of Listeria spp. Care should also be taken to adhere to the recommended incubation temperature as many listeria strains show an increased susceptibility to antibiotics when incubated at 37 rather than 30~ (Curtis et al., 1989b).
The various modifications of Lovett's broth are the results of different attempts to prevent a rapid drop in pH which occurs particularly after the inoculation of soft cheese and inhibits growth of listeria (Anon., 1991b; Walker et al., 1990). This does not happen with the UVM broths which are more strongly buffered. The unpublished modification of Knabel is designed to create a degree of anaerobiosis to enhance the recovery of injured cells (see below).
Two enrichment broths, Fraser's and L-PALCAMY, include an indicator system consisting of aesculin and ferric iron. Organisms which hydrolyse aesculin produce a blackening of the broth due to the formation of iron phenolic compounds derived from the aglucon. Broths without blackening are considered negative for listeriae and require no further examination. Up to 38% false positive results have been recorded with Fraser's broth but no false negative results were found provided incubation was continued for 48 h (Warburton et al., 1991a).
2. Recovery of injured Listeria monocytogenes
The selective agents in Lovett, Fraser, UVM and L-PALCAMY (Table 1) inhibit the growth and recovery of injured Listeria spp. from environmental and food samples (Busch and Donnelly, 1992). To overcome this problem, the UK Public Health Laboratory Service (Greenwood et al., 1991; Morris and Ribeiro, 1991) used the totally non-selective buffered peptone water incubated at 30~ for 24 h as the primary enrichment broth and the Lovett LEB incubated for 24 and 48 h for the secondary enrichment. Another nonselective universal pre-enrichment broth is that described for the recovery of both Listeria and Salmonella spp. from foods by Bailey and Cox (1992). Comparisons of these non-selective broths with other enrichment systems need to be made to assess their performance in the recovery of injured and uninjured Listeria from various naturally contaminated foods, particu- larly with regard to the possibility of overgrowth by competing organisms during the later stages of incubation.

67
Busch and Donnelly (1992) introduced a new concept, that of the dual purpose non-selective/selective broth. A non-selective 3-(N-morpholino)propanesulphonic acid (MOPS) buffered enrichment broth base (LRB)with magnesium sulphate and sodium pyruvate added to improve recovery of injured cells, was inoculated with the sample. After allowing 5 h for the injured Listeria to recover, the selective agents acriflavine and nalidixic acid were then added to the LRB base and incubation continued in the usual way. A similar technique may conveniently be used with some commercial presentations of other LEBs where the basal broth is prepared from a dehydrate and the selective supplements are supplied in vials for reconstitution and addition to the broth after sterilization. Delaying this addition for some hours after inoculation of the basal broth will allow resuscitation of injured bacteria.
Knabel et al. (1990) reported that strict anaerobic incubation aided in the recovery of severely injured L. monocytogenes. In 1992, Knabel (personal commu- nication) simplified this anaerobic recovery procedure by using a modified Lovett LEB (m-Lovett K), with 10 mg/1 of acriflavine and 0.5 g/1 filter sterilized cysteine added, then flushing the flasks with nitrogen to remove atmospheric oxygen. This was used as the primary enrichment step for the recovery of severely injured L. monocytogenes which had been heated to 62.8~ for 5 min and inoculated into raw milk. After 48 h incubation at 30~ the primary enrichment broth was transferred to Fraser's LEB. Knabel (personal communication) claimed that severely injured L. monocytogenes did not recover in the aerobic LRB of Busch and Donnelly (1992) or in nitrogen-flushed UVM I LEB with 0.5 g/1 filter sterilized cysteine added. It is possible that oxygen is toxic to the growth of severely injured listeriae, but not to the growth of moderately injured listeriae heated to 56~ for 50 min (Busch and Donnelly, 1992). Smith (1990) reported that injured L. monocytogenes did not recover in a non-selective medium at 5 or 12~ but Meyer and Donnelly (1992) stated that injured listeria can recover at 5~ in milk. Thus the media and conditions for the recovery of injured Listeria spp. remain uncertain and need further study and substantiation. The ability to isolate injured L. monocytogenes from foods and environments is important because if injured Listeria spp. are not detected, the bacteria may slowly recover and colonize foods and food plant environments.
3. Listeria selective agars (LSA)
The first significant step in producing a selective agar for listeria was that of Gray et al. (1950)who used potassium tellurite to inhibit Gram-negative organisms and examined the growth under a dissecting microscope. In 1960, McBride and Girard added lithium chloride, glycine and blood to phenyl ethanol agar to produce a medium which, until relatively recently, was the only readily available commercially produced selective agar for listeria isolation. Its chief drawback was its inability to inhibit enterococci. Ralovich et al. (1971) published the first truly selective agar using trypaflavine and nalidixic acid in a serum agar base and

68
Henry's illumination method to identify listeria colonies. Lee and McClain (1986) modified McBride's agar by substituting glycine anhydride for glycine, increasing the lithium chloride content and adding moxalactam. Omitting blood allowed Henry's method to be used for identification of listeria colonies. The discovery that listeriae tolerate up to 15 g/1 of lithium chloride, a concentration which inhibits almost all enterococci, led to many of the currently used formulations, most of which also incorporate the aesculin-ferric iron indicator system thus obviating the need for special illumination techniques to identify presumptive listeria colonies. Since the advent of Oxford agar (Curtis et al., 1989a) the trend has generally been towards a simplification of media, more reliance being placed on the use of second and third generation cephalosporin antibiotics and less on acriflavine, whilst polymyxins are still used for inhibiting most Gram-negative bacteria. The simplest LSA is the modified lithium chloride-ceftazidime agar (LCAM) developed by Lachica (1990 and personal communication). The selective agents in the commonly used agars are shown in Table 2. A problem in the use of cephalosporins in selective media for listeria was noted by Curtis et al. (1989b). They showed that the three commonly used cephalosporins, cefotetan, ceftazidime and moxalactam were more inhibitory to some strains of L. seeligeri when incubation was carried out at 37~ than at 30~ Two out of 14 strains of L. monocytogenes also showed this effect with ceftazidime but not with cefotetan or moxalactam. A similar effect has been observed with L. monocytogenes ATCC 19111, which is inhibited on agar at 35~ with ceftazidime but grows well at 30~ (Lee, unpublished results). Cef- tazidime at 20 mg/1 renders media such as PALCAM (van Netten et al., 1989) and modified Oxford (MC) agar (Lee, unpublished results)very selective and this may account for the occasional failure to recover strains of L. monocytogenes in comparative trials (Warburton et al., 1991b; Kovacs and Ralovich, 1991; Westoo and Peterz, 1992). These workers found Oxford agar best for isolation of Listeria spp. from enrichment broths followed by LPM, PALCAM and modified Oxford agar. A feature of PALCAM agar is the incorporation of a second indicator system, mannitol and phenol red, in addition to the aesculin-ferric iron complex. Microaerobic incubation is recommended by the authors for this agar. Dominguez-Rodriguez LSAMm agar includes potassium tellurite to enhance haemolysis by L. monocytogenes colonies in a secondary blood containing overlay (Blanco et al., 1989; Fernandez-Garayzabel et al., 1992). Whilst undoubtedly useful in differentiating haemolytic from non-haemolytic colonies the technique is time consuming and adds a further day's incubation to the process time. This is avoided in the enhanced haemolysis agar (EHA) of Cox et al. (1991a,b) but at the considerable expense of adding sheep blood, sphingomyelinase and 4-methyl umbelliferyl-/3-D-glucoside to a basal medium with PALCAM supplements. It is reported (Cox et al., 1991b) that this EHA has recovered significantly more Listeria spp. and L. monocytogenes from food samples than Oxford agar.
Modified Vogel Johnson agar (MVJ) has been further modified (Smith and Buchanan, 1990) by the addition of Tween 80 and egg yolk to promote the recovery of injured listeriae. However, fats, Tween 80 and especially Tween 20 can be inhibitory to growth of Listeria spp. if they are hydrolysed to fatty acids by the

Table 2 Selective agents used in Listeria selective agars
E HA LCA(M) LPM a
LiC1 Polymyxin (P) Acriflavine (g/1) or Colistin (mg/1)
(C) (mg/1)
10 P10 5 5 0 0 5 0 0
Tellurite (mg/1)
0 0 0
LSAMm b 15 P10 5 200 MC e 12 C10 0 0 MVJ(M) 5 0 0 105
Oxford c 15 C20 5 0
PALCAM d 10 P10 5 0
Other (mg/1)
Ceftazidime 30 Ceftazidime 50 Moxalactam 20,
Glycine anhydride 10 Phenylethanol 2500
Ceftazidime 20 Ceftazidime 20 Bacitracin 20
Nalidixic acid 50 Moxalactam 5 Glycine 10
Fosfomycin 10 Cefotetan 2 Cycloheximide 400
Ceftazidime 30
References
Cox et al. (1991b) Lachica (1990 & unpublished) Lee & McClain (1986)
Blanco et al. (1989) Lee, unpublished Smith & Buchanan (1990)
Curtis et al. (1989a)
van Netten et al. (1989)
a-e Monographs relating to these media are to be found in this volume.

70
microorganisms in the foods (Wang and Johnson, 1992). Seeliger and Jones (1986) reported that the growth of Listeria spp. was inhibited on the lactobacillus se|ective (MRS) medium of de Man et al. (1960), which contains 1 g/1 Tween and 5 g / l sodium acetate as the principle selective agents. The addition of 3 g/1 of sodium diacetate to processed meat products also controls the growth of Listeria spp. in processed meats (Schmidt and Leistner, 1991). Thus in medium develop- ment, there should be an awareness of compounds which may be used to inhibit the growth of listeriae in foods.
4. Procedures for the isolation of L. monocytogenes
Current procedures cannot recover very low levels (less than 0.3/g) of L. monocytogenes from foods (Hayes et al., 1991; Norrung et al., 1991; Warburton et al., 1991b) and the ability of present methods to achieve complete recovery of all injured cells is in doubt. For the best possible recovery a three stage procedure is recommended (i) pre-enrichment or resuscitation (ii) selective enrichment and (iii) selective plating. Media for each of these stages have been discussed above and consideration must now be given to the various combinations of broths and agars which have been assessed.
Two methods were developed in the US, the FDA method (Lovett, 1987; Lovett and Hitchens, 1988) and the USDA method (McClain and Lee, 1988). Both have been subject to a number of modifications and details of the original methods together with some modifications are given in tabular form by Warburton et al. (1991a). A group of experts from the International Dairy Federation (IDF), the International Standards Organisation and the Association of Official Analytical Chemists have also produced a provisional standard method (IDF, 143:1990). This recommends the use of a modification of Lovett's broth with reduced acriflavine for enrichment, followed by plating onto Oxford agar. The US CDC (Hayes et al., 1992) compared the recoveries of L. monocytogenes from naturally contaminated foods by the revised FDA procedure using Lovett LEB, with LPM and Oxford agars, the revised USDA procedure using UVMI and Fraser LEBs with modified Oxford (MC) agar and the Netherlands procedure using L-PALCAMY LEB with PALCAM agar. They found these procedures to be almost equally effective, but the recovery of L. monocytogenes by a combination of any two methods was significantly better than by one method used alone (P < 0.02). Westoo and Peterz (1992) found no difference in the recovery of inoculated L. monocytogenes when comparing Lovett's, UVMI and UVMII broths. Rollier et al. (1991) found that the use of UVMI and UVMII broths with plating on Oxford agar was more effective than using Lovett LEB and L-PALCAMY enrichment when examining fermented sausages. They found no difference between Oxford and PALCAM agars for plating.
An examination of commercial minced meat was carried out by Kokubo et al. (1990) in Japan using Lovett's (FDA) broth and trypaflavine nalidixic acid broth with Oxford, LPM, McBride and trypaflavine nalidixic acid agars. The best results

71
were obtained with Lovett's broth incubated for 7 days and subcultured onto Oxford agar. Subculture onto LPM was almost as successful.
Very extensive collaborative tests were carried out by Warburton et al. (1991a,b). Using both the original and revised FDA and USDA methods, they found that no single procedure recovered all of the L. monocytogenes in naturally contaminated samples. They recommended for all samples the use of the modified USDA method with subculture of UVMI broth to Fraser's broth and plating media at 24 and 48 h. They also recommended the use of Oxford and at least one other agar, either LPM or MOX (Warburton et al., 1991b). The use of several plating media was shown to give substantially improved isolation rates (up to 46%) in the recovery of L. monocytogenes (Warburton et al., 1991a).
5. Enumeration of L. monocytogenes in foods by direct plating
Several countries in Europe are considering setting limits for L. monocytogenes in certain categories of foods at the retail level (Anon., 1991a). Since many refrigerated ready-to-eat foods present no growth barrier to L. monocytogenes, it can grow to high numbers. In an extensive search for the possible causes of sporadic human listeriosis in the US, the CDC (Pinner et al., 1992) found strong positive correlation of human listeriosis to the foods in which high numbers of L. monocytogenes could be recovered by direct plating on LPM agar. Also, in the UK, pat6 with high counts of L. monocytogenes enumerated using Oxford agar was epidemiologically linked to a significant number of cases of listeriosis (McLauchlin et al., 1991; Morris and Ribeiro, 1991). High numbers of L. monocytogenes can easily be detected by plating the 1 in 10 food suspension in primary enrichment broth, immediately after preparation at 0 h onto LPM, Oxford, MC or PALCAM agars. This method assumes that L. monocytogenes growing in food samples is not injured and is able to form colonies on the selective plating media. However, in a few instances, high numbers of L. monocytogenes have been recovered by the MPN enrichment method but not by direct plating on LPM agar (Hayes et al., 1991, Pinner et al., 1992). Oxford agar has been used extensively by PHLS laboratories (Greenwood et al., 1991; McLauchlin et al., 1991; Morris and Ribeiro, 1991) for the direct enumeration of high numbers of L. monocytogenes in retail foods in the UK. However, the presence of large numbers of some competing bacteria can sometimes overwhelm the selective agents in this medium. Ringle et al. (1991) preferred to use the more selective PALCAM to Oxford agar for direct plating of soft cheeses. Vazquez-Boland et al. (1992) found that the more selective LSAMm and PALCAM agars were suitable for direct plating of silage samples for Listeria spp., but that Oxford agar was overwhelmed by the competing bacteria. Warburton et al. (1991b) drew attention to considerable (1-3 log)variat ion in recovery of certain strains on selective plating media and Tiwari and Aldenrath (1990) reported a strain which they were able to grow only on Oxford agar. Direct counting of listeriae in foods is therefore dependent on the nature and numbers of interfering organisms and on the ability of the chosen plating media to recover

72
quanti tat ively all of the strains sought. It should be no ted also that none of the med ia in common use at the m o m e n t will distinguish L. monocy togenes f rom o ther listeriae.
The growth of L. monocy togenes in natural ly con tamina ted food is both uneven and erratic. Michard et al. (1989) r epor t ed 100-fold higher numbers of L. monocy -
togenes on the surface of soft cheeses than in the centre , and a similar dis tr ibut ion of Listeria spp. has been no ted in retail vacuum packaged processed meats (Lee, unpubl i shed results). Thus counts of Listeria spp. in such foods may depend on which par ts of the sample are examined. Work by G r e e n w o o d et al. (1991) on soft cheeses and Schmidt and Kaya (1990) and Grau and Vander l inde (1992) on
processed meats found counts in excess of 1000 /g of L. monocy togenes in 8.4, 2.3 and 4% of samples, respectively, demons t ra t ing the need to examine large num- bers of samples in o rder to detec t sporadic contaminat ion . In this respect the use of the swabbing technique of Lee (1989) will allow investigation of large numbers of, for example, out of date vacuum packs of meat , detect ing counts in excess of 1 0 0 / g with comparat ively little effort or expense.
In summary, cur rent isolation media allow the recovery of L. monocy togenes
f rom a variety of foods and food plant envi ronments with relative ease. Yet, to date, ident if icat ion and recovery of con tamina ted foods responsible for h u m a n listeriosis ou tbreaks remains a slow and difficult process (Anon., 1992; McLauchl in et al., 1991).
References
Anon. (1991a) Emfehlungen des Bundesgesundheitsamtes zum nachweis zur bewertung von Listeria monocytogenes in Lebensmitteln. Bundesgesundheitsblatt 34, 227-229.
Anon. (1991b) Buffered listeria enrichment broth Code CM897 Oxoid folio No. 348 Unipath Limited, Basingstoke, UK.
Anon. (1992) L'6pid6mie de listeriose en France. Bull. Epidemiol. Hebdom. 38, 181-182. Bailey, J.S. and Cox, N.A. (1992) Universal pre-enrichment broth for the simultaneous detection of
Salmonella and Listeria in foods. J. Food Prot. 55, 256-259 Bille, J. (1990) Epidemiology of human listeriosis in Europe with special reference to the Swiss
outbreak. In: A.J. Miller, J.L. Smith and G.A. Somtukti, (Eds). Foodborne Listeriosis. Elsevier, Amsterdam, pp. 71-74.
Blanco, M.B., Fernandez-Garayzabal, J.F., Dominguez, L., Briones, V., Vazquez-Boland, J.A., Blanco, J.L., Garcia, J.A. and Suarez, G. (1989) A technique for the direct indentification of haemolytic- pathogenic Listeria on selective plating media. Lett. Appl. Microbiol. 9, 125-128.
Bojsen-Mr J. (1972) Human Listeriosis. Diagnostic, epidemiological and clinical studies. Acta Pathol. Microbiol. Scand. Suppl B. 229, 1-157.
Busch, S.V. and Donnelly, C.W. (1992) Development of a repair enrichment broth for resuscitation of heat-injured Listeria monocytogenes and Listeria innocua. Appl. Environ. Microbiol. 58, 14-20.
Cox, J.L., Siebenga, A., Pedrazzini, C. and Moreton, J. (1991a). Enhanced haemolysis agar (EHA): an improved selective and differential medium for isolation of Listeria monocytogenes. Food Microbiol. 8, 37-49.
Cox, J.L., Siebenga, A. and Pedrazzini, C. (1991b) Performance of enhanced haemolysis agar compared to Oxford medium for the isolation of Listeria monocytogenes from food enrichment broths. Food Microbiol. 8, 51-62.
Curtis, G.D.W., Mitchell, R.G., King, A.F. and Griffen, E.J. (1989a) A selective differential medium for the isolation of Listeria monocytogenes. Lett. Appl. Microbiol. 8, 95-98.

73
Curtis, G.D.W, Nichols, W.W. and Falla, T.J. (1989b) Selective agents for Listeria can inhibit their growth. Lett. Appl. Microbiol. 8, 169-172.
de Man, J.C., Rogosa, M. and Sharpe, M.E. (1960) A medium for the culture of Lactobacilli. J. Appl. Bacteriol. 23, 130-135.
Donnelly, C.W. and Baigent, G.J. (1986) Method for flow cytometric detection of Listeria monocyto- genes in milk. Appl. Environ. Microbiol. 689-695.
Farber, J.M. and Peterkin, P.I. (1991) Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 55, 476-511.
Fernandez-Garayzabal, J.F., Delgado, C., Blanco, M., Vazquez-Boland, J.A., Briones, V., Suarez, G. and Dominguez, L. (1992) Role of potassium tellurite and brain heart infusion in expression of the haemolytic phenotype of Listeria spp. on agar plates. Appl. Environ. Microbiol. 58, 434-438.
Fraser, J.A. and Sperber, W.H. (1988) Rapid detection of Listeria spp. in foods and environmental samples by esculin hydrolysis. J. Food Prot. 51, 762-765.
Goulet, V., Lepoutre, A., Rocourt, J., Courtieu, A.L., Dehaumont, P. and Veit, P (1993) Epid6mie de list6riose en France. Bull. Epidemiol. Hebdom. 4/93, 13-14.
Grau, F.H. and Vanderlinde, P.B. (1992) Occurrence, number and growth of Listeria monocytogenes on some vacuum-packaged processed meats. J. Food Prot. 55, 4-7.
Gray, M.L. (1960) Isolation of Listeria monocytogenes from oat silage. Science 132, 1767-1768. Gray, M.L., Stafseth, H.J. and Thorp, F. Jr. (1950) The use of potassium tellurite, sodium azide and
acetic acid in a selective medium for the isolation of Listeria monocytogenes. J. Bacteriol. 59, 443-444.
Gray, M.L., Stafseth, H.J., Thorp, F. Jr., Sholl, L.B. and Riley, W.F. Jr. (1948) A new technique for isolating Listerellae from the bovine brain. J. Bacteriol. 55, 471-476.
Greenwood, M.H., Roberts, D. and Burden P. (1991) The occurrence of Listeria species in milk and dairy products: a national survey in England and Wales. Int. J. Food Microbiol. 12, 197-206.
Hayes, P.S., Graves, L.M., Ajello, G.W., Swaminathan, B., Weaver, R.E., Wenger, J.D., Schuchat, A. and Broome, C.V. (1991) Comparison of cold enrichment and US Department of Agriculture methods for isolating Listeria monocytogenes from naturally contaminated foods. Appl. Environ. Microbiol. 57, 2109-2113.
Hayes, P.S., Graves, L.M., Swaminathan, B., Ajello, G.W., Malcolm, G.B., Weaver, R.E., Ransom, R., Deaver, K., Plikaytis, B.D., Schuchat A., Wenger, J.D., Pinner, R.W., Broome, C.V. and the Listeria study group. (1992) Comparison of three selective enrichment methods for the isolation of Listeria monocytogenes from naturally contaminated foods. J. Food Protect. 55, 952-959.
IDF (1990) Milk and milk products. Detection of Listeria monocytogenes. Provisional International Standard IDF 143:1990. Brussels, Belgium.
Kittson, E. (1992) A case cluster of listeriosis in Western Australia with links to pats consumption. Abstr. Eleventh International Symposium on Problems of Listeriosis, p. 18.
Knabel, S.J., Walker, H.W., Hartman, P.A. and Mendonca, A.F. (1990) Effects of growth temperature and strictly anaerobic recovery on the survival of Listeria monocytogenes during pasteurization. Appl. Environ. Microbiol. 56, 370-376.
Kokubo, Y., Iida, T., Kaneko, S. and Maruyama, T. (1990) Evaluation of enrichment and plating media for isolating Listeria monocytogenes from raw meat. J. Food Hyg. Soc. Japan 31, 51-56.
Kovacs, H.D. and Ralovich, B. (1991) Comparative examination of selective media for isolation of Listeria strains. Acta Microbiol. Acad. Sci. Hung. 38, 141-145.
Lachica, R.V. (1990) Selective plating medium for quantitative recovery of food-borne Listeria monocy- togenes. Appl. Environ. Microbiol. 56, 167-169.
Larsen, H.E. (1966) Isolation technique for Listeria monocytogenes: primary cultivation and cold incubation technique. Proc. Third Int. Symp. Listeriosis, Bilthoven, The Netherlands. pp 43-48.
Lee, W.H. and McClain, D. (1986) Improved Listeria monocytogenes selective agar. Appl. Environ. Microbiol. 52, 1215-1217.
Lee, W.H. (1989) Screening test to detect Listeria (> 100/g) in foods. Abstr. ASM Annual Meeting, New Orleans, LA, p. 325.
Lewis, S.J. and Corry, J.E.L. (1991) Comparison of a cold enrichment and the FDA method for

74
isolating Listeria monocytogenes and other Listeria spp. from ready to eat food on retail sale in the UK. Int. J. Food Microbiol. 12, 281-286.
Linnan, M.J., Mascola, L., Lou, X.D., Goulet, V., May, S., Salminen, C., Hird, D.W., Yonekura, L., Hayes, P., Weaver, R., Audurier, A., Plikaytis, B.D., Fannin, S.L., Kleks, A. and Broome, C.V. (1988) Epidemic listeriosis associated with Mexican-style cheese. N. Engl. J. Med. 319, 823-828.
Lovett, J. (1987) Lbteria isolation. In: Bacteriological Analytical Manual, AOAC. Arlington, VA. Lovett, J. and Hitchins, A.D. (1988) Listeria isolation. Revised October 13, 1988. Bacteriological
Analytical Manual, AOAC. Arlington, Va. Lovett, J., Francis, D.W. and Hunt, J.M. (1987) Lbteria monocytogenes in raw milk: detection,
incidence and pathogenicity. J. Food Prot. 50, 188-192. McBride, M.E. and Girard, K.F. (1960) A selective method for the isolation of Listeria monocytogenes
from mixed bacterial populations. J. Lab. Clin. Med. 55, 153-157. McClain, D. and Lee, W.H. (1988) Development of USDA-FSIS method for isolation of Lbteria
monocytogenes from raw meat and poultry. J. Assoc. Off. Anal. Chem. 71, 660-664. McLauchlin, J., Hall, S.M., Velani, S.K. and Gilbert, R.J. (1991) Human listeriosis and pate: a possible
association. Br. Med. J. 303, 773-775. Meyer, D. and Donnelly, C.W. (1992) Effect of incubation temperature on repair of heat injured listeria
in milk. J. Food Protect. 55, 579-582. Michard, J., Jardy, N. and Gey, J.L. (1989) D6nombrement et localisation de Listeria monocytogenes
dans les fromage a pat6 molle et a croute lav6e fabriques avec du cru lait en provenance d'une entreprise fromagbre. Microbiol. Alim. Nutrit. 7, 131-137.
Morris, I.J. and Ribeiro, C.D. (1991) The occurrence of Listeria species in pat6: the Cardiff experience 1989. Epidemiol. Infect. 107, 111-117.
Norrung, B., Solve, M., Ovesen, M. and Skovgaard, N. (1991) Evaluation of an ELISA test for detection of L&teria spp. J. Food Prot. 54, 752-755.
Pinner, R.W., Schuchat, A., Swaminathan, B., Hayes, P.S., Deaver, K.A., Weaver, R.E, Plitkaytis, B.D, Reaves, M., Broome, C.V., Wenger, J.D. and the L&teria Study Group. (1992) Role of foods in sporadic listeriosis. II. Microbiological and epidemiological investigation. J. Am. Med. Assoc. 267, 2046-2050.
Ralovich, B., Forray, A., Mero, E., Malovics, I. and Szazados, I. (1971) New selective medium for isolation of Listeria monocytogenes. Zbl. Bakteriol. I. Abt. Orig. 216, 88-89.
Ralovich, B., Emody, L., Malovics, I., Mero, E. and Forray, A. (1972) Methods to isolate L. monocytogenes from different materials. Acta Microbiol. Acad. Sci. Hung. 19, 367-369.
Ringle, P., Vincent, J.P. and Catteau, M. (1991) D6nombrement de Listeria dans les produit laitiers. In: A. Amgar (Ed.). Proceedings of the International Conference on Lbteria and Food Safety. p. 211. ASEPT, Laval, France.
Rollier, I., Croegaert, Th., Verplaetse, A. and Van Hoof, J. (1991) Comparison of three plating media for the enumeration and three media for isolation of Listeria spp. in fermented sausages. Arch. Lebensmittelhyg. 42, 65-69.
Schlech, W.F., Lavigne, P.M., Bortolussi, R.A., Allen, A.C., Haldane, E.V., Wort, A.J., Hightower, A.W., Johnson, S.E., King, S.H., Nicholls, E.S. and Broome, C.V. (1983) Epidemic listeriosis: evidence for transmission by food. N. Engl. J. Med. 308, 203-206.
Schmidt, U. and Kaya, M.K. (1990) Verhalten von listerien bei fleisch und fleischerzeugnissen in vakuum-verpacking. Mitteilungsbl. Bundesanstalt Fleischforschung. 29, 214-218.
Schmidt, U. and Leistner, L. (1991) Vorkommen and verhalten von listerien bei unverpaktem Bruh- wurstaufschnitt (Bediennungsware). Mitteilungsbl. Bundesanstalt Fleischforschung. 30, 178-185.
Schuchat, A., Deaver, K., Wenger, J.D., Plitkaytis, B.D., Mascola, L., Pinner, R.W., Reingold, A., Broome, C.V. and the Lbteria Study Group. (1992) Role of foods in sporadic listeriosis. I. Case-control study of dietary risk factors. J. Am. Med. Assoc. 267, 2041-2045.
Schwartz, B., Ciesielski, C.A., Broome, C.V., Gaventa, S., Brown, G.R., Gellin, B.G., Hightower, A.W., Mascola, L. and the listeriosis study group. (1992) Association of sporadic listeriosis with consump- tion of hot dogs and undercooked chicken. Lancet ii, 779-782.
Seeliger, H.P.R. and Jones, D. (1986) The genus Lbteria. In: P.H.A. Sneath, N.S. Mair, M.E. Sharpe

75
and J.G. Holt (Eds.). Bergey's Manual of Systematic Bacteriology 2. Williams and Wilkins, Baltimore, pp. 1235-1245.
Smith, J.L. (1990) Stress induced injury in Listeria monocytogenes. In: A.J. Miller, J.L. Smith and G.A. Somkuti (Eds.). Foodborne listeriosis. Elsevier, Amsterdam, pp. 203-209.
Smith, J.L. and Buchanan, R.L. (1990) Identification of supplements that enhance the recovery of Listeria monocytogenes on modified Vogel Johnson agar. J. Food Safety. 10, 155-163.
Tiwari, N.P. and Aldenrath, S.G. (1990) Isolation of Listeria monocytogenes from food products on four selective plating media. J. Food Prot. 53, 382-385.
van Netten, P., Perales, I., Curtis, G.D.W. and Mossel, D.A.A. (1989) Liquid and solid selective differential media for the detection and enumeration of Listeria monocytogenes and other Listeria spp. Int. J. Food. Microbiol. 8, 229-316.
Vasquez-Boland, J.A., Dominguez, L., Blanco, M., Rocourt, J., Fernandez-Garazabal, J.F., Gutierrez, C.B., Tascon, R.I. and Rodriguez-Ferri, E.F. (1992) Epidemiologic investigation of a silage-associ- ated epizootic ovine listeric encephalitis, using a new Listeria-selective medium and phage typing. Am. J. Vet. Res. 53, 368-371.
Walker, S.J., Archer, P. and Appleyard, J. (1990) Comparison of the Listeria-Tek ELISA kit with cultural procedures for the detection of Listeria species in foods. Food Microbiol. 7, 335-342.
Wang, L.L. and Johnson, E.A. (1992) Inhibition of Listeria monocytogenes by fatty acids and monoglyc- erides. Appl. Environ. Microbiol. 58, 624-629.
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Hunt, T., Messier, S., Plante, R., Tiwari, N.P. and Vinet, J. (1991a) A comparative study of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes in food. Int. J. Food Microbiol. 13, 105-118.
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Tiwari, N., Babiuk, T., LaCasse, P. and Read, S. (1991b) A Canadian comparative study of modified versions of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes. J. Food Prot. 54, 669-676.
Watkins, J. and Sleath, K.P. (1981) Isolation and enumeration of Listeria rnonocytogenes from sewage, sewage sludge and river water. J. Appl. Bacteriol. 50, 1-9.
Westoo, A. and Peterz, M. (1992) Evaluations of methods for the detection of Listeria monocytogenes in foods: NMKE collaboration study. J. Assoc. Off. Anal. Chem. Int. 75, 46-52.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
77
Chapter 6 Media used in the detection and enumeration
of Staphylococcus aureus
R o s a m u n d M. Ba i rd a, W . H . Lee b
a Summerlands House, Summerlands, Yeovil, Somerset BA21 3AL, England, UK b USDA-FSIS, Beltsville, MD 20705, USA
Abstract
Over the years a range of selective and diagnostic media has been developed to assist in the detection and enumeration of S taphy lococcus aureurs in routine food surveillance programmes and food poisoning investigations and these are reviewed here. Baird-Parker agar remains, however, the medium of choice for direct plating and enumeration of S. aureus in both Europe and the US. This paper also reports on a comparison of the productivity and selectivity of trypticase soy broth with 10% NaC1 and 1% sodium pyruvate (PTSBS) with trypticase soy broth (TSB) for the isolation of S. aureus. Using three strains of S. aureus and a strain of S. hyicus the productivity ratio of PTSBS to TSBS ranged from -0.17 to 0.57. In the recovery of heat-injured cells of S. aureus, PTSBS offered little or no improvement over TSB with sodium chloride but no pyruvate in a three-tube MPN assessment. Of the seven other bacterial cultures tested, none grew well on the PTSBS. Selectivity ratios of 4.4-7 were found.
S t a p h y l o c o c c u s a u r e u s is a well known and important cause of food poisoning. However, both animals and man are frequent carriers (20-50%) of this bacter ium in the nasopharynx. Low numbers of S. a u r e u s in foods are generally not consid- ered a hazard and thus are not of public health concern. The growth of S. a u r e u s
to high numbers ( > 106) in foods is a hazard because about half of the S. a u r e u s
strains produce heat stable enterotoxins when present at this level (Bergdoll, 1991). The detection and enumera t ion of S. a u r e u s and its enterotoxins in foods contin- ues to be a major task in food microbiology laboratories today.

78
S. aureus food poisoning arises from the ingestion of foods containing one or more extracellular heat-stable toxins. Seven serologically distinct enterotoxins have been identified (immunological types A, B, C1, C2, C3, D and E); although the amount required to cause illness in man is not known, at least 1 ~g toxin/100 g of food will induce clinical symptoms. The syndrome is characterised by nausea, vomiting, abdominal pain and diarrhoea, 2-6 h after eating contaminated food; recovery takes 1-3 days. Time to onset and severity of symptoms depend on the quantity of toxin consumed and the individual's susceptibility. Other coagulase- positive staphylococci, namely S. hyicus and S. in termedius , produce small amounts of enterotoxin but these organisms have not yet been implicated in food-poisoning outbreaks.
Foods most frequently incriminated in staphylococcal foodborne disease include cooked poultry meat, fish (and especially crustaceans such as prawns), bakery foods (particularly those with custard or cream fillings), dairy produce (particularly dried milk) and cooked vegetables. Poor handling of food in food service establish- ments and mishandling of food in the home account for the major causes of outbreaks; few outbreaks can be traced directly to contamination during food processing. Cross-contamination of cooked food by raw food, especially by meat, can occur, but contamination from human carriers during hand-processing is more important. In terms of control, correct processing, storage and food handling is critical, since cooking will not destroy the staphylococcal enterotoxins. Susceptible food should be rapidly cooled and kept refrigerated at all times.
Many foods will support growth of S. aureus and toxin formation with the exception of those with a lower pH (< 5.0) or a water activity (a w) below 0.86. Semi-preserved products (using salt or sugar) may favour the growth of staphylo- cocci, for unlike many other organisms, staphylococci can tolerate these relatively low water activities. In foods where there is no competing spoilage flora, staphylo- coccal growth will continue unchecked, unless prevented by low storage tempera- ture.
In a large US multi-location staphylococcal enterotoxin outbreak involving sterile canned mushrooms, Bennett (1992) found that some of the canned mush- rooms contained a biologically active but immunologically inactive staphylococcal enterotoxin that was not detected by an ELISA immunoassay. Schwabe et al. (1990) also found that heat inactivated staphylococcal enterotoxins could be reactivated by high pH treatment. The 90% inactivation time of 8.3 min at 120~ for staphylococcal enterotoxin A heated in PBS buffer with 2.5% normal rabbit serum added was reported by Modi et al. (1990). These reports suggest that the staphylococcal enterotoxins have a high degree of heat resistance and that heat inactivated enterotoxins can be reactivated under certain conditions. In addition, Bergdoll (1991) cautioned that most staphylococcal enterotoxin test kits are not sensitive enough to detect a level of < 1 ng /g enterotoxins in foods which is sufficient to cause human food poisoning.
These reports emphasise the need to test for viable S. aureus before the heat processing step. If that is not possible, it may be prudent to check for indications of high numbers of dead S. aureus in heat processed foods. The presence of

79
staphylococcal thermonuclease (Schmitt et al., 1990) is easily detected in foods and may be a useful marker for this, provided staphylococcal thermonuclease has similar heat resistance to staphylococcal enterotoxins (Modi et al., 1990). It may also be possible to use direct microscopic examination (Bouwer-Hertzberger and Mossel, 1982).
1. Selective culture media available for the isolation and enumeration of S. aureus
A selection of media has been developed and used over the past fifty years for the isolation and quantitative estimation of S. a u r e u s in food samples. Since most foods contaminated with S. a u r e u s also contain large numbers of other micro- organisms, a selective medium, with enrichment if necessary, must be used. The medium must have little or no inhibitory effect on S. aureus , particularly if the cells are injured from exposure to sublethal heating or preservatives in the food. Sublethally injured cells are known to be present in many semi-preserved and preserved foods. Once repaired such cells are known to be capable of producing enterotoxins (Smith et al., 1983). Furthermore the medium must also suppress as completely as possible the growth of any accompanying flora.
Selectivity has been attained by incorporating various substances, as shown in Tables 1 and 2. In the past high concentrations of sodium chloride (75-150 g/ l ) were widely used both in solid isolation media (Chapman, 1945) and in liquid enrichment broths (Buttiaux and Brogniart, 1947; Baer 1966; Baer et al., 1966; Gilden et al., 1966). However high concentrations of sodium chloride (i.e., > 40 g/ l ) are now known to be inhibitory to sublethally injured cells (Iandolo and Ordal, 1966; Hurst et al., 1973; Gray et al., 1974; Smolka et al., 1974; Idziak and Mossel, 1980) and use of such media is no longer commonplace. Potassium tellurite and lithium chloride have also been employed as selective agents in solid isolation media (Vogel and Johnson, 1960; Crisley et al., 1965). A combination of potassium tellurite (0.5 g / l )wi th anaerobic incubation in an enrichment broth was shown to be useful in detecting low numbers of S. a u r e u s in mixed cultures and also in suppressing the growth of common Gram-negative lactose-fermenting contaminants (Giolitti and Cantoni, 1966). Polymyxin has also been used (Orth and Anderson, 1970).
Besides the addition of selective agents, other substances have been employed to enhance productivity. The addition of pyruvate (Baird-Parker, 1962) and cata- lase (Flowers et al., 1977) is known to be especially useful in the recovery of stressed cells of S. a u r e u s in both solid isolation media and liquid enrichment media (Baird and van Doorne, 1982). Both pyruvate and catalase act to prevent cell death from H 2 0 2 accumulation during aerobic growth and repair (Brewer et al., 1977). The use of pyruvate is more convenient since it can be added to the medium before autoclaving.
For diagnostic purposes, mannitol (Vogel and Johnson, 1960), egg yolk (Baird- Parker, 1962), DNA (Andrews and Martin, 1978) pig plasma (Stadhouders et al.,

80
Table 1 Selective and diagnositic systems used in enrichment media for Staphylococcus aureus
Enrichment b r o t h Selective system Diagnostic system Reference
Tryptone soya Sodium chloride Buttiaux and broth (TSB) with Brogniart (1947) sodium chloride Baer (1966) (TSBS) Gilden et al. (1966)
TSB with sodium chloride and 1% sodium pyruvate (PTSBS)
Sodium chloride Lancette et al. (1986)
Giolitti and Cantoni Potassium tellurite Potassium tellurite Giolitti and Broth (GCB) Lithium chloride Cantoni (1966)
Glycine Anaerobic incubation
Liquid Baird-Parker Potassium tellurite Potassium tellurite Baird and van (LBP) Lithium chloride Doorne (1982)
Glycine Anaerobic incubation
1976) and fibrinogen (Hauschild et al., 1979) have been used in plating media. The appearance of S. aureus colonies on various isolation media is given in Table 3.
In practice direct plating techniques are most commonly used with the above media but a most probable number (MPN) technique using Giolitti and Cantoni Broth (GCB) and liquid Baird-Parker (LBP) has also been employed to detect low numbers of organisms. A combined plating procedure using a selective overlay above a Baird-Parker (BP) agar base has also been used (Isigidi et al., 1989) (see below).
C o m p a r a t i v e studies
A number of comparative studies have examined the relative merits of the various media used in the isolation and enumeration of S. aureus. Terplan et al. (1982) compared the recovery of heat-stressed cells on four selective agars and found that BP and Baird-Parker plasma (BP with 20% v / v pig plasma) agar were superior to potassium thiocyanate actidione sodium azide egg yolk pyruvate agar (KRANEP: Sinell and Baumgart, 1967) and modified Vogel Johnson agar (PCVJ: Andrews and Martin, 1978). Likewise Baird and van Doorne (1982) compared the recovery of S. aureus from naturally contaminated foods using a direct plating technique onto either BP or Vogel Johnson (VJ) agars and an enrichment technique in either TSB or LBP, followed by plating onto BP; the best recovery was found using the combination of LBP for enrichment and BP agar for plating.
Lancette (1986) reviewed the merits of three enrichment (LBP, GCB and PTSBS) and four plating media (BP, BP without egg yolk, single-step Staphylococ- cus-selective agar (4S-agar: Mintzer-Morgenstern and Katzenelson, 1982) and PCVJ). BP agar again emerged as the medium of choice to be used as the standard
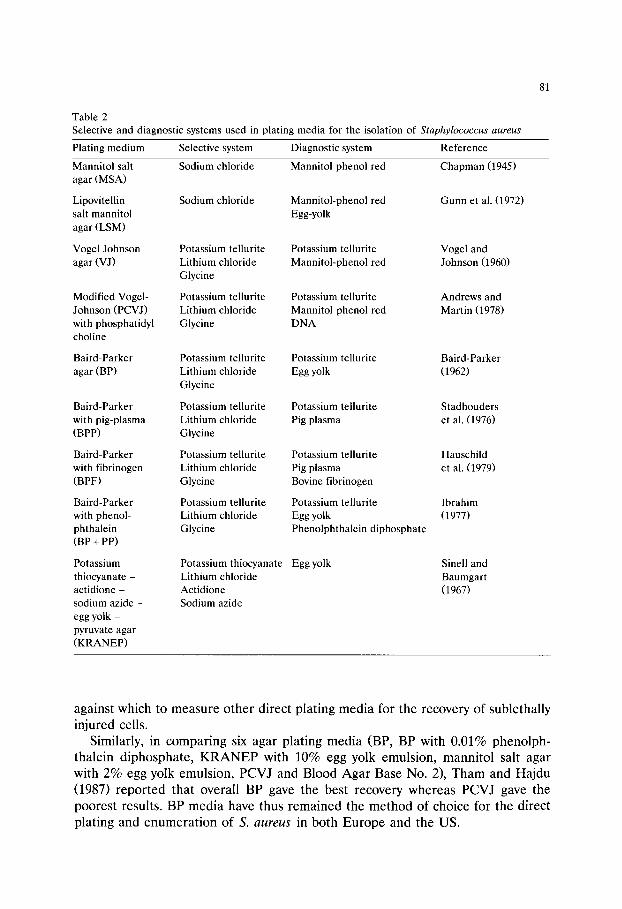
Table 2 Selective and diagnostic systems used in plating media for the isolation of Staphylococcus aureus
81
Plating medium Selective system Diagnostic system Reference
Mannitol salt Sodium chloride Mannitol-phenol red Chapman (1945) agar (MSA)
Lipovitellin Sodium chloride Mannitol-phenol red salt mannitol Egg-yolk agar (LSM)
Gunn et al. (1972)
Vogel Johnson Potassium tellurite Potassium tellurite Vogel and agar (VJ) Lithium chloride Mannitol-phenol red Johnson (1960)
Glycine
Modified Vogel- Potassium tellurite Potassium tellurite Andrews and Johnson (PCVJ) Lithium chloride Mannitol-phenol red Martin (1978) with phosphatidyl Glycine DNA choline
Baird-Parker Potassium tellurite Potassium tellurite Baird-Parker agar (BP) Lithium chloride Egg yolk (1962)
Glycine
Baird-Parker Potassium tellurite Potassium tellurite Stadhouders with pig-plasma Lithium chloride Pig plasma et al. (1976) (BPP) Glycine
Baird-Parker Potassium tellurite Potassium tellurite Hauschild with fibrinogen Lithium chloride Pig plasma et al. (1979) (BPF) Glycine Bovine fibrinogen
Baird-Parker Potassium tellurite Potassium tellurite Ibrahim with phenol- Lithium chloride Egg yolk (1977) phthalein Glycine Phenolphthalein diphosphate (BP+PP)
Potassium thiocyanate Egg yolk Sinell and Lithium chloride Baumgart Actidione (1967) Sodium azide
Potassium thiocyanate - actidione - sodium azide - egg yolk - pyruvate agar (KRANEP)
against which to measure other direct plating media for the recovery of sublethally injured cells.
Similarly, in comparing six agar plating media (BP, BP with 0.01% phenolph- thalein diphosphate, KRANEP with 10% egg yolk emulsion, mannitol salt agar with 2% egg yolk emulsion, PCVJ and Blood Agar Base No. 2), Tham and Hajdu (1987) reported that overall BP gave the best recovery whereas PCVJ gave the poorest results. BP media have thus remained the method of choice for the direct plating and enumeration of S. a u r e u s in both Europe and the US.

82
Table 3 Appearance of Staphylococcus aureus on various isolation media
Plating Colony appearance medium a
Reference
BP
BPP
BPF
B P + P P
KRANEP
LSM
MSA
VJ
PCVJ
Black, shiny, convex 1.0-1.5 mm with narrow white entire margin and surrounded by a clear zone extending 2-5 mm into opaque medium
Black, shiny, convex surrounded by a halo of precipitated fibrin
Black, shiny, convex surrounded by a halo of precipitated fibrin
As for BP, but surrounded by pale to dark pink zone of 4 mm
Yellow gold surrounded by milky turbid zone
Colony with opaque zone against yellow background
Colony surrounded by bright yellow zone
Black, convex, shiny surrounded by yellow zone
Black, convex, shiny with yellow clearing zones
Baird-Parker (1962)
Stadhouders et al. (1976)
Hauschild et al. (1979)
Ibrahim (1977)
Sinell and Baumgart (1967)
Gunn et al. (1972)
Chapman (1945)
Vogel and Johnson (1960)
Andrews and Martin (1975)
a See Table 2 for key to abbreviations.
Various modifications have, however, been made over the years in attempts to improve both recovery and selectivity of the original BP formula. Pig plasma (Devoyod et al., 1976; Stadhouders et al., 1976) rabbit and bovine plasma (Beckers et al., 1984) and fibrinogen (Hauschild et al., 1979) have been added as supple- ments but may be of variable quality (Mossel and Eelderink, 1979). Likewise the addition of sulphamethazine (Smith and Baird-Parker, 1964) and acriflavine, polymyxin and sulphonamide (Devriese, 1981) to increase selectivity may prove inhibitory to stressed S. aureus cells (Ray, 1979). Equally it is known that the selective capacity of BP agar is limited since only lipase-positive and lecithinase- positive S. aureus colonies are easily recognised. Egg yolk-negative strains require differentiation from other bacteria, such as Bacillus spp., Proteus spp., enterococci and micrococci by confirmatory tests. However, Isigidi et al. (1989) reported a two-layer plating technique for the enumeration of both egg-yolk-positive and egg-yolk-negative strains on BP agar. Following a one-hour recovery period on the basal medium, plates were overlayered with a highly selective supplemented BP agar (containing egg yolk emulsion, acriflavine, polymyxin E, potassium tellurite and sodium sulphamethazine). Both egg-yolk-positive and egg-yolk-negative strains
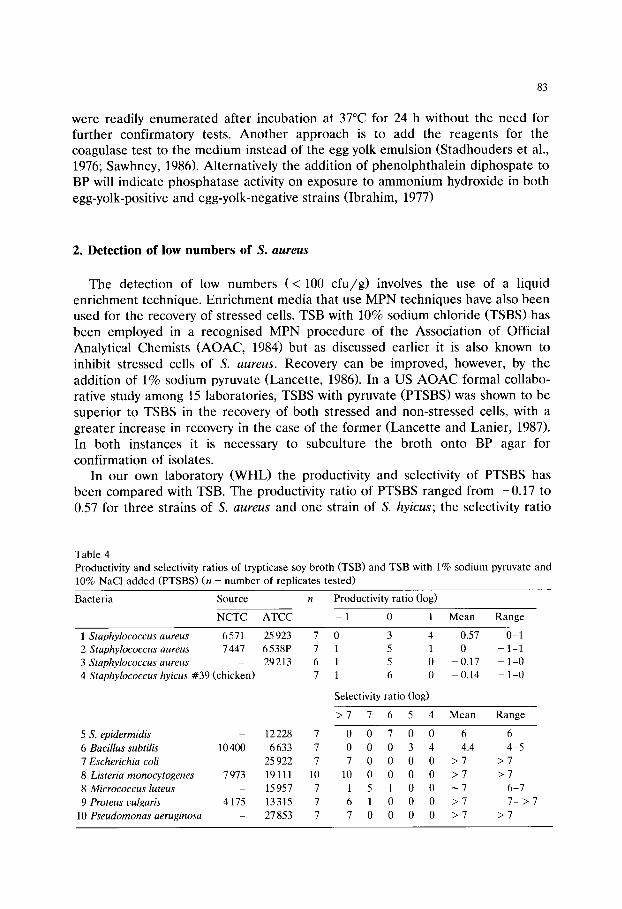
83
were readily e n u m e r a t e d after incubat ion at 37~ for 24 h without the need for fur ther confirmatory tests. A n o t h e r approach is to add the reagents for the coagulase test to the med ium instead of the egg yolk emulsion (S tadhouders et al., 1976; Sawhney, 1986). Alternatively the addit ion of pheno lph tha le in d iphospate to BP will indicate phospha tase activity on exposure to a m m o n i u m hydroxide in both
egg-yolk-positive and egg-yolk-negative strains (Ibrahim, 1977)
2. Detection of low numbers of S. aureus
The detect ion of low numbers ( < 100 c f u / g ) involves the use of a liquid enr ichment technique. Enr i chment media that use M P N techniques have also been used for the recovery of stressed cells. TSB with 10% sodium chloride (TSBS) has been employed in a recognised M P N procedure of the Associat ion of Official
Analytical Chemists (AOAC, 1984) but as discussed earlier it is also known to
inhibit stressed cells of S. aureus . Recovery can be improved, however, by the addit ion of 1% sodium pyruvate (Lancette , 1986). In a US A O A C formal collabo- rative study among 15 laboratories, TSBS with pyruvate ( P T S B S ) w a s shown to be superior to TSBS in the recovery of both stressed and non-s t ressed cells, with a
greater increase in recovery in the case of the former (Lancet te and Lanier, 1987). In both instances it is necessary to subculture the broth onto BP agar for
confirmation of isolates. In our own laboratory (WHL) the productivity and selectivity of PTSBS has
been compared with TSB. The productivity ratio of PTSBS ranged from - 0 . 1 7 to 0.57 for three strains of S. a u r e u s and one strain of S. hy icus; the selectivity ratio
Table 4 Productivity and selectivity ratios of trypticase soy broth (TSB) and TSB with 1% sodium pyruvate and 10% NaC1 added (PTSBS) (n = number of replicates tested)
Bacteria Source n Productivity ratio (log)
NCTC ATCC - 1 0 1 Mean Range
1 Staphylococcus aureus 6 571 2 Staphylococcus aureus 7 447 3 Staphylococcus aureus 4 Staphylococcus hyicus #39 (chicken)
5 S. epidermidis 6 Bacillus subtilis 10 400 7 Escherichia coli 8 Listeria monocytogenes 7 973 8 Micrococcus luteus 9 Proteus uulgaris 4175
10 Pseudomonas aeruginosa
25 923 7 0 3 4 0.57 0-1 6 538P 7 1 5 1 0 - 1-1 29213 6 1 5 0 -0.17 - 1-0
7 1 6 0 -0.14 - 1-0
Selectivity ratio (log)
> 7 7 6 5 4 Mean Range
12228 7 0 0 7 0 0 6 6 6633 7 0 0 0 3 4 4.4 4-5
25 922 7 7 0 0 0 0 > 7 > 7 19111 10 10 0 0 0 0 > 7 > 7 15 957 7 1 5 1 0 0 ~ 7 6-7 13 315 7 6 1 0 0 0 > 7 7- > 7 27853 7 7 0 0 0 0 > 7 > 7

84
Table 5 Recovery of S taphy lococcus aureus heated to 56~ for 15 min in trypticase soy broth (TSB), TSB with 10% NaC1 (TSBS), and TSB with 1% sodium pyruvate and 10% NaC1 added (PTSBS)
Heated Medium S. aureus S. aureus S. aureus
ATCC 25923 ATCC 6538P ATCC 25913 (Log cfu/ml) (Log cfu/ml) (Log cfu/ml)
No TSB 9.3 9.2 9.2 Yes TSB 5.8 7.4 8.0 Yes TSBS 3.6 7.4 7.0 Yes PTSBS 4.4 7.4 7.6 Pyruvate effect + 0 minimal
ranged from 4.4 to > 7 for seven other strains of bacteria tested, as shown in Table 4. In the recovery of heat injured cells of S. aureus (exposed to 56~ for 15 min), PTSBS offered little or no improvement over TSBS in a three-tube MPN assessment (Table 5).
LBP can be used in the recovery of stressed cells as well as for low numbers of S. aureus (Baird and van Doorne, 1982). Results can be reported at least one day earlier compared with the standard MPN method. In a trial using different isolation media, selective enrichment in LBP followed by subculture onto BP gave the highest recovery of cells. However, LBP is not widely used in the food industry, perhaps because aseptic additions of potassium tellurite and liquid paraffin must be made to each MPN tube and because high numbers of competing flora may well outgrow low numbers of S. aureus. Giolitti and Cantoni broth containing Tween 80 and pyruvate has also been used to recover small numbers of S. aureus (Chopin et al., 1985) but again the usefulness of the medium is limited by the need to add potassium tellurite to each MPN tube and also by its low selectivity.
3. Rapid S. aureus identification methods
S taphy lococcus spp., other than S. aureus, as well as other bacteria can form typical black colonies with or without halos on BP agar. Such colonies must be tested for coagulase to identify the S. aureus; commercially available kits can provide a rapid result in detecting the staphylococcal protein A as shown by agglutination of normal plasma coated latex beads. Thermostable DNase produc- tion has also been used to differentiate typical black colonies on BP agar (Lachica, 1980, 1984; Terplan et al., 1982). There is good qualitative correlation between thermonuclease and coagulase production, but not for staphylococcal enterotoxin production.

85
References
Andrews, G.P. and Martin, S.E. (1978) Modified Vogel and Johnson agar for Staphylococcus aureus. J.
Food Protect. 41, 530-532. Association of Official Analytical Chemists (1984) Official Methods of Analysis, 14th Edn. AOAC,
Arlington, VA, USA. Baer, E.F. (1966) Proposed method for isolating coagulase-positive staphylococci from food products:
report of a collaborative study. J. Assoc. Off. Anal. Chem. 49, 270-273. Baer, E.F., Franklin, M.K. and Gilden, M.M. (1966) Efficiency of several selective media for isolating
coagulase-positive staphylococci from food products. J. Assoc. Off. Anal. Chem. 49, 267-269. Baird, R.M. and van Doorne, H. (1982) Enrichment techniques for Staphylococcus aureus. Arch.
Lebensmittelhyg. 33, 146-150. Baird-Parker, A.C. (1962) An improved diagnostic and selective medium for isolating coagulase-positive
staphylococci. J. Appl. Bacteriol. 25, 12-19. Beckers, H.J., van Leusden, F.M., Bindschedler, O. and Guerraz, D. (1984) Evaluation of a pour plate
system with rabbit plasma-bovine plasma-agar for the enumeration of Staphylococcus aureus in food. Can. J. Microbiol. 30, 470-474.
Bennett, R.W. (1992) The bimolecular temperament of staphylococcal enterotoxin in thermally pro- cessed foods. J. Assoc. Off. Anal. Chem. 75, 6-12.
Bergdoll, M.S. (1991) Staphylococcus aureus. J. Assoc. Off. Anal. Chem. 74, 706-710. Bouwer-Hertzberger, S.A. and Mossel, D.A.A. (1982) Bacterioscopic examination of specimens possibly
involved in diseases of bacterial etiology transmitted by foods. In: Corry, J.E.L., Roberts, D. and Skinner F.A. (Eds.) Isolation and Identification Methods for Food Poisoning Organisms, pp. 25-33. Academic Press, London.
Brewer, D.G., Martin, S.E. and Ordal, Z.J. (1977) Beneficial effects of catalase or pyruvate in most-probable-number technique for the detection of Staphylococcus aureus. Appl. Environ. Micro- biol. 34, 797-800.
Buttiaux, R. and Brogniart, R. (1947) Techniques d'isolement des staphylocoques pathogens. Identifica- tion des staphylocoques enterotoxiques. Ann. Inst. Pasteur. 73, 830-834.
Chapman, G.H. (1945) The significance of sodium chloride in studies on staphylococci. J. Bacteriol. 50, 201-203.
Chopin, A., Malcolm, S., Jarvis, G., Asperger, H., Beckers, H.J., Berbona, A.M., Cominazzini, C., Carini, S., Lodi, R., Hahn, G., Heeschen, W., Jans, J.A., Jervis,-D.I., Lanier, J.M., O'Connor, F., Rea, M., Rossi, J., Seligmann, R., Tesone, S., Waes, G., Macquot, G. and Pivnick, H. (1985) ICMSF method studies XV. Comparison of four media and methods for enumerating Staphylococcus aureus
in powdered milk. J. Food Protect. 48, 21-27. Crisley, F.D., Peeler, J.J. and Angelotti, R. (1965) Comparative evaluation of five selective and
differential media for the detection and enumeration of coagulase positive staphylococci in foods. Appl. Microbiol. 13, 140-156.
Devoyod, J.J., Millet, L. and Hocquot, G. (1976) Un milieu gelos6 pour le d6nombrement direct de Staphylococcus aureus: milieu au plasma du porc pour S. aureus (PPSA). Can. J. Microbiol. 22, 1603-1611.
Devriese, L.A. (1981) Baird-Parker medium supplemented with acriflavine, polymyxins and sulphonamide for the selective isolation of Staphylococcus aureus from heavily contaminated materials. J. Appl. Bacteriol. 50, 351-357.
Flowers, R.S., Martin, S.E., Brewer, D.G. and Ordal, Z.J. (1977) Catalase and enumeration of stressed Staphylococcus aureus cells. Appl. Environm. Microbiol. 33, 1112-1117.
Gilden, M.M., Baer, E.F. and Franklin, M.K. (1966) Comparative evaluation of a direct plating procedure and an enrichment isolation procedure for detecting coagulase positive staphylococci in foods. J. Assoc. Off. Anal. Chem. 49, 273-275.
Giolitti, G. and Cantoni, C. (1966) A medium for the isolation of staphylococci from food stuffs. J. Appl. Bacteriol. 29, 395-398.
Gray, R.J.H., Gaske, M.A. and Ordal, Z.J. (1974) Enumeration of thermally stressed Staphylococcus
aureus MF31. J. Food Sci. 39, 844-846.

86
Gunn, B.A., Dunkelberg, W.E. and Creitz, J.R. (1972) Clinical evaluation of 2% LSM medium for primary isolation and identification of staphylococci. Am. J. Clin. Pathol. 57, 230-240.
Hauschild, A.H.W., Park, C.E. and Hilsheimer, R. (1979) A modified pork plasma agar for the enumeration of Staphylococcus aureus in foods. Can. J. Microbiol. 25, 1052-1057.
Hurst, A., Hughes, A., Baere-Rogers, J.L. and Collins Thompson, D.L. (1973) Physiological studies on the recovery of salt tolerance by Staphylococcus aureus after sublethal heating. J. Bacteriol. 116, 901-907.
Iandolo, J.J. and Ordal, Z.J. (1966) Repair of thermal injury of Staphylococcus aureus. J. Bacteriol. 91, 134-142.
Ibrahim, G.F. (1977) Detection and enumeration of coagulase positive staphylococci in dairy products. 3. Evaluation of the performance of the strip method. Aust. J. Dairy Technol. 32, 35-38.
Idziak, E.S. and Mossel, D.A.A. (1980) Enumeration of vital and thermally stressed Staphylococcus aureus in foods using Baird-Parker pig plasma agar (BPP). J. Appl. Bacteriol. 48, 101-113.
Isigidi, B.K., Devriese, L.A., Croegaert, T. and van Hoof, J. (1989) A highly selective two-stage isolation method for the enumeration of Staphylococcus aureus in foods. J. Appl. Bacteriol. 66, 379-384.
Lachica, R.V. (1980) Accelerated procedure for the enumeration and identification of food-borne Staphylococcus aureus. Appl. Environ. Microbiol. 39, 17-19.
Lachica, R.V. (1984) Egg-yolk free Baird-Parker medium for the accelerated enumeration of food borne Staphylococcus aureus. Appl. Environ. Microbiol. 48, 870-871.
Lancette, G.A. (1986) Current resuscitation methods for recovery of stressed Staphylococcus aureus
from foods. J. Food Protect. 49, 477-481. Lancette, G.A. and Lanier, J. (1987) Most probable number method for isolation and enumeration of
Staphylococcus aureus in food: collaborative study. J. Assoc. Off. Anal. Chem. 70, 35039. Mintzer-Morgenstern, L. and Katzenelson, E. (1982) A simple medium for the isolation of coagulase-
positive staphylococci in a single step. J. Food Protect. 45, 218-222. Modi, N.K., Rose, S.A. and Trantner, H.S. (1990) The effect of irradiation and temperature on the
immunological activity of staphylococcal enterotoxin A. Int. Food Microbiol. 11, 85-92. Mossel, D.A.A. and Elderink, I. (1979) A simple test for the assessment of the suitability of pork
plasma for incorporation into Baird-Parker base medium for the enumeration of Staphylococcus aureus in foods and other specimens. Lab. Pract. 28, 623.
Orth, D.S. and Anderson, A.W. (1970) Polymyxin-coagulase mannitol agar. 1. A selective isolation medium for coagulase positive staphylococci. Appl. Microbiol. 19, 73-75.
Ray, B. (1979) Current methods to detect stressed microorganisms. J. Food Protect. 42, 346-355. Sawhney, D. (1986) The toxicity of potassium tellurite to Staphylococcus aureus in rabbit plasma
fibrinogen agar. J. Appl. Bacteriol. 61, 149-155. Schmitt, M., Schuler-Schmid, U. and Schmidt-Lorenze, W. (1990) Temperature limits of growth, TNase
and enterotoxin production of Staphylococcus aureus strains isolated from foods. Int. J. Food Microbiol. 11, 1-20.
Schwabe, M., Notermans, S., Boot, R., Tatini, S.R. and Kramer, J. (1990) Inactivation of staphylococcus enterotoxins by heat and reactivation by high pH treatment. Int. J. Food Microbiol. 10, 33-42.
Smith, B.A. and Baird-Parker A.C. (1964) The use of sulphamezathine for inhibiting Proteus sp. on Baird-Parker isolation medium for Staphylococcus aureus. J. Appl. Bact. 27, 78-82.
Smith, J.L., Buchanan, R.L. and Palumbo, S.A. (1983) Effect of food environment on staphylococcal enterotoxin synthesis: a review. J. Food Protect. 46, 545-555.
Sinell, H.J. and Baumgart, J. (1967) Selektivnahrboden mit eigelb zur isolierung von pathogenen staphylokokken aus lebensmitteln. Zentalbl. Bakteriol. Parasitenk. Infektionskr. Hyg. Abt. 1: Orig. 204, 248-264.
Smolka, L.R., Nelson, F.E. and Kelly, L.M. (1974) Interaction of pH and NaC1 on enumeration of heat stressed Staphylococcus aureus. Appl. Microbiol. 27, 443-447.
Stadhouders, J., Hassing, F. and Vanaalst-Vanmaren, N.O. (1976) A pour-plate method for the detection and enumeration of coagulase positive Staphylococcus aureus in the Baird-Parker medium without egg yolk. Neth. Milk Dairy J. 30, 222-229.
Terplan, G., Zaadhof, K.-J. and Beckers, H. (1982) Quality assurance of newer media for the enumeration of Staphylococcus aureus in foods. Arch. Lebensmitelhyg. 33, 142-145.

87
Tham, W. and Hajdu, L.J. (1987) A comparison of six media for isolating Staphylococcus aureus from foods. Food Microbiol. 4, 133-146.
Vogel R.A. and Johnson, M. (1960) A modification of the tellurite-glycine medium for use in the identification of Staphylococcus aureus. Public Health Laboratory 18, 131-133.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
89
Chapter 7
Culture media for non-sporulating Gram-positive food spoilage bacteria
W . H . Holzapfel Institute of Hygiene and Toxicology, Federal Research Centre for Nutrition,
Engesserstrasse 20, D-7500 Karlsruhe 1, Germany
The spoilage association especially of protein-rich foods can be dominated by Gram-positive bacteria, notably lactic acid bacteria (LAB) which affect vacuum packaged refrigerated processed meats and some dairy products.
New food ecosystems are being created by novel packaging and processing technologies, resulting in spoilage associations differing from those previously reported. In addition, improvement in identifica- tion methods, allow the detection and isolation of 'novel' bacterial groups, e.g., Carnobacterium spp.
This review considers the genera Aerococcus, Brevibacterium, Brochothrix, Carnobacterium, Kurthia, Lactobacillus, Leuconostoc, Microbacterium, Micrococcus, Pediococcus and Propionibacterium.
Strictly selective procedures, including incubation temperature and atmosphere, are not yet available for the genera Aerococcus, Brevibacterium, Microbacterium and Micrococcus, and only with some limitations for Kurthia and Propionibacterium. On the other hand, a causative role in food spoilage has not been established clearly for all those groups, some of which may be 'opportunistic' in their behaviour.
The LAB groups Lactobacillus, Leuconostoc and Pediococcus (' LLP-Group') often share similar habitats and show similar physiological behaviour on a number of elective and selective media. Modifications to increase selectivity have been based mainly on de Man, Rogosa and Sharpe (MRS) or Rogosa agar, and include pH reduction, supplementation with chemical preservatives (e.g., sorbic acid and nitrate) and the use of reduced atmospheres or suboptimal incubation temperatures.
Carnobacteria differ from other LAB in their non-aciduric nature, and selective plating procedures use high-pH media (pH 8-9) by which competitors (mainly lactobacilli) are eliminated.
Introduction
Microbial spoilage of a food can be defined as the result of metabolic activities of one or more microbial groups, leading to adverse sensory changes in the specific product. The sum of those bacterial groups involved in spoilage is referred to as the 'spoilage association'.

90
The definition of a spoilage organism is especially difficult with respect to the lactic acid bacteria (LAB), because in some situations an organism can be used to yield desirable changes, and in others it can cause spoilage. For instance, Lacto- coccus lactis ssp. lactis can be responsible for the souring of fresh milk, but it is also used as the main component of dairy starter cultures for the manufacture of semi-hard cheese (Gouda, Cheddar, etc.). Lactobacillus sake is primarily responsi- ble for the spoilage of refrigerated vacuum packed processed meats, but is also a component of starter cultures for fermented meat products (dry and semi-dry sausages).
Within this review, the genera Aerococcus, Brevibacterium, Brochothrix, Carnobacterium, Kurthia, Lactobacillus, Leuconostoc, Microbacterium, Micrococcus, Pediococcus and Propionibacterium will be considered. Apart from Streptococcus, 'traditional' genera of the LAB include Lactobacillus, Leuconostoc and Pediococ- cus (sometimes referred to as the 'LLP-Group'). In addition, a 'new' genus Carnobacterium, formerly known as Lactobacillus divergens (Holzapfel and Ger- ber, 1983) and Lactobacillus piscicola (Hiu et al., 1984), shows phenotypic and phylogenetic relationship with Enterococcus (Hammes et al., 1992). Aerococcus seems to be more distantly related to these genera. The remaining LAB (e.g., Enterococcus, Lactococcus, Streptococcus, etc.) are considered elsewhere (see Reuter, this volume).
In spite of their fastidiousness, the LAB are of prime importance as spoilage bacteria in protein-rich and even non-animal food systems (e.g., vegetables, mayonnaises, etc.) and seem to be well adapted and competitive in a wide range of food substrates including ready-to-eat and delicatessen foods. Processing, packag- ing and refrigerated storage seem especially to favour some LAB, of which the lactobacilli often dominate the spoilage association of meat and vegetable systems (Reuter, 1985; Hammes et al., 1992) followed by Leuconostoc, Pediococcus and Carnobacterium spp. (Holzapfel and Schillinger, 1992). Among the other groups Brochothrix thermosphacta and Microbacterium lacticum in particular may play some role as spoilage bacteria in special situations (Gardner, 1981; Keddie and Jones, 1981 b).
The other genera appear to be of minor importance as spoilage bacteria, but will be included for completeness. The association of the respective bacterial groups with the spoilage of particular food products, is summarised in Table 1, including reference to the literature.
Commercial media
Most elective media and some selective media are commercially available in dehydrated form. Elective media satisfy the general nutritional requirements of one or more groups of bacteria, (e.g., de Man Rogosa and Sharpe (MRS) or Briggs agar for LAB (Baird et al., 1987; Curtis et al., 1993)) without completely inhibiting other groups (e.g., Micrococcus and Bacillus spp.). They normally allow higher recov- ery rates of the particular group(s) sought, but, because of their lower selectivity,

TABLE 1
Gram-positive non-sporulating bacteria associated with food spoilage (the numbers refer to literature references in the footnote; those in brackets reflect doubt with respect to the importance in spoilage)
Bacterial Group Product group
Dairy Meat Vegetable
Fresh Pro- Beverages Cheese Fresh milk cessed (ferm.)
Pro- Fer- Poultry Fresh Pro- Fer- cessed mented cessed mented
Alc. Fish Deli- beverages cates- (Wine/ sen Beer)
Other
Aerococcus viridans
Brevibacterium (2) Brochothrix (6) Carnobacterium Kurthia (13) Lactobacillus : 15
hetero (16) homo 16
Lactococcus 24 Leuconostoc 25 Microbacterium
(lacticum) 31 Micrococcus 25 Pediococcus Propioni-
bacterium
(1) 2 (4) 3
5 5 5 (6) 9/10 10/11 11 8
12 12 (13) 15 15 15 15 17 15 (15) (15) 21 (15)
16 (18) 18/19 17 18 (15) 15 22 16 18 18/19 17 18 (15) 22
4 7
(12)
20 23 15
24 30 26 19 30 8 27 28 29 30
31 31
(33)
(32) 32
Literature: 1, Deibel and Niven (1960); 2, Collins et al. (1983); 3, Cantoni et al. (1969); 4, Sharpe et al. (1977); 5, Gardner (1981); 6, Keddie and Jones (1981a); 7, Nickelson et al. (1980); 8, Thornley and Sharpe (1959); 9, Holzapfel and Gerber (1983); 10, Shaw and Harding (1984); 11, Borch and Molin (1988); 12, Hiu et al. (1984); 13, Gardner (1969); 14, Keddie (1981); 15, Hammes et al. (1992); 16, Fryer et al. (1970); 17, Coretti (1958); 18, Reuter (1975); 19, Von Holy and Holzapfel (1989); 20, McFeeters et al. (1984); 21, Dittrich (1977); 22, Back (1981); 23, Baumgart et al. (1983); 24, Mundt (1982); 25, Keller et al. (1987); 26, Sharpe and Pettipher (1983); 27, Vaughn (1985); 28, Rankine (1977); 29, Hatcher et al. (1984); 30, Holzapfel and Schillinger (1992); 31, Collins and Bradbury (1992); 32, Weiss (1992); 33, Cummins and Johnson (1992).

92
their 'selective' application is limited to spoilage associations typically dominated by one type or group of organisms, e.g., vacuum-packaged processed meats.
A selective medium typically contains one or more restrictive factors by which unwanted bacterial groups in a mixed population are inhibited but which still allow growth of the group(s) to be detected (e.g., Rogosa agar for LAB (Baird et al., 1987)); the medium contains the necessary growth factors even for fastidious groups. Several selective media only find application for specific food substrates or selected bacterial groups or species, and are not necessarily commercially avail- able. This applies especially to media to which one or more inhibitory factors are to be added, or to those requiring fortification, e.g., with fresh plant, vegetable or yeast extracts.
Catalase-positive groups
The presence of a cytochrome system is typically associated with aerobic bacteria, and allows the transfer of electrons during respiration. Toxic peroxide or superoxide radicals may be produced during aerobic growth but are detoxified by specific enzymes such as catalase. With the exception of Propionibacterium spp., which prefers an anaerobic or microaerophilic environment, the catalase-positive groups grow well under aerobic conditions. The groups of concern here, are the non- LAB: Brevibacterium, Brochothrix, Kurthia, Microbacterium and Micrococcus spp.
Brevibacterium
Brevibacteria do not represent a major spoilage group of foods. They can be associated with different food products, including cereals (Baumgart, 1990) milk and milk products (Collins et al., 1983) hams (Cantoni et al., 1969) and fish (Jones and Keddie, 1986), They grow well on most universal nutrient media and elective media. When present as a minor group, enrichment procedures can be applied.
Enrichment All strains of brevibacteria are able to produce methanethiol from L-methionine,
this feature forms the basis of an enrichment procedure described by Sharpe et al. (1977). The sample is incubated at 20-25~ in a mineral-mix medium containing (g/l): KzHPO4, 2.0; KHzPO4, 1.0; CaCI2" 2H20, 1.0; MgCI2" 6H20, 1.0; FeC12 �9 6H20, 0.01 and DL-methionine, 5.0. At different stages the sample is plated or streaked onto one of several non-selective media supporting brevibacterial growth.
Selective plating No strictly selective media are available but generally tryptone soya agar
supplemented with 4.0% NaC1 (TSAS) is used. It is based on commercially available tryptone soya broth and has the following composition (g/l): tryptone (Oxoid), 17.0; soya peptone (Oxoid), 3.0; NaC1, 9.0; KzHPO4, 2.5; glucose 2.5; agar, 12.0; pH 7.0.

93
Isolation is either by direct plating of a sample, or may follow enrichment procedures (Collins, 1992). Incubation for food samples is at 20-25~ for 5-7 days and should be in the light, typically inducing production of an orange pigment by B. linens (Jones and Keddie, 1986), although further tests on isolated colonies are still necessary for final identification. Apart from TSAS, Blood agar Base No. 2 (Difco) may be used for fish samples.
Brochothrix
Originally decribed by McLean and Sulzbacher (1953) as Microbacterium ther- mosphactum, this species was transferred to a new genus, Brochothrix, as Bro- chothrix thermosphacta (Sneath and Jones, 1976). A second species, B. campestris, was described by Talon et al. (1988), and differs from the former by its sensitivity to 8% NaC1 and 0.05% potassium tellurite, and its ability to hydrolyse hippurate. Although some uncertainty still exists about its natural habitat (Jones, 1992) indications are that Brochothrix spp. originate in soil and faeces from where the organisms are introduced into slaughterhouses and other food manufacturing environments (Skovgaard, 1985). B. thermosphacta is typically associated with meat and meat products, and especially in spoilage of prepacked meats and refrigerated meat products (Gardner, 1981). More recently, its association with fish and fish products has been studied (Nickelson et al., 1980). Both species have frequently been isolated by selective procedures from sources such as soil, grass, hay and faeces, and B. thermosphacta also from different vegetables, salads and milk products (Gardner, 1981).
Enrichment Not typically performed, but questions remain open about recovery of sub-
lethally damaged cells on selective media.
Selective plating The streptomycin-thallous acetate-actidione (STAA) agar medium suggested
by Gardner (1966) is still the medium of choice (see this volume for recipe).
Incubation At 22~ for up to 5 days, colonies visible after 48 h.
Selectivity Lactobacilli, streptococci, bacilli, coryneforms and microbacteria are unable to grow on the medium. A few pseudomonads are occasionally found, but are easily recognised by colony form and the use of Kovacs oxidase reagent (Gardner, 1966, 1985).
The selectivity is based on the high concentration of streptomycin sulphate, in addition to actidione and thallous acetate, which inhibits many Gram-negative and some Gram-positive bacteria, especially the coryneform bacteria morphologically resembling Brochothrix spp. (Skovgaard, 1985). Wilkinson and Jones (1977) using a

94
disc diffusion method reported one out of 25 strains to be sensitive to strepto- mycin. Thallous acetate and actidione serve to inhibit practically all yeasts, and many aerobic and facultatively anaerobic bacteria, with the exception of strepto- cocci and lactobacilli (Gardner, 1966; Skovgaard, 1985). STAA is therefore not absolutely selective, and difficulties may be encountered when only low numbers of Brochothrix spp. are present, as in faeces (Gardner, 1966).
Modifications of STAA Talon et al. (1988) supplemented the original STAA medium with nalidixic acid
(5 ~g/ml) and oxacillin (5/~g/ml) for isolation of both species from soil, grass, etc. Another modification of STAA has been used by Hechelmann (personal communi- cation) for enumeration of Brochothrix spp. in meat and meat products, and is based on blood agar base (Merck), supplemented with (g/l): yeast extract, 2; KzHPO4, 1; MgSO4" 7H20, 0.8; NazCO 3, 0.35; inositol, 10; and a 0.3% solution of neutral red (10 ml) as indicator. The medium is prepared in 1000 ml of distilled water and the pH adjusted to 7.0. After autoclaving 0.5 g of filter-sterilised streptomycin sulphate is added per litre. The selective agent is streptomycin and Brochothrix species ferment inositol to give pink colonies.
Kurthia
Relatively limited information is available on the occurrence of Kurthia spp. in the environment, but reports indicate that they are regularly isolated from meat and meat products, in addition to faeces and some clinical materials. Typically, Kurthia zopfii and K. gibsonii are associated with off-flavours and taints of fresh and cured meat products stored at elevated temperatures, and the psychrophilic K. sibirica with refrigerated samples (Keddie and Jones, 1992).
Enrichment This is not often practised, but partial enrichment of K. gibsonii is achieved by
preliminary enrichment in YNB (yeast extract nutrient broth) (see below for recipe) and incubation at 45~ for 24 h, followed by a second subculture at 45~ prior to inoculation onto YNG and YNA plates (Keddie and Jones, 1992: see below for recipes).
Selective plating Isolation methods rely on the unusual cultural properties of K. zopfii and K.
gibsonii, and involve either a (gelatin or agar) streak method or direct plating, on yeast nutrient gelatin medium (YNG) and YNA (Keddie, 1981; Keddie and Jones, 1992).
YNG (g/l): meat extract, 4; peptone, 5; yeast extract, 2.5; NaC1, 5; gelatin, 60. The pH is adjusted to 7.0, before sterilisation (of quantities up to 100 ml) at 115~ for 30 min. YNG plates are heavily inoculated by a single central streak of the sample and incubated at 20~ (with lids uppermost) and inspected daily for

95
filamentous outgrowths and gelatin liquefaction. A pure culture is obtained by streaking a small piece of gelatin with outgrowths onto YNA, usually after 4-5 days. YNA is similar to YNG, but contains 15 g of agar for solidification instead of gelatin.
Isolation by direct plating onto YNA may prove successful if Kurthia spp. constitute a significant proportion of the population, as may be the case for meat and meat products (Gardner, 1969).
For K. sibirica similar isolation methods may be used, in combination with low incubation temperatures, and taking into account the typical 'bird's feather' growth on YNG (Keddie and Jones, 1992).
Critical factors It should be kept in mind that the above isolation media are not strictly
selective. Careful observations are therefore necessary, e.g., of typical filamentous, or feathery growth in gelatin media, and also of 'medusa' head appearance of young colonies under low magnification, and rhizoid growth or even granular appearance on YNA (Keddie and Jones, 1992).
The quality and brand of gelatin may decisively influence the extent and character of outgrowth. To overcome the limiting effect of poor quality gelatin, YNG may be dissolved in mineral base E (Owens and Keddie, 1969) to give MYNG (Shaw and Keddie, 1983; Keddie and Jones, 1992). Overgrowth with fungi may be prevented by addition of 10 units of nystatin (Squibb) per ml of molten YNG (Keddie and Jones, 1992).
Microbacterium
Microbacterium lacticum appears to be the only species in the genus of impor- tance in foods, and is mainly associated with milk and dairy products and the dairy environment (Keddie and Jones, 1981b). Its thermoduric nature explains its survival in pasteurised milk (63~ for 30 min or 72~ for 15 s) (Abd-el-Malek and Gibson, 1952) and forms the basis for isolation procedures from dairy products. Laboratory pasteurised (63~ for 30 min) samples are plated onto non-selective media such as yeast extract milk agar (YMA) (Harrigan and McCance, 1976; Collins and Bradbury, 1992), and incubated at 30~ for up to 7 days. Cheese and butter samples are initially homogenised in 2% sodium citrate solution (Collins and Keddie, 1986).
Barnes and Corry (1969) reported the association of M. lacticum with the spoilage of egg albumen. Organisms phenotypically resembling microbacteria have been isolated from fresh beef, poultry giblets and raw and pasteurised eggs (Collins and Bradbury, 1992).
Micrococcus
Animal and dairy products may be considered as secondary sources of micrococ- ci (Keller et al., 1987; Kocur et al., 1992). This group does not appear to play an

96
important role in food spoilage, but is more or less ubiquitous, and often associ- ated with food or human environments (e.g., skin of man and animals). Micrococci surving in beer may probably be strains of M. kristinae, M. varians and M. luteus (Kocur et al., 1992).
Isolation procedures have been developed and are applied mainly to micrococci of human and mammalian skin. P agar is often used as a general isolating medium (Naylor and Burgi, 1956). FTO (furoxone trypticase oil red) agar permits the selective isolation of micrococci in presence of staphylococci, the latter being inhibited by nitrofuran (Furoxone) (Curry and Borovian, 1976).
Propionibacterium
The 'classical' propionibacteria are associated with dairy products and the dairy environment, especially cheese (Cummins and Johnson, 1992) but rarely dominate a spoilage association. Their presence in Emmental-type cheeses has been well studied, and has also been reported for fermenting olives (Cummins and Johnson, 1992).
Isolation procedures Most isolation methods are based on the ability of propionibacteria to ferment
lactate and their anaerobic to micro-aerophilic character. The yeast extract sodium lactate (YEL) medium of Malik et al. (1968) is generally used for isolation and maintenance of food and dairy associated propionibacteria, and contains (g/l): trypticase, 10; yeast extract, 10; sodium lactate, 10; KzHPO4, 2.5; MnSO4, 0.005 and agar, 15; in distilled water, with the final pH adjusted to 7.0. Different methods have been devised for obtaining anaerobic growth conditions, e.g., by using a plastic pouch, an agar overlay (Malik et al., 1968) or the addition of 0.5% sodium sulphite coupled with a overlay of paraffin oil (Demeter and Janoschek, 1941) or even by using candle oats jars (Vedamuthu and Reinbold, 1967). A recommended isolation method is to supplement YEL with 0.05% of cysteine and 0.05% of Tween 80, and to incubate the plates in a Brewer type anaerobic jar containing 10-20% CO 2 (Cummins and Johnson, 1992). Incubation temperatures range from 30-35~
Trypticase yeast extract glucose medium supports good growth of all Propioni- bacterium spp, and has the following composition (Johnson and Cummins, 1972) (g/l): trypticase, 10; yeast extract, 5; glucose, 10; CaC12, 0.02; MgSO 4, 0.02; NaC1, 0.02; Tween 80, 0.5; sodium formaldehyde sulfoxalate (Eastman Organic Chemi- cals), 0.5; NaHCO 3 (added as sterile solution at time of inoculation), 1.0; and 0.05 M of equimolar mono and dibasic potassium phosphate buffer. The final pH is adjusted to 7.0.
In an earlier study the suitability of four recommended media for the isolation and enumeration of propionibacteria from dairy products was evaluated (Britz and Holzapfel, 1973; Britz, 1975). None of these media, i.e. YEL (Malik et al., 1968), lactate acetate agar (LAA) (Nieuwenhof et. al., 1969), thioglycollate lactate agar (TLA) (Kamber et al., 1952) and Brewer's anaerobic agar 279 (BAA) (Niethammer

97
and Hitzler, 1960) proved satisfactory as selective medium for this purpose. The addition of 0.3% sodium azide to YEL improved selectivity to a satisfactory level (Britz, 1975).
Catalase-negative groups
Mainly represented by the LAB, these groups generally represent nutritionally more fastidious bacteria than the catalase-positive groups discussed earlier. Yet, the LAB as a whole can be considered a ubiquitous group, and representatives are associated with extreme habitats and environments, ranging from aerobic to strictly anaerobic atmospheres, the gastro-intestinal tract and vagina to plant leaf surfaces, from fermented food products of plant and animal origin to psychrotrophically spoiled meat, and from curing brines to secondary (malolactic) fermenting wines.
In general, the LAB are relatively resistant to inhibitory factors such as chemical preservatives (e.g., potassium sorbate) and curing salts (nitrite). These and other features, e.g., acid resistance (generally to pH 4.0-4.5) and general preference of anaerobic conditions, are taken into consideration in the formulation of selective and elective media for LAB, and in the modification of 'universal' LAB media such as MRS. The 'LLP-group' (Lactobacillus, Leuconostoc and Pediococcus) will be discussed together, since they frequently share common habitats, and their general physiological features allow their growth on the same selective media.
Aerococcus
The aerococci show some physiological and morphological resemblance to the pediococci, and are catalase negative or weakly catalase positive. Their importance in food spoilage is doubtful. The sparse information, however, may be due to the lack of sufficiently selective media for their detection, and possible incorrect identification of isolated colonies recovered from selective media for LAB. In addition, they may be overlooked in many isolation procedures because of their comparatively slower and weaker growth (Evans, 1986) and their frequent appear- ance as off-white pin-point colonies. Aerococcus strains can be isolated on a number of selective and elective media, generally employed for other Gram-posi- tive cocci. Blood agar supplemented with 0.001% potassium tellurite and 0.00025% crystal violet is recommended as selective medium (Evans, 1986). For culture maintenance APT (this volume) or tryptic soy broth give satisfactory results (Weiss, 1992).
Carnobacterium
The first representative of this relatively new genus was described as Lactobacil- lus divergens (Holzapfel and Gerber, 1983), suggesting 'deviating' physiological characteristics from other lactobacilli, e.g., inhibition by pH < 6.0 and acetate, their optimal growth at pH 8-9, and their inability to grow on Rogosa agar. The

98
original habitat of carnobacteria is not precisely known, but Carnobacterium divergens and some strains of C. piscicola (syn. C. carnis) are typically associated with meat and meat products and are probably similar to 'atypical' spoilage lactobacilli earlier isolated from chicken meat (Thornley and Sharpe, 1959; Barnes, 1976). They have also been isolated from refrigerated, vacuum-packaged unpro- cessed beef, pork and lamb (Hitchener et al., 1982; Shaw and Harding, 1984; Schillinger and Lficke, 1986) and CO2-packed pork (Borch and Molin, 1988). Some strains of C. piscicola appear to be fish pathogens (e.g., of trout and salmon) (Hiu et al., 1984), whilst two more species, C. gallinarum and C. mobile are associated with poultry (Collins et al., 1987). Two psychrotrophic species, C. funditum and C. alterfunditum, recently isolated from an antarctic lake (Franzmann et al., 1991) are probably not associated with food.
Isolation procedures On account of their non-aciduric nature, carnobacteria may be isolated directly
onto elective or non-selective media, from populations where either they or lactobacilli dominate, as may be the case for vacuum-packaged refrigerated (non- processed) meat or poultry meat, or for diseased fresh-water fish. Either streaking or standard plating procedures may be applied. Adjustment to elevated pH-values of 8-9 serves to inhibit lactobacillus growth and promote the development of carnobacterial colonies (Holzapfel and Long, 1984: unpubl, results).
For the non- or semi-selective recovery of carnobacteria from samples as described above, the following media may be used (Hammes et al., 1992): (1) Standard I nutrient agar (Merck), sucrose substituted for glucose, pH 8.0; (2) CASO-Agar (Merck), pH 7.5-8.0 (glucose should preferably be substituted by sucrose); (3) D-MRS (Bosch and Holzapfel, 1985, unpubl, results) a modified MRS-agar (de Man, Rogosa and Shape, 1960) from which acetate is omitted, glucose is substituted by sucrose and the pH adjusted to 8.5 before autoclaving at 121~ for 10 min. Carnobacterial colony growth is stimulated by increasing the MnSO 4 �9 7H 2 O concentration to 0.4% (Bosch and Holzapfel, 1985, unpubl, results); (4) Tryptic soy agar (TSA, Difco) supplemented with 0.3% yeast extract and adjusted to pH 7.5-8.0; (5) Tryptone glucose yeast extract agar (TGE, Oxoid) (pH 7.0) has been used by Borch and Molin (1988) for recovery of carnobacteria as part of the total count of refrigerated, prepacked meat and meat products.
Incubation should be at 25-28~ for 3 days, in either aerobic or reduced gas atmospheres (e.g., Anaerocult, Merck).
For selective detection and isolation of carnobacteria as a minor spoilage group cresol red thallous acetate sucrose agar (CTAS) has been suggested (this volume). Following conventional plating or streaking procedures, incubation is either at 30~ for 24-48 h or at 25~ for 3-4 days.
The selectivity of CTAS is related to its high pH (8.5 to 9.0), the presence of thallium acetate and nalidixic acid, and a relatively high concentration of sodium citrate. The recovery especially of C. divergens, from meat samples is unsatisfac- tory, and colonies often develop only to pin-point size or not at all. This may be overcome by reduction of the thallous acetate concentration to 0.5 g/1 or less,

99
which, however, may allow the growth of some leuconostocs and enterococci, which are not totally inhibited by CTAS. Substitution of sucrose by 2% inulin may be used to distinguish C. piscicola colonies from enterococci which do not ferment inulin. Strains of C. piscicola are characterised by small, bronze-metallic shiny yellowish to pinkish colonies, causing a yellow colour change of the medium and clearance of the precipitate. C. divergens strains show more sparse (pin-point colony) growth, with bronze-metallic sheen but often without colour change of the medium. Most enterococci produce yellow colour change and clearance of the precipitate, but without any metallic sheen. Growth of Listeria or Leuconostoc spp. is usually not observed, and phase contrast microscopy is an important tool to distinguish between colonies of rod-shaped cells (carnobacteria) and cocci (enter- ococci). When sucrose is substituted by inosine, especially C. piscicola strains often grow as dry, umbonate ('/3-type') colonies.
Restrictive factors The pH of 8-9 inhibits the growth of most lactobacilli, and especially Lacto-
bacillus curvatus and L. sake, associated with meat and processed meat spoilage. With the exception of enterococci, accompanying bacteria, such as Listeria spp., some leuconostocs and all Gram-negative bacteria, are inhibited by thallium acetate in combination with nalidixic acid.
Attempts to improve the selectivity of CTAS against enterococci have not been successful thus far. Tests on more than 60 different antibiotics showed similar resistance spectra for carnobacteria and enterococci, and the same was found for a number of heavy metal salts (Bosch and Holzapfel, 1985, unpublished results).
Lactobacillus, Leuconostoc and Pediococcus (LLP)
These genera often share similar habitats (e.g., fermenting plant materials, meat and dairy products) and show relatively similar growth requirements and physiolog- ical characteristics, and will therefore be discussed as one group under this heading. Quite pronounced differences, however, exist even within a genus in relation to resistance to reduced pH, acid, aerobiosis/anaerobiosis and inhibitory factors. Media for their isolation must therefore take into account their aciduric or acidophilic nature and their adaptation to specific environmental conditions: e.g., Leuconostoc oenos in wine and its adaptation to pH-values around 4.5; Pediococcus damnosus and other Pediococcus spp. resistant to hops in beer and brewery habitats; slime-producing strains of Leuconostoc mesenteroides ssp. mesenteroides in sugar refineries; the adaptation of Lactobacillus sake to (fermented) meat habitats and L. plantarum to vegetable and plant habitats.
The elective and selective media for lactic acid bacteria (including enterococci and lactococci) have been excellently reviewed by Reuter (1985). A number of these elective (APT, Briggs, MRS) and selective media (LaS, MRS-S, Rogosa and Rogosa 6.2) have been included as monographs in this volume.
In this review the main aspects presented by Reuter (1985) are summarised and amended with relevant information (see Tables 2 and 3). The composition of a representative number of media is presented in Table 4.

TABLE 2
Elective and selective media for isolation of lactic acid bacteria from food (modified from Reuter, 1985; Hammes et al., 1992)
Medium pH Abbre- Literature Application(s) b viation
Recovery c
Elective A ll-p urpose A gar with
Tween 6.7 APT Briggs-Agar 6.8 BRIGGS Modified Homohiochi 5.4 MHM
Medium L actob acillus-A gar 6.8 La de Man Rogosa, Sharpe- 6.4 MRS
Agar Brain-Heart-Infusion- 6.8 BHIYE
Yeast-E xtract-Agar LS-Differential-Agar 6.1 LSD Tryptose-P roteose-P-Yeast 6.8 TPPY
extract-Eriochrome- Agar
M 16-Agar 5.6 M 16 M 17-Agar 7.2 M 17
Rogosa-Agar (modified) 6.2 RA 6.2
Selective Acetate-Agar 5.6 AA MRS 5.5 5.5 MRSS Acidic Tomato Broth 4.8 ATB MRS + 0.1% sorbate 5.7 MRSS
Evans and Niven (1951) a Briggs (1953) a Kleynmans et al. (1989)
Reuter (1970b) de Man et al. (1960) a
Baird and Patterson (1980)
Eloy and Lacrosse (1976)a Braquart, 1981
Terzaghi and Sandine (1975) Terzaghi and Sandine (1975) a
a
Cavett (1963) Baird and Patterson (1980)a Garvie (1967) ISO-Discussion (1983) a
cured meats/general milk products, gut Lb/general general/vegetables/sour dough/
sake wine general general
meat lactics/general
yoghurt yoghurt/general
yoghurt
ferm. milk products general
psychr, meat lactics general/meats wine meats/general
Lb, Lc, Lt, (Pd), Cb Lb, Lb-t, Lc, Pd, Lt, Et, Cb Lb-hetero; lactics
Lb, Lc, Lt, Pd, (Cb?), Ec most lactics
most lactics
most lactics most lactics
Lb-t, lactics
Lt most lactics
Lb, Lc Lb (Lc?) Lc (oenos), Pd Lb, Lc

M R S + 0 . 1 % thallous 6.5 MRST Mol et al. (1971) general acetate
L actobacillus-A gar + 0.04% sorbate 5.0 LaS Reuter (1970b) a meats
Nitrite, Actidione- 5.5 NAP Davidson and Cronin (1973) meat lactics Polymyxin-Agar
Homoferm.-Heteroferm.- 7.0 HHM McDonald et al. (1987) fermented vegetables Differential Medium
Kefiranofaciens-Medium 5.5 KPL Toba et al. (1986) kefir Orange-Serum-Agar 5.5 OSA Murdock et al. (1952) fruit juices
Sucrose Agar for Brewery 6.2 SAB Boatwright and Kirsop beer Isolates (1976)
MRS + Beer 4.7 MRS-B Back (1978) beer NBB (Commercial only) NBB Back (1980) beer "Isolation Medium" 9 IM Villar et al. (1985) salted anchovies, soy sauce Rogosa-Agar 5.5 RA Rogosa et al. (1951) ~ general Tomato Juice Agar 5.0 TJA Yoshizumi (1975) wine Tetrazolium-Sucrose-Agar 6.0 TS Cavett et al. (1965) frozen peas and vegetables Thallous-Acetate-Tetra- 9 TATS Cavett et al. (1965) plant materials
zolium-S ucrose-Agar Fructose and Tween 80- 5.2 FT Cavin et al. (1989) wine
Medium Leuconostoc Mesen- 7.8 LMM Vrbaski et al. (1988) plant foods
teroides-Medium
most lactics at pH 6.5
Lb (Lc) Lb, Lt, (Lc)
homoferm./heteroferm, lactics
Lb. kefiranof aciens Lb, Lc, P d
Lb, Lc, P d
Pd beer lactics Ped. halophilus Lb, Lc, P d Lb (slow-growing) Lc, Lt, Ec Lc
Lc. oenos
Lc. mesenteroides ssp. mesenteroides
a Monographs in this volume. b With respect to food systems. c Lb, Lactobacillus; Lb-t, thermophilic lactobacilli; Lc, Leuconostoc; Pd, Pediococcus; Lt, Lactococcus; Ec, Enterococcus; Cb, Carnobacterium; Lactic
lactic acid bacteria (Lactobacillus, Lactococcus, Leuconostoc, Carnobacterium, Pediococcus, Streptococcus spp.); hetero = heterofermentative.

102
TABLE 3
A summary of media used for selective isolation of lactic acid bacteria with reference to particular food product groups (see Table 2 for explanations and key to abbreviations)
General MRS pH 5.5 (Baird and Patterson, 1980) MRSS (0.1% sorbate) (ISO-discussions 1983/86) a
MRST (0.1% thallous acetate) (Mol et al., 1971) RA (Rogosa et al., 1951) a
Meat AA pH 5.6 (Cavett, 1963) MRS 5.5 a MRSS a
LaS (0.04% sorbate, pH 5.0) (Reuter, 1970) a NAP (nitrite, polymyxin) (pH 5.5) (Davidson and Cronin, 1973)
Plant foods, vegetables, fruit juices HHM (pH 7.0)(McDonald et al., 1987) OSA (pH 5.5) (Murdock et al., 1952) TS (pH 6.0) (Cavett et al., 1965) TATS (Cavett et al., 1965) LMM (pH 7.8) (Vrbaski et al., 1988)
Wine ATB (pH 4.8) (Garvie, 1967) TJA (pH 5.0)(Yoshizumi, 1975) FT (pH 5.2) (Cavin et al., 1989)
Beer SAB (pH 6.2) (Boatwright and Kirsop, 1976) MRS + Beer (pH 4.7) (Back, 1978)
Salted anchovies, soy sauce 'IM' (Villar et al., 1985)
a Monographs in this volume.
Enrichment
Enrichment is commonly practised by supplementing a basal (acetate contain- ing, etc.) medium with the original substrate. Vital growth factors may thus be supplied for fastidious strains (e.g., tomato juice or vegetable or fresh meat extracts) and adaptation to the particular substrate accounted for.
When LLP are present as a minor group within a spoilage association their comparative aciduric nature and preference for microaerophilic to anaerobic conditions should be exploited. Special features for selective isolation of one group in presence of other organisms, particularly LLP, include: the relatively high resistance of leuconostocs to thallous acetate (Reuter, 1970a); the more aciduric nature of most homo- and facultatively heterofermentative lactobacilli, compared to leuconostocs; the resistance of several pediococci to hops extract; the aciduric nature of Leuconostoc oenos compared to other Leuconostoc spp.; the psy-

103
TABLE 4
Composition of representative elective and selective media for isolation and detection of lactobacilli, leuconostocs and pediococi in foods
Values are given in per cent (w/v) of each ingredient; 1.5% agar is added unless stated otherwise. To improve anaerobiosis, 0.05% cysteine hydrochloride (or 0.1% sodium ascorbate in some cases) may be added. Sterilisation is generally at 121~ for 15 min (exceptions, e.g., RA). For abbreviations of media and literature references see Table 2.
Ingredients Elective Selective
APT BRIGGS MHM La MRS ATB NAP OSA RA SAB
Glucose 1.0 2.0 0.5 Maltose 0.2 Fructose 0.5 N-Gluconate 0.2 Peptone 1.0 0.8 1.0 Peptonised milk Meat Extract 0.2 Yeast Extract 0.5 0.6 0.7 Tween 80 0.1 0.1 0.1 K 2 HPO4 0.5 KHzPO 4 MgSOa '7H20 0.08 0.02 MnSO4.4H20 0.014 0.005 FeSO 4. 7H 2 ~ 0.004 0.001 Na-acetate 0.5 Na-azide
Na2CO3 CaCO 3 NaNO 2 Na-citrate 0.5 NHa-citrate 0.2 Acetic acid Actidione Polymyxin B NaC1 0.5 0.5 Tomato Juice 40 Bromocresol
green Soluble starch 0.05 Sucrose Mevalonic acid
lactone 0.003 Orange Extract pH original 7.0 5.0 5.4 pH modified 6.7 6.8
2.0 2.0 1.0 0.75 0.4 2.0
1.0 1.0 1.0 1.0 1.0 1.0
1.0 1.0 0.25 0.5 0.5 0.5 1.0 0.3 0.1 0.1 0.1 0.2 0.3
0.02 0.058 0.2 0.0575 0.005 0.025 0.005 0.005
0.05 0.5
0.3 0.2
25
0.006
1.0 1.0
0.5 0.5 0.1 0.1
0.6 0.0575 0.05 0.012 0.05 0.0034 2.5
0.3
0.2 0.132
0.0001 0.01 0.00003
0.5 6.8 6.4 4.8 5.5 5.3 5.8
6.2 6.2
0.5
0.002
5.0
c h r o t r o p h i c n a t u r e o f t y p i c a l m e a t s p o i l a g e l a c t o b a c i l l i (e .g . , Lactobacillus sake) a n d l e u c o n o s t o c s (e .g . , Leuconostoc carnosum a n d Leuconostoc gelidum); t h e
t h e r m o p h i l i c n a t u r e o f m o s t o b l i g a t o r y h o m o f e r m e n t a t i v e l a c t o b a c i l l i c o m b i n e d
w i t h t h e i r a c i d t o l e r a n c e .
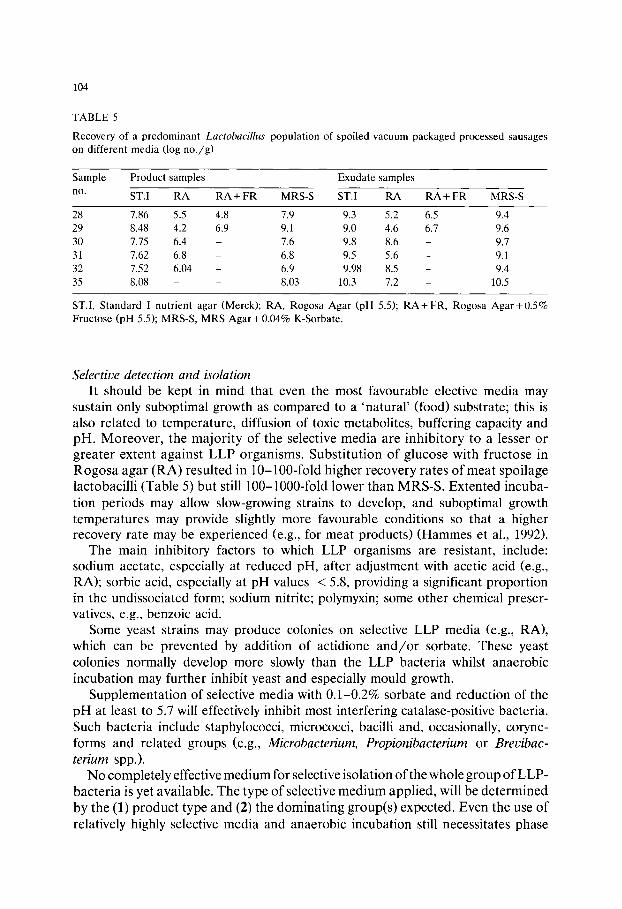
104
TABLE 5
Recovery of a predominant Lactobacillus population of spoiled vacuum packaged processed sausages on different media (log no . /g)
Sample Product samples Exudate samples
no. ST.I RA RA + FR MRS-S ST.I RA RA + FR MRS-S
28 7.86 5.5 4.8 7.9 9.3 5.2 6.5 9.4 29 8.48 4.2 6.9 9.1 9.0 4.6 6.7 9.6 30 7.75 6.4 - 7.6 9.8 8.6 - 9.7
31 7.62 6.8 - 6.8 9.5 5.6 - 9.1 32 7.52 6.04 - 6.9 9.98 8.5 - 9.4
35 8.08 - - 8.03 10.3 7.2 - 10.5
ST.I, Standard I nutrient agar (Merck); RA, Rogosa Agar (pH 5.5); R A + FR, Rogosa A g a r + 0 . 5 % Fructose (pH 5.5); MRS-S, MRS Agar +0.04% K-Sorbate.
Selective detection and isolation It should be kept in mind that even the most favourable elective media may
sustain only suboptimal growth as compared to a 'natural' (food) substrate; this is also related to temperature, diffusion of toxic metabolites, buffering capacity and pH. Moreover, the majority of the selective media are inhibitory to a lesser or greater extent against LLP organisms. Substitution of glucose with fructose in Rogosa agar (RA) resulted in 10-100-fold higher recovery rates of meat spoilage lactobacilli (Table 5) but still 100-1000-fold lower than MRS-S. Extented incuba- tion periods may allow slow-growing strains to develop, and suboptimal growth temperatures may provide slightly more favourable conditions so that a higher recovery rate may be experienced (e.g., for meat products) (Hammes et al., 1992).
The main inhibitory factors to which LLP organisms are resistant, include: sodium acetate, especially at reduced pH, after adjustment with acetic acid (e.g., RA); sorbic acid, especially at pH values < 5.8, providing a significant proportion in the undissociated form; sodium nitrite; polymyxin; some other chemical preser- vatives, e.g., benzoic acid.
Some yeast strains may produce colonies on selective LLP media (e.g., RA), which can be prevented by addition of actidione and/or sorbate. These yeast colonies normally develop more slowly than the LLP bacteria whilst anaerobic incubation may further inhibit yeast and especially mould growth.
Supplementation of selective media with 0.1-0.2% sorbate and reduction of the pH at least to 5.7 will effectively inhibit most interfering catalase-positive bacteria. Such bacteria include staphylococci, micrococci, bacilli and, occasionally, coryne- forms and related groups (e.g., Microbacterium, Propionibacterium or Brevibac- terium spp.).
No completely effective medium for selective isolation of the whole group of LLP- bacteria is yet available. The type of selective medium applied, will be determined by the (1) product type and (2) the dominating group(s) expected. Even the use of relatively highly selective media and anaerobic incubation still necessitates phase

105
con t r a s t mic roscopy of typical colonies , by which l euconos tocs , lac tobaci l l i and
ped iococc i may be d i s t ingu ished . A n exce l len t se lect ive and e lec t ive m e d i u m (NBB) for b e e r spo i lage bac t e r i a has
b e e n f o r m u l a t e d by Back (1978, 1980). N B B (NS.hrboden fiir bierschS.dliche Bak te - r ien) is b a s e d on c o n v e n t i o n a l cu l t iva t ion m e d i a for LAB, bu t also con ta ins
add i t i ona l g rowth fac tors as well as mal ic acid and hops. T h e s e g rowth fac tors a re
supp l i ed in c o n c e n t r a t e d mixture , the exact compos i t i on of which is conf iden t i a l
and for which a p a t e n t has b e e n r e q u e s t e d . N B B is u sed by ca. 95% of the
b r e w e r i e s in G e r m a n y and by severa l in o t h e r E u r o p e a n coun t r i e s and the USA,
for the d e t e c t i o n and e n u m e r a t i o n of b e e r spo i lage ped iococc i and lactobaci l l i . In
d i lu t ed fo rm it has also p roved useful for cu l t iva t ion of fas t id ious L A B f rom b e e r
and wine, e.g., Lactobacillus hilgardii, L. oenos and some s low-growing s t ra ins of
Pediococcus damnosus.
References
Abd-el-Malek, Y. and Gibson, T. (1952) Studies in the bacteriology of milk. III. The corynebacteria of milk. J. Dairy Res. 19, 153-159.
Back, W. (1978) Zur Taxonomie der Gattung Pediococcus. Ph~inotypische und genotypische Abgren- zung der bisher bekannten Arten sowie Beschreibung einer neuen biersch~idlichen Art: Pediococcus inopinatus. Brauwiss. 31,237-259, 312-320, 336-343.
Back, W. (1980) Biersch~idliche Bakterien. Nachweis und Kultivierung biersch~idlicher Bakterien im Betriebslabor. Brauwelt 120, 1562-1569.
Back, W. (1981) Sch~idliche Mikroorganismen in Fruchts~iften, Fruchtnektaren und siissen, alkohol- freien Erfrischungsgetr~inken. Brauwelt 121, 43-48.
Baird, K.J. and Patterson, J.T. (1980) An evaluation of media for the cultivation or selective enumera- tion of lactic acid bacteria from vacuum-packaged beef. Rec. Agric. Res. 28, 55-61.
Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (Eds.) (1987) Pharmacopoeia of Culture Media for Food Microbiology. Int. J. Food Microbiol. 5, 187-300.
Baird, R.M., Corry, J.E.L., Curtis, G.D.W., Mossel, D.A.A. and Skovgaard, N. (Eds.) (1989) Pharma- copoeia of Culture Media for Food Microbiology: additional monographs. Int. J. Food Microbiol. 9, 85-144.
Barnes, E.M. (1976) Microbiological problems of poultry at refrigerator temperatures: a review. J. Sci. Food Agric. 27, 777-782.
Barnes, E.M. and Corry, J.E.L. (1969) Microbial flora of raw and pasteurized egg albumen. J. Appl. Bacteriol. 32, 193-205.
Baumgart, J., Weber, B. and Hanekamp, B. (1983) Mikrobiologische Stabilit~it von Feinkosterzeugnis- sen. Fleischwirtschaft 63, 93-94.
Baumgart, J. (1990) Mikrobiologische Untersuchung von Lebensmitteln. 2. Aufl. Behr's Verlag, Ham- burg.
Boa.twright, J. and Kirsop, B.H. (1976) Sucrose agar: a growth medium for spoilage organisms. J. Inst. Brewing 82, 343-346.
Borch, E. and Molin, G. (1988) Numerical taxonomy of psychrotrophic lactic acid bacteria from prepacked meat and meat products. Antonie van Leeuwenhoek 54, 301-323.
Bracquart, P. (1981) An agar medium for the differential enumeration of Streptococcus thermophilus and Lactobacillus bulgaricus in yoghurt. J. Appl. Bacteriol. 51, 303-305.
Briggs, M. (1953) An improved medium for lactobacilli. J. Dairy Res. 20, 36-40. Britz, T.J. (1975)'n Studie van Propionibacterium spp. in suiwelprodukte. MSc.-Thesis, University of
Pretoria, Pretoria, South Africa.

106
Britz, T.J. and Holzapfel, W.H. (1973) The suitability of different media for the enumeration of propionibacteria. S. Afr. J. Dairy Technol. 5, 213-216.
Cantoni, C., Bianchi, M.A., Renon, P. and D'Autent, S. (1969) Ricerche sulla putrefazione del prosciutto crudo. Arch. Veterin. Ital. 20, 355-370.
Cavett, J.J. (1963) A diagnostic key for identifying the lactic acid bacteria of vacuum-packaged bacon. J. Appl. Bacteriol. 26, 453-470.
Cavett, J.J., Dring, H.J. and Knight, A.W. (1965) Bacterial spoilage of thawed frozen peas. J. Appl. Bacteriol. 28, 241-251.
Cavin, J.F., Schmitt, P., Arias, A., Lin, J. and Divies, C. (1988) Plasmid profiles in Leuconostoc species. Microbiol. Alim Nutr. 6, 55-62.
Cavin, J.F., Prevost, H., Lin, J., Schmitt, P. and Divies, C. (1989) Medium for screening Leuconostoc oenos strains defective in malo-lactic fermentation. Appl. Environ. Microbiol. 55, 751-753.
Collins, M.D. (1992) The genus Brevibacterium. In: A. Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn. Vol. II, Springer Verlag, New York, pp. 1351-1354.
Collins, M.D., Farrow, J.A.E., Goodfellow, M. and Minnikin, D.E. (1983) Brevibacterium casei sp. nov. and Brecibacterium epidermidis sp. nov. Syst. Appl. Microbiol. 4, 388-395.
Collins, M.D. and Keddie, R.M. (,1986) Genus Microbacterium. In: P.H.A. Sneath, N.S. Mair, M.E. Sharpe and J.G. Holt (Eds.). Bergey's Manual of Determinative Bacteriology, Vol. 2. Williams and Wilkins, Baltimore, pp. 1322-1323.
Collins, M.D., Farrow, A.E., Philips, B.A., Feresu, S. and Jones, D. (1987) Classification of Lactobacil- lus divergens, Lactobacillus piscicola and some catalase-negative, asporogenous, rod-shoped bacteria from poultry in a new genus, Carnobacterium. Int. J. Syst. Bacteriol. 37, 310-316.
Collins, M.D. and Bradbury, J.F. (1992) The genera Agromyces, Aureobacterium, Clavibacter, Curtobac- terium and Microbacterium. In: A. Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn. Vol. II, Springer Verlag, New York, pp. 1355-1368.
Coretti, K. (1958) Rohwurstfehlfabrikate durch Laktobazillen. Fleischwirtschaft 4, 218-225. Cummins, C.S. and Johnson, J.L. (1992) The genus Propionibacterium. In: A. Balows, H.G. Triiper, M.
Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn. Vol. I, Springer Verlag, New York, pp. 834-849.
Curry, J.C. and Borovian, G.E. (1976) Selective medium for distinguishing micrococci from staphylo- cocci in the clinical laboratory. J. Clin. Microbiol. 4, 455-457.
Curtis, G.D.W., Corry, J.E.L. and Baird, R.M. (1992) Pharmacopoeia of Culture Media for Food Microbiology: additional monographs (2). Int. J. Food Microbiol. 17 (in press).
Davidson, C.M. and Cronin, F. (1973) Medium for the selective enumeration of lactic acid bacteria from foods. Appl. Microbiol. 26, 439-440.
Deibel, R.H. and Niven, C.F. (1960) Comparative study of Gaffkya homari, Aerococcus viridans, tetrad-forming cocci from meat curing brines and the genus Pediococcus. J. Bacteriol. 79, 175-180.
de Man, J.C., Rogosa, M. and Sharpe, M.E. (1960) A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130-135.
Demeter, K.J. and Janoschek, A. (1941) Vorkommen und Entwicklung der Propions~iurebakterien in verschiedenen K~isearten. Zbl. Bakteriol. Abt. 2, 103, 257-271.
Dittrich, H.H. (1977) Mikrobiologie des Weines. Handbuch der Getr~inketechnologie. Eugen Ulmer Verlag, Stuttgart.
Eloy, C. and Lacrosse, R. (1976) Composition d'un milieu de culture destine a effecteur le denombre- ment des micro-organismes thermophiles du yoghurt. Bull. Rech. Agron. Gemblou 11 (1-2), 83-86.
Evans, J.B. and Niven, C.F. (1951) Nutrition of the heterofermenative lactobacilli that cause greening of cured meat products. J. Appl. Bacteriol. 62, 599-603.
Evans, J.B. (1986) Genus Aerococcus. In: P.H.A. Sneath, N.S. Mair, M.E. Sharpe and J.G. Holt (Eds.). Bergey's Manual of Systematic Bacteriology, Vol. 2. William and Wilkins, Baltimore, p. 1080.
Franzmann, P.D., H6pfl, P., Weiss, N. and Tindall, B.J. (1991) Psychrotrophic, lactic acid-producing bacteria from anoxic waters in Ace Lake, Antarctica; Carnobacterium funditum sp. nov. and Carnobacterium alterfunditum sp. nov. Arch. Microbiol. 156, 255-262.

107
Fryer, T.F., Sharpe, M.E. and Reiter, B. (1970) Utilisation of milk citrate by lactic acid bacteria and 'blowing' of film wrapped cheese. J. Dairy Res. 37, 17-28.
Gardner, G.A. (1966) A selective medium for the enumeration of microbacterium thermosphactum in meat and meat products. J. Appl. Bacteriol. 29, 455-460.
Gardner, G.A. (1969) Physiological and morphological characteristics of Kurthia zopfii isolated from meat products. J. Appl. Bacteriol. 32, 371-380.
Gardner, G.A. (1981) Brochothrix thermosphacta (Microbacterium thermosphactum) in the spoilage of meats: a review. In: T.A. Roberts, G.A. Hobbs, J.H.B. Christian and N. Skovgaard (Eds). Psy- chrotrophic Microorganisms in Spoilage and Pathogenicity. Academic Press, London, pp. 139-173.
Gardner, G.A. (1985) Streptomycin-thallous acetate-actidione (STAA) agar: a medium for the selective enumeration of Brochothrix thermosphacta. Int. J. Food Microbiol. 2, 69-70.
Garvie, E.J. (1967) Leuconostoc oenos sp. nov. J. Gen. Microbiol. 48, 431-438. Hammes, W.P., Weiss, N. and Holzapfel, W.H. (1992) Lactobacillus and Carnobacterium. In: A.
Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes. 2nd Edn. Vol. II, Springer Verlag, New York, pp. 1535-1594.
Harrigan, W.F. and McCance, M.E. (1976) Laboratory Methods in Food and Dairy Microbiology, revised Edn. Academic Press, London.
Hatcher, W.S., Jr., Hill, E.C., Splittstoesser, D.F. and Weihe, J.L. (1984) Fruit beverages. In: M.L. Speck (Ed.) Compendium of Methods for the Microbiological Examination of Foods. Am. Pub. Health Assoc., Washington, DC Chapter 47, p. 645.
Hiu, S.R., Holt, R.A., Sriranganathan, N., Seidler, R.J. and Fryer, J.L. (1984) Lactobacillus piscicola, a new species from salmonid fish. Int. J. Syst. Bacteriol. 34, 393-400.
Hitchener, B.J., Egan, A.F. and Rogers, P.J. (1982) Characteristics of lactic acid bacteria isolated from vacuum-packaged beef. J. Appl. Bacteriol. 52, 31-37.
Holzapfel, W.H. and Gerber, E.S. (1983) Lactobacillus dicergens sp. nov., a new heterofermentative Lactobacillus species producing L( + )-lactate. System. Appl. Microbiol. 4, 522-534.
Holzapfel, W.H. and Schillinger, U. (1992) The genus Leuconostoc. In: A. Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn., Vol. II, Springer Verlag, New York, pp. 1508-1534.
ISO (International Standards Organization) Discussion (1983) Report of the first meeting of ISO/TC 34/SC 6/WG 15 "Enumeration of Lactobacteriaceae in meat and meat products". Wageningen, The Netherlans 09.09.1983.
ISO (International Standards Organization) Discussion (1986) Third meeting 34/6 WG 15, Ghent, Belgium, 26.08.1986.
Johnson, J.L. and Cummins, C.S. (1972) Cell wall composition and deoxyribonucleic acid similarities among the anaerobic coryneforms, classical propionibacteria and strains of Arachnia propionica. J. Bacteriol. 109, 1047-1066.
Jones, D. (1992) The genus Brochothrix. In: A. Balows, H.G. Trfiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes. 2nd Edn. Vol. II, Springer Verlag, New York, pp. 1617-1628.
Jones, D. and Keddie, R.M. (1986) Genus BreL, ibacteriurn. In: P.H.A. Sneath, N.S. Mair, M.E. Sharpe and J.G. Holt (Eds.). Bergey's Manual of Determinative Bacteriology, Vol. 2, Williams and Wilkins, Baltimore, London, pp. 1301-1313.
Kamber, C.S., Reinbold, G.W. and Hussong, R.V. (1952) A plating method for the isolation and enumeration of propionibacteria. J. Dairy Sci. 35, 915-919.
Keddie, R.M. (1981) The genus Kurthia. In: M.P. Starr, H. Stolp, H.G. Triiper, A. Balows and H.G. Schlegel (Eds.). The Prokaryotes. A Handbook on Habitats, Isolation and Identification of Bacteria. Springer Verlag, Berlin, pp. 1888-1893.
Keddie, R.M. and Jones, D. (1981a) The genus Brochothrix (formerly Microbacterium thermosphactum, McLean and Sulzbacher). In: M.P. Starr, H. Stolp, H.G. Trfiper, A. Balows and H.G. Schlegel (Eds.). The Prokaryotes. A Handbook on Habitats, Isolation and Identification of Bacteria. Springer Verlag, Berlin, pp. 1866-1869.
Keddie, R.M. and Jones, D. (1981b) Aerobic saprophytic coryneform bacteria. In: M.P. Starr, H. Stolp, H.G. Triiper, A. Balows and H.G. Schlegel (Eds.). The Prokaryotes, Springer Verlag, New York, pp. 1838-1978.

108
Keddie, R.M. and Jones, D. (1992) The genus Kurthia. In: A. Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn. Vol. II, Springer Verlag, New York, pp. 1654-1662.
Keller, J.J., Holzapfel, W.H. and Steinman, M.A. (1987) The microbiological population differences between pasteurised and spoiled pasteurised milk. S. Afr. J. Dairy Technol. 19, 85-95.
Kleynmans, U., Heinzl, H. and Hammes, W.P. (1989) Lactobacillus suebicus sp. nov., an obligately heterofermentative Lactobacillus species isolated from fruit mashes. Syst. Appl. Microbiol. 11, 267-271.
Kocur, M., Kloos, W.E. and Schleifer, K.H. (1992) The genus Micrococcus. In: A. Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn. Vol. II. Springer Verlag, New York, pp. 1300-1311.
Malik, A.C., Reinbold, G.W. and Vedamuthu, E.R. (1968) An evaluation of the taxonomy of Propioni- bacterium. Can. J. Microbiol. 14, 1185-1191.
McDonald, L.C., McFeeters, R.F., Daeschel, M.A. and Fleming, H.P. (1987) A differential medium for the numeration of homofermentative and heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 53, 1382-1384.
McFeeters, R.F., Fleming, H.P and Daeschel, M.A. (1984) Malic acid degradation and brined cucumber bloating. J. Food Sci. 49, 999.
McLean, R.A. and Sulzbacher, W.L. (1953) Microbacterium thermosphactum spec. nov.: a nonheat resistant bacterium from fresh pork sausage. J. Appl. Bacteriol. 65, 428-433.
Mol, J.H.H., Hietbrink, J.E.A., Mollen, H.W.M. and van Tinteren, J. (1971) Observations on the microflora of vacuum-packed sliced cooked meat products. J. Appl. Bacteriol. 34, 377-397.
Mundt, J.O. (1982) The ecology of the streptococci. Microb. Ecol. 8, 355-369. Murdock, D.I., Folinazzo, J.F. and Troy, V.S. (1952) Evaluation of plating media for citrus concen-
trates. Food Technol. 6, 181-185. Naylor, H.B. and Burgi, E. (1956) Observations on abortive infections of Micrococcus lysodeikticus with
bacteriophage. Virology 2, 577-593. Nickelson, R., II, Finne, G., Hanna, M.O. and Vanderzant, C. (1980) Minced fish flesh from
nontraditional Gulf of Mexico finfish species: bacteriology. J. Food Sci. 45, 1321-1326. Niethammer, A. and Hitzler, M. (1960) Synthetic culture of Propionibacterium. Zbl. Bakteriol. Abt. 2,
113, 478. Nieuwenhof, F.F.J., Stadhouders, J. and Hup, G. (1969) Stimulating effect of lactobacilli on the growth
of propionibacteria in cheese. Ned. Melk Zuiveltijdschr. 23, 287-289. Owens, J.D. and Keddie, R.M. (1969) The nitrogen nutrition of soil and herbage coryneform bacteria.
J. Appl. Bacteriol. 32, 338-347. Rankine, B.C. (1977) Developments of malo-lactic fermentation of Australian red table wines. Am. J.
Enol. Vitic. 28, 27-33. Reuter, G. (1970a) Mikrobiologische Analyse von Lebensmitteln mit selektiven Medien. Arch. Lebens-
mittelhyg. 21, 30-35. Reuter, G. (1970b) Laktobazillen und eng verwandte Mikroorganismen in Fleisch und Fleischerzeugnis-
sen 2. Mitteilung: Die Charakterisierung der isolierten Laktobazillenst~imme. Fleischwirtschaft 50, 954-962.
Reuter, G. (1975) Classification problems, ecology and some biochemical activities of lactobacilli of meat products. In: J.G. Carr, C.V. Cutting and G.C. Whiting (Eds.). Lactic Acid Bacteria in Beverages and Food. Academic Press, London, pp. 221-229.
Reuter, G. (1985) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68. Reuter, G. (1992) Culture media for enterococci and group D-streptococci. Int. J. Food Microbiol. 17,
101-111. Rogosa, M., Mitchell, J.A. and Wisemann, R.F. (1951) A selective medium for the isolation and
enumeration of oral and faecal lactobacilli. J. Bacteriol. 62, 132-133. Schillinger, U. and Liicke, F.-K. (1986) Milchs~iurebakterien-Flora auf vakuumverpacktem Fleisch und
ihr Einfluss auf die Haltbarkeit. Fleischwirtschaft 66, 1515-1520. Sharpe, M.E., Law. B.A., Phillips, B.A. and Pitcher, D.G. (1977) Methanethiol production by coryne-
form bacteria: strains from dairy and human skin sources and Brevibacterium linens. J. Gen. Microbiol. 101,345-349.

109
Sharpe, M.E. and Pettipher, G.L. (1983) Food spoilage by lactic acid bacteria. Economic Microbiol. 8, 199-223.
Shaw, B.G. and Harding, C.D. (1984) A numerical taxonomic study of lactic acid bacteria from vacuum-packed beef, pork, lamb and bacon. J. Appl. Bacteriol. 56, 25-40.
Shaw, S. and Keddie, R.M. (1983) A numerical taxonomic study of the genus Kurthia with a revised description of Kurthia zopfii and a description of Kurthia gibsonii sp. nov. Syst. Appl. Microbiol. 4, 253-276.
Skovgaard, N. (1985) Brochothrix thermosphacta comments on its taxonomy, ecology and isolation. Int. J. Food Microbiol. 2, 71-79.
Sneath, P.H.A. and Jones, D. (1976) Brochothrix, a new genus tentatively placed in the family Lactobacillaceae. Int. J. Syst. Bacteriol. 26, 102-104.
Talon, R., Grimont, P.A.D., Grimont, F., Gasser, F. and Boeufgras, J.M. (1988) Brochothrix campestris sp. nov. Int. J. Syst. Bacteriol. 38, 99-102.
Terzaghi, B.E. and Sandine, W.E. (1975) Improved, medium for lactic streptococci and their bacterio- phages. Appl. Microbiol. 29, 807-813.
Thornley, M.J. and Sharpe, M.E. (1959) Microorganisms from chicken meat related to both lactobacilli and aerobic sporeformers. J. Appl. Bacteriol. 22, 368-376.
Toba, T., Abe, S., Arihara, K. and Adachi, S. (1986) A medium for the isolation of capsular bacteria from kefir grains. Agric. Biol. Chem. 50, 2673-2674.
Vaughn, R.H. (1985) The microbiology of vegetable fermentations. In: B.J.B. Wood (Ed.). Microbiology of Fermented Foods, Vol. 2, Elsevier, New York, pp. 49-109.
Vedamuthu, E.R. and Reinbold, G.W. (1967) The use of candle oats jar incubation for the enumera- tion, characterization and taxonomic study of propionibacteria. Milchwiss. 22, 428-431.
Villar, M., de Ruiz Holgado, A.P., Sanchez, J.J., Trucco, R.E. and Oliver, G. (1985) Isolation and characterization of Pediococcus halophilus from salt anchovies (Engraulis anchoita). Appl. Environ. Microbiol. 49, 664-666.
von Holy, A. and Holzapfel, W.H. (1989) Spoilage of vacuum packaged processed meats by lactic acid bacteria and economic consequences. Proc.: Xth International World Association of Veterinary Food Hygiene Symp., 2-7 July 1989, Stockholm, Sweden (pp. 185-190).
Vrbaski, L., Pekic, B. and Hauk, M. (1988) An improved medium for isolation of Leuconostoc mesenteroides. Mikrobiologija (Belgr) 25, 79-90.
Weiss, N. (1992) The genera Pediococcus and Aerococcus. In: A. Balows, H.G. Triiper, M. Dworkin, W. Harder and K.H. Schleifer (Eds.). The Prokaryotes, 2nd Edn. Vol. II, Springer Verlag, New York, pp. 1502-1507.
Wilkinson, B.J. and Jones, D. (1977) A numerical taxonomic survey of Listeria and related bacteria. J. Gen. Microbiol. 98, 399-421.
Yoshizumi, H. (1975) A malo-lactic bacterium and its growth factor. In: J.G. Carr, C.V. Cutting and G.C. Whiting (Eds). Lactic Acid Bacteria in Beverages and Food. Academic Press, London pp. 87-102.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
111
Chapter 8 Media for Aeromonas spp., Plesiomonas shigelloides and Pseudomonas spp. from food and environment
C l a u s J e p p e s e n
Environmental and Food Laboratory, Dyreg~rdsvej 1, 2740 Skovlunde, Denmark
Abstract
Several media are proposed particularly for the detection of Aeromonas spp. but also for Plesiomonas shigelloides and Pseudomonas spp. Some are for general purposes and others specifically for the examination of clinical, environmental or food samples.
All media are selective, due to antibiotics, bile salts, dyes and other selective agents, as well as differential, primarily based on the ability of the microorganisms to ferment/not ferment carbohydrates. As with all selective media, the recovery of stressed cells is sometimes prevented and the competing flora is not always completely inhibited so that confirmatory tests need to be made on presumptive positive colonies.
The choice of a specific medium for isolation of Aeromonas spp. will always depend on the type of sample to be examined and whether the investigator needs qualitative detection or quantitative recovery. The best medium for quantitative estimation of Aeromonas spp. from food and environmental samples seems to be starch ampicillin agar (SAA), though others might be recommended. There is a need for a comparative study including Rippey Cabelli agar (mA), ampicillin bile salts inositol xylose (MIX) agar, ampicillin dextrin agar (ADA), dextrin fuchsin sulphite agar (DFS) and starch glutamate ampicillin penicillin C-glucose agar (SGAP-10C) in addition to SAA.
For routine analysis of environmental and food samples for P. shigelloides, spread plating on inositol brilliant green bile salts (IBB) and plesiomonas (PL) agars is recommended.
For Pseudomonas spp., CFC agar permits quantitative recovery of both pigmented and non-pigmented strains from food and environmental samples, whilst at the same time inhibiting most other organisms.
1. Introduction
Aeromonas , Ples iomonas and many Pseudomonas spp. are ubiquitous but they
are most often associated with fresh surface water. These organisms have been

112
isolated from a variety of sources including wild and domestic animals, diarrhetic and asymptomatic humans as well as drinking water and various foods.
Aeromonas hydrophila is a facultatively anaerobic, Gram-negative rod. General characteristics of A. hydrophila include: motility by a single polar flagellum; metabolism of glucose by both fermentative and respiratory pathways; positive catalase and oxidase tests; production of exoenzymes such as amylase, protease, phospholipase and DNase; resistance to the vibriostatic agent O/129. Popoff (1984) lists four species in the genus Aeromonas: A. hydrophila, A. caviae, A. sobria and the A. salmonicida group consisting of A. salmonicida and its subspecies.
The A. hydrophila group (including also A. caviae and A. sobria) is character- ized by growth at 37~ and motility. A. salmonicida does not grow at 37~ is non-motile, usually produces a brown pigment and is a pathogen of salmonid fish. The A. hydrophila group is referred to as the motile aeromonads (or mesophilic aeromonads). It is this group that is of interest and importance to human public health.
Plesiomonas shigelloides is a motile, oxidase- and catalase-positive, facultatively anaerobic Gram-negative rod, having both a respiratory and a fermentative type of metabolism. Carbohydrates are catabolized with production of acid but no gas. Most strains are sensitive to vibriostatic agent O/129. P. shigelloides is an oppor- tunistic pathogen.
Pseudomonas spp. are motile (rarely non-motile), Gram-negative, catalase-posi- tive, oxidase-positive or -negative, straight or slightly curved rods. They are aerobic, having a strictly respiratory type of metabolism with oxygen as the terminal electron acceptor. In some cases nitrate can be used as an alternative electron acceptor, allowing growth to occur anaerobically. The predominant spoilage organisms in high water activity, proteinaceous foods belong to the genus Pseudomonas.
A variety of selective and differential media has been proposed, particularly for the detection of Aeromonas spp. but also for P. shigelloides and Pseudomonas spp. from clinical, environmental and food samples, but no single medium has received general acceptance. Problems may arise especially when quantitative recovery is required or in cases ~here large numbers of competing flora are present.
Depending on the purpose of the examination, the following factors will influence the choice of medium: (1) selectivity, i.e., the ability to suppress back- ground flora; (2) ability to differentiate between the organisms looked for and the competing flora on basis of colonial morphology; (3) quantitative recovery of the desired organism, including recovery of injured cells.
In clinical samples the organisms in question are usually present in high numbers. In food and environmental samples on the other hand they might only be present in low concentrations and are often accompanied by a competing mi- croflora that is physiologically similar to the organisms sought. Furthermore the organisms may often be injured, e.g., due to refrigeration or heating. Food and water microbiologists are concerned not only about the presence of various organis~s, but also about the quantity since the risk of spoilage and the degree of hazard is related to the number of organisms present in the samples.

113
The aim of this paper is to review media proposed in the literature for isolation of Aeromonas, Plesiomonas and Pseudomonas spp.
2. Isolation and enumeration of Aeromonas spp.
2.1. Plating media
Numerous procedures and media have been proposed for the isolation of Aeromonas spp. from clinical, environmental and food samples. Many investiga- tions of methods for the isolation of Aeromonas spp. have been made by clinical microbiologists and often the same media have been used for isolation from food and the environment.
Though often used, the usual enteric media such as Hektoen enteric agar, MacConkey agar, eosin methylene blue agar and Salmonella-Shigella agar are not suitable for selection of Aeromonas and Plesiomonas spp. as both genera include lactose-negative as well as lactose-positive strains (von Graevenitz and Bucher, 1983; Desmond and Janda, 1986).
Several plating media (listed in Table 1) have been developed for the isolation of Aeromonas spp. from clinical, environmental and food samples. The selectivity of most media depends on ampicillin and/or inhibitors used in media for Enter- obacteriaceae (bile salts, brilliant green and sodium lauryl sulphate). Differentia- tion most often exploits amylase or carbohydrate metabolism.
Plating media for clinical samples Von Graevenitz and Bucher (1983) evaluated two enrichment broths and nine
plating media for their efficiency in recovering Aeromonas spp. from human faeces and correctly differentiating them from Enterobacteriaceae and P. shigel- loides. They found both alkaline peptone water (APW) and trypticase soy broth with ampicillin (TSBA) suitable for enrichment. As differential plating media the best were inositol brilliant green bile salts agar (IBB), dextrin fuchsin sulphite agar (DFS) (originally developed for isolating Aeromonas spp. from water (Schubert, 1967)), xylose sodium deoxycholate citrate agar (XDC) and pril xylose ampicillin agar (PXA). DNase-toluidine blue-ampicillin agar (DNTA)performed poorly compared to the media listed above, peptone beef extract glycogen agar (PBG) could not differentiate Aeromonas spp. from normal stool flora and Rimler-Shotts agar (RS) was not sufficiently inhibitory to other Gram-negative organisms. Salt starch xylose lysine sodium deoxycholate agar (SSXLD) was eliminated because starch hydrolysis by Aeromonas strains could not be clearly seen and the specificity of Rippey Cabelli agar (mA)was insufficient, because it could not clearly distin- guish aeromonads from Enterobacteriaceae.
A very simple direct plating medium developed for clinical samples is blood agar with ampicillin (BA-AMP) (Millership et al., 1983). Ampicillin prevents growth of most of the competing faecal flora whilst allowing efficient recovery of

114
Aeromonas spp. Another advantage of BA-AMP is that it is a good medium to use for the oxidase test.
Plating media for food and environmental samples The detection of Aeromonas spp. in foods and environmental samples is usually
quite easy. However, difficulties may arise when quantitative recovery is required or in cases where large numbers of other microorganisms are present, e.g., in minced meat. Rimler-Shotts agar (RS) has often been used in environmental investigations but studies have demonstrated that the recovery rate is low, partly due to the use of novobiocin and also because the detection of lysine decarboxylat- ing Aeromonas spp. is not satisfactory (Rippey and Cabelli, 1979; Kaper et al., 1981; Arcos et al., 1988).
Palumbo et al. (1985) tested some of the media described by von Graevenitz and Bucher (1983) on food samples and found that the clinical media were not suitable for use on food products. Media containing bile salts, deoxycholate and novobiocin were too inhibitory, Aeromonas spp. could not be readily differentiated and the competing microflora of food overgrew the plates. Palumbo et al. (1985) developed a new medium specifically for the quantitative recovery of Aeromonas spp. from foods: starch ampicillin agar (SAA). SAA is composed of phenol red agar base (Difco), ampicillin (10 mg/1) to suppress the background microflora and soluble starch (10 g/ l ) as a differential substance. Relatively few organisms in food except for Aeromonas and most Vibrio spp. are amylase-positive. When pure cultures or food samples were surface plated on SAA and incubated at 28~ for 24 h, colonies of Aeromonas spp. appeared yellow to honey-coloured with a diameter of 2-3 mm. Production of amylase was determined by flooding the plates with Lugol's iodine solution. Amylase-positive colonies were surrounded by a clear zone, indicative of starch hydrolysis. Palumbo et al. (1992) modified SAA by replacing the soluble starch with amylose azure (Sigma) eliminating the need for flooding with iodine. Amylase-positive colonies appeared as surrounded by a light halo against a blue background. Amylose azure is rather expensive ($350/25 g) and at the Environ- mental and Food Laboratory we have observed, that a light halo with a width of 2-3 mm surrounds aeromonas colonies on unmodified SAA, even when the plates are not flooded with any reagent. This observation is in agreement with Hudson and de Lacy (1991) and the method seems to be reliable and convenient. Our experience with SAA is that it is a very effective medium for the isolation of Aeromonas spp. from food and water, enabling rapid differentiation of these microorganisms from the background flora. For food samples we normally find confirmation rates, i.e., presumptive isolates verified as Aeromonas spp., of nearly 100% (results not shown).
Palumbo et al. (1985) surveyed retail foods of animal origin using SAA and found Aeromonas spp. in all foods examined, despite of the presence of very large numbers of competing microflora.
Other investigators have also utilized SAA to examine various food samples for the presence of Aeromonas spp. (Callister and Agger, 1987; Okrend et al., 1987; Stern et al., 1987). All investigators found SAA particularly useful, especially in

115
terms of confirmation rates. Kn6chel (1989) surveyed environmental as well as food samples and observed a 85% confirmation rate on SAA compared with only 18% on dextrin fuchsin sulphite agar (DFS) and 36% and 40% on blood agar with 10 mg and 30 mg respectively of ampicillin/1.
The incorporation of ampicillin in SAA suppresses Enterobacteriaceae and the ampicillin resistant competing flora, primarily psychrotrophic Pseudomonas spp., can mostly be eliminated by incubating at 35-37~ A few motile Aeromonas spp. (< 1%) are sensitive to ampicillin whereas more than 10% are sensitive to novobiocin (Rippey and Cabelli, 1979; Palumbo et al., 1985). SAA does not differentiate Aeromonas spp. from Vibrio spp., but it is our experience that none of the presumptive Aeromonas isolates from food are identified as Vibrio spp. This may be explained by the fact that Aeromonas spp. are only of interest in chilled foods, where most Vibrio spp. will not grow. Furthermore those Vibrio spp. capable of growing at 4~ are amylase-negative and therefore would not be confused with motile Aeromonas spp. on SAA (Baumann et al., 1984). For environmental samples Vibriostatic agent O/129 might be incorporated in SAA.
Nishikawa and Kishi (1987) designed bile salt brilliant green starch agar (BBS) for the isolation of Aeromonas spp. and found it to be suitable for the isolation and quantitative recovery of motile Aeromonas spp. from foods and environmental samples, detecting Aeromonas spp. by their amylase reaction.
Rippey and Cabelli (1979) developed a method for the recovery and enumera- tion of Aeromonas spp. from environmental samples using membrane filters. Their medium (mA) contains ampicillin, sodium deoxycholate and ethanol as selective agents and trehalose as a differential agent. Suspect colonies of Aeromonas spp. on mA are yellow (trehalose-positive), circular and 1-3 mm in diameter. Arcos et al. (1988) evaluated eleven media for their suitability for the selective isolation of Aeromonas spp. from polluted water, mA showed the best recovery rate, but its selectivity was low.
Havelaar et al. (1987) evaluated seven selective media for the isolation of Aeromonas spp. from environmental samples by membrane filtration and only obtained satisfactory recoveries with the mA agar of Rippey and Cabelli (1979) and dextrin fuchsin sulphite agar (DFS, Schubert, 1967). Glutamate starch peni- cillin agar (GSP, Kielwein, 1969) gave recoveries as good as mA and DFS. Despite this, GSP was not tested further because the yellow colour resulting from starch hydrolysis tended to express itself only as a halo in the medium and this made accurate counting difficult.
Neither Arcos et al. (1988) nor Havelaar et al. (1987) included SAA in their evaluation. In an examination of wastewater Poff6 and de Beeck (1991) found that in the presence of a high level of competing flora (e.g., Escherichia coli and Ps. fluorescens) recovery on mA was distinctly better than on SAA, both by plate count and membrane filtration.
Havelaar et al. (1987) designed a modification of mA-agar and DFS agar. The positive aspects of these two media were combined in a new medium, ampicillin dextrin agar (ADA). Recovery of Aeromonas spp. from environmental samples was optimal at an ampicillin concentration of 10 mg/1 and incubation for 24 h at

Table 1 Media for selection of Aeromonas spp.
Medium Selective agents Differential agents Original purpose Reference
Ampicillin bile salts inositol Ampicillin (20 mg/l); Inositol; xylose Environmental Cunliffe and Adcock (1989) xylose agar ( 'MIX agar') bile salts
Ampicillin blood agar (BA-AMP) Ampicillin (10 mg/1)
Ampicillin dextrin agar (ADA) Ampicillin (10 mg/l)
Amylose ampicillin agar (AAA) Ampicillin (10 mg/1)
Bile salts brilliant green Bile salts; brilliant green starch agar (BBS)
Dextrin fuchsin sulphite agar (DFS)
DNase toluidine blue ampicillin agar (DNTA)
Inositol brilliant green bile salts agar (IBB)
Peptone beef extract glycogen agar (PBG)
Sodium sulphite; fuchsin
Ampicillin (30 mg/1)
Brilliant green; bile salts
Sodium lauryl sulphate
Erythrocytes
Dextrin
Amylose azure
Starch
Dextrin
DNA
Inositol
Glycogen
Clinical
Environmental
Food
Food/environmental
Environmental
Clinical
Clinical
Environmental
Millership et al. (1983)
Havelaar et al. (1987)
Lachica (1990)
Nishikawa and Kishi (1987)
Schubert (1967)
von Graevenitz and Zinterhofer (1970)
Schubert (1977)
McCoy and Pilcher (1974)
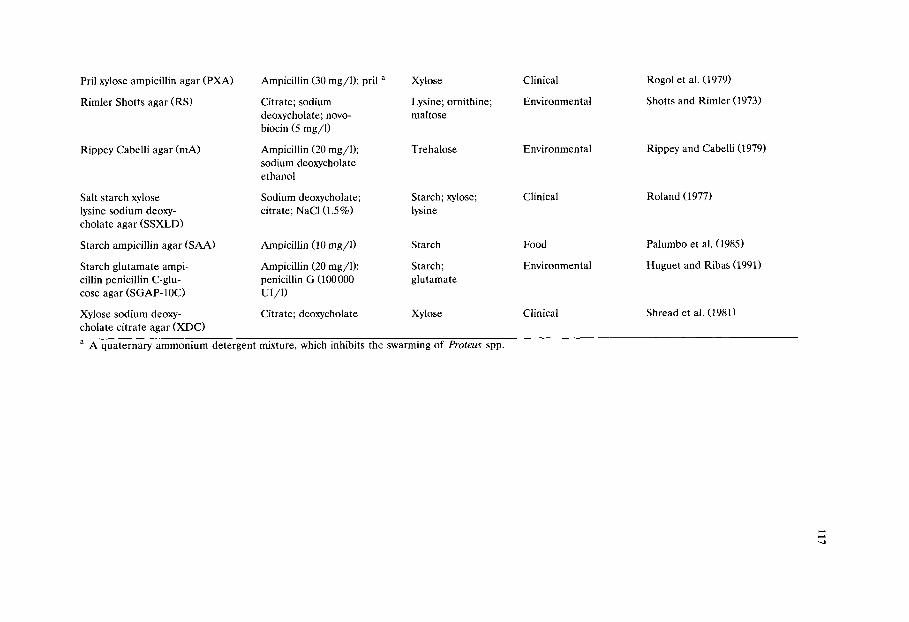
Pril xylose ampicillin agar (PXA)
Rimler Shotts agar (RS)
Rippey Cabelli agar (mA)
Salt starch xylose lysine sodium deoxy- cholate agar (SSXLD)
Starch ampicillin agar (SAA)
Starch glutamate ampi- cillin penicillin C-glu- cose agar (SGAP-10C)
Ampicillin (30 mg/1); pril a
Citrate; sodium deoxycholate; novo- biocin (5 mg/l)
Ampicillin (20 mg/1); sodium deoxycholate ethanol
Sodium deoxycholate; citrate; NaCI (1.5%)
Ampicillin (10 mg/l)
Ampicillin (20 mg/1); penicillin G (100000 UI/1)
Xylose
Lysine; ornithine; maltose
Trehalose
Starch; xylose; lysine
Starch
Starch; glutamate
Xylose sodium deoxy- Citrate; deoxycholate Xylose cholate citrate agar (XDC)
Clinical
Environmental
Environmental
Clinical
Food
Environmental
Clinical
Rogol et al. (1979)
Shotts and Rimler (1973)
Rippey and Cabelli (1979)
Roland (1977)
Palumbo et al. (1985)
Huguet and Ribas (1991)
Shread et al. (1981)
a A quaternary ammonium detergent mixture, which inhibits the swarming of Proteus spp.

118
30~ The confirmation rate was > 90% and no false-negative samples were encountered. They recommended the medium also for isolation of Aeromonas spp. from sea water, provided that the vibriostatic agent O/129 was added in a concentration of 50 mg/1. ADA is an unbuffered medium and consequently, the pH may easily vary. It is important to use freshly prepared medium because the pH may fall during storage in a refrigerator. The use of ADA at a final pH 7.8 + 0.2 (at 25~ is recommended. The suitability of different brands of dextrin was tested and optimum results were obtained with Difco 161 and Merck 3006 (Havelaar and Vonk, 1988). Aeromonas spp. ferment dextrin and produce yellow colonies, 1-2 mm in diameter. The suppression of competing flora from environmental samples is variable but in general sufficient for practical purposes, as the differences in size and colour between typical and atypical colonies make it quite easy to detect presumptive Aeromonas spp.
Cunliffe and Adcock (1989) designed a 'MIX agar' with two selective agents: ampicillin and bile salts and two differential agents: meso-inositol and xylose. The carbohydrates were included for negative selection, as neither is utilized by Aeromonas spp. Typical Aeromonas colonies grown on MIX agar are circular and blue-green. The method utilizes anaerobic incubation as a means of reducing the growth of non-aeromonads while allowing the growth of the facultatively anaerobic Aeromonas spp. Recoveries on MIX agar were comparable to those on nutrient agar and the confirmation rate of presumptive Aeromonas spp. exceeded 92%. The best results were obtained when samples of surface water and drinking water were membrane filtered and incubated anaerobically (10% H 2 90% N 2) at 30~ for 18 h and then aerobically at 35~ for 24 h.
As a means of reducing background flora of drinking and surface water on ADA, Havelaar et al. (1987) also examined anaerobic incubation (100% N 2 for 48 h at 30~ They found that anaerobic incubation lowered the rate of dextrin fermentation and thereby reduced the specificity of the isolation medium. On A D A Aeromonas spp. are identified by fermentation of dextrin, whereas on MIX agar Aeromonas spp. are identified by an inability to ferment meso-inositol as well as xylose. The contrasting results on anaerobic incubation may be explained by the different criteria for recognition of presumptive colonies on the two media as well as by the different conditions of incubation (time/atmosphere).
Huguet and Ribas (1991) found that by modification of glutamate starch penicillin medium (GSP) the isolation and enumeration of Aeromonas spp. from environmental samples could be improved. In a preliminary experiment Huguet and Ribas (1991) found that GSP, ADA and SAA gave equal results for the selection and enumeration of Aeromonas spp. They modified the original GSP medium (Kielwein, 1969) by addition of 10/~g of glucose-C(arbon)/1 (as nutrient addition) and ampicillin (20 mg/1). This medium, SGAP-10C, permitted better recovery of stressed cells of Aeromonas spp., and sodium G penicillin in combina- tion with ampicillin inhibited Pseudomonas spp. and the remaining background flora efficiently. Furthermore the majority of the background flora, i.e., Pseu- domonas spp., do not use the hydrocarbon sources as easily as Aeromonas spp., which can make use of very low concentrations of carbon (van der Kooij et al.,

119
1980). SGAP-10C gave significantly better recovery of Aeromonas spp. compared to GSP, especially when large numbers of Pseudomonas spp. were present. Hugnet and Ribas (1991) concluded that SGAP-10C incubated at 28~ for 48 h was an efficient selective medium for the isolation and enumeration of Aeromonas spp. from environmental samples. In Table 1 only the SGAP-10C-medium is included.
An incubation temperature of 35-37~ has been suggested for clinical samples whilst 25-30~ is recommended for environmental and food samples. The choice of temperature must depend on the purpose of the examination and especially if the goal is to detect potentially human infective Aeromonas spp. an incubation temperature of 35~176 is optimal.
2.2. Enrichment procedures
The use of enrichment procedures depends on the samples to be examined. It is well known that injured cells cannot be quantitatively recovered on selective media, but only very few studies have examined the recovery of injured Aeromonas spp. As previously mentioned, von Graevenitz and Bucher (1983) recommended trypticase soy broth with ampicillin (TSBA) and alkaline peptone water (APW) as enrichment broths for the isolation of Aeromonas spp. Abeyta et al. (1989) compared TSBA with modified Rimler Schotts Broth (MRSB) and found that TSBA was far superior to MRSB for isolating Aeromonas spp. from environmental samples, especially when large numbers of other competing organisms were present. Palumbo et al. (1985) suggested a most probable number enrichment with APW followed by detection on SAA for stressed organisms present in low levels.
2.3. Identification
The classification of motile Aeromonas spp. is complex. A summary of impor- tant tests that differentiate motile Aeromonas spp. from other organisms fre- quently encountered in the same environment is given in Table 2 and confirmation tests for presumptive Aeromonas spp. are listed in Table 3.
Table 2 Important tests differentiating motile Aeromonas spp. from other organisms in the same environment (modified after Popoff, 1984)
Aeromonas spp. Distinguishes motile Aeromonas spp. from:
Oxidase-positive
Fermentation of carbohydrates
Growth without added NaCl and resistance to Vibriostatic agent O/129
DNase production, failure to metabolize inositol and resistance to Vibriostatic agent O/129
Motility and growth at 37~
Enterobacteriaceae
Pseudomonas spp.
Halophilic Vibrio spp.
Plesiomonas shigelloides
Aeromonas salmonicida

120
Table 3 Confirmat ion of presumptive Aeromonas spp. (Modified after Palumbo, 1990)
Gram-negative, short rod, motile, oxidase- and catalase-positive
Starch (amylase) +
Manni tol +
DNase +
Gas from glucose + a
Beta haemolytic + a
Resistant to Vibriostatic agent O / 1 2 9 +
Growth at 37~ +
a Aeromonas caviae negative.
The motile Aeromonas spp. can be identified to genus level by conventional biochemical tests, using the Aeromonas hydrophila (AH)medium of Kaper et al. (1979) or by some of the commercially available diagnostic kits, e.g., API 20 E (Abeyta et al., 1986), Minitek (Holmes and Humphry, 1988) or Mast-ID 15 (Holmes and Dawson, 1987). Further identification to species level can however be of great importance in some cases as the biological properties of each group differ significantly (Janda, 1985).
In Table 4 differential characters for distinguishing between A. hydrophila, A. caviae and A. sobria are listed.
3. Isolation, enumeration and identification of Plesiomonas shigel lo ides
C l i n i c a l i s o l a t i o n s o f P. sh ige l lo ides h a v e b e e n m a d e u s i n g m e d i a d e s i g n e d f o r
E n t e r o b a c t e r i a c e a e , b u t t h e s e m e d i a a r e , a s p r e v i o u s l y m e n t i o n e d , n o t i d e a l a n d
Table 4 Different ia t ion between A. hydrophila, A. caviae, A. sobria and Plesiomonas shigelloides (modified after
Popoff, 1984)
Characteristics A. hydrophila A. caviae A. sobria P. shigelloides
Monotr ichous flagellation a + + + _
Lophotr ichous flagellation a _ _ _ +
Esculin hydrolysis + + - -
Growth in KCN broth b + + _ _
Arginine dihydrolase + + - +
L-Arabinose utilization + + - -
Fe rmenta t ion of salicin + + - -
Fe rmenta t ion of sucrose + + + -
Fermenta t ion of manni tol + + + -
Breakdown of inositol - - - +
Acetoin from glucose c + - d -
Gas from glucose + - + -
H 2 S from cysteine + - + -
Lysine decarboxylase + - + +
Orni th ine decarboxylase - - - +
a In liquid medium; ~ Moiler technique; c Voges-Proskauer; d Differs among strains.

121
some medium components may even be toxic (Millership and Chattopadhyay, 1984; Schubert, 1984).
Von Graevenitz and Bucher (1983) evaluated nine solid and two liquid media (the same as for Aeromonas spp.) for their suitability to select P. shigelloides from human faeces and recommended enrichment in alkaline peptone-water (APW), followed by plating on inositol brilliant green bile salts agar (IBB). They found that more than 50% of P. shigelloides strains were inhibited by ampicillin, which precludes the use of media containing this antibiotic. In contrast to von Gravenitz and Bucher, Millership and Chattopadhyay (1984) did not find APW a useful enrichment for P. shigelloides.
Millership and Chattopadhyay (1984) compared xylose deoxycholate citrate agar (XDCA) to bile salts brilliant green agar (BBG) (with inositol omitted) for isolation of P. shigelloides and found that plating on BBG was the preferred method.
Miller and Koburger (1985) designed a plating medium specifically for P. shigelloides and in a preliminary study involving environmental samples, this Plesiomonas agar (PL) seemed comparable to IBB. PL agar gives optimum results after 24 h of incubation, whereas 48 h is recommended for the incubation of IBB agar (Schubert, 1984).
In a later study Miller and Koburger (1986) again evaluated the recovery of P. shigelloides from environmental samples using PL and IBB agars. IBB was then found more effective compared to PL in recovering the organisms from environ- mental samples and IBB also gave fewer false-positive isolates. The growth of P. shigelloides is favoured on IBB agar by providing inositol as a carbon source which can be used by only a few competing bacteria. P. shigelloides is capable of fermenting inositol and will form on IBB red to pinkish colonies in the presence of neutral red indicator, however in over-crowded plates, colonies can appear small and white (Schubert, 1984).
The environmental samples had high levels of contaminating bacteria which were inhibited on IBB, allowing easier recognition and isolation of P. shigelloides than on PL. PL contains low concentrations of bile salts and is not highly inhibitory and relies on differentiating the organisms by alkaline and acid reactions in presence of phenol red indicator; incorporation of lysine and nonfermentable carbohydrates allows for the selection of lysine decarboxylase-positive, nonfer- menting organisms such as P. shigelloides. P. shigelloides forms pink colonies surrounded by a red zone on PL. Growth of pseudomonads can be a problem on PL agar where Pseudomonas spp. also form pink colonies, although less opaque than P. shigelloides. Pseudomonas spp. can easily be distinguished on triple sugar iron agar (TSI) slants.
One important feature of PL medium compared to IBB agar is its ability to recover heat- and cold stressed cells. However recovery of lightly injured cells from environmental samples was apparently not a problem on IBB (Miller and Koburger, 1986). Hence, only if injured cells are known to be present is the routine use of PL agar indicated.
For routine analysis of environmental samples Miller and Koburger (1986) recommend surface plating on both IBB and PL agars with incubation at 35~ for

122
Tab le 5
Dis t ingu ish ing charac ter i s t ics of m e m b e r s of the family V ib r ionaceae (modif ied af ter Mil ler and
Koburger , 1985)
Plesiomonas A eromonas Vibrio
Oxidase + + +
Lysine + - +
Mann i to l - + +
Inosi tol (acid) + - -
O / 1 2 9 Sensit ive Res i s t an t Sensit ive
Ge l a t i na se - + +
Amylase - + +
24-48 h. Suspect colonies should be inoculated into both TSI slants and inositol gelatine deeps. P. shigelloides is oxidase-positive, alkaline over acid on TSI, ferments inositol and does not hydrolyse gelatine. Some of the more important diagnostic characteristics are listed in Table 5. For further confirmatory tests see Table 4.
3.1. Enrichment procedures
The controversial results on the use of APW have already been mentioned (von Graevenitz and Bucher, 1983; Millership and Chattopadhyay, 1984). This contro- versy is further stressed by Freund et al. (1988) who found that APW gave a high recovery rate for P. shigelloides from fresh water but still significantly lower than with tetrathionate broth, whereas van Damme and Vandepitte (1980) on the other hand reported no increase in isolation rate using tetrathionate broth (with and without iodine) for P. shigelloides from freshwater fish.
It seems that the choice of enrichment medium depends on factors such as the level and composition of competing flora and choice of solid medium, and further studies are needed to find out whether an optimal enrichment medium can be recommended.
Incubation at 40~ has been proposed as a selective enrichment step resulting in significantly higher recovery of P. shigelloides (Miller and Koburger, 1986; Freund et al., 1988).
4. Media for Pseudomonas spp.
In some habitats, e.g., in various foods where they are important spoilage organisms, Pseudomonas spp. may represent a minority of the total microflora at the beginning of shelf-life. However under certain conditions, such as high water activity, neutral pH, suitable temperature and a good supply of oxygen, their capacity for rapid growth decides their dominance. Therefore a selective medium

123
for their detection may be of great value in assessing the potential shelf-life of a product.
Pseudomonas spp. grow well on non-selective media (e.g., plate count agar and blood agar) and on routine primary isolation media (e.g., MacConkey agar and eosine methylene blue agar)when incubated at a temperature suitable for their growth.
A medium that is frequently used for the isolation of fluorescent Pseudomonas spp. is medium B of King et al. (1954), which enhances pyoverdin production and the colonies can be identified on plates by a characteristic diffusible pigment under UV-light. Some selective solid media have been developed on the basis of medium B. An example is the medium proposed by Sands and Rovira (1970) containing penicillin G, novobiocin and cycloheximide, which do not inhibit the fluorescent pseudomonads.
A problem associated with the use of media developed for isolation of Pseu- domonas spp. from foods is the considerable interference from non-pseudomonads (Brant, 1975).
Mead and Adams (1977) investigated the selectivity and productivity of four existing media for isolation of Pseudomonas spp. (ALVC, MGV, Difco heart infusion agar with diamide and CETCH). They found that ALCV medium (Gyllen- berg et al., 1960a,b), CETCH medium (Solberg et al., 1972) and MGV medium (Masurovsky et al., 1963) all gave adequate recoveries of 12 pure cultures of pseudomonads isolated from poultry. These media effectively suppressed the growth of Gram-positive organisms, but there were marked differences in their ability to suppress unwanted Gram-negative microorganisms. ALVC medium per- mitted growth of most strains of Gram-negative bacteria, whereas Serratia liquefa- ciens could grow on MGV and Shewanella putrefaciens, Aeromonas spp. and S. liquefaciens grew on CETCH. On MGV the majority of Pseudomonas spp. pro- duced only small colonies and were difficult to count. The Difco medium (Verbov- sky and Collins, 1973) was the least successful, inhibiting most of the non-pig- mented Pseudomonas spp. whilst supporting the growth of certain yeasts.
Mead and Adams (1977) developed a new selective medium based on Difco heart infusion agar with cephaloridine, fucidin and cetrimide as selective agents (CFC), which permits the enumeration of both pigmented and non-pigmented Pseudomonas spp. They found that the medium effectively suppressed Gram-posi- tive bacteria and supported good growth of Pseudomonas spp. (including Pseu- domonas aeruginosa)whilst inhibiting other Gram-negative bacteria, although S. putrefaciens was only partly inhibited.
Mead (1985) pointed out that CFC medium can be used for enumeration of Pseudomonas spp. in various types of food. The medium is inoculated by surface plating and incubated for 48 h at 25~ The medium aims to suppress virtually all unwanted organisms. There are two types of colonies: pigmented and non-pig- mented, 2-5 mm in diameter. Confirmatory tests are not usually required but in cases of doubt flooding the plates with oxidase reagent can be used to distinguish Pseudomonas spp. from other organisms which may be present (e.g., in some cases S. liquefaciens and yeasts).

124
5. Conclusion
Several media are proposed for the detection especially of Aeromonas spp. but also for P. shigelloides and Pseudomonas spp. Some for general purposes and others specifically for isolation from clinical, environmental or food samples.
The selective media generally give a good recovery but, as for all selective media, problems concerning the recovery of injured ceils and inhibition of compet- ing flora are considerable. The media vary in sensitivity and selectivity and their suitability may vary according to the source of the samples and whether the investigator needs qualitative detection or quantitative recovery. The final choice of media and incubation temperature will thus depend on the purpose of the investigations. Achieving an increase in one regard may result in reduction in another.
The best medium for quantitative detection of Aeromonas spp. from food and environmental samples seems to be SAA, although mA and MIX agar as well as ADA, DFS and SGAP-10C appear promising. One essential shortcoming of most of the published comparative studies on selective media for Aeromonas spp. is the omission of one or more apparently suitable media. A reliable standard evaluation of the above mentioned media needs to be done with food and environmental samples.
Conditions of incubation such as temperature and time should be seriously considered in such an evaluation. When examining food and environmental sam- ples, where the competing flora are dominated by Pseudomonas spp. incubation at 37~ for 24 h gives the best results due to an effective inhibition of Pseudomonas spp. (as proposed by Rippey and Cabelli, 1979). At lower temperature the growth of Pseudomonas spp., especially P. fluorescens, will often interfere in a degree that makes quantitative recovery impossible.
For P. shigelloides media and methods for isolation have not been fully investi- gated. For routine analyses of environmental samples surface plating on both IBB and PL at 35~ is recommended.
Specific biochemical tests for confirmation of presumptive isolates of Aeromonas spp. and P. shigelloides are of great importance since food and environmental samples often contain a complex microflora, which can interfere with isolation and confuse identification.
For Pseudomonas spp. none of the media reviewed are sufficiently selective, especially when the organisms are present in low numbers. However CFC medium seems to be recommendable for isolation of Pseudomonas spp. from foods (Baird et al., 1985).
References
Abeyta, C. Jr., Kaysner, C.A., Wekell, M.M., Sullivan, J.J. and Stelma, G.N. (1986) Recovery of Aeromonas hydrophila from oysters implicated in an outbreak of foodborne illness. J. Food Protect. 49, 643-646.

125
Abeyta, C. Jr., Weagant, S.D., Kaysner, C.A., Wekell, M.M., Stott, R.F., Krane, M.H. and Peeler, J.T. (1989) Aeromonas hydrophila in shellfish growing waters: incidence and media evaluation. J. Food Protect. 52, 7-12.
Arcos, M.L., de Vincente, A., Morinigo, M.A., Romero, P. and Borrego, J.J. (1988) Evaluation of several selective media for recovery of Aeromonas hydrophila from polluted waters. Appl. Environ. Microbiol. 54, 2786-2792.
Baird, R.M., Corry, J.E.L. and Curtis, C.D.W. (Eds.) (1987) Pharmacopoeia of Culture Media for Food Microbiology Int. J. Food Microbiol. 5, 187-300.
Baumann, P., Furniss, A.L., and Lee, J.V. (1984) Vibrio. In: N.R. Krieg and J.G. Holt (Eds.), Bergey's Manual of Systematic Bacteriology, Vol. 1, pp. 518-538. Williams & Wilkins, Baltimore, USA.
Brant, A.W. (1975) Selective media for psychrotrophic spoilage bacteria of fresh poultry. 2nd Eur. Symp. Poult. Meat, Oosterbeek, 31.
Callister, S.M. and Agger, W.A. (1987) Enumeration and characterization of Aeromonas hydrophila and Aeromonas caviae isolated from grocery store produce. Appl. Environ. Microbiol. 53, 249-253.
Cunliffe, D.A. and Adcock, P. (1989) Isolation of Aeromonas spp. from water by using anaerobic incubation. Appl. Environ. Microbiol. 55, 2138-2140.
Desmond, E. and Janda, M. (1986) Growth of Aeromonas species on enteric agars. J. Clin. Microbiol. 23, 1065-1067.
Freund, S.M., Koburger, J.A. and Wei, C.-I. (1988) Enhanced recovery of Plesiomonas shigelloides following an enrichment technique. J. Food Protect. 51, 110-112.
Gyllenberg, H., Eklund, E., Antila, M. and Vartiovaara, U. (1960a) Contamination and deterioration of market milk. II. Significance of pseudomonads as deteriorating bacteria. Acta Agric. Scand. 10, 50-64.
Gyllenberg, H., Eklund, E., Antila, M. and Vartiovaara, U. (1960b) Contamination and deterioration of market milk. III. A selective plating test for the demonstration of significant numbers of pseu- domonads. Acta Agric. Scand. 10, 65-73.
Havelaar, A.H., During, M. and Versteech, J.F. (1987): Ampicillin-dextrin agar medium for the enumeration of Aeromonas species in water by membrane filtration. J. Appl. Bacteriol. 62, 279-287.
Havelaar, A.H. and Vonk, M. (1988) The preparation of ampicillin dextrin agar for the enumeration of Aeromonas in water. Lett. Appl. Microbiol. 7, 169-171.
Holmes, B. and Dawson, C.A. (1987) Evaluation of Mast-ID 15 system for identifying Enterobacteri- aceae, some Vibrionaceae, and Acinetobacter. J. Clin. Pathol. 40, 1168-1173.
Holmes, B. and Humphry, P.S. (1988) Identification of Enterobacteriaceae with the Minitek system. J. Appl. Bacteriol. 64, 151-161.
Hudson, J.A. and de Lacy, K.M. (1991) Incidence of motile aeromonads in New Zealand retail foods. J. Food Protect. 54, 696-699.
Huguet, J.M. and Ribas, F. (1991) SGAP-10C agar for the isolation and quantification of Aeromonas from water. J. Appl. Bacteriol. 70, 81-88.
Janda, M.J. (1985) Biochemical and exoenzymatic properties of Aeromonas species. Diagnost. Micro- biol. Infect. Dis. 3, 223-232.
Kaper, J.B., Lockman, H. and Colwell, R.R. (1981) Aeromonas hydrophila: ecology and toxigenicity of isolates from an estuary. J. Appl. Bacteriol. 50, 359-377.
Kaper, J., Seidler, R.J., Lockman, H. and Colwell, R.R. (1979) Medium for the presumptive identifica- tion of Aeromonas hydrophila and Enterobacteriaceae. Appl. Environ. Microbiol. 38, 1023-1026.
Kielwein, G. (1969) Ein N~ihrboden zur selektiven Ziichtung von Pseudomonaden und Aeromonaden. Arch. Lebensmittelhyg. 20, 131-138.
King, E.O., Ward, M.K. and Raney, D.E. (1954) Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 44, 301-307.
Kn6chel, S. (1989) The suitability of four media for enumerating Aeromonas spp. from environmental samples. Lett. Appl. Microbiol. 9, 67-69.
Masurovsky, E.B., Goldblith, S.A. and Voss, J. (1963) Differential medium for selection and enumera- tion of the genus Pseudomonas. J. Bacteriol. 85, 722-723.
McCoy, R.H. and Pilcher, K.S. (1974) Peptone beef extract glycogen agar, a selective and differential Aeromonas medium. J. Fish. Res. Board Can. 31, 1553-1555.

126
Mead, G.C. (1985) Enumeration of pseudomonads using cephaloridine-fucidin-cetrimide agar (CFC). Int. J. Food Microbiol. 2, 21-26.
Mead, G.C. and Adams, B.W. (1977) A selective medium for the rapid isolation of pseudomonads associated with poultry meat spoilage. Br. Poult. Sci. 18, 661-670.
Miller, M.L. and Koburger, J.A. (1985): Plesiomonas shigelloides: an opportunistic food and waterborne pathogen. J. Food Protect. 48, 449-457.
Miller, M.L. and Koburger, J.A. (1986) Evaluation of inositol brilliant green bile baits and Plesiomonas agars for recovery of Plesiornonas shigelloides from aquatic samples in a seasonal survey of the Suwannee river estuary. J. Food Protect. 49, 274-277.
Millership, S.E. and Chattopadhyay, B. (1984) Methods for the isolation of Aeromonas hydrophila and Plesiomonas shigelloides from faeces. J. Hyg., Cambridge 92, 145-152.
Millership, S.E., Curnow, S.R. and Chattopadhyay, B. (1983) Faecal carriage rate of Aeromonas hydrophila. J. Clin. Pathol. 36, 920-923.
Nishikawa, Y. and Kishi, T. (1987) A modification of bile salts brilliant green agar for isolation of motile Aeromonas from foods and environmental specimens. Epidemiol. Infect. 98, 331-336.
Okrend, A.J.G., Rose, B.E. and Bennett, B. (1987) Incidence and toxigenicity of Aeromonas species in retail poultry, beef and pork. J. Food Protect. 50, 509-513.
Palumbo, S.A., Maxino, F., Williams, A.C., Buchanan, R.L. and Thayer, D.W. (1985) Starch-ampicillin agar for the quantitative detection of Aeromonas hydrophila. Appl. Environ. Microbiol. 50, 1027- 1030.
Palumbo, S. (1990) A review of methods for detection of the psychrotrophic foodborne pathogens Listeria monocytogenes and Aeromonas hydrophila. J. Food Safety, 11, 105-122.
Palumbo, S., Abeyta, C. and Stelma Jr., G. (1992) The Aeromonas hydrophila group. In: Compendium of Methods for the Microbiological Examination of Foods. American Public Health Association, Washington, DC, pp. 497-515.
Poff~, R. and Op de Beeck, E. (1991) Enumeration of Aeromonas hydrophila from domestic wastewater treatment plants and surface waters. J. Appl. Bacteriol. 71, 366-370.
Popoff, M. (1984) Genus III. Aeromonas. In: N.R. Krieg and J.G. Holt (Eds.) Bergey's Manual of Systematic Bacteriology, Vol. 1, pp. 545-548. Williams & Wilkins, Baltimore, USA.
Rippey, S.R. and Cabelli, V.J. (1979) Membrane filter procedure for enumeration of Aeromonas hydrophila in fresh waters. Appl. Environ. Microbiol. 38, 108-113.
Rogol, M., Sechter, I., Grinberg, L. and Gerichter, C.B. (1979) Pril-xylose-ampicillin agar, a new selective medium for the isolation of Aeromonas hydrophila. J. Med. Microbiol. 12, 229-231.
Roland, F.P. (1977) Salt-starch xylose deoxycholate agar. A single medium for the isolation of sodium and non-sodium dependent enteric gram-negative bacilli. Med. Microbiol. Immunol. 163, 241-249.
Sands, D.C. and Rovira, A.D. (1970) Isolation of fluorescent pseudomonads with a selective medium. Appl. Microbiol. 20, 513-514.
Schubert, R.H.W. (1967) Das Vorkommen der Aeromonaden in oberirdischen Gew~issern. Arch. Hyg. 150, 688-708.
Schubert, R.H.W. (1977) Ueber den Nachweis von Plesiomonas shigelloides Habs und Schubert, 1962, und ein Elektivmedium, den Inositol-Brillantgriin-Gallesalz-Agar. E. Rodenwaldt-Arch. 4, 97-103.
Schubert, R.H.W. (1984) Genus IV. Plesiomonas. In: Krieg, N.R. and J.G. Holt (Eds.) Bergey's Manual Volume I, pp. 548-550. Williams and Wilkins, Baltimore, USA.
Shotts, E.B., Jr. and Rimler, R. (1973) Medium for the isolation of Aeromonas hydrophila. Appl. Microbiol. 26, 550-553.
Shread, P., Donovan, T.J. and Lee, J.V. (1981) A survey of the incidence of Aeromonas in human faeces. Soc. Gen. Microbiol. Q. 8, 184.
Solberg, M., O'Leary, V.S. and Riha, W.E. (1972) New medium for the isolation and enumeration of pseudomonads. Appl. Microbiol. 24, 544-550.
Stern, N.J., Drazek, E.S. and Joseph, S.W. (1987) Low incidence of Aeromonas sp. in livestock feces. J. Food Protect. 50, 66-69.
van Damme, L.R. and Vandepitte, J. (1980) Frequent isolations of Edwardsiella tarda and Plesiomonas shigelloides form healthy Zairese freshwater fish: a possible source of sporadic diarrhea in the tropics. Appl. Environ. Microbiol. 39, 475-479.

127
van der Kooij, D., Visser, A. and Hijne n, W.A.M. (1980) Growth of Aeromonas hydrophila at low concentrations of substrates added tap water. Appl. Environ. Microbiol. 39, 1198-1204.
Verbovsky, P.D. and Collins, E.B. (1973) Effect of diamide on growth of selected bacteria. J. Dairy Sci. 56, 1180-1182.
von Graevenitz, A. and Bucher, C. (1983) Evaluation of differential and selective media for isolation of Aeromonas and Plesiomonas spp. from human faeces. J. Clin. Microbiol. 17, 16-21.
von Graevenitz, A. and Zinterhofer, L. (1970) The detection of Aeromonas hydrophila in stool specimens. Health Lab. Sci. 7, 124-126.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Edsl ) �9 1995 Elsevier Science B.V. All rights reserved
129
Chapter 9 Culture media for the isolation of campylobacters
J a n e t E .L . C o r r y a, D . E . P o s t b, p. C o l i n ~, M. J . L a i s n e y c
a Department of Clinical Veterinary Science, University of Bristol, Langford, Avon, BS18 7DU, UK b Unipath Limited, Wade Road, Basingstoke, Hants. RG24 8PW, UK
c National Centre for Veterinary and Food Studies (CNEVA), Central Laboratory for Poultry and Pig Research (LCRAP), Beausemaine, B.P. 53-22440, Ploufragan, France
Abstract
The history of the development of selective media for isolation of campylobacters, including the rationale for choice of selective agents is described. Developments have included modifications to allow incubation at 37~ instead of 42 or 43~ and changes in the types and concentrations of antibiotics in order not to inhibit organisms such as Campy- lobacter upsaliensis, C. jejuni subsp, doylei and some strains of C. coli and C. lari. When examining foods, plating media originally developed for isolation from faeces are normally used, sometimes after liquid enrichment. Most of the media include ingredients intended to protect campylobacters from the toxic effect of oxygen derivatives. Most commonly used are lysed or defibrinated blood; charcoal; a combination of ferrous sulphate, sodium metabisul- phite and sodium pyruvate (FBP); and haemin or haematin. To date no medium includes an indicator system - for instance a pH indicator to show whether colonies produce acid or alkali from particular substrates. The manner in which liquid enrichment media are used has been modified for food samples to avoid inhibitory effects on sublethally damaged cells by toxic components in the formula. This is done by a preliminary period of incubation at reduced temperature and sometimes by delayed addition of antibiotics. Expensive and time-consuming methods have been proposed to achieve a microaerobic atmosphere while using liquid enrichment media. To date there is no generally accepted 'standard' method of isolating campylobacters from food.

130
Campylobacters were originally classified within the genus Vibrio, but differ from vibrios in a number of respects, particularly in their DNA base composition and their ability to grow under conditions of reduced oxygen tension. They are Gram-negative, oxidase-positive, curved or spiral rod-shaped bacteria 0.2-0.5 ~m wide and 0.5-8 /zm long. They possess one polar flagellum, which gives them a very characteristic 'cork-screw' motility. Survival of campylobacters is poor under most conditions and particularly in dry, relatively warm and aerobic situations. As cultures lose their viability the rod-shape changes to a coccoid form which has been described as 'viable but non-culturable'. This is because, although these forms cannot be grown using normal media, at least some have been shown to be capable of initiating infections in animals (e.g. Jones et al., 1991)
The genus Campylobacter has recently been subdivided into three related genera: Campylobacter, which contains the species C. fetus, C. jejuni, C. coli, C. lari (NARTC, formally known as 'nalidixic acid resistant thermophilic campylobac- ters'), C. hyointestinalis, C. upsaliensis, C. mucosalis, C. concisus, C. sputorum; Helicobacter which contains H. pylori (previously C. pyloridis, responsible for human gastric infection and gastric ulcers) H. cinaedi, H. fennelliae and H. mustelae; Arcobacter which contains A. cryaerophilus, A. butzleri, A. nitrofigilis and A. skirrowii (Vandamme et al., 1991, 1992; Vandamme and Goossens, 1992; Tenover and Fennell, 1992). Table 1 lists the most important species of this group, which are associated with human and /o r animal diarrhoea together with their important characteristics.
Selective media were originally designed to isolate C. jejuni from faeces, by use of a cocktail of antibiotics in a rich basal medium and exploiting the ability of this organism to grow at 42 or 43~ (Butzler and Skirrow, 1979). Later C. coli and C. lari (formerly C. laridis) were distinguished, although many workers either failed to differentiate between these three species when monitoring the functioning of selective media or only tested for C. jejuni (e.g. Park et al., 1981; Rosef, 1981; Acuff et al., 1982a,b; Bolton et al., 1982; Goossens et al., 1983; Martin et al., 1983; Wesley et al., 1983; Barot and Bokkenhauser, 1984; Stern et al., 1984; Beuchat et al., 1985; Furlanetto et al., 1991; Holler, 1991; Jacob and Stelzer, 19~2; Stern et al., 1992; Stern and Line, 1992; Scotter et al., 1993).
There is evidence that some strains of C. coli and even a few strains of C. jejuni are likely to have been missed due to their sensitivity to cephalothin (Brooks et al., 1986; Ng et al., 1985, 1988). More recent work has indicated that other species of campylobacter of importance in human intestinal disease will not be isolated by the usual selective media because of their susceptibility to the antibiotics used in most media e.g.C, upsaliensis (Warmsley and Karmali, 1989; Goossens et al., 1990a). In addition a closely related group of organisms unable to grow at 42 or 43~ the normal isolation temperature, and recently reclassified as Arcobacter rather than Campylobacter spp. on the basis of their aerotolerance and various genotypic tests (Vandamme et al., 1991, 1992), are increasingly being implicated as important causes of human gastro-intestinal disease (Patton et al., 1989; Lastovica et al., 1989; Kiehlbauch et al., 1991; Taylor et al., 1991). At present, therefore, Arcobacter species and C. upsaliensis are unlikely to be detected as causes of

131
human (or animal) diarrhoea, neither would they be detected in routine examina- tion of human or animal faeces, foods, water or environmental specimens unless membrane filtration and non-selective media at 37~ are used. Recently, however, a modification to blood-free selective agar has been described which enables C. upsaliensis to be isolated, thus removing the practical difficulties of the filtration method (Aspinall et al., 1993). Between 1989 and 1991 only 15% of thermophilic campylobacters reported as causes of human diarrhoea in England and Wales were identified to species level. Of these 89-93% were C. jejuni and 7-10% were C. coli or C. lari, with 0.2-0.4% other species of campylobacter (Pearson and Healing, 1992). However, Goossens et al. (1990b) reported C. upsaliensis to be the cause of 13% of campylobacter diarrhoea cases in Belgium. C. jejuni subsp, doylei is another strain implicated as a cause of diarrhoea that frequently does not grow at 42~ and is sensitive to cephalothin (Taylor et al., 1991; Bolton et al., 1992). These considerations should be borne in mind when reading this review, which will be concerned almost exclusively with the thermophilic campylobacters.
The media used for isolating campylobacters from foods and water have been derived from those first developed for the isolation of campylobacters from faeces. In some cases the same plating media are still used for both purposes. In line with techniques developed for isolating other pathogens, such as salmonellas, from foods and other environments, liquid (enrichment) media have been developed, and also pre-enrichment media, intended to aid recovery of sublethally damaged campylobacters. Incubation has usually been carried out at 42 or 43~
Besides a variety of selective agents, almost all of which are antibiotics, media for campylobacter usually contain ingredients to neutralise the toxic effects of substances that form in the presence of oxygen and light. In addition, almost all workers have found it necessary to incubate plates in an atmosphere of about 5-7% oxygen, 10% carbon dioxide and 80% nitrogen and /or hydrogen. This can be achieved by using a bottled gas mixture of those proportions, by using the appropriate gas-generating envelope or by replacing two thirds of the atmosphere with a mixture of either nitrogen or hydrogen plus 5-15% carbon dioxide (Skirrow et al., 1982). Skirrow et al. (1991) reported that the presence of hydrogen at not less than 7% improved the primary isolation of C. jejuni from faeces. Candle jars have been reported to be successful for incubation of plates of Butzler's medium Virion, provided 37~ is used (Goossens et al., 1983). Ribeiro et al. (1985) compared growth of C. jejuni, C. coli and C. lari in atmospheres generated by burning ethanol (methylated spirit) rather than a candle (paraffin wax) and found higher colony counts and larger colonies using ethanol. Colony counts for C. lari were particularly low when using candles. Similar comparisons by Skirrow et al. (1987) confirmed these results. Best results were obtained when the spirit was burned in a 90 mm diameter Petri dish with the jar not more than half full with dishes. Pennie et al. (1984) found that a satisfactory atmosphere could be obtained by using a mixture of grade 0 steel wool, previously soaked in cupric sulphate solution and an Alka-Seltzer (sodium bicarbonate) tablet in water, all placed in a plastic bag! Jacob and Stelzer (1992) reported that this method worked well.
Recent work by Jones et al. (1993) has shown that C. jejuni may adapt to grow
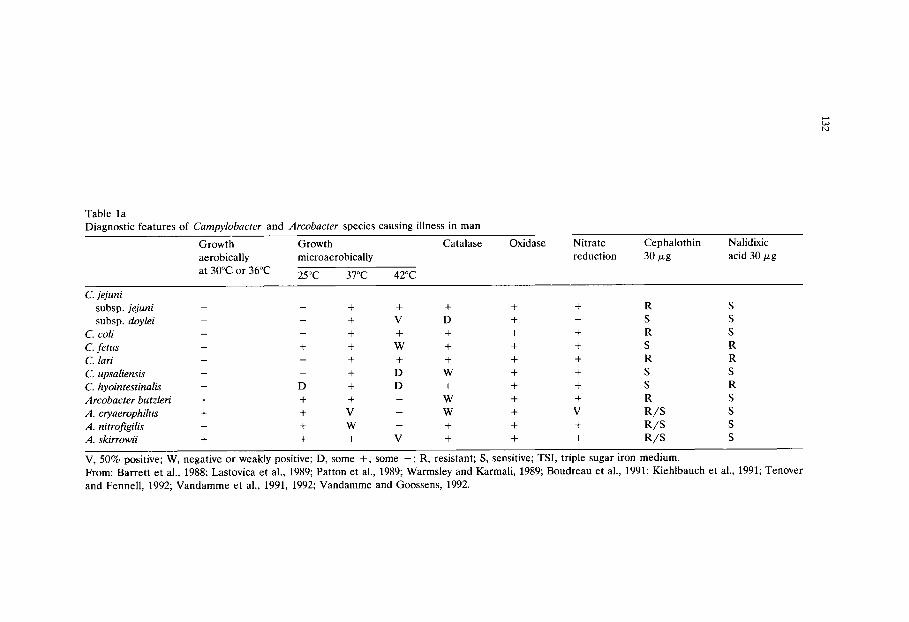
Table la Diagnostic features of C a m p y l o b a c t e r and A r c o b a c t e r species causing illness in man
Growth Growth aerobically microaerobically
at 30~ or 36~ 25oc 37oc 42oc
Catalase Oxidase Nitrate Cephalothin Nalidixic reduction 30/zg acid 30/zg
C. j e j u n i
subsp, j e j u n i - - + + + + + R S
subsp, d o y l e i - - + V D + - S S
C. col i - - + + + + + R S
C. f e t u s - + + W + + + S R
C. lari - - + + + + + R R
C. u p s a l i e n s i s - - + D W + + S S
C. h y o i n t e s t i n a l i s - D + D + + + S R
A r c o b a c t e r b u t z l e r i + + + - W + + R S
A . c r y a e r o p h i l u s + + V - W + V R / S S
A . n i t ro f ig i l i s + + W - + + + R / S S
A . s k i r r o w i i + + + V + + + R / S S
V, 50% positive; W, negative or weakly positive; D, some +, some - ; R, resistant; S, sensitive; TSI, triple sugar iron medium. From: Barrett et al., 1988; Lastovica et al., 1989; Patton et al., 1989; Warmsley and Karmali, 1989; Boudreau et al., 1991; Kiehlbauch et al., 1991; Tenover and Fennell, 1992; Vandamme et al., 1991, 1992; Vandamme and Goossens, 1992.
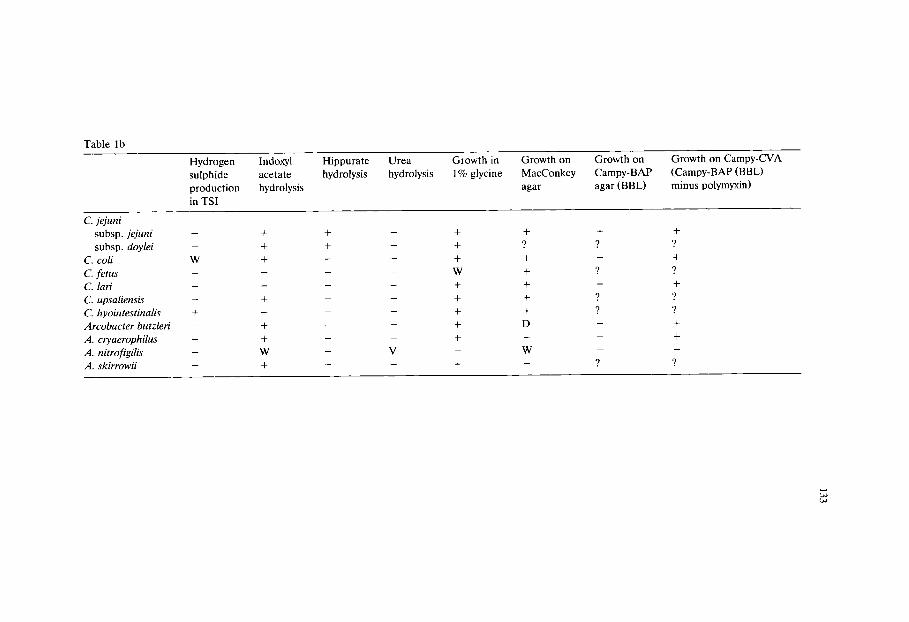
Table lb
Hydrogen sulphide production in TSI
Indoxyl acetate hydrolysis
Hippurate hydrolysis
Urea hydrolysis
Growth in 1% glycine
Growth on MacConkey agar
Growth on Campy-BAP agar (BBL)
Growth on Campy-CVA (Campy-BAP (BBL) minus polymyxin)
C. jejuni subsp, jejuni
subsp, doylei
C. coli W
C. fe tus C. lari
C. upsaliensis
C. hyointestinalis +
Arcobacter butzleri -
A. cryaerophilus
A. nitrofigilis A. skirrowii
+
+
+
+
+
+
W +
+
+
+
W +
+
+
+
+
+
+
9 +
+
+
+
+
D
W

134
Table 2 Most widely used isolation media for thermophilic campylobacters
Plating Media
Skirrow (Skirrow, 1977; see also this volume) Butzler (Lauwers et al., 1978) Campy BAP (Blaser et al., 1978) Preston agar (Bolton and Robertson, 1982; this volume) Butzler (Virion) (Goossens et al., 1983) Butzler (Oxoid) (Goossens et al., 1983) mCCD agar (Bolton et al., 1984 modified according to Hutchinson and Bolton, 1984; see also this volume) Karmali agar (Karmali et al., 1986) Semi-solid medium (Goossens et al., 1989)
Enrichment Media
Preston broth (Bolton et al., 1982; see also this volume) Doyle and Roman broth (1982) VTP FBP broth (a modification by Lovett et al., 1983, of the medium of Park et al., 1981) mCCD broth (Bolton et al., 1984, modified according to Hutchinson and Bolton, 1984; see also this volume) Park and Sanders broth (1991) Exeter broth (De Boer and Humphrey, 1991) Hunt and Radle broth (Hunt, 1992)
in a normal air atmosphere. The implications of this observation are not yet clear, but may explain to some extent the 'viable but non-culturable' phenomenon. Fraser et al. (1992) reported that, provided the relative humidity was 99%, normal air containing 10% carbon dioxide gave larger colonies of C. jejuni and C. coli than the conventional gas mixture. However at 95% RH growth was poor. Other workers have reported that C. jejuni, as evidenced by its colonial morphology, is sensitive to the moisture level in plating media (Buck and Kelly, 1981), although changes in colonial morphology could also be due to inhibitory oxygen derivatives formed during storage and drying of plates.
No campylobacter media with indicator systems for identification of presump- tive campylobacter colonies have yet been developed. Experienced workers gener- ally claim to be able to recognise the organism by the typical appearance of colonies - flat, glossy and effuse, thinly spreading if the agar is moist. However, colonies are sometimes atypical, especially if plates are rather dry, so Gram staining of oxidase-positive colonies and examination by microscope for the charac- teristic morphology is advisable.
The number of formulations proposed for the isolation of thermophilic campy- lobacters probably exceeds that for any other group of bacteria, especially if one considers that all have been published since 1972 and almost all since 1977. Table 3 summarises many of these. Before the medium of Dekeyser et al. (1972) was available, isolation of 'related Vibrio' (as campylobacters were then known) de- pended on the use of membrane filtration followed by subculture onto nutritionally

135
rich blood agar. The method of Dekeyser and co-workers used a combination of centrifugation, filtration through a 0.65 ~m membrane filter and plating of the filtrate onto a selective agar.
Even if all the campylobacter media formulae had been published with a full description of the rationale used in their development, there would be insufficient space to consider them in detail. This review will examine the basic principles the media have in common and then concentrate on the details of the most widely used ones. The commonly used plating and liquid media are listed in Table 2. Filtration methods have frequently been used to concentrate campylobacters in samples of water (Bolton et al., 1982; 1987; Ribeiro and Price, 1984; Rosef et al., 1987; Stelzer et al., 1988; Brennhovd et al., 1992). Filtration is sometimes followed by liquid enrichment, sometimes by direct plating onto selective agar. Centrifuga- tion has also been used when examining food samples (Lovett et al., 1983).
1. Basal media
Few publications provide an explanation for the choice of basal medium and many have been used. Although campylobacters will grow in relatively simple media such as nutrient agar, most workers have used basal media developed for other fastidious capnophilic or anaerobic pathogens such as brucella or thioglycol- late medium, Columbia or blood agar base and Miiller Hinton broth. However, Bolton and Robertson (1982) chose nutrient broth no. 2, an unsophisticated medium, as the basis for their Preston media because it contained less thymidine, an antagonist to the activity of the selective agent trimethroprim, used in most selective solid and liquid media (see Table 3).
2. Blood
Most solid media incorporate blood at levels between 5 and 15%. Some media use defibrinated blood from various animals and others lysed horse blood. Only one uses lysed sheep blood (Stelzer and Jacob, 1992). Skirrow et al. (1982) stated that lysed horse blood was needed to neutralise trimethoprim antagonists which are present in most media. Many media which incorporate trimethoprim do not contain lysed horse blood although blood of some type is usually present. The media of Martin et al. (1983) and Goossens et al. (1989), neither of which contain blood, have the highest concentrations of trimethoprim. Presumably this is neces- sary to counteract the thymidine. Blood is also thought to be active in neutralising toxic oxygen derivatives (Juven et al., 1985; Weinrich et al., 1990).
3. FBP and other bloodless supplements
A combination of ferrous sulphate (FeSO4.7H20), sodium metabisulphite and sodium pyruvate (FBP), each at 0.25 or 0.5 g per litre was suggested by George et

Table 3
Formulat ion of liquid and solid selective media for thermophilic campylobacters (concentrations in mg per litre unless otherwise stated)
Medium n a m e / Basal Antioxygen Cephalo- Trimetho- Polymyxin B or Vanco- reference, medium system sporins prim Colistin (C) mycin or
Other details in iu Teico-
planin (T)
Dekeyser A 1 T A 15% S 10000 Skirrow A 2,a BAB 7% LH 5 2500 10 Blaser A 3 BA 10% S 5 2500 10 Butzler A 4 TB 10% S 15 cthin * 10000 (C) CAMPY-BAP A 5 BA 10% S 15 cthin 5 2500 10 Blaser-Wang A 6 BAB 7% LH 15 cthin 5 2500 10 C-2 A 7 BA 5% H / F B P 5 cthin 5 8000 10 C-3 A 7 BA 5% H / F B P 1 cthin 5 2500 10 Park (1981) B 8 BB - 4 8000 8 BU40 B 9 TB 10% S 15 cthin 40000 (C) Rosef B 10,b NB - 5 2500 10 Preston A ll(a) NB 5% LH 10 5000 Preston B 11(b) NB 5% L H / F B P 10 5000 Doyle-Roman B 12, c BB 7% LH 5 20000 15 Chris topher B + A 13 BB 0.5g P 15 cthin 5 2500 10 FBP-AM B / A 14 B B / A FBP (B) 15 cthin 5 2500 10
5% DH (A) Lander B 15 VI 5g C 10 50000 40 Park-Stankiewicz B 16,d BB 2g F, 0.25g B, 30 cthin 4 3 mg (C) 8
0.5g P Butzler medium Virion A 17 CA 5 - 7 % S 15 czone 10000 (C) Butzler medium Oxoid A 18 CA 5 - 7 % S 15 czolin 10000 (C) 5 Modified Park (1981) B 19 BB FBP 7.5 5000 15 CEB B 20 BB - 32 czone 32 VTP-FBP B 21 BB FBP, 5% LH 10 5000 20 CAK A 22 GA 7% H 15 cthin Rosef-Kapperud A 23 GA, IVx - 15 cthin 10 000 (C) A T B 24,e T; YE, FBP, HT 6.25 codin 20000
NaC1 CCD A 25,f NB 4g C, 0.25g F, 10 czolin
0.25g P CCD B 25,f NB 4g C, 0.25g 10 czolin
FBP
Rifam- Novo- 5-Fluoro- picin biocin uracil or
Na-deoxy- cholate (D)
5
2 5
5
10 10
100
10
333
25
1000 (D)
lOOO (D)
Bacitracin Anti- in iu fungals
25000 50 CY
25000 50 CY 2 AM 2 AM
25000
25000
50 CY
100 CY 100 CY 50 CY
2 AM 2 AM
100 CY
2 AM 50 CY
1 AM 25 000 iu NY
mCCD A 26,f NB
Doyle-Roman B 27, g BB Steele-McDermott 28 Waterman A 29 BAB Waterman B 29 TB Virion 30 CA Exeter A + B 31,h NB
Karmali A 32,i CA
4g C, 0.25g F, 0.25g P
10%S + FBP 5% DH 5% DH 5 -7 % S 5% LH, 0.5g F, 0.2g B, 0.2g P 4g C, 0.32g HT, 0.1g P
32 czone
30 czone 15 czone
32 czone
5 5
20
10
2.9 mg (C) 2500
10 000
4 mg (C)
10
10 x
20
10
10 10 y
1000 (D) 10 AM
100 CY
2 AM
CAR B / A 33 B A / B B FBP, 3% LH 32 czone 10 2.5 AM Exeter A + B 34 NB FBP, 5% LH 15 czone 4 mg 10 2 AM SSM B + A 35 MHB + - 30 czone 50
4 g/1 A A Bolton 36,j 5% LH, 0.1g HE, 20 czone 20 50 CY
0.5g P, 0.5g B Park-Sanders B 37 BB 0.25 P, 1 g Na 32 czone * * 10 10 100 CY * *
citrate, 5% LH Hunt-Radle B 38 NB, YE FBP, 5%LH 15 czone ** * 12.5 10 100 CY
or 2 AM 200 CY Campy Cefex A 39 BA 0.5 g F, 0.2 g B, 33 czone
0.5 gP , 5% LH CA T A 40,f NB 4g C 8 czone 4 (T) 1000 (D) 10 AM
Key: C O L U M N 1. References: 1 Dekeyser et al., 1972; 2 Skirrow, 1977; 3 Blaser et al., 1978; 4 Lauwers et al., 1978; 5 Blaser et al., 1979; 6 Blaser et al., 1980; 7 Gilchrist et al., 1981; 8 Park et al., 1981; 9 Patton et al., 1981" 10 Rosef, 1981; ll(a)Bolton and Robertson, 1982; ll(b)Bolton et al., 1982; 12 Doyle and Roman, 1982; 13 Christopher et al., 1982; 14 Ehlers et al., 1982; 15 Lander, 1982; 16 Park and Stankiewicz, 1982; 17 Butzler et al., 1983; 18 Goossens et al., 1983; 19 Lovett et al., 1983; 20 Martin et al., 1983" 21 Park et al., 1983; 22 Rosef et al., 1983a; 23 Roseff and Kapperud, 1983; 24 Wesley et al., 1983" 25 Bolton et al., 1984b; 26 Hutchinson and Bolton, 1984; 27 Ray and Johnson, 1984b; 28 Steele and McDermott, 1984; 29 Waterman et al., 1984; 30 Goossens et al., 1986; 31 Humphrey, 1986b; 32 Karmali et al., 1986; 33 Weber et al., 1987; 34 De Boer and Humphrey, 1989; 35 Goossens et al., 1989; 36 Bolton, personal communication; 37 Park and Sanders, 1981" 38 Hunt and
Radle, 1992 (cited by Stern and Line, 1992); 39 Stern et al., 1992; 40 Aspinall et al., 1993. A, agar; B, broth; SSM, semi-solid selective motility medium; ATB, alkaline tryptone broth. Other details (pH, growth factors, other additives): a pH 7.3-7.4; b yeast extract, resazurin; c Na succinate, cysteine HC1; d 3% calf serum; e 10 g bicine, pH 8.0; f casein hydrolysate; g modified protocol: add polymyxin after 6 h at 37~ for freeze-stressed campylobacters; h antibiotics added after 4 h at 37~ i pH 7.4, 1 g Na
deoxycholate; j a-ketoglutamic acid, NaCO 3. C O L U M N 2. Basal medium: TA, thioglycollate agar; TB, thioglycollate broth base; BA, brucella agar base; BB, brucella broth base; NB, nutrient broth; BAB, blood agar base; VI, veal infusion; CA, Columbia agar base; GA, gonococcus agar base; T, tryptose; MHB, Mueller-Hinton broth; YE, yeast extract; IVx, IsoVitalex (vitamin and nutrient mixture (BBL)); A, 10 g meat peptone, 5 g lactoalbumin hydrolysates, 5 g yeast extract, 5 g NaC1, 0.6 g NaCO 3. C O L U M N 3. Antioxygen system: S, sheep blood; H, horse blood; LH, lysed horse blood; DH, defibrinated horse blood; C, charcoal; F, ferrous sulphate (FeSO4-7H20) ; B, sodium metabisulphite (NaHSO0; P, sodium pyruvate. At 0.5 g per litre (unless otherwise stated): HT, haematin; HE, haemin. C O L U M N 4. Cepha lospor ins ; cthin, caphalothin ; ezolin, cephazolin; czone,cefoperazone; codin, cefsulodin. * 15 mg cefalexin instead of cephalothin in recipe
of Butzler and Ski r row, 1979 (used wi thout cephalospor ins by Butzler et al., 1973). ** added later. *** + 15 later. x broth only. y agar only. C O L U M N 12. Antifungals: CY, cycloheximide (actidione); AM, amphotericin B; NY, nystatin.

138
al. (1978) and Hoffman et al. (1979a) as an addition to campylobacter media to counteract the toxic effect of oxygen (Hoffman et al., 1979b). Many selective media contain some or all of these compounds but concentrations vary. A few media contain both FBP supplement and blood (e.g. Gilchrist et al., 1981; Weber et al., 1987; Stern et al., 1992) but in most cases media contain either blood or FBP. Alternatives to blood or FBP include haematin (Razi and Park, 1979; Wesley et al., 1983; Karmali et al., 1986) and charcoal (Bolton et al., 1984; Karmali et al., 1986). Lignite-derived humic acids plus 0.05% FeSO4.7H20 have been proposed as an alternative system (Weinrich et al., 1990). The mode of action of these supple- ments is not clear, but they may help to neutralise hydrogen peroxide, singlet oxygen and /o r superoxide ions.
4. Storage of campylobacter plating media
Hoffman et al. (1979a) found that growth of campylobacters was substantially reduced when plates of nutrient medium were stored in the presence of light and, in particular, air. Similar results were obtained by Bolton et al. (1984) and Juven and Rosenthal (1985). The incorporation of aerotolerant supplements such as FBP or blood into campylobacter media reduces this effect but does not completely eliminate it (Fricker, 1985; Weinrich et al., 1990). Preston agar stored aerobically at room temperature inhibits the growth of C. lari to a greater extent than C. jejuni or C. coli (Fricker, 1985). Loss of moisture is probably also a factor (Buck and Kelly, 1981; Fraser et al., 1992). Campylobacter plates prepared in the laboratory should therefore either be used immediately or after storage in the dark anaerobi- cally at room temperature, or aerobically in the refrigerator with precautions to prevent dehydration. A maximum storage time of 5 days for laboratory prepared plates is probably advisable.
5. pH
The pH of many of the isolation media is not specified, but presumably approximates to that of the basal medium used, normally near neutrality. Some liquid enrichment media have a pH ca. 8.0 (Wesley et al., 1983) although Park et al. (1983) found that their medium functioned best at pH 7.0. High pH media could be used to counteract acid production by competitive flora. The ability of competitive flora to acidify the enrichment media may depend on the food under examination (Humphrey, 1986b).
6. Choice of antibiotics
Antibiotics in selective media developed for campylobacters were chosen on the basis of those to which test strains were resistant and those most effective in

139
inhibiting competitive flora. Table 4 lists the results of various published studies on the resistance of campylobacters to antibiotics. Neither Butzler (Dekeyser et al., 1972; Butzler et al., 1973; Lauwers et al., 1978), Skirrow (Skirrow, 1977; Butzler and Skirrow, 1979) nor Blaser (Blaser et al., 1979) published the full rationale for the development of their media. Probably these authors used data from Plastridge et al. (1964), Butzler et al. (1974) and Vanhoof et al. (1978). Later workers will have had access to other studies listed in Table 4. Plastridge et al. examined 57 strains of 'related vibrio' bacteria that probably belonged to the C. jejuni/C, coli group, finding them resistant to bacitracin, novobiocin and polymyxin B. Butzler et al. (1974) found over 90% of 114 strains of related vibrios resistant to cephalothin and all were resistant to vancomycin and rifampicin. With few exceptions (Ng et al., 1985 and presumably Plastridge et al., 1964) the strains tested had been isolated using antibiotic-containing selective media and so the possibility of missing strains sensitive to the antibiotic used in the selective media will have been perpetuated.
7. Antibiotic activity
Polymyxin is generally active only against Gram-negative bacteria, and Proteus spp. are sometimes resistant. Trimethoprim usually inhibits Proteus spp. as well as other Gram-negative bacteria. Colistin is closely related to polymyxin, has a similar spectrum of activity and is used in some media instead. Vancomycin and rifampicin are both used, but not usually together. They are effective against Gram-positive bacteria. Rifampicin is also active against Gram-negative organisms. Butzler's media have generally incorporated bacitracin instead of vancomycin. The cephalosporins (cephalothin, cefaperazone) have a wide spectrum of activity against Gram-positive bacteria. Cefoperazone is now usually preferred because some campylobacters, especially C. coli, are sensitive to cephalothin (Brooks et al., 1986; Ng et al., 1985, 1988; Burnens and Nicolet, 1992). Cycloheximide (actidione), amphotericin B or nystatin are present in many media to inhibit yeasts and moulds.
8. Development of plating media (Table 3)
Bolton and co-workers gave the most detailed rationale for their formulation of Preston and CCD (charcoal, cefazolin (later replaced by cefoperazone) deoxy- cholate) media. When developing Preston medium, Bolton and Robertson (1982) carried out a survey of the MIC's of 104 strains of Campylobacter spp. and a variety of competitive organisms against four antibiotics (polymyxin, rifampicin, vancomycin and trimethoprim). They chose polymyxin because of its activity against Gram-negative bacteria (except for Proteus spp., which were suppressed by the trimethoprim). Because rifampicin showed a wide spectrum of activity against Gram-positive and Gram-negative bacteria it was chosen in preference to van- comycin, which has limited activity against Gram-negative organisms. Ampho-

Table 4a Campylobacter antibiotic resistance profiles obtained by various authors (mic, mg/1 required to inhibit)
Authors Plastridge et al., 1964 Vanhoof et al., 1978 Walder et al., 1979
Species of jejuni (57) * coli (61) jejuni (95) jejuni (100) Campylobacter (grp II) (grps III & IV)
Karmali et al., 1980
jejuni (60) fetus (12)
ampicillin cloxacillin moxalactam cefazolin cefotaxime cephalothin cefoxitin cephalexin cefuroxime cefoperazone cefamandole chloramphenicol polymyxin-B colistin bacitracin trimethoprim rifampicin vancomycin novobiocin sulphamethoxazole METHOD
1024 4-16
128 64
128 256-512
Nutrient agar+ 5% ox blood
< 0.195-50 2.5-40 > 100
6.25- > 100 6.25- > 100 6.25- > 100 6.25- > 100
0.39-6.25 1.56-100 6.25- > 100
50-200 6.25- > 100
50- > 100
1.95-250 Mueller-Hinton agar (for trimethoprim & sulphamethoxazole plus 5% lysed horse blood)
5-40 > 160
40-160 40-160
3.1-12.5
> 160
5-80 Mueller-Hinton agar + 5% human blood
Key: DST, diagnostic sensitivity test agar, IST, isosensitivity test agar; * number of strains tested; ** iu per ml.
128- >~ 512 8-32 2-32 4-16
128- >I 512 16-32 32- >1 512 32-64 16- >~ 512 32-128
128 >~ 512 16-64
DST + 5% lysed horse blood

Table 4b
Authors
Species of Campylobacter
ampicillin cloxacillin moxalactam cefazolin cefotaxime cephalothin cefoxitin cephalexin cefuroxime cefoperazone cefamandole chloramphenicol polymyxin-B colistin bacitracin trimethoprim rifampicin vancomycin novobiocin sulphamethoxazole METHOD
See Table 4a for key.
Ahonkai et al., 1981
jejuni (36) *
2-8
4-32
2-4
128
2-8
32-128 128
Mueller-Hinton broth + laked blood
Karmali et al., 1981 Michel et al., 1983
jejuni
(157) * (105)
(95) (60)
(106) (108) (107) (104)
(107) (142)
(97)
jejuni (103) *
0.125-64
Ng et al., 1985
1-256 64- >/512
2-128 128- >/512
1-32 64- >1 512 32- >/512 16- >~ 512
128- >f 512 0.5-8 1-32
(139) (56) (91) (77) (88) (55)
0.5- >~ 64 1-32 0.5-32
256- ~ 512
~512
128- ~ 256
jejuni (6) *
>/512 256- >~ 512
8-128 128- ~ 512
64- ~512 8-256
DST + 5 % lysed horse blood
8-512 IST + 7% lysed human blood
2-8 (16-64iu) 4-32
> 512 256
32-128 > 128
32-64
Muller-Hinton agar
coli (24) *
32-256
0.5-8 (4.6iu) 2-32
> 512 128-256
2-128 > 128
1-128

Table 4c
Au thor s
Species of Campylobacter
G e b h a r t et al., 1985
coli (10) * hyointest- inalis (1 O) *
Ng et al., 1988 Kiehlbauch et al., 1992
jejuni (60) * coli (101) * cryoaerophila (12) * butzleri (64) *
Pat ton et al., 1989
upsaliensis (12) *
ampicill in cloxacillin
moxalac tam cefazolin cefotaxime cepha lo th in
cefoxitin cephalexin cefuroxime ce foperazone ce famandole ch lo ramphen ico l polymyxin-B
colistin baci t racin t r imethopr im rifampicin vancomycin novobiocin su lphamethoxazo le
M E T H O D
4.8 0.25-0.5
128
128 128
32-64
1 6 - 6 4
16-64
64-128 32 - > 256
2 - 1 6 2 -16 6 4 - >/128 64 - >/128 < 1-8 < 1 -8
>/128 ** >~ 128 ** 1-16 < 1 -16 >~ 128 ** >~ 128 ** 256- > 512 256-512 >~ 128 64 - >1 128 256- > 256 256- > 256
32 -64 32-64 4-128 16- > 128 >/128 >t 128 128- > 128 128- > 128
16-32 32-64 16-128 8 -128
Mue l l e r -H in ton agar Muel le r -Hin ton agar
8 - > 32 4 - > 32
8 - > 64 4 - > 64 > 32 3 2 - > 32
16- > 32 8 - > 32 16- > 32 16- > 32
4 - > 32 8 - > 32 2.5 ** ~ 2 . 5 **
> 50 > 50 10 * * ~ 10 * *
> 10 > 10
Muel le r -Hin ton broth + 5%
lysed horse blood
1-32
0 .5-4 1-32 1-8
1-16 2 - 8
32
See Table 4a for key.

143
tericin B was added to inhibit yeasts. The level of polymyxin was set at 5000 iu per litre on the basis of Vanhoof's results, which showed that some campylobacters were sensitive to 10000iu per litre. When developing their CCD medium Bolton and co-workers determined that campylobacters grow best on solidified nutrient broth no. 2, after comparison with other media such as Columbia blood agar base, veal or brain heart infusion and diagnostic sensitivity test agar. They then carried out a systematic survey of alternatives to blood for neutralising oxygen toxicity by comparison with the growth of campylobacter obtained on nutrient agar plus 5% lysed horse blood (Bolton and Coates, 1983; Bolton et al., 1984a). A combination of 0.4% charcoal, 0.25% ferrous sulphate and 0.25% sodium pyruvate was best. A further study (Bolton et al., 1984b) surveyed the effect of 11 dyes, 17 chemical compounds and 14 chemotherapeutic agents on one strain each of C. jejuni biotype 1, C. jejuni biotype 2, C. coli and a NARTC (nalidixic acid resistant thermophilic campylobacter - C. lari) and a selection of Gram-positive and Gram-negative competitive bacteria. No details were given of either the competi- tive test strains or the inhibitors examined, but deoxycholate and cefazolin were chosen as the most effective inhibitory agents. Casein hydrolysate was found necessary to stimulate the growth of environmental NARTC (C. lari) strains. CCDA, apart from Karmali's medium, is the only widely used plating medium not using blood. This is useful, particularly for laboratories specialising in food microbiology in which the use of blood in media is uncommon. Blood is expensive, has a short shelf life and is easily contaminated. Later Hutchinson and Bolton (1984) replaced cefazolin (10 mg/1) by cefoperazone (32 mg/1). This allowed fewer contaminants to grow, and permitted the modified medium (mCCDA) to be used at 37~ However, amphotericin B was needed to prevent overgrowth by yeasts able to grow at 37~ but not at 42~
No rationale is provided by Blaser et al. (1979) for their 'Campy-BAP' medium, which has a similar formulation to Skirrow (1977) agar, but with added cephalothin, as used by Lauwers et al. (1978).
In 1983, Butzler and co-workers produced a second selective plating medium called Butzler medium Virion (Goossens et al., 1983). This was said to be superior to a slightly modified version of their 1978 medium (Lauwers et al., 1978), 'Butzler medium Oxoid', which had a different basal medium, less blood and cephazolin instead of cephalothin. Butzler medium Virion substituted cephoperazone for cefazolin, rifampicin for vancomycin, colistin for polymyxin and trimethroprim and amphotericin B for cycloheximide. The Virion medium was said to suppress competing flora more effectively, especially pseudomonads and Enterobacteri- aceae. Campylobacter colonies were easier to recognise. Incubation temperature was reduced from 42~ to 37~ (Butzler et al., 1983). Goossens et al. (1986) modified the Virion medium, omitting the colistin and doubling the concentration of cefoperazone in order to detect colistin-sensitive campylobacters, including some strains of C. coli, reported by Ng et al. (1985) to be sensitive to colistin and polymyxin. Goossens and co-workers found no significant improvement over their Butzler Virion agar. However, they recommended its use in preference to the earlier medium, presumably because they felt it would be able to isolate colistin-

144
and polymyxin-sensitive campylobacters. Burnens and Nicolet (1992)were able to isolate C. upsaliensis after changing to the medium of Goossens et al. (1986).
The medium of Karmali et al. (1986) is a variation of modified CCD medium (Hutchinson and Bolton, 1984) using haematin rather than ferrous sulphate, vancomycin instead of deoxycholate and cycloheximide instead of amphotericin B. Vancomycin at 20 mg per litre rather than the more common 10 mg per litre was chosen for better suppression of Gram-positive competitors (Bacillus spp. and enterococci). Cycloheximide was chosen because of superior yeast suppression. In agreement with Hutchinson and Bolton (1984) cefoperazone was found superior to cephalothin. The most numerous contaminants were found to be Enterobacteri- aceae which are resistant to cefoperazone when present in high numbers, espe- cially Klebsiella oxytoca. The rationale for the modification to the oxygen quench- ing system was not stated.
The semi-solid medium of Goossens et al. (1989) relies upon the ability of campylobacters to swarm, analogous to the system used in semi-solid Rappaport Vassiliadis media for salmonellas (Busse, 1993). The use of a semi-solid medium, where growth and swarming is mostly below the surface apparently dispenses with the need for blood, charcoal or other anti-oxygen system other than a microaerobic atmosphere. The medium has to be stored in the dark at 4~ and prepared twice weekly. Cefoperazone (30 mg/1) and a high level of trimethoprim (50 mg/1) are used as selective agents. Three strains of C. jejuni and 7 strains of C. coli with MIC's to cefoperazone < 100 p,g/ml (screened in a previous study of 200 in- hibitors) were tested for their ability to swarm in the new medium; all gave satisfactory results. Eleven different campylobacter strains were tested for their ability to initiate swarming in the medium. A range of 1-15 cells was necessary, with a mean of 6.7. The medium functioned very well by comparison with other plating media. Neat faecal samples were inoculated using a loop at the edge of 50 mm diameter Petri dishes of the medium. Incubation was at 42~ for 42 h in a microaerobic atmosphere or a candle jar. Positive campylobacter cultures were recognised by the characteristic swarming growth. Presumably isolation required subculture to selective or non-selective plates to obtain a pure growth. Polymyxin or other Gram-positive inhibitors, as well as anti-fungal antibiotics were not necessary because competitor bacteria did not swarm. The medium had a number of advantages: cheapness because of the small volumes needed, no necessity for blood, only two antibiotics in the formula, easy interpretation of swarming, low incidence of contaminants, and the ability to fit large numbers of 50 mm plates into gas jars. A disadvantage would be the unsuitability of semi-solid agar for quantitative estimates of campylobacters, unless MPN techniques were used.
Recently Aspinall et al. (1993) have developed a modification of mCCD agar designed to isolate C. upsaliensis as well as the other thermophilic campylobacters at 37~ Of 51 strains of C. upsaliensis tested, 47 were resistant to 8 mg per litre of cefoperazone in CCD agar base and all were resistant to 64 mg per litre of teicoplanin. The medium contains 8 mg/1 cefoperazone and 4 mg/1 teicoplanin, replacing 32 mg/1 cefoperazone in mCCD agar (Table 3). Teicoplanin has an antimicrobial spectrum similar to that of vancomycin, active mainly against Gram-

145
positive bacteria. By comparison with mCDD agar the final formulation isolated equivalent numbers of Campylobacter spp. other than C. upsaliensis from faeces and was superior to mCCD agar for C. upsaliensis, with slightly more growth of competitors. These results were confirmed using faeces artificially inoculated with C. upsaliensis.
9. Membrane filter method of Steele and McDermott (1984)
This method is a simplification of the filtration method of Dekeyser et al. (1972). It employs 47 mm 0.45 /~m cellulose triacetate membrane filters, placed centrally on a 90 mm 6% sheep blood agar plate. Ten to twelve drops of a 1 in 10 suspension of faeces in physiological saline solution are placed onto the membrane filter using a Pasteur pipette and taking care not to let the drops spill over the edge of the membrane. The membrane is removed after 30 min, by which time the fluid (and presumably sufficient campylobacters)will have passed through. The plates are then incubated micro-aerobically at 41~ +_ 1~ for 3-5 days examining daily. Cellulose nitrate filters were unsatisfactory and 0.8 /~m pore size gave unacceptable levels of contamination; 0.65/~m filters were not tested.
Comparison of the filter method with a selective medium containing sheep blood, FBP supplement, trimethoprim, vancomycin and colistin gave 45 positive with the selective medium and 50 with the filter method out of a total of 56 positive samples from the 1000 tested. Six strains of C. jejuni were not isolated by the filter method and 11 Campylobacter spp. were not isolated with the selective medium. Six of these were C. jejuni. None of the non-C, jejuni strains grew in the selective agar because of the colistin present (they were also sensitive to polymyxin). These authors found that the filter method usually gave either campylobacter colonies or no colonies at all. However, only about 10% of campylobacters in the faecal suspension actually passed through the filter. This method or a similar one (e.g. using 0.65 ~m filters) has been used by various workers, often in parallel with selective agar media or after selective enrichment (Megraud and Gavinet, 1987; Goossens et al., 1990b; De Boer and Reitsma, 1991; Moreno et al., 1993; Scotter et al., 1993). Shanker et al. (1991), working with 0.45 ~m cellulose acetate or mixed cellulose ester filters and artificially contaminated faeces, found that 104/g of most strains of campylobacter could be detected. In their procedure the filter was removed 10 minutes after applying the faecal suspension.
10. Enrichment methods for isolation from food
The Preston and mCCD media of Bolton and co-workers have been utilised as enrichment media using the same inhibitors, and incubation at 37 or 42~ Liquid Preston medium contains 0.25 g/1 FBP in addition to 5% horse blood (Bolton et al., 1982; this Volume ). Both media have been found useful for isolating cam- pylobacters from samples in which campylobacter numbers are comparatively

146
low (e.g. faeces of normal animals, water and drain swabs - Bolton et al., 1982, 1983).
Park et al. (1981) developed an enrichment broth based on Brucella broth, containing vancomycin, trimethroprim and polymyxin B (VTP). It was later modi- fied (Park et al., 1983) by adding FBP supplement and lysed horse blood, reducing the amount of the polymyxin B and increasing that of vancomycin and trimetho- prim (VTP-FBP) - see Table 3. The media were used to examine raw chickens. Carcasses were rinsed with nutrient broth, which was then filtered through cheesecloth and centrifuged. The sediment was resuspended in Brucella broth and then either plated directly onto selective agar or added to the enrichment broth. Incubation was carried out at 37~ under microaerobic conditions. VPT broth was found to detect 0.2 campylobacters per g of sediment in the presence of 104-106 competitors per g (Park et al., 1981). Enrichment showed 62% of chickens to be positive for campylobacters, compared with 32% by direct plating. VPT-FBP broth functioned better at 37~ than 42~ and 48 h incubation was better than 24 h or 72 h. pH 7.0 was optimal.
Christopher et al. (1982) used a liquid version of Campy-BAP agar (see Table 3) with added pyruvate. This medium contained cephalothin and amphotericin B in addition to vancomycin, polymyxin and trimethroprim. Ehlers et al. (1982) modi- fied this medium by adding FBP. The broth was incubated at 42~ in microaerobic atmosphere. The limit of detection in cheese artificially contaminated with C. jejuni and using an MPN method was about 0.3 campylobacters per g. Acuff et al. (1982) published a similar modification containing FBP, but without blood. Doyle and Roman (1982) developed a broth similar to that of Park and co-workers, but with a very high level of polymyxin (20,000 iu per litre), cycloheximide and an oxygen quenching system of 7% lysed horse blood, as well as added succinate and cysteine HC1. Incubation was microaerobic at 42~ for 16-18 h. The medium was evaluated using a variety of foods artificially inoculated with 46 strains of C. jejuni 2 of C. coli and 2 of C. lari. 10 or 25 g of food were homogenised directly in 90 or 100 ml of enrichment broth. Campy-BAP agar was used as the plating medium. All strains in raw hamburger and raw milk were detected at 1-4 cells per g and most at 0.1 to 0.4 cells per g. The broth was less effective for chicken, probably due to the numbers and types of competitors.
Wesley et al. (1983) used a medium (alkaline tryptone broth) containing rifampicin, polymyxin at 20,000 iu per litre and 6.25 mg/1 of cefsulodin. The levels of polymyxin and cefsulodin chosen were necessary to inhibit Pseudomonas aerugi- nosa contaminants. FBP and haematin were included as oxygen quenchers and the optimum pH level was 8.0. 100 ml of broth was inoculated with 10 ml of carcass rinse fluid. Incubation was microaerobic at 42~ for 48 h. Comparison of this medium with the medium and sampling method of Park et al. (1981) for isolation of campylobacters from naturally contaminated chicken gave a much higher isolation rate (25 versus 6 out of 50 chickens using the Wesley or Park medium respectively).
Lovett et al. (1983) modified the medium of Park et al. (1981). They added FBP, and as a result of testing growth of C. jejuni strains and competitors, reduced the

147
level of polymyxin and increased the levels of the other antibiotics (Table 3). The 8,000 iu per litre of polymyxin used in the Park formulation was found to be inhibitory to many strains of C. jejuni.
11. Effect of damage
Ray and Johnson (1984a,b) observed that viability of freeze-injured C. jejuni was reduced when cells were incubated in selective broth or (to a lesser extent) on selective agar at 42~ They used the liquid medium of Acuff et al. (1982) and Campy-BAP agar. The basal media without antibiotics were also inhibitory, an effect attributed partly to the high incubation temperature. Blood reduced the toxicity, as also did succinate and cysteine. Polymyxin was toxic to the injured cells but the other antibiotics were not. As a result of these studies a medium was proposed using a modification of the broth of Ehlers et al. (1982), which contained succinate and cysteine but no FBP supplement. Incubation was at 37~ for the first 6 h to resuscitate injured cells before adding the polymyxin and raising the incubation temperature to 42~
Humphrey and Cruickshank (1985) investigated the effect of a variety of inhibitors used in selective media on 6 serotypes of C. jejuni uninjured or injured by freezing, heating or EDTA. Bacitracin, trimethoprim, cefoperazone, colistin, novobiocin, vancomycin and deoxycholate were tested in blood agar, all at levels used in selective media. The growth rate and colony size of undamaged strains were reduced by rifampicin. Injured cells were more affected. Deoxycholate was toxic for some but not all injured cells. The toxic effect of rifampicin and deoxycholate in complete medium was confirmed for one strain when freeze- damaged. Counts on Preston medium were reduced compared to Skirrow, Campy- BAP or blood agar for one of the 6 strains. Further work (Humphrey 1986a,b) confirmed the observations of Ray and Johnson (1984) that damaged C. jejuni recovered better in media at 37~ than 42 or 43~ even in the absence of inhibitors. An enrichment medium and method was subsequently proposed which had nutrient broth as the base. To this was added 5% lysed horse blood and FBP. Antibiotics were omitted for the initial period of incubation. Food samples were incubated in the basal broth for 2 h at 37~ before addition of the antibiotics and incubation was continued at 43~ for up to 48 h. The antibiotics were trimetho- prim, cefoperazone, colistin, amphotericin B and either vancomycin or rifampicin (see Table 3). Some inhibition of C. jejuni was observed even after the 2 h 37~ resuscitation period. Vancomycin was less inhibitory than rifampicin. No informa- tion was supplied about the incubation atmosphere. The plating medium used following enrichment had a similar formulation, but substituted rifampicin for vancomycin in order to suppress competing flora. In a later paper Humphrey (1989) suggested that pre-enrichment at 37~ should be continued for 4 h and that addition of all antibiotics should be delayed until the 4 h pre-enrichment had been completed. The rifampicin-containing enrichment medium was recommended in parallel with a similar plating medium (Humphrey 1986b; Table 3). Pre-enrichment

148
at 37~ for 4 h was recommended for all types of sample, even those heavily contaminated, such as sewage and chicken skin, although for these, selective medium rather than the basal enrichment broth, was recommended. Later the broth was modified to contain polymyxin instead of colistin, because of difficulty in obtaining colistin (De Boer and Humphrey, 1991; Humphrey, personal communi- cation).
A similar enrichment medium for use with food was suggested by Bolton (Table 3; personal communication; Anon., undated). It has a rich basal medium to aid resuscitation of sublethally damaged campylobacters. Similarly to Humphrey's Exeter enrichment medium, preliminary incubation of the medium complete with antibiotics, for 4 h at 37~ was recommended to aid resuscitation of injured organisms, followed by 42~ for 14-48 h. The combination of some of the FBP ingredients with lysed horse blood and haemin, together with a small headspace in the culture vessels was designed to avoid the need for a microaerobic atmosphere. Sodium carbonate was added, presumably to prevent acidification and /o r provide a source of carbon dioxide. The broth is used in a 1:4 (w/v) ratio of food to medium, with a 1.5 cm headspace in a screw-capped bottle (presumably with the cap tightened). It can also be used to enrich faeces (1 ml of 10% suspension added to 5 ml broth).
Another enrichment method employing similar principles was published in abstract only by Park and Sanders (1991) (see Table 3 for formulation). No rationale was supplied for the choice of antibiotics. The method involved incuba- tion of the broth containing vancomycin and trimethoprim for 4 h at 31-32~ followed by addition of cefoperazone and cycloheximide. Incubation temperature was raised to 37~ for 2 h and then 42~ for 40-42 h. All incubation was microaerobic; static at 30-32 ~ and 37 ~ and shaking at 42~
Yet another enrichment medium, a variation of Park and Sanders broth, also employing a period of incubation at reduced temperature, and delayed addition of antibiotics was devised by Hunt and Radle (cit. Stern and Line, 1992; Hunt, 1992). This medium contains 0.25 g/1 FBP and 5% lysed horse blood, cefoperazone, trimethoprim, vancomycin, and amphotericin B or cycloheximide (see Table 3).
Pre-enrichment is carried out in the medium containing all the antibiotics (except for 15 mg/1 cefoperazone of the final total of 30 mg/ l ) at 32~ in a microaerobic atmosphere in flasks sealed in plastic bags and shaken in a water bath. After 3 h the remainder of the cefoperazone is added and the temperature of the water bath raised to 37~ After a further 2 h at 37~ the bath temperature is raised to 42~
The FDA Bacteriological Analytical Manual (1992) recommends use of Hunt and Radle broth for most types of food, modified Exeter broth for water and environmental samples and Hunt and Radle broth and modified Preston agar for dairy products. Frozen products or products stored chilled for 10 days or more are pre-enriched according to the Hunt and Radle protocol or the Humphrey (1989) protocol. Foods not chill-stored are examined by the Hunt and Radle protocol, but adding all antibiotics at the beginning of incubation. Shaking water baths are recommended for incubation, with continuous gas flow (except during incubation

149
at 30~ if it is static). Plating is on mCCDA and Campy-Cefex directly and after enrichment. In general 25 g food is added to 100 ml of enrichment broth. Liquid products such as milk and ice cream are centrifuged. Chickens are rinsed, the rinse fluid centrifuged and the pellet added to the enrichment broth.
12. Atmosphere during incubation of enrichment media
No systematic investigation of the effect of atmosphere during incubation of liquid media has been carried out. Some earlier workers used static incubation in normal air in screw-capped bottles, sometimes with a small airspace, sometimes with the airspace undefined (Bolton et al., 1982b; Fricker et al., 1983; Martin et al., 1983; Waterman et al., 1984). Others used static incubation in bottles or tubes in a microaerobic atmosphere (Acuff et al., 1982; Ehlers et al., 1982; Megraud, 1987; De Boer and Reitsma, 1991). Doyle and Roman (1984)used 250 ml conical flasks possessing side-arms and replaced the atmosphere with the microaerobic gas mixture. They were evacuated and filled three times. Park et al. (1981, 1983) and Lovett et al. (1983) bubbled microaerobic gas mixture through their enrichment broths. Park and Sanders (1991) used static incubation of plastic storage bags flushed with microaerobic atmosphere. The report of Ribeiro and Price (1984) who used Preston broth containing FBP supplement (George et al., 1978) to isolate campylobacters from water indicated that a microaerobic atmosphere during incubation gave a higher isolation rate. Wesley et al. (1983) reported that better results were obtained with a high surface area of broth in a flask compared to a lower surface area in tubes, in both cases using a microaerobic atmosphere. Comparison of Doyle and Roman's evacuation-replacement procedure with a continuous gas flow procedure using Doyle and Roman's enrichment broth for examining inoculated hamburgers indicated that the continuous gas flow gave counts 1-2 log cycles higher for 6 out of 8 strains of C. jejuni for both tests (Heisick et al., 1984). There appears to be no published justification for the FDA recommendation of continuous gas flow with agitation (Hunt, 1992).
13. Comparisons of media
The most important comparisons will be discussed in chronological order. Of course the earlier comparisons could not examine media only later developed, so that the most recent comparisons are likely to be of greatest interest. Comparisons published to support the development of a new medium have only been cited where several other media have been compared. There are few comparisons of media for isolation from food, so this review will also include studies for isolation from faeces and water.
Morris et al. (1982) compared plating media for isolating C. jejuni from (1) dog and cat faeces and (2) human diarrhoeal faeces. In the first study BU40 (a modification of Butzler's medium (Butzler and Skirrow, 1979) with 40,000 rather

150
than 10,000 iu per litre colistin (Patton et al., 1981), was compared with Skirrow and Butzler medium. Eighty-seven out of 347 samples were positive for C. jejuni. BU40 gave significantly higher numbers of positive samples than Butzler's or Skirrow's media but Butzler and Skirrow media were not significantly different from one another. No two media detected all the positive samples but Skirrow + BU40 detected 98% of them. Detection of C. jejuni on Skirrow medium was sometimes hampered by swarming Proteus spp.. Coliforms were sometimes a problem on Skirrow and Butzler media, but not on BU40, and Providencia spp. were inhibited only by Skirrow's medium. In the second study, Campy-BAP, Skirrow, Butzler and BU40 agars were compared. No significant differences were found between these four plating media. A greater number of contaminants was found on Skirrow's medium. The difference between the results in the two studies may have been because the human faeces examined in the second study were from subjects with diarrhoea and hence campylobacters were present in high numbers. In both surveys it was necessary to check the plates at 24, 48 and 72 h in order to detect all positive samples.
Bolton et al. (1983) compared Skirrow, Butzler (with cephazolin), Campy-BAP, Blaser agar (a modification of Campy-BAP, with 5% horse blood and blood agar base No. 2) and Preston media. Pure cultures of two strains each of C. jejuni, C. coli and NARTC (C. lari)were tested. All the media reduced (i.e. up to 1 log cycle) the counts of pure cultures of at least some strains compared with a non-selective (5% lysed blood) agar. Butzler medium was judged unsuitable for cultivating C. coli and C. lari (reduction of over 3 log cycles). Examination of campylobacter-positive human faeces and faecal samples from sheep, pigs, cattle and chickens, plus abattoir drain swabs indicated that no medium detected all positive samples. The order of efficiency of isolation was Preston (84%), Blaser (76%), Skirrow (74%), Campy-BAP (69%), Butzler (68%). In terms of rate of contamination with unwanted organisms, Preston agar had fewest followed by Butzler, Skirrow, Blaser and Campy-BAP. The most frequent contaminants were coliforms, Pseudomonas spp. and streptococci. Growth of yeasts occurred on Skirrow, Butzler and Preston media. Plates read after 48 h at 42~ gave consis- tently higher isolation rates than plates read at 24 h. Isolation rates were not significantly higher after 72 h, because of overgrowth by contaminants. The use of Preston enrichment broth improved isolation rates from samples with compara- tively low numbers of campylobacters - e.g. sheep and cattle rectal swabs and abattoir drain swabs. The observation that Blaser agar gave better results than Campy-BAP when the two media were identical except for differing basal media and blood content is of interest. Campy-BAP was also the only medium supplied ready-made and this might have had an effect. Concerning poor growth of C. lari and C. coli Bolton et al. commented that they had found that Butzler's medium based on Columbia agar and 5% horse blood, instead of blood agar base no. 2 and 5% horse blood, recovered campylobacters much more effectively from avian and porcine samples, where C. lari and C. coli respectively predominate. The later Butzler media (Oxoid and Virion) used Columbia agar base.
Fricker et al. (1983) compared Preston, Skirrow, Butzler (with cephazolin) and

151
Blaser (as used by Bolton et al. 1983) agars, for plating seagull faeces directly or after enrichment in Preston or Roman and Doyle's enrichment medium. Enrich- ment greatly increased the isolation rate. The highest rate of isolation occurred using Preston enrichment onto Preston agar. There was little difference in the performance of the four selective agar media, except that C. lari and C. coli strains grew very poorly on Butzler medium, confirming the observations of Bolton et al. (1983).
Merino et al. (1986) tested seven plating media for the examination of 263 samples of diarrhoeal faeces from humans. Butzler (with cephazolin, basal medium not clear), Blaser, Skirrow, Preston, mCCDA, Butzler Virion and Preston with 2 mg/1 amphotericin B. All the media isolated campylobacter at similar frequency, but mCDDA was preferred because it gave the lowest number of plates with contaminating flora. Incubation at 43~ for 24 h was considered optimal. Further incubation until 48 h resulted in overgrowth of contaminants.
Rosef et al. (1987) compared methods for isolating campylobacters from natu- rally contaminated water. In the first part of their study they filtered the water through 0.45 mm and 0.22 mm membranes and compared recovery when the filters were placed in Preston, Rosef or Rosef+ FBP supplement enrichment broth (42~176 incubated in microaerobic atmosphere for 24-48 h), with the recovery obtained when the filters were laid face up on Skirrow or CAK agar. Of forty positive samples, 39 were detected without enrichment (using CAK and Skirrow agar), 27 were detected by use of Skirrow agar alone and 35 using CAK agar alone. All positive samples were detected using 0.45 /xm filters and only one using the 0.22/xm filter. The best enrichment medium was Preston, followed by Rosef + FBP. In the second part of the study the water was passed through various combinations of 0.65, 0.45 and 0.22 ~ m filters which were then placed face up on CAK agar. The 0.45/xm filter used alone gave the highest isolation rate followed by the 0.65/~m filter used alone. In the third part Preston, CAK and Skirrow agars were com- pared, inoculating with 0.45 /~m filters, face up. Preston detected the greatest number of positive samples, followed by CAK, with Skirrow detecting substantially fewer. All three media needed to be incubated for 48 h in order to detect the maximum number of positive samples. No bias was observed in proportions of C. jejuni, C. coli and C. lari isolated on the three plating media. In a similar study, using 0.45 ~m filters, Brennhovd and Kapperud (1991) compared enrichment in Preston broth followed by plating on Preston agar or CAK agar, with direct plating of the filters onto Preston or CAK agar. Preston agar gave higher numbers of positive samples than CAK agar by direct plating. Direct plating on Preston agar, together with enrichment in Preston broth with subculture to Preston agar, detected 93% of positive samples. It was necessary to examine plates at 48 h as well as 24 h incubation.
Gun-Munro et al. (1987) compared counts of pure cultures of 70 strains of C. jejuni on Skirrow, Butzler, Blaser-Wang, Preston, mCCDA and Karmali media. All except Skirrow, mCCDA and Karmali gave slightly but significantly lower mean counts than non-selective blood agar. 'Simulated faeces' samples were prepared by inoculating each of seventy different faeces samples, previously found to be

152
negative for campylobacters, with one of the seventy pure cultures of C. jejuni at a level of 105 per ml faeces. The six media were scored in terms of numbers of simulated faeces samples found positive, as well as their ability to inhibit the inoculated faecal flora. The percent positive samples detected at 48 h for each medium was as follows: Blaser-Wang 41%; Butzler 86%; Preston 91%; Skirrow 96%; Karmali 97%; mCCDA 99%. Karmali and mCCDA suppressed the competi- tive flora more effectively. Karmali was chosen rather than mCCDA to compare with Skirrow medium against almost 2,556 human faecal specimens and 224 animal and bird specimens. This was because the authors found campylobacter colonies easier to recognise on Karmali than on mCCDA. There was little to choose between Skirrow and Karmali medium in terms of productivity, with Karmali isolating a few more strains from human faeces in transport media, and from chicken faeces. Skirrow medium gave better results on faeces from other animals. However, Karmali medium was much more selective and therefore easier to use. For instance at 48 h the faecal flora was suppressed by 75% or more with 57% of the samples on Karmali, compared with only 32% suppression on the Skirrow medium.
The study by Albert et al. (1992) on human diarrhoeal stools compared mCCDA incubated at 42~ with the membrane filter method of Steele and McDermott (1984) but using a 0.65/zm pore membrane filter and incubating at 37~ The filter method isolated many more campylobacters, mostly uncharacterised Campylobac- ter spp., C. upsaliensis and C. jejuni subsp, doylei, but also significantly more C. jejuni. Multiple strains of campylobacter were often isolated from one faecal sample using the filter method. The differing performance of the two methods could have been due to the different incubation temperatures and /o r the antibi- otics in the selective medium. Megraud and Gavinet (1987) compared isolation rates from human faeces after direct plating on Butzler medium Virion at 42~ for 2 days with membrane filtration using a 0.65/zm cellulose acetate filter (incubating at 37~ or with membrane filtration after enrichment in Preston broth (all incubated at 37~ and plates incubated for 5-7 days). The enrichment filtration method was most sensitive and direct plating least sensitive. Better results were obtained with Millipore membranes (cellulose nitrate and acetate) than Sartorius membranes (cellulose nitrate).
Moreno et al. (1993) examined faecal samples from various domestic pets and compared with direct plating on mCCDA two methods of filtration through a 0.65 /~m pore (Millipore) filters onto 5% horse blood agar. Incubation was at 37~ for periods up to 8 days. A filtration procedure was superior to direct plating for isolating campylobacters (particularly C. upsaliensis) from this type of specimen. A method that involved initial low speed centrifugation of the faecal suspension (10%) in order to prevent clogging of the filter, followed by filtration through a 0.65 ~m filter and high speed centrifugation in order to concentrate the organisms, was superior to the Steele and McDermott (1984) method.

153
14. Isolation from foods
Beuchat (1985) compared Campy-BAP, CCDA (unmodified - Table 3 - Bolton and Coates (1983)) and Butzler agar (as in Table 3: with cephalothin), plus five enrichment media (by MPN) for the isolation of campylobacters from chill-stored chicken. The chicken had been artificially inoculated with five strains of C. jejuni. Recovery was poor with all the enrichment/plating combinations, but the enrich- ment media of Christopher et al. (1981) and Park et al. (1983) combined with Butzler's or CCDA media gave best results. The medium of Martin et al. (1983) performed particularly poorly. Preston broth and the medium of Rosef and Kapperud (1983) gave intermediate results. Direct plating of chicken samples diluted in 0.1% peptone water gave better recovery than enrichment, with CCDA and Campy-BAP yielding higher counts than Butzler agar. Preliminary study of Doyle and Roman's enrichment broth indicated that it was superior to other media.
Fricker (1984) undertook a comparison of three enrichment media: Preston, Lander and Doyle and Roman all incubated at 42~ one transport/storage medium stored at 4~ (Campy-thio: Blaser et al. 1979) and two plating media - Preston and Campy-BAP. All media had 0.05% FBP added and were incubated aerobically in 6 ml quantities in quarter oz (ca. 7.5 ml) bottles. Naturally contami- nated giblets from frozen chickens were examined. 177/198 samples were positive for campylobacters. Doyle and Roman and Preston media both detected 176 positive samples while Lander medium detected 159 and the Campy-thio medium 69. Samples were not plated directly, but the result using Campy-thio, in which multiplication of campylobacters would not have occurred, was probably similar to the result that would have been found by direct plating. Preston agar isolated campylobacters more frequently than Campy-BAP (577 v. 562 times). 48 h was the optimal time for incubation at 42~ Enrichment in either Doyle and Roman or Preston broth followed by plating onto Preston agar was recommended.
A comparison of the Doyle and Roman enrichment method with that of Lovett et al. (1983) for isolation from milk was carried out by Hunt et al. (1985). The Lovett method was significantly better. Heisick et al. (1984) and Heisick (1985) also compared the Doyle and Roman broth with the Lovett et al. medium, using artificially inoculated beefburgers or milk-puddings and various strains of C. jejuni. The two methods gave similar results, although higher numbers of C. jejuni were sometimes achieved in the Lovett medium, especially when added in the absence of food.
Peterz (1991)conducted a collaborative trial amongst 6 laboratories in Scandi- navia, testing chicken liver artificially inoculated with two strains of C. jejuni at levels between 0.3 and 59 per g. 2.5 g were enriched in 50 ml of Preston broth, without FBP but incubated microaerobically, at 42~ for 24 h, followed by plating onto mCCDA and Preston agar. A detection limit of about one campylobacter per g of liver was determined. The two plating media gave similar isolation rates, although mCCDA contained fewer contaminants.
De Boer and Humphrey (1991) compared Park and Sanders method to Preston

154
broth and mCCD broth for chilled or frozen chicken and found Park and Sanders superior, but gave no details. Exeter medium (modified from that described by Humphrey (1986b) see Table 3) was reported to give good results when used with a 37~ 18 h pre-enrichment step.
De Boer and Reitsma (1991) compared the effectiveness of Preston broth (without FBP) and mCCD broth in combination with plating onto Skirrow, Preston, mCCDA and filtration through an 0.6 /xm pore membrane used with blood agar (modified Steele and McDermott, 1984). All incubation was micro- aerobic and naturally contaminated chilled and frozen chickens were examined. The two enrichment broths gave similar results, most positives occurring with mCCD broth plated via the membrane filter. Direct plating of frozen chicken gave much lower numbers of positive results than when samples were enriched (e.g. 8% positive versus 27% on mCCDA with or without enrichment in Preston broth respectively). With chilled chicken, direct plating on mCCDA gave 32% positive, compared with 47% positive after enrichment. Direct plating using the membrane filter method was completely ineffective.
The study of Furanetto et al. (1991), also examining naturally contaminated chilled chicken carcasses, compared direct plating on Campy-BAP or VTP-FBP agar with plating on the same media after enrichment in Doyle and Roman or Preston broth. All incubation was static in a micro-aerobic atmosphere. The combination of Doyle and Roman enrichment with Campy-BAP gave the highest number (16/42) of positive samples, although the results were not statistically significantly different from direct plating on either Campy-BAP (13/42) or VTP- FBP agar (11/42). The combination of Doyle and Roman broth with VTP-FBP agar gave particularly poor results (1/42).
Stern and Line (1992) compared the enrichment broths of Doyle and Roman, Park and Sanders and Hunt and Radle (plating onto Campy-BAP, mCCDA and Campy-Cefex) for examining whole chilled, naturally contaminated chickens. Inoc- ula were prepared by rinsing the carcasses with 200 ml buffered peptone, filtering the rinse through cheesecloth, centrifuging, discarding the supernatant, streaking some of the pellet onto the selective agars, resuspending the rest of the pellet in 5 ml buffered peptone water and inoculating 1 ml of the resuspension to 100 ml of enrichment broth. Microaerobic atmosphere was obtained by replacing the air in polythene bags with the appropriate gas mixture (flushing 3 times). All bags were incubated in water baths with shaking. The Doyle and Roman broth was incubated at 42~ The Park and Sanders and Hunt and Radle broths were incubated at 32~ for 3.5 h, 37~ for 2.5 h and then at 42~ Cefoperazone supplement was added to the Hunt and Radle broth and cefoperazone and cycloheximide to the Park and Sanders broth before raising the temperature from 32 to 37~ Re-gassing of the enrichment broths was carried out after adding the supplements. Ten out of 50 chicken carcasses were positive for campylobacter by direct plating. Forty-nine out of 50 were detected by all the enrichment/plating methods combined. 23, 40 and 43 positive chickens were detected using Doyle and Roman, Park and Sanders and Hunt and Radle broths respectively, mCCD agar gave most positive samples

155
overall, and with the Park and Sanders method, although all three selective media detected similar numbers of positive samples from Hunt and Radle broth.
The study of Scotter et al. (1993) used frozen samples of naturally contaminated chicken skins, in which the campylobacters were more likely to be damaged, and therefore require pre-enrichment. Twelve laboratories in the UK participated in the trial. Tests were carried out using two methods defined in a draft International Standards Organisation Method. The three enrichment methods used 10 g quanti- ties of food in 90 ml of enrichment medium, under a small headspace. They were as follows: (1) Preston broth without FBP supplement, incubated in micro-aerobic atmosphere at 42~ for 48 h; (2) Park and Sanders broth incubated aerobically, with antibiotic supplement and at times and temperatures prescribed by Park and Sanders (1989); (3) in Preston broth containing FBP supplement, aerobically, but according to the temperatures and times prescribed by Humphrey (1989) - 37~ for 4 h and then 42~ for 44 h. All broths were plated directly onto Skirrow agar. Method 1 used any selective medium chosen by the participant, method 2 used a 0.65 /tm membrane on non-selective blood agar and method 3 used Preston agar in addition. All plates were incubated at 42~ At 10 cells per 10 g chicken skin all three methods were equally effective. At about 2 cells per 10 g, method 2 gave significantly higher isolation rates. Skirrow agar was frequently overgrown with pseudomonads and proteus. The membrane plus non-selective agar was as effective as selective agars such as MCCDA. No advantage was gained by pre- enriching at 37~ for 4 h (method 3). However, pre-enrichment at 37~ for 4 h was intended by Humphrey (1989) for use with his (Exeter) medium (Humphrey, 1986b), not Preston broth.
15. Conclusions
Media for isolating campylobacters and related bacteria from faeces and other environments such as food and water are not yet optimal, and there is so far no consensus concerning the best media and methods. The key to elucidating efficient and economic media and methods might lie in more investigation into the phenomenon of 'viable but not culturable' campylobacters and the ability of at least some strains of C. jejuni to adapt to grow in air (Jones et al., 1993). Future developments could include the use of aerobic incubation and/or different media for some groups, as well as 37~ instead of 42 ~ or 43~ Some plating media are already incubated at 37~
Of the plating media listed in Table 2, probably the most widely used today are Skirrow, Campy-BAP, Preston, mCCD, Butzler (Virion) and Karmali. mCCD and Karmali agars have performed best in comparative studies using faecal samples. Other media - e.g. the semi-solid medium of Goossens et al. (1989) and the medium of Aspinall et al. (1993), have yet to be tested in comparative studies. If cefoperazone- and polymyxin- or colistin-sensitive campylobacters are sought and/or arcobacters and C. jejuni subsp, doylei, which do not grow at 42~ or 43~ media such as mCCD, those of Goossens et al. (1986) and Aspinall et al. (1993)

156
and the semi-solid medium of Goossens et al. (1989) incubated at 37~ might be useful. Use of modifications of the membrane filter method of Steele and McDer- mott (1984) in combination with non-selective blood agar are useful for this group, but only if relatively high numbers of campylobacters are present in the faecal suspension or enrichment culture.
Development of liquid enr ichment media has so far been confined to the isolation of thermophil ic campylobacters, predominant ly C. jejuni subsp, jejuni. Future work may be directed towards modifying these media and current methods in order to detect the mesophilic and antibiotic-sensitive types ment ioned above. This may, in turn, depend on whether those types are perceived to be an important cause of diarrhoea, and whether contaminated food is shown to be a vector.
More investigation is needed concerning the optimal atmospheric conditions necessary for thermophilic campylobacters in liquid medium, including whether an aerobic a tmosphere might be advantageous for some applications, possibly in parallel with micro-aerobic conditions.
Acknowledgements
We would like to thank Margare t Smart for typing the manuscript and David Corry for translating the paper by Brennhovd and Kapperud (1991) from Norwe- gian.
References
Acuff, G.R., Vanderzant, C., Gardner, F.A. and Golan, F.A. (1982) Evaluation of an enrichment plating procedure for the recovery of Campylobacter jejuni from turkey eggs and meat. J. Food Prot. 45, 1276-1278.
Ahonkhai, V.I., Cherubin, C.E., Sierra, M.F., Bokkenheuser, V.D., Shulman, M.A. and Mosenthal, A.C. (1981) In vitro susceptibility of Campylobacter fetus subsp, jejuni to N-formimidoyl thien- amycin, rosaramicin, cefoperazone and other antimicrobial agents. Antimicrob. Agents Chemother. 20, 850-851.
Albert, M.J., Tee, W., Leach, A.S., Asche, V. and Penner, J.L. (1992) Comparison of a blood-free medium and a filtration technique for the isolation of Campylobacter spp. from diarrhoeal stools of hospitalised patients in central Australia. J. Med. Microbiol. 37, 176-179.
Anon. (undated) Lab-M Culture Media. Lab-M, Bury BL9 6AU, UK, p. 36. Aspinall, S.T., Wareing, D.R.A., Hayward, P.G. and Hutchinson, D.N. (1993) Selective medium for
thermophilic campylobacters including Campylobacter upsaliensis. J. Clin. Pathol. 46, 829-831. Barot, M.S. and Bokkenheuser, V.D. (1984) Systematic investigation of enrichment media for wild-type
Campylobacter jejuni strains. J. Clin. Microbiol. 20, 77-80. Baird, R.M., Corry, J.E.L. and Curtis, G.D., editors (1987) Pharmacopoeia of culture media for food
microbiology. Int. J. Food Microbiol. 5, 212-213 (mCCD agar), 250-253 (Preston media), 268-269 (Skirrow agar).
Beuchat, L.R. (1985) Efficacy of media and methods for detecting and enumerating Campylobacter jejuni in refrigerated chicken meat. Appl. Environ. Microbiol. 50, 934-939.
Blaser, M.J., Cravens, J., Powers, R. and Wang, W.L. (1978) Campylobacter enteritis associated with canine infection. Lancet 2, 979-981.

157
Blaser, M.J., Berkowicz, I.D., Laforce, F.M., Cravens, J., Relier, L.B. and Wang, W.L. (1979) Campylobacter enteritis: clinical and epidemiological features. Ann. Intern. Med. 91, 179-185.
Blaser, M.J., Hardesty, H.L., Powers, B. and Wang, W.L.L. (1980) Survival of Campylobacter fetus subsp, jejuni in biological milieus. J. Clin. Microbiol. 11, 309-313.
Bolton, F.J., Hinchcliffe, P.M., Coates, D. and Robertson, L. (1982) A most probable number method for estimating small numbers of campylobacters in water. J. Hyg. Camb. 89, 185-190.
Bolton, F.J. and Robertson, L. (1982) A selective medium for isolating Campylobacter jejuni/coli. J. Clin. Pathol. 35, 462-467.
Bolton, F.J. and Coates, D. (1983) Development of a blood-free campylobacter medium: screening tests on basal media and supplements, and the ability of selected supplements to facilitate aerotolerance. J. Appl. Bacteriol. 54, 115-125.
Bolton, F.J., Coates, D., Hinchcliffe, P.M. and Robertson, L. (1983) Comparison of selective media for isolation of Campylobacter jejuni/coli. J. Clin. Pathol. 36, 78-83.
Bolton, F.J., Coates, D. and Hutchinson, D.N. (1984a) The ability of Campylobacter media supplements to neutralise photochemically induced toxicity and hydrogen peroxide. J. Appl. Bacteriol. 56, 151-157.
Bolton, F.J., Hutchinson, D.N. and Coates, D. (1984b) A blood-free selective medium for the isolation of Campylobacter jejuni from faeces. J. Clin. Microbiol. 19, 169-171.
Bolton, F.J., Coates, D. Hutchinson, D.N. and Godfree, A.F. (1987) A study of thermophilic campy- lobacters in a river system. J. Appl. Bacteriol. 62, 167-176.
Bolton, F.J., Wareing, D.R.A., Skirrow, M.B. and Hutchinson, D.N. (1992) Identification and biotyping of campylobacters. In: Board, R.D., Jones, D. and Skinner, F.A. (Editors) Identification Methods in Applied and Environmental Microbiology. Society for Applied Bacteriology Technical Series, 29. Blackwell, Oxford, pp. 151-161.
Boudreau, M., Higgins, R. and Mittal, K.R. (1991) Biochemical and serological characterisation of Campylobacter cryoaerophila. J. Clin. Microbiol. 29, 54-58.
Brennhovd, O. and Kapperud, G. (1991) Comparison of three methods of isolation of thermotolerant Campylobacter spp. from water In: Brennhovd, O. (Editor) Termotolerante Campylobacter spp. og Yersinia spp. i noem norske vannforekomster. Ph.D. thesis, Norwegian College of Veterinary Medicine, Oslo (in Norwegian).
Brennhovd, O., Kapperud, G. and Langeland, G. (1992) Survey of thermotolerant Campylobacter spp. and Yersinia spp. in three surface water sources in Norway. Int. J. Food Microbiol. 15, 327-338.
Brooks, B.W., Garcia, M.M., Frazer, A.D.E., Lior, H., Stewart, R.B. and Lammerding, A.M. (1986) Isolation and characterisation of cephalothin-susceptible Campylobacter coli from slaughtered cattle. J. Clin. Microbiol. 24, 591-595.
Buck, G.E. and Kelly, M.T. (1981) Effect of moisture content of the medium on colony morphology of Campylobacter fetus subsp, jejuni. J. Clin. Microbiol. 14, 585-586.
Burnens, A.P. and Nicolet, J. (1992) Detection of Campylobacter upsaliensis in diarrheic dogs and cats, using a selective medium with cephoperazone. Am. J. Vet. Res. 53, 48-51.
Busse, M. (1994) Media for salmonellae. Int. J. Food Microbiol. in press. Butzler, J.P., Dekeyser, P. and Detrain, M. (1973) Related Vibrio in stools. J. Pediatr. 82, 493-495. Butzler, J.P., Dekeyser, P. and Lafontaine, T. (1974) Susceptibility of related vibrios and Vibrio fetus to
twelve antibiotics. Antimicrob. Agents Chemother. 5, 86-89. Butzler, J.P. and Skirrow, M.B. (1979) Campylobacter enteritis. Clin. Gastroenterol. 8, 737-765. Butzler, J.P., De Boeck, M. and Goossens, H. (1983) New selective medium for isolation of Campy-
lobacter jejuni from faecal specimens. Lancet i, 818. Christopher, F.M., Smith, G.C. and Vanderzant, C. (1982) Examination of poultry giblets, raw milk and
meat for Campylobacter fetus subsp, jejuni. J. Food Prot. 45, 260-262. De Boer, E. and Humphrey, T.J. (1991) Comparison of methods for the isolation of thermophilic
campylobacters from chicken products. Microb. Ecol. Health Dis. 4 (Special Issue) $43. De Boer, E. and Reitsma, W. (1991) Isolation of Campylobacter from poultry products using membrane
filtration. In: Ruiz-Palacios G.M., Calva, F. and Ruiz-Palacios, B.R. (Editors) Campylobacter V, Proc. 5th International Workshop on Campylobacter Infection, Instituto Nacional de la Nutricion, Vasco de Quiroga 15, Mexico 1400, D.F., pp. 113-115.

158
Dekeyser, P., Gossuin-Detrain, M., Butzler, J.P. and Sternon, J. (1972) Acute enteritis due to related vibrio: first positive stool cultures. J. Infect. Dis. 125, 390-392.
Doyle, M.P. and Roman, D.J. (1982) Recovery of Campylobacter jejuni and Campylobacter coli from inoculated foods by selective enrichment. Appl. Environ. Microbiol. 43, 1343-1353.
Ehlers, J.G., Chapparo-Serrano, M., Richter, R.L. and Vanderzant, C. (1982) Survival of Campylobac- ter fetus subsp, jejuni in cheddar and cottage cheese. J. Food Prot. 45, 1018-1021.
FDA (Food and Drug Administration) Bacteriological Analytical Manual (1992) 7th edition. AOAC, Arlington, VA.
Fraser, A.D.E., Chandan, V., Yamazaki, H., Brooks, B.W. and Garcia, M.M. (1992) Simple and economical culture of Campylobacter jejuni and Campylobacter coli in CO 2 in moist air. Int. J. Food Microbiol. 15, 377-382.
Fricker, C.R. (1984) Procedures for the isolation of Campylobacter jejuni and Campylobacter coli from poultry. Int. J. Food Microbiol. 1, 149-154.
Fricker, C.R. (1985) A note on the effect of different storage procedures on the ability of Preston medium to recover campylobacters. J. Appl. Bacteriol. 58, 57-62.
Fricker, C.R., Girdwood, R.W.A. and Munro, D. (1983) A comparison of procedures for the isolation of campylobacters from seagull faeces. J. Hyg. Camb. 91, 445-450.
Furanetto, S.M.P., Nascimento, D.D., Cerqueira-Campos, M.L. and Iaria, S.T. (1991) Efficacy of direct plating and selective enrichment media for detecting Campylobacter jejuni in fresh eviscerated whole market chickens - S~o Paulo, Brazil. Rev. Microbiol. Silo Paulo 22, 303-307.
Gebhart, C.J., Ward, G.E. and Kurtz, H.J. (1985) In vitro activities of 47 antimicrobial agents against three Campylobacter spp. from pigs. Antimicrob. Agents Chemother. 27, 55-59.
George, H.A., Hoffman, P.S., Smibert, R.M. and Krieg, N.R. (1978) Improved media for growth and aerotolerance of Campylobacter fetus. J. Clin. Microbiol. 8, 36-41.
Gilchrist, M.J.R., Grewell, C.M. and Washington, J.A. (1981) Evaluation of media for isolation of Campylobacter fetus subsp, jejuni from faecal specimens. J. Clin. Microbiol. 14, 393-395.
Goossens, H., De Boeck, M. and Butzler, J.P. (1983) A new selective medium for the isolation of Campylobacter jejuni from human faeces. Eur. J. Clin. Microbiol. 2, 389-394.
Goossens, H. De Boeck, M., Coignau, H., Vlaes, L., Van Den Borre, C. and Butzler, J.P. (1986) Modified selective medium for isolation of Campylobacter spp. from faeces: comparison with Preston medium and blood-free medium and a filtration system. J. Clin. Microbiol. 24, 840-845.
Goossens, H., Vlaes, L., Galand, I., Van Den Borre, C. and Butzler, J.P. (1989) Semisolid blood-free selective motility medium for the isolation of campylobacters from stool specimens. J. Clin. Microbiol. 27, 1077-1080.
Goossens, H., Pot, B., Vlaes, L. et al. (1990a) Characterisation and description of 'Campylobacter upsaliensis' isolated from human faeces. J. Clin. Microbiol. 28, 1039-1046.
Goossens, H., Vlaes, L., DeBoeck, M. et al. (1990b) Is 'Campylobacter upsaliensis' an unrecognised cause of human diarrhoea? Lancet 335, 584-586.
Gun-Munro, J., Rennie, R.P., Thornley, J.H., Richardson, H.L., Hodge, D. and Lynch, J. (1987) Laboratory and clinical evaluation of isolation media for Campylobacter jejuni. J. Clin. Microbiol. 25, 2274-2277.
Heisick, J. (1985) Comparison of enrichment broths for isolation of Campylobacter jejuni. Appl. Environ. Microbiol. 50, 1313-1314.
Heisick, J., Lanier, J. and Peeler, J. (1984) Comparison of enrichment methods and atmosphere modification procedures for isolating Campylobacter jejuni from foods. Appl. Environ. Microbiol. 48, 1254-1255.
Hoffman, P., Krieg, N.R. and Smibert, R.M. (1979a) Studies of the microaerophilic nature of Campylobacter fetus subsp, jejuni. I. Physiological aspects of enhanced aerotolerance. Can. J. Microbiol. 25, 1-7.
Hoffman, P.S., George, H.A., Krieg, N.R. and Smibert, R.M. (1979b) Studies of the microaerophilic nature of Campylobacter fetus subsp, jejuni. II. Role of exogenous superoxide anions and hydrogen peroxide. Can. J. Microbiol. 25, 8-16.
H611er, C. (1991) A note on the comparative efficacy of three selective media for isolation of Campylobacter species from environmental samples. Zbl. Hyg. Unweltmed. 192, 116-123.

159
Humphrey, T.J. (1986a) Injury and recoyery in freeze-or-heat damaged Campylobacter jejuni. Lett. Appl. Microbiol. 3, 81-84.
Humphrey, T.J. (1986b) Techniques for optimum recovery of cold-injured Campylobacter jejuni from milk and water. J. Appl. Bacteriol. 61, 125-132.
Humphrey, T.J. (1989) An appraisal of the efficacy of pre-enrichment for the isolation of Campylobac- ter jejuni from water and food. J. Appl. Bacteriol. 66, 119-126.
Humphrey, T.J. and Cruikshank, J.G. (1985) Antibiotic and deoxycholate resistance in Campylobacter jejuni following freezing or heating. J. Appl. Bacteriol. 59, 65-71.
Hunt, J.M. (1992) Campylobacter. In: Food and Drug Administration Bacteriological Analytical Manual, 7th edition. AOAC, Arlington, VA, pp. 77-94
Hunt, J.M., Francis, D.W., Peeler, J.T. and Lovett, J. (1985) Comparison of methods for isolating Campylobacter jejuni from raw milk. Appl. Environ. Microbiol. 50, 535-536.
Hutchinson, D.N. and Bolton, F.J. (1984) Improved blood-free selective medium for the isolation of Campylobacter jejuni from faecal specimens. J. Clin. Pathol. 37, 956-957.
Jacob, J. and Stelzer, W. (1992) Comparison of two media for the isolation of thermophilic campylobac- ters from waste waters of different quality. Zbl. Mikrobiol. 147, 42-44.
Jones, D.M., Sutcliffe, E.M. and Curry, A. (1991) Recovery of viable but non-culturable Campylobacter jejuni. J. Gen. Microbiol. 137, 2477-2482.
Jones, D.M., Sutcliffe, E.M., Rios, R., Fox, A.J. and Curry, A. (1993) Campylobacter jejuni adapts to aerobic metabolism in the environment. J. Med. Microbiol. 38, 145-150.
Juven, B.J. and Rosenthal, I. (1985) Effect of free-radical and oxygen scavengers on photochemically generated oxygen toxicity and on the aerotolerance of Campylobacter jejuni. J. Appl. Bacteriol. 59, 413-419.
Karmali, M.A., De Grandis, S. and Fleming, P.C. (1980) Antimicrobial susceptibility of Campylobacter jejuni and Campylobacter fetus subsp, fetus to eight cephalosporins with special reference to species differentiation. Antimicrob. Agents Chemother. 18, 948-951.
Karmali, M.A., De Grandis, S. and Fleming, P.C. (1981) Antimicrobial susceptibility of Campylobacter jejuni with special reference to resistance patterns of Canadian isolates. Antimicrob. Agents Chemother. 19, 593-597.
Karmali, M.A., Simor, A.E., Roscoe, M., Fleming, P.C., Smith, S.S. and Lane, J. (1986) Evaluation of a blood-free, charcoal-based, selective medium for the isolation of Campylobacter organisms from faeces. J. Clin. Microbiol. 23, 456-459.
Kiehlbauch, J.A. Brenner, D.J., Nicholson, M.A., Baker, C.N., Patton, C.M., Steigerwalt, A.B. and Wachsmuth, I.K. (1991) Campylobacter butzleri sp. nov. isolated from humans and animals with diarrhoeal illness. J. Clin. Microbiol. 29, 376-385.
Kiehlbauch, J.A., Baker, C.N. and Wachsmuth, I.K. (1992) In vitro susceptibilities of aerotolerant Campylobacter isolates to 22 antimicrobial agents. Antimicrob. Agents Chemother. 36, 717-722.
Lander, K.P. (1982) A selective, enrichment and transport medium for campylobacters. In: D.G. Newell (Editor) Campylobacter: Epidemiology, Pathogenesis and Biochemistry. MTP Press, Lancaster, pp. 77-78.
Lastovica, A.J., Le Roux, E. and Penner, J.L. (1989) 'Campylobacter upsaliensis' isolated from blood cultures of paediatric patients. J. Clin. Microbiol. 276, 657-659.
Lauwers, S. De Boeck, M. and Butzler, J.P. (1978) Campylobacter enteritis in Brussels. Lancet i, 604-605.
Lovett, J., Francis, D.W. and Hunt, J.M. (1983) Isolation of Campylobacter jejuni from raw milk. Appl. Environ. Microbiol. 46, 459-462.
Martin, W.H., Patton, C.M., Morris, G.K., Potter, M.E. and Puhr, N.D. (1983) Selective enrichment broth medium for isolation of Campylobacter jejuni. J. Clin. Microbiol. 17, 853-855.
Megraud, F. (1987) Isolation of Campylobacter spp. from pigeon faeces by a combined enrichment- filtration technique. Appl. Environ. Microbiol. 53, 1394-95.
Megraud, F. and Gavinet, A.M. (1987) Comparison of three protocols to isolate Campylobacter species from human faeces. In: Kaijser, B. and Falsen, E. (Editors) Campylobacter IV, Proc. 4th Interna- tional Workshop on Campylobacter Infections, Univ. G6teborg, Sweden, pp. 90-91.

160
Merino, F.J., Agulla, A., Villasante, P.A., Diaz, A., Saz, J.V. and Velasco, A.C. (1986) Comparative efficacy of seven selective media for isolating Campylobacter jejuni. J. Clin. Microbiol. 24, 451-452.
Michel, J., Rogol, M. and Dickman, D. (1983) Susceptibility of clinical isolates of Campylobacter jejuni to sixteen antimicrobial agents. Antimicrob. Agents Chemother. 23, 796-797.
Moreno, G.S., Griffiths, P.L., Connerton, I.F. and Park, R.W.A. (1993) Occurrence of campylobacters in small domestic and laboratory animals. J. Appl. Bacteriol. 75, 49-54.
Morris, G.K., Bopp, C.A., Patton, C.M. and Wells, J.G. (1982) Media for isolating campylobacter. Arch. Lebensmittelhyg. 33, 151-153.
Ng, L.K., Stiles, M.E. and Taylor, D.E. (1985) Inhibition of Campylobacter coli and Campylobacter jejuni by antibiotics used in selective growth media. J. Clin. Microbiol. 22, 510-514.
Ng, L.K., Taylor, D.E. and Stiles, M.E. (1988) Characterisation of freshly isolated Campylobacter coli strains and suitability of selective media for their growth. J. Clin Microbiol. 26, 518-523.
Park, C.E., Stankiewicz, Z.K., Lovett, J. and Hunt, J. (1981) Incidence of Campylobacter jejuni in fresh eviscerated whole market chickens. Can. J. Microbiol. 27, 841-842.
Park, C.E. and Stankiewicz, Z.K. (1982) A selective enrichment procedure for the isolation of Campylobacter jejuni from foods. In: Newell, D.G. (Editor) Campylobacter: Epidemiology, Patho- genesis and Biochemistry. MTP, Lancaster, UK, p. 79.
Park, C.E., Stankiewicz, Z.K., Lovett, J., Hunt, J. and Francis, D.W. (1983) Effect of temperature, duration of incubation and pH of enrichment culture on the recovery of Campylobacter jejuni from eviscerated market chickens. Can. J. Microbiol. 29, 803-806.
Park, C.E. and Sanders, G.W. (1991) A sensitive enrichment procedure for the isolation of Campy- lobacter jejuni from frozen foods. In: Riuz-Palacios, G.M., Calva, F. and Ruiz-Palacios, B.R. (Editors) Campylobacter V, Proc. 5th International Workshop on Campylobacter Infections, Na- tional Institute of Nutrition, Puerto Vallarta, Mexico, p. 102.
Patton, C.M., Mitchell, S.W., Potter, M.E. and Kaufman, A.F. (1981) Comparison of selective media for primary isolation of Campylobacter fetus subsp, jejuni. J. Clin. Microbiol. 13, 326-330.
Patton, C.M., Shaffer, N., Edmonds, P. et al. (1989) Human disease associated with Campylobacter upsaliensis (catalase-negative or weakly positive Campylobacter species) in the United States. J. Clin. Microbiol. 27, 66-73.
Pearson, A.D. and Healing, T.D. (1992) The surveillance and control of campylobacter infection. Communicable Disease Report Review, 2. Public Health Laboratory Service, London, R133-140.
Pennie, R.A., Zurino, J.N., Rose, C.E. and Guerrant, R.L. (1984) Economical simple method for production of the gaseous environment required for cultivation of Campylobacter jejuni. J. Clin. Microbiol. 20, 320-322.
Peterz, M. (1991) Comparison of Preston agar and a blood-free selective medium for detection of Campylobacter jejuni in food. J. Assoc. Off. Anal. Chem. 74, 651-654.
Plastridge, W.N., Williams, L.F. and Trowbridge, D.G. (1964) Antibiotic sensitivity of physiologic groups of microaerophilic vibrios. Am. J. Vet. Res. 25, 1295-1299.
Ray, B. and Johnson, C. (1984a) Sensitivity of cold-stressed Campylobacter jejuni to solid and liquid enrichments. Food Microbiol. 1, 173-176.
Ray, B. and Johnson, C. (1984b) Survival and growth of freeze-stressed Campylobacter jejuni cells in selective media. J. Food Safety 6, 183-195.
Razi, M.H.H. andPark, R.W.A. (1979) Studies of media for the isolation and storage of Campylobacter spp. J. Appl. Bacteriol. 47, x.
Ribeiro, C.D. and Price, T.H. (1984) The use of Preston enrichment broth for the isolation of 'thermophilic' campylobacters from water. J. Hyg., Camb. 92r
Ribeiro, C.D., Marks, J. and Grimshaw, A.D. (1985) Economic cultivation of 'thermophilic' Campy- lobacter spp. J. Clin. Pathol. 38, 1311-1313.
Rosef, O. (1981) Isolation of Campylobacter fetus subsp, jejuni from the gall bladder of normal slaughter pigs, using an enrichment procedure. Acta Vet. Scand. 22, 149-151.
Rosef, O. and Kapperud, G. (1983) House flies (Musca domestica) as possible vectors of Campylobacter fetus subsp, jejuni. Appl. Environ. Microbiol. 45, 381-383.
Rosef, O., Gondrosen, B., Kapperud, G. and Underdal, B. (1983) Isolation and characterisation of Campylobacter jejuni and Campylobacter coli from domestic animals and wild animals in Norway. Appl. Environ. Microbiol. 46, 855-859.

161
Rosef, O., Kapperud, G. and Skjerve, E. (1987) Comparison of media and filtration procedures for qualitative recovery of thermotolerant Campylobacter spp. from naturally contaminated surface water. Int. J. Food Microbiol. 5, 29-39.
Scotter, S.L., Humphrey, T.J. and Henley, A. (1993) Methods for the detection of thermotolerant campylobacters in foods: results of an inter-laboratory study. J. Appl. Bacteriol. 74, 155-163.
Shanker, S., Gordon, S.W., Fuller, H. and Gilbert, G.L. (1991) Sensitivity of a filtration method for the detection of Campylobacter/Helicobacter spp. in faeces. Microb. Ecol. Health Dis. 4, $47.
Skirrow, M.B. (1977) Campylobacter enteritis: a new disease. Brit. Med. J. 2, 9-11. Skirrow, M.B., Benjamin, J., Razi, M.H.H. and Waterman, S. (1982) Isolation, cultivation and identifi-
cation of Campylobacter jejuni and Campylobacter coli. In: Corry, J.E.L., Roberts, D. and Skinner, F.A. (Editors) Isolation and Identification Methods for Food Poisoning Organisms. Academic Press, London, pp. 313-328.
Skirrow, M.B., Vickery, C.R., Lovelock, M.R. and Evans, J.J. (1987) Cheap microaerobic atmospheres: modification of the methylated spirit burn method. In: Kaijser, B. and Falsen, E. (Editors) Campylobacter IV, Proc. 4th International Workshop on Campylobacter Infections, Univ. G6teborg, Sweden.
Skirrow, M.B., Merry, T.J. and Vickery, C.R. (1991) Hydrogen facilitates the isolation of Campylobacter jejuni. In: Ruiz-Palacios, G.M., Calva, F. and Ruiz-Palacios, B.R. (Editors) Campylobacter V, Proc. 5th International Workshop on Campylobacter Infections, Instituto Nacional de la Nutricion, Vasco de Quiroga 15, Mexico 1400, D.F., p. 109.
Steele, T.W. and McDermott, S.W. (1984) The use of membrane filters applied directly to the surface of agar plates for the isolation of Campylobacter jejuni from faeces. Pathology 16, 263-265.
Stelzer, W. and Jacob, J. (1992) Das Vorkommen von Campylobacter in einem Mittelgebirgsbach. Zbl. Mikrobiol. 147, 45-50.
Stelzer, W., Mochmann, H.P., Richter, U. and Dobberkau, H.J. (1988) Characterisation of Campylobac- ter jejuni and Campylobacter coli isolated from waste water. Zbl. Bakteriol. Hyg. A 269, 188-196.
Stern, N.J., Green, S., Thaker, N., Krout, D. and Chju, H. (1984) Recovery of Campylobacter jejuni from fresh and frozen meat and poultry collected at slaughter. J. Food Prot. 47, 372-374.
Stern, N.J. and Line, J.E. (1992) Comparison of three methods for recovery of Campylobacter spp. from broiler carcasses. J. Food Prot. 55, 663-666.
Stern, N.J., Wojton, B. and Kwiatek, K. (1992) A differential-selective medium and dry ice-generated atmosphere for recovery of Campylobacter jejuni. J. Food Prot. 55, 514-517.
Taylor, D.N., Kiehlbauch, J.A., Tee, W., Pitarangsi, C. and Echeverria, P. (1991) Isolation of group 2 aerotolerant Campylobacter species from Thai children with diarrhoea. J. Infect. Dis. 163, 1062-1067.
Tenova, F.C. and Fennell, C.L. (1992) The genera Campylobacter and Helicobacter. In: Balows, A., Truper, H.G., Dworkin, M., Hurder, W. and Schleifer, K.H. (Editors) The Prokaryotes IV, 2nd edition. Springer, New York, pp. 3488-3511.
Vandamme, P., Falsen, E., Rossau, R., Hoste, B., Segers, P., Tytgat, R. et al. (1991) Revision of Campylobacter, Helicobacter and Wolinella taxonomy: amendation of generic descriptions and proposal of Arcobacter gen. nov. Int. J. Syst. Bacteriol. 41, 88-103.
Vandamme, P. and Goossens, H. 1992 Taxonomy of campylobacter, arcobacter and helicobacter. Zbl. Bakteriol. 276, 447-472.
Vandamme, P., Vancanneyt, M., Pot, B., Mels, L., Hoste, B., Dewettinck, D. et al. (1992a) Polybasic taxonomic study of the amended genus Arcobacter with Arcobacter butzleri comb. nov. and Arcobacter skirrowi sp. nov., an aerotolerant bacterium isolated from veterinary specimens. Int. J. Syst. Bacteriol. 42, 344-356.
Vandamme, P., Pugina, P., Benzi, G., Van Etterijck, R., Vlaes, L., Kersters, K. et al. (1992b) Outbreak of recurrent abdominal cramps associated with Arcobacter butzleri in an Italian school. J. Clin. Microbiol. 30, 2335-2337.
Vanhoof, R., Vanderlinden, M.P., Diereckx, R., Lauwers, S., Yourassowsky, E. and Butzler, J.P. (1978) Susceptibility of Campylobacter fetus subsp, jejuni to twenty nine antimicrobial agents. Antimicrob. Agents Chemother. 14, 553-556.
Walder, M. (1979) Susceptibility of Campylobacter fetus subsp, jejuni to twenty antimicrobial agents. Antimicrob. Agents Chemother. 16, 37-39.

162
Warmsley, S.L. and Karmali, M.A. (1989) Direct isolation of atypical thermophilic Campylobacter species from human faeces on selective agar medium. J. Clin. Microbiol. 27, 668-670.
Waterman, S.C., Park, R.W.A. and Bramley, A.J. (1984) A search for the source of Campylobacter jejuni in milk. J. Hyg., Camb. 92, 333-337.
Weber, G., Manafi, M. and Reisinger, H. (1987) Die Beduntung von Yersinia enterocolitica und thermophilen Campylobactern ffir die Wasserhygiene. Zbl. Bakteriol. Hyg. B 184, 501-514.
Weinrich, K., Winkler, K. and Heberer, E. (1990) Untersuchungen zur Eignung ausgew~ihlter Humin- saurenchargen als Nahbodenzusatz fiir Medien sur Isolierung thermophiler Campylobacterspecies. Deusche Tierarztliche Wochenschrift 97, 511-515.
Wesley, R.D., Swaminathan, B. and Stadelman, W.J. (1983) Isolation and enumeration of Campylobac- ter jejuni from poultry products by a selective enrichment method. Appl. Environ. Microbiol. 46, 1097-1102.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
163
Chapter 10 Media for 'total' Enterobacteriaceae, coliforms
and Escherichia coli
R u t h M. Blood a, G . D . W . Curt is b
a 91 Little Bookham Street, Bookham, Leatherhead, Surrey KT23 3AF, UK. b Bacteriology Department, John Radcliffe Hospital, Oxford OX 3 9DU, UK
Abstract
Selective and enrichment media for the isolation and enumeration of 'total' Enterobacte- riaceae, coliforms, faecal coliforms and E. coli from foods are described. The effects of time and temperature of incubation are discussed. Reports of some comparative studies of these media and factors to be considered in the selection of media for this group of organisms are noted and the difficulties associated with the isolation and enumeration of pathogenic serotypes of E. coli considered.
The family Enterobacteriaceae consists of Gram-negative facultatively anaero- bic non-spore-forming bacteria that grow well on peptone, meat extract and, usually, MacConkey agar (Brenner, 1984). Members of the family constitute a large biochemically and genetically related group that shows substantial heterogeneity in its ecology, host range and pathogenic potential for man, animals, insects and plants. The family is subdivided into a number of genera, based on genetic and antigenic properties and, more traditionally, on biochemical characteristics and pathogenicity, as follows: Escherichia, Shigella, Salmonella, Citrobacter, Klebsiella, Enterobacter, Erwinia, Serratia, Hafnia, Edwardsiella, Proteus, Providencia, Mor- ganella, Yersinia and others. All ferment glucose. Those which ferment lactose are grouped together as 'coliform bacteria'. Table 1 shows the genera in which lactose fermenting species occur and known associations of pathogenicity.

164
Table 1 The Enterobacteriaceae and the "coliform" group (lactose-positive strains)
Genus Detected by Faecal "coliform test" origin
Enteropathogenic for man
b Citrobacter yes a no Edwardsiella no yes
b Enterobacter yes no Erwinia no d no b Escherichia yes yes Hafnia no c no b
b Klebsiella yes no b Proteus no no
Salmonella no yes Serratia no no Shigella no yes Yersinia no yes
n o
e some no no some e no
e some e some
yes n o
yes e some
a Except slow lactose-positive strains. b Some strains of faecal origin but also grow in other environments. c Except occasional strains. d Except strains adapted to rapid growth near 37~ e Some serotypes contain enteropathogenic strains. From ICMSF, 1978
En te robac te r i aceae are widely distr ibuted and found in soil, water, vegeta t ion and the intestinal tract of animals. Examinat ion of foods, ingredients and raw materials , for the presence of marke r groups such as coliforms or total Enterobac- ter iaceae, is one of the most common tests in the food microbiology laboratory, partly because of the relative ease and speed with which the tests can be accomplished. Where it is claimed that a food has been processed for safety, the finding of such organisms demons t ra tes a failure of the process or post-process contamina t ion (indicator function; Mossel, 1982). The presence of E. coli in such a food shows the possibility of the presence of enter ic pa thogens in the processing area (index function).
Several factors have influenced deve lopment of media for the detect ion, isola- tion and enumera t ion of members of the Enterobac te r iaceae . These include the derivation of the family name (entero = intestinal), the role of several genera as enter ic pa thogens and the long-known existence of E. coli as par t of the normal gut flora of man. Most cul ture media a t t empt to mimic the envi ronment of the intest ine by inclusion of bile salts or more def ined components with similar propert ies .
Media for specific pa thogens of the family En te robac te r i aceae (other than E. coli) are considered by o ther contr ibutors and this review will deal with media for the two groups of En te robac te r i aceae used as marke r organisms. These are i) the lac tose- ferment ing but otherwise ill-defined 'coliform' group, including ' faecal ' coliforms (supposedly of faecal origin and capable of growth at 44-45.5~ and E. coli and ii) the ' total ' En te robac te r iaceae . Media for E. coli in its role as an

165
enteropathogen will also be considered. The principal aspects to be discussed will include formulation and the effect of incubation temperature.
The origins of today's media go back to the early part of the century when MacConkey, reporting experiments to determine the distribution of 'Bacillus coli' in nature, published papers on lactose fermenting bacteria in faeces (1905) and on the use of bile salt media and their advantages in some bacteriological examina- tions. In his paper of 1908, MacConkey referred to earlier studies he had made in 1897 using potato juice medium with bile salts, and to studies with a medium containing bile salts (0.5%), peptone (2%), lactose (1%), neutral red (0.0025%) and agar (1.5%) in tap water. He found it was essential to use bile salts which were neutral in reaction to neutral red if acid production was to be easily recognised. The absence of sodium chloride from these early formulations results in a low electrolyte content which inhibits the swarming of Proteus spp. Such media are still marketed today under the name 'MacConkey agar w / o salt' and are used extensively in clinical laboratories. MacConkey concluded that the various lactose- fermenting organisms found in faeces could be further distinguished by their action on other carbohydrates incorporated either singly or in combination in bile salts-neutral red media and thus the foundations of the selective-indicator media in use today were laid.
An ideal medium for use in food microbiology, whether broth or agar, would selectively grow only the target organism or group of organisms. With a group of organisms so closely related as the Enterobacteriaceae it is unlikely that such a medium could be developed for a single genus such as Escherichia or even the less well defined coliform group and recourse must be had to the incorporation of substrates to provide the means of differentiating the target from the unwanted strains. Where selective indicator~ media have been formulated specifically with Enterobacteriaceae in mind, steps may need to be taken to prevent the growth of unwanted organisms by methods such as overlaying of the inoculated surface with agar (Mossel et al., 1979). Even with such precautions recovery of Enterobacteri- aceae will be affected by the incubation temperature used and the nature of the organisms present in the inoculum i.e. psychrophilic, mesophilic or thermophilic (Mossel et al., 1979). The difficulty of recovering sublethally injured organisms on selective media must also be considered when choosing suitable media for these bacteria.
1. Liquid media
Table 2 shows the main features of liquid media formulated for the growth of Enterobacteriaceae. Selectivity is provided by surface active agents, alone or in combination, in all but three of these broths. Lactose broth contains no selective agents or indicator and is recommended only for use in testing normally sterile material such as eggs and egg products (AOAC, 1990). Minerals modified gluta- mate (MMG) broth is a chemically defined, elective medium derived from formate lactose glutamate medium (Gray, 1964). Selectivity in boric acid lactose broth is
Table 2 Principal features of liquid media for Enterobacteriaceae
Medium Target Incubation
Group a (~ (h)
Selective agents Fermentable carbohydrate(s)/ differential system(s)
Reference
A-1
Boric acid lactose broth
Brilliant green bile broth (syn. Brilliant green lactose bile broth; BGBB)
EC broth
fc 35 3 followed by 44.5 21
Ec 43 48
cfm 35 48 Ec 44 18
cfm 37 48 Ec 45.5 48
Triton-X 100
boric acid
brilliant green, bile
bile salts
lactose, salicin, gas production
lactose
lactose, gas production
lactose, gas production
Andrews et al., 1981
Vaughn et al., 1951
Dunham and Schoenlein, 1926
Hajna and Perry, 1943
EC-MUG broth Ec 44.5 bile salts lactose, MUG hydrolysis Rippey et al., 1987

Enterobacteriaceae Ent b enrichment (EE) broth
4 32 44
10d brilliant green, 24-48 ox bile 18
glucose
GN broth - modified EEc 37 18-24 sodium citrate, sodium deoxycholate
lactose, arabinose
Lactose broth cfm
Lauryl tryptose broth cfm (syn. lauryl sulphate broth; Ec LST)
35
35 44
Lauryl tryptose - MUG broth Ec 35
24-48 (none)
24-48 lauryl sulphate
24-48 lauryl sulphate
lactose
lactose, gas and indole production
lactose, gas and indole production, MUG hydrolysis
MacConkey broth cfm 30/37 48 ox bile lactose, gas production
Minerals modified glutamate cfm 30 48 (chemically defined lactose, gas production (MMG) broth elective medium)
a cfm = coliforms, Ec = E. coli, EEc = enteropathogenic E. coli, Ent = Enterobacteriaceae, fc = faecal coliforms. b Incubation conditions will determine which group of Enterobacteriaceae is detected.
Mossel et al., 1963
AOAC., 1990
Mallman and Darby, 1941
Feng and Hartman, 1982
MacConkey, 1905
Abbiss et al., 1981

168
obtained by the use of boric acid. The triphenylmethane dye, brilliant green, is used to inhibit Gram-positive organisms and the combination of sodium deoxy- cholate and sodium citrate in GN broth inhibits Gram-positive and some Gram- negative organisms.
Various differential systems are found in these broths. With the exception of Enterobacteriaceae enrichment (EE) broth which contains glucose, all the media incorporate lactose, with or without a pH indicator to demonstrate acid produc- tion. Other carbohydrates readily utilised by the target organisms are also found in some formulations. Production of gas from carbohydrate fermentation may be detected with the aid of an inverted Durham's tube. Lauryl (sulphate) tryptose (LST) broth (Mailman and Darby, 1941)was said to be suitable for the detection of indole production after addition of the appropriate reagents.
/3-D-Glucuronidase activity is a specific marker for E. coli and some Shigella and Salmonella strains. 4-Methylumbelliferyl-/3-D-glucuronide (MUG) incorporated in a broth will be cleaved by the enzyme present in these strains, releasing the compound 4-methylumbelliferone which fluoresces when exposed to long-wave- length (about 360 nm) UV light. The benefits of MUG incorporation into media for MPN procedures were demonstrated by Moberg et al. (1988)who found that use of LST-MUG allowed the omission of the confirmation step for faecal coliforms in the examination of chilled and frozen foods that were not highly contaminated, thus reducing the overall time required for the test. The problem of endogenous /3-D-glucuronidase in molluscan shellfish, which causes a high rate of false positive results, can be overcome by incorporation of the fluorescent sub- strate into the confirmation broths rather than in the primary growth medium (Koburger and Miller, 1985; Rippey et al., 1987). For a review of the use of /3-D-glucuronidase in the detection and identification of E. coli, see Frampton and Restaino (1993). It is important to note that enterohaemorrhagic E. coli, including O157 :H7 strains, do not express this enzyme. The absence of a positive MUG test has been used as a method of selectively isolating such strains (Szabo et al., 1986).
The examination of shellfish and shellfish-growing waters may require a special technique due to the salinity of the natural environment and the adverse effect this may have on the target organisms. Medium A-1 (Andrews and Presnell, 1972)was designed for this purpose and the A-1 technique has been adopted as an approved method by the AOAC for the estimation of faecal coliforms in shellfish growing waters. The medium contains, in addition to the lactose, tryptone and sodium chloride found in most media for these organisms, the surfactant Triton X-100 and salicin which is a carbohydrate readily utilised by E. coli.
2. Solid media
The principal features of agars formulated for this group of organisms are shown in Table 3. As with the broths, bile salts are used in many of these media to inhibit Gram positive organisms but dyes are also used, alone or in combination with bile salts or, in Endo agar, sodium sulphite. The intrinsic toxicity of some

169
batches of media to non-stressed Enterobacteriaceae was shown by Mossel et al. (1979) to be a problem which could be overcome by careful selection of crystal violet and, particularly, bile salts.
The most common differential system is a fermentable carbohydrate together with a pH indicator. Tryptone bile agar (TBA) is the only medium not containing a fermentable carbohydrate. Lactose is incorporated in all the others, except violet red bile glucose (VRBG) agar which contains glucose and is designed as a 'total Enterobacteriaceae' medium. Where total Enterobacteriaceae are sought using such MacConkey type agars Mossel et al. (1979) have shown that glucose may be substituted for lactose and that this may result in higher and more consistent counts.
Other differential systems are found in some media. Hydrogen sulphide produc- tion is detected in deoxycholate hydrogen sulphite lactose agar by the blackening of H2S-producing colonies due to the formation of iron sulphide. In Endo agar coliform bacteria produce aldehyde and acid from lactose. The aldehyde liberates fuchsin from the fuchsin-sulphite compound resulting in a red coloration of the colonies due to the fuchsin, a reaction so intense with E. coli that the fuchsin crystallizes out and gives to the colonies a characteristic green metallic sheen.
Production of gas at 44.5~ by faecal coliform organisms is demonstrated in an unusual way in fecal coliform agar (FCA; Chen and Wu, 1992). Calcium lactate in the medium reacts with CO 2 to form precipitates producing yellow to yellow-green colonies surrounded by pale yellow zones. The detection of indole production by E. coli in the system of Holbrook et al. (1980) requires a special technique (vide inffa). In the pour-plate with overlay method of Hall (1984) the inoculum is mixed with a small volume of resuscitant (non-selective) agar containing yeast extract, tryptone, sodium pyruvate, sodium glycerophosphate and magnesium sulphate. This is subsequently overlaid with the selective agar which is formulated so that the lactose content, whilst adequate for acid production, does not interfere with the utilization of tryptophan. Bromothymol blue is used as the pH indicator to avoid interference with the indole test which is done using a rapid micro-tube technique.
Incorporation of MUG into violet red bile agars allows the detection of /3-D-glucuronidase activity by exposure of the incubated plates to UV light (Feng and Hartman, 1982).
3. Membrane methods
A procedure for the estimation of E. coli in foods using cellulose membranes was developed by Anderson and Baird-Parker (1975) from a method described by Delaney et al. (1962). The method did not include a resuscitation step and was subsequently modified (Holbrook et al., 1980) to make provision for this by pre-incubation of the inoculated membrane on a non-inhibitory agar before transfer to TBA, a technique which also reduces the carbohydrate content in the inoculum by allowing diffusion into the non-inhibitory agar. This modification not only enables resuscitation of damaged cells but also renders the test for indole
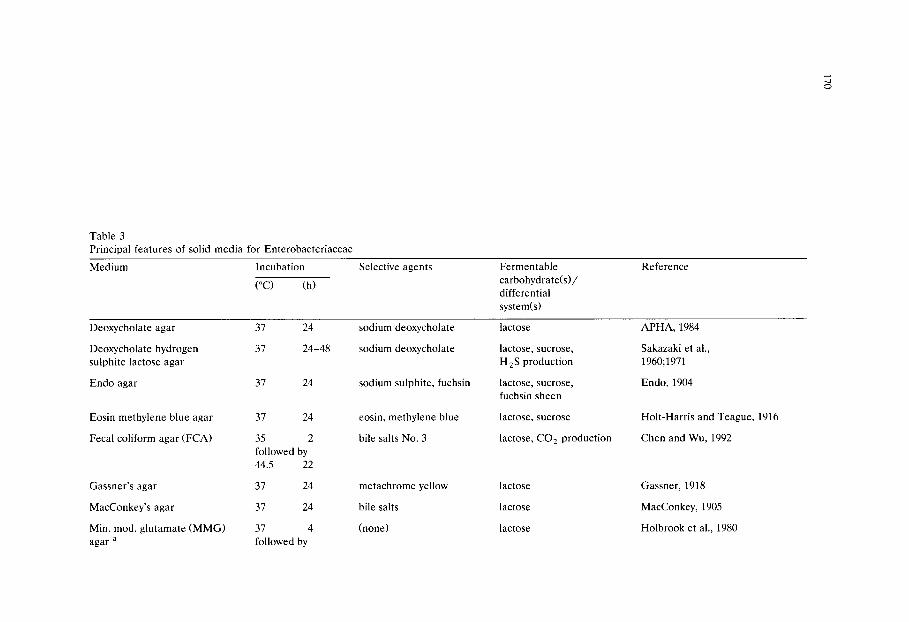
Table 3 Principal features of solid media for Enterobacteriaceae
Medium Incubation Selective agents
(~ (h)
Fermentable carbohydrate(s)/ differential system(s)
Reference
Deoxycholate agar 37 24
Deoxycholate hydrogen 37 24-48 sulphite lactose agar
Endo agar 37 24
Eosin methylene blue agar
Fecal coliform agar (FCA)
Gassner's agar
MacConkey's agar
Min. mod. glutamate (MMG) a agar
37 24
35 2 followed by 44.5 22
37 24
37 24
37 4 followed by
sodium deoxycholate
sodium deoxycholate
sodium sulphite, fuchsin
eosin, methylene blue
bile salts No. 3
metachrome yellow
bile salts
(none)
lactose
lactose, sucrose, H 2 S production
lactose, sucrose, fuchsin sheen
lactose, sucrose
lactose, CO 2 production
lactose
lactose
lactose
APHA, 1984
Sakazaki et al., 1960;1971
Endo, 1904
Holt-Harris and Teague, 1916
Chen and Wu, 1992
Gassner, 1918
MacConkey, 1905
Holbrook et al., 1980

Tryptone bile agar (TBA)
b Resuscitant agar
44 18-24 bile salts No. 3
30 0.5 (none) followed by
indole production
(none) Hall, 1984
Yeast extract lactose tryptone bile agar
Violet red bile glucose agar (VRBG)
44 18-20 bile salts No. 3
4 c 10d bile salts No. 3, crystal 32 24-48 violet 42-44 18
lactose
glucose Mossel et al., 1978
Violet red bile (VRB) agar 4 c 10d bile salts No. 3, crystal 32 24-48 violet 42-44 18
lactose APHA, 1984
Violet red bile M U G agar 37 18-24 bile salts No. 3, crystal lactose, MUG hydrolysis Feng and Hartman, 1982 violet
a Inoculum is spread over the surface of an acetate membrane overlaid on MMG agar. After the initial incubation period the membrane is transferred to
TBA. b Inoculum is mixed with molten resuscitant agar which is allowed to set. After the initial incubation period the plates are overpoured with yeast extract
lactose tryptone bile agar. c Incubation conditions will determine which group of organisms is detected.

172
production after incubation on TBA more reliable. The modified method is as follows. A cellulose acetate membrane is laid on the surface of a pre-dried plate of MMG agar and the inoculum is spread over the membrane. After the membrane surface has dried the plate is incubated at 37~ for 4 h and the membrane is then transferred to a pre-dried TBA plate and incubation continued for a minimum of 18 h at 44.5~ At the end of this second incubation the membranes are examined. Those with growth are removed from the agar into a Petri dish lid containing 2 ml of indole reagent (Vracko and Sherris, 1963) so that the whole of the lower surface is wetted. The excess reagent is removed after 5 min and the reaction developed in bright sunlight or under a UV lamp. Indole positive colonies, which appear pink, may then be counted to give the total E. coli count. Further confirmation of identity is generally regarded as unnecessary. The cost of the examination can be greatly reduced if cellulose nitrate membranes are used instead of cellulose acetate. Holbrook and Anderson (1982) specify either.
A similar system is the hydrophobic grid membrane filter technique (Entis, 1984). This uses a membrane printed with hydrophobic lines in a grid pattern which acts as a barrier to the spread of colonies. Dilutions of food to be examined are filtered through the membrane which is then placed on the surface of the appropriate agar plate. Resuscitation by the use of a short incubation period on non-selective agar prior to overnight incubation on selective agar is possible and testing for indole production can also be accomplished in much the same manner as Holbrook et al. The number of squares occupied by colonies is counted and converted to MPN values of organisms by the use of a formula. By the use of different media and incubation conditions counts may be made of total coliforms, faecal coliforms or E. coli. Filters and equipment are available commercially.
4. Incubation conditions
There are many recommendations for incubation temperatures and times. Apart from the use of refrigeration temperatures to isolate psychrotrophs, the choice is of temperatures between 30 and 45.5~ When examining dairy products for coliform organisms it is standard practice to incubate at 30~ rather than 35 or 37~ With most other foods the choice lies between 35~ favoured by American workers, and 37~ more frequently used by Europeans. A common practice is to incubate primary tests at 35 or 37~ and confirmatory tests for coliforms at 35-37~ and for faecal coliforms, including E. coli, at 44-45.5~ depending on the medium used. It should be noted that aberrant strains may be encountered, e.g.E. coli O157:H7, which behave atypically at elevated temperatures.
Weiss et al. (1983) studied MPN counts of faecal coliforms and E. coli using LST at 35~ for primary culture and with confirmation in EC at 44.5, 45.0 and 45.5~ Only 29% of raw milk and 33% of raw meat samples gave equal recovery rates of faecal coliforms at the three temperatures when gas production was the criterion of positivity (standard MPN). Where growth, with or without gas produc- tion, was the criterion (total MPN), 36% of the milk and 47% of the meat samples

173
showed no difference in recovery at the three temperatures. They concluded that incubation at 44.5~ resulted in the greatest sensitivity for E. coli whilst the use of 45.5~ gave the greatest specificity.
With regard to the length of incubation Mehlman (1984) stated that the total incubation time may be limited to 24 h as in water or shellfish bacteriology or extended to 48 h as in the classical identification of the coliform group. The longer period will recover those E. coli isolates which develop after 24 h and thus the 48 h test generally produces higher counts than similar tests of only 24 h duration (Moussa et al., 1973; Pierson et al., 1978; Bindschedler et al., 1981). Weiss et al. (1983) found that confirmed coliform MPN counts at 24 and 48 h differed by more than 0.6 log cycle in 23% of raw milk and 48% of raw meat samples. Methods specifying a total of 48 h incubation often recommend examination of tubes at 24 h when confirmatory tests on presumptive positive tubes can be initiated. More confirmatory tests may need to be set up if extended incubation reveals more presumptive positive tubes.
Temperatures and duration of incubation recommended for plated media are less diverse than those for broths although similar reasoning has been applied. Pre-incubation for 2-4 h at a lower temperature prior to overnight incubation at 44 or 44.5~ is said to allow recovery of injured cells on FCA and TBA. Incubation of VRBG and VRB agars at 4~ for 10 days is recommended for optimal growth of psychrotrophic coliforms whilst the same agars should be incubated at 32~ for 24-48 h or 42-44~ for 18 h for mesophilic or thermotrophic coliforms respec- tively (Anon., 1987). Mossel et al. (1979) have shown that incubation at 30~ led to higher confirmed colony counts in minced meat than at 37~ but this was not the case when samples of drip from frozen broilers were examined. This difference was explained by the predominance of psychrotrophic types (up to 75% of isolates) in minced meat whereas in chicken drip E. coli was the predominant type with psychrotrophs making up only 30% of isolates. A further study (Mossel et al., 1986) showed that psychrotrophs were not markedly inhibited at 37~ although growth was variable at this temperature. They recommended the use of 30~ for incuba- tion of violet red bile agars where psychrotrophs are sought and 42-43~ where it is desired to suppress their growth.
5. Comparative studies
Numerous comparative studies of media have been made but their value is limited by the differences in sample materials, conditions of testing and formula- tions surveyed. In the examination of frozen food, both Shelton et al. (1962) and Hall (1964) preferred LST broth to lactose broth for detection of coliforms using the MPN technique, gas positive tubes being subcultured to brilliant green bile lactose broth (BGBB) for confirmation. Further comparisons of media for the isolation of coliform organisms from a wide variety of dehydrated and deep frozen foods were reported by Moussa et al. (1973). They found that for the detection of coliforms and Enterobacteriaceae, MMG broth was significantly better than BGBB,

174
LST and EE broths for all types of samples tested except frozen poultry where the difference was not significant. For the detection of faecal coliforms MMG broth was again significantly better than the other media both for dehydrated products and deep frozen foods. For deep frozen poultry the best results were obtained with lactose broth but this was not a good medium for deep frozen products as a whole. In their hands BGBB performed poorly in the determination of coliforms and Enterobacteriaceae as did EE broth for faecal coliforms. In the examination of ground beef Pierson et al. (1978) found LST broth gave higher coliform counts than either MacConkey broth or BBGB whilst EC broth and BGBB did not differ significantly in their ability to detect faecal coliforms. Further studies on raw meats and poultry were made by Rayman et al. (1979)who found the membrane method of Anderson and Baird-Parker (1975) was less variable, more rapid and economical than the A O A C MPN method using LST broth. When examining frozen samples they incorporated a resuscitation step by spreading the inocula on the surface of membranes overlying tryptic soy agar and incubating for 4 h at 35-37~ The membranes were then aseptically transferred to TBA and incubation continued for 24 h at 44.5~ The inclusion of this resuscitation step in the membrane method may account for the significantly higher counts which were obtained compared to the A O A C MPN technique.
In a collaborative study involving five laboratories (Sharpe et al., 1983) mem- brane filter methods gave significantly higher counts of E. coli in ground beef, cheese and cut green beans than an MPN technique using LST broth at 35~ with confirmation in EC broth at 44.5~ Results for the membrane techniques were obtained within 24 h whereas the MPN method, which required confirmation using IMViC tests, might require 10-14 days. These workers felt unable to recommend the filter methods for bean or alfalfa sprouts however, noting that these often contained high levels of Klebsiella spp.
Soft cheeses, cooked meats and pate were examined by Abbiss et al. (1981) using MMG broth, LST, MacConkey broth and BGBB. They found that with an MPN technique MMG broth was superior in sensitivity for coliform counts and that it compared favourably in specificity to the other broths. Other dairy samples were examined by Cooke and Jorgenson (1977)who found BGBB equal in sensitivity to MMG broth and better than lactose broth for the detection of coliforms in dried milk and butter. Work with dairy and cocoa products (Bindsche- dler et al., 1981) comparing BGBB, LST, lactose broth, EE broth and MMG broth for the determination of coliforms and E. coli showed that MMG broth had the highest sensitivity for coliform detection. However, it was also the medium which gave rise to the most false positive results and confirmatory tests were therefore required. In general, these studies showed that whenever a significant difference existed in the sensitivity to determine E. coli, MMG broth was better than the other media. Frequently EE broth was less satisfactory for the detection of Enterobacteriaceae than the other media. None of these methods included a specific resuscitation step although Bindschedler et al. carried out their first incubation at 30~ for 48 h and MMG is an elective medium which should allow recovery of damaged cells (Holbrook et al., 1980).

175
Andrews et al. (1981) evaluated the A-1 and LST broths for faecal coliform detection in a variety of foods and showed that the efficiency of the A-1 procedure was dependent on the type of food analyzed. They recommended it only for use as a screening test in food examinations.
Motes et al. (1984) compared BGBB, LST, direct plating on membranes placed on TBA and a roll tube technique with MacConkey agar for the examination of shellfish. The best recoveries were obtained using LST and BGBB with the lowest recovery on direct plating. The membrane method did not include the resuscita- tion step now in common use (Holbrook et al., 1980) and the poor success of the method may have been due to the failure to recover stressed organisms exposed directly to the selective TBA. A similar explanation was suggested by West and Coleman (1986) for the poor performance of direct plating methods in their comparison of techniques for the detection of E. coli in shellfish. An MPN method using MMG broth gave higher recoveries of confirmed E. coli than either a roll tube or pour plate technique with MacConkey agar No.3 even when a 2 h resuscitation at a lower temperature was introduced. They concluded that for both heavily (> 1000 E. coli/lO0 g) and lightly (100-500/100 g) contaminated shellfish, MPN procedures had greater sensitivity and that the MPN is technically superior to plate methods for E. coli since it determines gas as well as acid production. Nevertheless they recognised that confirmation was required. By inoculating tryp- tone water in parallel with BGBB and incubating both at 44~ their system allowed the completion of the examination within 48 h.
A collaborative and comparative study of coliform determination by MPN techniques (Silliker et al., 1979) examined three methods: i) presumptive determi- nation of coliforms using LST broth with subsequent confirmation of gas-positive tubes, ii) determination of coliforms with MacConkey broth, and iii) presumptive determination with BGBB followed by confirmation. Differences between the methods were relatively small when compared with the differences between laboratories using the same methods and following a common protocol. They were therefore of the opinion that there was no basis for selecting a single procedure as the 'best' method.
In a variety of fresh and processed foods examined without resuscitation Oblinger et al. (1982) found no significant differences between recoveries on VRB and VRBG agars. The most productive temperature for incubation was 20~ followed by 35, 7, 45 and I~ VRBG was not significantly better than VRB for recovery of total Enterobacteriaceae but typical colonies were more easily recog- nised and enumerated on VRBG than on VRB. Recovery of total Enterobacteri- aceae on both agars at 35~ was similar to that at 20~ Klein and Fung (1976) showed that on VRB agar incubated at 44.5~ faecal coliforms from sewage or water could be quantified and accurately separated from non-faecal coliforms by differences in colony size. They found no significant differences between recover- ies using the elevated temperature VRB method, a membrane filtration method and a five tube MPN system with EC broth. A comparison of pour-plate with overlay and spread plate techniques for counting Enterobacteriaceae in minced meat using VRBG agar (Murthy and Bachhil, 1982) demonstrated significantly

176
higher recoveries with the spread plate technique. They suggested that the better performance of the spread plates was due to the oxygen requirements of the bacteria and cited the work of Hechelmann et al. (1973)who reported low Enterobacteriaceae counts under anaerobic conditions. However, Mossel et al. (1978) considered that the overlay procedure, whilst establishing a sufficiently reduced oxygen tension to suppress the growth of strictly aerobic Gram negative organisms, also enhanced anaerobic glucose utilization by the Enterobacteriaceae resulting in colonies of more characteristic appearance.
The problem of resuscitation of stressed organisms was addressed by the VRB-2 pour-plate procedure (Hartman et al., 1975) which used a base layer of plate count agar (PCA) overlayered with double strength VRB (VRB-2) agar. Recoveries using this system were greater than with the conventional system using VRB agar for both layers, increases of 31% with raw milk, 70% with ice cream and 61% with cottage cheese being reported from a multicentre study (Marshall et al., 1978). Reber and Marshall (1982) compared the performance of a pour-plate system using VRB for both layers and PCA/VRB-2 agar for the recovery of stressed coliforms from stored acidified half-and-half (a cream product) and showed that PCA/VRB-2 recovered 20% more coliforms than VRB agar. Comparisons of recoveries of heat- and chlorine-injured E. coli cells by an MPN method using lactose tryptone-MUG broth and the VRB-2 method incubated at 35~ for 24-48 h showed the MPN method to be superior (Feng and Hartman, 1982).
A comparison of two plate methods incorporating resuscitation steps with direct counting on VRB agar and MPN estimations in MacConkey broth together with confirmation tests (Hall, 1984) showed that the membrane transfer method of Holbrook et al. (1980) gave recoveries almost as high as the pour-plate with overlay method of Hall.
The membrane method has the advantage of direct indole testing on all colonies in one simple operation whereas Hall's method involves removal of the agar from the base of the Petri dish, its inversion into the lid and subsequent selection of colonies for individual testing. The sometimes anomalous results of the indole reaction may have accounted for the slightly lower counts obtained by the mem- brane method in Hall's study when compared with his medium. The relative simplicity of the membrane method makes it less time consuming. Hall's medium provides information on lactose fermentation, but the exposure of organisms which may have been stressed, to molten agar at 45~ before resuscitation is a disadvan- tage. Both two-stage methods gave higher recoveries of E. coli than the other methods, with counts approximately three times higher than on VRB agar and twice the MPN estimation.
Chen and Wu (1992) in their report on FCA (Table 3) found that recovery of freeze-stressed E. coli on FCA was about 1 log cycle lower than on nonselective media. Although their technique involves preincubation at 35~ for two hours prior to overnight incubation at 44.5~ this preincubation is carried out on the selective medium which may account for the reduction in the count on FCA compared to PCA. When the method was compared with the FDA MPN method

177
(FDA, 1984), which is without any resuscitation step, counts on 32 food samples were not significantly different.
Bredie and de Boer (1992)compared commercially available/3-D-glucuronidase based methods with the ISO standard MPN (1991a) method and the Anderson and Baird-Parker (1975) procedure for enumeration of E. coli in naturally contami- nated foods of animal origin. They found the Anderson and Baird-Parker and the commercially produced Petrifilm R E. coli methods useful for routine counting of E. coli in raw meat, poultry and meat products. The MPN procedure was more sensitive but impractical and considerably more expensive. A collaborative study in 24 laboratories (Entis, 1989) examined a hydrophobic grid membrane filter method incorporating the use of MUG for the enumeration of total coliform and E. coli in foods by comparing its performance against the AOAC 3-tube MPN method. The total coliform methods did not differ significantly in the examination of raw milk, raw ground poultry, whole egg powder and cheese powder but the MPN method detected a significantly higher number of organisms in ground black pepper. The hydrophobic grid membrane method detected significantly higher numbers of E. coli present in egg powder, cheese powder and ground black pepper samples while not differing significantly from the 3-tube method for the raw milk and raw ground poultry samples.
An assessment of media containing MUG and a chromogenic substrate, 5- bromo-4-chloro-3-indolyl-/3-D-galactoside (X-gal), demonstrated that cleavage of X-gal is a quicker and more sensitive parameter for total coliforms than gas production and that the combination of fluorescence and indole production is slightly superior to fluorescence and gas production for the identification of E. coli (Hahn and Wittrock, 1991).
West and Coleman's (1986)work on the isolation of E. coli from shellfish has been referred to above. Their study included a comparison of various solid media used as pour plates, spread plates, sandwich plates and roll tubes. With some systems a resuscitation step consisting of 2 h incubation at 30~ prior to overnight incubation at 44~ was included. No significant differences were found between recoveries on the agar media, whether used with or without this resuscitation step, although counts were higher with the methods that used resuscitation. Failure to achieve a significant increase in the counts despite the pre-incubation at a lower temperature is probably due to the presence of bile salts in the medium on which resuscitation was attempted, or possibly to the use of molten agar to overlay plates (Murthy and Bachhil, 1982).
6. Media for enteropathogenic E. coli
In 1971 a large outbreak of gastroenteritis in the USA was traced to the consumption of imported soft cheeses (Marier et al., 1973). E. coli O124 : H17 was isolated from the faeces of the affected patients and from samples of the cheeses. This first reported isolate of enteropathogenic E. coli from food exemplifies many of the problems encountered in the detection of this group of organisms. The

178
strain was atypical in that it was late in fermenting lactose and it also shared O antigens with Shigella dysenteriae 3 thus adding to the difficulties of identification. Mehlman and Romero (1982) showed that traditional enrichment may not be suitable for the recovery of pathogenic types of E. coli and that many of these strains did not produce gas or indole at 44~ They found that only 31% of pathogenic strains could be quantitatively recovered by the A O A C (1980) method, that 30% were inhibited on MacConkey agar, 43% on eosin methylene blue agar and 17% by TBA. Substitution of Tween 80 for bile salts No. 3 in TBA and the addition of yeast extract (0.1%) resulted in an agar which supported growth at 44~ of all pathogenic strains tested.
The enteropathogenicity of certain serovars of E. coli is now well established but specific techniques for their isolation are confined to those serovars, the most important of which is O157 : H7, which do not produce acid from sorbitol within 24 h. This property led to the formulation of a MacConkey-type agar with lactose replaced by sorbitol for isolation of such strains from faeces (Rappaport and Henig, 1952) and this sorbitol MacConkey (SMAC) agar is the basis of all agars subsequently developed for this group of organisms. Other serovars have been incriminated in foodborne outbreaks of gastroenteritis (Mehlman and Romero, 1982), but because of the severity of the illness it causes O157:H7 has attracted most attention and other serovars will not be further considered in this review.
E. coli O157:H7 differs from most E. coli in its inability to ferment sorbitol but also fails to produce/3-o-glucuronidase and does not grow well at temperatures of 44-45.5~ (Doyle and Schoeni, 1984). Raghubeer and Matches (1990) showed that the temperature range for growth and gas production in EC broth within 48 h was 19.3-41~ Since incorporation of MUG into media, incubation at 44-45.5~ and observation of gas production at 44~ are commonly used in the enumeration of E. coli in foods, it is important to realise that such techniques will not detect this pathogenic serotype. Isolation of the organism from stools of infected patients, where large numbers are usually present, is easy to accomplish using SMAC (March and Ratnam, 1986) but recovery from foods is much more difficult since numbers are much lower and the use of selective enrichment favours strains of environmental origin compared to strains of human origin and may also result in loss of plasmids which encode for virulence (Hill et al., 1985). Szabo et al. (1986) addressed these problems by modifying TBA to produce haemorrhagic coli (HC) agar. This medium has a reduced amount of bile salts No. 3 (0.112%) and contains additionally, sodium chloride (0.5%), sorbitol (2%), MUG (0.01%) and bromocre- sol purple (0.0015%). The reduced bile salt content and the addition of sodium chloride allowed O157 :H7 strains to grow at 44.5~ None of the O157 :H7 strains which they tested fermented sorbitol within 48 h and the O157:H7 colonies thus appeared blue in contrast to colonies of E. coli types I and II which produced a yellow colour due to the formation of acid as a result of sorbitol fermentation. Since O157 :H7 strains do not produce/3-D-glucuronidase their colonies did not fluoresce under UV light. The medium was designed for use with a membrane technique, either that of Anderson and Baird-Parker (1975) or the hydrophobic grid membrane method (Entis, 1984) and testing for indole production can be done

179
in the usual way for these methods. However, since sorbitol positive organisms will metabolise sorbitol preferentially to tryptophan, type I organisms remain yellow in this test, i.e. apparently negative, even though they are true indole producers. As with the modification to the Anderson and Baird-Parker method (Holbrook et al., 1980) incubation of the membrane on the selective HC medium can be preceded by a period of resuscitation on a non-selective agar. A modification of this method using hydrophobic grid membranes incubated on HC agar followed by specific staining with a monoclonal antibody against E. coli O157:H7 conjugated to horseradish peroxidase has reliably demonstrated the absence of E. coli O157:H7 at a level of 10 cells per g of food within one day (Todd et al., 1988). Conner (1992) was unable to recover cells of E. coli O157:H7 which had been salt- or cold-in- jured before plating direct to SMAC and suggested that incorporation of Mg ++ ions into SMAC might enhance the recovery of such cells. In a later publication however (Ahmed and Conner, 1993), the addition of Mg ++ to SMAC for the recovery of heat injured E. coli O157 :H7 was said to compromise the medium and the authors recommended phenol red sorbitol agar with MUG or 1% pyruvic acid. Improved selectivity and differentiation was claimed for the CR-SMAC agar of Chapman et al. (1991)who used it to isolate O157 strains from faeces. Cefixime was incorporated to eliminate some Proteus spp. which would otherwise mimic the target organisms and other non-O157 E. coli strains which did not ferment sorbitol were differentiated by the addition of rhamnose which, like sorbitol, was not attacked by O157 :H7 strains. A benefit of the inclusion of rhamnose in this medium is that it is fermented by E. hermanni thus eliminating another possible source of error. Addition of rhamnose to SMAC more than doubles the cost of the medium and the authors make no comment on the effect of the addition of cefixime on the shelf life of the prepared agar. In summary it can be said that media for the isolation of pathogenic serotypes of E. coli from foods are not well developed and a combination of culture and one of the newer, non-cultural techniques may be the most promising way to achieve specific results.
7. Concluding remarks
There can be little surprise that in the course of nearly a hundred years of bacteriology, during which time basic techniques have changed very little, numer- ous formulations of culture media have been recommended for growing the most studied of all organisms, E. coli. Perhaps the greatest surprise is that a formula first published by MacConkey in 1905 should still be in use in a recognisable form today. With such a wide field from which to choose it is difficult to make recommendations for particular situations but it is possible to make some observa- tions of a general nature from a study of the literature and many situations are covered by standard methods, some promoted by regulatory agencies. Many authorities e.g. ICMSF (Table 4) and ISO (Table 5) offer a choice of more than one technique for the enumeration of this group of organisms detailing both MPN and solid media methods. In deciding which type of method to use consideration

Table 4 Media recommended by ICMSF (1978) for examinations for coliforms and Enterobacteriaceae in foods (details of media are shown in Tables 2 and 3)
Organisms or Medium (incubation temperature)
group sought Coliform isolation Coliform Faecal coliform E. coli Enterobacteriaceae (technique) confirmation confirmation confirmation isolation
Coliforms LST broth BGBB EC broth (MPN: N. America) (35-37~ (35-37~ (44.5~
Coliforms MacConkey broth BGBB IMViC tests (MPN: British) (35-37~ Peptone water a
(44~ Coliforms BGBB VRB or Endo BGBB IMViC tests (MPN: confirmed) (35-37~ agar (35-37~ Peptone water a
(44~ Coliforms VRB agar b IMViC tests (colony count) (35-37~
Enterobacteriaceae (presence/absence; preenriched in BPW c)
Enterobacteriaceae (colony count)
EE broth o (35-37~
VRBG agar (35-37~
a For indole test. b With overlay. c Buffered peptone water. o Confirm by oxidase test and mode of attack on glucose.
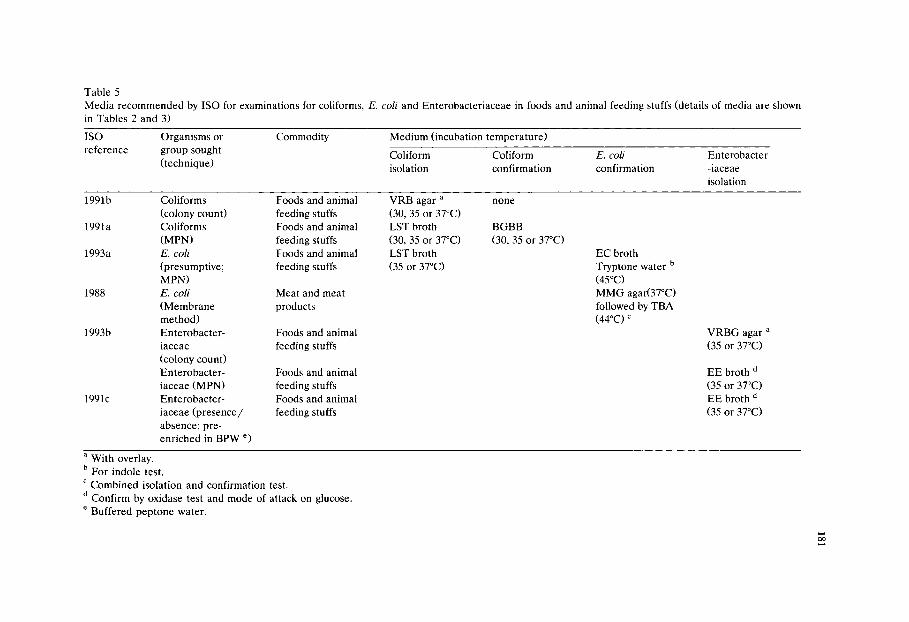
Table 5 Media recommended by ISO for examinations for coliforms, E. coli and Enterobacteriaceae in foods and animal feeding stuffs (details of media are shown
in Tables 2 and 3)
ISO Organisms or Commodity Medium (incubation temperature)
reference group sought Coliform Coliform E. coli Enterobacter (technique) isolation confirmation confirmation -iaceae
isolation
199 lb Coliforms Foods and animal (colony count) feeding stuffs
1991 a Coliforms Foods and animal (MPN) feeding stuffs
1993a E. coli Foods and animal (presumptive; feeding stuffs MPN)
1988 E. coli Meat and meat (Membrane products method)
1993b Enterobacter- Foods and animal iaceae feeding stuffs (colony count) Enterobacter- Foods and animal iaceae (MPN) feeding stuffs
1991c Enterobacter- Foods and animal iaceae (presence/ feeding stuffs absence; pre- enriched in BPW e)
VRB agar a
(30, 35 or 37~ LST broth (30, 35 or 37~ LST broth (35 or 37~
none
BGBB (30, 35 or 37~
EC broth Tryptone water b (45~ MMG agar(37~ followed by TBA (44oc) c
VRBG agar (35 or 37~
EE broth d (35 or 37~ EE broth d (35 or 37~
a With overlay. b For indole test. c Combined isolation and confirmation test. d Confirm by oxidase test and mode of attack on glucose. e Buffered peptone water.

182
should be given to the fact that, where low numbers are sought, M P N techniques are more sensitive than plating methods. They are also subject to greater variation, slower to arrive at a confi rmed result and more expensive and labour-intensive. Plating methods , particularly when they incorporate a rapid confirmation step as in
the technique of Holbrook et al. (1980) produce confirmed results more speedily at the expense of some sensitivity. A fur ther point to be considered is that the use of a resuscitat ion step is essential if injured organisms are likely to be encountered . This may simply involve pre- incubat ion of a selective med ium at a lower tempera- ture before the definitive incubation at an elevated (44-45.5~ t empera tu re or,
preferably, the use of one of the m e m b r a n e transfer methods with pre- incubat ion on a non-selective agar at 35 or 37~ for two to four hours followed by transfer to
selective agar at 44-45.5~ for overnight incubation.
Pa thogenic serotypes of E. coli present a part icular problem and no universally applicable culture me thod has yet been evolved for their enumera t ion in food. It may well be that isolation and differentiat ion of these organisms will best be
accomplished by a combinat ion of culture and molecular biological methods .
References
Abbiss, J.S., Wilson, J.M., Blood, R.M. and Jarvis, B. (1981) A comparison of minerals modified glutamate medium with other media for the enumeration of coliforms in delicatessen foods. J. Appl. Bacteriol. 51, 121-127.
Ahmed, N. and Conner, D.E. (1993) Evaluation of different media for recovery of thermally-injured Escherichia coli O157:H7. J. Food Prot. 56, 913.
Anderson, J.M. and Baird-Parker, A.C. (1975) A rapid direct plate method for enumerating Escherichia coli biotype I in foods. J. Appl. Bacteriol. 39, 111-117.
Andrews, W.H. and Presnell, M.W. (1972) Rapid recovery of Escherichia cold from estuarine waters. Appl. Microbiol. 23, 521-523.
Andrews, W.H., Wilson, C.R., Poelma, P.L., Bullock, L.K., McClure, F.D. and Gentile, D.E. (1981) Interlaboratory evaluation of the AOAC method and the A-1 procedure for the recovery of fecal coliforms from foods. J. Assoc. Off. Anal. Chem. 64, 1116-1121.
Anon. (1987) Pharmacopoeia of culture media for food microbiology. Int. J. Food Microbiol. 5, 187-300.
A O A C (1980) Official methods of analysis, 13th edition. Association of Official Analytical Chemists, Arlington, VA.
AOAC (1990) Official methods of analysis, 14th edition. Association of Official Analytical Chemists, Arlington, VA.
APHA (1984) Compendium of methods for the microbiological examination of foods, 2nd edition. American Public Health Association, Washington, DC.
Bindschedler, O., deMan, J.C. and Curiat, G. (1981) Comparative study of several culture media to determine coliforms and E. coli in dairy and cocoa products. Zbl. Bakteriol. II. Abt. 136, 146-151.
Bredie, W.L. and de Boer, E. (1992) Evaluation of the MPN, Anderson-Baird-Parker, Petrifilm E. coli and Fluorocult ECD method for enumeration of Escherichia coli in foods of animal origin. Int. J. Food Microbiol. 16, 197-208.
Brenner, D.N. (1984) In: Bergey's Manual of Systematic Bacteriology, Vol. 1. Williams and Wilkins, Baltimore, MD, pp. 408-420.
Chapman, P.A., Siddons, C.A., Zadik, P.M. and Jewes, L. (1991) An improved selective medium for the isolation of Escherichia coli O157. J. Med. Microbiol. 35, 107-110.

183
Chen, H-C. and Wu, S-D. (1992) Agar medium for enumeration of fecal coliforms. J. Food Sci. 57, 1454-1457.
Conner, D.E. (1992) Temperature and NaC1 affect growth and survival of Escherichia coli O157:H7 in poultry-based and laboratory media. J. Food Sci. 57, 532-533.
Cooke, B.C. and Jorgenson, M. (1977) An evaluation of modified minerals glutamate medium for use in the presumptive coliform test on dairy products. N.Z.J. Dairy Sci. Technol. 12, 272-273.
Delaney, J.E., McCarthy, J.A. and Grasso, R.J. (1962) Measurement of Escherichia coli type 1 by the membrane filter. Water and Sewage Works 109, 289-294.
Doyle, M.P. and Schoeni, J.L. (1984) Survival and growth characteristics of Escherichia coli associated with hemorrhagic colitis. Appl. Environ. Microbiol. 48, 855-856.
Dunham, H.G. and Schoenlein, H.W. (1926) Brilliant green bile media. Stain Technol. 1, 129-134. Endo, S. (1904) Uber ein Verfahren zum Nachweis von Typhusbacillen. Centralbl. Bakt. I. Orig. 35,
109-110. Entis, P. (1984) Enumeration of total coliforms, fecal coliforms and Escherichia coli in foods by
hydrophobic grid membrane filter: collaborative study. J. Assoc. Anal. Chem. 67, 812-823. Entis, P. (1989) Hydrophobic grid membrane filter/MUG method for total coliform and Escherichia
coli enumeration in foods: collaborative study. J. Assoc. Anal. Chem. 72, 936-950. FDA. (1984) Bacteriological Analytical Manual, 6th edition. Food and Drugs Administration, Washing-
ton, DC. Feng, P.C.S. and Hartman, P.A. (1982) Fluorogenic assays for immediate confirmation of Escherichia
coli. Appl. Environ. Microbiol. 43, 1320-1329. Frampton, E.W. and Restaino, L. (1993) Methods for Escherichia coli identification in food, water and
clinical samples based on beta-glucuronidase detection. J. Appl. Bacteriol. 74, 223-233. Gassner, G. (1918) Ein neuer Dreifarbennahrboden zur Typhus-Ruhr-Diagnose. Centralbl. Bakt. I.
Orig. 80, 219-222. Gray, R.D. (1964) An improved formate lactose glutamate medium for the detection of Escherichia coli
and other coliform organisms in water. J. Hyg. 62, 495-508. Hahn, G. and Wittrock, E. (1991) Comparison of chromogenic and fluorogenic substances for differen-
tiation of coliforms and Escherichia coli in soft cheese. Acta Microbiol. Hung. 38, 265-271. Hajna, A.A. and Perry, C.A. (1943) Comparative study of presumptive and confirmative media for
bacteria of the coliform group and for fecal streptococci. Am. J. Publ. Health 33, 550-556. Hall, H.E. (1964) Methods of isolation and enumeration of coliform organisms. In: Examination of
Foods for Enteropathogenic and Indicator Bacteria; Review of methodology and manual of selective procedures, edited by K.H. Lewis and R. Angelotti. Public Health Serv. Publ. 1142, Division of Environmental Engineering and Food Protection, U.S. Department of Health Education and Welfare, Washington, DC.
Hall, L.P. (1984) A new direct plate method for the enumeration of Escherichia coli in frozen foods. J. Appl. Bacteriol. 56, 227-235.
Hartman, P.A., Hartman, P.S. and Lanz, W.W. (1975) Violet red bile 2 agar for stressed coliforms. Appl. Microbiol. 29, 537-539.
Hechelmann, H., Rossmanith, E., Periec, M. and Leistner, L. (1973) Untersuchung zur Ermittlung der Enterobacteriaceae zahl bei schlachtgelflugel. Fleischwirtschaft 53, 107-113.
Hill, W.E., Ferreira, J.L., Payne, W.L. and Jones V.M. (1985) Probability of recovering pathogenic Escherichia coli from foods. Appl. Environ. Microbiol. 49, 1374-1378.
Holbrook, R. and Anderson, J.M. (1982) The rapid enumeration of E. coli in foods by direct plating method. In: Isolation and Identification Methods for Food Poisoning Organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner. SAB Technical Series, 17. Academic Press, London, U.K.
Holbrook, R., Anderson, J.M. and Baird-Parker, A.C. (1980) Modified direct plate method for counting Escherichia coli in foods. Food Technol. Aust. 32, 78-83.
Holt-Harris, J.E. and Teague, O.A. (1916) A new culture medium for the isolation of Bacillus typhosus from stools. J. Infect. Dis. 18, 596-600.
ICMSF (1978) Microorganisms in foods I. Their significance and methods of enumeration, 2nd edition. University of Toronto Press.

184
ISO (1988) Meat and meat products - Enumeration of Escherichia coli - Colony count technique at 44~ using membranes. ISO 6391-1988, International Standards Organisation.
ISO (1991a) Microbiology - General guidance for the enumeration of coliforms - Most probable number technique. ISO 4831-1991, International Standards Organisation.
ISO (1991b) Microbiology - General guidance for the enumeration of coliforms - Colony count technique. ISO 4832-1991, International Standards Organisation.
ISO (1991c) Microbiology - General guidance for the detection of Enterobacteriaceae with pre-enrich- ment. ISO 8523-1991, International Standards Organisation.
ISO (1993a) Microbiology - General guidance for enumeration of presumptive Escherichia coli - Most probable number technique. ISO 7251-1993, International Standards Organisation.
ISO (1993b) Microbiology - General guidance for the enumeration of Enterobacteriaceae without resuscitation - MPN technique and colony count technique. ISO 7402-1993, International Standards Organisation.
Klein, H. and Fung, D.Y.C. (1976) Identification and quantification of fecal coliforms using violet red bile agar at elevated temperature. J. Milk Food Technol. 39, 768-770.
Koburger, J.A. and Miller, M.L. (1985) Evaluation of a fluorogenic MPN procedure for determining Escherichia coli in oysters. J. Food Protect. 48, 244-245.
MacConkey, A. (1905) Lactose fermenting bacteria in faeces. J. Hyg. Camb. 5, 333-379. MacConkey, A.T. (1908) Bile salt media and their advantages in some bacteriological examinations. J.
Hyg. Camb. 8, 322-334. Mailman, W.L. and Darby, C.W. (1941) Use of a lauryl sulphate tryptose broth for the detection of
coliform organisms. Am. J. Publ. Health 31, 127-134. March, S.B. and Ratnam, S. (1986) Sorbitol-MacConkey medium for detection of Escherichia coli
O157:H7 associated with hemorrhagic colitis. J. Clin. Microbiol. 23, 869-872. Marier, R., Wells, J.G., Swanson, R.C., Callahan, W. and Mehlman, I.J. (1973) An outbreak of
enteropathogenic Escherichia coli foodborne disease traced to imported French cheese. Lancet ii, 1376-1378.
Marshall, R.T., Hartman, P.A., Cannon, R.Y., Lambeth, L., Richardson, G.H., Spurgeon, K.R., Weddle, D.B., Wingfield, M. and White, C.H. (1978) Group comparative study of VRB-2 agar in the recovery of coliforms from raw milk, ice cream and cottage cheese. J. Food Prot. 41, 544-545.
Mehlman, I.J. (1984) Coliforms, fecal coliforms, Escherichia coli and enteropathogenic E. coli. In: Compendium of Methods for the Microbiological Examination of Foods, 2nd edition, edited by M.L.Speck. APHA, Washington, DC.
Mehlman, I.J. and Romero, A. (1982) Enteropathogenic Escherichia coli: methods for recovery from foods. Food Technol. 36, 73-79.
Moberg, L.J., Wagner, M.K. and Kellen, L.A. (1988) Fluorogenic assay for rapid detection of Escherichia coli in chilled and frozen foods: collaborative study. J. Assoc. Off. Anal. Chem. 71, 589-602.
Mossel, D.A.A. (1982) Marker (index and indicator) organisms in food and drinking water. Semantics, ecology, taxonomy and enumeration. Antonie van Leeuwenhoek 48, 609-611.
Mossel, D.A.A., Eelderink, I., Koopmans, M. and van Rossem, F. (1978) Optimalisation of a Mac- Conkey-type medium for the enumeration of Enterobacteriaceae. Lab. Pract. 27, 1049-1050.
Mossel, D.A.A., Eelderink, I., Koopmans, M. and van Rossem, F. (1979) Influence of carbon source, bile salts and incubation temperature on recovery of Enterobacteriaceae from foods using Mac- Conkey-type agars. J. Food Prot. 42, 470-475.
Mossel, D.A.A., Visser, M. and Cornelissen, A.M.R. (1963) The examination of foods for Enterobacte- riaceae using a test of the type generally adopted for the detection of salmonellae. J. Appl. Bacteriol. 26, 444-452.
Mossel, D.A.A., van der Zee, H., Hardon, A.P. and van Netten, P. (1986) The enumeration of thermotrophic types amongst the Enterobacteriaceae colonizing perishable foods. J. Appl. Bacteriol. 60, 289-295.
Motes, M.L., McPhearson, R.M. and DePaola, A. (1984) Comparison of three international methods with APHA method for enumeration of Escherichia coli in estuarine waters and shellfish. J. Food Prot. 47, 557-561.

185
Moussa, R.S., Keller, N., Curiat, G. and de Man, J.C. (1973) Comparison of five media for the isolation of coliform organisms from dehydrated and deep frozen foods. J. Appl. Bacteriol. 36, 619-629.
Murthy, T.R.K. and Bachhil, V.N. (1982) Comparison of pour plate with overlay and spread plating for Enterobacteriaceae count in minced meat. J. Food Sci. Technol. India 19, 37-39.
Oblinger, J.L., Kennedy, J.E. and Langston, D.M. (1982) Microflora recovered from foods on violet red bile agar with and without glucose and incubated at different temperatures. J. Food Prot. 45, 948-952.
Pierson, C.J., Emswiler, B.S. and Kotula, A.W. (1978) Comparison of methods for estimation of coliforms, fecal coliforms and enterococci in retail ground beef. J. Food Prot. 41, 263-266.
Raghubeer, E.V. and Matches, J.R. (1990) Temperature range for growth of Escherichia coli serotype O157:H7 and selected coliforms in E. coli medium. J. Clin. Microbiol. 28, 803-805.
Rappaport, F. and Henig, E. (1952) Media for the isolation and differentiation of pathogenic Esch. coli (serotypes O l l l and 055). J. Clin. Pathol. 5, 361-362.
Rayman, M.K., Jarvis, G.A., Davidson, C.M., Long, S., Allen, J.M., Tong, T., Dodsworth, P., McLaugh- lin, S., Greenberg, S., Shaw, B.G., Beckers, H.J., Qvist, S., Nottingham, P.M. and Stewart, B.J. (1979) ICMSF methods studies. XIII. An international comparative study of the MPN procedure and the Anderson-Baird-Parker direct plating method for the enumeration of Escherichia coli biotype I in raw meats. Can. J. Microbiol. 25, 1321-1327.
Reber, C.L. and Marshall, R.T. (1982) Comparison of VRB and VRB-2 agars for recovery of stressed coliforms from stored acidified half-and-half. J. Food Prot. 45, 584-585.
Rippey, S.R., Chandler, L.A. and Watkins, W.D. (1987) Fluorimetric method for enumeration of Escherichia coli in molluscan shellfish. J. Food Prot. 50, 685-690, 710.
Sakazaki, R., Namioka, S., Osada, A. and Yamada, C.A. (1960) A problem on the pathogenic role of Citrobacter of enteric bacteria. Jpn. J. Exp. Med. 30, 13-22.
Sakazaki, R., Tamura, K., Prescott, L.M., Benzic, Z., Sanyal, C. and Sinha, R. (1971) Bacteriological examination of diarrheal stools in Calcutta. Indian J. Med. Res. 59, 1025-1034.
Sharpe, A.N., Rayman, M.K., Burgener, D.M., Conley, D., Loit, A., Milling, M., Peterkin, P.I., Purvis, U. and Malcolm, S. (1983) Collaborative study of the MPN, Anderson-Baird-Parker direct plating, and hydrophobic grid-membrane filter methods for the enumeration of Escherichia coli biotype I in foods. Can. J. Microbiol., 29, 1247-1252.
Shelton, L.R., Leininger, H.V., Surkiewicz, B.F., Boer, E.F., Elliott, R.P., Hyndman, J.B. and Kramer, N. (1962) A bacteriological survey of the frozen pre-cooked food industry. U.S. Department of Health, Education and Welfare, Food and Drug Administration. Washington, DC.
Silliker, J.H., Gabis, D.A. and May, A. (1979) ICMSF methods studies. XI. Collaborative/comparative studies on determination of coliforms using the most probable number procedure. J. Food Prot. 42, 638-644.
Szabo, R.A., Todd, E.C.D. and Jean, A. (1986) Method to isolate Escherichia coli O157:H7 from food. J. Food Prot. 49, 768-772.
Todd, E.C.D., Szabo, R.A., Peterkin, P., Sharpe, A.N., Parrington, L., Bundle, D., Gidney, M.A.J. and Perry, M.B. (1988) Rapid hydrophobic grid membrane filter-enzyme-labeled antibody procedure for identification and enumeration of Escherichia coli O157 in foods. Appl. Environ. Microbiol. 54, 2536-2540.
Vaughn, R.H., Levine, M. and Smith, H.A. (1951) A buffered boric acid lactose medium for enrichment and presumptive identification of Escherichia coli. Food Res. 16, 10-19.
Vracko, R. and Sherris, J.C. (1963) Indole spot test in bacteriology. Am. J. Clin. Pathol. 39, 429-432. Weiss, K.F., Chopra, N., Stotland, P., Riedel, G.W. and Malcolm, S. (1983) Recovery of fecal coliforms
and of Escherichia coli at 44.5, 45.0 and 45.5~ J. Food Prot. 46, 172-177. West, P.A. and Coleman, M.R. (1986) A tentative national reference procedure for isolation and
enumeration of Escherichia coli from bivalve molluscan shellfish by most probable number method. J. Appl. Bacteriol. 61, 505-516.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
187
Chapter 11 Media for salmonella
M a r t i n Busse
Institut fiir Mikrobiologie, Forschungszentrum fiir Lebensmittel und Milch Weihenstephan, D-8050 Freising, Germany
Abstract
The main categories of selective plating media are reviewed. The selective enrichment media are considered to be of critical importance in salmonella isolation. During the last 10 years Rappaport-Vassiliadis broth has emerged as a new reliable enrichment medium. Likewise motility enrichment is a very promising technique, but will no doubt have its limitations. However, since it is based on a different principle compared with traditional enrichment, it may complement traditional techniques in a useful way. Most important, however, is the insight that the selectivity of a selective procedure is not solely defined by the medium and by incubation conditions. The food sample and its microflora apparently influences the result considerably. This finding may open a new and fascinating field of scientific research; the ecology of selective media.
The process of isolating salmonellas from foods is to some extent prone to failure. The main risk is probably in sampling, but even if a contaminated portion has been drawn, salmonellas can be lost during enrichment. If salmonella domi- nates the flora of the enrichment broth it will be found in any event. However, if salmonella forms only a minority among competitors, successful detection may well be a mat ter of chance. In this case plating media may become important. Depend- ing on the types of competitive bacteria, detection of occasional colonies of salmonella may be easier if the appropriate plating medium has been used. Unfortunately, the composition of the flora is never known in advance. Therefore the appropriate plating medium will be unknown too.
Successful enrichment is the decisive step in the detection of salmonellas, and great efforts have been made in recent years to improve media and procedures for enrichment, which will be the main topic of this review. A short survey will also be made of the most important plating media currently used. Finally, the effect of competitors on the detection of salmonellas will be discussed.
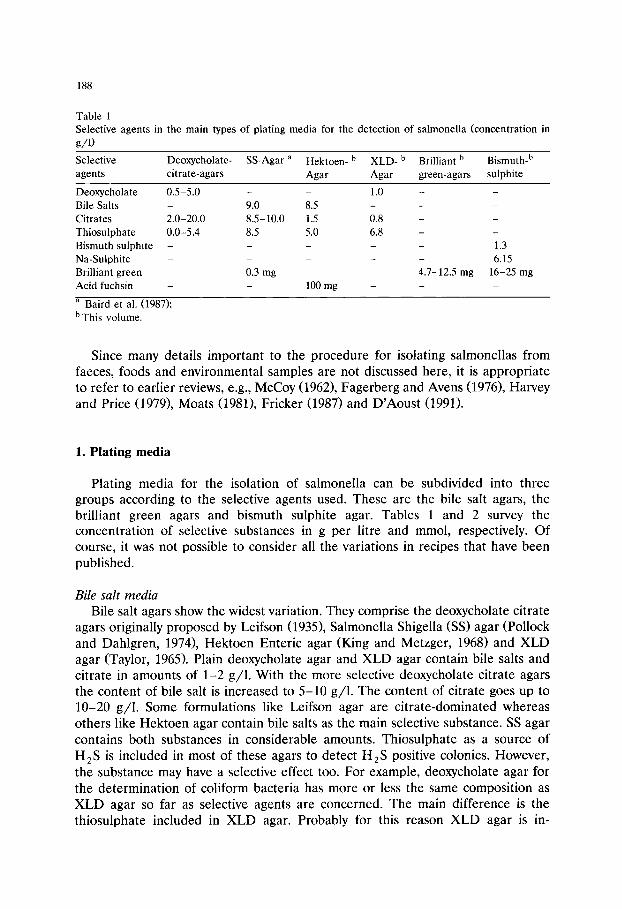
188
T a b l e 1
S e l e c t i v e a g e n t s in t h e m a i n t y p e s o f p l a t i n g m e d i a fo r t h e d e t e c t i o n o f s a l m o n e l l a ( c o n c e n t r a t i o n in
g/l)
S e l e c t i v e D e o x y c h o l a t e - S S - A g a r a H e k t o e n - b X L D - b Br i l l i an t b B i smuth_b
a g e n t s c i t r a t e - a g a r s A g a r A g a r g r e e n - a g a r s s u l p h i t e
D e o x y c h o l a t e 0 . 5 - 5 . 0 - - 1.0 - -
Bi le Sa l t s - 9.0 8.5 - - -
C i t r a t e s 2 . 0 - 2 0 . 0 8 . 5 - 1 0 . 0 1.5 0.8 - -
T h i o s u l p h a t e 0 . 0 - 5 . 4 8.5 5.0 6.8 - -
B i s m u t h s u l p h i t e . . . . . 1.3
N a - S u l p h i t e . . . . . 6.15
B r i l l i a n t g r e e n - 0.3 m g - - 4 . 7 - 1 2 . 5 m g 1 6 - 2 5 m g
A c i d f u c h s i n - - 100 m g - - -
a B a i r d e t al. (1987);
b T h i s v o l u m e .
Since many details important to the procedure for isolating salmonellas from faeces, foods and environmental samples are not discussed here, it is appropriate to refer to earlier reviews, e.g., McCoy (1962), Fagerberg and Avens (1976), Harvey and Price (1979), Moats (1981), Fricker (1987) and D'Aoust (1991).
1. Plating media
Plating media for the isolation of salmonella can be subdivided into three groups according to the selective agents used. These are the bile salt agars, the brilliant green agars and bismuth sulphite agar. Tables 1 and 2 survey the concentration of selective substances in g per litre and mmol, respectively. Of course, it was not possible to consider all the variations in recipes that have been published.
Bile salt media Bile salt agars show the widest variation. They comprise the deoxycholate citrate
agars originally proposed by Leifson (1935), Salmonella Shigella (SS) agar (Pollock and Dahlgren, 1974), Hektoen Enteric agar (King and Metzger, 1968) and XLD agar (Taylor, 1965). Plain deoxycholate agar and XLD agar contain bile salts and citrate in amounts of 1-2 g/1. With the more selective deoxycholate citrate agars the content of bile salt is increased to 5-10 g/1. The content of citrate goes up to 10-20 g/1. Some formulations like Leifson agar are citrate-dominated whereas others like Hektoen agar contain bile salts as the main selective substance. SS agar contains both substances in considerable amounts. Thiosulphate as a source of He S is included in most of these agars to detect H eS positive colonies. However, the substance may have a selective effect too. For example, deoxycholate agar for the determination of coliform bacteria has more or less the same composition as XLD agar so far as selective agents are concerned. The main difference is the thiosulphate included in XLD agar. Probably for this reason XLD agar is in-
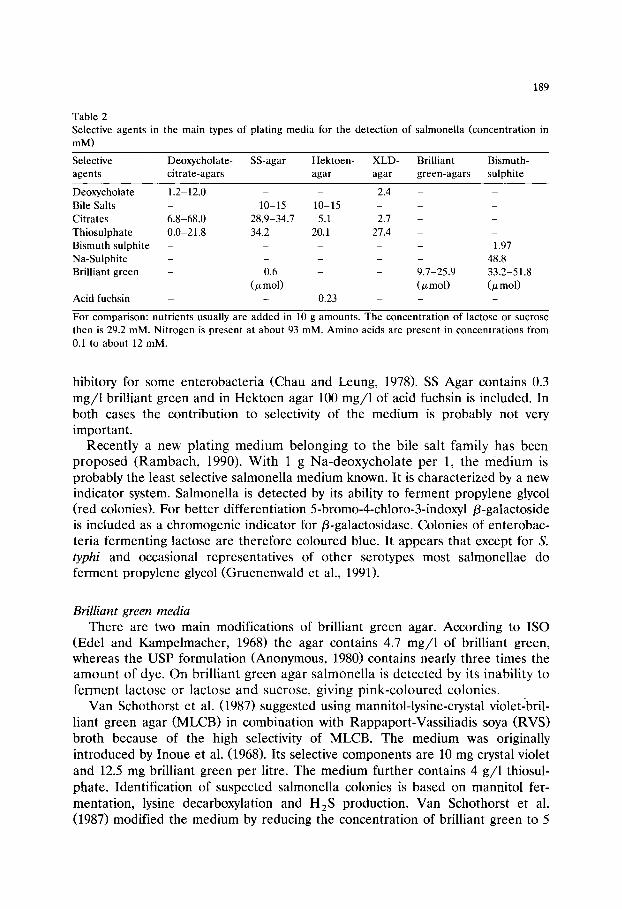
189
Table 2 Selective agents in the main types of plating media for the detection of salmonella (concentration in raM)
Se lec t ive Deoxychola te - SS-agar Hektoen- X L D - Brilliant Bismuth- agents citrate-agars agar agar green-agars sulphite
Deoxycholate 1.2-12.0 - - 2.4 - - Bile Salts - 10-15 10-15 - - - Citrates 6.8-68.0 28.9-34.7 5.1 2.7 - - Thiosulphate 0.0-21.8 34.2 20.1 27.4 - - Bismuth sulphite . . . . . 1.97 Na-Sulphite . . . . . 48.8 Brilliant green - 0.6 - - 9.7-25.9 33.2-51.8
(/xmol) (/~mol) (~mol) Acid fuchsin - - 0.23 - - -
For comparison: nutrients usually are added in 10 g amounts. The concentration of lactose or sucrose then is 29.2 mM. Nitrogen is present at about 93 raM. Amino acids are present in concentrations from 0.1 to about 12 mM.
hibitory for some en te robac te r ia (Chau and Leung, 1978). SS Agar contains 0.3 mg /1 brilliant green and in H e k t o e n agar 100 mg /1 of acid fuchsin is included. In both cases the contr ibut ion to selectivity of the m e d i u m is probably not very important .
Recen t ly a new p la t ing m e d i u m be long ing to the bile salt family has been p r o p o s e d ( R a m b a c h , 1990). W i t h 1 g N a - d e o x y c h o l a t e per 1, the m e d i u m is probably the least selective salmonel la m e d i u m known. It is charac ter ized by a new indicator system. Salmonel la is de tec ted by its ability to fe rment propylene glycol (red colonies). For be t te r different ia t ion 5-bromo-4-chloro-3-indoxyl /3-galactoside is included as a chromogenic indicator for /3-galactosidase. Colonies of enterobac- teria fe rment ing lactose are there fore coloured blue. It appears that except for S. typhi and occasional representa t ives of o ther serotypes most sa lmonel lae do fe rment propylene glycol (Gruenenwald et al., 1991).
Brilliant green media There are two main modificat ions of brilliant green agar. According to ISO
(Edel and Kampe lmacher , 1968) the agar contains 4.7 mg /1 of brill iant green, whereas the USP formulat ion (Anonymous, 1980) contains nearly three t imes the a m o u n t of dye. On br i l l iant green aga r s a lmone l l a is de tec ted by its inabi l i ty to f e rmen t lac tose or lac tose and sucrose, giving p i n k - c o l o u r e d colonies .
Van Schothorst et al. (1987) suggested using mannitol-lysine-crystal violet-bril- liant green agar (MLCB) in combinat ion with Rappapor t -Vass i l iadis soya (RVS) broth because of the high selectivity of MLCB. The m e d i u m was originally in t roduced by Inoue et al. (1968). Its selective componen ts are 10 mg crystal violet and 12.5 mg brilliant green per litre. The m e d i u m fur ther contains 4 g/1 thiosul-
phate . Ident if icat ion of suspected salmonel la colonies is based on manni tol fer- menta t ion , lysine decarboxylat ion and H zS product ion. Van Schothorst et al. (1987) modif ied the m e d i u m by reducing the concent ra t ion of brilliant green to 5

190
mg/1. Using pure strains the authors showed that MLCB is more selective than brilliant green agar. Salmonella strains grew well, whereas certain competitive enterobacteria which still grow on brilliant green agar, did not develop on MLCB. Van Schothorst et al. (1987) found that productivity of MLCB greatly depends on the quality of agar used.
Bismuth sulphite agar Bismuth sulphite agar is usually said to be particularly useful for detecting S.
typhi. Interestingly, it is very difficult to get clear information on the exact composition with respect to bismuth. According to Leininger (1976) the medium contains 1.85 g/1 of ammonium bismuth citrate and 6.15 g/1 sodium sulphite. This corresponds to the 8 g/1 of 'bismuth sulphite indicator' usually found in the handbooks of medium manufacturers. If Leininger is correct then about 4 mM bismuth (or 1.97 mM bismuth sulphite) and close to 50 mM sulphite are added to the medium. In Table 1 therefore the bismuth content of the medium is indicated in terms of bismuth sulphite and additional NazSO 3. Even that may be misleading since during preparation of the medium a precipitate is formed which probably contains part of the bismuth. As far as the surplus sulphite is concerned, an unknown amount will be oxidized during the preparation of the medium. Accord- ing to McCoy (1962) 3 g of the ammonium bismuth citrate is added per litre which would be 1.6 times the amount of bismuth. Finally, it should be noted that bismuth sulphite agar is a brilliant green agar par excellence. Even if the lower amount of 16 mg/1 is considered (Table 4), the selectivity conferred by the dye is definitely higher than that of the brilliant green agars. It is puzzling why bismuth sulphite agar is considered to select for S. typhi, when this serotype is said not to grow on brilliant green agar. A specific trait of bismuth sulphite agar is that on this medium salmonella is detected by H zS-formation and bismuth reduction only.
In Table 2 the concentration of selective agents is given in mM or IzM. In some cases, for example with bile salt mixtures or ill defined iron citrates, the molar concentrations given are at best plausible guesses. Bile salts, citrates and thiosul- phate are used at concentrations of 10 to about 30 mM which is equivalent to nutrient levels. For example with lactose this would correspond to 3-10 g/1. By contrast, brilliant green acts as a true poison and is used at the tzM level. The same applies to malachite green (Table 4). Fuchsin seems to be less poisonous. For example, Endo agar contains 400 mg/1, which corresponds to 1 mM of basic fuchsin. Bismuth sulphite too is added at this level. Undoubtedly, it is more informative if the composition of selective media is expressed on a molar basis.
Finally, the following remark should be made. Originally plating media for salmonella were devised mainly for direct plating of faeces. The 'old' media like the deoxycholate citrate agar of Leifson (1935) or the bismuth sulphite agar of Wilson and Blair (1927) had to be rather selective. Today, detecting salmonella generally includes selective enrichment. Under those conditions, selectivity of plating does not seem to be of major importance. Of course whatever enrichment procedure is used, Gram-negative competitors are always encountered at varying rates. However, there is no indication that this can be overcome by the selectivity

Table 3 Selective agents in the main media for enrichment of salmonella (concentrations in g/l)
191
Selective agent Selenite Selenite Tetrathionate a Rappaport RV a Rappaport broth brilliant broth (original) semisolid a Leifson green USP ISO
NaHSeO 3 4.0 4.0 . . . . . Na-Thiosulphate - - 40.7 30.0 - - - MgClz-6H20 . . . . 28.6 28.6-36.0 17.25-23.34 Malachite
green oxalate . . . . 108 mg 36 mg 37-65 mg Brilliant green - 5 mg (10 mg) 10 mg - - - Na-Taurocholate - 1.0 . . . . . Bile, bile salts - - 4.75 1.0 - - -
The USP formulation of tetrathionate broth does not necessarily contain brilliant green. a See monographs in this volume.
of plat ing media . Thus, plat ing med ia for salmonel las become increasingly less selective. On the o the r hand, much effort has been made to improve the indicator systems for ident if icat ion of sa lmonel la colonies. Of course, some authors still feel that selectivity of plat ing media is an impor tan t fea ture (D 'Aous t 1991).
2. Selective enrichment media
Media conta in ing e i ther seleni te or t e t r a th iona te plus bril l iant g reen and bile or else malachi te green in combina t ion with high amount s of magnes ium chlor ide are at p resen t the p r e d o m i n a n t tools for selective enr ichment . The malachi te green containing R a p p a p o r t med ia have been deve loped during the last 25 years. In cer ta in par ts of the world there is a s t rong tendency to replace t e t r a th iona te b ro th with modif ied R a p p a p o r t broth. A n o t h e r m o d e r n deve lopmen t is the t echn ique of motili ty enr ichment .
Seleni te and t e t r a th iona te bro ths have been extensively discussed in ear l ier reviews. We the re fo re concen t ra te on the more recent work compar ing R a p p a p o r t b ro th with the t radi t ional media .
General remarks on composition Tables 3 and 4 summar ize the concent ra t ions of the inhibitors in the commonly-
used en r i chmen t media in g /1 and mM, respectively. As m e n t i o n e d already, th ree
groups of med ia can be dist inguished. These are seleni te b ro th (Leifson, 1936), t e t r a th iona te b ro th (Muller , 1923; Kauffmann, 1930) and the R a p p a p o r t family. Table 3 shows that selective agents are used in ra the r high amounts . For example,
t e t r a th iona te b ro th is fortif ied with 10 mg bril l iant green, which is close to the uppe r limit in bril l iant g reen agars. Also the amounts of th iosu lphate in this m e d i u m are very high. Of course, th iosu lpha te is oxidized by iodine to te t ra th ion-

192
Table 4 Selective agents in the main media for enrichment of salmonella (concentration in mM)
Selective agent Selenite Selenite Tetrathionate Rappaport RV broth brilliant broth (original) Leifson green USP ISO
Rappaport semisolid
NaHSeO 3 26.5 26.5 . . . . . Thiosulphate (as thionate) - - 82.0 60.4 - - - MgCI2.6H20 . . . . 141 141-177 84.8-114.8 Malachite green oxalate . . . . 123 a 41.1 a 42.2-74.2 Brilliant green - 10.4 a (20.7) a 20.7 a _ _ _
Na-Taurocholate - 2.3 . . . . . Bile, bile salts - - 5-7 1.0-1.3 - - -
a Brilliant green and malachite green (/~M).
ate which will reduce the molar i ty by half. However , the concent ra t ion of te- t ra th ionate , amount ing to 60-80 mM, is still considerable (Table 4). The molar i- ty of selenite in selenite broth , again is within the nutr ient range. Selectivity of Rappapor t media depends on malachite green, a relatively high ionic strength and a low pH. The Rappaport-Vassiliadis medium contains 41 /xM of malachite green and 141-177 mM MgC12 (Table 4). The pH is 5.2. Some authors seem to assume that the relatively high content of MgC12 leads to hypertonic conditions or to a 'high' osmotic pressure (Harvey and Price, 1978; Peterz et al., 1989). It should be mentioned, however, that with MgCI 2 a reduction of the a w value to 0.95 requires a concentration of 840 mM. That is 4.7 times the concentration used in the Rappapor t media.
Rappaport- Vassiliadis medium (R V) The original Rappapor t medium (Rappaport et al., 1956) contains rather high
amounts of malachite green and magnesium chloride (Table 3). In addition the medium has a very low pH of 5.2. The medium was developed for the enrichment of S. paratyphi and other serotypes known to be relatively resistant to brilliant green. MgC12 was shown to counteract the toxic effect of the dye for salmonella (Rappapor t and Konforti, 1959). Vassiliadis et al. (1976) modified the medium by reducing the concentration of the dye to one third. At 41/xM, the concentration is still equivalent to the level of brilliant green used in bismuth sulphite agar. In the semisolid modifications of Rappapor t medium the concentrations of selective agents have been lowered further (see the m o n o g r a p h s in this volume).
Rappaport-Vassiliadis medium (Vassiliadis et al., 1976) has been generally accepted as a useful tool beside selenite and tetrathionate broths. At present in many countries the use of selenite in combination with RV is preferred as a standard method. The reasons for this have been summarized by Beckers et al. (1987a). Tetrathionate (MK) has been compared with RV in several studies. Vassiliadis (1983) summarized the results obtained in his department since 1976. Out of 2000 samples of meat, meat products, pig faeces and sewage 17% were found positive with MK and 25% with RV. Van Schothorst and Renaud (1983)

Table 5 Comparison of tetrathionate broth (MK) and Rappaport-Vassiliadis broth (RV)
193
Authors and types of sample Number of Percentage positive samples MK RV
Vassiliadis (1983) meat, pig faeces sewage
Pietzsch (1984) pig faeces chicken skin, minced meat
Beckers et al. (1986 a) minced meat, spices, dried vegetables, poultry, egg products, mayonnaise
2109 17 25
701 48 92
590 47 63
tested environmental samples from food factories. With MK about 40% of the positive samples went undetected. Similar results were obtained by Pietzsch (1984). He investigated 701 samples of pig faeces, chicken skin and minced pork. Of 324 positive samples 48% were found with MK and 92% with RV. With meat products Tongpim et al. (1984) found that RV gave more positive results with artificially contaminated samples. Some of these results are summarized in Table 5. In a series of papers Beckers at al. (1986 b,c) showed that differences be tween the media were predominant ly due to the fact that the prepara t ion of MK medium was a critical factor. Table 6 shows the results obtained in one of these studies. Apparent ly it is much easier to obtain reproducible results with RV medium. Northol t et al. (1985) came to the same conclusion. Testing artificially contami- nated caseinate or milk powder, MK medium prepared under everyday routine conditions gave far fewer positives compared with RV. If, however, MK was prepared very carefully both media proved to be equally effective.
The method used by Rappapor t et al. (1956) and also by Vassiliadis et al. (1976) to prepare Rappapor t media caused confusion with respect to the concentrat ion of MgC12. According to the original recipe a MgC12 stock solution is p repared by adding 400 g of MgC12 �9 6 H 2 0 to 1000 ml of water. Of this stock solution 100 ml
Table 6 Productivity of enrichment according to the method of medium preparation: comparison of tetrathion- ate brilliant green bile (MK) and Rappaport-Vassiliadis (RV)
Seletive enrichment medium Percentage positive
(1) Series (2) Series
MK distributed by organizing lab. MK laboratory made (Oxoid) MK laboratory made (Merck)
RV distributed by organizing lab. RV laboratory made (from ingredients) RV laboratory made (Oxoid)
98.5 98.4 68.5 69.5
98.5 98.4 - 96.3 - 91.2
Collaborative study in 20 laboratories. Milk powder contaminated with salmonella reference sample. Data from Beckers et al. (1986c).

194
are used to prepare the medium. The final volume of the medium is 1110 ml. As Peterz et al. (1989) pointed out, the stock solution has a volume of 1260 ml and therefore it will contain 31.7 g MgC12 �9 6 H 2 0 per 100 ml. The final medium will therefore contain 28.6 g / l MgC12 �9 6 H20. If, however, the concentration per litre is calculated without considering the increase in volume of the stock solution a value of 36 g MgC12. 6 H 2 0 is obtained. Some manufacturers use anhydrous MgC12. The correct concentration then is 13.4 g/1 (or 13.58 g/1 if the hexahydrate concentration is rounded up to 29 g/l). The equivalent of the higher concentration will be 17 g/1. All these values will be found in the literature and in the handbooks of medium manufacturers. However, there is increasing awareness that the correct amount is 29 g of the hexahydrate of MgC12 or 13.58 g of the anhydrous salt per litre. This is equivalent to 143 mM (Table 4). Finally it should be pointed out that van Schothorst et al. (1987) tested various MgC12 concentrations in the range of 29 to 36 g/1 hexahydrate. The results clearly showed that 36 g/1 is too high a concentration.
Van Schothorst and Renaud (1983) reported that using soya peptone instead of tryptone improves recovery rates of salmonella. Similar results have been obtained by McGibbon et al. (1984), Fricker et al. (1985), Fricker and Girdwood (1985) and Quail et al. (1986). The effect observed in Fricker's laboratory, however, was slight.
A specific trait of all malachite green/MgC12 media seems to be that better results are obtained if the selective broth is inoculated from the pre-enrichment solution not at a ratio of 1:10 but at higher ratio, e.g. 1:100. This effect was demonstrated by Rappaport et al. (1956) and by Harvey and Price (1980)with the original formulation of this medium. Testing modified RV, Tongpim et al. (1984) made similar observations. This effect might be predicted because inoculating fewer bacteria into the selective medium means that more divisions will occur before multiplication ceases. As a consequence, small differences in generation times of competing bacteria may have a more pronounced effect (Jameson, 1961). Interestingly, raising the ratio of inoculum to tetrathionate medium had no effect (Tongpim et al., 1984). In tetrathionate broth differences in the lag phases of competitors are possibly more important. It should be pointed out, however, that the situation is quite unclear. For example, Kafel and Pogorzelska (1987)working with selenite and tetrathionate broth also found pronounced effects of transfer volumes for these media. Recovery of salmonella from minced meat was optimal if the pre-enrichment culture was diluted by a factor of 102-104. Obviously, such discrepancies illustrate how little is known about the black box called enrichment (Beckers et al., 1987b).
Vassiliadis et al. (1976) recommended incubation of RV at 43~ This achieved maximum selectivity, but any deviation above 43~ may be lethal for salmonella. For this reason most workers follow the ISO recommendation to incubate at 42~ (Anon, 1990; ISO 6579: 1990).
Motility enrichment on semisolid media Motility enrichment is an old technique (Craigie, 1931). Originally it was
performed in U-tubes where the one side was inoculated and after appropriate

195
incubation fast moving bacteria could be isolated from the other side. Stuart and Pivnick (1965) for example tried to isolate salmonella from faeces by this tech- nique. Other authors using the procedure were Banwart (1968), Harper and Shortridge (1969) and Fung and Kraft (1970). Chau and Huang (1974; 1976) tried to optimize the method by testing the effect of pH, ionic strength and triphenyl- methane dyes on motility of enterobacteria. A more modern application of the method was introduced by Goossens et al. (1984) who replaced U-tubes by Petri dishes. They used a modified Rappaport medium (modified semisolid Rappaport: MSR) containing 3.2 g agar, 65 mg malachite green oxalate and 17.25 g MgC12 per litre (Table 3). The nutrients were reduced to 60% of the Rappaport formulation. A similar medium was proposed by De Smedt et al. (1986) containing 2.7 g agar, 37 mg of malachite green oxalate and 23.3 g MgC12 (Table 3). The nutrient content was also reduced. As a semisolid modification of RV, the agar is called MSRV (modified semisolid Rappaport-Vassiliadis). The more selective MSR is incubated at 35-37~ wheras for MSRV 43~ was proposed (reduced tot 42~ this volume). Like the whole Rappapor t family these media do not contain sugars.
The technique is rather simple. A drop of the pre-enrichment solution or of selective enrichment broth is spotted on the semisolid medium. Motile bacteria will spread radially and form a large colony. Provided the motility of the competi- tive bacteria is sufficiently reduced, salmonella will be the dominating organism at the edge of the colony. In model experiments using a limited number of pure cultures De Smedt and Bolderdijk (1987) demonstrated that small numbers of salmonella (60/ml) can be recovered even if outnumbered by competitive bacteria by a factor of 10 v. In a collaborative study (De Smedt and Bolderdijk, 1990) artificially contaminated cocoa powder and milk chocolate was tested comparing the semisolid Rappaport-Vassiliadis (MSRV) medium with the traditional RV enrichment procedure. Differences in favour of MSRV were highly significant. This effect, however, was due to the fact that three out of 11 salmonella strains used were lactose positive. Some of the laboratories had difficulties in recognizing lactose positive strains of salmonella with the traditional procedure. With motility enrichment these strains could be easily detected. When lac+ strains were ex- cluded from the statistical evaluation no significant differences in recovery rates were observed. In another collaborative study (De Zutter et al., 1991) RV and MSRV were applied to a wide variety of foods. Four trials were performed. In the first trial pre-enrichment broth was inoculated with a salmonella reference sample, a gelatin capsule containing five cells of S. typhimurium II 505 (Beckers et al., 1985). In the second trial the salmonella reference sample was combined with a reference sample of competitive bacteria (ca. 104 cells per sample). In a third trial foods were contaminated with the salmonella reference sample and in the last series naturally contaminated food was tested. Without food material added, the reference salmonella was easily recovered by both methods. Adding competitive bacteria had no significant effect. Adding food to the pre-enrichment generally lowered the recovery rates. The effect was a little more pronounced with RV. The same result was obtained with naturally contaminated foods. MSRV detected 96% of all contaminated samples, whereas with RV 90% of these samples were found.

196
None of the differences proved to be statistically significant. Apparently motility enrichment is a reliable technique. Finally it should be mentioned that Perales and Erkiaga (1991) compared both semi-solid media for motility enrichment. A total of 154 samples of high- and low-moisture foods and feeds were tested. Motility enrichment was applied to the pre-enrichment solution and to selective enrichment in selenite and tetrathionate. In this study MSRV proved to be very inferior, detecting only 15 out of 62 positive samples. With MSR Salmonella was recovered from 60 samples. Further, plating out of selenite and tetrathionate enrichments in the traditional manner detected only 19 and 18 positive samples, respectively. Again, research on the methods of salmonella detection does not yield a consistent picture. Procedures which according to Perales and Erkiaga are rather inferior in other laboratories were shown to be quite reliable.
The principle behind motility enrichment clearly differs from that of traditional enrichment. In motility enrichment the motility of salmonellas is thought to be less impaired by the selective medium than the motility of competitive bacteria. In contrast, with traditional enrichment it is the growth rate which is influenced by enrichment conditions. It might be useful to include motility enrichment as a plating medium in the standard procedure. Of course, experience will show the extent to which strains of salmonella too are susceptible to motility inhibition by malachite green. It also remains to be seen how frequently competitors will be encountered that are highly motile on semisolid Rappaport media. Non-motile salmonellas, of course, will be found by traditional plating procedures only.
According to De Smedt and co-workers (1986, 1987) motility enrichment should be considered as a 'rapid' method. This refers to the fact that a high proportion of all positive samples is found by plating pre-enrichment cultures. 'Rapid methods' currently do have a considerable scientific appeal. In addition, for some applica- tions fast tests, if reliable enough, will be important. It should be borne in mind, however, that at least in food production, quality assurance is not so much impeded by lack of rapidity but rather by heterogeneous distribution of pathogens, if present. The number of samples which should be examined to draw safe conclusions is most important, rather than the speed. It is true, however, that motility enrichment has the advantage of being relatively cost efficient.
3. Enrichment and competitive bacteria
Selective enrichment media are usually supposed to suppress the accompanying flora. As far as Gram-positive bacteria are concerned, selective media for salmonella are quite efficient. There are, however, truly competitive bacteria. Everybody knows that streaks consisting solely or predominantly of salmonella are satisfying but rare. Complete suppression of all unwanted organisms is sort of a mystic dream of microbiologists and, of course, an empty phrase in the handbooks of media producers. In the early days of salmonella research, scientists found that upon continued incubation a succession of dominant species occurred in broth cultures of faeces. It was hoped to find conditions under which salmonella would
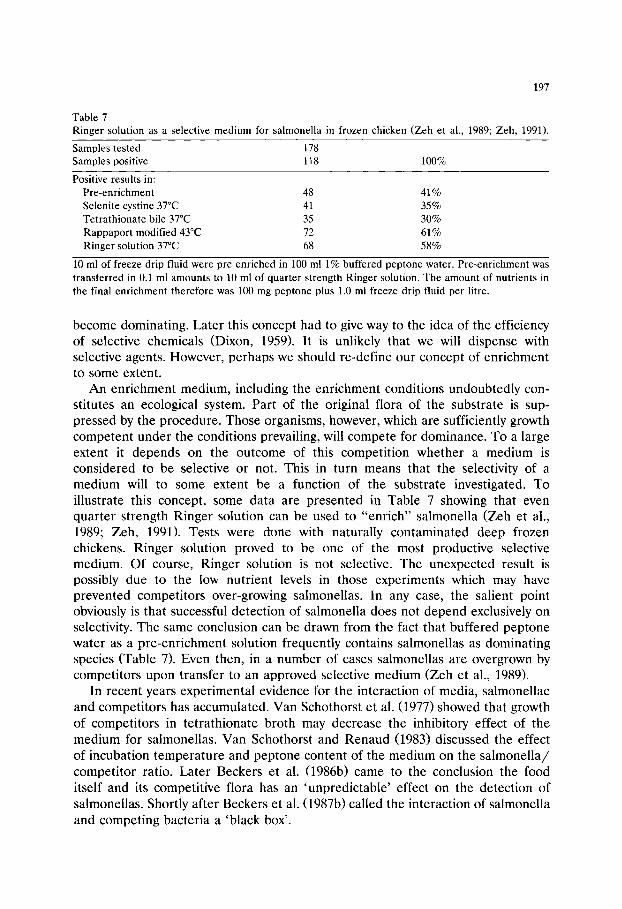
197
Table 7 Ringer solution as a selective medium for salmonella in frozen chicken (Zeh et al., 1989; Zeh, 1991).
Samples tested 178 Samples positive 118 100%
Positive results in: Pre-enrichment 48 41% Selenite cystine 37~ 41 35% Tetrathionate bile 37~ 35 30% Rappaport modified 43~ 72 61% Ringer solution 37~ 68 58%
10 ml of freeze drip fluid were pre-enriched in 100 ml 1% buffered peptone water. Pre-enrichment was transferred in 0.1 ml amounts to 10 ml of quarter strength Ringer solution. The amount of nutrients in the final enrichment therefore was 100 mg peptone plus 1.0 ml freeze drip fluid per litre.
become dominating. Later this concept had to give way to the idea of the efficiency of selective chemicals (Dixon, 1959). It is unlikely that we will dispense with selective agents. However, perhaps we should re-define our concept of enrichment to some extent.
An enrichment medium, including the enrichment conditions undoubtedly con- stitutes an ecological system. Part of the original flora of the substrate is sup- pressed by the procedure. Those organisms, however, which are sufficiently growth competent under the conditions prevailing, will compete for dominance. To a large extent it depends on the outcome of this competition whether a medium is considered to be selective or not. This in turn means that the selectivity of a medium will to some extent be a function of the substrate investigated. To illustrate this concept, some data are presented in Table 7 showing that even quarter strength Ringer solution can be used to "enrich" salmonella (Zeh et al., 1989; Zeh, 1991). Tests were done with naturally contaminated deep frozen chickens. Ringer solution proved to be one of the most productive selective medium. Of course, Ringer solution is not selective. The unexpected result is possibly due to the low nutrient levels in those experiments which may have prevented competitors over-growing salmonellas. In any case, the salient point obviously is that successful detection of salmonella does not depend exclusively on selectivity. The same conclusion can be drawn from the fact that buffered peptone water as a pre-enrichment solution frequently contains salmonellas as dominating species (Table 7). Even then, in a number of cases salmonellas are overgrown by competitors upon transfer to an approved selective medium (Zeh et al., 1989).
In recent years experimental evidence for the interaction of media, salmonellae and competitors has accumulated. Van Schothorst et al. (1977) showed that growth of competitors in tetrathionate broth may decrease the inhibitory effect of the medium for salmonellas. Van Schothorst and Renaud (1983) discussed the effect of incubation temperature and peptone content of the medium on the salmonella/ competitor ratio. Later Beckers et al. (1986b) came to the conclusion the food itself and its competitive flora has an 'unpredictable' effect on the detection of salmonellas. Shortly after Beckers et al. (1987b) called the interaction of salmonella and competing bacteria a 'black box'.

198
It is quest ionable, therefore , whe ther it is appropr ia te to discuss the problems of enr ichment solely from the aspect of 'selectivity' and 'productivity' . Usually the wanted organism has to compete with other bacteria and this may affect the final result considerably. To cope with this complication, research on the competi t ive ability of salmonellae as well as typical competi tors might be useful. The idea of
Beckers et al. (1985), to evaluate methods of enr ichment with the help of a
salmonella reference sample, may realise its full impact if the reference sample too
is carefully defined. The same probably applies to any effort to install a system of
quality assurance for selective enr ichment media (Mackey, 1985). The ideas put forward here must not be misunders tood as a resigned at t i tude
towards the problem of salmonella detection. On the contrary, the fact that selective media do have an ecology and that compet i t ion is an impor tant factor in enr ichment , yields a rational explanat ion for " the vast and often bewildering amount of information on isolation me thods" which, as Fricker (1987) put it, "is daunt ing" . Fur the r research on the ecology of enr ichment media, may very well provide a more appropr ia te unders tand ing of the process of enr ichment .
References
Anon. (1980) United States Pharmacopeia XX. Microbiological limit tests. Mack Publishing Co., Easton.
Anon. (1990) The Oxoid Manual, 6th Edn., Unipath, Basingstoke, England. Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (Eds.) (1987) Pharmacoepeia of Culture Media for Food
Microbiology. Int. J. Food Microbiol. 5, 187-300. Baird, R.M., Corry, J.E.L., Curtis, G.D.W., Mossel, D.A.A. and Skovgaard, N. (Eds.) (1989) Pharma-
coepeia of Culture Media, Additional Monographs. Int. J. Food. Microbiol. 9, 85-144. Banwart, G.J. (1968) Glassware apparatus for determining motile bacteria. I. Salmonella. Poult. Sci. 47,
1209-1212. Beckers, H.J., van Leusden, F.M., Meijssen, M.J.M. and Kampelmacher, E.H. (1985) Reference
material for the evaluation of a standard method for the detection of Salmonella in foods and feedstuffs. J. Appl. Bacteriol. 59, 507-512.
Beckers, H.J., van Leusden, F.M., Roberts, D. Pietzsch, O. Price, T.H., van Schothorst, M, Beumer, R.R., Peters, R. and Kampelmacher, E.H., (1986a) Evaluation of reference material for the detection of Salmonella. Int. J. Food Microbiol. 3, 287-298.
Beckers, H.J., van Leusden, F.M. and Peters, R. (1986b) Comparison of Muller-Kauffmann's te- trathionate broth and modified Rappaport's medium for isolation of Salmonella. J. Food Safety 8, 1-9.
Beckers, H.J. Peters, R. and Pateer, P.M. (1986c) Collaborative study on the isolation of Salmonella from reference material using selective enrichment media prepared from individual ingredients and from commercial dehydrated media. Int. J. Food Microbiol. 4, 1-11.
Beckers, H.J., Roberts, D., Pietzsch, O., van Schothorst, M., Vassiliadis, P. and Kampelmacher, E.H. (1987a) Replacement of Muller-Kauffmann's tetrathionate brilliant green bile broth by Rappaport- Vassiliadis's magnesium malachite green broth in the standard method for the detection of salmonellas. J. Food Microbiol. 4, 59-64.
Beckers, H.J., Heide, J.V.D., Fenigsen-Narucka, U. and Peters, R. (1987b) Fate of salmonellas and competing flora in meat sample enrichments in buffered peptone water and in Muller-Kauffmanns' tetrathionate medium. J. Appl. Bacteriol. 62, 97-104.
Chau, P.Y. and Huang, C.T. (1974) A one-day selective migration procedure for detecting salmonellae in faeces. J. Clin. Pathol. 27, 405-407.

199
Chau, P.Y. and Huang, C.T. (1976) A simple procedure for screening of Salmonella using a semi-solid enrichment and a semi-solid indicator medium. J. Appl. Bacteriol. 41, 283-294.
Chau, P.Y. and Leung, Y.K. (1978) Inhibitory action of various plating media on the growth of certain Salmonella serotypes. J. Appl. Bacteriol. 45, 341-345.
Craigie, J. (1931) Studies on serological reactions of flagella of B. typhosus. J. Immunol. 21, 417-511. Curtis, G.D.W., and Baird R.M. (Eds.) 1993 Pharmacoepeia of Culture Media for Food Microbiology.
Additional monographs II. Int. J. Food Microbiol. 17, 201-268. D'Aoust, J.Y. (1991) Salmonella In: Doyle, M.P. (Ed.) Foodborne bacterial pathogens, pp. 428-445,
Marcel Dekker, inc., New York, Basel. De Smedt, J.M. and Bolderdijk, R.F. (1987) Dynamics of Salmonella isolation with modified semi-solid
Rappaport-Vassiliadis medium. J. Food Prot. 50, 658-661. De Smedt, J.M. and Bolderdijk, R.F. (1990). Collaborative study of the international office of cocoa,
chocolate, and sugar confectionery on the use of motility enrichment for Salmonella detection in cocoa and chocolate, J. Food Prot. 53, 659-664.
De Zutter, L., De Smedt, J.M., Abrams, R. Beckers, H., Catteau, M., de Borchgrave, J., Debevre, J., Hoekstra, J., Jonkers, F., Lenges, J., Notermans, S., van Damme, L., Vandermeersch, R., Ver- braeken, R. and Waes, G. (1991) Collaborative study on the use of motility enrichment on modified semisolid Rappaport-Vassiliadis medium for the detection of Salmonella from foods. Int. J. Food Microbiol. 13, 11-20.
Dixon, J.M.S. (1959) Observations on the growth in nutrient broth of mixtures of Escherichia coli strains, and of faecal specimens harbouring pathogenic Escherichia coli serotypes. J. Hyg. Cam- bridge 57, 174-183.
Edel, W. and Kampelmacher, E.H. (1968) Comparative studies on Salmonella-isolation in eight European laboratories. Bull Word Health Org. 39, 487-491.
Fagerberg, D.J. and Avens, J.S. (1976) Enrichment and plating methodology for Salmonella detection in food. A review. J. Milk Food Technol. 39, 628-646.
Fricker, C.R. and Girdwood, R.W.A. (1985) A note on the isolation of salmonellas from environmental samples using three formulations of Rappaport's broth. J. Appl. Bacteriol. 58, 343-346.
Fricker, C.R., Quail, E., McGibbon, L. and Girdwood, R.W.A. (1985) An evaluation of commercially available dehydrated Rappaport-Vassiliadis medium for the isolation of salmonellae from poultry, J. Hyg. Cambridge 95, 337-344.
Fricker, C.R. (1987) A Review. The isolation of salmonellas and campylobacters. J. Appl. Bacteriol. 63, 99-116.
Fung, F.Y.C. and Kraft, A.A. (1970) A rapid and simple method for the detection and isolation of Salmonella from mixed cultures and poultry products. Poult. Sci. 49, 46-54.
Goossens, H., Wauters, G., De Boeck, M., Jansens, M. and Butzler, J.P. (1984) Semisolid selective- motility enrichment medium for the isolation of salmonellae from fecal specimens. J. Clin. Microbiol. 19, 940-941.
Gruenenwald, R., Henderson, R.W. and Yappow, S. (1991) Use of Rambach propylene glycol containing agar for identification of Salmonella spp. J. Clin. Microbiol. 29, 2354-2356.
Harper, J. and Shortridge, F.K. (1969) A selective motility medium for routine isolation of Salmonella. J. Hyg. 67, 181-186.
Harvey, R.W.S. and Price, T.H. (1979) A review. Principles of Salmonella isolation. J. Appl. Bacteriol. 46, 27-56.
Harvey, R.W.S. and Price, T.H. (1980)Salmonella isolation with Rappaport's medium after pre-enrich- ment in buffered peptone water using a series of inoculum ratios. J. Hyg. 85, 125-128.
Inoue, T., Takagi, S., Ohnishi, A., Tamura, K. and Suzuki, A. (1968) Food-borne disease Salmonella isolation medium (MLCB). Paper presented at the 66th Annual Meeting of the Society Veterinary Science of Japan.
Jameson, J.E. (1961) A discussion of dynamics of Salmonella enrichment. J. Hyg. 60, 193-207. Kafel, S. and Pogorzelska, E. (1987) Transfer volume-dependent recovery of Salmonella from minced
meat. J. Food Prot. 50, 584-586. Kauffmann, F. (1935) Ein kombiniertes Anreicherungsverfahren fiir Thyphus- und Paratyphusbazillen.
Zbl. Bakt. I. Orig. 119, 148-152.

200
King, S. and Metzger, W.J. (1968) A new plating medium for the isolation of enteric pathogens. Appl. Microbiol. 16, 577-578.
Leifson, E. (1935) New culture media based on sodium deoxycholate for the isolation of intestinal pathogens and for the enumeration of the colon bacillus in milk and water. J. Pathol. Bacteriol. 40, 581-599.
Leifson, E. (1936) New selenite enrichment media for the isolation of typhoid and paratyphoid bacilli. Am. J. Hyg. 24, 423-432.
Leininger, H.V. (1976) Equipment, media, reagents, routine tests and strains. In: Speck, M.L. (Ed.) Compendium of methods for the microbiological examination of foods, p. 24 Am. Publ. Health Assoc.
Mackey, B.M. (1985) Quality control monitoring of liquid selective media used for isolating salmonel- lae. Int. J. Food Microbiol. 2, 41-48.
McCoy, J.H. (1962) The isolation of Salmonella. J. Appl. Bacteriol. 25, 213-224. McGibbon, L., Quail, E. and Fricker, C.R. (1984) Isolation of salmonellae using two forms of
Rappaport-Vassiliadis medium and brilliant green agar. Int. J. Food Microbiol. 1, 171-177. Moats, W.A. (1981) Update on Salmonella in foods: selective plating media and other diagnostic media.
J. Food Prot. 44, 375-380. Muller, L. (1923) Un nouveau milieu d'enrichissement pour la recherche du bacille typhique et des
paratyphique. Comp. Rend. Soc. Biol. 89, 424-437. Northolt, M.D., Stadhouders, J. and van Asseldonk, W. (1985) Comparison of Rappaport-Vassiliadis
medium and Mfiller-Kauffmann medium for the detection of salmonellae in caseinate and milk powder. Neth. Milk Dairy J. 39, 49-55.
Perales, I. and Erkiaga, E. (1991) Comparison between semisolid Rappaport and modified semisolid Rappaport-Vassiliadis media for the isolation of Salmonella spp. from foods and feeds. Int. J. Food Microbiol. 14, 51-58.
Peterz, M., Wiberg, C. and Norberg, P. (1989) The effect of incubation temperature and magnesium chloride concentration on growth of Salmonella in home made and in commercially available dehydrated Rappaport-Vassiliadis broths. J. Appl. Bacteriol. 66, 523-528.
Pietzsch, O. (1984) Comparative studies on Salmonella enrichment procedures. In: Agriculture. Priority aspects of Salmonella research. Larsen, H.E. (Ed.) Commission of the European Communities, pp. 85-95, Luxembourg, Office for official publications of the European Communities.
Pollock, H.M. and Dahlgren, B.J. (1974) Clinical evaluation of enteric media in the primary isolation of Salmonella and Shigella. Appl. Microbiol. 27, 197-201.
Quail, E., McGibbon, L. and Fricker, C.R. (1986) A study of the relative efficiencies of three commercially available dehydrated Rappaport-Vassiliadis media. J. Hyg. Cambridge 96, 425-429.
Rambach, A. (1990) New plate medium for facilitated differentiation of Salmonella spp. from Proteus spp. and other enteric bacteria. Appl. Environ. Microbiol. 56, 301-303.
Rappaport, F., Konforti, N. and Navon, B. (1956) A new enrichment medium for certain salmonellae. J. Clin. Pathol. 9, 261-266.
Rappaport, F. and Konforti, N. (1959) Selective enrichment medium for paratyphoid bacteria. In- hibitory and growth promoting factors. Appl. Microbiol. 7, 63-66.
Stuart, P.F. and Pivnick, H. (1965) Isolation of Salmonella by selective motility systems. Appl. Microbiol. 13, 365-372.
Taylor, W.F. (1965) Isolation of shigellae. I. Xylose-lysine agars: new media for the isolation of enteric pathogens. Am. J. Clin. Pathol. 44, 471-475.
Tongpim, S., Beumer, R.R., Tamminga, S.K. and Kampelmacher, E.H. (1984) Comparison of modified Rappaport's medium (RV) and Muller-Kauffmann medium (MK-ISO) for the detection of Salmonella in meat products. Int. J. Food Microbiol. 1, 33-42.
Van Schothorst, M., van Leusden, F.M., Jeunink, J. and De Dreu, J. (1977) Studies on the multiplica- tion of salmonellae in various enrichment media at different incubation temperatures. J. Appl. Bacteriol. 42, 157-163.
Van Schothorst, M. and Renaud, A.M. (1983) Dynamics of Salmonella isolation with modified Rappaport's medium. J. Appl. Bacteriol. 54, 209-215.

201
Van Schothorst, M., Renaud, A.M. and van Beek, C. (1987) Salmonella isolation using RVS broth and MLCB agar. Food Microbiol. 4, 11-18.
Vassiliadis, P., Paternaki, E., Papaiconomon, N., Papadakis, J.A. and Trichopoulos, D. (1976) Nouveau proc~d~ d'enrichessement de Salmonella. Ann. Microbiol. Inst. Pasteur 127B, 195-200.
Vassiliadis, P. (1983) The Rappaport-Vassiliadis (RV) enrichment medium for the isolation of Salmonellas: an overview. J. Appl. Bacteriol. 54, 69-76.
Wilson, W.J. and Blair, E.M.McV. (1927) Use of glucose bismuth sulphite iron medium for the isolation of Bacillus typhosa and Bacillus proteus. J. Hyg. 26, 374-391.
Zeh, G., Albrecht, M. and Busse, M. (1989) Enrichment of Salmonella at low nutrient levels. In: Modern Microbiological Methods for Dairy Products, pp. 424-428. Int. Dairy Fed., Brussels.
Zeh, G. (1991) Nachweis von Salmonellen in Milchpulver. Verbesserung der Methode fiir die Zwecke der Betriebskontrolle. Dissertation, Landwirtschaftliche Fakult~it, Technische Universit~it Mfinchen.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
203
Chapter 12 Culture media for the isolation and enumeration
of pathogenic Vibrio species in foods and environmental samples
T e r e n c e J. D o n o v a n a, Peter van Ne t t en b
a Area Public Health Laboratory, William Harvey Hospital, Ashford, Kent TN24 OLZ, UK b Q Laboratories Ltd, Quayside, Navigation Way, Preston PR2 2YP, UK
Abstract
The genus Vibrio now includes a large number of species. Clear evidence is only available for the aetiological role of V. cholerae, V. vulnificus and V. parahaemolyticus in foodborne diseases. Until recently, V. cholerae serogroup 0:1 was accepted as the cause of epidemic cholera. However, the designation of outbreaks of diarrhoeal diseases caused by V. cholerae 0:139 as clinical cholera has lead to renewed interest in Non 0:1 serogroups of V. cholerae.
A wide range of enrichment and selective media for the isolation of vibrios has been developed. These media are reviewed with respect to their ability to recover and differenti- ate the target vibrios. Alkaline peptone water (APW) remains the recommended enrich- ment mendium for vibrios in parallel with either salt polymyxin broth (SPB) or glucose salt teepol (or sodium dodecylsulphate) broth (GSTB) when tests for V. parahaemolyticus are required.
Thiosulphate citrate bile salt agar (TCBS) in parallel with polymyxin mannose tellurite (PMT) or sodium dodecylsulphate polymyxin sucrose agar (SPS) are the recommended selective plating media.

204
The family Vibrionaceae includes the genera, Vibrio, Aeromonas, Plesiomonas and Photobacterium. Members of the family are usually found in aquatic environ- ments but can also be isolated from man and other animals in which they may be pathogenic (Baumann et al., 1984). Vibrio spp. occur often in association with plankton and their numbers increase in parallel with seasonal planktonic bloom (Oliver, 1989; Twedt, 1989; West, 1989).
The genus Vibrio includes at least 30 species. Evidence from epidemiological and ecological studies has established the aetiological role of V. cholerae, V. vulnificus and V. parahaemolyticus in foodborne diseases (Oliver, 1989; Twedt, 1989; Roberts, 1992). V. cholerae serogroup 0:1 is best known as the cause of epidemic cholera. Recently, a non 0:1 V. cholerae serogroup 0:139 has been incriminated in a large outbreak of clinical cholera in Bangladesh (Bhattacharya et al., 1993). There are now 140 O serogroups of V. cholerae (Shimada et al., 1994). Gastroenteritis due to V. parahaemolyticus associated with the consumption of contaminated seafoods is found in several areas of the world. Strains of V. parahaemolyticus isolated from patients with gastroenteritis are Kanagawa-posi- tive; this phenomenon of haemolysis of human red blood cells is thought to be associated with enterotoxin production (Miyamoto et al., 1969). However, most environmental isolates are Kanagawa-negative (Twedt, 1989) and the mechanism of pathogenicity of V. parahaemolyticus is not fully understood. The role of other Vibrio spp. such as V. alginolyticus, V. fluvialis, V. furnissii, V. hollisae, V. mimicus, V. metchnikovii and V. damsela has not been firmly established (Stiles, 1989) but they have been implicated as opportunistic pathogens (West, 1989).
1. Rationale for screening foods for V. cholerae, V. parahaemolyticus and V. vulnificus
1.1. Ecology
V. cholerae is a non-halophilic organism and does not grow at 10~ or below. It occurs world-wide in fresh water, brackish water and coastal water. In areas where cholera is endemic, V. cholerae can frequently be found in the environment due to contamination via irrigation water, sewage and the use of untreated night soil to increase land fertility (Fersenfeld, 1965). However, sporadic outbreaks of cholera in non-endemic areas has lead to the detection of natural aquatic reservoirs for V. cholerae 0:1. The viable but non-culturable phenomenon may also explain the source of infection in these areas (West, 1989).
V. parahaemolyticus has similar temperature requirements to V. cholerae, and is also found worldwide, but because of its halophilic character, the organism is usually only resident in the sea and brackish waters (Twedt, 1989; Doyle, 1991).
V. vulnificus has been detected in areas with warm seawater temperatures (> 20~ (Kelly, 1982; Morris, 1988; Oliver, 1989) with a greater frequency of infection during the warmer months of the year (Tilton and Ryan, 1987).

205
1.2. Mode of infection
In the natural habitat food, such as seafood, is thought to be contaminated with pathogenic Vibrio spp. via water. The numbers of Vibrio spp. may increase in the seafood due to biological concentrat ion in fish and shellfish, especially bivalve
molluscs. Cholera is not always a waterborne disease (Roberts , 1992). Contaminat ion of
food followed by an increase in the number of vibrios may lead to foodborne cholera. In endemic areas food, such as vegetables, can become contaminated with V. cholerae via irrigation water, human faeces or sewage. Direct contaminat ion by food handlers excreting the organisms with or without symptoms is another route of transmission to a wide variety of foods (Table 1). Outside the natural habitat V. cholerae can grow above 10~ on non-acid foods with a low number of competit ive organisms (cooked foods) and a water activity greater than 0.93 (Fersenfeld, 1965;
Roberts, 1992). V. parahaemolyticus infection is particularly associated with consumption of raw
or undercooked seafood often p repared in the traditional Japanese culinary style (Table 1). Short periods of t empera ture abuse allow a rapid increase in numbers of V. parahaemolyticus in seafood.
V. vulnificus foodborne illness is invariably associated with consumption of raw or improperly cooked bivalve shellfish (Morris and Blake, 1985). Most infections
Table 1 Foods implicated in foodborne vibrio infections
Source Food V. cholerae V. paraheamolyticus V. vulnificus
Bivalve Mussels + molluscan Clams + shellfish Oysters +
Cockles + Curstacean Prawns + shellfish Crab +
Lobster + Fish Sardines +
Lagoon fish + Raw fish + Dried fish
Water Vegetables + Lettuce + Bottled water + Cooked rice + Potato salad +
Infected Food Cooked meats + Handlers Hors d'oevres +
Prawns + Chicken/rice + Cooked Squid +
Adapted from Oliver (1989), Twedt (1989) and Roberts (1992).

206
with V. vulnificus are rapidly invasive, often with septicaemia. Gastroenteritis is uncommon. There is a trend towards greater frequency of infection during the warmer weather when sea temperatures are high (Tilton and Ryan, 1987).
1.3. Methods of detection and control
Surveillance for pathogenic Vibrio spp. must be aimed at monitoring and controlling not only contamination of the end product, but also the natural habitat, the ingredients and the process environment. A qualitative enrichment procedure is normally performed for the detection of foodborne V. cholerae, V. para- haemolyticus and V. vulnificus. Quantitative procedures, either direct plating or most probable number (MPN), are occasionally used (Kaysner et al., 1989; Klontz et al., 1993). When V. cholerae has been isolated serotyping is used to determine if the isolate belongs to serogroup 0:1, and tests for cholera toxin, cholera-like toxin or cytotoxin production may be performed. A special medium, Wagatsuma agar, may be used to check the Kanagawa reaction of V. parahaemolyticus (Miyamoto et al., 1969).
Different selective systems are required when isolating Vibrio spp. to suppress the microbial populations characteristic of different foods and sampling sites. Non-fermentative Gram-negative bacteria, other Vibrio spp., Enterobacteriaceae and to a lesser extent Enterococcus, Staphylococcus and Micrococcus spp. are the most frequent competitors. Lactic acid bacteria, pseudomonads, Bacillus spp., yeasts and moulds are important competitive organisms in vegetables.
The protocol for the detection of pathogenic Vibrio spp. should include a procedure to recover sublethally injured organisms. An effective selective enrich- ment should suppress interfering organisms so that during enrichment competitive organisms will not overgrow pathogenic Vibrio spp. Selective plating media must allow an easy and reliable detection of suspect colonies of pathogenic Vibrio spp. These conditions can generally be achieved during the isolation of pathogenic Vibrio spp. from cooked foods, but their detection in heavily contaminated raw seafood and vegetables can be hindered by large numbers of competitors.
The isolation of the pathogenic vibrios, V. cholerae, V. parahaemolyticus and V. vulnificus, is frequently complicated by the presence of V. alginolyticus and V. metschnikovii in the marine environment, hence the effect of other Vibrio spp. needs to be evaluated when assessing detection methods.
2. Enrichment media for Vibrio cholerae
Culture media which were developed for the isolation of vibrios from faeces in clinical laboratories have also been employed for the isolation of IA. cholerae from foods. The main enrichment broths are listed in Table 2. Alkaline peptone water (APW) has been used from 1887 to the present time, as the standard medium for the enrichment of V. cholerae. A wide range of different formulae have been used for APW in the last 100 years. The variations include: (i) the type of peptone; (ii)
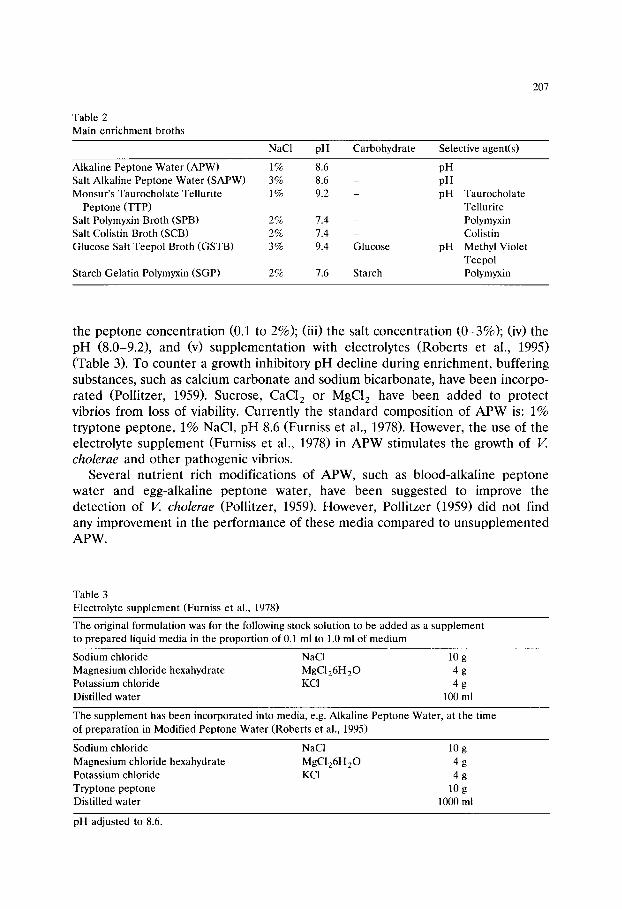
Table 2 Main enrichment broths
207
NaCI pH Carbohydrate Selective agent(s)
Alkaline Peptone Water (APW) 1% 8.6 Salt Alkaline Peptone Water (SAPW) 3% 8.6 Monsur's Taurocholate Tellurite 1% 9.2
Peptone (TTP) Salt Polymyxin Broth (SPB) 2% 7.4 Salt Colistin Broth (SCB) 2% 7.4 Glucose Salt Teepol Broth (GSTB) 3% 9.4
Starch Gelatin Polymyxin (SGP) 2% 7.6
Glucose
Starch
pH pH pH Taurocholate
Tellurite Polymyxin Colistin
pH Methyl Violet Teepol Polymyxin
the peptone concentration (0.1 to 2%); (iii) the salt concentration (0-3%); (iv) the pH (8.0-9.2), and (v) supplementation with electrolytes (Roberts et al., 1995) (Table 3). To counter a growth inhibitory pH decline during enrichment, buffering substances, such as calcium carbonate and sodium bicarbonate, have been incorpo- rated (Pollitzer, 1959). Sucrose, CaC12 or MgC12 have been added to protect vibrios from loss of viability. Currently the standard composition of APW is: 1% tryptone peptone, 1% NaC1, pH 8.6 (Furniss et al., 1978). However, the use of the electrolyte supplement (Furniss et al., 1978) in APW stimulates the growth of V. cholerae and other pathogenic vibrios.
Several nutrient rich modifications of APW, such as blood-alkaline peptone water and egg-alkaline peptone water, have been suggested to improve the detection of V. cholerae (Pollitzer, 1959). However, Pollitzer (1959) did not find any improvement in the performance of these media compared to unsupplemented APW.
Table 3 Electrolyte supplement (Furniss et al., 1978)
The original formulation was for the following stock solution to be added as a supplement to prepared liquid media in the proportion of 0.1 ml to 1.0 ml of medium
Sodium chloride NaCI 10 g Magnesium chloride hexahydrate MgCI26H2 ~ 4 g Potassium chloride KCI 4 g Distilled water 100 ml
The supplement has been incorporated into media, e.g. Alkaline Peptone Water, at the time of preparation in Modified Peptone Water (Roberts et al., 1995)
Sodium chloride NaC1 10 g Magnesium chloride hexahydrate MgC126H2 ~ 4 g Potassium chloride KCI 4 g Tryptone peptone 10 g Distilled water 1000 ml
pH adjusted to 8.6.

208
The use of gelatine as sole energy and nitrogen source to stimulate the growth of gelatinase-positive Vibrio spp. has been exploited in gelatine-phosphate-saline water (Spira, 1984). The presence of numerous other gelatinase-positive organisms in ecological niches where V. cholerae may be found reduces the usefulness of this medium for the selective enrichment of V. cholerae (Spira, 1984).
The overgrowth of V. cholerae by non-vibrios and other vibrios in APW and the antagonism of these organisms towards V. cholerae has led to the development of selective and differential-selective V. cholerae enrichment broths (Table 2). Apart from taurocholate tellurite peptone water (TTP) (Monsur, 1963), none of the selective media has replaced APW as the enrichment broth of choice for clinical specimens. The reasons are (i) the complexity of formulae; (ii) the difficulties in standardizing selective agents eg bile salts in APW; (iii) the inhibition of the growth of V. cholerae 0:1, particularly biotype E1Tor on bismuth sulphite type media; and (iv) differential systems outdated by the modern classification (Pol- litzer, 1959).
Monsur's TTP has been recommended as an alternative to APW for clinical samples, because of (i) the simplicity of its preparation; (ii) its usefulness as a transport medium; (iii) its better suppression of interfering intestinal organisms; and (iv) its similar or better support of the growth of V. cholerae (Pal et al., 1967; Furniss et al., 1978).
Other alternatives to APW are obtained by the addition either of tellurite (alkaline tellurite peptone water (ATPW); Pal et al., 1967)or of taurocholate (alkaline taurocholate peptone water (ABPW); Spira, 1984). A salt-free APW has been suggested to overcome the overgrowth of V. cholerae by halophilic Vibrio spp. (Spira, 1984). Starch-gelatin-polymyxin broth was suggested by Kitaura et al. (1983) for isolating V. cholerae 0:1 biotype E1Tor. The selective system consisted of 0.5% NaC1 and 1500 iu /ml polymyxin B.
3. Enrichment of Vibrio parahaemolyticus
The recognition of V. parahaemolyticus infections as food poisoning particularly associated with seafoods stimulated the development of selective enrichment media for use in food laboratories. These exploit the ability of V. parahaemolyticus to grow at alkaline pH (8.6-9.4), its halophilism (1-7% NaC1) and its resistance to a wide variety of selective agents. These include detergents, such as teepol (0.2-0.4%), se, dium dodecylsulphate (SDS; 0.13%), antibiotics, such as colistin (5000 iu/ml) and polymyxin B (100-1000 iu/ml), chemotherapeutic agents, such as al l- benzene sulphonate (0.2%), bile salts such as sodium lauryl sulphate (0.05%) and metachrome yellow II RD (0.125%). Some broths are made differen- tial by exploitation of activities common to Vibrio spp., such as gelatin hydrolysis, -" rch h?drolysis and sucrose fermentation. The need for strongly selective enrich- merit broths is not generally accepted. There is a consensus that the following enrichment media are effective for the isolation of V. parahaemolyticus: APW;

209
glucose salt teepol broth (GSTB); salt polymyxin broth (SPB) or salt colistin broth (SCB). GSTB can be modified by the substitution of SDS for teepol.
There are however differences in the type of polymyxin used, colistin (polymyxin E) or polymyxin B, whether the concentration used is expressed in/zg or units and whether the polymyxin is added before or after sterilisation. We suggest 100 iu of polymyxin B per ml of medium added after sterilisation and immediately before use as this reduces the loss of polymyxin activity during sterilisation and storage.
4. Enrichment media for Vibrio vulnificus
The only specific enrichment broth formulated for the enrichment of V. vulnificus from food is a starch gelatine polymyxin B broth. The selective system of the broth consists of NaC1 (2%) and polymyxin B (1500 iu /ml) (Kitaura et al, 1983). APW has been successfully used as an enrichment medium for pathogenic vibrios including V. vulnificus (Klontz, 1993).
5. Plating agars for Vibrio cholerae (Tables 4 and 5)
A wide range of plating agars has been formulated using, as in enrichment broths, the preference of V. cholerae for alkaline conditions, its resistance to bile salts, sodium tellurite, bismuth sulphite and some dyes. Alkaline nutrient-rich agars, such as alkaline blood, haemoglobulin, egg white, whole egg and casein were popular in the first part of this century. However, the poor selectivity of most of these media have made them of only historical interest (Pollitzer, 1959).
Thiosulphate citrate bile salts sucrose (TCBS)(Kobayashi et al., 1963) is a highly selective differential medium that is widely used not only for V. cholerae but all other pathogenic vibrios except V. holliseae. Its use was also recommended for the isolation of V. parahaemolyticus (Furniss et al., 1978) and V. vulnificus (Varnam and Evans, 1991). Its selective system consists of ox bile (0.8%), NaC1 (1%) and alkaline pH (8.6) which suppress the growth of most interfering organ- isms such as Enterobacteriaceae, pseudomonads, aeromonads, and Gram-positive bacteria (Kobayashi et al., 1963). The advantage of TCBS is its sucrose/ bromothymol blue diagnostic system which readily distinguishes sucrose-positive vibrios such as V. cholerae from other colonies (Table 5). This, its availability in commercially dehydrated form, and the fact that it does not have to be autoclaved have led to its widespread use. The disadvantages of TCBS are: its lot-to-lot and brand-to-brand variation in quality (West, 1984); its relatively high cost and the difficulty of identifying V. cholerae by direct agglutination due to sticky colonies that do not form stable suspension in saline. Oxidase reactions should not be performed on colonies from media such as TCBS containing fermentable carbohy- drates. The appearance of TCBS agar is blue green at pH 8.6. Monsur's gelatin taurocholate tellurite (GTT) agar exploits the selective system of tellurite (0.0005- 0.0001%), taurocholate (0.5%) and alkaline pH (8.5) (Monsur, 1961). In contrast to

210
Table 4 Main selective agars
NaC1 pH Carbohydrate Selective agents Indicators
1% 8.6 Sucrose Thiosulphate BTB Sodium + Ferric citrate TB
1% 8.5 - Taurocholate Tellurite
Thiosulphate Citrate Bile Salt Agar (TCBS)
Monsur's Gelatin Taurocholate Tellurite Agar (GTT)
Polymyxin Mannose Tellurite 1% (PMT)
SDS Polymyxin Sucrose 2% (SPS)
Cellobiose Polymyxin B 2% Colistin (CPC)
Bromothymol Blue Teepol 4% Agar (BTBT)
Sucrose Tellurite Teepol 0% Agar (STT)
Trypticase Soya Agar 3% Triphenyl tetrazolium (TSAT)
8.4 Mannose Polymyxin Tellurite SDS
7.6 Sucrose SDS Polymyxin CR
7.6 Cellobiose Colistin Polymyxin
7.8 Sucrose Teepol
8.0 Sucrose Teepol Tellurite
7.1 Sucrose Bile Salt
BTB
BTB
BTB CR BTB TB BTB
TTC
Vibrio vulnificus Agar (VV) 1% 8.6 Salicin Tellurite Crystal Violet Ox Gall
Vibrio vulnificus 2% 8.5 Cellobiose Ox Gall X Gal enumeration (VVE) Lactose Taurocholate
X-Gal Tellurite
Key: Thiosulphate = Sodium thiosulphate, Tellurite = Potassium tellurite, SDS = Sodium dodecylsul- phate, TTC = Triphenyl tetrazolium chloride, X-Gal = 5 bromo-4-chloroindoxyl-/3-o-galactopyranoside, BTB= Bromothymol Blue, TB= Thymol Blue, CR =Cresol Red, Taurocholate= Sodium tauro- cholate.
TCBS, GTT agar is an inexpensive medium which requires autoclaving but is not commercially available. Agglutination and oxidase tests can be performed directly on colonies from the plate. GTT agar has a light brown colour.
The difficulty of distinguishing V. cholerae colonies from other sucrose-ferment- ing organisms, such as Aeromonas and Proteus spp. on TCBS and problems with agglutination of colonies with V. cholerae 0:1 antiserum were the reasons for developing a modified vibrio agar (Tamura et al., 1971). The selective system of vibrio agar which consists of sodium deoxycholate (0.1%), ox bile (0.5%) and sodium lauryl sulphate (0.2%), makes it less selective than TCBS. The diagnostic system of the sucrose/waterblue-cresol red combination, however, is a feature of vibrio agar. It allows the bluish-grey, translucent and sticky V. cholerae colonies to be easily distinguished from the smaller blue colonies of proteus and aeromonas.
The variable performance of TCBS agar led to the development of the less complex sucrose tellurite teepol (STT) agar (Chatterjee et al., 1977). It combines the diagnostic system of TCBS with a selective system that comprises Teepol and tellurite (0.005%). Teepol replaced bile salts which are responsible for variation in the quality of TCBS and to a lesser extent TTG agar. STI' agar is a salt-free agar

211
which exploits the non-halophilic nature of V. cholerae. The sucrose concentration in STT agar is relatively low compared to that in TCBS. V. cholerae colonies on STT agar, like those on GTT agar, can be used directly for agglutination and oxidase tests. STT agar does not require autoclaving but is not commercially available.
Polymyxin mannose tellurite (PMT) agar was devised for easy differentiation of colonies of V. cholerae 0:1 from those of non-0:1 (Shimada et al., 1990). The differentiation is based on mannose/bromothymol blue-cresol red medium on which V. cholerae 0:1 gives yellow colonies with brown centre and most non-0:1 strains give dark violet colonies. The selective system consists of sodium dodecyl sulphate (0.02%) and polymyxin B (180 iu/ml) . PMT agar is commercially avail- able from Nissui Seiyaku Co. Tokyo. However, large numbers of non 0:1 V. cholerae isolates including the newly described 0:139 strain are mannose fer- menters and are not differentiated with PMT. Direct 0 agglutination reactions are more easily performed from PMT than TCBS. Sodium dodecylsulphate polymyxin sucrose (SPS) agar (Kitaura, 1983) and cellobiose-polymyxin B colistin (CPC) agar (Massad and Oliver, 1987)which were developed for the isolation of V. vulnificus have also been used for the isolation of V. cholerae. SPS has been used for the isolation of V. cholerae from environmental samples. Colonies of V. cholerae 0:1 produce a halo due to sulphatase activity and a yellow (acid) reaction, in contrast to V. vulnificus which produces colonies with halos and purple green (sucrose- negative) reactions. Other serogroups of V. cholerae show variable halo production (Bryant et al., 1987). CPC has been reported to be of potential value for isolating V. cholerae when incubated at 40~ (Massad and Oliver, 1987). Colonies of V. cholerae are purple (cellobiose-negative) compared with V. vulnificus colonies which are yellow (acid from cellobiose).
6. Plating media for Vibrio parahaemolyticus
After the recognition of V. parahaemolyticus as a halophile it was at first common practice to isolate the organism on conventional culture media supple- mented with 3% salt (Twedt, 1989). Competing microflora, however, obscured V. parahaemolyticus colonies on such media. As a consequence, a range of more selective plating media was developed, some of which are summarised in Table 4. Selective agents, such as bile salts, tellurite and Teepol, used in enrichment media (Table 2) have also been exploited in plating media. The diagnostic system used for most plating media is sucrose/bromothymol blue.
TCBS and Monsur's GTT which were developed for the isolation of V. cholerae have also been found useful for the isolation of V. parahaemolyticus (Furniss et al., 1978).
Bromothymol blue Teepol (BTBT) agar (Sakazaki, 1972) is another alternative to TCBS agar. It uses a diagnostic system similar to TCBS but a selective system in which ox bile is replaced by Teepol (0.2%) and the salt concentration is 4%. BTBT agar is commercially available from Nissui Seiyaku Co. Ltd. Tokyo.

212
Table 5 Colonial size and appearance of vibrios
Medium Vibrio Size (mm) Appearance
TCBS V. choleare 2-3 Yellow V. parahaemolyticus 3-5 Blue green V. vulnificus 2-3 Blue green V. alginolyticus 3-5 Yellow
GTT V. cholerae 2-3 Transparent grey with surrounding halo and faint black centre
V. parahaemolyticus 2-5 Opaque grey with surrounding halo and black centre
V. vulnificus V. alginolyticus 2-5 Opaque grey with surrounding halo
and black centre
PMT V. cholerae (mannose + ) 2-3 Yellow V. cholerae (mannose - ) 2-3 Dark Violet V. parahaemolyticus 3-4 Yellow V. vulnificus 1-2 Yellow V. alginolyticus 1-3 Yellow
SDS V. cholerae 1-3 Yellow with variable halo V. parahaemolyticus 2-3 Purple green no halo V. culnificus 1-3 Purple green with halo V. alginolyticus 1-2 Yellow no halo
CPC V. cholerae - Purple surrounded by blue zone V. parahaemolyticus NG V. vulnificus V. alginolyticus NG Yellow surrounded by yellow zone
BTBT V. cholerae 2-3 Yellow V. parahaemolyticus 3-5 Blue Green V. vulnificus - Blue Green V. alginolyticus 3-5 Yellow
STT V. cholerae 3-5 Yellow V. parahaemolyticus 3-5 Blue V. vulnificus V. alginolyticus 3-5 Yellow
TSAT V. cholerae V. parahaemolyticus 3-5 Red V. vulnificus V. alginolyticus 2-5 Pink swarming
VV V. cholerae NG V. parahaemolyticus V V V. vulnificus 2-4 Light grey translucent with black centre
V. alginolyticus
Size in mm of colonial diameter after incubation at 35-37~ for 20-24 h, except for CPC where results are from incubation at 40~ NG = No growth, - = no data, V = variation in size and appearance.
Difficulty in distinguishing V. parahaemolyticus from V. alginolyticus led to the development of trypticase soy agar triphenyltetrazolium (TSAT) agar (Kourany, 1983). This medium uses as a novel differential system, the combination of sucrose and triphenyltetrazolium. V. parahaemolyticus colonies on TSAT are large and red, thus allowing a better and more reliable differentiation from the smaller white

213
colonies with pink centres, of V. alginolyticus. Proteus colonies appear similar to V. parahaemolyticus on this medium and need to be distinguished by their appear- ance and reaction on triple sugar iron agar. The selective system consists of bile salts (0.05%) and salt (2.5%). The appearance of TSAT agar is straw yellow at pH 7.1.
7. Plating media for Vibrio vulnificus
TCBS has been used for the isolation of V. vulnificus from seafood and waters. V. vulnificus produces greenish blue sucrose-negative colonies, slightly smaller than V. parahaemolyticus colonies. To distinguish between V. vulnificus and V. para- haemolyticus colonies on TCBS further testing is required.
Concern about the recovery of V. vulnificus using TCBS led to the development of Vibrio vulnificus (VV) agar (Brayton et al., 1983). VV was developed by a step-wise modification to the nutrients and inhibitory compounds originally devel- oped for TCBS. Sodium citrate used in TCBS agar was found to inhibit V. vulnificus and so it was omitted. Salicin is employed as the source of carbohydrate with tellurite (0.0005%) crystal violet (0.00015%), ox gall (0.8%) and a pH of 8.6 to inhibit the growth of Gram-positive and Gram-negative organisms other than V. vulnificus. Crystal violet also provides a blueish-green tint to the medium for ease of colony recognition. VV agar is claimed to be more effective than TCBS agar in inhibiting Enterobacteriaceae, Pseudomonas spp. and staphylococci (Brayton et al., 1983). V. vulnificus appears as large (2-4 mm) light grey colonies with dark grey or black centre. Some strains of V. parahaemolyticus and V.fluvialis will grow on VV agar and form colonies similar to V. vulnificus (Brayton et al., 1983). Other Vibrio spp. give pinpoint opaque colonies which do not reduce tellurite.
Kitaura et al. (1983) emphasised the need for a new medium that differentiates V. vulnificus from similar sucrose-negative vibrios. The sulphatase activity of V. vulnificus was exploited in sodium dodecyl polymyxin sucrose (SPS) agar for the detection of V. vulnificus in the environment. When grown on SPS agar V. vulnificus colonies appear blue (sucrose-negative) surrounded by an opaque halo (sulphatase zone). Shewanella (formerly Alteromonas) putrefaciens also forms sucrose non-fermenting colonies with a halo and may be confused with V. vulnifi- CUS.
The lack of selectivity of TCBS and VV agar motivated Massad and Oliver (1987) to develop cellobiose, polymyxin B, colistin (CPC) agar for the isolation of V. vulnificus. The medium exploits the polymyxin (colistin and polymyxin B) resistance of Vibrio species. Incubation is carried out at 40~ to exclude marine isolates of the genera Pseudomonas, Flavobacterium, Photobacterium and other vibrios. However, the inclusion of two polymyxins, polymyxin B and colistin (polymyxin E), with the same spectrum of antimicrobial activity suggests a empiri- cal rather than a scientific basis for their use. The fermentation of cellobiose by V. vulnificus produces yellow colonies surrounded by a yellow halo. These colonies

214
are easily differentiated from the colonies of V. cholerae and other cellobiose- negative organisms which produce purple colonies surrounded by a blue zone.
Miceli et al. (1993) recommended a direct plating procedure using Vibrio vulnificus enumeration medium (VVE) for the enumeration of V. vulnificus. This complex medium contains ox gall, sodium cholate, sodium taurocholate and potassium tellurite as selective agents with a chromogenic substate 5 bromo-4-chlo- roindoxyl-/~-ogalactopyranoside (X-Gal) for the detection of /~-galactosidase colonies of V. vulnificus which are blue-green on this medium. VVE is claimed to be reliable, simple and inexpensive for the enumeration of V. vulnificus in oysters.
8. Mode of action of selective systems in some Vibrio culture media
The range of selective agents employed in recommended vibrio culture media is limited to alkaline pH, salts, bile salts, tellurite, Teepol, polymyxin B and colistin (polymyxin E). Bile salts inhibit most Gram-positive bacteria, some Gram-negative bacteria other than Enterobacteriaceae and occasionally Vibrio spp. Teepol has been used as a substitute for bile salts for Enterobacteriaceae and vibrio media (Jameson and Emberley, 1956; Chatterjee et al., 1977). Tellurite inhibits some Gram-positive bacteria but some bacteria frequently occurring in food, such as staphylococci, streptococci, enterococci and lactic acid bacteria are resistant. Polymyxins are bactericidal towards Gram-negative bacteria and interfere with the structure and function of their outer and cytoplasmic membranes. Polymyxins suppress the growth of Acinetobacter spp., Escherichia coli, Enterobacter spp., Klebsiella spp., Pseudomonas aeruginosa and Shigella spp. but resistant strains of Pseudomonas spp., Proteus spp. and Serratia marcescens are common. Polymyxin B and colistin (polymyxin E) have similar antibacterial spectra.
pH values of media in the alkaline range 8.0-9.4 have been used to exploit the tolerance of vibrios to alkaline growth conditions compared with most other bacteria found in the environment. Vibrios are able to continue to utilise sub- strates for growth at alkaline pH values and outgrow other bacteria. This combined with inhibitors, eg bile salts, leads to the selective growth of vibrios. Liquid media which do not contain buffering salts, eg APW, may not be able to retain an alkaline pH due to an inoculum of acidic or neutral samples.
The consensus view for the optimum pH value for the selective growth of vibrios is 8.4-8.6.
The presence of salts such as sodium chloride stimulates the growth of all vibrios; however, some vibrios require salts other than sodium chloride. The ionic requirements for the growth of marine bacteria were studied by MacLeod (1965). The electrolyte supplement as described by Furniss et al. (1978)with additional magnesium and potassium salts overcomes these difficulties. Vibrios can be di- vided into groups dependent on their salt requirements for growth. Tests such as salt tolerance are used in identification. Sodium chloride concentrations of 1-2% stimulate the growth of most vibrios but not most competitive bacteria. Sodium

215
chloride concentrations of over 5% make the medium selective for halophilic vibrios only eg V. parahaemolyticus and V. alginolyticus.
9. Recommended culture media
9.1. Vibrio cholerae
The combination of enrichment using APW with TCBS as the selective plating medium is the most common method. The performance of APW is rendered less selective by the presence of large numbers of competing flora. A short incubation period at 35-37~ with a subculture onto plating medium at that time followed by subculture after a further 18 h is thought to be an advantage when there are high numbers of competing flora and low numbers of V. cholerae.
The use of Monsur's TTP as an alternative enrichment broth is recommended, as is the possible use of a second selective plating medium such as PMT or Monsur's GTT. Further reports on the successful use of SPS and CPC agars for the isolation of V. cholerae are needed before they can be recommended.
9.2. Vibrio parahaemolyticus
Since the world-wide interest in V. parahaemolyticus in the 1970's, three enrichment broths are now most frequently used. These are glucose salt teepol salt broth (GSTB), salt polymyxin broth (SPB) and salt colistin broth (SCB). As with V. cholerae, short incubation (24 h) at 37~ is recommended to prevent overgrowth of competitors.
Nakanishi and Murase (1974) found both salt colistin broth (SCB) and salt polymyxin broth (SPB) were superior to glucose salt teepol broth (GSTB) for the enrichment of V. parahaemolyticus from raw fish and environmental swabs.
APW has been found to be most effective for the isolation of V. parahaemolyti- cus from oysters (Eyles et al., 1985) and prawns (Oscroft, 1987). The addition of 3% sodium chloride or electrolyte supplement should increase the efficiency of APW for V. parahaemolyticus (Furniss et al., 1978).
9.3. Vibrio vulnificus
The use of alternative plating media such as SPS, VVE and CPC should be considered for V. vulnificus. SPS is relatively easy to prepare and V. vulnificus colonies produce a halo due to sulphatase activity. V. cholerae and V. para- haemolyticus can also be isolated using this medium.
The use of SGP broth and SPS agar is suggested for the surveillance of seafood and sea water for V. vulnificus and V. cholerae 0:1 (Kitaura et al., 1983). The combination of polymyxin B broth and TCBS agar was found to be unsuccessful by these authors.

216
References
Baumann, P., Furniss, A.L. and Lee, J.V. (1984) Genus Vibrio. In: Bergeys's Manual of Systematic Bacteriology, 9th edition. Williams and Wilkins, Baltimore, MD, pp. 518-538.
Bhattacharya, S.K., Bhattacharya, M.K., Balakrish, G., Dutta, D., Deb, A., Ramamurthy, Garg, S., Saha., P.K., Dutta, P., Moitra, A., Mandal, B.K., Shimada, T., Takeda, Y. and Deb, B.C. (1993) Clinical profile of acute diarrhoea cases infected with the new epidemic strain of Vibrio cholerae 0:139: designation of the disease as cholera. J. Infect. 27, 11-15.
Brayton, P.R., West, P.A., Russek, E. and Collwell, R.R. (1983) New selective plating medium for isolation of Vibrio vulnificus biogroup 1. J. Clin. Microbiol. 17, 1039-1044.
Bryant, R.G., Jarvis, J. and Janda, J.M. (1987) Use of sodium dodecylsulphate-polymyxin B sucrose medium. Isolation of Vibrio vulnificus from shellfish. Appl. Environ. Microbiol. 53, 1556-1559.
Chatterjee, B.D., Kumar, P. and Tarun, S. (1977) Sucrose Teepol Tellurite agar: a new selective indicator medium for the isolation of Vibrio species. J. Infect. Dis. 135, 654-658.
Doyle, M.P. (1991) Pathogenic Escherichia coli, Yersinia enterocolitica and Vibrio parahaemolyticus foodborne illness. In: Foodbourne Illness: A Lancet Review, edited by W.M. Waites and J.P. Arbuthnott. Edward Arnold, London, pp. 77-85.
Eyles, M.J., Davey, G.R., Arnold, G. and Ware, H.M. (1985) Evaluation of methods for the enumera- tion and identification of Vibrio parahaemolyticus in oysters. Food Technol. Aust. 37, 302-304.
Fersenfeld, O. (1965) Notes on food, beverages and fomites contaminated with Vibrio cholerae. Bull. WHO 33, 725-734.
Furniss, A.L., Lee, J.V. and Donovan, T.J. (1978) The Vibrios. Monograph Series, Public Health Laboratory Service, HMSO, London.
Jameson, J.E. and Emberley, H.W. (1956) A substitute for bile salts in culture media. J. Gen. Microbiol. 15, 198.
Kelly M.J. (1982) Effect of temperature and salinity on Vibrio (Beneckia) vulnificus in a Gulf Coast environment. Appl. Environ. Microbiol. 44, 820-824.
Kitaura, T., Doke, S., Azuma, I., Imaida, K., Miyano, K., Harada, K. and Yabuuchi, E. (1983) Halo production by sulphatase activity in V. vulnificus and V. cholerae 0:1 on a new selective sodium dodecyl sulphate containing medium: a screening marker in environmental surveillance. FEMS Microbiol. Lett. 17, 205-209.
Klontz, K.C., William, L., Baldy, L.M. and Campos, M. (1993) Raw oyster-associated vibrio infections: linking epidemiological data with laboratory testing of oysters obtained from a retail outlet. J. Food Prot. 50, 977-979.
Kobayashi, T., Enomoto, S., Sakazaki, R. and Kuwahara, S. (1963) A new selective isolation medium for vibrio group on a modified Nakanishi's medium (TCBS agar medium). Jpn. J. Bacteriol. 18, 387-392.
Kourany, M. (1983) Medium for isolation and differentiation of Vibrio alginolyticus. Appl. Environ. Microbiol. 45, 310-312.
Macleod, R.A. (1965) The question of the existence of specific marine bacteria. Bacteriol. Rev. 29, 9. Massad, G. and Oliver, J. (1987) New selective differential medium for Vibrio cholerae and Vibrio
vulnificus. Appl. Environ. Microbiol. 53, 2262-2264. Miceli, G.A., Watkins, W.D. and Rippey, S.R. (1993) Direct plating procedures for enumerating Vibrio
vulnificus in oysters (Crussostrea virginica). Appl. Environ. Microbiol. 59, 3519-3524. Miyamoto, Y., Kato, T., Obara, Y., Akiyama, S., Takizawa, K. and Yamai, S. (1969) In vitro haemolytic
characteristic of Vibrio parahaemolyticus: its close association with human pathogenicity. J. Bacte- riol. 100, 1147-1149.
Monsur, K.A. (1961) A highly selective gelatin-taurocholate-tellurite medium for the isolation of Vibrio cholerae. Trans. R. Soc. Trop. Med. Hygiene 55, 440-442.
Monsur, K.A. (1963) Bacteriological diagnosis of cholera under field conditions. Bull. WHO 28, 387-389.
Morris, J.G. (1988) Vibrio vulnificus ndash; a new monster of the deep? Ann. Int. Med. 109, 261-262. Morris, J.G. and Blake, R.E. (1985) Cholera and other vibrios in the United States. New Engl. J. Med.
312, 343-350.

217
Nakanishi, H. and Murase, M. (1974) Enumeration of Vibrio parahaemolyticus in raw fish meat. In: International Symposium on Vibrio parahaemolyticus, edited by R. Fujino, R. Sakagachi, R. Sakazaki and Y. Takeda. Saikon Publishing Co., Tokyo.
Oliver, J.D. (1989) Vibrio vulnificus. In: Foodborne Bacterial Pathogens, edited by M.P. Doyle. Marcel Dekker, New York, pp. 569-600.
Oscroft, C.A. (1987) Effects of freezing on the survival of Vibrio parahaemolyticus and the efficiency of enumeration methods to recover this organism from frozen and non frozen prawn homogenates. CFPRA Tech. Memo 457, Chipping Campden.
Pal, S.C., Murty, G.V.S., Pandit, C.G., Murty, D.K. and Shrivastav, J.B. (1967) A comparative study of enrichment media in the bacteriological diagnosis of cholerae. Indian J. Med. Res. 55, 318-324.
Pollitzer, R. (1959) Cholera. WHO Monogr. Series 43, Geneva. Roberts, D. (1992) Growth and survival of Vibrio cholerae in foods. PHLS Microbiol. Digest 9, 24-31. Roberts, D., Hooper, W. and Greenwood, M. (1995) Practical Food Microbiology: Practical Method
6.14, Vibrio spp. Public Health Laboratory Service, London, pp. 155-158. Sakazaki, R. (1972) BTB teepol agar in media for bacteriological examination (2) Kindai Igaku Co.,
Tokyo. Shimada, T., Sakazaki, R., Fujimura, S., Niwano, K., Mishina, M. and Takizawa, K. (1990) A new
selective differential agar medium for isolation of Vibrio cholerae 0:1: PMT (polymyxin-mannose- tellurite) agar. Jpn. J. Med. Microbiol. 43, 37-41.
Shimada, T., Arakawa, E., Itoh, K., Okitsu, T., Matsushima, A., Asai, Y., Yamai, S., Nakazato, T., Nair, G.B., Albert, M.J. and Takeda, Y. (1994) Extended serotyping scheme for Vibrio cholerae Curr. Microbiol. 28, 175-178.
Spira, W.M. (1984) Tactics for detecting pathogenic vibrios in the environment. Chapter 17 in: Vibrios in the Environment, edited by R.R. Collwell. Wiley, New York, pp. 251-268.
Stiles, M.E. (1989) Less recognised or presumptive foodborne pathogenic bacteria. In: Foodborne Bacterial Pathogens, edited by M.P. Doyle. Marcel Dekker, New York, pp. 673-733.
Tamura, K., Shimada, S. and Prescott, L.M. (1971) Vibrio agar: a new plating medium for isolation of Vibrio cholerae. Jpn. J. Med. Sci. Biol. 24, 125-127.
Tilton, R.C. and Ryan, R.W. (1987) Clinical and ecological characteristics of Vibrio vulnificus in north eastern United States. Diagnostic Microbiology and Infectious Diseases 6, 109-117.
Twedt, R.M. (1989) Vibrio parahaemolyticus in foodborne bacterial pathogens In: Foodborne Bacterial Pathogens, edited by M.P. Doyle. Marcel Dekker, New York, pp. 544-568.
Varnam, A.H. and Evans, M.G. (1991) Foodborne pathogens: an illustrated text. Wolfe Publishing, London, pp. 176-180.
West, P.A. (1984) Statistical evaluation of a quality control method for isolation of pathogenic Vibrio species on selected thiosulphate-citrate-bile salts sucrose agars. J. Clin. Microbiol. 16, 1110-1116.
West, P.A. (1989) The human pathogenic vibrios. A public health update with environmental perspec- tives. Epidemiol. Infect. 103, 1-34.

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
219
Chapter 13
Isolation of Yersinia enterocolitica from foods
E. de Boer Inspectorate for Health Protection - Food Inspection Service, PO Box 9012, 7200 GN Zutphen, The Netherlands
Many selective enrichment and plating media for the isolation of Yersinia enterocolitica from foods are described. However, at present no single isolation procedure is available for the recovery of all pathogenic strains of Yersinia enterocolitica. Cold enrichment in phosphate-buffered saline plus 1% sorbitol and 0.15% bile salts (PBSSB) and two-step enrichment with tryptone soy broth (TSB) and bile oxalate sorbose (BOS) broth are very efficient methods for the recovery of a wide spectrum of serotypes of Y. enterocolitica. Enrichment in irgasan ticarcillin chlorate (ITC) broth was found to be the most efficient method for the recovery of strains of serotype 0:3, which is the most common clinical serotype of Y. enterocol#ica in Europe. Post-enrichment alkali treatment often results in higher isolation rates. Cefsulodin irgasan novobiocin (CIN) agar and Salmonella-Shigella deoxycholate calcium chloride (SSDC) agar are the most commonly used plating media. For the recovery of serotype 0:8 strains, the common clinical isolates in North America, enrichment in BOS and plating on CIN seems the most efficient procedure. Selection of the proper enrichment procedure will depend on the bio/serotypes of Yersinia spp. sought and on the type of food to be examined. The use of more than one medium for both enrichment and plating will result in higher recovery rates of Yersinia spp. from foods. Parallel use of the following two isolation procedures is recommended. (1) Enrichment in ITC for 2 days at 24~ plating on SSDC agar (2 days at 30~ (2) Pre-enrichment in TSB for 1 day at 24~ enrichment in BOS for 5 days at 24~ alkali treatment (mixing 0.5 ml enriched broth with 4.5 ml of 0.5% KOH in 0.5% NaC1 for 5 s); plating on CIN agar (2 days at 24~
Introduction
Yersinia enterocolitica is a coccoid-shaped Gram-negative, facultatively anaero- bic bacterium belonging to the genus Yersinia in the Enterobacteriaceae family. The organism is recognised as a foodborne pathogen and some large food-associ- ated outbreaks of yersiniosis have been reported. In developed countries, Y. enterocolitica can be isolated from 1-2% of all human cases of acute enteritis (Kapperud, 1991).
Yersinia enterocolitica and related species have been isolated from many types of food. The majority of these food isolates differ in biochemical and serological characteristics from typical clinical strains and are usually classified as 'non-patho-

220
genic' or 'environmental' Yersinia strains. These strains, which include the 'related species' Y. frederiksenii, Y. kristensenii, Y. intermedia, Y. aldovae, Y. rohdei, Y. mollaretii and Y. bercovieri, are ubiquitous and lack clinical significance with the exception of a few atypical cases (Kapperud, 1991). The strains associated with human disease mainly belong to the serogroups 0:3, 0:5, 27, 0:8 and 0:9 of Y. enterocolitica sensu stricto.
The epidemiology of Y. enterocolitica infections is, for the greater part, not understood. There is an association with consumption of contaminated foods, especially pork (De Boer and Nouws, 1991).
The increasing interest in Y. enterocolitica infections and the role of foods in some outbreaks of yersiniosis has led to the development of improved procedures for the isolation of this organism from foods during the last 10-15 years.
As the numbers of Y. enterocolitica organisms in foods are usually low and there is often a great variety of background flora, direct isolation on selective plating media is seldom successful. Isolation methods usually involve enrichment of the sample followed by plating onto selective agar media and confirmation of typical colonies.
Enrichment
As a psychrotrophic organism Y. enterocolitica is able to multiply at 4~ and enrichment at this temperature for 2-4 weeks is widely used. At this low tempera- ture, the growth rate of competitive bacteria is slowed sufficiently to enable Y. enterocolitica to multiply to numbers necessary for isolation on plating media.
Media used for this 'cold enrichment' include simple buffers like phosphate- buffered saline (PBS), PBS modified by addition of 1% sorbitol and 0.15% bile salts (PBSSB) (Mehlman et al., 1978), PBS supplemented with 1% mannitol (Schiemann, 1979a), PBS with 0.5% peptone (Weagant and Kaysner, 1983), PBS with peptone and cycloheximide (Vidon and Delmas, 1981), tryptone soy broth (Van Pee and Stragier, 1979), trypticase soy broth (Schiemann, 1983a) and tris- buffered peptone water, pH 8.0 (Greenwood and Hooper, 1989).
The long period required for cold enrichment is often unacceptable for quality assurance of foods. Schiemann and Olson (1984)showed that an incubation at 15~ for 2 days was as efficient as enrichment at 4~ for some weeks. Doyle and Hugdahl (1983) incubated PBS for 1-3 days at 25~ and Greenwood and Hooper (1989) incubated tris-buffered peptone water at 9~ for 11-14 days.
Several other enrichment procedures involving incubation at higher tempera- ture for shorter periods and using selective media have been proposed. Modified Rappaport broth (Wauters, 1973) has been used for many years as the first-choice medium for the isolation of the major pathogenic serotypes of Y. enterocolitica in Europe. This medium has been shown to inhibit the common North American serotype 0:8 strains of Y. enterocolitica (Schiemann, 1983a; Walker and Gilmour, 1986). Carbenicillin in modified Rappaport broth was shown to inhibit the growth of certain serotype 0:3 strains (Schiemann, 1982). However, Wauters et al. (1988)

221
stated that serogroup 0:3 strains are not inhibited by carbenicillin and that omis- sion of this antibiotic results in decrease of selectivity of the enrichment medium.
Lee et al. (1980) described two modified selenite media that were effective for recovery of certain strains of Y. enterocolitica from meats. They found it critical to limit the sample size of the blended meat suspension to 0.2 g per 100 ml enrichment medium to restrict the growth of competitive bacteria. Otherwise the slower growing Y. enterocolitica would be overgrown by the faster growing normal bacterial flora of the meat.
Schiemann (1982) developed a two-step enrichment procedure for recovery of Y. enterocolitica from food. In this procedure pre-enrichment for 9 days at 4~ in yeast extract rose bengal broth is followed by selective enrichment with bile oxalate sorbose (BOS) broth at 22~ for 5 days. As the pre-enrichment medium is less selective, it will allow multiplication of small inocula and repair of injured cells. BOS broth was found especially useful for the isolation of serotype 0:8 strains, but strains of serotype 5, 27 were more difficult to recover (Schiemann, 1983a).
Recently Wauters et al. (1988) developed a new enrichment broth, named ITC, derived from modified Rappaport broth and based on the selective agents irgasan, ticarcillin and potassium chlorate. In comparative studies ITC broth was especially effective for the recovery of Y. enterocolitica 0:3 from pork and porcine tonsils, while cold and two-step enrichments yielded better results for non-pathogenic strains (Wauters et al., 1988a; Kwaga et al., 1990; De Boer and Nouws, 1991).
Aulisio et al. (1980) found that strains of Y. enterocolitica are more tolerant of alkaline solutions than other Gram-negative bacteria. By treating food enrichments with potassium hydroxide (KOH) solutions before plating, the background flora is markedly reduced, making selection of Yersinia colonies from similar colonies on the isolation medium easier. However, Schiemann (1983b) found that various factors, including medium, temperature and growth phase, influence alkalotoler- ance and reduce the effectiveness of alkali treatment. Weagant and Kaysner (1983) concluded that a specific treatment time cannot be recommended. They found that streaking three to four successive plates from the KOH rinse at 10-s intervals enhanced the probability of obtaining isolated colonies of Y. enterocolitica even when growing in the presence of numerous organisms.
Direct KOH treatment of meat samples proved to be a valuable rapid method for direct isolation of Yersinia from meat contaminated with more than 102 cells per g (Fukushima, 1985). Based on results of comparative experiments the use of 0.125% KOH with an exposure time of 5 min was recommended for the direct detection of Y. enterocolitica in foods (Schraft and Untermann, 1989). Direct plating after KOH treatment is only suitable for strongly positive material (Wauters et al., 1988a).
Plating media
Different agar plating media have been used to isolate Y. enterocolitica from clinical specimens and food (Table 1). Initially media like MacConkey, Salmonella-

222
TABLE 1
Selective agents in enrichment and plating media for Yersinia enterocolitica
Medium a Selective agents
Enrichment MRB
Selenite Media
PBSSB
BOS
ITC
Plating CIN
VYE
SSDC
MacConkey
magnesium chloride (2.8%), malachite green (0.0013%), carbenicillin (0.00025%)
sodium selenite (0.15 or 0.25%), malachite green (0.002%), carbenicillin (0.001%)
bile salts (0.15%)
sodium oxalate (0.5%), bile salts (0.2%), irgasan (0.0004%), sodium furadantin (0.001%)
magnesium chloride (6%), malachite green (0.001%), irgasan (0.00001%), ticarcillin (0.00001%), potassium chlorate (0.1%)
sodium deoxycholate (0.05%), crystal violet (0.0001%), irgasan (0.0004%), cefsulodin (0. 0015%), novobiocin (0. 00025%)
sodium deoxycholate (0.1%), crystal violet (0.0001%), irgasan (0.0004%), cefsulodin (0.0004%), oleandomycin (0.001%), josamycin (0. 002%)
sodium deoxycholate (0. 85%)
bile salts (0.15%), crystal violet (0.0001%)
a MRB, modified Rappaport broth (Wauters, 1973); Selenite Media (Lee et al., 1980); PBSSB, phosphate-buffered saline with sorbitol and bile salts (Mehlman et al., 1978); BOS, bile oxalate sorbose broth (Schiemann, 1982; this volume); ITC, irgasan ticarcillin chlorate (Wauters et al., 1988a; this volume); CIN, cefsulodin irgasan novobiocin agar (Schiemann, 1979b; this volume); VYE, virulent Yersinia enterocolitica agar (Fukushima, 1987); SSDC, Salmonella-Shigella deoxycholate calcium chloride agar (Wauters, 1973; this volume), MacConkey Agar no. 3 (Oxoid CMll5).
Shigella, desoxycholate citrate and bismuth sulphite agars, designed for the isola- tion of enteropathogens, were used.
Lee (1977) modified MacConkey agar with Tween 80 to improve differentiation of Yersinia colonies from other lactose-negative colonies. However, lipolytic Yersinia strains which are easily recognized on this medium as white wrinkled colonies surrounded by a sheen, are usually non-pathogenic (De Boer and Seldam, 1987).
Salmonella-Shigella agar was made more selective for Y. enterocolitica by addition of sodium deoxycholate and CaC12 (SSDC)(Wauters, 1973, 1988a). Colonies of Y. enterocolitica on this medium are small, round and colourless. Some species of Morganella, Proteus, Serratia and Aeromonas can grow on SSDC and differentiation of Yersinia from these competing organisms may be difficult.
Schiemann (1979b) developed cefsulodin-irgasan-novobiocin (CIN) agar, a se- lective and differential agar medium for Y. enterocolitica. Organisms capable of
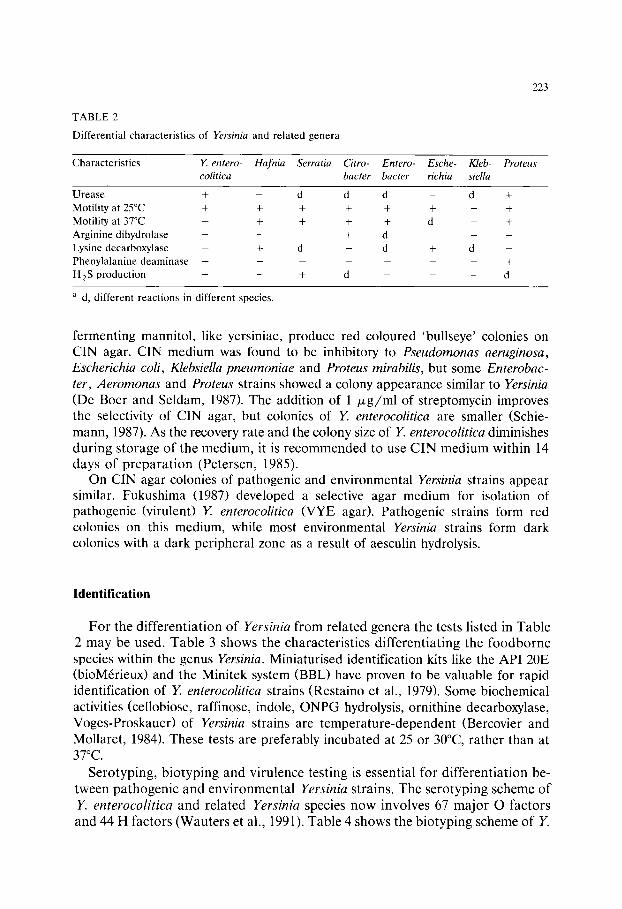
T A B L E 2
D i f f e r e n t i a l c h a r a c t e r i s t i c s o f Yersinia a n d r e l a t e d g e n e r a
2 2 3
C h a r a c t e r i s t i c s Y. entero- Hafnia Serratia Citro- Entero- Esche- Kleb- Proteus colitica bacter bacter richia siella
U r e a s e + - d d d - d +
M o t i l i t y a t 2 5 ~ + + + + + + - +
M o t i l i t y a t 3 7 ~ - + + + + d - +
A r g i n i n e d i h y d r o l a s e - - - + d - - -
L y s i n e d e c a r b o x y l a s e - + d - d + d -
P h e n y l a l a n i n e d e a m i n a s e . . . . . . . +
H 2 S p r o d u c t i o n - - + d - - - d
a d, d i f f e r e n t r e a c t i o n s i n d i f f e r e n t s p e c i e s .
fermenting mannitol, like yersiniae, produce red coloured 'bullseye' colonies on CIN agar. CIN medium was found to be inhibitory to Pseudomonas aeruginosa, Escherichia coli, Klebsiella pneumoniae and Proteus mirabilis, but some Enterobac- ter, Aeromonas and Proteus strains showed a colony appearance similar to Yersinia (De Boer and Seldam, 1987). The addition of 1 /xg/ml of streptomycin improves the selectivity of CIN agar, but colonies of Y. enterocolitica are smaller (Schie- mann, 1987). As the recovery rate and the colony size of Y. enterocolitica diminishes during storage of the medium, it is recommended to use CIN medium within 14 days of preparation (Petersen, 1985).
On CIN agar colonies of pathogenic and environmental Yersinia strains appear similar. Fukushima (1987) developed a selective agar medium for isolation of pathogenic (virulent) Y. enterocolitica (VYE agar). Pathogenic strains form red colonies on this medium, while most environmental Yersinia strains form dark colonies with a dark peripheral zone as a result of aesculin hydrolysis.
Identification
For the differentiation of Yersinia from related genera the tests listed in Table 2 may be used. Table 3 shows the characteristics differentiating the foodborne species within the genus Yersinia. Miniaturised identification kits like the API 20E (bioMdrieux) and the Minitek system (BBL) have proven to be valuable for rapid identification of Y. enterocolitica strains (Restaino et al., 1979). Some biochemical activities (cellobiose, raffinose, indole, ONPG hydrolysis, ornithine decarboxylase, Voges-Proskauer) of Yersinia strains are temperature-dependent (Bercovier and Mollaret, 1984). These tests are preferably incubated at 25 or 30~ rather than at 37~
Serotyping, biotyping and virulence testing is essential for differentiation be- tween pathogenic and environmental Yersinia strains. The serotyping scheme of Y. enterocolitica and related Yersinia species now involves 67 major O factors and 44 H factors (Wauters et al., 1991). Table 4 shows the biotyping scheme of Y.
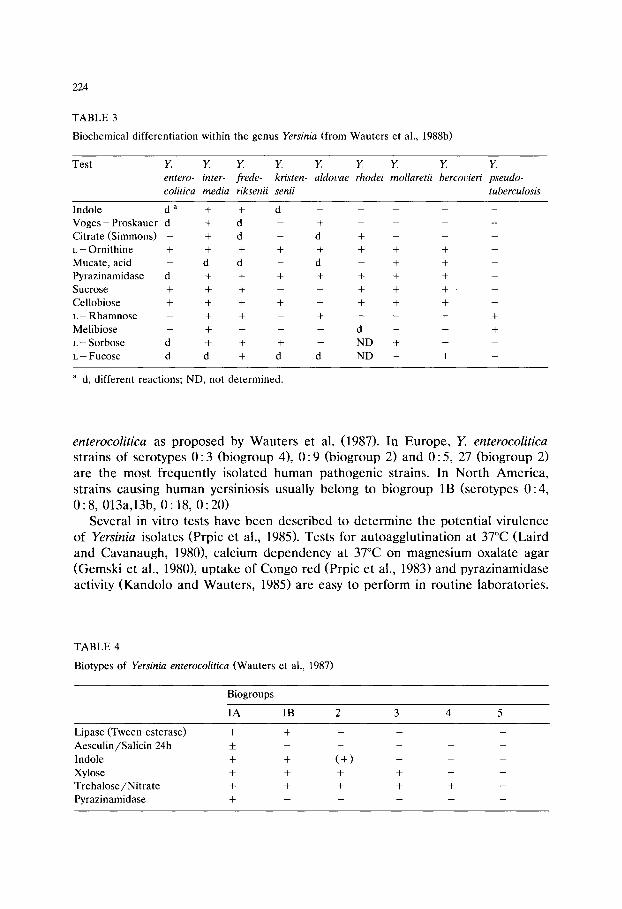
2 2 4
T A B L E 3
B i o c h e m i c a l d i f f e r e n t i a t i o n w i t h i n t h e g e n u s Yersinia ( f r o m W a u t e r s e t a l . , 1 9 8 8 b )
T e s t Y. Y. Y. Y. Y Y. Y. Y. Y.
entero- inter- frede- kristen- aldovae rhodei mollaretii bercovieri pseudo- colitica media riksenii senii tuberculosis
I n d o l e d a + + d . . . . .
V o g e s - P r o s k a u e r d + d - + . . . .
C i t r a t e ( S i m m o n s ) - + d - d + - - -
L - - O r n i t h i n e + + + + + + + + -
M u c a t e , a c i d - d d - d - + + -
P y r a z i n a m i d a s e d + + + + + + + -
S u c r o s e + + + - - + + + . -
C e l l o b i o s e + + + + - + + + -
L - - R h a m n o s e - + + - + - - - +
M e l i b i o s e - + - - - d - - +
L- - S o r b o s e d + + + - N D + - -
L - - F u c o s e d d + d d N D - + -
a d, d i f f e r e n t r e a c t i o n s ; N D , n o t d e t e r m i n e d .
enterocolitica as proposed by Wauters et al. (1987). In Europe, Y. enterocolitica strains of serotypes 0:3 (biogroup 4), 0:9 (biogroup 2) and 0:5, 27 (biogroup 2) are the most frequently isolated human pathogenic strains. In North America, strains causing human yersiniosis usually belong to biogroup 1B (serotypes 0:4, 0: 8, 013a,13b, 0: 18, 0: 20)
Several in vitro tests have been described to determine the potential virulence of Yersinia isolates (Prpic et al., 1985). Tests for autoagglutination at 37~ (Laird and Cavanaugh, 1980), calcium dependency at 37~ on magnesium oxalate agar (Gemski et al., 1980), uptake of Congo red (Prpic et al., 1983) and pyrazinamidase activity (Kandolo and Wauters, 1985) are easy to perform in routine laboratories.
T A B L E 4
B i o t y p e s o f Yersinia enterocolitica ( W a u t e r s e t a l . , 1 9 8 7 )
B i o g r o u p s
1 A 1B 2 3 4 5
L i p a s e ( T w e e n - e s t e r a s e ) +
A e s c u l i n / S a l i c i n 2 4 h _+
I n d o l e +
X y l o s e +
T r e h a l o s e / N i t r a t e +
P y r a z i n a m i d a s e +
+ (+) - _ -I- + + --
+ + + +

225
Comparative studies
From a comparison of pre-enrichment media, Schiemann (1983a) proposed tryptone soy broth as the medium of choice. Comparison of the two-step enrich- ment against cold enrichment and modified Rappaport broth showed improved recovery of human pathogenic strains of Y. enterocolitica from inoculated foods using pre-enrichment in tryptone soy broth followed by enrichment in BOS (Schiemann, 1982, 1983a; Walker and Gilmour, 1986).
In a study on the occurrence of Y. enterocolitica in foods Delmas and Vidon (1985) evaluated seven enrichment procedures. Most positive samples obtained within the shortest time were found using enrichment in PBSSB and BOS, with alkali treatment before plating onto CIN agar. However, only environmental serogroups of Y. enterocolitica were isolated.
The enrichment medium ITC was found superior to enrichment in BOS for isolation of pathogenic Y. enterocolitica from pork products, though enrichment in BOS isolated more non-pathogenic strains (Wauters et al., 1988a; Kwaga et al., 1990; De Boer and Nouws, 1991).
In different studies cold enrichment in PBSSB gave high isolation rates of Yersinia spp., which mainly proved to be non-pathogenic environmental strains (De Boer et al., 1982, 1986; De Boer and Seldam, 1987). In these studies MRB was found a suitable medium for the isolation of serogroup 0:3 and 0:9 strains, but much less efficient for the isolation of environmental strains.
Alkali treatment often resulted in a marked increase in the number of Yersinia isolations. This occurred especially when high numbers of competing organisms were present, as was usually the case for PBSSB, and when a plating medium with low selectivity, such as MacConkey agar, was used (De Boer et al., 1982; De Boer and Seldam, 1987).
MacConkey agar has been demonstrated to be a very productive medium for Y. enterocolitica (Mehlman et al., 1978; Schiemann, 1979b; De Boer and Seldam, 1987). However, Yersinia colonies are hard to recognize on this medium because of its low selectivity.
In several comparative studies CIN agar was found to be the most selective plating medium for Yersinia spp. (Aldova et al., 1990; Cox et al., 1990; Harmon et al., 1983; Schiemann, 1983a; Walker and Gilmour, 1986). Fukushima (1987)found that biotype 3B, serogroup 0:3 strains were inhibited on CIN agar. Differentiation of Yersinia colonies from other colonies on CIN agar is not always easy (Fukushima, 1987; De Boer and Seldam, 1987).
Plating ITC enrichments onto SSDC isolated more serogroup 0:3 strains than plating onto CIN agar (Wauters et al., 1988; De Boer and Nouws, 1991). SSDC proved to be unsuitable after alkali treatment (Wauters et al., 1988a).
Discussion
Addition of selective agents such as magnesium chloride, malachite green, bile salts, irgasan, and the antibiotics carbenicillin, ticarcillin and cefsulodin, to Yersinia

226
isolation media results in growth inhibition of the Gram-positive and part of the Gram-negative flora (Table 1). With regard to the differential properties of the Yersinia isolation media now in use, it is evident that the colony appearance of Yersinia on these media is not very characteristic and confirmation tests of presumptive colonies are always necessary.
At present, no single isolation procedure is available for the recovery of all pathogenic strains of Y. enterocolitica from foods. Cold enrichment in PBSSB, two-step enrichment with TSB and BOS, and enrichment in ITC are the most commonly used enrichment procedures. CIN and SSDC agars are the most commonly used plating media and are also commercially available. These enrich- ment and plating media are not very selective for Y. enterocolitica as they support the growth of several other members of the family of Enterobacteriaceae. This makes the isolation of low numbers of Yersinia in products containing many other contaminants rather difficult. Moreover, non-pathogenic environmental Y. entero- colitica strains are very common in many raw foods and may greatly hinder the isolation of pathogenic Yersinia strains from these products.
Cold enrichment and two-step enrichment are very efficient methods for the recovery of a wide spectrum of serotypes of Y. enterocolitica. However, usually one is only interested in the pathogenic serotypes and as the colony appearance of pathogenic and environmental strains on CIN and SSDC agar is similar, it requires much work to select pathogenic strains from these media. This highlights the need for the development and evaluation of agar media selecting for pathogenic serotypes, like VYE agar (Fukushima, 1985).
Enrichment in ITC was found to be the most efficient method for the recovery of strains of serotype 0:3, which is the most common clinical serotype of Y. enterocolitica in Europe. The usefulness of this enrichment broth for the isolation of other pathogenic serotypes remains to be determined. This also holds for SSDC agar, which is probably less suitable for pathogenic strains other than serotype 0:3.
For the recovery of the North American serotype 0:8 strains, enrichment in BOS and plating on CIN seems the most efficient procedure.
Selection of the proper enrichment procedure will depend on the bio/serotypes of Yersinia spp. sought and on the type of food to be examined. The use of more than one medium for both enrichment and plating will obviously result in higher recovery rates of Yersinia spp. from foods. Parallel use of the following two isolation procedures is recommended.
(1) Enrichment in ITC for 2 days at 24~ plating on SSDC agar (2 days at 30~
(2) Pre-enrichment in TSB for 1 day at 24~ enrichment in BOS for 5 days at 24~ alkali treatment (mixing 0.5 ml enriched broth with 4.5 ml of 0.5% KOH in 0.5% NaC1 for 5 s); plating on CIN agar (2 days at 24~
Monographs are described for CIN, BOS, ITC and SSDC in this volume.

227
References
Aldova, E., Svandova, E., Votypka, J. and Sourek J. (1990) Comparative study of culture methods to detect Yersinia enterocolitica serogroup 0:3 on swine tongues. Zbl. Bakteriol. 272, 306-312.
Aulisio, C.C.G., Mehlman, I.J. and Sanders, C. (1980) Alkali method for rapid recovery of Yersinia enterocolitica and Yersinia pseudotuberculosis from foods. Appl. Environ. Microbiol. 39, 135-140.
Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (Eds.) (1987) Pharmacopoeia of culture media for food microbiology. Int. J. Food Microbiol. 5, 208-209.
Bercovier, H. and Mollaret, H.H. (1984) Yersinia. In: Krieg, N.R. and Holt, J.G. (Eds.), Bergey's Manual of Systematic Bacteriology, Vol. 1, Williams and Wilkins, Baltimore and London.
Cox, N.A., Bailey, J.S., Del Corral, F., Shotts, E.B. (1990) Comparison of enrichment and plating media for isolation of Yersinia. Poultry Sci. 69, 686-693.
Curtis, G.D.W. and Baird, R.M. (Eds.) (1993) Pharmacopoeia of culture media for food microbiology: additional monographs (2). Int. J. Food Microbiol. 17, in press.
De Boer, E., Hartog, B.J. and Oosterom, J. (1982) Occurrence of Yersinia enterocolitica in poultry products. J. Food Prot. 45, 322-325.
De Boer, E., Seldam, W.M. and Oosterom, J. (1986) Characterization of Yersinia enterocolitica and related species isolated from foods and porcine tonsils in the Netherlands. Int. J. Food Microbiol. 3, 217-224.
De Boer, E. and Seldam, W.M. (1987) Comparison of methods for the isolation of Yersinia enterocolit- ica from porcine tonsils and pork. Int. J. Food Microbiol. 5, 95-101.
De Boer, E. and Nouws, J.F.M. (1991) Slaughter pigs and pork as a source of human pathogenic Yersinia enterocolitica. Int. J. Food Microbiol. 12, 375-378.
Delmas, C.L. and Vidon, D.J.M. (1985) Isolation of Yersinia enterocolitica and related species from foods in France. Appl. Environ. Microbiol. 50, 767-771.
Doyle, M.P. and Hugdahl, M.B. (1983) Improved procedure for recovery of Yersinia enterocolitica from meats. Appl. Environ. Microbiol. 45, 127-135.
Fukushima, H. (1985) Direct isolation of Yersinia enterocolitica and Yersinia pseudotuberculosis from meat. Appl. Environ. Microbiol. 50, 710-712.
Fukushima, H. (1987) New selective agar medium for isolation of virulent Yersinia enterocolitica. J. Clin. Microbiol. 25, 1068-1073.
Gemski, P., Lazere, J.R. and Casey, T. (1980) Plasmid associated with pathogenicity and calcium dependency of Yersinia enterocolitica. Infect. Immun. 27, 682-685.
Greenwood, M.H. and Hooper, W.L. (1989) Improved methods for the isolation of Yersinia species from milk and foods. Food Microbiol. 6, 99-104.
Harmon, M.C., Yu, C.L. and Swaminathan, B. (1983) An evaluation of selective differential plating media for the isolation of Yersinia enterocolitica from experimentally inoculated fresh ground pork homogenate. J. Food Sci. 48, 6-9.
Kandolo, K. and Wauters, G. (1985) Pyrazinamidase activity in Yersinia enterocolitica and related organisms. J. Clin. Microbiol. 21,980-982.
Kapperud, G. (1991) Yersinia enterocolitica in food hygiene. Int. J. Food Microbiol. 12, 53-66. Kwaga, J., Iversen, J.O. and Saunders, J.R. (1990) Comparison of two enrichment protocols for the
detection of Yersinia in slaughtered pigs and pork products. J. Food Prot. 53, 1047-1049. Laird, W.J. and Cavanaugh, D.C. (1980) Correlation of autoagglutination and virulence of yersinae. J.
Clin. Microbiol. 11, 430-432. Lee, W.H. (1977) Two plating media modified with Tween 80 for isolating Yersinia enterocolitica. Appl.
Environ. Microbiol. 33, 215-216. Lee, W.H., Harris, M.E., McClain, D., Smith, R.E. and Johnston, R.W. (1980) Two modified Selenite
media for the recovery of Yersinia enterocolitica from meats. Appl. Environ. Microbiol. 39, 205-209. Mehlman, I.J., Aulisio, C.C.G. and Sanders, A.C. (1978) Problems in the recovery and identification of
Yersinia from food. J. Assoc. Off. Anal. Chem. 61,761-771. Petersen, T. (1985) Keeping quality of cefsulodin-irgasan-novobiocin (CIN) medium for detection and
enumeration of Yersinia enterocolitica. Int. J. Food Microbiol. 2, 49-54.

228
Prpic, J.K., Robins-Browne, R.M. and Davey, R.B. (1983) Differentiation between virulent and avirulent Yersinia enterocolitica isolates by using Congo red agar. J. Clin. Microbiol. 18, 486-490.
Prpic, J.K., Robins-Browne, R.M. and Davey, R.B. (1985) In vitro assessment of virulence in Yersinia enterocolitica and related species. J. Clin. Microbiol. 22, 105-110.
Restaino, L., Grauman, G.S., McCall, W.A. and Hill, W.M. (1979) Evaluation of the Minitek and API 20E systems for identification of Yersinia enterocolitica. J. Food Prot. 42, 120-123.
Schiemann, D.A. (1979a) Enrichment methods for recovery of Yersinia enterocolitica from foods and raw milk. Contr. Microbiol. Immunol. 5, 212-227.
Schiemann, D.A. (1979b) Synthesis of a selective agar medium for Yersinia enterocolitica. Can. J. Microbiol. 25, 1298-1304.
Schiemann, D.A. (1982) Development of a two-step enrichment procedure for recovery of Yersinia enterocolitica from food. Appl. Environ. Microbiol. 43, 14-27.
Schiemann, D.A. (1983a) Comparison of enrichment and plating media for recovery of virulent strains of Yersinia enterocolitica from inoculated beef stew. J. Food Prot. 46, 957-964.
Schiemann, D.A. (1983b) Alkalotolerance of Yersinia enterocolitica as a basis for selective isolation from food enrichments. Appl. Environ. Microbiol. 46, 22-27.
Schiemann, D.A. (1987) Yersinia enterocolitica in milk and dairy products. J. Dairy Sci. 70, 383-391. Schiemann, D.A. and Olson, S.A. (1984) Antagonism by Gram-negative bacteria to growth of Yersinia
enterocolitica in mixed cultures. Appl. Environ. Microbiol. 48, 539-544. Schraft, H. and Untermann, F. (1989) Use of KOH treatment for direct detection of Yersinia
enterocolitica in foods. 10th WAVFH Int. Symp., Stockholm, Abstracts, p. 31. Van Pee, W. and Stragier, J. (1979) Evaluation of some cold enrichment and isolation media for the
recovery of Yersinia enterocolitica. Antonie van Leeuwenhoek J. Microbiol. Serol. 45, 465-477. Vidon, D.J.M. and Delmas, C.L. (1981) Incidence of Yersinia enterocolitica in raw milk in Eastern
France. Appl. Environ. Microbiol. 41, 355-359. Walker, S.J. and Gilmour, A. (1986) A comparison of media and methods for the recovery of Yersinia
enterocolitica and Yersinia enterocolitica-like bacteria from milk containing simulated raw milk microfloras. J. Appl. Bact. 60, '175-183.
Wauters, G. (1973) Improved methods for the isolation and the recognition of Yersinia enterocolitica. Contr. Microbiol. Immunol. 2, 68-70.
Wauters, G., Kandolo, K. and Janssens, M. (1987) Revised biogrouping scheme of Yersinia enterocolit- ica. Contr. Microbiol. Immunol. 9, 14-21.
Wauters, G., Goossens, V., Janssens, M. and Vandepitte, J. (1988a) New enrichment method for isolation of pathogenic Yersinia enterocolitica serogroup 0:3 from pork. Appl. Environ. Microbiol. 54, 851-854.
Wauters, G., Janssens, M., Steigerwalt, A.G. and Brenner, D.J. (1988b) Yersinia mollaretii sp.nov, and Yersinia bercovieri sp. nov., formerly called Yersinia enterocolitica biogroups 3A and 3B. Int. J. Syst. Bacteriol. 38, 424-429.
Wauters, G., Aleksic, S., Charlier, J. and Schulze, G. (1991) Somatic and flagellar antigens of Yersinia enterocolitica and related species. Contr. Microbiol. Immunol. 12, 239-243.
Weagant, S.D. and Kaysner, C.A. (1983) Modified enrichment broth for isolation of Yersinia enterocolit- ica from nonfood sources. Appl. Environ. Microbiol. 45, 468-471.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
229
Chapter 14 Media for detecting and enumerating yeasts
and moulds
Larry R. Beuchat Food Safety and Quality Enhancement Laboratory, Department of Food Science and Technology,
University of Georgia, Griffin, GA 30223-1797, USA
Dilution plating techniques are designed to determine populations of viable fungal propagules per unit weight or volume of food. Direct plating techniques, on the other hand, are designed to assess the internal mycoflora of individual pieces of foods, e.g., seeds or dried fruits, and results are expressed as a percentage of infected pieces. Both techniques are used by industry and regulatory agencies to monitor levels of fungal contamination at various stages of food handling, storing, processing and marketing. Peptone (0.1%) water is commonly used as a diluent for samples to be homogenized or blended. Buffered diluents containing up to 30% glucose or 60% sucrose are recommended for enumerating xerophiles. No one medium is satisfactory for detection or enumeration of yeasts and moulds in all foods. Dichloran rose bengal chloramphenicol agar, oxytetracycline glucose yeast extract agar and rose bengal chloramphenicol agar are superior to acidified potato dextrose agar for enumeration of yeasts and moulds. Dichloran 18% glycerol agar performs well for enumerating moderately xerophilic yeasts and moulds. Fastidious xerophiles require media containing high concentrations of sugars and /o r sodium chloride. Media have been formulated to detect potentially aflatoxigenic aspergilli and mycotox- igenic strains of penicillia and fusaria, but increased selectivity and specificity of media for detecting mycotoxigenic moulds are needed. Heat-resistant mould ascospores often require heat treatment prior to plating in order to activate the germination process. The spread-plate technique is strongly preferred over the pour-plate technique for enumerating yeasts and moulds. The recommended incubation temperature is 25~ but incubation time between plating and counting colonies ranges from 5 days for determination of general populations of mycoflora to 4 weeks or more for fastidious xerophiles. There is a need for new and improved media for selectively isolating various groups, genera, species and /o r strains of fungi capable of growing only under specific environmental conditions, e.g., low a w or, in the case of sublethally injured cells, under conditions which facilitate resuscitation. Improved media are needed which accurately detect moulds producing specific mycotoxins in a wide range of food types.
Introduction
Yeasts and moulds are distributed widely in soil, water and air. Consequently, unprocessed materials of both plant and animal origin are contaminated with fungi

230
at the time they reach the food manufacturer. Processing schemes can either render the finished food product free of fungi or merely reduce populations. Given enough time, survivors may grow and eventually spoil the product. The detection and enumeration of viable yeasts and moulds in unprocessed and processed foods is an integral part of total quality management programs, and can be used to monitor the effectiveness of sanitation practices at each step during post-harvest and post-slaughter handling, processing and distribution of foods.
Dilution plating techniques are designed to determine populations of viable fungal propagules per unit weight or volume of food. Direct plating techniques, on the other hand, are designed to assess the internal mycoflora of individual pieces of foods, e.g., seeds or dried fruits, and results are expressed as a percentage of infected pieces. The direct plating technique is qualitative, then, rather than quantitative. Both techniques are used by industry as well as regulatory agencies to monitor relative levels of fungal contamination at various stages of food handling, storing, processing and marketing. Procedures for analysis of any given product may differ from laboratory to laboratory. Certainly, procedures differ for analysing various types of foods. There are excellent reviews and descriptions of dilution and direct plating procedures in the literature (Booth, 1971a,b; Corry, 1982a; Jarvis et al., 1983; Koburger and Marth, 1984; Mislivec and Stack, 1984; King et al., 1986; Jarvis and Williams, 1987; Beuchat and Hocking, 1990) and the reader is referred to these publications. For detailed descriptions of common foodborne fungi, see Pitt and Hocking (1985a) and Samson and van Reenen-Hoekstra (1988). The following text provides essentially only an introduction to procedures for dilution and direct plating for the purpose of determining mycological quality of foods.
Sampling procedure
Procedures for selecting samples of foods for mycological analyses differ de- pending upon the consistency or physical state of the food. Replicate samples representative of the lot to be examined must be obtained. The size of the sample for dilution plating may range from 5 to 250 g (ml). Samples selected for direct plating should consist of at least 100 pieces, six to ten pieces per 9-cm plate. All foods, whether solid or liquid, should be thoroughly mixed before samples are taken. Dry foods such as nuts and cereal grains are usually ground to a powder if they are to be analysed using the dilution plating technique. Sampling procedures are outlined by the International Commission on Microbiological Specifications for Foods (ICMSF, 1986).
Preparation of samples
Preparation of samples for dilution plating may consist of manually shaking with a known volume of diluent, direct blending with a diluent in a blender or

231
pummelling with a diluent in a Stomacher. In some instances, rinsing of the entire sample with a diluent may be necessary. Jarvis et al. (1983) reported that higher populations are usually obtained by stomaching or blending than by shaking techniques. Hastings et al. (1986) observed that there were no significant differ- ences between mould populations detected in samples prepared by blending or stomaching. Methods by which samples are homogenized prior to secondary dilution are described by Jarvis and Williams (1987).
Soaking foods in diluent prior to mixing and diluting can have at least three effects (Seiler, 1986). First, soaking may allow resuscitation of sublethally damaged cells from dried, intermediate moisture or acidic foods. Soaking may also facilitate better release of cells which are present within tissues. Thirdly, with hard or sharp materials such as cereals and nuts, the softening effect of soaking will help prevent damage to stomacher bags used for preparing primary dilutions. After homogeniz- ing, samples should not be allowed to stand for more than a few minutes before removing a portion for subsequent dilution and plating, since fungal propagules may settle to the bottom of the container, resulting in under- or overestimation of populations, depending upon the location in the mixture from which the portion is withdrawn.
To assess the internal mycoflora of cereal grains, nuts and dried fruits and vegetables by direct plating, various chemicals are used for surface disinfection. These include 3% hydrogen peroxide, 2% potassium permanganate, 75% ethanol, 0.001% mercuric chloride and 0.35% sodium or calcium hypochlorite (Booth, 1971a). Contact time ranges from 1 to 5 min, followed by rinsing with sterile distilled water. Andrews (1986) stated that most food particles should be effec- tively surface disinfected by immersion in a 0.35% (w/v) sodium hypochlorite solution for 2 min. Rinsing with sterile water is not essential, provided samples are adequately drained after disinfection.
Diluent
Peptone (0.1%) water and 0.05-0.1 M potassium phosphate buffer (pH 7.0) are among the most commonly used diluents for yeast and mould enumeration. However, several other diluents may be employed, depending upon the type of food under investigation. For example, it is important to use a diluent containing a sufficient amount of solute to minimize osmotic shock to xerophilic mould and osmophilic yeast cells when serial dilutions are made prior to plating. Buffered diluents containing up to 30% glucose or 60% sucrose have been used for this purpose. Death of halophilic fungi is minimized by incorporating 6 to 10% sodium chloride into diluents.
Surface active agents may be added to diluents to reduce clumping of mould spores and conidia. For a discussion of diluents suitable for various foods, see King et al. (1986) and Jarvis and Williams (1987).

232
Culture media
General purpose media Because foods differ widely in composition and the environments to which they
have been exposed, the types and populations of mycoflora also differ. Pitt (1986) characterized the ideal enumeration medium as having the following attributes. It should suppress bacterial growth completely, without affecting growth of food fungi. It should be nutritionally adequate and support the growth of relatively fastidious fungi. The radial growth of colonies should be constrained but spore germination should not be inhibited.
Unfortunately, no one medium is satisfactory for detection or enumeration of yeasts and moulds in all foods. Traditionally, acidified potato dextrose agar (PDA) has been used for enumeration, but this medium is not an exceptionally good nutrient source and it may inhibit resuscitation of injured cells due its low pH (3.5). Antibiotic supplemented media such as oxytetracycline glucose yeast extract (OGY) agar (Mossel et al., 1962, 1970), rose bengal chlortetracycline (RBC) agar (Jarvis, 1973) and dichloran rose bengal chloramphenicol (DRBC) agar (King et al., 1979) are superior to acidified media for both dilution and direct plating techniques. Monographs have been published for all these media in the ICFMH Pharma- copoeia (see this volume). They are less inhibitory to injured yeasts and moulds, more effective in inhibiting bacterial growth and less likely to cause precipitation of food particles because of their higher pH (5-6). Basal media to which chloram- phenicol, chlortetracycline, oxytetracycline, gentamicin, kanamycin or streptomycin is added at concentrations up to 100 ~g /ml include OGY, DRBC, plate count agar, mycophil agar, malt agar and tryptone glucose yeast extract agar. The effectiveness of most antibiotics is diminished at alkaline pH, so if highly alkaline foods are being analysed, adjustment of the medium pH to less than 8.0 may be necessary to minimize bacterial growth.
Rose bengal (Jarvis, 1973; King et al., 1979) and dichloran (2,6-dichloro-4- nitroaniline) have been successfully used to control growth of spreading moulds (King et al., 1979). Caution should be taken to avoid exposure of rose bengal supplemented media to light, since cytotoxic breakdown products (Banks et al., 1985) may result in underestimation of mycoflora in samples.
Two techniques have been developed for general enumeration of yeasts and moulds which are a clear departure from traditional methods. The Hydrophobic Grid Membrane Filter (HGMF) technique, using an automated ISO-GRID TM
sample processor counting system was developed by Brodsky et al. (1982). The HGMF is essentially a membrane base upon which is applied a hydrophobic grid. The food sample is passed through the filter before transferring it to the surface of PDA (pH 5.6) supplemented with 100 ppm chloramphenicol and chlortetracycline. Plates are incubated for 48 + 4 h at 25~ before colonies are counted.
A culture film (Petrifilm TM YM) method has been developed for enumerating yeasts and moulds in foods. A dry selective indicator medium on a special support film is rehydrated upon application of 1 ml of diluted sample. The sample is then covered with a second film to prevent drying during incubation at 25~ for 4 or 5

233
days. This system has been demonstrated to perform well for enumerating yeasts and moulds in a wide range of types of foods (Beuchat et al., 1990, 1991).
Xerophilic fungi While many yeasts and moulds capable of growing at low a w can also grow at
high a w, some species require low a w in recovery media to enable growth and colony development. Reduction of a w is achieved by supplementing basal media with sodium chloride or polyhydric alcohols such as glycerol, glucose, fructose or sucrose. Dichloran 18% glycerol (DG18) agar (Hocking and Pitt, 1980; Baird et al., 1987), glucose yeast extract sucrose agar, yeast extract sucrose agar, malt salt agar (King et al., 1986), 25% glycerol nitrate agar (Pitt, 1979) and malt yeast 5% salt 12% glucose agar (Pitt and Hocking, 1985b) are but a few of the mycological media formulated to select for fungi capable of growing at low a w. Others are described by Beuchat and Hocking (1990).
DG18 agar (a w 0.95) was developed for enumeration of moderately xerophilic moulds and osmophilic yeasts in products such as grains, flours, nuts and spices. The medium supports growth of the Aspergillus restrictus series, Wallemia sebi, Zygosaccharomyces rouxii, Debaryomyces hansenii and many penicillia and other aspergilli (Beuchat and Hocking, 1990). Unfortunately, some Eurotium species grow too rapidly on DG18 agar, obscuring the growth of other xerophiles. The use of surfactants and fungicides to control spreading of Eurotium colonies on DG18 agar has been recommended (Beuchat and Tapia de Daza, 1992; Tapia de Daza and Beuchat, 1992).
Differences exist in tolerance of xerophilic fungi to various solutes and hydro- gen ion concentration. In an investigation of germination and growth of six xerophilic moulds, Pitt and Hocking (1977) concluded that a universal isolation medium for enumerating xerophilic fungi could be based on glycerol or glucose/fructose but not on sodium chloride as the aw-limiting solute.
Apart from species of the Aspergillus glaucus and A. restrictus groups, Xe- romyces bisporus, Wallemia sebi, Chrysosporium species and some Eurotium and Scopulariopsis species, the vast majority of moulds are not able to grow in the a w range of 0.61-0.70. Likewise, only a few species of yeasts will grow in this a w range. Those that can grow have been described as osmophilic and can cause spoilage of high-sugar products. Particularly troublesome is Zygosaccharomyces rouxii. Raw sugar cane, maple, chocolate and fruit syrups, honey, confectionery products, jams and jellies, fruit concentrates and dried fruits are most vulnerable to degradation by Z. rouxii. Spoilage of these products may also be due to growth of Z. bailii, but this yeast is more likely to be implicated in acidic foods such as salad dressings, mayonnaise and pickles with somewhat higher a w.
As with xerophilic moulds, plating media with reduced a w are required for recovering osmophilic yeasts from foods. Potato dextrose agar supplemented to contain 60% sucrose (Restaino et al., 1985), a yeast extract medium containing 50% glucose (Jermini et al., 1987) and a wide range of other sugar-supplemented media (Scarr, 1959; King et al., 1986; Beuchat and Hocking, 1990) are suitable for

234
isolating and enumerating osmophilic yeasts. Often, xerophilic moulds will also develop on these media.
Heat-resistant moulds Spoilage of thermally processed fruits and fruit products by heat-resistant
strains of Byssochlamys fulva, B. nivea, Neosartorya fischeri, Talaromyces flavus, Talaromyces bacillisporus and Eupenicillium brifeldianum have been recognized for some time (Beuchat and Rice, 1979; Beuchat and Pitt, 1992). Ascospores of these moulds show high heat resistance, in some instances comparable to bacterial spores. Because of their low incidence in fruit products (less than 10 per 100 g or ml), relatively large samples must be analysed. Centrifugation may be used to concentrate ascospores in liquid fruit products, the force and time necessary being influenced by volume, viscosity and specific gravity of the sample.
Heat-resistant ascospores may require heat activation before germination and growth will occur (Splittstoesser et al., 1972; Katan, 1985; Beuchat, 1987). Samples are heated at 75-80~ for 1 h before 50-ml quantities are combined with 100 ml of 1.5 • strength potato dextrose agar in 150-mm Petri plates. Plates are incubated at 30~ for up to 30 days before colonies are counted. Most viable ascospores will germinate and form visible colonies within 10 days; however, heat-injured and other debilitated ascospores may require additional time to form colonies. An alternative method which avoids error due to aerial contamination involves heating samples in flat-sided bottles followed by incubation on their sides. Colonies develop on the surface of the fruit product. This system is suitable for pulps and homogenates.
Mycotoxigenic moulds Several media have been formulated for use in recovering specific genera or
groups of yeasts and moulds based on metabolic activities. Among these are media suitable for enumeration of moulds capable of producing mycotoxins. These media are of value to any quality assurance program concerned with mycological quality of foods from a public health standpoint. The presence of mycotoxin producers in a food does not necessarily mean that mycotoxins are present. On the other hand, the absence of mycotoxin producers is not evidence that mycotoxins are not present in the food, since growth followed by death of mycotoxigenic moulds may have occurred at some point prior to analysis. Nevertheless, media formulated to select for mycotoxin producers are of interest to the food mycologist and consider- able effort is being expended in several laboratories to develop these media for routine use.
Detection of Aspergillus flavus and Aspergillus parasiticus in foods indicates the possibility of aflatoxin contamination. In connection with their studies on aflatoxin contamination of grains, Bothast and Fennell (1974) developed a diagnostic medium which could be used to rapidly differentiate between members of the A. flavus group and other common storage fungi. The medium contains ferric citrate which promotes the formation of a persistent bright yellow-orange reverse pigmen- tation of colonies. The medium (Aspergillus differential medium, ADM) also

235
effectively distinguishes between members of the A. flavus group and other Aspergillus species of interest in medical mycology (Salkin and Gordon, 1975). Noting that supplementation of rose bengal streptomycin agar with botran (2,6-di- chloro-4-nitroaniline) facilitated the isolation and enumeration of A. flavus from peanuts (Bell and Crawford, 1967), ADM was modified by Hamsa and Ayres (1977) to yield a medium with greater selectivity.
Hara et al. (1974) developed a medium to simplify the screening of large numbers of A. flavus isolates for aflatoxin production. Detection of aflatoxin-posi- tive strains utilized ultraviolet-induced fluorescence of aflatoxin produced in a Czapek's solution agar containing corn steep liquor, HgC12 and (NH4)HzPO 4 instead of NaNO 3. Pitt et al. (1983) developed a selective medium for enumerating A. flavus and A. parasiticus based on the medium of Bothast and Fennell (1974). Results can be obtained after 42 h incubation at 30~ making the medium suitable for use in industrial quality control laboratories. Results were reproducible and comparable with those obtained using standard fungal enumeration media incu- bated for much longer periods. A very low percentage of false positives or negatives was reported. Aspergillus flavus and parasiticus agar (AFPA) is inclu- ded in this volume.
A simple method for screening aflatoxin-producing A. flavus and A. parasiticus using ultraviolet photography has been developed (Yabe et al., 1987). In ultraviolet photographs of colonies, aflatoxin producers appear as grey or black colonies, whereas non-producers appear as white colonies. The technique may be useful as a simple, safe and rapid method of screening aflatoxin-producing moulds.
A selective medium was developed by Frisvad (1983, 1986) to screen for Penicillium viridicatum and Penicillium verruculosum as an aid in the examination of stored cereal products for toxic metabolites. The medium, pentachloronitroben- zene rose bengal yeast extract sucrose (PRYES) agar, contains 15% sucrose, chloramphenicol and chlortetracycline (50 mg/1), rose bengal (25 mg/1) and pentachloronitrobenzene (100 mg/1) as selective agents. Members of the Muco- rales are completely inhibited, allowing important storage moulds to grow. Ochra- toxin A and citrinin producers in the P. viridicatum group II appear as violet brown reverse on PRYES agar, whereas producers of xanthomegnin and viomellein (P. viridicatum group I and P. aurantiogriseum) are indicated by their yellow reverse and obverse colours. The medium appears to be quite suitable for screen- ing for nephrotoxin-producing fungi in cereals.
Dichloran rose bengal yeast extract sucrose (DRYES) agar was originally formulated as a screening medium to detect Penicillium verrucosum, but may be used for a more general determination of toxigenic Penicillium and Aspergillus species (Frisvad et al., 1990a). They recommend the use of both DG18 agar and DRYES agar if populations and identity of fungi and their mycotoxin production is important in the examination of dried products.
Three media have been developed for selective isolation and direct identifica- tion of various groups of moulds associated with cereal products, meat and cheese (Frisvad et al., 1990b). Acetic acid dichloran yeast extract sucrose (ADYES) agar is useful for selective and indicative isolation of moulds spoiling acid-preserved

236
bread. Roquefortine C and PR-toxin produced by Penicillium roqueforti var. roqueforti on ADYES agar can be detected by an agar plug method. Dichloran creatine (DC) agar is an effective screening medium for penicillia associated with meat, cheese, nuts and other lipid and /o r protein foods. The third medium, dichloran yeast extract sucrose (DYES) agar, was developed for qualitative deter- mination of toxigenic penicillia and aspergilli.
Creatine sucrose agar, containing creatine as a sole nitrogen source and bromocresol purple as a pH indicator, is reported to be a good medium to differentiate subgenera of penicillia and to subdivide taxa and subgroups based on mycotoxin production into approximately two equal groups (Frisvad, 1985).
Andrews and Pitt (1986) developed dichloran chloramphenicol peptone agar (DCPA), a selective medium for isolating Fusarium species and some dematia- ceous hyphomycetes (e.g., Alternaria spp.) from cereals. The medium was shown to select against species of Aspergillus, Penicillium, Cladosporium and mucoraceous fungi. Fusarium species produced well-formed colonies with good conidial produc- tion, permitting rapid identification. The medium can also be used for the identification of Fusarium species (Hocking and Andrews, 1987). Conner (1990) reported that DCPA and DCPA supplemented with 0.5 txg/ml crystal violet were effective in suppressing Aspergillus and Penicillium while allowing selective growth of Fusarium species.
The possibility of using the growth response of Fusarium species on tannin sucrose agar as an aid to identification was investigated by Thrane (1986). Of the eleven Fusarium species investigated, eight were able to grow on tannin sucrose agar. The difference in growth response proved to be an additional character usable for identification of species. A selective medium for Fusarium species was developed using Czapek-Dox agar containing iprodione [3-(3,5-dichlorophenyl)-N- (1-methyl-ethyl)-2,4-dioxo-l-imidazolidine-carboxamide] (3 mg/1) and dichloran (2 mg/1) (Abildgren et al., 1987). This medium (CZID agar) is selective against numerous species of Alternaria, Epicoccum, Penicillium and mucoraceous fungi. Fusaria produce large, easily recognizable colonies on CZID agar, facilitating isolation and subculture.
Andrews (1990) evaluated media for differentiation of potentially toxigenic Alternaria alternata from the Alternaria state of Pleospora infectoria. The two species can be differentiated on dichloran chloramphenicol peptone agar (DCPA) but are more readily differentiated on dichloran chloramphenicol malt agar (DCMA).
Selective media for yeasts Relatively few media are available specifically for enumerating or at least
facilitating the growth of yeasts at the expense of moulds and bacteria. However, since yeasts may be the predominant microflora in some foods, e.g., confectionery products and fruit juice concentrates, special media for enumeration of yeasts are desirable.
Oxytetracycline glucose yeast extract (OGYE) agar (Mossel et al., 1970; Baird et a|., 1987) has been widely used for many years, especially in European countries, as

237
a general medium for enumerating yeasts. The medium loses its bacteriostatic effect if incubated at temperatures greater than 25~ Moulds will also develop colonies on OGYE agar. For selective recovery of psychrotrophic yeasts from chilled proteinaceous foods, oxytetracycline and gentamicin seem to be the bacte- riostatic combination of choice (Dijkmann et al., 1979). Tryptone glucose yeast extract (TGY) agar supplemented with antibiotics can also be used successfully for products containing high populations of yeasts. Davenport (1980) outlined a guide to media and methods for studying yeasts and yeast-like organisms in foods.
Media have also been developed for enumerating yeasts in specific food products. For example, Kish et al. (1983) formulated a selective medium for wine yeasts. The medium, containing 150 mg/1 bisulphite and 12% (by volume) ethanol, was suitable for the enumeration of wine yeasts when present at low populations in the natural microflora during early stages of grape juice fermentation. In the brewing industry, lysine agar (Walters and Thiselton, 1953; Lin, 1975), Schwarz differential medium, and other selective media (Lin, 1973) are used to detect wild yeasts. Heard and Fleet (1986) evaluated selective media for enumeration of yeasts during wine fermentation. Colonies of Saccharomyces cerevisiae dominated on malt extract agar and sometimes masked the presence of other genera. Lysine agar suppressed the growth of S. cerevisiae and enabled the enumeration of non-Sac- charomyces species such as Kloeckera apiculata, Candida stellata and Saccharomy- codes ludwigii. Growth of non-Saccharomyces yeasts on ethanol sulphite agar was variable.
Molybdate agar fortified with 0.125% calcium propionate was successfully used by Rale and Vakil (1984) to selectively isolate and identify yeasts from tropical fruits. This medium requires a longer incubation period for full expression of the natural pigmentation of yeasts.
Plating technique
The spread-plate technique is strongly preferred to the pour-plate technique for enumeration of yeasts and moulds in foods using dilution plating. Spread plating avoids any risk of thermal inactivation of fungal propagules which may be associ- ated with the pour-plate technique and facilitates maximum exposure of cells to atmospheric oxygen, thus allowing sporulation to proceed unencumbered in those instances when identification of mycoflora is desired.
Media should be prepared and poured (ca. 20 ml per plate) 16-40 h in advance of use to facilitate 'drying' of the surface. Generally, 0.1 ml of appropriately diluted sample is deposited in duplicate or triplicate on the surface of media and then spread evenly over the surface using a sterile bent glass rod. Rods should not exceed 2 mm in diameter in order to minimize the amount of sample adhering to them at the end of the spreading procedure. Larger samples, e.g., up to 0.3 ml of a 10-1 dilution of sample, can be spread on each plate to facilitate enumeration of low populations of yeasts and moulds.

238
Incubation
Media used for general enumeration of yeasts and moulds should be incubated at 25~ for 4 or, preferably, 5 days before colonies are counted. Higher tempera- tures, e.g., 40-45~ must be used for thermophiles whereas incubation at 5-15~ is needed for psychrotrophic fungi. The incubation time necessary before counting colonies of specific genera, species or groups of fungi varies. For example, accurate enumeration of moderate xerophiles on DG18 agar may require 6-8 days incuba- tion and mycelial development by fastidious xerophiles may require 4 weeks or longer.
Petri plates should be incubated in an upright position and should not be disturbed until colonies are ready to count. Movement of plates can result in release of mould conidia or spores and subsequent development of satellite colonies which would give an overestimation of population in the test sample. If spreading moulds are a problem, one may be forced to count colonies after 3 and again after 5 days of incubation.
No attempt should be made to select plates containing 30-300 colonies for counting. Rather, select dilutions giving 10-100 colonies on 9-cm plates or 20-200 colonies on 14-cm plates. Several colonies should be picked from countable plates periodically and examined microscopically by wet mount to ensure that bacteria are not growing. Development of bacterial colonies is more likely to occur on acidified media than on antibiotic-supplemented media.
Non-linearity of counts from dilution plating often occurs, i.e., 10-fold dilutions of samples often do not result in 10-fold reductions in numbers of colonies recovered on plating media. This has been attributed to fragmentation of mycelia and breaking of spore clumps during dilution in addition to competitive inhibition when large numbers of colonies are present on plates (Jarvis et al., 1983).
Interpretation of data
Unprocessed fruits, vegetables, grains and nuts may harbour large populations of yeasts and /o r moulds without apparent spoilage. Counts exceeding 106 C F U / g are not uncommon. The same populations in processed ingredients or finished products may indicate spoilage. The significance of high populations in either unprocessed or processed foods and food ingredients can only be assessed by reference to the history of the product and its intended treatment before it reaches the consumer.
Results from direct plating reveal the extent of internal infection of samples and therefore may give an indication of the environmental conditions under which the product was stored prior to analysis. The major genera of moulds detected using the direct plating techniques may also give some indication concerning time at which infection occurred, i.e., in the field or during storage. The presence of potentially mycotoxigenic moulds in the internal areas of grains or nuts, however, is not evidence that mycotoxins are present. Conversely, the absence or low

239
incidence of mycotoxigenic moulds is not evidence that the product does not contain mycotoxins.
Precautions
Mycological media should be moni tored for performance by comparing recovery of yeasts and moulds as well as unwanted bacterial colonies. Seiler (1985) con- cluded that the Miles-Misra method (Corry, 1982b) can be used for assessing recovery of certain yeasts and for ensuring that bacteria are adequately inhibited. The method is not suitable for use with the more rapidly spreading yeasts and moulds which require a stab inoculation method and measurement of rate of colony diameter increase with incubation time. Strains of microorganisms selected for monitoring performance of media should exhibit stable characteristics which do not change after repea ted subculturing. A range of microorganisms represent ing rapid- and slow-growing yeasts, moulds and bacteria should be included. Microor- ganisms likely to be present in the food under study should be selected for the evaluation.
A proport ion of mould and yeast cells in any given food being analysed may be metabolically or structurally injured as a result of physical or chemical stress (Beuchat, 1984). Such cells require opt imum nutrient, pH, osmotic and tempera- ture conditions for recovery if subsequent repair and colony formation are to occur on enumera t ion media.
Infections, intoxications and respiratory problems as a result of inhaling fungal spores can occur in laboratory personnel working with fungi. It is important to not remove lids from Petri plates containing colonies of moulds unless absolutely necessary for purposes of identification. All plates on which colonies have devel- oped should be properly enclosed and heat sterilized before discarding. Anyone who has mishandled moulds in a laboratory will be r emembered by those who work in that laboratory for many years to come.
References
Abildgren, M.P., Lund, F., Thrane, U. and Elmholt, S. (1987) Czapek-Dox agar containing iprodione and dichloran as a selective medium for the isolation of Fusarium species. Lett. Appl. Micriobiol. 5, 83-86.
Andrews, S. (1986) Optimization of conditions for the surface disinfection of sorghum and sultanas using sodium hypochlorite solutions. In: A.D. King, J.I. Pitt, L.R. Beuchat and J.E.L. Corry (Eds.), Methods for Mycological Examination of Foods. NATO ASI Series A: Life Sciences, Vol 122. Plenum Press, New York. pp. 28-32.
Andrews, S. (1990) Differentiation of Alternaria species isolated from cereals on a dichloran malt extract agar. International Workshop on Standardization of Methods for the Mycological Examina- tion of Foods, Progr. and Abstracts. Baarn, The Netherlands. p. 25.
Andrews, S. and Pitt, J.I. (1986) Selective medium for isolation of Fusarium species and dematiaceous hyphomycetes from cereals. Appl. Environ. Microbiol. 51, 1235-1238.

240
Baird, R.M., Corry, J.E.L. and Curtis, G.D.W. (Eds.) (1987) Pharmacopoeia of Cultive Media for Food Microbiology. Int. J. Food Microbiol. 5, 187-300.
Banks, J.G., Board, R.G., Carger, J. and Dodge, A.D. (1985) The cytotoxic and photodynamic inactivation of microorganisms by rose bengal. J. Appl. Bacteriol. 58, 392-400.
Bell, D.K. and Crawford, J.L. (1967) A botran-amended medium for isolating Aspergillus flavus from peanuts and soil. Phytopathology 57, 939-941.
Beuchat, L.R. (1984) injury and repair of yeasts and moulds. In: Revival of Injured Microbes, M.H.E. Andrews and A.D. Russell (Eds.), Academic Press, London. pp. 293-308.
Beuchat, L.R. (1987) Influence of organic acids on the heat resistance characteristics of Talaromyces flavus ascospores. Int. J. Food Microbiol. 6, 97-105.
Beuchat, L.R. and Hocking, A.D. (1990) Some considerations when analysing foods for the presence of xerophilic fungi. J. Food Prot. 53, 984-989.
Beuchat, L.R., Nail, B.V., Brackett, R.E. and Fox, T.L. (1990) Evaluation of a culture film (PetrifilmTMyM) methods for enumerating yeasts and molds in dairy and high-acid foods. J. Food Prot. 53, 864, 869-874.
Beuchat, L.R., Nail, B.V., Brackett, R.E. and Fox, T.L. (1991) Comparison of the Petrifilm T M yeast and mold culture film method to conventional methods for enumerating yeasts and molds in foods. J. Food Prot. 54, 443-447.
Beuchat, L.R. and Pitt, J.I. (1992) Detection and enumeration of heat resistant molds. In: C. Vanderzant and D. Splettstoesser (Eds.), Compendium of Methods for the Microbiological Exami- nation of Foods. Am. Public Health Assoc., Washington, DC, pp. 239-249.
Beuchat, L.R. and Rice, S.L. (1979) Byssochamys spp. and their importance in processed fruits. Advan. Food Res. 25, 237-288.
Beuchat, L.R. and Tapia de Daza, M.S. (1992) Evaluation of chemicals for restricting colony spreading by a xerophilic mold, Eurotium amstelodami, on dichloran-18% glycerol agar. Appl. Environ. Microbiol. 58, 2093-2095.
Booth, C. (1971a) Fungal culture media. In: C. Booth, (Ed.) Methods in Microbiology, Vol. 4., Academic Press, London. pp. 50-93.
Booth, C. (1971b) introduction to general methods. In: C. Booth (Ed.), Methods in Microbiology, Vol. 4. Academic Press, London.
Bothast, R.J. and Fennell, D.I. (1974) A medium for rapid identification and enumeration of As- pergillus flavus and related organisms. Mycologia 66, 365-369.
Brodsky, M.N., Entis, P., Entis, M.P., Sharpe, A.N. and Jarvis, G.A. (1982) Determination of aerobic plate and yeast mold counts in foods using an automated hydrophobic grid-membrane filter technique. J. Food. Prot. 45, 301-304.
Conner, D.E. (1990) Evaluation of methods for selective enumeration of Fusarium spp. in foodstuffs. International Workshop on Standardization of Methods for the Mycological Examination of Foods, Progr. and Abstracts. Baarn, The Netherlands. p. 26.
Corry, J.E.L. (1982a) Assessment of the selectivity and productivity of media used in analytical mycology. Arch. Lebensm. Hyg. 33, 160-164.
Corry, J.E.L. (1982b) Quality assessment of culture media by the Miles-Misra method. In: J.E.L. Corry (Ed.), Quality Assurance and Quality Control in Microbiological Culture Media. G.I.T.-Verlag, Darmstadt, pp. 21-37.
Curtis, G.D.W. and Baird, R.M. (Eds.) (1993) Pharmacopoeia of cultive media for food microbiology: additional monographs (2). Int. J. Food Microbiol. 17, in press.
Davenport, R.R. (1980) An outline to media and methods for studying yeasts and yeast-like organisms. In: F.A. Skinner, S.M. Passmore and R.R. Davenport (Eds.), Biology and Activities of Yeasts. Soc. Appl. Bacteriol. Series No. 9, Academic Press, London, pp. 261-278.
Dijkmann, K.E., Koopmans, J. and Mossel, D.A.A. (1979) The recovery and identification of pschrotrophic yeasts from chilled and frozen comminuted fresh meats. J. Appl. Bacteriol. 47, ix.
Frisvad, J.C. (1983) A selective and indicative medium for groups of Penicillium viridicatum producing different mycotoxins in cereals. J. Appl. Bacteriol. 54, 409-416.
Frisvad, J.C. (1985) Creatine sucrose agar, a differential medium for mycotoxin producing terverticillate Penicillium species. Lett. Appl Micriobiol. 1, 109-113.

241
Frisvad, J.D. (1986) Selective medium for Penicillium viridicatum in cereals. In: A.D. King, J.I. Pitt, L.R., Beuchat and J.E.L. Corry (Eds.), Methods for the Mycological Examination of Food, NATO ASI Series A: Life Sciences, Vol. 122. Plenum Publ. Co., New York, pp. 132-135.
Frisvad, J.C., Filtenborg, O. and Thrane, U. (1990a) Collaborative study on media for detecting and enumerating toxigenic fungi. 2nd International Workshop on the Standardization of Methods for the Mycological Examination of Foods, Progr. and Abstracts. Baarn, The Netherlands, p. 27.
Frisvad, J.C., Lund, F., Filtenborg, O. and Thrane, U. (1990b) Selective media for the detection of the associated toxigenic mycoflora of cereal products, meat and cheese. 2nd International Workshop on the Standardization of Methods for the Mycological Examination of Foods, Progr. and Abstracts. Baarn, The Netherlands. pp. 27-28.
Hamsa, T.A.P. and Ayres, J.C. (1977) A differential medium for the isolation of Aspergillus flavus from cottonseed. J. Food Sci. 42, 449-453.
Hara, S., Fennell, D.I. and Hesseltine, C.W. (1974) Aflatoxin-producing strains of Aspergillus flavus detected by fluorescence of agar medium under ultraviolet light. Appl. Microbiol. 27, 1118-1123.
Hastings, J.W., Tsai, W.Y.J. and Bullerman, L.B. (1986) Comparison of stomaching versus blending in sample preparation for mold enumeration. In: A.D. King, J.I. Pitt, L.R. Beuchat and J.E.L. Corry (Eds.), Methods for Mycological Examination of Foods, NATO ASI Series A: Life Sciences, Vol. 122, Plenum Press, New York, pp. 7-9.
Heard, G.M. and Fleet, G.H. (1986) Evaluation of selective media for enumeration of yeasts during wine fermentation. J. Appl. Bacteriol. 60, 477-481.
Hocking, A.D. and Andrews, S. (1987) Dichloran chloramphenicol peptone agar as an identification medium for Fusarium species and some dematiaceous hyphomycetes. Trans. Br. Mycol. Soc. 89, 239-244.
Hocking, A.D. and Pitt, J.I. (1980) Dichloran glycerol medium for enumeration of xerophilic fungi from low-moisture foods. Appl. Environ. Microbiol. 39, 488-492.
International Commission on Microbiological Specification for Foods. (1986). Microorganisms in Foods. 2. Sampling for Microbiological Analysis: Principles and Specific Applications. University of Toronto Press, Toronto.
Jarvis, B. (1973) Comparison of an improved rose-bengal-chlortetracycline agar with other media for the selective isolation and enumeration of moulds and yeasts in food. J. Appl. Bacteriol. 36, 723-727.
Jarvis, B., Seiler, D.A.L., Ould, A.J.L. and Williams, A.P. (1983) Observations on the enumeration of moulds in food and feeding stuffs. J. Appl. Bacteriol. 55, 325-336.
Jarvis, B. and Williams, A.P. (1987) Methods for detecting fungi in foods and beverages. In: L.R. Beuchat (Ed.), Food and Beverage Mycology, Van Nostrand Reinhold, New York, pp. 599-636.
Jermini, M.F.G., Geiges, O. and Schmidt-Lorenz, W. (1987) Detection isolation and identification of osmotolerant yeasts from high-sugar products. J. Food Prot. 50, 468-472.
Katan, T. (1985) Heat activation of dormant ascospores of Talaromyces flavus. Trans. Br. Mycol. Soc. 84, 748-750.
King, A.D., Hocking, A.D. and Pitt, J.I. (1979) Dichloran rose bengal medium for enumeration and isolation of molds from foods. Appl. Environ. Microbiol. 37, 959-964.
King, A.D., Pitt, J.I., Beuchat, L.R. and Corry, J.E.L. (Eds.) (1986) Methods for the Mycological Examination of Food. NATO ASI Series A, Life Sciences, Vol. 122. Plenum Press, New York.
Kish, S., Shaft, R. and Margalith, P. (1983) A note on a selective medium for the wine yeasts. J. Appl. Bacteriol. 55, 177-179.
Koburger, J.A. and Marth, E.H. (1984) Yeasts and molds. In: M. Speck (Ed.), Compendium of Methods for the Microbiological Examination of Foods, Am. Public Health Assoc. Washington, DC, pp. 197-202.
Lin, Y. (1973) Detection of wild yeasts in the brewery: a new criterion. Brewers Digest 48, 60-69. Lin, Y. (1975) Detection of wild yeasts in the brewery: efficiency of differential media. J. Inst. Brew. 81,
410-417. Mislivec, P.B. and Stack, M.E. (1984) Enumeration of yeasts and molds and production of toxins. In:
Bacteriological Analytical Manual, 6th Edn. US Food and Drug Administration, Washington, DC. Mossel, D.A.A., Visser, M. and Mengerink, W.H.J. (1962) A comparison of media for the enumeration
of moulds and yeasts for foods and vegetables. Lab. Prac. 11, 109-112.

242
Mossel, D.A.A., Kleynen-Semmeling, A.M.C., Vincentie, H.M., Beerens, H. and Catsavas, M. (1970) Oxytetracycline glucose yeast extract agar for selective enumeration of moulds and yeasts in foods and clinical material. J. Appl. Bacteriol. 33, 454-457.
Pitt, J.I. (1979) The Genus Penicillium and its Teleomorphic States Eupenicillium and Talaromyces. Academic Press, London.
Pitt, J.I. (1986) Properties of the ideal enumeration medium. In: A.D. King, J.I. Pitt, L.R. Beuchat and J.E.L. Corry (Eds.), Methods for the Mycological Examination of Food, NATO ASI Series A: Life Sciences, Vol. 122. Plenum Publ. Co., New York, pp. 132-132.
Pitt, J.I. and Hocking, A.D. (1977) influence of solute and hydrogen ion concentration on the water relations of some xerophilic fungi. J. Gen. Microbiol. 101, 35-40.
Pitt, J.I. and Hocking, A.D. (1985a) Fungi and Food Spoilage. Academic Press, Sydney. Pitt, J.I. and Hocking, A.D. (1985b) New species of fungi from Indonesian dried fish. Mycotaxon 22,
197-208. Pitt, J.I., Hocking, A.D. and Glenn, D.R. (1983) An improved medium for the detection of Aspergillus
flauus and Aspergillus parasiticus. J. Appl. Bacteriol. 54, 109-114. Rale, V.B. and Vakil, J.R. (1984) A note on an improved molybdate agar for the selective isolation of
yeasts from tropical fruits. J. Appl. Bacteriol. 56, 409-4)3. Restaino, L., Bills, S. and Lenovich, L.M. (1985) Growth response of an osmotolerant, sorbate-resistant
Saccharomyces rouxii strain: evaluation of plating media. J. Food Prot. 48, 207-209. Salkin, I.F. and Gordon, M.A. (1975) Evaluation of Aspergillus differential medium. J. Clin. Microbiol.
2, 74-75. Samson, R.A. and van Reenen-Hoekstra, E.S. (1988) Introduction to Food-Borne Fungi. 3rd Edn.
Centraalbureau voor Schimmelculturees. Baarn, The Netherlands. Scarr, M.P. (1959) Selective media used in the microbiological examination of sugar products. J. Sci.
Food Agric. 10, 678-681. Seiler, D.A.L. (1985) Monitoring mycological media. Int. J. Food Microbiol. 2, 123-131. Seiler, D.A.L. (1986) Effect of presoaking on recovery of fungi from cereals and cereal products. In:
A.D. King, J.I. Pitt, L.R. Beuchat and J.E.L. Corry (Eds.), Methods for the Mycological Examina- tion of Food. NATO ASI Series A., Life Sciences, Vol. 122. Plenum Press, New York. pp. 26-28.
Splittstoesser, D.F., Wilkison, M. and Harrison, N. (1972) Heat activation of Byssochlamys fulua ascospores. J. Milk Food Technol. 35, 399-401.
Tapia de Daza, M.S. and Beuchat, L.R. (1992) Suitability of modified dichloran glycerol (DG18) agar for enumerating unstressed and temperature-stressed xerophilic molds. Food Microbiol. 10, in press.
Thrane, U. (1986) The ability of common Fusarium species to grow on tannin-sucrose agar. Lett. Appl. Microbiol. 2, 33-35.
Waiters, L.S. and Thiselton, M.A. (1953) Utilization of lysine by yeasts. J. Inst. Brew. 59, 401-404. Yabe, K., Ando, Y., Ito, M. and Terakado, N. (1987) Simple method for screening aflatoxin-producing
molds by UV photography. Appl. Environ. Microbiol. 53, 230-234.

Part 2
Pharmacopoeia of Culture Media

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
243
Notes on the use of the monographs
Composition: Under this heading will be found all the ingredients of the medium in the concentrations present in the finished product. Where salts are available in both anhydrous and hydrated forms the quantities given refer to the anhydrous form unless otherwise stated. The quantity of agar specified assumes the use of a product that produces a satisfactory gel in nutrient broth when used at a concen- tration of 15 g/1. Local circumstances may require variations in this quantity depending on the gelling properties of the agar used. In media such as semi-solid agars where the gel strength is critical, the specification of the agar should be followed exactly.
Bile and its derivatives are subject to considerable variation and where these substances are specified in these monographs they should be tested in trial batches of media before general use.
Preparation: For optimal performance media should be prepared as directed. With some media it is possible to prepare and sterilize basal ingredients for subsequent re-melting and addition of other ingredients to produce the final product. Whilst this practice may be convenient, particularly in small laboratories, in general the re-heating of media should be avoided.
Autoclaving is sometimes undesirable and often unnecessary in the preparation of media. The total heat input is difficult to standardize in general purpose laboratory autoclaves and more consistent results are obtained by the use of a purpose-built medium sterilizer (Corry et al., 1986).
Physical properties: pH values given are for readings made at 20-25~ unless otherwise stated.
No values have been given for gel strength measurements. Several machines are available which can be used to measure this property but none is in general use. Costin (1978) gave details of a method using the Gelomat R machine (Heinrich Bareiss Apparatebau Company, 7931 Oberdischingen, Germany) and stated that satisfactory batches of media should have a gel rigidity of between 50 and 80 g. Users of the Van der Bijl penetrometer regard a penetration of less than 2 mm within 10 min as satisfactory but, whilst this ensures a minimum strength of gel, it

244
provides no indication of an excessively stiff matrix. Other machines which may be used to measure gel strengths are the Gelometer (Marine-Colloids Inc., 2 Edison Place, Springfield, NJ 07081, U.S.A.) and the LFRA Texture Analyser (C.S. Stevens & Son Ltd., Unit 4, Executive Park, Hatfield Road, St. Albans, Herts, ALl 4TA, U.K.) but no standards have been published for these machines.
Shelf life: Dehydrated media and dry ingredients should be stored according to the manufacturer's instructions and with regard to the stated shelf life or expiry date. Some ingredients, particularly dyes, undergo changes when exposed to light which result in the formation of substances toxic to microorganisms or reduce their selectivity. For this reason the shelf life given for ready to use media applies to poured plates or plugged tubes stored in the dark. Shelf life can be maximized by packing poured plates in sealed bags and tubed media in screw capped containers to reduce the rate of water loss due to evaporation.
Resuscitation of stressed organisms: Test strains will not normally have been stressed (i.e. exposed to sublethal heating, drying, chilling, acid) but when using these media for samples where stressing may have occurred appropriate resuscita- tion procedures should be carried out. For a review of this problem see Mossel and van Netten (1984) and Ray (1986).
Drying of poured plates: Facilities for drying vary from laboratory to laboratory. It is not possible therefore to recommend a single procedure to obtain a uniform dryness of agar surfaces. Where local production methods are consistent it should be possible to adopt a standardized method to give uniform results. Over-drying should be avoided.
Incubation temperatures: Strict adherence to the stated incubation temperature is recommended as variations in temperature can affect the selectivity of many media. Monitoring of incubator temperatures, preferably by means of a thermo- graph with a sensor in the medium, will provide warnings of inconsistencies in performance. Loading with many plates in high stacks or the use of high sided non-perforated trays can result in extremely long delays in achieving the desired temperature.
Reading of results and interpretation: Many of the media described in these monographs are designed for the isolation of a single genus or species. Very few however are entirely specific and the identity of characteristic colonies should be regarded as 'presumptive' until confirmatory tests for identity have been carried out.
Quality assessment: Test strains. For each medium a minimum set of test strains is given which should be used on all batches of media prepared in the laboratory. Where other strains are known to perform satisfactorily with a given medium these are also listed under the heading of Supplementary Strains. These extra strains can

245
be used for more extensive testing undertaken when assessing new batches of dehydrate from manufacturers or when investigating problems. The occasional use of freshly isolated 'wild' strains may also prove valuable.
Where strains of Proteus sp. are used for quantitative testing of agars, a refer- ence medium on which the organism does not swarm should be used. Alternatively the ability of a test medium to prevent swarming of Proteus sp. may be checked qualitatively and another organism (e.g. Morganella morganii) used in quantitative tests.
Strains are identified by their numbers in the National Collection of Industrial and Marine Bacteria (NCIMB) Quality Control catalogue. Cross references to numbers in other collections will be found in Appendix II. Those strains which are not available from NCIMB are listed in each monograph with the appropriate number(s) in other collections.
Methods: Recommended methods are given in Appendix I.
Criteria: Unless otherwise stated in individual monographs the criteria for assess- ment of prepared media are as follows. For the modified Miles-Misra and dilution to extinction methods, recovery of all strains on non-selective media should be within 0.7 lOgl0 of the recovery of the same strains on the reference medium. When ecometry is used the Absolute Growth Index (AGI) should be at least 3 and an equivalent score should be obtained with the streaking method.
For unwanted strains on selective media recovery by modified Miles-Misra or dilution to extinction methods should be more than 5 lOgl0 below the recovery with the reference medium. For wanted strains recovery should be less than 1 lOgl0 below that with the reference medium. Ecometry should yield AGIs not greater than 2.0 for unwanted strains and not less than 2.5 for wanted strains. Equivalent scores should be obtained with the streaking method. The reference medium, unless otherwise stated, is tryptone soya agar or broth. Standards for mycological media are given in individual monographs.
The quality assessment procedures detailed in these monographs relate primar- ily to productivity, selectivity and colonial appearance. With some media other methods may also be appropriate and these have been reviewed by Curtis (1985).
References
Corry, J.E.L., Leclerc, M., Mossel, D.A.A., Skovgaard, N., Terplan, G. and van Netten, P. (1986) An investigation into the quality of media prepared and poured by an automatic system. Int. J. Food Microbial. 3, 109-120.
Costin, I.D. (1978) A semiautomatic instrument for the determination of gel rigidity in microbiological nutrients and gelling agents. In: Mechanizing microbiology, edited by A.N. Sharpe and D.S. Clark, Charles C. Thomas, Springfield, IL, pp. 170-196.
Curtis, G.D.W. (1985) A review of methods for quality control of culture media. Int. J. Food Microbiol. 2, 13-20.

246
Mossel, D.A.A. and van Netten, P. (1985) Harmful effects of selective media on stressed microorgan- isms: nature and remedies. In: The revival of injured microorganisms, edited by M.H.E. Andrew and A.D. Russell, Academic Press, London, pp. 329-369.
Ray, B. (1986) Impact of bacterial injury and repair in food microbiology: its past, present and future. J. Food Protect. 49, 651-655.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
247
Summary of organisms and recommended media
Bacillus cereus
Brochothrix thermosphacta
Campylobacter spp.
Carnobacterium spp.
Clostridium perfringens
Coli-aerogenes group (see also Enterobacteriaceae)
Enterobacteriaceae
Enterococcus spp.
Escherichia coli
Lactic acid bacteria
Mannitol egg yolk polymyxin agar Polymyxin pyruvate egg yolk mannitol bromo- thymol blue agar
Streptomycin thallous acetate actidione agar
Charcoal cefoperazone deoxycholate agar Charcoal cefoperazone deoxycholate broth Preston campylobacter selective agar Preston enrichment broth Skirrow campylobacter selective agar
Cresol red thallium acetate sucrose agar
Differential reinforced clostridial agar (modified) Iron sulphite agar Oleandomycin polymyxin sulphadiazine perfringens agar Rapid perfringens medium Sulphite cycloserine azide agar Tryptose sulphite cycloserine agar
Brilliant green bile broth Lauryl tryptose broth
Enterobacteriaceae enrichment broth Violet red bile glucose agar Violet red bile agar
Citrate azide tween carbonate agar Kanamycin aesculin azide agar M-enterococcus agar
Brilliant green bile broth Enterobacteriaceae enrichment broth Lauryl tryptose broth Tryptone bile agar Violet red bile glucose agar Violet red bile agar
All purpose tween agar-modified Briggs agar L-S differential agar M-17 agar de Man, Rogosa and Sharpe agar de Man, Rogosa and Sharpe agar with sorbic acid (pH 6.2)

248
Lactobacilli (see also Lactic acid bacteria)
Lactococcus lactis ssp. lactis
Leuconostocs
Listeria spp.
Moulds
Pediococci
Pseudomonas spp.
Salmonella spp. (see also Entero- bacteriaceae)
Shigella spp. (see also Enterobac- teriaceae)
Briggs agar Lactobacillus sorbic acid agar de Man, Rogosa and Sharpe agar Rogosa agar
Briggs agar de Man, Rogosa and Sharpe agar de Man, Rogosa and Sharpe agar with sorbic acid Rogosa agar (pH 6.2)
see Lactic acid bacteria
Dominguez Rodriguez LSAMm agar FDA listeria enrichment broth Fraser broth Levinthal broth with acriflavine and nalidixic acid Lithium chloride ceftazidime tween 80 agar Lithium chloride phenylethanol moxalactam agar Oxford agar Oxford agar-modified Polymyxin acriflavine lithium chloride ceftazidime aesculin mannitol agar Polymyxin acriflavine lithium chloride ceftazidime aesculin mannitol egg yolk broth University of Vermont broths I & II
Aspergillus flavus and parasiticus agar Dichloran glycerol (DG 18) agar Dichloran rose bengal chloramphenicol agar Oxytetracycline glucose yeast extract agar
Briggs agar de Man, Rogosa and Sharpe agar de Man, Rogosa and Sharpe agar with sorbic acid Rogosa agar (pH 6.2)
Cephaloridine fucidin cetrimide agar
Bismuth sulphite agar Diagnostic salmonella selective semisolid medium Hektoen enteric agar Lysine iron cystine neutral red broth Mannitol lysine crystal violet brilliant green agar Muller Kauffmann tetrathionate broth Phenol red brilliant green agar Rambach agar Rappaport medium-semisolid modification Rappaport-Vassiliadis broth Rappaport-Vassiliadis medium-semisolid modifi- cation Selenite cystine broth Xylose lysine deoxycholate agar
Violet red bile agar Xylose lysine deoxycholate agar

Staphylococcus aureus
Streptococcus bouis
Streptococcus thermophilus
Vibrio spp.
Yersinia spp. (see also Enterobac- teriaceae)
249
Baird-Parker agar Baird-Parker liquid medium Giolitti and Cantoni broth KRANEP agar Tryptone soya broth with 10% NaC1 & 1% sodium pyruvate
Kanamycin aesculin azide agar M-enterococcus agar
Briggs agar
Cellobiose polymyxin B colistin agar Thiosulphate citrate bile salts agar
Bile oxalate sorbose broth Cefsulodin irgasan novobiocin agar Irgasan ticarcillin chlorate broth Salmonella shigella deoxycholate calcium agar

This Page Intentionally Left Blank

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
251
All Purpose Tween (APT) agar-modified ( for H 20 2 detection)
Description and history
This medium was devised by Evans and Niven (1951) and a modification developed by Shipp (1964) for the rapid non-selective enumeration of bacteria capable of elaborating H 2 0 2 during growth under aerobic conditions. Such reaction on cooked cured meats results in a "green" discolouration. This principle was also used by Whittenbury (1964) and Reuter (1970).
Composition (grams)
Peptone (tryptic digest of casein) Yeast extract Glucose Sodium chloride Tri-sodium citrate Di-potassium hydrogen orthophosphate Sorbitan monooleate (Tween 80) Magnesium sulphate- 7H2 O Manganese (II) chloride �9 4 H 2 0 Iron (II) s u l p h a t e - 7 H 2 0 Thiamine hydrochloride Agar Distilled or deionized water
12.5 7.5
10.0 5.0 5.0 5.0 0.2 0.8 0.14 0.04 0.001
13.5 1000.0
Preparation
Double layer plates are prepared. Bottom layer: APT agar (15 ml). Top layer: 20 g of MnO 2 are added to 200 ml APT broth, dispensed in 10 ml amounts and sterilized by autoclaving at 121~ for 15 min. To each 100 ml amount of sterile, cooled (45~ APT agar is added 10 ml of MnO 2 suspension. When the bottom layer is firmly set a thin layer of APT agar with MnO~ is added.

252
Physical properties
Appearance
pH
Bottom layer: light brown, clear. Top layer: black, with the suspended MnO 2. 6.7 +0 .2
Shelf life
Ready to use medium At least 7 days at 4 + 2~
Inoculation method for samples
Sample dilutions (in 0.1% peptone + 0.85% NaC1) are surface plated to obtain discrete colonies.
Incubation method
At 30~ for a minimum of 5 days in air or 25~ for 5 days or 7~ for 10 days.
Reading of results and interpretation
H202 forms soluble compounds with suspended MnO 2. Colonies surrounded by a clear zone are regarded as potentially positive. These may be inoculated onto the surface of a sterile cooked, cured meat and after incubation in the dark at 20~ for 24-72 h, green discolouration should be apparent for a positive result.
Quality assessment
(i) Positive reaction Test strains
Inoculation method
Criteria
(ii) Negative reaction Test strain
Inoculation method
Criteria
Lactobacillus viridescens 50057 Lactobacillus brevis 50043
Broth cultures streaked for discrete colonies.
Growth with clear zones around colonies.
Lactobacillus sake 50056
Broth cultures streaked for discrete colonies.
Growth without clear zones around colonies.
(iii) Characteristic appearance of colonies
All H202 producing colonies are surrounded by a clear zone on the medium.
References
Evans, J.B. and Niven, C.F. (1951) Nutrition of the heterofermentative lactobacilli that cause greening of cured meat products. J. Bacteriol. 62, 599-603.

253
Reuter, G. (1970) Laktobazillen und eng v erwandte Mikroorganismen in Fleisch und Fleischerzeugnis- sen. 2. Mitteilung: Die Charakterisierung der isolierten Laktobazillenstamme. Fleischwirtschaft 50, 954-962.
Shipp, H.L. (1964) The green discolouration of cooked cured meats of bacterial origin. Technical circular No. 266, British Food Manufacturing Industries Research Association, Leatherhead, U.K.
Whittenbury, R. (1964) Hydrogen peroxide formation and catalase activity in the lactic acid bacteria. J. Gen. Microbiol. 35, 13-26.

254 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Aspergillus flauus and parasiticus agar (AFPA)
Description and history
This is a selective medium for the enumeration in foods of the mycotoxin producing fungi Aspergillus flauus and Aspergillus parasiticus. Bothast and Fennell (1974) developed Aspergillus differential medium (ADM), containing 1.0% yeast extract, 1.5% tryptone and 0.5% ferric chloride, recommending incubation at 28~ for 3 days. Hamsa and Ayres (1977) incorporated streptomycin and dichloran into ADM and recommended incubation at 28~ for 5 days. Both media relied upon the formation of a bright orange-yellow reverse pigment by Aspergillus flauus and related species. Pitt et al. (1983) further refined the formulation, producing Aspergillus flauus and parasiticus agar (AFPA), which gives sufficient colour development to enable recognition of Aspergillus flauus (or Aspergillus parasiticus) colonies within 42-48 h at 30~
Composition (grams)
Yeast extract Peptone (bacteriological) Iron (III) ammonium citrate Dichloran (2,6-dichloro-4-nitroaniline) Chloramphenicol Agar Distilled or deionized water
20.0 10.0
0.5 0.002 0.1
15.0 1000.0
Preparation
Dissolve yeast extract, peptone, iron (III) ammonium citrate and agar in the water by heating. Add 1.0 ml of a 0.2% (w/v) ethanolic solution of dichloran and 10 ml of a 1% ethanolic solution of chloramphenicol. Adjust to pH 6.2 + 0.2 and sterilize by autoclaving at 121~ for 15 min. Cool to 50~ and dispense 15 ml amounts into

255
9 cm diameter Petri dishes. Dry and use immediately or store at 4 _+ 2~ in the dark for up to 4 weeks before using.
Physical properties
Appearance Amber, clear. pH 6.2 +_ 0.2
Shelf life
Ready to use medium 4 weeks at 4 _+ 2~
Inoculation method for samples
Surface spread 0.1 or 0.2 ml of diluted sample per 9 cm diameter plate.
Incubation
At 30~ for 42-48 h.
Reading of results and interpretation
Aspergillus flavus and Aspergillus parasiticus produce an orange-yellow (chrome yellow) reverse colony pigmentation. Aspergillus niger sometimes produces colonies with a light yellow reverse, but is readily distinguished from Aspergillus flavus after further 24-48 h incubation by the production of black conidial heads. Aspergillus ochraceus produces an orange-yellow reverse, but only forms colonies after pro- longed incubation.
Quality assessment
Use a stab inoculation procedure. Examine reverse of colonies after 42-48 h incubation at 30~ for typical pigmentation (see above). If doubt exists over Aspergillus niger, continue incubation for a further 24-48 h. Prolonged incubation is not recommended.
Test strains CSIRO * A-NRRL + Aspergillus flavus 3084 3251 Aspergillus parasitieus 2744 2999 Aspergillus niger 2522 3361
* CSIRO Food Research Laboratory, PO Box 52, North Ryde, New South Wales, 2113, Australia. + USDA Northern Utilization Research and Development Division, Peoria, Illinois, U.S.A.

256
References
Bothast, R.J. and Fennell, D.I. (1974) A medium for rapid identification and enumeration of As- pergillus flauus and related organisms. Mycologia 66, 365-369.
Hamsa, T.A. and Ayres, J.C. (1977) A differential medium for the isolation of Aspergillus flavus from cottonseed. J. Food Sci. 42, 449-453.
Pitt, J.I., Hocking, A.D. and Glenn, D.R. (1983) An improved medium for the detection of Aspergillus flauus and A. parasiticus. J. Appl. Bacteriol. 54, 109-114.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
257
Baird-Parker agar
Description and history
This is a selective and diagnostic medium for the enumeration of Staphylococcus aureus in foods which was first reported by Baird-Parker in 1962. It is now widely recommended by national and international bodies. Selectivity is attained with potassium tellurite and lithium chloride and sulphamezathine is advised to sup- press Proteus spp. (Baird-Parker, 1969) The egg yolk clearing reaction is the diagnostic feature of characteristic colonies of Staphylococcus aureus. Sodium pyruvate is a critical component essential to both recovery of damaged Staphylo- coccus aureus and their subsequent growth (Baird-Parker and Davenport, 1965).
Composition (grams)
Tryptone 8.014 Beef extract 4.007 Yeast extract 0.801 Li th ium chlor ide . H 20 4.007 Sodium pyruvate 8.9 Glycine 11.2 Potass ium tellurite 0.89 Sulphameza th ine 0.089 Egg yolk emulsion 20% (ml) 44.5 Agar 17.8 Distil led or deionized water to 1000.0
In practice this medium is prepared as under.
Preparation
Basal Agar
Add tryptone 10 g, beef extract 5 g, yeast extract 1 g, lithium chloride 5 g and agar 20 g to 1 litre of distilled or deionized water, soak then heat to dissolve. Adjust pH to 7.2 and dispense in 90 ml amounts in 4 oz bottles and autoclave at 121~ for 15 min.

258
Complete Medium
Prepare sterilized (membrane-filtered) aqueous solutions of sodium pyruvate 20% w/v, glycine 20% w/v, potassium tellurite (Merck) 1% w/v, and sulphamezathine 0.2% w/v. It is important to use a satisfactory brand of potassium tellurite. Supplies from Merck and Sigma have been found adequate. The salt must be completely dissolved prior to filtration. The sulphamezathine solution is prepared by dissolving 0.5 g of pure sulphamezathine in 25 ml of N/10 NaOH and diluting to 250 ml with distilled water.
Melt and cool base to 50~ to each 90 ml add the above pre-warmed solutions: 20% sodium pyruvate solution 5 ml, 20% glycine solution 6.3 ml, 1% potassium tellurite solution 1 ml, 0.2% sulphamezathine solution 5 ml and egg yolk emulsion 5 ml. Mix and pour in approximately 12 ml amounts into 9 cm diameter Petri dishes.
Commercially available egg yolk emulsion or egg yolk emulsion/tellurite mix- ture may be used. Egg yolk emulsion prepared by the method of Billing and Luckhurst (1957), has also been used in international trials (Chopin et al., 1985).
Physical properties
Appearance pH
Complete medium opaque, cream/pale fawn in colour. Basal agar 6.9 _+ 0.1
Shelf life
Prepared basal medium
Complete medium Solutions
3 months at 20~ Commercial media containing sodium pyruvate may have a shorter shelf life. Use within 24 h of preparation. Glycine and potassium tellurite-6 months at 20~ Dis- card tellurite if a white precipitate forms. Sodium pyruvate and egg yolk emulsion- 1 month at 5~
Inoculation method for samples
Dry plates for minimal period at 45-50~ to remove surface moisture prior to use. Spread plate technique using 0.1 ml inocula on 9 cm diameter Petri dishes or 0.5 m! on 14 cm diameter dishes. Spread inoculum over the whole plate until surface appears dry.
Incubation method
At 37~ for 48 h in air.

259
Reading of results and interpretation
Plates should be first examined after 20-24 h incubation when Staphylococcus aureus appear as black shiny colonies 1.0-1.5 mm diameter with a very narrow colourless margin surrounded by clear zones extending into a still opaque medium. These are marked and counted. Plates are re-incubated for a further day and additional black shiny egg yolk clearing colonies identified. Confirmatory tests for coagulase production are required. Colonies typical of Staphylococcus aureus but without an egg yolk reaction should also be tested for coagulase and if positive confirmed by further tests (Devriese and Hajek, 1980). Egg yolk negative strains of Staphylococcus aureus may occur in some foods particularly those from dairy sources.
Quality assessment
(i) Productivity Test strains Staphylococcus aureus 50081
Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra onto Baird-Parker agar and Heart Infusion agar.
Criteria Counts on Baird-Parker agar should be within 0.5 lOg l0 of the counts on the non-selective medium. 50081 will show a reduction in count (> 0.5 log10) if the Baird-Parker agar containing sulphamezathine is deficient in pyruvate.
(ii) Selectivity Test strains Escherichia coli 50034
Proteus mirabilis (ATCC 29906/CECT 4168 /NCTC 11938)
Inoculation method As above.
Criteria Recovery on Baird-Parker agar should be 3.0 lOg l0 below the recovery on Heart Infusion agar.
(iii) Characteristic appearance of Staphylococcus aureus
Black shiny colonies up to 3 mm diameter (at 48 h) with coiourless margin and typically surrounded by an egg yolk clearing zone, the majority of strains subsequently producing an area of precipitate within the cleared zone.

260
References
Baird-Parker, A.C. (1962) An improved diagnostic and selective medium for isolating coagulase positive staphylococci. J. Appl. Bacteriol. 25, 12-19.
Baird-Parker, A.C. (1969) Isolation methods for microbiologists, edited by D.A. Shapton and G.W. Gould. SAB Technical Series No. 3. Academic Press, London, pp. 1-8.
Baird-Parker, A.C. and Davenport, E. (1965) The effect of recovery medium on the isolation of Staphylococcus aureus after heat treatment and after storage of frozen or dried cells. J. Appl. Bacteriol. 28, 390-402.
Billing, E. and Luckhurst, E.R. (1957) A simplified method for the preparation of egg yolk media. J. Appl. Bacteriol. 20, 90.
Chopin, A., Malcolm, S., Jarvis, G., Asperger, H., Beckers, H.J., Bertona, A.M., Cominazzini, C., Carini, S., Lodi, R., Hahn, G., Heeschen, W., Jans, J.A., Jervis, D.I., Lanier, J.M., O'Connor, F., Rea, M., Rossi, J., Seligmann, R., Tesone, S., Waes, G., Mocquot, G., and Pivnick, H. (1985) ICMSF Methods studies XV. Comparison of four media and methods for enumerating Staphylococcus aureus in powdered milk. J. Food Protect. 48, 21-27.
Devriese, L.A. and Hajek, V. (1980) Identification of pathogenic staphylococci isolated from animals and foods derived from animals. J. Appl. Bacteriol. 49, 1-11.
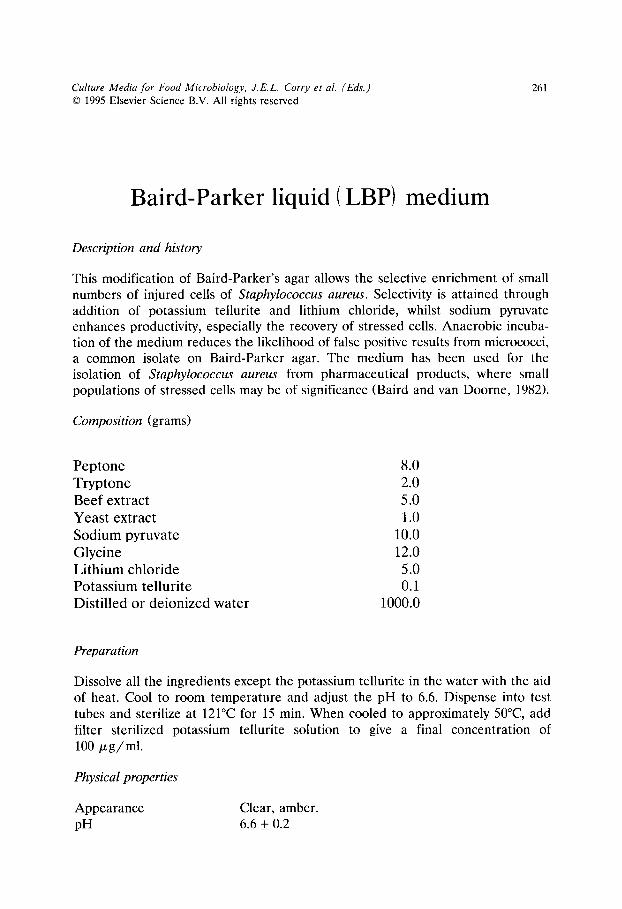
Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
261
Baird-Parker liquid (LBP) medium
Description and history
This modification of Baird-Parker's agar allows the selective enrichment of small numbers of injured cells of Staphylococcus aureus. Selectivity is attained through addition of potassium tellurite and lithium chloride, whilst sodium pyruvate enhances productivity, especially the recovery of stressed cells. Anaerobic incuba- tion of the medium reduces the likelihood of false positive results from micrococci, a common isolate on Baird-Parker agar. The medium has been used for the isolation of Staphylococcus aureus from pharmaceutical products, where small populations of stressed cells may be of significance (Baird and van Doorne, 1982).
Composition (grams)
Peptone 8.0 Tryptone 2.0 Beef extract 5.0 Yeast extract 1.0 Sodium pyruvate 10.0 Glycine 12.0 Lithium chloride 5.0 Potassium tellurite 0.1 Distilled or deionized water 1000.0
Preparation
Dissolve all the ingredients except the potassium tellurite in the water with the aid of heat. Cool to room temperature and adjust the pH to 6.6. Dispense into test tubes and sterilize at 121~ for 15 min. When cooled to approximately 50~ add filter sterilized potassium tellurite solution to give a final concentration of 100/xg/ml.
Physical properties
Appearance Clear, amber. pH 6.6 _+ 0.2

262
Shetf
Basal medium without potassium tellurite Complete medium
1 month at 4~ Use the same day.
Before use, stored basal medium should be heated to 100~ for 15 min to remove oxygen, cooled to 50~ and the potassium tellurite solution added.
Inoculation method for samples
Use food macerates or decimal dilutions in peptone saline and inoculate broths in the proportion 1:10. Layer tubes with sterile liquid paraffin.
Incubation method
At 37~ for 48 h anaerobically.
Reading of results and interpretation
Turbidity and the production of a black deposit provide presumptive evidence of Staphylococcus aureus. Isolation on Baird-Parker agar and subsequent confirma- tory tests for coagulase production must be carried out.
Quality assessment
(i) Productivity Test strain Staphylococcus aureus 50080
Inoculation method Dilution to extinction.
Criteria Recovery in LBP should be within 1 titre unit of the recovery in TSB after 48 h at 37~
(ii) Selectivity Test strain Micrococcus luteus 50063
Escherichia coli 50034
Inoculation method Dilution to extinction.
Criteria Recovery in LBP should be less than 5 titre units of recovery in TSB after 48 h at 37~
Reference
Baird, R.M. and van Doorne, H. (1982) Enrichment techniques for Staphylococcus aureus. Arch. Lebensmittelhyg. 33, 146-150.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
263
Bile Oxalate Sorbose (BOS) broth
Description and history
A two-step enrichment procedure for the recovery of Yersinia enterocolitica from foods was developed by Schiemann (1982). In this procedure pre-enrichment in yeast extract rose bengal broth or in tryptone soya broth is followed by selective enrichment in BOS broth. The selective agents in BOS broth are sodium oxalate, bile salts, irgasan and sodium furadantin. BOS broth was found especially useful for the isolation of serotype 0:8 strains (Schiemann, 1983).
Composition (grams)
Di-sodium hydrogen o r t h o p h o s p h a t e . 7H2 ~ 17.25 Sodium oxalate 5.0 Bile salts no. 3 2.0 Sodium chloride 1.0 Magnes ium sulphate �9 7H 2 O 0.01 Sorbose * 10.0 Asparag ine * 1.0 Meth ion ine * 1.0 Metani l yellow * 0.025 Yeas t extract * 0.025 Sodium pyruvate * 0.05 2,3,4'-trichloro-2'-hydroxyl diphenyl e ther *
(Irgasan; Ciba-Ceigy) 0.004 Sodium furadant in * 0.01 Disti l led or deionized water 659.0
Preparation
Dissolve ingredients except those marked * in distilled water. Adjust pH to 7.6. Autoclave at 121~ for 15 min. Cool to 50~ and add the following filter sterilized solutions:

264
Sorbose (10%) Asparagine (1.0%) Methionine (1.0%) Metanil yellow (2.5 mg /ml ) Yeast extract (2.5 mg /ml ) Sodium pyruvate (0.5%) Irgasan (0.4% in 95% ethanol)
Adjust pH to 7.6. On day of use add the following solution"
Sodium furadantin (1.0 mg /ml )
100.0 ml 100.0 ml 100.0 ml
10.0 ml 10.0 ml 10.0 ml
1.0 ml
10.0 ml
Distribute aseptically in the required volumes in flasks.
Physical properties
Appearance Yellow, clear. pH 7.6 +_ 0.1
Shelf life
Prepared medium (without sodium furadantin) 7 days at 4 _+ 2~
Inoculation method for samples
Add food sample to pre-enrichment medium in proportion 1"10. Incubate for 9 days at 4~ or for 20-24 h at 24 _+ 2~ Inoculate pre-enriched medium in BOS broth in proportion 1"10.
Incubation method
At 24 +_ 2~ for 5 days in air.
Reading of results and interpretation
Growth of yersiniae usually results in obvious turbidity. The medium should always be cultured by streaking one loopful on CIN agar.
Quality assessment
(i) Productivity Test strains Yersinia enterocolitica, biotype 4 serotype 0" 3 50087
Yersinia enterocolitica, biotype 1 serotype 0"8 50085

265
Inoculation method Dilution to extinction.
Criteria Growth should be within 2 titre units of the growth in tryptone soya broth after 5 days at 24~
(ii) Selectivity Test strains Proteus mirabilis (ATCC 29906/CECT 4168/NCTC
11938) Pseudomonas aeruginosa 50067
Inoculation method Dilution to extinction.
Criteria Difference in growth should be equal to or less than 5 titre units of the growth in tryptone soya broth after 5 days at 24~
References
Schiemann, D.A. (1982) Development of a two-step enrichment procedure for recovery of Yers&ia enterocolitica from food. Appl. Environ. Microbiol. 43, 14-27.
Schiemann, D.A. (1983) Comparison of enrichment and plating media for recovery of virulent strains of Yersinia enterocolitica from inoculated beef stew. J. Food Protect. 46, 957-964.

266 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Bismuth sulphite agar
Description and history
This is a modification of the original Wilson and Blair (1927) selective diagnostic medium used for the isolation of Salmonella typhi and other salmonellae from food and other materials e.g. sewage, water, etc. Bismuth sulphite and brilliant green are the selective agents. The medium also contains indicators for sulphide production. The use of this medium is advocated by several authorities (Anon, 1981; ICMSF, 1978; Speck, 1984) and it may be particularly useful when lactose fermenting strains of salmonellae are sought.
Composition (grams)
Beef extract Peptone Glucose Di-sodium hydrogen orthophosphate �9 12H20 Iron (II) sulphate. 7H2 O Bismuth ammonium citrate Sodium sulphite Brilliant green Agar Distilled or deionized water
5.0 10.0 5.0 4.0 0.3 1.85 6.15 0.016 *
20.0 1000.0
* concentration varies with formulation and activity of the dye
Preparation
Suspend the ingredients in the water. Mix and heat very carefully to boiling, with frequent agitation, to dissolve soluble material. An insoluble precipitate is formed. Cool to 45-50~ Adjust pH to 7.6 _+ 0.2. Keeping the precipitate in suspension, pour 15-20 ml quantities into 9 cm diameter Petri dishes or about 40 ml into 14 cm diameter Petri dishes.

Physical properties
Appearance
pH
267
Pale green/s t raw, opaque with flocculent precipitate which must be uniformly dispersed in the liquid medium. 7.6 +0 .2
Shelf life
Ready to use medium 5 days at 4~ dependent on formulation and specific application. Manufacturers of some dehydrated prepara- tions recommend storage of the plated medium for a minimum period of one to three days, others recommend use on the day of preparation. In general the medium becomes less inhibitory on storage.
Inoculation method for samples
Streak inoculation on pre-dried plates (see above) from selective enrichment broths by method specified in ISO 6579-1981(E) (Anon, 1981; see also van Leusden et al., 1982). Other streaking techniques which result in well-isolated colonies may be satisfactory.
Incubation method
At 37 _+ 1~ for 24-48 h in air.
Reading of results and interpretation
After 24 h well-isolated colonies of most salmonellae are small, black, flat or raised, with blackening of the medium and characteristic metallic sheen. These reactions are intensified on a further 24 h incubation. Non-HzS producing strains of salmonellae grow as green colonies with no blackening. Because of this and other variations in characteristics of salmonellae on bismuth sulphite agar, almost any growth on the medium should be subject to further tests.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077
Supplementary strain Salmonella saintpaul 50075
Inoculation method Modified Miles-Misra or streaking/ecometry.

268
(ii) Selectivity T e s t s t r a ins Escherichia coli 50034
Proteus mirabilis ( A T C C 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C
11938) - qua l i t a t i ve tes t fo r c o n t r o l o f s w a r m i n g .
I n o c u l a t i o n m e t h o d M o d i f i e d M i l e s - M i s r a o r s t r e a k i n g / e c o m e t r y .
(iii) Characteristic appearance o f colonies See above .
References
Anon. (1981) Microbiology - general guidance on methods for the detection of Salmonellae. Interna- tional Standard ISO 6579-1981(E) Geneva: International Organisation for Standardisation.
ICMSF. (1978) Microorganisms in foods. 1. Their significance and methods of enumeration. 2nd edn. University of Toronto Press, Toronto, Ont.
Speck, M.L. (1984) Compendium of methods for the microbiological examination of foods. 2nd edn. American Public Health Association.
van Leusden, F.M., van Schothorst, M. and Beckers, H.J. (1982) The standard Salmonella isolation method. In: Isolation and identification methods for food poisoning organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner. SAB Technical Series No. 17. Academic Press, London, pp. 35-49.
Wilson, W.J. and Blair, E.M.M'V. (1927) Use of a glucose bismuth sulphite iron medium for the isolation of B. typhosa and B. proteus. J. Hyg. 26, 374-391.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
269
Briggs agar
Description and history
Briggs tomato juice agar (Briggs, 1953) was developed primarily for the cultivation of lactobacilli from milk and dairy products and has since been successfully used for strains from other sources (Cox and Briggs, 1954; Lerche and Reuter, 1960; Reuter, 1964). It may be used for the non-selective enumeration of the whole group of lactic acid bacteria and has better productivity for some strains of Lactobacillus delbrueckii ssp. bulgaricus, Streptococcus salivarius ssp. thermophilus and Lactococcus lactis ssp. lactis than MRS agar.
Composition (grams)
Tryptone (tryptic digest of casein) Peptone (peptic digest of meat) Yeast extract Glucose Starch, soluble Sorbitan monooleate (Tween 80) Sodium chloride Agar
8.0 8.0 6.0
20.0 0.5 1.0 5.0
20.0
Distilled or deionized water 600.0, 800.0 or 900.0 (depending on tomato juice used, a, b or c)
additionally:
a) Tomato juice dilution One part of the commercial product without salt and preserving agents is diluted with two parts distilled or deionised water, boiled and filtered. 400 ml of this dilution is added to 600 ml basal agar.
b) Fresh tomato juice Filter the juice of freshly pulped tomatoes to remove seeds and coarse particles. Autoclave and add 200 ml to 800 ml of the basal agar.

270
c) Tomato extract dilution Mix 40 g tomato extract (triple), 5.75 g Na2HPO4, 0.8 g K H z P O 4 in 500 ml distilled or deionised water, filter and autoclave at 121~ for 15 min. Add 100 ml to 900 ml basal agar. The remainder may be stored for future use.
Preparation
Suspend the ingredients in the appropriate amount of water and add the diluted tomato juice or tomato extract dilution. Heat to boiling to dissolve completely. Adjust pH to 6.8 with 10% NaOH. Sterilize in the autoclave for 15 min at 121~ Cool to 50~ and distribute into sterile Petri dishes. Avoid overheating this medium which hydrolyses the agar, resulting in a soft medium. N.B. The tomato juice must be free of Cu contamination.
Physical properties
Appearance
pH
Medium to dark amber, slightly opalescent, may have a slight precipitate. 6.8 + 0.2
Shelf life
Tomato extract dilution Ready to use medium
4-6 weeks at 4 + 2~ 14 days at 4 + 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
This depends on the particular habitat of the organisms to be cultivated. Dairy strains should be incubated at 30~ for 2 days followed by 1 day at 22~ intestinal or yoghurt strains for 2 days at either 37 or 42~ All incubation should be performed under anaerobic or microaerobic (6% 02: 10% CO 2 in N 2) conditions.
Reading of results and interpretation
Briggs agar is an elective medium that gives good colony counts and a similar colony size and appearance for all lactic acid bacteria. Other microorganisms may be distinguished by pigments and extraordinary colonial morphology (micrococci, Gram negative species, yeasts). Confirmation tests e.g. Gram stain, catalase reac- tion, are necessary to distinguish mixed microflora. A selective medium for

271
lactobacilli or lactic acid bacter ia should be used in paral lel where very mixed microf lora are present .
Quality assessment
(i) Productivity Test strains Lactobacillus delbrueckii ssp. bulgaricus 50050
Streptococcus salivarius ssp. thermophilus 50083 Lactococcus lactis ssp. lactis 50058
Inocula t ion m e t h o d Modif ied Miles-Misra or s t r e a k i n g / e c o m e t r y .
Cri ter ia Recovery on Briggs agar should be within 1 log m0 of recovery on M R S agar.
(ii) Characteristic appearance of colonies
Small greyish-white colonies, 1 -3 m m diameter , flat or raised, smooth, rough or in te rmedia te . Non-lact ic acid bacter ia e.g. yeasts and Gram-nega t ive species, may be dis t inguished by colony size and appea rance af ter fu r ther incubat ion at room t empera tu re .
References
Briggs, M.J. (1953) An improved medium for lactobacilli. J. Dairy Res. 20, 36-40. Cox, C.P. and Briggs, M.J. (1954) Experiments on growth media for lactobacilli. Appl. Bacteriol. 17,
18-26. Lerche, M. and Reuter, G. (1960) Beitrag zur Methodik der Isolierung und Differenzierung von aerob
wachsenden Laktobazillen. Zbl. Bakteriol. I. Orig. 179, 354-370. Reuter, G. (1964) Vergleichsuntersuchungen an 90 thermophilen Lactobazillen Stammen verschiedener
Herkunft. Zbl. Bakteriol. I. Orig. 193, 454-466.

272 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Brilliant Green Bile (BGB) broth
Description and history
This broth is a modification of MacConkey's liquid medium for the isolation of Enterobacteriaceae, formulated by Dunham and Schoenlein in 1926 to attain maximum recovery of bacteria of the coli-aerogenes group, while inhibiting most Gram positive organisms which might hinder the development of the bacteria sought. It contains brilliant green and bile as the inhibitory agents for Gram positive organisms and lactose as the carbon source, which is dissimilated rapidly by the coli-aerogenes group, mostly by a heterofermentative pathway, leading to gas formation. MacKenzie et al. (1948) found it markedly superior to MacConkey broth for confirmation of Escherichia coli at 44~ and it is now common practice to carry out preliminary MPN tests using a less selective medium such as lauryl tryptose broth or minerals modified glutamate medium and confirm any tubes showing a positive reaction by subculture to BGB broth.
Composition (grams)
Pep tone 10.0 Lactose 10.0 Ox bile 20.0 Brilliant green 0.0133 Distil led or deionized water 1000.0
Preparation
Dissolve the ingredients in water, distribute in the required volumes in flasks or tubes and heat at 100~ for 30 min. Although lactose media are markedly less sensitive to heat damage than those containing glucose, the performance of the medium, with respect to both selectivity and productivity is much more consistent when it is decontaminated by standardised pasteurization as recommended.
Physical properties
Appearance Green, clear. pH 7.4 _+ 0.2

273
Shelf life
Ready to use medium 1 month at 4 _+ 2~ in screw capped containers.
Inoculation method for samples
Use food macerates or decimal dilutions and inoculate the broth in the proportion 1:9. BGB broth can be used at double strength. In this instance, it must not be autoclaved. Equal volumes of food macerates, or decimal dilutions (or fluid samples) are added to the double strength BGB broth.
Incubation method
To detect members of the coli-aerogenes group incubate at 35 or 37~ for 24-48 h. For dairy purposes and where it is desired to detect the full range of coliform bacteria, incubate at 30~ for 24-48 h. For Escherichia coli the temperature of 44 _+ 0.1~ for 18 h is specifically recommended. Psychrotrophic coliforms can be detected by incubation at 4~ for 10 days.
Reading of results and interpretation
Turbidity and often change of colour of the medium towards yellowish-green provides presumptive evidence of the presence of bacteria of the coli-aerogenes group, particularly when accompanied by copious gas formation. This should be confirmed by isolation on violet red bile glucose agar and subsequent study of the mode of attack on glucose and negative oxidase reaction, preferably by the technique described in the monograph on VRB agar.
Quality assessment
(i) Productivity Test strains Escherichia coli 50034
Citrobacter freundii 50025
Inoculation method Dilution to extinction.
Criteria Recovery in BGB broth should be within one titre unit of the recovery in tryptone soya broth and copi- ous gas formation should occur with Escherichia coli and Citrobacter freundii within 18-24 h at 30~ the same should apply for Escherichia coli only after incubation for 24 h at 44 + O.I~

274
(ii) Selectivity Test strains Enterococcus faecalis 50030
Staphylococcus aureus 50080
Supp lemen ta ry strains Bacillus cereus 50014 Lactococcus lactis ssp. lactis 50058
Inocula t ion m e t h o d Dilut ion to extinction.
Cri ter ia Recovery in BGB should be less than 5 t i tre units of the recovery in t ryp tone soya bro th after 18-24 h at 30~ At 44 + 0.1~ the m e d i u m should be even more selective.
References
Dunham, H.G. and Schoenlein, H.W. (1926) Brilliant green bile media. Stain Technol. 1, 129-134. MacKenzie, E.F.W., Taylor, E.W. and Gilbert, W.E. (1948) Recent experiences in the rapid identifica-
tion of Bacterium coli, type 1. J. Gen. Microbiol. 2, 197-204.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
275
Cefsulodin Irgasan Novobiocin (CIN) agar
Description and history
The medium was originally formulated by Schiemann (1979) for detection of Yersinia enterocolitica. He subsequently (1982) revised it by substituting sodium deoxycholate for bile salts and reducing the novobiocin content. It relies on the use of the selective inhibitory components sodium deoxycholate, crystal violet, cefsu- lodin, irgasan and novobiocin. The indicative principle is fermentation of mannitol with localised pH reduction which forms a red colony due to the neutral red, and a zone of precipitation due to the deoxycholate.
Composition (grams)
Peptone 20.0 Yeast extract 2.0 Mannitol 20.0 Sodium pyruvate 2.0 Sodium chloride 1.0 Magnesium sulphate- 7H2 O 0.01 Sodium deoxycholate 0.5 Neutral red 0.03 Crystal violet 0.001 Cefsulodin * 0.015 2,4,4'-trichloro-2'-hydroxyl diphenyl ether 0.004
(Irgasan, Ciba-Geigy) * Novobiocin * 0.0025 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients (except those marked *) in 970 ml water. Bring to the boil to dissolve completely. Allow to cool to 50~ and aseptically add the following filter-sterilized solutions: (i) 10 ml of cefsulodin (150 mg/100 ml ethanol/water 1:1); (ii) 10 ml of irgasan (40 mg/100 ml ethanol); (iii) 10 ml of novobiocin (25 mg/100 ml water). Mix gently and pour into sterile Petri dishes.

276
Physical properties
Appearance Pink, clear. pH 7.4 _+ 0.2
Shelf life
Ready to use medium 7 days at 4 _+ 2~
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per 9 cm diameter pre-dried plate.
Incubation method
At 30~ for 18-24 h or at 22~ for 48 h in air.
Reading of results and interpretation
Yersinia enterocolitica appears as round pink colonies about 2 mm diameter, with dark pink centres surrounded by a zone of precipitated bile. Confirmatory tests are required.
Quality assessment
(i) Productivity Test strains Yersinia enterocolitica, biotype 4 serotype 0"3 50087
Yersinia enterocolitica, biotype 1 serotype 0"8 50085
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Escherichia coli 50034
Pseudomonas aeruginosa 50067
Supplementary strains Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 19938) Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or streaking/ecometry.
(iii) Characteristic appearance of colonies
Round pink, about 2 mm in diameter with a dark pink centre and surrounded by a precipitation zone.

277
References
Schiemann, D.A. (1979) Synthesis of a selective agar medium for Yersinia enterocolitica. Can. J. Microbiol. 25, 1298-1304.
Schiemann, D.A. (1982) Development of a two step enrichment procedure for recovery of Yersinia enterocolitica from food. Appl. Environ. Microbiol. 43, 14-27.

278 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Cellobiose Polymyxin B Colistin (CPC) agar
Description and history
The medium was developed for the isolation of Vibrio vulnificus and Vibrio cholerae from environmental sources (Massad and Oliver, 1987). It was shown to be superior to other media in its ability to select and differentiate V. vulnificus from other species found in the environment (Oliver et al., 1992). Its use in the examination of oysters was reported by Kaysner et al. (1989) and Tamplin et al. (1991). The medium is less inhibitory to V. vulnificus than is TCBS, the medium used for isolation of most other pathogenic Vibrio spp. The selective components of CPC agar are the antibiotics colistin and polymyxin B, and a high incubation temperature (40~ Differentiation is based on cellobiose fermentation which is detected by the pH indicators.
Composition (grams)
Solution 1 Peptone 10.0 Beef extract 5.0 Sodium chloride 20.0 Bromothymol blue 0.04 Cresol red 0.04 Agar 15.0 Distilled or deionised water 900.0
Solution 2 Cellobiose Colistin methanesulphonate (i.u.) Polymyxin B (i.u.) Distilled or deionized water
15.0 1 360 000.0
100 000.0 100.0
Preparation
Suspend the ingredients for solution 1 in water. Adjust to pH 7.6. Autoclave at 121~ for 15 min. Cool to 55~ For solution 2, dissolve cellobiose in water by heating gently. Cool. Add antibiotics. Filter sterilize solution 2. Add to cooled solution 1. Pour plates.

279
mCPC Modification
CPC agar has been used in a modified form to make it less inhibitory to Vibrio vulnificus. Modified CPC (mCPC) is used by the U.S. FDA. The modifications are a reduction of cellobiose concentration to 10 g/1 and colistin methanesulphonate to 400000 i.u./1. Solution 1 may be boiled rather than autoclaved and filter sterilization of Solution 2 is not necessary.
Physical properties
Appearance pH
Olive green to light brown-purple. 7.6 + 0.2
Shelf life
Use freshly prepared medium. Old plates of CPC are more inhibitory, especially to Vibrio cholerae, than is fleshly prepared medium.
Inoculation method for samples
Surface spreading or streaking over plate from sample, homogenate or alkaline peptone water enrichment broth culture.
Incubation method
At 39-40~ for 24 to 48 h in air. Lower temperature incubation decreases the inhibitory nature of the medium and allows growth of some other species of Vibrio.
Reading of results and interpretation
The medium is very selective and most other Vibrio spp. will not grow on it. Vibrio vulnificus produces flat yellow (cellobiose fermenting) colonies with opaque cen- tres and translucent peripheries, about 2 mm in diameter. Vibrio cholerae E1 Tor produces raised purple or green colonies. Classical Vibrio cholerae are sensitive to polymyxin B and will not grow on the medium.
Quality assessment
(i) Productivity Test strains Vibrio vulnificus 50104
Vibrio cholerae (NCTC 11348)
Inoculation method Modified Miles-Misra or streaking/ecometry.

280
(ii) Selectivity Tes t s t ra ins Vibrio fluvialis 50084
Vibrio parahaemolyticus ( N C T C 11344)
Photobacterium leiognathi 50103
Pseudomonas aeruginosa 50067
I n o c u l a t i o n m e t h o d M o d i f i e d M i l e s - M i s r a or s t r e a k i n g / e c o m e t r y .
(iii) Characteristic appearance o f colonies See above.
References
Kaysner, C.A., Tamplin, M.L., Wekell, M.M., Stott, R.F. and Colburn, K.G. (1989) Survival of Vibrio vulnificus in shellstock and shucked oysters (Crassostrea gigas and Crassostrea virginica) and effects of isolation medium on recovery. Appl. Environ. Microbiol. 55, 3072-3079.
Massad, G. and Oliver, J.D. (1987) New selective and differential medium for Vibrio cholerae and Vibrio vulnificus. Appl. Environ. Microbiol. 53, 2262-2264.
Oliver, J.D., Guthrie, K., Preyer, J., Wright, A., Simpson, L.M., Siebeling, R. and Morris, Jr., J.G. (1992). Use of colistin-polymyxin B-cellobiose agar for isolation of Vibrio vulnificus from the environment. Appl. Environ. Microbiol. 58, 737-739.
Tamplin, M.L., Martin, A.L., Ruple, A.D., Cook, D.W. and Kaspar, C.W. (1991) Enzyme immunoassay for identification of Vibrio vulnificus in seawater, sediment, and oysters. Appl. Environ. Microbiol. 57, 1235-1240.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
281
Cephaloridine Fucidin Cetrimide (CFC) agar
Description and history
The selectivity of CFC agar depends upon a specific combination of antibacterial compounds: cephaloridine, fucidin and cetrimide (Mead and Adams, 1977). The medium has no differential properties and aims to suppress virtually all unwanted organisms. Although developed for use in the microbiological examination of poultry-meat products, CFC agar can also be applied to other foods for isolating Pseudomonas spp. (Gardner, 1980; Banks and Board, 1983)
Composition (grams)
Heart infusion agar Cephaloridine (Ceporin) Sodium fusidate (Fucidin) Cetyltrimethyl ammonium bromide (Cetrimide) Distilled or deionized water
40.0 0.05 0.01 0.01
1000.0
Preparation
Prepare 1 litre of heart infusion agar. Dispense into bottles (or final containers) and sterilize by autoclaving at 121~ for 15 min. Cool the sterile basal medium to 50-52~ and add the required amounts of the following filter-sterilized aqueous solutions: (i) 5 ml of 1% cephaloridine; (ii) 1 ml of 1% sodium fusidate; (iii) 1 ml of 1% cetyltrimethyl ammonium bromide.
Physical properties
Appearance Straw coloured. pH 7.4 + 0.2
Shelf life
Ready to use medium At least 1 month at 20 + 2~

282
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per 9 cm diameter pre-dried plate. Ensure adequate ventilation of plates during incubation.
Incubation method
At 25~ for 48 h in air.
Reading of results and interpretation
There are two types of colonies, pigmented and non-pigmented. Round, white or cream colonies or greenish, fluorescent colonies, 2-5 mm diameter are considered to be pseudomonads. Confirmatory tests are not usually required but in cases of doubt plates can be flooded with oxidase reagent (1% tetramethyl-p-phenylene- diamine dihydrochloride). Colonies showing an outer dark-purple ring within 10 seconds are counted as pseudomonads.
Quafity assessment
(i) Productivity Test strains Pseudomonas fluorescens 50068
Pseudomonas fragi 50069
Supplementary strain Pseudomonas putida 50070
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(ii) Selectivity Test strains Acinetobacter sp. 50011
Escherichia coli 50034
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(iii) Characteristic appearance of colonies
Round, white or cream colonies or greenish, fluorescent colonies, 2-5 mm diameter.
References
Banks, J.G. and Board, R.G. (1983) The classification of pseudomonads and other obligately aerobic Gram-negative bacteria from British pork sausage and other ingredients. Syst. Appl. Microbiol. 4, 424-438.

283
Gardner, G.A. (1980) The occurrence and significance of Pseudomonas in Wiltshire bacon brines. J. Appl. Bacteriol. 48, 69-74.
Mead, G.C. and Adams, B.W. (1977) A selective medium for the rapid isolation of pseudomonads associated with poultry meat spoilage. Brit. Poult. Sci. 18, 661-670.

284 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Charcoal Cefoperazone Deoxycholate (CCD) agar- modified
Description and history
This blood-free medium is an improved version of the original CCD agar (Bolton et al., 1984). Modified CCD agar (Hutchinson and Bolton, 1984)was developed for the direct isolation of campylobacters from human and animal faeces and its major constituents are charcoal, cefoperazone and sodium deoxycholate. It has subse- quently proved to be a very successful subculture medium when used in conjunc- tion with Preston enrichment broth (Bolton et al., 1986).
Composition (grams)
Beef extract 10.0 Peptone 10.0 Sodium chloride 5.0 Charcoal 4.0 Casein hydrolysate 3.0 Iron (II) sulphate 0.25 Sodium pyruvate 0.25 Sodium deoxycholate 1.0 Cefoperazone * 0.032 Amphoter ic in * 0.01 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend the above ingredients except those marked * in the water and bring to the boil to dissolve completely. Sterilize by autoclaving at 121~ for 15 min. Cool to 50~ and aseptically add the cefoperazone, and amphotericin if required, to give the final concentration(s)stated.

285
Physical properties
Appearance Black, opaque. pH 7.4 _+ 0.2
Shelf life
Ready to use medium 10 days at 4 _+ 2~
Inoculation method for samples
Plates should not be overdried but preferably left overnight at room temperature. This medium can be inoculated as a surface spread plate or conventionally to produce discrete colonies.
Incubation method
At 42~ for 48 h microaerobically in a jar with an atmosphere containing approxi- mately 5% O 2, 10% CO 2 and 85% N 2 or H 2.
Reading of results and interpretation
Campylobacter jejuni strains produce grey, moist, flat and occasionally spreading growth which may be accompanied with a green hue a n d / o r a metallic sheen. Campylobacter coli strains tend to be creamy-grey in colour, moist and often produce a more discrete type of colony. Campylobacter lari strains are more varied and produce both types of colonial morphology.
Quality assessment
(i) Productivity Test strains Campylobacter jejuni 50091
Campylobacter coli 50092
Inoculation method
(ii) Selectivity Test strains
Modified Miles-Misra or streaking/ecometry.
Proteus mirabilis (ATCC 29906, CECT 4168 /NCTC 11938) Pseudomonas aemginosa 50067
Supplementary strains Bacillus cereus 50014 Escherichia coli 50034 Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or streaking/ecometry.

286
(iii) Characteristic appearance of colonies
See above. Occas iona l ly c o n t a m i n a t i n g o rgan i sms may grow on this m e d i u m .
T h e s e inc lude c e f o p e r a z o n e res i s t an t Pseudomonas spp. and E n t e r o b a c t e r i -
aceae , some s t r ep tococc i and yeasts .
References
Bolton, F.J., Hutchinson, D.N. and Coates, D. (1984) Blood-free selective medium for isolation of Campylobacter jejuni from faeces. J. Clin. Microbiol. 19, 167-171.
Bolton, F.J., Hutchinson, D.N. and Coates, D. (1986) Comparison of three selective agars for the isolation of campylobacters. Eur. J. Clin. Microbiol. 5, 466-468.
Hutchinson, D.N. and Bolton, F.J. (1984) An improved blood-free selective medium for the isolation of Campylobacter jejuni from faecal specimens. J. Clin. Pathol. 37, 956-957.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
287
Charcoal Cefoperazone Deoxycholate (CCD) broth
Description and history
This blood-free broth is the liquid version of modified CCD agar (Bolton et al., 1984; Hutchinson and Bolton, 1986). Charcoal, ferrous sulphate and sodium pyruvate replace blood, while sodium deoxycholate and cefoperazone are the selective agents in this enrichment broth. It is used in conjunction with modified CCD agar or Skirrow's agar for the isolation of low numbers of thermophilic campylobacters from food and environmental samples (Bolton et al., 1986; Korho- nen and Martikainen, 1990).
Composition (grams)
Beef extract 10.0 Peptone 10.0 Sodium chloride 5.0 Charcoal 4.0 Casein hydrolysate 3.0 Iron (II) sulphate 0.25 Sodium pyruvate 0.25 Sodium deoxycholate 1.0 Cefoperazone * 0.032 Amphoter ic in * 0.01 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients except those marked * in the water and bring to the boil. Sterilize by autoclaving at 121~ for 15 min. Cool to 50~ and aseptically add the required amount of cefoperazone and amphotericin to give the final concentra- tions stated. Distribute aseptically in the required volumes in flasks or tubes.
Physical properties
Appearance Black, opaque. pH 7.4 _+ 0.2

288
Shelf life
Ready to use medium 14 days at 4 + 2~
Inoculation method for samples
Use macerates of foods or environmental samples and inoculate broth in propor- tion 1"10.
Incubation method
At 42~ for 24-48 h microaerobically in a jar with an atmosphere containing approximately 5% 0 2, 10% CO 2 and 85% N 2 or H 2.
Reading of results and interpretation
Growth of campylobacters in the broth is usually not visible, but can be detected by microscopic examination. The medium should always be subcultured by streak- ing one loopful on CCD agar or Skirrow's medium.
Quality assessment
(i) Productivity Test strains Campylobacter jejuni 50091
Campylobacter coli 50092
Inoculation method Dilution to extinction.
Criteria Growth should be within 2 titre units of the growth in tryptone soya broth after 48 h at 42~
(ii) Selectivity Test strains Proteus mirabilis (ATCC 29906/CECT 4168 /NCTC
11938) Pseudomonas aeruginosa 50067
Inoculation method Dilution to extinction.
Criteria Difference in growth should be equal to or less than 5 titre units of the growth in tryptone soya broth.
References
Bolton, F.J., Hutchinson, D.N. and Coates, D. (1984) Blood-free selective medium for isolation of Campylobacter jejuni from faeces. J. Clin. Microbiol. 19, 167-171.

289
Bolton, F.J., Hutchinson, D.N. and Coates, D. (1986) Comparison of three selective agars for the isolation of campylobacters. Eur. J. Clin. Microbiol. 5, 466-468.
Hutchinson, D.N. and Bolton, F.J. (1984) An improved blood-free selective medium for the isolation of Campylobacter jejuni from faecal specimens. J. Clin. Pathol. 37, 956-957.
Korhonen, L.K. and Martikainen, P.J. (1990) Comparison of some enrichment broths and growth media for the isolation of thermophilic campylobacters from surface water samples. J. Appl. Bacteriol. 68, 593-599.

290 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Citrate Azide Tween Carbonate (CATC) agar
Description and history
CATC agar is a selective medium for the isolation and cultivation of enterococci. High concentrations of citrate and azide achieve almost complete suppression of other bacterial flora and allow good growth of enterococci (Enterococcus faecalis ssp. and Enterococcus faecium). (Reuter, 1968; 1978; 1985).
Composition (grams)
Peptone (tryptic digest of casein) Yeast extract Potassium di-hydrogen or thophosphate Sodium citrate Sorbitan monooleate (Tween 80) Sodium carbonate * 2,3,5,-Triphenyltetrazolium chloride * Sodium azide * Agar Distilled or deionized water
15.0 5.0 5.0
15.0 1.0 2.0 0.1 0.4
15.0 1000.0
Preparation
Suspend the ingredients except those marked * in the water and heat to boiling to dissolve completely. Sterilize in the autoclave for 15 min at 121~ Cool to about 50~ and add:
20 ml of 10% solution of sodium carbonate (prepared in sterile water without heating)
10 ml of 1% solution of 2,3,5,-triphenyltetrazolium chloride
4 ml of 10% solution of sodium azide.

291
Adjust pH to 7.0 _+ 0.1 at 50~ Distribute into sterile Petri dishes.
Physical properties
Appearance Pale pink. pH 7.0 +_ 0.1 at 37-40~
Shelf lif
Ready to use medium 7 days at 4 + 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
24 h at 37-40~ in air. If plates show no growth or unusually small colonies, then incubation for a further 24 h is advisable.
Reading of results and interpretation
Distinctly red well-developed colonies can be considered as Enterococcus faecalis whilst pink colonies or colonies with a red centre are likely to be Enterococcus faecium. Since other enterococci or streptococci may appear as pink colonies, though smaller in size ( < 0.5 mm diameter) confirmation tests should be carried ou t .
Quality assessment
(i) Productivity Test strains Enterococcus faecalis 50030
Enterococcus faecium 50032
Supplementary strain Enterococcus hirae 50031
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(ii) Selectivity Test strains Escherichia coli 50034
Lactococcus lactis ssp. lactis 50058
Supplementary strains Lactobacillus casei ssp. rhamnosus 50045 Pediococcus damnosus 50065 Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or s t reaking/ecometry.

292
(iii) Characteristic appearance of colonies
Red , mostly smoo th colonies: Enterococcus faecalis. Pink, par t ly rough colonies, or red centre: Enterococcus faecium. Small colourless or p ink colonies ( < 0.5 m m diameter ) : o the r s t reptococci or enterococci , which might grow at a
s lower rate.
References
Reuter, G. (1968) Erfahrungen mit Nahrboden fiir die selektive mikrobiologische Analyse von Fleisch- erzeugnissen. Arch. Lebensmittelhyg. 19, 53-57 u. 84-89.
Reuter, G. (1978) Selektive Kultivierung von "Enterokokken" aus Lebensmitteln tierischer Herkunft. Arch. Lebensmittelhyg. 29, 84-91.
Reuter, G. (1985) Selective media for group-D-streptococci. Int. J. Food Microbiol. 2, 103-114.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
293
Cresol red Thallium Acetate Sucrose (CTAS) agar
Description and history
This medium was devised for the detection of carnobacteria (syn. Lactobacillus divergens) which fail to grow well on conventional selective media for lactobacilli (Holzapfel and Gerber, 1983). Investigations showed pH values below 6.0 to be inhibitory to carnobacteria especially in the presence of acetate. Growth of carnobacteria was found to take place at pH values of up to 11.0 (Holzapfel and Long, 1984; unpublished results) compared to a maximum pH of 7.2 for growth of practically all lactobacilli. Also, relatively high concentrations of between 0.4 and 1% manganese sulphate have been found to stimulate growth of carnobacteria more than that of enterococci (Bosch and Holzapfel, 1985; unpublished results). In order to minimise Maillard reactions during heat sterilization of such alkaline media, glucose was substituted by sucrose. Thus far, all carnobacteria have been found to ferment sucrose. The selectivity of the CTAS medium is based on its high pH (8.5 to 9.0) and the presence of thallium acetate, nalidixic acid and a relatively high concentration of sodium citrate.
Composition (grams)
Peptone from casein Yeast extract Sucrose Sorbitan monooleate (Tween 80) Sodium citrate Manganese (II) sulphate �9 4 H 2 0 Di-potassium hydrogen orthophosphate Thallium (I) acetate Nalidixic acid Cresol red 2,3,5-Triphenyltetrazolium chloride Agar Distilled or deionized water
10.0 10.0 20.0
1.0 15.0 4.0 2.0 1.0 0.04 0.004 0.01
15.0 1000.0

294
Preparation
Add all the components except triphenyl-tetrazolium chloride to 990 ml of water and bring to the boil to dissolve completely. Cool to 55~ and adjust the pH of the medium to 9.1 with 1N NaOH. Autoclave at 121~ for 10 min, cool to 55~ and add 10 ml of a 10% solution of triphenyl-tetrazolium chloride. Pour about 15 ml of medium per Petri dish.
Physical properties
Appearance pH
Red to purple-red with precipitate. 9.0 + 0.2
Shelf life
Ready to use medium Freshly prepared plates are preferred but the medium may be stored for 1-2 weeks at 4 + 2~ in sealed plastic bags.
Inoculation method for samples
Surface spreading or direct plating of dilution series.
Incubation method
At 30~ for 24-48 h or at 25~ for 3-4 days in air.
Reading of results and interpretation
Strains of Carnobacterium piscicola (syn. Carnobacterium carnis) appear as small, bronze-metallic shiny yellowish to pinkish colonies, causing a yellow colour change of the medium and clearance of the precipitate. The metallic shine is best observed on the periphery of growth. Growth of Carnobacterium divergens strains is more sparse resulting mainly in pin-point colonies, often without colour change of the medium, and showing a bronze-metallic shine. Good growth of most Enterococcus spp. is observed with yellow colour change and clearance of precipitate in the medium. These small to pin-point (0.5 to 0.1 mm) colonies are convex, shiny and pinkish to yellowish, and without metallic shine. Listeria spp. grow sparsely, if at all, with a bronze metallic shine resembling Carnobacterium divergens. Phase contrast microscopy for determination of cell morphology is a useful tool to distinguish rods (carnobacteria) from cocci (enterococci).

295
Quafity assessment
(i) Productivity Test strains Carnobacterium piscicola 50020
Carnobacterium divergens 50021
Inoculation method Modified Miles-Misra or s t reaking/ecometry .
(ii) Selectivity Test strains Lactobacillus sake 50056
Inoculation method Modified Miles-Misra or s t reaking/ecometry .
Criteria No growth after 48-72 h at 30~
(iii) Characteristic appearance of colonies See above.
Note
Tests on more than 60 different antibiotics showed carnobacteria to have similar resistance spectra as enterococci; the same was found for heavy metal salts (Bosch and Holzapfel, 1985; unpublished results). However, substitution of sucrose by 2% inulin may be used to distinguish colonies of 'non-divergens' carnobacteria from enterococci. Unlike enterococci these carnobacteria (e.g. Carnobacterium piscicola) ferment inulin causing a change in colour of the medium from red to yellow.
Reference
Holzapfel, W.H. and Gerber, E.S. (1983) Lactobacillus divergens sp. nov., a new heterofermentative Lactobacillus species producing L(+ )-lactate. System. Appl. Microbiol. 4, 522-534.

296 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Diagnostic Salmonella Selective Semisolid Medium (DIASALM)
Description and history
DIASALM is a diagnostic selective motility agar in Petri dishes to be used for the isolation of Salmonella spp. from food and environmental samples (van Netten and van der Zee, 1991). The basal medium is a semi-solid indole motility medium (SIM). The selective system exploits the resistance of Salmonella spp. as compared to other Enterobacteriaceae to relatively high osmotic pressures, relatively low pH, malachite green and novobiocin. The diagnostic system of DIASALM consists of sucrose, lactose and bromocresol purple. This differentiates salmonellae not only from the motile non-pathogenic lactose fermenters but also from many non-patho- gens which ferment lactose or sucrose. The use of discs impregnated with iron (II) ammonium sulphate identifies suspected non-motile salmonellae.
The semi-solid approach simultaneously enriches salmonellae and separates motile salmonellae from most competitive organisms resistant to the selective system. As a result salmonellae are mostly isolated in pure culture but sometimes occur mixed with Proteus, Hafnia or Enterobacter spp. as interfering motile Enterobacteriaceae.
Where Salmonella enteritidis is specifically sought, the addition of nitrofurantoin increases the selectivity of the medium by suppressing the growth of other salmonella serogroups (Rampling et al., 1990).
Composition (grams)
Pancreat ic digest of casein * Peptic digest of animal tissue * Iron (II) ammon ium sulphate * Sodium thiosulphate * Sucrose Lactose Potass ium di-hydrogen o r thophospha te Malachi te green oxalate Magnes ium chlor ide . 6 H 2 O Bromocresol purple Sodium novobiocin Agar * Distil led or deionized water
20.0 6.1 0.2 5.0 7.5 0.5 1.47 0.037
23.34 0.08 0.010 3.5
1000.0

297
* Ingredients of BBL SIM-medium. It is imperative to use the BBL version (Cat. no. 11578).
Nitrofurantoin 0.015 g/1 may be added if Salmonella enteritidis only is sought.
Preparation
Solution A
Suspend 30 g SIM-medium (ingredients marked *), 7.5 g sucrose, 0.5 g lactose, 1.47 g potassium dihydrogen orthophosphate and 23.34 g magnesium chloride in 980 ml water. This suspension is made on the day of preparation of DIASALM.
Solution B
Dissolve 3.7 g of analytically pure malachite green oxalate in 100 ml of distilled water. Solution B can be kept for at least 8 months at room temperature in a dark bottle.
Solution C
Dissolve 0.8 g bromocresol purple in 18.5 ml 0.01N NaOH and 82.5 ml water. Store in refrigerator in dark bottle.
Solution D
Dissolve 100 mg of sodium novobiocin in 100 ml of distilled water. Sterilize by filtration. Store in refrigerator in an airtight dark bottle.
Solution E (optional)
Completely dissolve 150 mg nitrofurantoin in 10 ml acetone and sterilize by filtration. Store in refrigerator in an airtight dark bottle.
The medium is prepared by adding to 990 m of solution A, 1 ml of solution B and 10 ml of solution C. Heat the suspension in a steamer or waterbath to about 80~ until the ingredients are completely dissolved. Cool to 50~ and add 10 ml of solution D and, if desired, 10 ml of solution E. Mix and pour 20 ml in 9 cm diameter Petri dishes. After solidification plates should be stored in a refrigerator. Before inoculation plates are dried at 55~ for 5 min to remove surface moisture.
10% iron (II) ammonium sulphate discs
Immerse paper discs such as those used for antibiotic sensitivity testing in a 10% solution of iron (II) ammonium sulphate. Dry discs by placing in an open Petri dish in a 55~ incubator.

298
Physical properties
Appearance pH
Dark green-blue, transparent. 5.5 _+0.1
Shelf life
Ready to use medium 3 days at 4 _+ 2~
Inoculation method for samples
The plate is inoculated either with three drops (3 x 0.033 ml) in separate spots or with 0.1 ml in one spot in the middle. After inoculation place an iron (II) ammonium sulphate impregnated disc on the surface of the agar approximately 2 cm from the edge of the plate.
Incubation
18-24 h at 42~
Reading of results and interpretation
After incubation plates with a purple migration zone with black V-shaped feathers within it require further investigation for the presence of motile salmonellae. A black centre suggests the presence of non-motile salmonellae. The edge of the migration zone and the purple zone are not always identical. To obtain the purest subculture it is imperative to take a loopful of culture from the edge of the migration zone onto a salmonella selective diagnostic agar inoculated in such a way as to obtain well-isolated colonies. Where a black centre is apparent subculture should be made from this in the same way. The identity of suspected salmonella colonies is confirmed by appropriate biochemical and serological tests which can also be made directly with growth from the edge of the migration on green plates with black V-shaped zones.
Quality assessment
(i) Productiuity Test strains
Inoculation method
Salmonella enteritidis 50073 Salmonella uirchow 50077 Salmonella dublin 50072 Miles-Misra (without spreading of drop).

299
Cri ter ia Typical appea rance and motility (zone should extend at least 2 cm from point of inoculation). The Salmonella uirchow and Salmonella dublin strains should not grow on m e d i u m containing ni t rofuran- toin.
(ii) Selectivity Test strains Enterobacter cloacae (ATCC 23355)
Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 11938) Citrobacter freundii 50025 Pseudomonas aeruginosa 50067
Inocula t ion m e t h o d Miles-Misra (without spreading of drop).
Cri ter ia No growth or growth conf ined to cent re of plate.
(iii) Characteristic appearance of growth See above.
References
Rampling, A., Upson, R. and Brown, D.F.J. (1990) Nitrofurantoin resistance in isolations of Salmonella enteritidis phage type 4 from poultry and humans. J. Antimicrob. Chemother. 25, 285-290.
van Netten, P. and van der Zee, H. (1991) The use of a diagnostic semi-solid medium for the isolation of Salmonella enteritidis from poultry-meat. Quality of Poultry Products, Safety and Marketing aspects, Proceedings of the 10th Symposium on the quality of poultry-meat, Doorwerth, May 12-17, 1991. Spelderholt, Beekbergen.

300 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Dichloran Glycerol (DG18) agar
Description and history
This is a low water activity medium (a w 0.95) described by Hocking and Pitt (1980) suitable for the enumeration and isolation of xerophilic moulds in foods. The inclusion of dichloran serves to inhibit the rapid spreading of mucoraceous fungi and restrict colony size of other genera thus enabling more accurate counting of plates. The addition of chloramphenicol and the reduced a w prevent the growth of bacteria. Although specifically developed to enumerate xerophilic fungi in dried and semi-dried foods it has proved to be a useful general purpose medium for counting yeasts and moulds in a range of foodstuffs (Beckers et al., 1982).
Composition (grams)
Mycological peptone Glucose Potassium di-hydrogen orthophosphate Magnesium sulphate- 7 H 2 0 Dichloran (2,6-dichloro-4-nitroaniline) Chloramphenicol Glycerol Agar Distilled or deionized water
5.0 10.0
1.0 0.5 0.002 0.1
220.0 15.0
1000.0
Preparation
Suspend the ingredients except dichloran, chloramphenicol and glycerol in about 800 ml of water and heat to dissolve completely. Add 1 ml of a 0.2% (w/v) ethanolic solution of dichloran and 10 ml of a 1% ethanolic solution of chloram- phenicol and bring to 1000 ml with water. Add glycerol, mix and sterilize by autoclaving at 121~ for 15 min. Cool to 50~ mix well and dispense 15 ml amounts into sterile Petri dishes. Dry and use immediately or store dried plates at 4 _+ 2~ until required.

301
Physical properties
Appearance pH
Shelf life
Ready to use medium
Medium amber, slightly opalescent. 5.6 _+ 0.2
7 days at 4 _+ 2~
Inoculation method for samples
Spread 0.1 or 0.2 ml dilutions of the food sample over the whole of the surface of 9 cm diameter plates.
Incubation
At 25~ for 5 days in the dark. Where identification is required, prolong incuba- tion until characteristic colonies are formed.
Reading of results and interpretation
Where separate counts of moulds and yeasts are required, identify by morphologi- cal appearance and, where necessary, microscopic examination of the two groups of microorganisms.
Quality assessment
Moulds Use the stab inoculation procedure. Growth rate should be within 30% of the figure given.
Test strains CMI ATCC
Rhizopus stolonif er Aspergillus jqa vus Eurotium amstelodami
Growth rate (mm per day at 25~
61269 12.0 91856ii 11.0 17455 16018 3.3
Yeasts Use the stab inoculation procedure. Growth rate should be within 30% of the figure given.
Test strains
Saccharomyces cerevisiae Zygosaccharomyces rouxii
Growth rate (mm per day at 25~
50105 50106
0.6 0.5

302
Bacteria Use streaking procedure (Appendix I, method B) with a loopful (10/xl) from a 24 h broth culture. No growth should be evident on sectors C, D or E of the streaked plate after incubation at 25~ for 5 days.
Test strain
Bacillus subtilis 50018
References
Hocking, A.D. and Pitt, J.I. (1980) Dichloran-glycerol medium for enumeration of xerophilic fungi from low moisture foods. Appl. Environ. Microbiol. 39, 488-492.
Beckers, H.J., Boer, E., Van Eikelenboom, C., Hartog, B.J., Kuik, D., Mol, N., Nooitgedagt, A.J., Northold, M.O., and Samson, R.A. (1982) Inter. Stand. Org. Document ISO/TC34/SC9/N151.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
303
Dichloran Rose Bengal Chloramphenicol (DRBC) agar
Description and h&tory
This is a selective medium for the enumeration of moulds and yeasts in foods. King et al. (1979) showed that the combination of dichloran and rose bengal markedly restricts the size and height of mould colonies thus preventing overgrowth of luxuriant species and assisting accurate counting of colonies. The addition of chloramphenicol and the reduced pH of 5.6 serve to prevent the growth of most bacteria. This medium can be used to enumerate both toxigenic and non-toxigenic moulds. It is not diagnostic for detecting specific mycotoxin producers.
Composition (grams)
Mycological pep tone Glucose Potass ium di-hydrogen o r thophospha te Magnes ium sulphate �9 7H 20 Dichloran (2-6-dichloro-4-ni t ro-ani l ine) Rose Bengal Chloramphenico l Agar Distilled or deionized water
5.0 10.0
1.0 0.5 0.002 0.025 0.1
15.0 1000.0
Preparation
Suspend the ingredients except chloramphenicol in the water and bring to the boil to dissolve completely. Add 10 ml of a 1% ethanolic solution of chloramphenicol, mix and sterilize by autoclaving at 121~ for 15 min. Cool to below 50~ and dispense 15 ml amounts into sterile Petri dishes. Use immediately or store at 4 + 2~ in the dark until required.
Physical properties
Appearance pH
Deep pink, without any significant precipitate. 5.6 + 0.2 at 25~

304
Shelf life
Ready to use medium 7 days at 4 _+ 2~ in the dark.
Inoculation method for samples
Surface spreading over whole plate using 0.1 or 0.2 ml per 9 cm diameter plate.
Incubation method
At 25~ for 5 days in the dark. Where identification is required, prolong incuba- tion until characteristic colonies are formed.
Reading of results and interpretation
Where separate counts of moulds and yeasts are required, identify by morphologi- cal appearance and, where necessary, microscopic examination of the two groups of microorganisms. Colonies of yeasts and bacteria can be confused and should be checked microscopically.
Quality assessment
Moulds Use stab inoculation procedure. Growth rate should be within 30% of the figure given.
Test strains CMI ATCC
Rhizopus stolonif er Aspergillus fla vus Penicillium cyclopium
Growth rate (mm per day at 25~
61269 8.1 91856ii 4.5 19759 16025 3.5
Yeasts Use stab inoculation procedure. Growth rate should be within 30% of figure given.
Test strain
Saccharomyces cereuisiae
Growth rate (mm per day at 25~
50105 1.4
Bacteria Use streaking procedure (Appendix I, method B) with a loopful (10 /xl) from a 24 h broth culture. No growth should be evident on sectors C, D or E of the streaked plate after incubation at 25~ for 5 days.

305
Test strain Bacillus subtilis 50018
Supplementary strain
Escherichia coli 50034
Reference
King, D.A., Hocking, A.D. and Pitt, J.I. (1979) Dichloran-rose bengal medium for enumeration and isolation of molds from foods. Appl. Environ. Microbiol. 37, 959-964.

306 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Differential Reinforced Clostridial agar-Modified (mDRCM)
Description and history
Modified Differential Reinforced Clostridial agar was developed for the enumera- tion of spores of sulphite reducing clostridia in dried foods (Gibbs and Freame, 1965; Weenk et al., 1991; Weenk et al., 1995). It consists of a nutritionally rich basal medium, including starch to promote spore germination. Resazurin is added as redox indicator, turning red at high redox potentials, indicating aerobic condi- tions (i.e. failure to achieve anaerobic atmosphere). Sulphite and an iron salt are added as indicators. Sulphite reducing clostridia produce sulphide from sulphite, which gives a black precipitate with the iron present in the medium. Sulphite reducing clostridia are enumerated as black colonies. The current version of this medium omits acetate (Weenk et al., 1995).
Composition (grams)
Starch 1.0 Casein peptone 5.0 Meat peptone 5.0 Meat extract powder 8.0 Yeast extract powder 1.0 Resazurin 0.002 Cysteine HC1 0.5 Glucose 1.0 Iron (III) ammonium citrate 1.0 Sodium sulphite �9 7 H 2 0 0.75 Agar 20.0 Distilled or deionized water 1000.0
Preparation
Dissolve all ingredients in the water, except the iron (III) ammonium citrate and sodium sulphite. Autoclave at 121~ for 15 min, cool to about 48~ and aseptically add just before use, appropriate volumes of freshly prepared solutions of iron (III)

307
ammonium citrate (20% w/v, heat sterilized: 15 min, 121~ and sodium sulphite (10% w/v, filter sterilized).
Physical properties
Appearance Pale straw. pH 7.6 _+ 0.2
Shelf life
Basal medium Complete medium
At least 2 weeks at 4~ Use the same day (do not remelt).
Inoculation method for samples
Pour-plate procedure, using 1 ml per 9 cm diameter plate. When set, plates are overlaid with sterile mDRCM.
Incubation method
At 30~ for 3 days anaerobically.
Reading of results and interpretation
Discrete black colonies of 1-5 mm diameter are considered to be presumptive sulphite reducing clostridia.
Quality assessment
(i) Productivity Test strains Clostridium perfringens 50027
Clostridium bifermentans 50026
Supplementary strain Clostridium sporogenes 50099
Inoculation method Modified Miles-Misra or streaking method using surface inoculation of pre-poured plates with subse- quent overlay.
Criteria Recovery on mDRCM should be within 0.3 lOg l0 of the recovery on reinforced clostridial agar after 3 days anaerobic incubation at 30~

308
(ii) Characteristic appearance of colonies
Discrete black colonies of 1-5 mm diameter .
Comments
In order to facilitate spore germinat ion, a heat t r ea tmen t of the s p o r e s / s a m p l e of
10 min at 80~ may be given before inoculation of the agar.
Note
m D R C M can be used as an overlayer (ca. 5 mm thick) on surface- inoculated
modif ied M E Y P agar (q.v.). This enables a prel iminary presumptive identification
of the Clostridium species isolated (Weenk et al., 1995).
References
Gibbs, B.M. and Freame, B. (1965) Methods for the recovery of clostridia from foods. J. Appl. Bacteriol. 28, 95-111.
Weenk, G., Fitzmaurice, E. and Mossel, D.A.A. (1991) Selective enumeration of spores of Clostridium species in dried foods. J. Appl. Bacteriol. 70, 135-143.
Weenk, G.H., van den Brink, J.A., Struijk, C.B. and Mossel, D.A.A. (1995) Modified methods for the enumeration of spores of Clostridium species in dried foods. Int. J. Food Microbiol. In press.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
309
Dominguez Rodriguez LSAMm agar
Description and history
LSAMm listeria selective agar was developed by Blanco et al. J(1989) and Dominguez et al. (1990) for the detection of /3-haemolysis of Listeria spp. by a blood overlay procedure. This medium is a modification of the selective plating medium proposed by Dominguez et al. (1984) incorporating more effective selec- tive agents proposed by other authors. LSAMm has proved to be a highly selective agar inhibiting the natural microflora of many different samples (raw milk, cheese, meat, faeces, silage). Direct plating of silage samples naturally contaminated with Listeria monocytogenes on Oxford, MOX and LPM agars showed only a 4.4-4.8 log reduction of non-listeria bacteria from the silage, while LSAMm and PALCAM agars yielded a 6.8 log reduction of the non-listeria background bacteria (Fernan- dez-Garayzabal et al., 1992a; Vazquez-Boland et al., 1992). LSAMm promotes the growth of listeria present in the samples with minimal inhibition and simultane- ously provides enough information to allow an easy and reliable identification of the listeria colonies, which facilitates the isolation and counting of listeria directly on this selective agar (Dominguez et al., 1990). The use of potassium tellurite and brain heart infusion agar in LSAMm greatly enhances the/3-haemolysis of Listeria monocytogenes due to listeriolysin O (Fernandez-Garayzabal et al., 1992b). LSAMm agar should therefore be a useful medium for the detection and enumeration of Listeria spp. in highly contaminated food and feedstuffs.
Composition (grams)
Brain heart infusion agar (Follow specific BHI medium instructions) Aesculin Iron (III) ammonium citrate * Lithium chloride Potassium tellurite * Acriflavine HC1 Polymyxin B sulphate Ceftazidime pentahydra te * Distilled or deionized water
52.0
0.75 0.5
15.0 0.04 0.005 0.01 0.02
1000.0

310
Preparation
Suspend the ingredients, except those marked * in 900 ml of distilled water in a bottle or flask containing a magnetic stirring bar and autoclave the medium for 12 min at 121~ Iron (III) ammonium citrate is sterilized (121~ 12 rain) separately in 85 ml of distilled water. Cool the basal medium in a water bath to 46~ then add the heat sterilized iron (III) ammonium citrate and the following filter sterilized solutions: 10 ml of 0.2% sodium ceftazidime and 5 ml of 0.8% potassium tellurite. Dispense 15 ml per 9 cm diameter Petri dish.
Physical properties
Appearance
pH
Light yellow or tan. Dark colour indicates overheating and decomposition of either aesculin or tellurite. 7.4 +0 .2
Shelf life
Ready to use medium Component parts
3 weeks at 4 _+ 2~ Ceftazidime solution can be stored at - 20~ for several months, potassium tellurite solution at 4 + 2~ for one month.
Inoculation method for samples
1. From enrichment broths, streak a loopful of the enrichment broth on LSAMm agar to obtain isolated colonies on the plates.
2. For direct plating and counting of > 100 c f u / g of Listeria spp. growing on solid samples such as silage and many kinds of foods at the retail level, make homogenate suspensions of the food or silage samples, and then spread plate 0.1 ml of each dilution on LSAMm agar. In liquid samples the detection limit is > 10 c fu /ml of Listeria spp.
Incubation method
At 37~ for 48 h in air.
Reading of results and interpretation
After 48 h incubation Lbsteria spp. typically form 1.5 mm grey-green colonies with a dark brown centre surrounded by a black halo. Catalase, tellurite and aesculin positive colonies on LSAMm are in most cases Lister& spp.

311
Quafity assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152 /
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria ivanovii serovar 5 50095
Supplementary strains Listeria innocua serovar 6a 50094 Listeria monocytogenes serovar 4a (ATCC 19114/ NCTC 5214) Listeria seeligeri serovar 1/2b 50000 Listeria welshimeri 50001
Inoculation method
(ii) Selectivity Test strains
Modified Miles-Misra or streaking.
Enterococcus faecalis 50030 Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938)
Supplementary strains Bacillus cereus 50014 Brochothrix thermosphacta 50019 Carnobacterium piscicola 50024
Inoculation method Modified Miles-Misra or streaking.
(iii) Characteristic appearance of colonies
Grey-green colonies 1.5 mm diameter with a central dark brown area, sur- rounded by a black aesculin positive zone.
Determination of haemolytic activity by an overlay technique
This technique is based on the addition of a sheep blood (SRBC) top-layer to the selective plating medium after growth of listeria.
Composition of the top layer (grams)
BHI broth Agar Sodium chloride Sheep blood red cells suspension (ml) Distilled or deionized water
37.0 7.0 8.0
50.0 1000.0

312
Preparation of the SRBC suspension
25 ml of twice-washed SRBC are mixed with 25 ml saline solution (0.8% NaC1). This suspension is stored at 4~ and used within 1 week.
Preparation of the top layer
All ingredients, except the SRBC suspension are sterilized (121~ 15 min) and stored at 4~ Just before use, base top layer is boiled, cooled to 45~ and mixed with the SRBC suspension.
Reading of results and interpretation
After incubation (37~ 48 h) and recognition of listeria colonies by their morpho- logical characteristics on LSAMm, plates are cooled (4~ 2 h) and subsequently covered gently by an 8 ml SRBC top layer prepared as above. After the addition of the top layer, plates are incubated again (overnight at 30~ for haemolysis screening.
On LSAMm all colonies morphologically resembling listeria and positive for tellurite, aesculin and catalase (tested by removing a piece of colony prior to the top layer addition), and displaying typical haemolysis correspond in most cases to Listeria monocytogenes.
Comments
The overlay technique allows the discrimination and specific enumeration by their haemolysis of presumptive haemolytic/pathogenic listeria colonies, avoiding the possibility of overlooking the presence of small numbers of pathogenic listeria colonies on agars with a heavy non-pathogenic listeria growth, when only a few colonies are subcultured for further biochemical identification.
This overlay technique, when used with LSAMm has proved to be an effective procedure for the measurement of Listeria monocytogenes in foods.
Recently published results (Fernandez-Garayzabal et al., 1992b) suggest that the performance of the SRBC top layer could be improved by decreasing the concentration of potassium tellurite in LSAMm to 40 mg/1, as the haemolysis of Listeria monocytogenes is slightly higher at this concentration and the selectivity of LSAMm is not affected.
Selective properties of acriflavine may vary from lot to lot and manufacturer to manufacturer. Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the optimum concentration for use, with regard to the efficiency of selectivity and absence of inhibition of Listeria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.

313
References
Blanco, M., Fernandez-Garayzabal, J.F., Dominguez, L., Briones, V., Vazquez-Boland, J.A., Blanco, J.L., Garcia, J.A. and Suarez, G. (1989) A technique for the direct identification of haemolytic-path- ogenic Listeria on selective plating media. Lett. Appl. Microbiol. 9, 125-128.
Dominguez Rodriguez, L., Suarez Fernandez, G., Fernandez-Garayzabal, J.F., and Rodriguez Ferri, E. (1984) New methodology for the isolation of Listeria microorganisms from heavily contaminated environments. Appl. Environ. Microbiol. 47, 1188-1190.
Dominguez, L., Fernandez-Garayzabal, J.F., Blanco, M., Briones, V., Vazquez-Boland, J.A., Blanco, J.L. and Suarez, G. (1990) Overlay technique for direct detection and identification of haemolytic Listeria on selective plating medium: comparison of five media. Z. Lebensm. Unters. Forsch. 191, 16-19.
Fernandez-Garayzabal, J.F., Blanco, M., Vazquez-Boland, J.A., Briones, V., Garcia, J.A., Delgado, C., Domingo, M., Marco, J. and Dominguez, L. (1992a) A direct plating method for monitoring the contamination of Listeria monocytogenes in silage (Unpublished results).
Fernandez-Garayzabal, J.F., Delgado, C., Blanco, M., Vazquez-Voland, J.A., Briones, V., Suarez, G. and Dominguez, L. (1992b) Role of potassium tellurite and brain heart infusion in expression of the haemolytic phenotype of Listeria spp. on agar plates. Appl. Environ. Microbiol. 58, 434-438.
Vazquez-Boland, J.A., Dominguez, L., Blanco, M., Rocourt, J., Fernandez-Garayzabai, J.F., Gutierrez, C.B., Tascon, R.I. and Rodriguez Ferri, E. (1992) Epidemiologic investigation of a silage-associated epizootic of ovine listeric encephalitis using a new Listeria-selective enumeration medium and phage typing. Am. J. Vet. Res. 53, 368-371.

314 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Enterobacteriaceae Enrichment (EE) broth
Description and history
The medium is, in essence, a modification of brilliant green bile lactose broth, which in turn was a modification of MacConkey's liquid medium (Mossel et al., 1963). EE broth therefore contains brilliant green and bile as the inhibitory agents for Gram positive organisms, glucose as the main energy source to give all Enterobacteriaceae an equal chance of rapid development and avoid a bias towards lactose positive types, and an increased amount of buffer to control inhibition of growth in the earlier stages of enrichment and autosterilization at the end.
Composition (grams)
Pep tone Glucose Di-sodium hydrogen o r thophospha te Potass ium di-hydrogen o r thophospha te Ox bile Brilliant green Disti l led or deionized water
10.0 5.0 6.45 2.0
20.0 0.0135
1000.0
Preparation
Dissolve the ingredients in the water, distribute in the required volumes in flasks or tubes and heat at 100~ for 30 min only. Further heating must be avoided. Do not autoclave. Cool rapidly in cold running tap water.
Physical properties
Appearance Green, clear. pH 7.2 +_ 0.2
Shelf life
Ready to use medium 1 month at 4 _+ 2~ in screw capped containers.

315
Inoculation method for samples
Use food macerates or decimal dilutions and inoculate broth in the proportion 1:10. When large volumes of enrichment fluid are not appropriate, EE broth can be used at double strength; this makes strict adherence to the instructions for heat treatment (cf. above) imperative.
Incubation method
To detect mesophilic members of the Enterobacteriaceae incubate at 30~ for 24-48 h. Psychrotrophic Enterobacteriaceae can be detected by incubation at 4~ for 10 days.
Reading of results and interpretation
Turbidity and often change of colour of the medium towards yellowish-green provides presumptive evidence of the presence of Enterobacteriaceae. Isolation on a MacConkey type agar, e.g. violet red bile (lactose) agar and subsequent confir- matory tests must be carried out.
Quality assessment
(i) Productivity Test strains Escherichia coli 50034
Hafnia aluei 50037
Inoculation method Dilution to extinction.
Criteria
(ii) Selectiuity Test strains
Recovery in EE should be within one titre unit of the recovery in tryptone soya broth after 18-24 h at 30~
Enterococcus faecalis 50030 Staphylococcus aureus 50080
Inoculation method Dilution to extinction.
Criteria Recovery in EE should be less than 5 titre units of the recovery in tryptone soya broth after 18-24 h at 30~
References
Mossel, D.A.A., Visser, M. and Cornelissen, A.M.R. (1963) The examination of foods for Enterobacte- riaceae using a test of the type generally adopted for the detection of salmonellae. J. Appl. Bacteriol. 26, 444-452.

316
Mossel, D.A.A., Harrewijn, G.A. and Nesselrooy-van Zadelhoff, C.F.M. (1974) Standardisation of the selective inhibitory effect of surface active compounds used in media for the detection of Enterobac- teriaceae in foods and water. Health Labor. Sci. 11, 260-267.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
317
FDA Listeria enrichment broth
Description and history
This medium was originally developed from Levinthal broth with acriflavine and nalidixic acid (Ralovich et al., 1971; 1972) to enrich Listeria spp. present in low numbers in raw milk (Lovett et al., 1987). Subsequently it has been modified and used for the isolation of Listeria spp. from dairy products, vegetables, meat products and seafoods (Walker et al., 1990; Cox et al., 1991; Warburton et al., 1991).
Composition (grams)
Trypticase soy bro th Yeast extract Acriflavine HC1 Nalidixic acid (sodium salt) C~cloheximide (Actidione) Distil led or deionized water
30.0 6.0 0.015 0.04 0.05
1000.0
Preparation
Dissolve the trypticase soy broth and yeast extract in the water and sterilize by autoclaving at 121~ for 15 min. Cool the medium and add the last three ingredients just before use. Acriflavine and nalidixic acid are dissolved in distilled water and filter sterilized. Cycloheximide is dissolved in 40% ethanol and water before filter sterilization.
Physical properties
Appearance Amber, translucent with greenish tinge. pH 7.3 _+ 0.2
Shelf life
Base medium 6 months at 4 _+ 2~

318
Inoculation method for samples
Add 25 g of food to 225 ml of enrichment broth. Blend or stomach as required and then transfer to a 500 ml Erlenmeyer flask.
Incubation method
At 30~ for 24 and 48 h. After incubation, the enrichment broth culture should be streaked directly onto Oxford agar and lithium chloride phenylethanol moxalactam (LPM) agar.
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152 /
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria iuanouii serovar 5 50095
Inoculation method Dilution to extinction.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 29906/CECT 4168 /NCTC 11938)
Inoculation method Dilution to extinction.
Comments
Selective properties of acriflavine may vary from lot to lot and manufacturer to manufacturer. Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the optimum concentration for use, with regard to the efficiency of selectivity and absence of inhibition of L&teria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.
References
Cox, L.J., Siebenga, A. and Pedrazzini, C. (1991) Performance of enhanced haemolysis agar compared to Oxford medium for the isolation of Listeria monocytogenes from food enrichment broths. Food Microbiol. 8, 51-62.
Lovett, J., Francis, D.W. and Hunt, J.M. (1987) Listeria monocytogenes in raw milk: Detection, incidence and pathogenicity. J. Food Protect. 50, 188-192.

319
Ralovich, B., Forray, A., Mero, E., Malovics, I., and Szazados, I. (1971) New selective medium for isolation of L. monocytogenes. Zentralbl. Bakteriol. I.Abt.Orig. 216, 88-91.
Ralovich, B., Emody, L., Malovics, I., Mero, E. and Forray, A. (1972) Methods to isolate L. monocytogenes from different materials. Acta Microbiol. Acad. Sci. Hung. 19, 367-369.
Walker, S.J., Archer, P. and Appleyard, J. (1990) Comparison of the Listeria-Tek Elisa Kit with cultural procedures for the detection of Listeria species in foods. Food Microbiol. 7, 335-342.
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Tiwari, N.P., Babiuk, T., Lacasse, P. and Read, S. (1991) A Canadian comparative study of modified versions of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes. J. Food Protect. 54, 669-696.

320 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
F r a s e r b r o t h - m o d i f i e d
Description and history
Fraser and Sperber (1988) developed the secondary enrichment broth (UVM Listeria Enrichment Broth II)used in the United States Department of Agricul- ture procedure for isolating Listeria monocytogenes from meats (McClain and Lee, 1988). The present formula is a subsequent modification with an increased amount of acriflavine. It gives a presumptive test for Listeria spp. as it contains an indicator for the hydrolysis of aesculin. This reaction is not exclusive to Listeria spp. so any microorganisms giving a positive reaction (blackening) must be isolated and further identified.
Composition (grams)
Proteose peptone (peptic digest of animal tissue) Tryptone (pancreatic digest of casein) Beef extract Yeast extract Sodium chloride Di-sodium hydrogen or thophosphate" 2H2 O Potassium di-hydrogen or thophosphate Aesculin Lithium chloride Iron (III) ammonium citrate Nalidixic acid Acriflavine Distilled or deionized water
5.0 5.0 5.0 5.0
20.0 12.0
1.35 1.0 3.0 0.5 0.02 0.025
1000.0
Preparation
Basal broth
Suspend all of the ingredients, except the acriflavine and iron (III) ammonium citrate, in the water and heat to dissolve. Dispense 10 ml portions into 16 x 150 mm culture tubes with screw cap closures and sterilize at 121~ for 15 min.

321
Supplements
To each tube of the cooled basal broth, immediately before use, add 0.1 ml portions of filter sterilized aqueous solutions of 0.25% of acriflavine and 5.0% iron (III) ammonium citrate.
Physical properties
Appearance Light straw colour, clear. pH 7.2 _+ 0.2
Shelf life
Basal broth and supplement 14 days at 4 + 2~ Use complete medium immedi- ately after addition of the supplements.
Inoculation method for samples
Food or environmental samples are cultured in a primary listeria enrichment broth and incubated at 30~ for 24 h. These cultures are gently mixed and 0.1 ml portions are added to 10 ml of complete Fraser broth.
Incubation
At 35~ for 24 and 48 h (Warburton et al., 1991) in air and in the dark.
Reading of results and interpretation
After incubation the tubes are compared to an uninoculated control against a white background. Blackened cultures are considered as presumptively positive for Listeria spp. However, confirmatory tests are necessary on all positive cultures. Cultures which retain the original straw colour are considered to be negative.
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152 /
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria ivanovii serovar 5 50095
Inoculation method Dilution to extinction.

322
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 11938)
Inocula t ion me thod Dilut ion to extinction.
Comments
Selective proper t ies of acriflavine may vary f rom lot to lot and manufac tu r e r to manufac tu re r . Each new batch must be assayed in combinat ion with o ther selective agents to be used in the med ium to de te rmine the op t imum concen t ra t ion for use, with regard to the efficiency of selectivity and absence of inhibit ion of Listeria spp. S torage of stock solutions of acriflavine for per iods of more than one mon th is not r e c o m m e n d e d .
References
Fraser, J.A. and Sperber, W.H. (1988) Rapid detection of Listeria spp. in food and environmental samples by esculin hydrolysis. J. Food Protect. 51, 762-765.
McClain, D. and Lee, W.H. (1988) Development of USDA-FSIS method for isolation of Listeria monocytogenes from raw meat and poulty. J. Assoc. Off. Anal. Chem. 71, 660-664.
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Tiwari, N.P., Babiuk, T., Lacasse, P. and Read, S. (1991) A Canadian comparative study of modified versions of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes. J. Food Protect. 54, 669-676.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
323
Giolitti and Cantoni Broth (GCB)
Description and history
Giolitti and Cantoni broth was developed for the recovery of low numbers of stressed Staphylococcus aureus in foods (Giolitti and Cantoni, 1966). It has been adopted by the International Dairy Federation for the selective enrichment of stressed Staphylococcus aureus cells in dried milk (Anon., 1978). It contains mannitol, glycine and sodium pyruvate to enhance repair of stressed Staphylococ- cus aureus cells and to promote growth. Lithium chloride and potassium tellurite inhibit competitive Gram positive and negative flora. Tellurite is reduced by Staphylococcus aureus and related species to tellurium giving the medium a black colour. Anaerobic growth conditions increase the selectivity of the medium.
Composition (grams)
Tryptone 10.0 Meat extract 5.0 Yeast extract 5.0 Lithium chloride 5.0 Mannitol 20.0 Sodium chloride 5.0 Glycine 1.2 Sodium pyruvate 3.0 Potassium tellurite 0.05 Distilled or deionized water 1000.0
Preparation
Dissolve the ingredients, except the potassium tellurite, in the water, heat and shake to obtain a complete solution. Cool to room temperature and adjust the pH to 6.9 +_ 0.1. Dispense in 19 ml amounts in 20 x 200 mm tubes. Autoclave 20 min at 115~ Before use, expel the air by heating to 100~ for 10 min. Cool and add 0.1 ml of a filter sterilized 1% (w/v) solution of potassium tellurite per tube.

324
Physical properties
Appearance pH
Clear, light brown. 6 .9+0.1
Shelf life
Basal medium Complete medium
2 weeks at 4~ Use the same day.
Inoculation method for samples
1 ml of reconstituted sample is inoculated into the GCB. Mix the inoculum gently with the broth and overlay the broth with liquid paraffin or agar.
Incubation method
At 37~ for 24-48 h.
Reading of results and interpretation
Tubes showing any blackening or black precipitate are considered presumptive positive.
Quality assessment
(i) Productivity Test strain Staphylococcus aureus 50081
Inoculation method Dilution to extinction.
Criteria
(ii) Selectivity Test strains
Recovery in GCB should be within one titre unit of recovery in tryptose soya broth.
Escherichia coli 50034 Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938)
Supplementary strain Bacillus licheniformis 50016
Inoculation method Dilution to extinction.
Criteria Recovery in GCB should be less than 5 titre units of the recovery in tryptone soya broth.

325
References
Anon. (1978) Detection of coagulase positive staphylococci in dried milk. International Dairy Federa- tion Standard 60A.
Giolitti, G. and Cantoni, C. (1966) A medium for the isolation of staphylococci from foodstuffs. J. Appl. Bacteriol. 29, 395-398.

326 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Hektoen Enteric (HE) agar
Description and history
This medium for the isolation of Shigella and Salmonella spp. was originally developed by King and Metzger (1968) of the Hektoen Institute. It relies on the use of bile salts for selective inhibition and two indicator systems: (i) bromothymol blue and acid fuchsin as indicators of carbohydrate dissimilation and (ii) ferric iron as an indicator of the formation of hydrogen sulphide from thiosulphate. HE agar allows good growth of Shigella spp. because the inhibition of these organisms by bile salts is reduced by the addition of relatively large amounts of peptone and carbohydrates. The medium provides good colonial differentiation and inhibits some co|iforms and other nonlactose-fermenting bacteria, thereby facilitating the identification of colonies of Salmonella and Shigella spp. from food products.
Composition (grams)
Proteose peptone 12.0 Yeast extract 3.0 Sodium chloride 5.0 Lactose 12.0 Sucrose 12.0 Salicin 2.0 Bromothymol blue 0.065 Acid fuchsin 0.08 Sodium thiosulphate 5.0 Iron (III) ammonium citrate 1.5 Bile salts 9.0 Agar 14.0 Distilled or deionized water 1000.0
Preparation
Suspend the components in the water. The medium is boiled for a few seconds until the ingredients are completely dissolved. Do not autoclave or overheat. Cool to 47~ and pour into sterile Petri dishes. Final pH should be 7.5 _+ 0.2.

327
For increased selectivity novobiocin can be added at a final concentration of 0.01 g/1. For this dissolve 100 mg of novobiocin in 10 ml of distilled water, sterilize by filtration and aseptically add 1 ml of this solution to the cooled medium, mix well and distribute into sterile Petri dishes.
Physical properties
Appearance Blue-green, transparent. pH 7.5 _+ 0.2
SheZf Zife
Ready to use medium 3 weeks at 4 + 2~
Inoculation method for samples
1. Subculturing after enrichment" A loopful of an incubated selective broth is streaked onto the medium to attain isolated colonies.
2. Direct counting: Surface spreading over whole plate using 0.5 ml of decimal dilutions of food homogenates.
Incubation method
At 35-37~ for 18-24 h in air.
Reading of results and interpretation
On HE agar, Salmonella spp. produce transparent green or blue-green colonies, with or without black centres or may appear as almost completely black colonies (HzS production). Shigella spp. produce green, transparent colonies. As other organisms can form colonies similar to Salmonella and Shigella spp., biochemical and serological confirmatory tests are necessary. Lactose, sucrose or salicin fer- menting Gram negative bacteria may be inhibited or may produce salmon-coloured colonies.
Quality assessment
(i) Producticity Test strains Salmonella circhow 50077
Salmonella enteritidis 50073 Shigella flexneri 50079

328
I n o c u l a t i o n m e t h o d
(ii) Selectivity Tes t s t ra in
M o d i f i e d M i l e s - M i s r a or s t r e a k i n g / e c o m e t r y .
Enterococcus faecalis 50030
I n o c u l a t i o n m e t h o d M o d i f i e d M i l e s - M i s r a or s t r e a k i n g / e c o m e t r y .
(iii) Characteristic appearance of colonies See above.
References
Andrews, W.H., Poelma, P.L. and Wilson, C.R. (1981). Comparative efficiency of brilliant green, bismuth sulfite, Salmonella-Shigella, Hektoen enteric, and xylose lysine deoxycholate agars for the recovery of Salmonella from foods: collaborative study. J. Assoc. Anal. Chem. 64, 899-928.
King, S. and Metzger, W. (1968). A new plating medium for the isolation of enteric pathogens. I. Hektoen Enteric Agar. Appl. Microbiol. 16, 577-578.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
329
Irgasan Ticarcillin Chlorate (ITC) broth
Description and history
This medium was formulated by Wauters et al. (1988) for the enrichment of Yersinia enterocolitica from meat products. It is derived from the enrichment broth for salmonellae described in 1956 by Rappaport et al.. The basic selective agents are malachite green and magnesium chloride, however in a different ratio from that used in the original medium. Selectivity for Yersinia enterocolitica is enhanced by triclosan (irgasan), ticarcillin and chlorate, the latter being inhibitory for Enterobacteriaceae possessing type A nitratase. The medium performs well for the pathogenic Yersinia enterocolitica biotype 4 serotype 0:3, but is less appropriate for other serotypes.
Composition (grams)
Tryptone Yeast extract Magnesium chlor ide. 6H2 O Sodium chloride Malachite green Potassium chlorate 2,3,4'-Trichloro-2'-hydroxyldiphenyl ether
(Irgasan, Ciba-Geigy) * Ticarcillin * Distilled or deionized water
10.0 1.0
60.0 5.0 0.01 1.0 0.001
0.001 1000.0
Preparation
Dissolve the ingredients except those marked * in water. Sterilize by autoclaving. Allow the medium to cool and add aseptically ticarcillin and irgasan. Distribute aseptically Suitable volUmes, usually 100 ml, in sterile containers. The surface should be small in comparison to the volume to minimise aeration. Erlenmeyer flasks form suitable containers.

330
Physical properties
Appearance Blue-green, clear. pH 6.8
Shelf life
Ready to use medium At least 1 month at 4 _+ 2~ in screw-capped containers.
Inoculation method for meat samples
A 20% (w/v) suspension of minced meat is prepared in peptone water and gently shaken without homogenizing by mixer or stomacher. The suspension is filtered through gauze to trap solid particles and a 5 ml volume of the filtrate is inoculated into 100 ml ITC broth. Inoculation of the meat itself, either minced or homoge- nized, results in a decreased recovery. Incubation should be carried out at 24 + I~ for 2 or 3 days.
Reading of results and interpretation
Since growth of yersiniae does not always result in obvious turbidity, the medium should always be subcultured by streaking one loopful either on SSDC agar (q.v.) (preferably) or on CIN agar.
Quality assessment
(i) Productivity Test strain Yersinia enterocolitica biotype 4, serotype 3 50087
Inoculation method Dilution to extinction.
Criteria Growth should be within 2 titre units of the growth in tryptone soya broth after 2 days, but even a slight turbidity in ITC should be recorded.
(ii) Selectivity Test strains Escherichia coli 50034
Pseudomonas aeruginosa 50067
Supplementary strains Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938) Staphylococcus aureus 50080
Inoculation method Dilution to extinction.

331
Cr i t e r i a D i f f e r e n c e in g rowth shou ld be equa l to or less t h a n 5
t i t re uni t s of t he g rowth in t r y p t o n e soya b ro th .
References
Wauters, G., Goossens, V., Janssens, M. and Vandepitte, J. (1988) New enrichment method for isolation of pathogenic Yersinia enterocolitica serogroup 0:3 from pork. Appl. Environ. Microbiol. 54, 851-854.
Rappaport, F., Konforti, N. and Navon, B. (1956) A new enrichment medium for certain salmonellae. J. Clin. Pathol. 9, 261-266.

332 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Iron sulphite agar
Description and history
The medium was first developed by Wilson and Blair (1924) to enumerate Clostridium perfringens (B. welchii) in water. The medium utilizes the ability of the genus Clostridium to reduce sulphite which reacts with the iron citrate to form ferrous sulphide, staining the colonies black (Pr6vot and Thouvenot, 1954; Skov- gaard, 1958).
Composition (grams)
Tryptone Yeast extract Sodium sulphite (anhydrous) Iron (III) ci trate. 5H2 O Distilled or deionized water
15.0 10.0 0.5 0.5
1000.0
Preparation
Suspend ingredients other than sodium sulphite and iron (III) citrate in 980 ml distilled water and boil to dissolve completely. Dispense in bottles or tubes and sterilize at 120~ for 10 min. Cool to about 50~ and add 10 ml of a 5% sodium sulphite solution and 10 ml of 5% iron (III) citrate solution.
Physical properties
Appearance Pale straw. pH 7.0 _+ 0.2
Shelf life
Ready to use medium Day of preparation.

333
Inoculation method for samples
1. Pour plate procedure, using 1 ml per 9 cm diameter plate. When set, plates are overlaid with sterile medium.
2. Inoculate 1 ml of appropriate dilutions into test tubes and pour about 12-15 ml of the finished medium into the tubes. Mix carefully and allow agar to solidify, then add a further 2-3 ml of liquified medium as a cover to prevent uptake of oxygen.
Incubation method
At 37~ for 48 h. Pour plates should be incubated anaerobically. If thermophilic clostridia are expected, incubate also at 55~
Reading of results and interpretation
Read the cultures after 1 and 2 days incubation. Discrete black colonies of 1-5 mm diameter are considered to be presumptive clostridia.
Quality assessment
(i) Productivity Test strains Clostridium sporogenes 50099
Clostridium perfringens 50027
Inoculation method Surface plating, modified Miles-Misra or streaking technique using pre-poured plates with subsequent overlay.
(ii) Selectivity Test strains Bacillus subtilis 50018
Escherichia coli 50034
Inoculation method As above.
(iii) Characteristic appearance of colonies Discrete black colonies of 1-5 mm diameter.
References
Prevot, A.R. and Thouvenot, H. (1954) Valeur de l'azide de sodium pour l'isolement des anaerobes. Ann. Inst. Pasteur, 86, 236-237.

334
Skovgaard, N. (1958). Specific enumeration of clostridia in foods. VIII Nordiska Veterinarmotet, Sektion E, rapport 2, 1-7.
Wilson, W.J. and Blair, E.M.M'V. (1924) The application of a sulphite-glucose-iron agar medium to the quantitative estimation of B. welchii and other reducing bacteria in water supplies. J. Pathol. Bacteriol. 27, 119-121.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
335
Kanamycin Aesculin Azide (KAA) agar
Description and history
The history of this medium is summarized by Mossel et al. (1978). It utilises (i) the selective inhibitory components azide and kanamycin and (ii) the indicator system aesculin and ferric iron for the isolation and differentiation of group D strepto- cocci. Growth of the majority of unwanted organisms is suppressed, while sought organisms hydrolyse aesculin, producing black zones around the colonies due to the formation of black iron phenolic compounds derived from the aglucon. The selectivity of the medium can be increased by incubation at 42 _+ 0.3~
Composition (grams)
Tryptone 20.0 Yeast extract 5.0 Sodium chloride 5.0 Sodium citrate 1.0 Aesculin 1.0 Iron (III) ammonium citrate 0.5 Sodium azide 0.15 Kanamycin sulphate 0.02 Agar No. 1 (Oxoid) 12.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients in the water. Bring to the boil to dissolve completely. Dispense into bottles (or final containers) and sterilize by autoclaving at 121~ for 15 min.
Physical properties
Appearance Light pale straw. pH 7.2 _+ 0.2

336
Shelf life
Ready to use medium 7 days at 4 + 2~
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per pre-dried 9 cm diameter plate.
Incubation method
At 37~ or 42 + 0.3~ in air for 18-24 h.
Reading of results and interpretation
Round white or grey colonies, about 2 mm diameter surrounded by black haloes are considered to be group D streptococci ( 'presumptive ' count).
Quality assessment
(i) Productivity Test strains Enterococcus faecalis 50030
Enterococcus faecium 50032 Streptococcus boris (DSM 2 0 4 8 0 / N C D O 5 9 7 / NCTC 8177)
Inoculation method Modified Miles-Misra or s t reak ing /ecomet ry .
(ii) Selectivity Test strains Escherichia coli 50034
Lactococcus lactis ssp. lactis 50058
Supplementary strains Aerococcus viridans 50012 Bacillus cereus 50014 Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or s t reak ing /ecomet ry .
(iii) Characteristic appearance of colonies
Round, white or grey, about 2 mm diameter surrounded by black haloes of at least 1 cm diameter.
Reference
Mossel, D.A.A., Bijken, P.H.G., Eelderink, I. and van Spreekens, K.A. (1978) Streptococci of Lance- field groups A, B and D and those of buccal origin in foods: their public health significance, monitoring and control. In: Streptococci edited by F.A. Skinner and L.B. Quesnel, SAB Symposium Series No. 7. Academic Press, London, pp. 315-347.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
337
KRANEP-agar (Kalium-Rhodanid Actidione Natriumazid Eigelb Pyruvat agar)
Description and history
KRANEP-agar is a selective medium for the enumeration of Staphylococcus aureus in foods which was first described by Sinell and Baumgart (1967). Selectivity is attained with potassium thiocyanate, sodium azide, lithium chloride and cyclo- heximide (Reuter, 1968). The egg yolk precipitation reaction is the diagnostic growth feature of characteristic colonies of Staphylococcus aureus. Sodium pyru- vate is an essential component for the recovery of damaged Staphylococcus aureus. Mannitol is a special growth component for most species of staphylococci.
Composition (grams)
Peptone (tryptic digest of beef) Beef extract Sodium chloride Di-sodium hydrogen or thophosphate �9 2H2 O Sodium pyruvate Potassium thiocyanate Lithium chloride Mannitol Sodium azide * Cycloheximide (Actidione) * Egg yolk emulsion (ml) * Agar Distilled or deionized water
10.0 5.0 3.0 2.0 8.2
25.5 5.1 5.1 0.05 0.041
50.0 20.0
930.0
Preparation
Suspend the ingredients except those marked * in the water and heat to boiling to dissolve completely. Adjust pH to 6.8-7.0. Heat at 100~ for 20 to 30 min. Cool to about 50~ and add freshly prepared solutions and egg yolk emulsion:
10 ml of a 0.05% membrane-filtered aqueous solution of sodium azide. 10 ml of a 0.041% membrane-filtered aqueous solution of cycloheximide.

338
50 ml of a freshly prepared egg yolk emulsion, diluted 1" 1 with 0.9% sodium chloride solution.
Physical properties
Appearance pH (complete medium)
Opaque, pale yellow. 6.8
Shelf life
Ready to use medium 7 days at 4 + 2~ 2 weeks after packaging into plastic bags at 4 + 2~ Commercial media, which contain sodium azide should be used within 2 days.
Inoculation method
Spread plate technique using 0.1 ml inocula on 9 cm diameter Petri dishes or 0.05 ml each from different decimal dilution steps in drop plate technique.
Incubation method
At 37~ for 48 h, in air, if necessary followed by incubation at room temperature for 24 h.
Reading of results and interpretation
After an incubation of 48 h well-grown golden yellow colonies with a precipitation zone of egg yolk in a medium which remains opaque are considered as Staphylo- coccus aureus. Confirmatory tests for coagulase production are required. Colonies typical for Staphylococcus aureus but without an egg yolk reaction should also be tested for coagulase and if positive their identity should be confirmed by further tests (Kusch and Reuter, 1971; Devriese and Hajek, 1980).
Quality assessment
(i) Productivity Test strains Staphylococcus aureus 50081
Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra.
Criteria Recovery on KRANEP Agar should be within 0.5 lOg l0 of the counts on the non-selective medium (Kusch and Reuter, 1971).

339
(ii) Selectivity Test strains Bacillus cereus 50014
Proteus mirabilis ( A T C C 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 11938) Staphylococcus warneri 50082
Inocu la t ion m e t h o d Modi f i ed Miles-Misra .
Cr i ter ia Recovery on K R A N E P - A g a r should be at least 3.0
lOg l0 below the recovery on non-se lec t ive agar. Pro- teus mirabilis should show no growth at all (Te rp lan et al., 1981).
(iii) Characteristic appearance of Staphylococcus au reus colonies
Well grown go lden yellow colonies up to 3 m m d i a m e t e r (at 48 h) typically s u r r o u n d e d by an egg yolk prec ip i ta t ion zone.
References
Devriese, L.A. and Hajek, V. (1980) Identification of pathogenic staphylococci isolated from animals and foods derived from animals. J. Appl. Bact. 49, 1-11.
Kusch, D. and Reuter, G. (1971) Vergleichende Untersuchungen iiber die Eignung des KRANEP- und des Baird-Parker Mediums fiir die selektive Isolierung koagulasepositiver Staphylokokken. Zbl. Bakt. I Abt. Orig. 217, 23-34.
Reuter, G. (1968) Erfahrungen mit N~ihrb6den fiir die selektive mikrobiologische Analyse von Fleisch- erzeugnissen. Arch. Lebensmittelhyg. 19, 53-57 & 84-89.
Sinell, H.-J. and Baumgart, J. (1967) Selektivn~ihrb6den mit Eigelb zur Isolierung von pathogenen Staphylokokken aus Lebensmitteln. Zbl. Bakt. I Abt. Orig. 204, 248-264.
Terplan, G., Becker, H. and Zaadhof, K.-J. (1981) Eignung gebr~iuchlicher und neuer Medien zum Nachweis von Staph. aureus and M6glichkeiten ihrer Qualit~itskontrolle. Arch. Lebensmittelhyg. 32, 126-130.

340 Culture Media for Food Microbiology, J.E.L. Corry et at (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Lactobacillus Sorbic acid (LaS) agar (syn. Sorbic acid agar base)
Description and history
LaS agar is prepared according to the formulation of Reuter (1968; 1970). The combination of anaerobic incubation and reading of the results after 2 days enables selective enumeration of 'lactobacilli' (genus Lactobacillus). Acetate, citrate, sorbic acid and low pH suppress other bacterial flora, including enterococci and Streptococcus salivarius ssp. thermophilus. Productivity for Lactobacillus del- brueckii ssp. bulgaricus may be slightly reduced. This medium is good for isolating lactobacilli from a mixed flora, but can be inhibitory for some yoghurt cultures and for sublethally damaged bacteria (Reuter, 1985).
Composition (grams)
Peptone (tryptic digest of casein) Beef extract Yeast extract Glucose Sorbitan monooleate (Tween 80) Sodium acetate �9 3 H 2 0 Tri-sodium citrate Magnesium sulphate . 7 H 2 0 * Manganese (II) sulphate- 4H 20 § Agar Sorbic acid Distilled or deionized water
10.0 10.0 5.0
20.0 1.0 5.0 3.0 0.2 0.05
20.0 0.4
1000.0
* 20 ml of 1% solution + 5 ml of 1% solution
Preparation
Suspend all the ingredients except the sorbic acid in the water and boil to dissolve completely. Heat at 100~ for 1 h. Add the sorbic acid dissolved in about 10 ml 1N NaOH, adjust pH to 5.2 with 10% HC1, boil for 5 min. Final pH 5.0 at 30~ This medium must not be autoclaved.

341
Physical properties
Appearance pH (final)
Pale with slight opalescence. 5.0 at 30~
Shelf life
Ready to use medium Up to 1 week at 4 + 2~ Up to 2 weeks in sealed plastic bags at 4 _+ 2~
Inoculation method
Surface spreading over whole plate or modified Miles-Misra.
Note: pH of a representative plate should be checked before use to confirm selective potential.
Incubation method
37~ for thermophilic and mesophilic species ] 2 days under anaerobic or 30~ for mesophilic and psychrotrophic species J microaerobic (6% O2: 10% CO2 in N 2)
conditions.
Reading of results and interpretation
All well grown colonies after 48 h incubation at 30 or 37~ are considered as lactobacilli. Pin point colonies should be ignored. Confirmation by Gram stain may be necessary to exclude some strains of lactic streptococci, leuconostocs and pediococci from certain habitats. Carnobacterium piscicola does not grow.
Quality assessment
(i) Productivity Test strains Lactobacillus gasseri 50040
Lactobacillus plantarum 50054 Lactobacillus sake 50056
Supplementary strain Lactobacillus brevis 50043
Inoculation method Modified Miles-Misra or streaking/ecometry.
Criteria Recovery on LaS agar should be within 1.0 lOg l0 of the recovery on Briggs agar after 48 h at 30 or 37~

342
(ii) Selectivity Test strains Staphylococcus aureus 50080
Saccharomyces cerevisiae 50105
Supp lemen ta ry strains Bacillus cereus 50014 Enterococcus hirae 50031 Escherichia coli 50034
Inocula t ion m e t h o d Modif ied Miles-Misra or s t r e a k i n g / e c o m e t r y .
Cri ter ia Recovery on LaS-Agar should be 5.0 log10 below the recovery on Briggs agar. Enterococcus hirae pro- duces pin-point colonies.
(iii) Characteristic appearance of colonies
Small white or greyish colonies, 0 .5-2.5 m m diameter , flat or raised, smooth or rough. Non-lact ic acid bacter ia may be dis t inguished by colony size of > 2.5 m m after p ro longed incubat ion at room t empera tu re . Enterococc i may appea r as medium-s ized colonies of 0 .5-1 m m diameter .
References
Reuter, G. (1968) Erfahrungen mit N~ihrb6den fiir die selektive mikrobiologische Analyse von Fleis- cherzeugnissen. Arch. Lebensmittelhyg. 19, 53-57 and 84-89.
Reuter, G. (1970) Laktobazillen und eng verwandte Mikroorganismen in Fleisch und Fleischerzeugnis- sen, 2. Mitteilung: Die Charakterisierung der isolierten Laktobazillenstamme. Fleischwirtsch. 50, 954-962.
Reuter, G. (1985) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
343
Lauryl tryptose broth
Description and history
A selective medium for the detection of coli-aerogenes bacteria in water, dairy products and other foods (Mallman and Darby, 1941). The surface active agent lauryl sulphate acts as the selective agent in restricting the growth of bacteria other than the coli-aerogenes group. The broth is specifically designed to allow rapid multiplication and copious gas production from a small inoculum of target organ- isms. Unlike other media it is possible to test directly for indole production.
Composition (grams)
Tryptose Lactose Sodium chloride Di-potassium hydrogen orthophosphate Potassium di-hydrogen orthophosphate Sodium lauryl sulphate Distilled or deionized water
20.0 5.0 5.0 2.75 2.75 0.1
1000.0
Preparation
Add the ingredients to the water and mix until completely dissolved. Dispense in 10 ml amounts into suitable plugged test tubes or screw cap bottles and add a Durham tube to each tube or bottle. Sterilize by autoclaving at 121~ for 15 min. To avoid bubbles in the Durham tubes, allow the autoclave temperature to drop to 75~ before opening.
Physical properties
Appearance pH
Light amber, clear to slightly opalescent. 6.8 + 0.2

344
Shelf life
Ready to use medium 1 month at 4 _+ 2~ in screw capped containers.
Inoculation method for samples
Use MPN tests for presumptive enumera t ion of coli-aerogenes bacteria and Escherichia coli as described in the American Public Heal th Association Standard Methods for water and waste water (1971), dairy products (1967) and foods (1966).
Incubation method
To detect members of the coli-aerogenes group incubate at 35 or 37~ for 24-48 h. For dairy purposes and where it is desired to detect the full range of coliform bacteria, incubate at 30~ for 24-48 h. For Escherichia coli the t empera ture of 44 _+ 0.1~ for 18 h is specifically recommended. Psychrotrophic coliforms can be detected by incubation at 4~ for 10 days.
Reading of results and interpretation
Turbidity provides presumptive evidence of the presence of coli-aerogenes bacte- ria, particularly when accompanied by gas formation. Fur ther confirmation of the presence of Escherichia coli may be obtained from the indole reaction and subculture to VRB agar (q.v.)
Quality assessment
(i) Productivity Test strains
Inoculation method
Escherichia coli50034 Hafnia aluei 50037
Dilution to extinction.
(ii) Selectivity Test strain
Inoculation method
Enterococcus faecalis 50030
Dilution to extinction.
References
American Public Health Association (1971) Standard Methods for the Examination of Water and Waste Water, 13th edn., pp. 651, 664-665.
American Public Health Association (1967) Standard Methods for the Examination of Dairy Products, 12th edn., edited by W.G. Walter, New York NY.
American Public Health Association (1966) Recommended Methods for the Microbiological Examina- tion of Foods, 2nd edn., edited by J.M. Scharf, New York NY.
Mailman, W.L. and Darby, C.W. (1941) Use of lauryl sulphate tryptose broth for the detection of coliform organisms. Am. J. Publ. Hlth, 31, 127-134.

Culture Media for Food Microbiology, J.E.L. Corrv et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
345
Levinthal broth with acriflavine and nalidixic acid
Description and history
This selective enrichment medium was originally developed to assist the isolation of Listeria spp. from faeces and nasal or vaginal discharges (Ralovich et al., 1972). Recent studies (Ralovich, 1989) have also demonstrated its effectiveness in the examination of foods containing low numbers of the organism.
After incubation of the broth subcultures may be made on any suitable selective agar.
Composition (grams)
Lab-Lemco powder (Oxoid) Bacto pep tone Sodium chloride Def ibr ina ted sheep blood (ml) Acriflavine Nalidixic acid Distilled or deionized water
10.0 10.0
5.0 50.0
0.015 0.04
1000.0
Preparation
Put all the ingredients except acriflavine and nalidixic acid into an Erlenmeyer flask and steam for one hour. Filter and adjust pH to 7.4. Sterilize by autoclaving at 121~ for 15 min. Add 1.5 ml of sterile 1% acriflavine and 20.0 ml of sterile 0.2% nalidixic acid solutions. Mix well, and dispense aseptically into tubes or flasks in convenient amounts. The broth can be stored in sealed containers.
Physical properties
Appearance pH
Amber, translucent with greenish-yellow colour. 7.3 +0.1
Shelf life
Ready to use medium 14 days in dark at 4 _+ 2~

346
Inoculation method for samples
Add 25 g of homogenized or 25 ml of liquid food to 225 ml of the enrichment broth. Swabs may be placed directly into tubes containing 5-10 ml of the enrich- ment broth.
Incubation method
At 30~ for 48 h and for a further 5 days at room temperature in the dark. Subculturing should be performed at intervals throughout the incubation.
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152/
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ CECT 935 /NCTC 10527) Listeria ivanovii serovar 5 50095
Inoculation method Dilution to extinction.
(ii) Selectivity Test strains Proteus mirabilis (ATCC 29906/CECT 4168/NCTC
11938) Enterococcus faecalis 50030
Inoculation method Dilution to extinction.
Comments
Selective properties of acriflavine may vary from lot to lot and manufacturer to manufacturer. Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the optimum concentration for use, with regard to the efficiency of selectivity and absence of inhibition of Listeria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.
References
Levinthal, W. (1918) Bakteriologische und serologische Influenzastudien. Z. Hyg. Infektionskr. 86, 1-24.
Ralovich, B., Emody, L., Malovics, I., Mero, E. and Forray, A. (1972) Methods to isolate L. monocytogenes from different materials. Acta Microbiol. Acad. Sci. Hung. 19, 367-369.
Ralovich, B. (1989) Data on the enrichment and selective cultivation of listeriae. Int. J. Food Microbiol. 8, 205-217.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
347
Lithium chloride Ceftazidime Tween 80 (LCT) agar
Description and history
LCT agar, developed by Lachica (1992) is a selective medium for the isolation and direct counting of Listeria spp. from food samples. It is an improved modification of the LC agar (Lachica, 1990a) in that it promotes the formation in 24 h instead of 40 h of listeria colonies large enough for use in a 6 h identification scheme (Lachica, 1990b). Selectivity is achieved by the use of lithium chloride and ceftazidime (Bannerman and Bille, 1988). Glycine anhydride was deleted since it was observed to enhance rather than inhibit the growth of some food microflora. Good growth is promoted by the use of brain heart infusion agar, Tween 80 and aerobic incubation. A listeria colony is differentiated from other bacterial colonies by its distinct prismatic pattern when observed under various angles of oblique transillumination (Lachica, 1991); a simplified Henry test (Lachica, 1990c) based on a fixed angle of transillumination to observe the characteristic bluish hue of listeria colonies was originally used. The original LC agar is currently used as a secondary enrichment medium in one commercial application and for growth of immuno-adsorbed listeria cells in another commercial application.
Composition (grams)
Brain hear t infusion agar
Li th ium chloride Ceftazidime (Fortaz) Sorbi tan monoolea te (Tween 80) Distil led or deionized water
47 or 52 (according to manufac turer )
5.0 0.05 1.0
1000.0
Preparation
Suspend the ingredients except ceftazidime in water, and bring to the boil to dissolve completely. Autoclave at 121~ for 15 min. Cool to 45-48~ and asepti- cally add 2.5 ml of an aqueous solution of 20 m g / m l ceftazidime which has been sterilized by filtration through a 0.2/zm membrane filter. The ceftazidime solution may be stored at -20~

348
Physical properties
Appearance pH
Transparent, light to medium amber. 7.4 + 0.2
Shelf life
Ready to use medium 4 weeks at 4 + 2~
Inoculation method for samples
Surface streaking of dry plates for isolation or modified Miles-Misra.
Incubation method
At 35~ for 24 h in air.
Reading of results and interpretation
After 24 h incubation, colonies of Listeria monocytogenes are ca. 1.5 mm in diameter with finely granular, entire to slightly undulate edge and appear white under reflected light. Depending on the angle of transmitted light, the iridescence of a listeria colony can vary from blue to red (Lachica, 1991).
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152/
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ CECT 9 3 5 / N C T C 10527)
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 29906/CECT 4168 /NCTC 11938)
Inoculation method Modified Miles-Misra or streaking/ecometry.
References
Bannermann, E.S. and Bille, J. (1988) A new selective medium for isolating Listeria spp. from heavily contaminated material. Appl. Environ. Microbiol. 54, 165-167.

349
Lachica, R.V. (1990a) Selective plating, medium for quantitative recovery of foodborne Listeria monocytogenes Appl. Environ. Microbiol. 56, 67-169.
Lachica, R.V. (1990b) Same-day identification scheme for colonies of Listeria monocytogenes Appl. Environ. Microbiol. 56, 1166-1168.
Lachica, R.V. (1990c) Simplified Henry technique for the initial recognition of Listeria colonies. Appl. Environ. Microbiol. 56, 1164-1165.
Lachica, R.V. (1991) Spectral pattern-oblique transillumination (SPOT) device for recognition of Listeria colonies. Ann. Mtg., Amer. Soc. Microbiol., Dallas, TX, May 5-9, Abstr. P-25.
Lachica, R.V. (1992) Improved selective medium for the quantitative detection of Listeria monocyto- genes in foods. Ann. Mtg., Inst. Food Technol., New Orleans, LA, June 20-24, Abstr. 74.

350 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Lithium chloride Phenylethanol Moxalactam (LPM) agar
Description and history
LPM agar was developed in 1986 to recover Listeria spp. from very mixed microflora in UVM enrichment broths (q.v.) and it is more selective for listeria than other media developed previously (Lee and McClain, 1986). LPM's selective nature is used successfully by the USDA (McClain and Lee, 1988) and the FDA (Datta et al., 1988) to enumerate > 100/g of Listeria spp. or Listeria monocyto- genes from many kinds of contaminated foods by direct plating without enrich- ment. Listeria spp. will form colonies on LPM agar in 24 h at 30~ while most, but not all, of the competing microflora are suppressed. Listeria colonies can be recognised and selected for identification using the 45 ~ transillumination of Henry (1933). Listeria spp. which have been frozen grow well on LPM agar but heat injured cells do not.
Composition (grams)
Pancreatic digest of casein Peptone (peptic digest of animal tissue) Beef extract Sodium chloride Lithium chloride Glycine anhydride * (Sigma G-7251) 2-phenylethanol (Sigma P-6134) Sodium or ammonium moxalactam Agar Distilled or deionized water
5.0 5.0 3.0 5.0 5.0
10.0 2.5 0.02
15.0 1000.0
* Note: Do not substitute glycine anhydride with glycine, which makes the LPM very inhibitory to growth of Listeria spp.
Preparation
Base
Suspend the ingredients except the moxalactam in bottles or flasks containing a magnetic stirring bar and autoclave the medium for 12 min at 121~ Mix the

351
medium gently after autoclaving. LPM is heat sensitive and so it is important to cool it promptly to 46~ in a water bath.
Moxalactam solution
Sodium or ammonium moxalactam (Eli Lilly or Sigma M-1900) 1 g Potassium buffer (0.1 M) pH 6.0 100 ml (Made by adding 0.1 M mono-potassium di-hydrogen orthophosphate solution to 0.1 M di-potassium hydrogen orthophosphate solution to reach pH 6.0). Sterilize by filtration through 0.2/zm filter. Store both the powder and 3-5 ml aliquots of the 1% moxalactam in a freezer at -60~
Complete medium
Add 2 ml 1% moxalactam solution per litre of the autoclaved and cooled molten base while stirring with a magnetic mixer. Dispense 12 ml per 9 cm Petri dish.
Physical properties
Appearance Almost clear and colourless. pH 7.3 _+ 0.2
Shelf life
Dehydrated agar base
Ready to use medium
Store at 2-8~ to prevent loss of selectivity. Use before manufacturer's expiry date. 2-3 weeks at 4 _+ 2~ It is essential to store plates in sealed plastic bags as the phenylethanol is volatile.
Inoculation method for samples
Surface streaking or modified Miles-Misra. For rapid semiquantitative detection of > 100/g of Listeria spp. in foods, swab the surface of solid samples with a moistened (dry for liquid samples) swab, and inoculate half of a LPM plate. Streak the other half of the plate with a sterile loop (McClain and Lee, 1988). Liquid samples can be concentrated by centrifugation before testing.
Incubation method
LPM plates are usually placed in thin polyethylene plastic bags and incubated at 30~ in air for 20-24 h for isolation, and 40 h for counting colonies without magnification.

352
Reading of results and interpretation
Under optimum 45 ~ transillumination the more isolated and larger (24 h old) listeria colonies appear as whitish piles of crushed glass often showing mosaic-like internal structures occasionally having blue-grey iridescent areas. Smaller, more crowded, colonies have a more pronounced blue-grey iridescence that tends to sparkle. When growth becomes near confluent an even blue-grey iridescent sheen can be observed. For quantitative determination, count all the colonies that grow on LPM agar in 40 h and then identify the percentage of these colonies that are Listeria spp. or Listeria monocytogenes. For semiquantitative estimation, simply compare the density of all the colonies in the swabbed area with standard density photos of 102, 103, 104, 105, 106 and 107 cfu/ml of Listeria monocytogenes swabbed and streaked on LPM agar (similar to Oxoid Dip Slide) and then identify from the streaked area the percentage of colonies as Listeria spp. or Listeria monocytogenes. Alternatively, scan the swab and streak plates at 24 h using 45 ~ transillumination, and quantitatively enumerate the positive samples using the decimal dilution plating technique.
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152 /
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria iuanovii serovar 5 50095
Inoculation method Modified Miles-Misra.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 29906/CECT 4168 /NCTC 11938)
Inoculation method Modified Miles-Misra.
(iii) Characteristic appearance of colonies See above.
References
Datta, A.R., Wentz, B.A., and Hill, W.E. (1988) Identification and enumeration of beta-haemolytic Listeria monocytogenes in naturally contaminated dairy products. J. Assoc. Off. Anal. Chem. 71, 673-675.

353
Domjan Kovacs, H. and Ralovich, B. (1991) Model examination of selective media for isolation of Listeria strains. Acta Microbiol. Hung. 38, 141-145.
Henry, B.S. (1933) Dissociation in the genus Brucella. J. Infect. Dis. 52, 374-402. Lee, W.H. and McClain, D. (1986) Improved Listeria monocytogenes selective agar. Appl. Environ.
Microbiol. 52, 1215-1217. McClain, D. and Lee, W.H. (1988) Development of the USDA-FSIS method for the isolation of
Listeria monocytogenes. J. Assoc. Off. Anal. Chem. 71, 660-664.
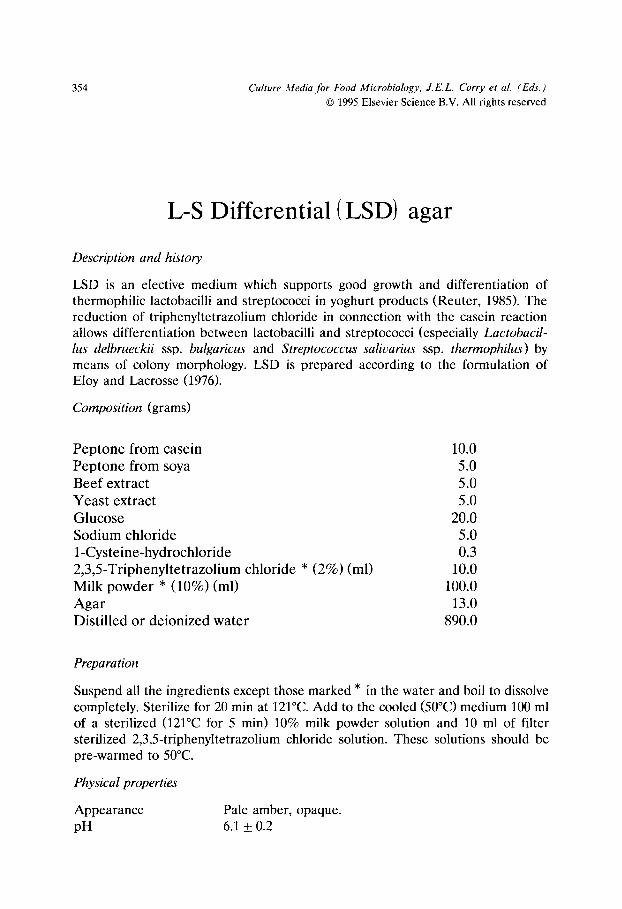
354 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
L-S Differential (LSD) agar
Description and history
LSD is an elective medium which supports good growth and differentiation of thermophilic lactobacilli and streptococci in yoghurt products (Reuter, 1985). The reduction of triphenyltetrazolium chloride in connection with the casein reaction allows differentiation between lactobacilli and streptococci (especially Lactobacil- lus delbrueckii ssp. bulgaricus and Streptococcus saliuarius ssp. thermophilus) by means of colony morphology. LSD is prepared according to the formulation of Eloy and Lacrosse (1976).
Composition (grams)
Peptone from casein Peptone from soya Beef extract Yeast extract Glucose Sodium chloride 1-Cysteine-hydrochloride 2,3,5-Triphenyltetrazolium chloride * (2%) (ml) Milk powder * (10%) (ml) Agar Distilled or deionized water
10.0 5.0 5.0 5.0
20.0 5.0 0.3
10.0 100.0
13.0 890.0
Preparation
Suspend all the ingredients except those marked * in the water and boil to dissolve completely. Sterilize for 20 min at 121~ Add to the cooled (50~ medium 100 ml of a sterilized (121~ for 5 min) 10% milk powder solution and 10 ml of filter sterilized 2,3,5-triphenyltetrazolium chloride solution. These solutions should be pre-warmed to 50~
Physical properties
Appearance Pale amber, opaque. pH 6.1 +0.2

355
Shelf life
Ready to use medium 10 days at 4 ___ 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
At 43~ for 48 h in air.
Reading of results and interpretation
Brilliant red colonies with clear halo can be considered as lactic streptococci (Streptococcus salivarius ssp. thermophilus and Lactococcus lactis ssp. lactis) whilst pink colonies with a red centre are likely to be Lactobacillus delbrueckii ssp. bulgaricus. Lactobacillus acidophilus and other lactobacilli from milk sources may appear as red colonies with an opaque halo.
Quality assessment
(i) Productivity Test strains Streptococcus salivarius ssp. thermophilus 50083
Lactococcus lactis ssp. lactis 50058 Lactobacillus delbrueckii ssp. bulgaricus 50050 Lactobacillus acidophilus 50101
Inoculation method Modified Miles-Misra or s t reaking/ecometry .
Criteria Recovery on LSD agar should be within 1.0 lOg l0 of the recovery on MRS agar or Briggs agar for Strepto- coccus salivarius ssp. thermophilus.
(ii) Characteristic appearance of colonies
Brilliant red colonies with clear halo"
Pink colonies with red centre"
Red colonies with opaque halo:
Streptococcus salivarius ssp. thermo- philus Lactococcus lactis ssp. lactis and ssp. diacetilactis. Lactobacillus delbrueckii ssp. bulgari- CUS.
Lactobacillus acidophilus and other lactobacilli from milk sources.

356
Unwanted species may be distin- guished by colony appearance after further incubation at room tempera- ture.
References
Eloy, C. and Lacrosse, R. (1976) Composition d'un milieu de culture destine ~ effectuer le denombre- ment des micro-organismes thermophiles du yoghourt. Bull. Rech. Agron. Gemblou 11, 83-86.
Reuter, G. (1985) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
357
Lysine-Iron-Cystine-Neutral Red (LICNR) broth
Description and history
Lysine iron cystine neutral red broth was developed by Hargrove et al. (1971) as a presumptive test for salmonellae in dairy products. This medium was further investigated and modified by Hoben et al. (1973)who found it useful for a variety of foods and food ingredients. Salmonellae decarboxylate lysine but do not ferment salicin and hence produce an alkaline reaction. In addition they produce hydrogen sulphide from cystine which reacts with the ferric salt to give a black precipitate of ferric sulphide. D'Aoust (1977) and Morgan-Jones (1982) have both reported that the colour change could only be used as a guide and the presence of salmonellae should be confirmed by conventional methods.
Composition (grams)
L-Lysine 10.0 Tryptone 5.0 Yeast extract 3.0 Mannitol 5.0 Glucose 1.0 Salicin 1.0 Iron (III) ammonium citrate (brown) 0.5 Sodium thiosulphate 0.1 L-Cystine 0.1 Neutral red 0.025 Distilled or deionized water 1000.0
Preparation
Suspend all the ingredients in the water and heat to dissolve, mix well and adjust pH to 6.2. Dispense into tubes in appropriate quantities. Either use immediately or sterilize at 121~ for 15 min.
Physical properties
Appearance Red, clear. pH 6.2 + 0.2

358
Shelf life
Ready to use medium 7 days at 4 + 2~ in the dark.
Inoculation method for samples
Inoculate broths with liquid food or macerates of food in appropriate volumes of the appropriate decimal dilution.
Using a Most Probable Number (MPN) method this medium can be used to enumerate salmonellae in a food, the accuracy required determining whether 3, 5 or 10 tubes are inoculated at each decimal dilution.
Incubation method
At 37~ for 24-48 h in air.
Reading of results and interpretation
The colour change of the medium to yellow together with a black deposit, provides presumptive evidence for the presence of salmonellae. Those broths showing the colour change should be confirmed by streaking a loopful of the broth on a medium selective for salmonellae and confirming characteristic colonies by bio- chemical and serological tests.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella typhimurium 50100 Salmonella virchow 50077
Inoculation method Dilution to extinction.
Criteria
(ii) Selectivity Test strains
Recovery in LICNR should be within 1 titre unit of the recovery in tryptone soya broth with colour change to yellow and the formation of a black precipitate after 48 h at 37~
Escherichia coli 50034 Pseudomonas aeruginosa 50067
Inoculation method Dilution to extinction.

359
Cr i t e r i a G r o w t h is u n i n h i b i t e d in L I C N R . A t 24 h no co lour
c h a n g e shou ld be s een wi th e i t h e r o rgan i sm. A f t e r
48 h Escherichia coli may tu rn the i nd i ca to r faint ly
o r a n g e bu t no c h a n g e shou ld be o b s e r v e d wi th Pseu- domonas aeruginosa. N o black p r e c i p i t a t e shou ld be
fo rmed .
References
D'Aoust, J.Y. (1977) Limitations on lysine-iron-cystine-neutral red broth in the presumptive identifica- tion of salmonella. Appl. Environ. Microbiol. 34, 595-596.
Hargrove, R.C., McDonough, F.C. and Reomen, R.H. (1971) A selective medium and presumptive procedure for detecting salmonella in dairy products. J. Milk Food Technol. 34, 6-11.
Hoben, D.A., Ashton, D.H. and Peterson, A.C. (1973) A rapid presumptive procedure for the detection of Salmonella in foods and food ingredients. Appl. Microbiol. 25, 123-129.
Morgan-Jones, S.C. (1982) A method for enumerating salmonellas from environments in the poultry industry. In: Isolation and identification methods for food poisoning organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner. Academic Press, London, pp. 83-90.

360 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1994 Elsevier Science B.V. All rights reserved
M 17 agar
Description and history
M 17 agar is an elective medium for the detection and differentiation of lactic streptococci and is prepared according to Terzaghi and Sandine (1975). The special buffering capacity of disodium-/3-glycerophosphate enables better growth of the lactic streptococci (Reuter, 1985). The addition of lactose allows differentiation between lactose-positive streptococci and lactose-negative mutants. Bacteriophages can also be detected by plaque formation. The medium is used for cultivation procedures in plasmid demonstration and molecular based investigations.
Composition (grams)
Peptone from casein 5.0 Peptone from meat 5.0 Peptone from soya 5.0 Yeast extract 2.5 Lactose 5.0 Disodium-/3-glycerophosphate 19.0 Magnesium sulphate 0.25 Ascorbic acid 0.5 Agar 11.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients in the water and boil to dissolve completely. Sterilize for 20 min at 121~
Physical properties
Appearance Light amber, clear. pH (final) 7.2 _+ 0.2
Shelf life
Ready to use medium 7 days at 4 _+ 2~

361
Inoculation method
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
At 37~ for 48 h in air. For mesophilic strains incubation at 30~ for 48 h in air is recommended.
Reading of results and interpretation
M 17 agar is an elective medium that gives good colony counts and a characteristic colony size for thermophilic streptococci, lactococci and enterococci. Other micro- organisms must be excluded by specific colony appearance and by confirmation tests e.g. Gram stain and catalase test.
Quality assessment
(i) Productivity Test strains Streptococcus salivarius ssp. thermophilus 50083
Lactococcus lactis ssp. lactis 50058 Enterococcus faecalis 50030
Inoculation method Modified Miles-Misra.
Criteria Recovery on M17 agar should be within 1.0 lOgl0 of the recovery on Briggs agar or another non selective medium.
(ii) Characteristic appearance of lactic streptococci Usually well-grown white colonies. Size: Streptococcus salivarius ssp. thermophilus and Lactococcus lactis with
subspecies except ssp. cremoris > 3 mm diameter lactose-negative mutants < 1.0 mm diameter enterococci 1.0-1.5 mm diameter unwanted species < 1.0 mm diameter
References
Reuter, G. (1985) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68. Terzaghi, B.E. and Sandine, W.E. (1975) Improved medium for lactic streptococci and their bacterio-
phages. Appl. Microbiol. 29, 807-813.

362 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
de Man, Rogosa and Sharpe (MRS) agar
Description and history
MRS agar was developed by de Man, Rogosa and Sharpe (1960) primarily for the cultivation of lactobacilli from various sources with the intention of producing a defined medium as a substitute for tomato juice agar. It may be used for the cultivation of the whole group of lactic acid bacteria. The medium shows good productivity for nearly all lactic acid bacteria but the original version is not selective. It may be made selective for lactic acid bacteria by lowering the pH to 5.7 and the addition of 0.14% sorbic acid (see MRS-S agar). Some strains from dairy sources may show reduced growth rates (see Briggs agar).
Composition (grams)
Peptone (tryptic digest of casein) Beef extract Yeast extract Glucose Sorbitan monooleate (Tween 80) Di-potassium hydrogen or thophosphate Magnesium sulphate . 7 H 2 0 * Manganese (II) sulphate �9 4 H 2 0 § Ammonium citrate Sodium acetate �9 3 H 2 0 Agar Distilled or deionized water
10.0 8.0 4.0
20.0 1.0 2.0 0.2 0.05 2.0 5.0
15.0 1000.0
* 20 ml of 1% solution + 5 ml of 1% solution
Preparation
Suspend the ingredients in the water and boil to dissolve completely. Sterilize for 15 min at 121~
Physical properties
Appearance pH
Light amber, clear. 6.2 Adjust pH after dissolving completely at about 50~

363
Shelf life
Ready to use medium 14 days at 4 _+ 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
This depends on the particular habitat of the organisms to be cultivated. Dairy strains should be incubated at 30~ for 2 days followed by 1 day at 22~ meat strains at 25~ for 2 days, intestinal or yoghurt strains for 2 days at either 37 or 42~ All incubations should be performed under anaerobic or microaerobic (5% 02: 10% CO2 in N 2) conditions.
Reading of results and interpretation
MRS agar is an elective medium that gives good colony counts and a characteristic colony size and morphology for lactobacilli and for other lactic acid bacteria. Other microorganisms must be excluded by specific colonial appearance and by confirma- tion tests e.g. Gram-stain and catalase test. Carnobacterium spp. either show weak growth or fail to grow.
Quality assessment
(i) Productivity Test strains Lactobacillus gasseri 50040
Lactobacillus sake 50056
Inoculation method Modified Miles-Misra or streaking/ecometry.
Criteria Recovery on MRS agar should be within 1.0 log10 of the recovery on Briggs agar.
(ii) Characteristic appearance of colonies
Usually small greyish white colonies, flat or raised, smooth, rough or interme- diate.
Reference
de Man, J.C., Rogosa, M. and Sharpe, M.E. (1960) A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130-135.

364 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
de Man, Rogosa and Sharpe agar with sorbic acid (MRS-S agar)
Description and history
MRS agar was developed by de Man, Rogosa and Sharpe (1960) primarily for the cultivation of lactobacilli from various sources with the intention of producing a defined medium as a substitute for tomato juice agar. It may be used for the cultivation of the whole group of lactic acid bacteria. The medium results in good productivity for nearly all lactic acid bacteria but it is not selective in the original version. Some strains from milk sources may show reduced growth rates (see Briggs agar). It becomes selective for lactic acid bacteria if the pH is lowered to 5.7 and sorbic acid is added in a concentration of 0.14% (= 0.2% potassium sorbate) (ISO, 1984; Reuter, 1985).
Composition (grams)
Peptone (tryptic digest of casein) Beef extract Yeast extract Glucose Sorbitan monooleate (Tween 80) Di-potassium hydrogen or thophosphate . 3H2 O Magnesium sulphate. 7 H 2 0 * Manganese (II) sulphate �9 4 H 2 0 + Ammonium citrate Sodium acetate �9 3 H 2 0 Sorbic acid Agar Distilled or deionized water
10.0 8.0 4.0
20.0 1.0 2.0 0.2 0.05 2.0 5.0 1.4
15.0 1000.0
* 20 ml of 1% solution + 5 ml of 1% solution
Preparation
Suspend the ingredients except the sorbic acid in the water and boil to dissolve completely. Heat at 100~ for 1 h. Add the sorbic acid in about 10 ml 1N NaOH,

365
adjust pH to 5.8 with 10% HC1 and boil for 5 min. Final pH should be 5.7 at 30~ This medium must not be autoclaved.
Physical properties
Appearance Light amber, clear. pH 5.7 at 30~
Shelf life
Ready to use medium 7 days at 4 _+ 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
This depends on the particular habitat of the organisms to be cultivated. Dairy strains should be incubated at 30~ for 2 days followed by 1 day at 22~ meat strains at 25~ for 3 days, intestinal or yoghurt strains for 2 days at either 37 or 42~ All incubations should be performed under anaerobic or microaerobic (5% O2: 10% CO 2 in N 2) conditions.
Reading of results and interpretation
Reading the results should be performed after a defined incubation time (see above). All well grown colonies are considered as lactic acid bacteria. Some enterococci may show reduced growth. Some yeast strains may occur with delayed growth. Carnobacterium spp. either show weak growth or fail to grow.
Quality assessment
(i) Productivity Test strains Lactobacillus gasseri 50040
Lactobacillus sake 50056 Pediococcus damnosus 50065
Supplementary strains Lactococcus lactis ssp. lactis 50058 Leuconostoc mesenteroides 50060
Inoculation method Modified Miles-Misra or streaking/ecometry.
Criteria Recovery on MRS-S agar should be within 1.0 lOg l0 of recovery on MRS agar. Lactococcus lactis ssp. lactis produces pin-point colonies.

366
(ii) Selectivity Test strains Staphylococcus aureus 50080
Bacillus cereus 50014
Supplementa ry strain Escherichia coli 50034
Inoculat ion me thod Modif ied Miles-Misra or s t r e ak ing / ecome t ry .
Criteria Recovery on MRS-S should be 5.0 lOg l0 below the recovery on MRS. Staphylococcus aureus produces pin-point colonies.
(iii) Characteristic appearance of colonies
Small greyish-white colonies, flat or raised, smooth, rough or in termediate .
References
de Man, J.C., Rogosa, M. and Sharpe, M.E. (1960) A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130-135.
ISO/TC 34/SC 6/WG 15, No. 3 and No. 5 (1984). Draft report: Enumeration of Lactobacteriaceae in meat and meat products.
Reuter, G. (1985) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
367
Mannitol Egg Yolk Polymyxin (MEYP) agar
Description and history
This medium was devised by Mossel et al. (1967) for the isolation of Bacillus cereus. It relies on the inhibitory component polymyxin and two indicator systems, mannitol and phenol red and egg yolk. Growth of many unwanted organisms is suppressed. Bacillus cereus does not attack mannitol but dissimilates egg yolk and consequently gives rise to typical bacilliform colonies with purple-red zones and white haloes.
Since its introduction, MEYP agar has been adopted as a productive medium by Lancett et al. (1980) and Harmon et al. (1984) and is popular in both the United States and Europe (Kramer and Gilbert, 1989).
Composition (grams)
Peptone 10.0 Meat extract 1.0 Sodium chloride 10.0 Mannitol 10.0 Phenol red 0.025 Polymyxin B (i.u.) * 100 000 Egg yolk emulsion * - 2 0 % (ml) 100.0 Agar 15.0 Distilled or deionized water 900.0
Preparation
Suspend the ingredients except those marked * in 900 ml of water. Bring to the boil to dissolve completely and sterilize by autoclaving at 121~ for 15 min. Cool to about 50~ and add the polymyxin B suspended in 2 ml of sterile distilled water and 100 ml of a sterile 20% egg yolk emulsion, using aseptic precautions through- out.
Physical properties
Appearance Light pink, opaque. pH 7.2 _+ 0.2

368
Shelf life
Ready to use medium 4 days at 4 + 2~
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per 9 cm diameter plate.
Incubation method
At 30~ for 24-30 h in air.
Reading of results and interpretation
Crenated colonies, about 5 mm in diameter, on a distinct red background and surrounded by an intense precipitate are virtually always isolates of Bacillus cereus.
Quality assessment
(i) Productivity Test strain
Inoculation method
(ii) Selectivity Test strains
Bacillus cereus 50014
Modified Miles-Misra or streaking/ecometry.
Escherichia coli 50034 Micrococcus luteus 50063
This medium can also be used as the bottom layer, with mDRCM (q.v.) as the top layer, for the enumeration of Clostridium species in foods. For this purpose phenol red is replaced by bromocresol purple (25 mg 1-1) and incubation is anaerobic (Weenk et al. 1995).
References
Kramer, J.M. and Gilbert, R.J. (1989) Bacillus cereus and other Bacillus species. In: Foodhome Bacterial Pathogens edited by M.P. Doyle. Marcel Dekker, New York, pp. 21-70.
Harmon, S.M., Kautter, D.A. and McClure, F.D. (1984) Comparison of selective plating media for enumeration of Bacillus cereus in foods. J. Food Protect. 47, 65-67.
Lancett, G.A. and Harmon, S.M. (1980) Enumeration and confirmation of Bacillus cereus in foods. J. Assoc. Offic. Anal. Chem. 63, 581-586.
Mossel, D.A.A., Koopman M.J. and Jongerius, E. (i967) Enumeration of Bacillus cereus in foods. Appl. Microbiol. 15, 650-653.
Weenk, G.H., van den Brink, J.A., Struijk, C.B. and Mossel, D.A.A. (1995) Modified methods for the enumeration of spores of Clostridium species in dried foods. Int. J. Food Microbiol. in press.
Inoculation method Modified Miles-Misra or streaking/ecometry.
(iii) Characteristic appearance of colonies
Crenated, about 5 mm in diameter on a distinctly red background and surrounded by a copious egg yolk precipitate.
Note

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
369
Mannitol Lysine Crystal violet Brilliant green (MLCB) agar
Description and history
MLCB agar is a selective and diagnostic agar for the isolation of salmonellae, other than Salmonella typhi and Salmonella paratyphi, from faeces and foods. First described by Inoue et ai. (1968), it was not until van Schothorst et al. (1987) reported favourably on the use of the medium that it became better known. The inhibitory properties of the medium are conferred by the inclusion of brilliant green and crystal violet. The recognition of salmonella colonies depends on the fermention of mannitol and the decarboxylation of lysine together with an indica- tor system (sodium thiosulphate and iron (III) ammonium citrate) for hydrogen sulphide detection. The medium is not suitable for the isolation of Salmonella typhi or Salmonella paratyphi, because of the inhibitory concentration of brilliant green, nor for other brilliant green-sensitive strains (Curtis and Clarke, 1994). Van Schothorst et al. (1987) found the medium excellent and with enhanced selectivity for the isolation of hydrogen sulphide positive salmonellae after enrichment in Rappaport-Vassiliadis broth containing soya peptone (RVS) in place of tryptone. Salmonellae grow as large purple-black colonies due to H zS production. Atypical salmonellae that produce little or no H zS grow as mauve-grey colonies and may develop a central black "bull's-eye". Contaminating organisms may grow as small colourless colonies although some strains of Citrobacter species may mimic the appearance of Salmonella spp. and some Proteus spp. may swarm (van Schothorst et al., 1987; Bridson, 1990).
Composition (grams)
Yeast extract 5.0 Pep tone 10.0 'Lab-Lemco ' powder 2.0 Sodium chloride 4.0 Manni to l 3.0 L-Lysine hydrochlor ide 5.0 Sodium thiosulphate 4.0 Iron (III) a m m o n i u m citrate 1.0 Brilliant green 0.005 * Crystal violet 0.01 Agar 15.0 Distilled or deionized water 1000.0

370
* concentra t ion varies with formulat ion and activity of dye - see note at the end of this monograph .
Preparation
Brilliant green solution
Prepare 0.5% w/v aqueous solution of brilliant green.
Crystal violet solution
Prepare 0.5% w/v aqueous solution of crystal violet.
Complete medium
Suspend all the ingredients except brilliant green and crystal violet in the water. Add 1 ml brilliant green solution and 10 ml crystal violet solution. Mix and heat very carefully to boiling with frequent agitation to dissolve the ingredients com- pletely. Cool to 50~ and aseptically adjust the pH value to 6.8 _+ 0.2. Pour 15-20 ml quantities into 9 cm diameter Petri dishes and allow to set. Dry the prepared plates before use.
Do not autoclave or overheat the medium.
Physical properties
Appearance Violet, clear. pH 6.8 _+ 0.2
Shelf life
Ready to use medium 5 days at 4 _+ 2~
Inoculation method for samples
Surface streaking of the sample, preferably after enrichment in a salmonella selective enrichment broth such as RVS broth, to obtain well-isolated colonies. A heavy inoculum may be advantageous.
Incubation method
At 35 (or 37)~ for 18-24 h in air.
Reading of results and interpretation
Typical strains of salmonellae produce large purple-black colonies. Atypical (non- H2S-producing) strains grow as mauve-grey colonies and may develop a central black "bull's eye". Contaminating organisms may grow on the medium as small colourless colonies.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077

371
Supplementary strains
Inoculation method
(ii) Selectivity Test strains
Salmonella saintpaul 50075 Salmonella typhimurium (NCTC 12190)
Modified Miles-Misra or s t r eak ing /ecomet ry .
Escherichia coli 50034 Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4168 / NCTC 11938) - qualitative test for control of swarming.
Inoculation method Modified Miles-Misra or s t r eak ing /ecomet ry .
(iii) Characteristic appearance of colonies Large purple-black colonies, 2 -3 mm in diameter.
References
Bridson, E.Y. (1990) MLCB agar. In "The Oxoid Manual", 6th edition, pp 2-157-158. Curtis, G.D.W. and Clarke, L.A. (1994) Comparison of the MSRV method with an in-house conven-
tional method for the detection of Salmonella in various high and low moisture foods. (Corresp.) Lett. Appl. Microbiol. 18, 239-240.
Inoue, T., Takagi, S., Ohnishi, A., Tamura, K. and Suzuki, A. (1968) Food-borne disease Salmonella isolation medium (MLCB). Paper presented at the 66th Meeting of the Society Veterinary Science of Japan.
van Schothorst, M., Renaud, A. and van Beek, C. (1987) Salmonella isolation using RVS broth and MLCB agar. Food Microbiol. 4, 11-18.
Specification for brilliant green
Bacteriological per formance
Brilliant green should suppress the spreading of most Proteus species while not inhibiting the growth of salmonellae (other than Salmonella typhi and Salmonella paratyphi and other brilliant green-sensitive strains).
Method of test
Prepare MLCB agar containing various concentrat ions of brilliant green viz. 4.5 rag/1 to 6 rag/1.
Procedure
Inoculate a set of plates with different brilliant green concentrat ions with a pure culture of a swarming Proteus sp. and another set with a pure culture of a Salmonella sp. and incubate these plates at 37~ for no longer than 24 h.
A satisfactory concentrat ion of the dye should allow typical growth of salmonellae and limited growth of proteus. Prepare the brilliant green solution to contain one thousand times the final concentrat ion in the complete medium and add 1 m l / l of medium e.g. if the selected concentrat ion is confirmed as 0.005 g/1 prepare 0.5% w / v solution of brilliant green.

372 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
M-Enterococcus (ME) agar
Description and history
In 1957 Slanetz and Bartley described this improved version of the original medium of Slanetz et al. (1955). The medium relies upon the selective inhibitory properties of sodium azide and the incorporation of tetrazolium which most organisms growing on this medium will reduce to some extent under the conditions provided. Although devised originally for use with membrane filters, the medium can also be used for direct plating. Selectivity is increased by incubation at 37~ for 4 h followed by 44 + l~ for 44 h.
Composition (grams)
Tryptose Yeast extract Glucose Di-sodium hydrogen o r thophospha te . 2H2 O Sodium azide 2,3,5-Triphenyltetrazolium chloride Agar Distilled or deionized water
20.0 5.0 2.0 4.0 0.4 0.1
10.0 1000.0
Preparation
The first five ingredients are dissolved in the water and the pH value adjusted to 7.2. The agar is then added and the solution heated sufficiently to dissolve the agar. When cooled to about 50~ add 1 ml of 1% filter-sterilized 2,3,5-triphenylte- trazolium chloride solution per 100 ml of molten medium. Do not autoclave or overheat this medium.
Physical properties
Appearance Pale straw. pH 7.2 _+ 0.2
Shelf life
Ready to use medium 7 days at 4 _+ 2~

373
Inoculation method for samples
Surface spreading over whole pla te using 0.1 ml per pre -dr ied 9 cm plate.
Incubation method
At 37~ for 48 h or at 37~ for 4 h followed by 44 _+ 1~ for 44 h in air.
Reading of results and interpretation
Round , pink to dark maroon-co lou red colonies, 0 .5 -3 m m diameter , are consid-
e red to be Lancef ie ld Group D streptococci ( including enterococci) .
Quality assessment
(i) Productiuity Test strains Enterococcus faecalis 50030
Enterococcus faecium 50032
Inocula t ion m e t h o d Modif ied Miles-Misra or s t r e a k i n g / e c o m e t r y .
(ii) Selectiuity Test strains Lactococcus lactis ssp. lactis 50058
Escherichia coli 50034
Supp lemen ta ry strains Bacillus cereus 50014 Staphylococcus aureus 50080
Inocula t ion m e t h o d Modif ied Miles-Misra or s t r e a k i n g / e c o m e t r y .
(iii) Characteristic appearance of colonies
Round , pink to dark maroon-co lou red colonies, 0 .5-3 m m in d iameter .
References
Burkwall, M.K. and Hartman, P.A. (1964) Comparison of direct plating media for the isolation and enumeration of enterococci in certain frozen foods. Appl. Microbiol. 12, 18-23.
Slanetz, L.W. and Bartley, C.H. (1957) Numbers of enterococci in water, sewage and faeces determined by the membrane filter technique with an improved medium. J. Bacteriol. 74, 591-595.
Slanetz, L.W., Bent, D.F. and Bartley, C.H. (1955) Use of membrane filter technique to enumerate enterococci in water. Publ. Hlth. Reports 70, 67-72.
Taylor, E.W. and Burman, N.P. (1964) The application of membrane filtration techniques to the bacteriological examination of water. J. Appl. Bacteriol. 27, 294-303.

374 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Muller Kauffmann tetrathionate broth
Description and history
This medium is used for the selective isolation of salmonellae. Selectivity is conferred by tetrathionate (from the reaction of thiosulphate and iodine) and by the addition of ox bile and brilliant green which are modifications introduced by Kauffmann (1935). The medium has been the subject of European trials (Edel and Kampelmacher, 1968; 1969) and is used in International Standards Organisation (ISO) methods (Anon., 1975; 1981; van Leusden et al., 1982).
Composition (grams)
Meat extract 4.5 Peptone 9.0 Sodium chloride 2.7 Calcium carbonate 40.5 Sodium thiosulphate �9 5 H 2 0 50.0 Iodine 4.0 Potassium iodide 5.0 Ox bile desiccated 5.0 Brilliant green 0.01 * Distilled or deionized water 1072.0
* concentration varies with activity of dye.
Preparation
Base
Add the meat extract, peptone, sodium chloride and calcium carbonate or the dehydrated complete base to 900 ml water and boil until completely dissolved. Sterilize at 121~ for 20 min.
Sodium thiosulphate solution
Dissolve the sodium thiosulphate in about 50 ml of water. Dilute to 100 ml. Sterilize at 121~ for 20 min.

375
Iodine solution
Dissolve 25 g potassium iodide in a minimal volume of water and add 20 g iodine. Dilute to 100 ml. Store the solution in a tightly closed container. Add 20 ml to the cooled base.
Brilliant green solution
Add 0.5 g brilliant green to 100 ml water. Store the solution for at least one day in the dark to allow auto-sterilization to occur. Add 2 ml to the cooled base.
Ox bile solution
Dissolve 10 g ox bile in 100 ml water by boiling. Sterilize at 121~ for 20 min. Add 50 ml to the cooled base.
Complete medium
Using aseptic precautions add the other ingredients (in the order listed above) to the base. Mix well. Distribute required volumes aseptically into sterile containers of appropriate size, keeping calcium carbonate suspended during distribution.
Physical properties
Appearance
pH
Pale green milky opaque suspension which on standing gives a pale liquid over a heavy precipitate. 7.4 +0 .2
Shelf life
Ready to use medium 7 days at 4 _+ 2~ (Anon., 1981)
Inoculation method for samples
1. Where damaged cells are sought transfer food-inoculated and incubated pre- enrichment medium (PEM) into MK tetrathionate broth in ratio 1 part PEM to 10 parts tetrathionate broth. Use 10 ml PEM to 100 ml tetrathionate broth (or 1 ml to 10 ml).
OF
2. Add comminuted sample to medium in the ratio 1 part sample to 9 parts medium e.g. 25 g food to 225 ml medium.
Incubation method
At 42 +_ I~ for 24-48 h in air.

376
Reading of results and interpretation
Inoculated incubated medium is subcultured after 24 and 48 h onto selective diagnostic agar in such a way as to obtain well-isolated colonies. Suspect colonies are subcultured and their identity confirmed by appropriate biochemical and serological tests.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077
Inoculation method Dilution to extinction.
Criteria Recovery in Muller-Kauffmann tetrathionate broth should be within 3 titre units of the recovery in tryptone soya broth after 48 h at 42~
(ii) Selectivity Test strains Enterobacter cloacae (ATCC 23355)
Pseudomonas aeruginosa 50067
Inoculation method Dilution to extinction.
Criteria Recovery in Muller-Kauffmann tetrathionate broth should be less than 5 titre units of the recovery in tryptone soya broth after 48 h at 42~
Comments
This medium may be markedly influenced by the following:
1. Source of ingredients or dehydrated complete medium 2. The presence of food or other organic material 3. The physiological state of the organisms sought 4. The competing microflora 5. Temperature of incubation - 43~ may be very inhibitory to salmonellae; 41.5~
may be preferred 6. Possibly by combinations of the above
Successful performance of the medium tested in one set of standard conditions may not be achieved if any of those conditions are altered.

377
References
Anon. (1975) International Organization for Standardization. Meat and meat products - detection of salmonellae (Reference method). ISO 3565-1975(E).
Anon. (1981) International Organization for Standardization. Microbiology - General guidance on methods for the detection of Salmonella. ISO 6579(E).
Anon. (1985) International Organization for Standardization. Milk and milk products - Detection of Salmonella. ISO 6785-1985(E).
Edel, W. and Kampelmacher, E.H. (1968) Comparative studies on Salmonella isolations in eight European laboratories. Bull. Wld. Hlth. Org. 39, 487-491.
Edel, W. and Kampelmacher, E.H. (1969) Salmonella isolation in nine European laboratories using a standardized technique. Bull. Wld. Hlth. Org., 41, 297-306.
Kauffmann, F. (1935) Weitere Erfahrungen mit dem kombinierten Anreicherungsverfahren fiir Salmo- nellabacillen. Ztschr. Hyg., 117, 26-32.
van Leusden, F.M., van Schothorst, M. and Beckers, H.J. (1982) The standard Salmonella isolation method. In: Isolation and identification methods for food poisoning organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner. SAB Technical Series, No. 17. Academic Press, London, pp. 35-49.

378 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Oleandomycin Polymyxin Sulphadiazine Perfringens Agar (OPSPA)
Description and history
This medium was devised by Handford (1974). It depends upon (i) the selective inhibitory properties of oleandomycin, polymyxin and sulphadiazine and (ii) an indicator system involving sulphite and ferric iron. Most unwanted organisms are suppressed while Clostridium perfringens and related species will reduce the sulphite and form black colonies due to the production of ferrous sulphide (Mead et al., 1982).
Composition (grams)
Tryptose Soya peptone Yeast extract Sodium metabisulphite Iron (III) ammonium citrate Sulphadiazine Oleandomycin phosphate Polymyxin B sulphate (i.u.) Agar Distilled or deionized water
15.0 5.0 5.0 1.0 1.0 0.1 0.0005
10 000 15.0
1000.0
Preparation
Suspend ingredients other than the oleandomycin and polymyxin in 1 litre of distilled water. Bring to the boil to dissolve completely. Dispense 100 ml amounts into bottles (or final containers) and sterilize at 121~ for 15 min. Concentrated stock solutions of the antibiotics are made by dissolving 0.5 g of oleandomycin phosphate in 100 ml of sterile distilled water and by dissolving 500,000 i.u. of polymyxin B sulphate in 5 ml of sterile distilled water. These concentrated solutions are further diluted 1 in 100 and just before pouring 1 ml of each diluted solution is added to the molten basal medium.

379
Physical properties
Appearance pH
Pale straw. 7.6 + 0.2
Shelf life
Ready to use medium Concentrated antibiotic solutions Diluted antibiotic solutions
Day of preparation. 2 months at 4 _+ 2~ 1 week at 4 _+ 2~
Inoculation method for samples
Pour-plate procedure, using 1 ml per 9 cm diameter plate. When set, plates are overlaid with sterile medium.
Incubation method
At 37~ for 18-24 h anaerobically.
Reading of results and interpretation
Discrete black colonies of 1-5 mm diameter are considered to be presumptive Clostridium perfringens.
Quality assessment
(i) Productivity Test strains Clostridium perfringens (haemolytic) 50027
Clostridium perfringens (non-haemolytic) 50028
Inoculation method Surface plating using modified Miles-Misra or ecom- etry, using pre-poured plates with subsequent over- lay.
Criteria Recovery on OPSPA of typical colonies (see below) should be within 0.7 log10 of the recovery on Rein- forced Clostridial Medium after 18-24 h at 37~ anaerobically.
(ii) Selectivity Test strains Clostridium bifermentans 50026
Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938)

380
Inoculation method As (i) above.
Criteria Recovery on OPSPA should be less than 5 lOg l0 of the recovery on Reinforced Clostridial Medium after 18-24 h at 37~ anaerobically.
(iii) Characteristic appearance of colonies
Discrete black colonies of 1-5 mm diameter.
References
Handford, P.M. (1974) A new medium for the detection and enumeration of Clostridium perfringens in food. J. Appl. Bacteriol. 37, 559-570.
Mead, G.C., Adams, B.W., Roberts, T.A. and Smart, J.L. (1982) Isolation and enumeration of Clostridium perfringens. In: Isolation and identification methods for food poisoning organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner. SAB Technical Series No. 17. Academic Press, London, pp. 99-110.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
381
Oxford agar
Description and history
The medium was developed by Curtis et al. (1989a) for the isolation of Listeria monocytogenes from faeces and other clinical samples. It has also been recom- mended for the isolation of Listeria spp. from foods (Anon., 1990; Warburton et al., 1991a; 1991b). It utilises (i) the selective inhibitory components lithium chlo- ride (Ludlam, 1949), acriflavine, colistin, fosfomycin, cefotetan and cycloheximide and (ii) the indicator system aesculin and ferric iron for the isolation and differen- tiation of listeria. Listeria spp. hydrolyse aesculin, producing black zones around the colonies due to the formation of black iron phenolic compounds derived from the aglucon. Gram negative bacteria are completely inhibited. Most unwanted Gram positive species are suppressed but some coagulase negative staphylococci may appear as aesculin negative colonies. Some strains of enterococci grow poorly and exhibit a weak aesculin reaction, usually only after 40 h incubation.
Typical Listeria monocytogenes colonies are usually visible after 24 h but incuba- tion should be continued for a further 24 h to detect slow growing strains.
Composition
Columbia agar base 39.0 Aesculin 1.0 Iron (III) ammonium citrate 0.5 Lithium chloride 15.0 Cycloheximide 0.4 Colistin 0.02 Acriflavine (Sigma) 0.005 Cefotetan 0.002 Fosfomycin 0.01 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients except the cefotetan and fosfomycin in the water. Bring to the boil to dissolve completely. Sterilize by autoclaving at 121~ for 15 min. Cool to 50~ then add the cefotetan and fosfomycin aseptically as filter-sterilized solutions.

382
Physical properties
Appearance Clear, yellow. pH 7.0 + 0.2
Shelf life
Ready to use medium 14 days in the dark at 4 + 2~
Inoculation method for samples
As the medium is highly selective a heavy inoculum should be spread over the whole plate.
Incubation method
At 30 or 37~ in air for 48 h. Where Listeria seeligeri or Listeria ivanovii are sought incubation at 30~ has been shown to give better results (Curtis et al., 1989b).
Reading of results and interpretation
After 24 h colonies of Listeria monocytogenes are small (1 mm) and surrounded by black haloes. Incubation should be continued for a further 24 h to allow slow growing strains to develop. Confirmatory tests are necessary as other Listeria species may grow with similar colonial appearances.
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar l / 2 a (ATCC 35152/
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria ivanovii serovar 5 50095
Supplementary strain Listeria monocytogenes sv 4a (ATCC 19114/NCTC 5214)
Inoculation method Modified Miles-Misra.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938)

383
Supplementary strains Bacillus cereus 50014 Micrococcus luteus 50063 Pseudomonas aeruginosa 50067 Staphylococcus warneri 50082
Inoculation method Modified Miles-Misra.
(iii) Characteristic appearance of colonies
L&teria monocytogenes forms 1 mm diameter black colonies surrounded by black haloes after 24 h. At 48 h colonies are 2 -3 mm in diameter, black with a black halo and sunken centre. Other Listeria species show a similar appear- ance. When examined before 24 h growth of Listeria spp. is sometimes apparent but without the characteristic blackening. Some strains of species other than Listeria monocytogenes are inhibited on this medium when incu- bated at 37~ Incubation at 30~ will allow growth of these strains.
Note:
Where greater selectivity is required, e.g. in the examination of environmental samples, the cefotetan content may be increased (up to 0.01 g/ l ) . However, this will result in slower growth of Listeria spp. and incubation for 48 h becomes essential.
Comments
Selective propert ies of acriflavine may vary from lot to lot and manufac turer to manufacturer . Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the opt imum concentrat ion for use, with regard to the efficiency of selectivity and absence of inhibition of L&teria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.
References
Anon. (1990) FDA Bacteriological Analytical Manual, Chapter 29. Listeria isolation; Culture Medium Substitution in Method of Analysis. Federal Register (Docket No. 88D-0367) 55, 38753-38754.
Curtis, G.D.W., Mitchell, R.G., King, A.F. and Griffin, E.J. (1989a) A selective differential medium for the isolation of Listeria monocytogenes. Lett. Appl. Microbiol. 8, 95-98.
Curtis, G.D.W., Nichols, W.W. and Falla, T.J. (1989b) Selective agents for listeria can inhibit their growth. Lett. Appl. Microbiol. 8, 169-172.
Domjan Kovacs, H. and Ralovich, B. (1991) Model examination of selective media for isolation of Listeria strains. Acta Microbiol. Hung. 38, 141-145.
Ludlam, G.B. (1949) A selective medium for the isolation of Staph. aureus from heavily contaminated material. Monthly Bull. Min. Hlth. Publ. Hlth. Lab. Ser. 8, 15-20.

384
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Hunt, T., Messier, S., Plante, R., Tiwari, N.P. and Vinet, J. (1991a) A comparative study of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes in foods. Int. J. Food Microbiol. 13, 105-118.
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Tiwari, N.P., Babiuk, T., Lacasse, P. and Read, S. (1991b) A Canadian comparative study of modified versions of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes. J. Food Protect. 54, 669-676.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
385
Oxford agar-Modified (MC)
Description and history
Modified Oxford agar (MC) is similar to Oxford agar (q.v.) developed by Curtis et al. (1989a). In MC the lithium chloride content is reduced to 12 g/1 to permit the growth of some of the more sensitive Listeria monocytogenes strains (e.g. ATCC 19111 and CDC F4561). Ceftazidime (20 mg/1) replaces the cycloheximide, acri- flavine, cefotetan and fosfomycin used in Oxford agar. Limited experience has indicated that Staphylococcus spp. and fungal growth from food samples are not a problem with MC. However, the Listeria monocytogenes strain ATCC 19111 and some strains of Listeria seeligeri and Listeria ivanovii are inhibited on MC at 35~ (Curtis et al., 1989b). Thus incubation at 30~ is preferred for the optimal recovery of Listeria spp. on this medium. A suitable MC agar should support good growth of Listeria monocytogenes strain ATCC 19111 at 30~ and suppress the growth of the more resistant strains of Enterococcus faecalis (e.g. FSIS 16a).
Composition (grams)
Columbia agar base (Oxoid CM331) Aesculin Iron (III) a m m o n i u m citrate Li th ium chloride Colistin, me thane su lphonate (Sigma) or sulphate Ceftazidime pen tahydra te (Glaxo) Distilled or deionized water
39.0 1.0 0.5
12.0 0.01 0.02
1000.0
Preparation
Suspend all the ingredients except ceftazidime in a bottle or flask containing a magnetic stirring bar and autoclave the medium for 12 min at 121~ Mix the medium after autoclaving and cool to 46~ Add 2 ml of filter sterilized 1% ceftazidime solution per litre of the autoclaved, cooled molten base while stirring with a magnetic mixer. Dispense 12 ml per 9 cm diameter Petri dish.

386
Physical properties
Appearance
pH
Light yellow or tan. Dark colour indicates overheating and decomposition of aesculin. 7.2 + 0.2
m
Shelf life
Ready to use plates 14 days at 4 + 2~
Inoculation methods for samples
1)
2)
3)
From enrichment broths, soak a sterile cotton swab in the enrichment broth and swab it on half of an MC agar plate, then streak for isolation with a sterile loop in two 90 ~ directions. For direct plating and counting of > 100/g of Listeria spp. in foods, make decimal dilutions of the foods and spread plate 0.1 ml of each dilution on to MC agar. For the rapid detection of > 100/g of Listeria spp. growing in many kinds of refrigerated foods, wipe a cotton swab moistened with sterile pH 7.2 PBS or similar phosphate buffer over the interface of vacuum packaged refrigerated foods or just under the surface of soft ripened cheeses. Swabs may also be dipped into food homogenates. MC agar plates should then be inoculated by the swab and streak procedure described above. Isolated listeria-like colonies on the swab and streak plate can be recognised and picked for further identification. The number of listeria colonies on the swab and streak MC plates can be estimated by comparing the density of colonies on the inoculated area with that of standard inocula in a similar manner to the Dip Slide procedure of Guttmann and Naylor (1967).
Incubation method
At 30~ for 26 h and 40 h in air.
Reading of results and bzterpretation
Typical Listeria spp. form distinctive small (1 mm)white hemi-spherical colonies after 26 h incubation at 30~ which can be recognized easily with some practice. It is best to ignore the blackening or black zones on MC agar entirely when working with mixed cultures as this can be caused by other bacteria and become confusing. In limited observations, no additional Listeria monocytogenes colonies were recov- ered from MC agar after 26 h incubation. The 40 h incubation is used to detect other, slow growing, Listeria spp. Additional confirmatory tests are necessary to identify various Listeria spp.

387
Quality assessment
(i) Productivity Test strains Listeria ivanovii serovar 5 50095
Listeria monocytogenes serovar 1 /2a (ATCC 35152 / NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527)
Supplementary strain Listeria monocytogenes sv 4a (ATCC 1 9 1 1 4 / N C T C 5124)
Inoculat ion me thod Modif ied Miles-Misra.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 11938)
Supplementa ry strain Staphylococcus aureus 50080
Inoculat ion me thod Modif ied Miles-Misra.
References
Curtis, G.D.W., Mitchell, R.G., King, A.F. and Griffin, E.J. (1989a) A selective differential medium for the isolation of Listeria monocytogenes. Lett. Appl. Microbiol. 8, 95-98.
Curtis, G.D.W., Nichols, W.W. and Falla, T.J. (1989b) Selective agents for listeria can inhibit their growth. Lett. Appl. Microbiol. 8, 169-172.
Guttmann, D. and Naylor, G.R.E. (1967) Dip Slide: an aid to quantitative urine culture in general practice. Br. Med. J. 3, 343-345.

388 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1993 Elsevier Science B.V. All rights reserved
Oxytetracycline Glucose Yeast extract (OGY) agar
Description and history
A selective medium for the enumeration of moulds and yeasts in foods. Mossel et al. (1962; 1970) showed that this medium gave better recovery of fungi from a variety of foodstuffs than media which rely on low pH to suppress bacterial growth. It is not suitable for use with certain high protein foods. In addition, it may not restrict adequately the growth of fast spreading moulds. It neither selects for specific groups of fungi, nor is diagnostic for detecting specific mycotoxin produc- ers.
Composition (grams)
Yeast extract 5.0 Glucose 20.0 Biotin 0.0001 Oxytetracycline 0.1 Agar 12.0 Distilled or deionized water 1000.0
Preparation
Suspend the basal ingredients in the water and bring gently to the boil to dissolve completely. Sterilize by autoclaving at 115~ for 10 min. Cool to 50~ and aseptically add 10 ml of oxytetracycline solution (10 mg/ml). Mix well and dispense 15 ml amounts into 9 cm diameter Petri dishes. Use immediately or store at 4 + 2~ until required.
Physical properties
Appearance
pH
Medium amber, slightly opalescent without significant precipitate. 7.0 + 0.2 at 25~

389
Shelf life
Ready to use medium 7 days at 4 +_ 2~
Inoculation method for samples
Surface spreading over whole plate using 0.1 or 0.2 ml per 9 cm diameter plate.
Incubation
At 25 _+ 0.5~ for 5 days. Where identification is required, prolong incubation until characteristic colonies are formed.
Reading of results and interpretation
Where separate counts of moulds and yeasts are required, identify the two groups of microorganisms by morphological appearance and, where necessary, micro- scopic examination.
Quality assessment
Moulds Use stab inoculation procedure. within 30% of the figure given.
Growth rate should be
Test strains CMI ATCC
Rhizopus stolonifer Aspergillus fla vus Penicillium cyclopium
Growth rate (mm per day
at 25~
61269 91856ii 19759 16025
25.0 8.6 3.9
Yeasts Use stab inoculation procedure. within 30% of the figure given.
Growth rate should be
Test strain
Saccharomyces cerevisiae
Growth rate (mm per day
at 25~
50105 2.5
Bacteria Use streaking procedure (Appendix I, method B) with a loopful (10 txl) from a 24 h broth culture. No growth should be evident on sectors C, D or E of the streaked plate after incubation at 25~ for 5 days.

390
Test strain
Bacillus subtilis 50018
References
Mossel, D.A.A., Visser M. and Mengerink, W.H.J. (1962) A comparison of media for the enumeration of moulds and yeasts in foods and beverages. Lab. Prac. 11, 109-112.
Mossel, D.A.A., Kleynen-Semmeling, A.M.C., Vincentie, H.M., Beerens, H. and Catsaras, M. (1970) Oxytetracycline-glucose-yeast extract agar for selective enumeration of moulds and yeasts in foods and clinical material. J. Appl. Bacteriol., 33, 454-457.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
391
Phenol red brilliant green agar (modified brilliant green agar)
Description and history
This is a selective and diagnostic agar for the isolation, following pre-enrichment and selective enrichment, of salmonellae other than Salmonella typhi from foods and feeds. Emanating from the Netherlands (Edel and Kampelmacher, 1968; 1969) this formulation has been widely assessed in Europe. It is used in the method of the International Standards Organization (Anon., 1975; 1981; 1985).
Composition (grams)
Meat extract 5.0 Peptone 10.0 Yeast extract 3.0 Di-sodium hydrogen orthophosphate 1.0 Sodium di-hydrogen orthophosphate 0.6 Lactose 10.0 Sucrose 10.0 Phenol red 0.09 Brilliant green 0.005 * Agar 15.0 Distilled or deionized water 1000.0
concentration varies with formulation and activity of dye - see note at end of this monograph.
Preparation
Base
Dissolve the meat extract, peptone, yeast extract and buffer salts in 900 ml water by boiling. Adjust the pH value so that it is 7.0 +_ 0.1 at 25~ after sterilization. Sterilize the base at 121~ for 15 min.

392
Sugar/phenol red solution
Dissolve the ingredients in 100 ml water. Heat in a waterbath at 70~ for 20 min. Cool to 55~ and use immediately.
Brilliant green solution
Dissolve 0.5 g brilliant green in 100 ml water. Store the solution for at least one day in the dark to allow auto-sterilization to occur.
Complete medium
Under aseptic conditions add 1 ml brilliant green solution to the sugar/phenol red solution cooled to about 55~ Add to the base at 55~ and mix. Distribute quantities of about 40 ml of the freshly prepared medium cooled to 45~ into sterile 14 cm diameter Petri dishes. If large dishes are not available pour about 15 ml into 9 cm Petri dishes.
Physical properties
Appearance Red, clear. pH 6.9 _+ 0.2
Shelf life
Ready to use medium 4 days at 4~ + 2~
Inoculation method for samples
Streak inoculation from selective enrichment broth by method specified by ISO 6579 1981(E) (see also van Leusden et al., 1982). Other streak techniques which yield well-isolated colonies may also be used.
Incubation method
At 37~ for 20-24 h in air.
Reading of results and interpretation
Typical colonies of Salmonella spp. are mostly smooth, very low convex, moist p ink/red. Subculture for further biochemical and serological tests.

393
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077
Supplementary strains Salmonella dublin 50072 Salmonella saintpaul 50075
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Proteus mirabilis (ATCC 29906/CECT 4168/NCTC
11938) - qualitative test for control of swarming. Escherichia coli 50034
Supplementary strain Morganella morgani 50064
Inoculation method Modified Miles-Misra or streaking/ecometry.
(iii) Characteristic appearance of colonies
Assess only on well-isolated colonies. Colonies of Salmonella are red, sur- rounded by bright red medium.
References
Anon. (1975) International Organization for Standardization. Meat and meat products - detection of Salmonella. (Reference method). ISO 3565-1975(E).
Anon. (1981) International Organization for Standardization. Microbiology - General guidance on methods for the detection of Salmonella. ISO 6579-1981(E).
Anon. (1985) International Organization for Standardization. Milk and milk products - detection of Salmonella. ISO 6785-1985.
Edel, W. and Kampelmacher, E.H. (1968) Comparative studies on Salmonella isolation in eight European laboratories. Bull. Wld. Hlth. Org. 39, 487-491.
Edel, W. and Kampelmacher, E.H. (1969) Salmonella isolation in nine European laboratories using a standardized technique. Bull. Wld. Hlth. Org. 41, 297-306.
van Leusden, F.M., van Schothorst, M. and Beckers, H.J. (1982) The standard Salmonella isolation method. In: Isolation and identification methods for food poisoning organisms edited by J.E.L. Corry, D. Roberts and F.A. Skinner. SAB Technical Series, No. 17. Academic Press, London, pp. 35-49.

394 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Specification for brilliant green
Bacteriological performance
Brilliant green should suppress the spreading of Proteus spp. while not inhibiting the growth of salmonellae (other than Salmonella typhi, Salmonella paratyphi and other brilliant green-sensitive strains).
Method of test
Prepare phenol red brilliant green agar containing various concentrations of brilliant green viz. 4.5 mg/1 to 6 mg/1.
Procedure
Inoculate a set of plates with different brilliant green concentrations with a pure culture of a swarming Proteus and another set with a pure culture of a Salmonella and incubate these plates at 37~ for no longer than 24 h.
A satisfactory concentration of the dye should allow growth of salmonellae with typical pink colonies, 1 to 2 mm in diameter, and limited growth of proteus i.e. no spreading.
Prepare the brilliant green solution to contain one thousand times the concentra- tion in the complete medium and add 1 ml/1 of medium e.g. if the selected concentration is confirmed as 0.005 g/l , prepare a 0.5% w/v solution of brilliant green.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
395
Polymyxin Acriflavine Lithium Chloride Ceftazidime Aesculin Mannitol (PALCAM)
agar
Description and history
PALCAM, developed by van Netten et al. (1989), is a selective differential medium for the isolation and direct counting of Listeria spp. from faecal and biological specimens as well as food and heavily contaminated environmental samples. It is an improved modification of RAPAMY (van Netten et al., 1988a), ALPAMY (van Netten et al., 1988b) and Oxford (Curtis et al., 1989) media. Selectivity is achieved by the use of microaerobic incubation and the inhibitory components lithium chloride, acriflavine, polymyxin and ceftazidime. Its diagnostic properties are based on the use of two indicator systems: (i) aesculin and ferric iron and (ii) D-mannitol combined with phenol red. The selective system aims to suppress virtually all unwanted organisms, including most Enterococcus spp. The double diagnostic system allows easy distinction of Listeria spp. from enterococci and staphylococci. Listeria spp. are aesculin positive, but mannitol negative, producing green colonies with black haloes against a pink-purple background. When direct counting is applied to foods processed for safety, solid medium repair must precede the use of PALCAM.
Composition (grams)
Columbia agar 39.0 Glucose 0.5 Mannitol 10.0 Aesculin 0.8 Iron (III) ammonium citrate 0.5 Phenol red 0.08 Polymyxin B * (i.u.) 100000.0 Acriflavine HC1 * 0.005 Lithium chloride 15.0 Ceftazidime * 0.02 Distilled or deionized water 1000.0

396
Preparation
Suspend the above ingredients except those marked * in 960 ml water and bring to the boil to dissolve completely. Adjust pH to 7.2 + 0.1. Sterilize by autoclaving at 121~ for 15 min. Cool to 47~ and aseptically add the required amount of the following filter-sterilized solutions" (i) 10 ml of 0.1% polymyxin B sulphate (Pfizer), (ii) 10 ml of 0.05% ethanolic acriflavine (Sigma) and (iii) 20 ml of 0.116% sodium ceftazidime pentahydrate (Glaxo). Mix gently and pour about 40 ml of freshly prepared medium into sterile 14 cm diameter Petri dishes or 15 ml into 9 cm dishes.
Physical properties
Appearance Pink-purple, opaque. pH 7.2 _+ 0.1
Shelf life
Ready to use medium At least 4 weeks at 4 _+ 2~
Inoculation method for samples
Dry plates for 20 min at 55~ to remove surface moisture prior to use.
1. Subculturing after enrichment A loopful of an incubated non-selective or selective enrichment broth is streaked onto PALCAM to attain isolated colonies.
2. Direct counting Spread plate technique using 0.1 ml of decimal dilutions of food homogenates on 9 cm diameter Petri dishes or 1.0 ml on 14 cm dishes. Spread inoculum over the whole plate until surface appears dry.
Incubation method
At 30~ for 24 to 48 h, microaerobically in a jar containing 5-12% CO2, 5-15% O 2 in N 2.
Reading of results and interpretation
After microaerobic incubation allow plates tO regain their pink-purple colour by exposure to air for 1 h. Well-isolated greenish, smooth colonies of approximately 1.5 to 2 mm with black haloes against pink-purple background are virtually always Listeria spp. Further biochemical and serological identification is performed ac- cording to recognised microbiological procedures.

397
Quafity assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1 /2a (ATCC 35152 /
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria ivanovii serovar 5 50095
Inoculation method Modified Miles-Misra or s t r eak ing /ecomet ry .
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 11938)
Inoculation method Modified Miles-Misra or s t r eak ing /ecomet ry .
(iii) Characteristic appearance of colonies
Smooth green colonies, 1.5-2 mm in diameter sometimes with black centres, but always with black haloes are Listeria spp.
White or yellow colonies, 1.5-3 mm in diameter with yellow haloes are staphylococci.
Round smooth white-grey colonies < 1 mm in diameter with green haloes are Enterococcus spp.
Comments
Selective propert ies of acriflavine may vary from lot to lot and manufac turer to manufacturer . Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the opt imum concentrat ion for use, with regard to the efficiency of selectivity and absence of inhibition of L&teria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.
References
Curtis, G.D.W., Mitchell, R.G., King, A.F. and Griffin, E.J. (1989) A selective differential medium for the isolation of Listeria monocytogenes. Lett. Appl. Microbiol. 8, 95-98.
van Netten, P., van der Ven, A., Perales, I. and Mossel, D.A.A. (1988a) A selective and diagnostic medium for use in the enumeration of Listeria spp. in foods. Int. J. Food Microbiol. 6, 187-198.

398
van Netten, P., Perales, I. and Mossel, D.A.A. (1988b) An improved selective and diagnostic medium for isolation and counting of Listeria spp. in heavily contaminated foods. Lett. Appl. Microbiol. 7, 17-21.
van Netten, P., Perales, I., Curtis, G.D.W. and Mossel, D.A.A. (1989) Liquid and solid selective differential media for the detection and enumeration of L. monocytogenes and other Listeria spp. Int. J. Food Microbiol. 8, 299-316.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
399
Polymyxin Acriflavine Lithium Chloride Ceftazidime Aesculin Mannitol Egg Yolk
(L-PALCAMY) broth
Description and history
L-PALCAMY developed by van Netten et al. (1989), is a selective differential enrichment medium used in procedures for the detection and isolation of Listeria spp. from faecal and biological specimens as well as food and heavily contaminated environmental samples. The selectivity of L-PALCAMY is dependent on the inhibitory action of lithium chloride, acriflavine, polymyxin and ceftazidime. The diagnostic traits of L-PALCAMY are based on the use of (i) aesculin and ferric iron and (ii) D-mannitol combined with phenol red. The selective system aims to suppress virtually all unwanted organisms, including most Enterococcus spp. Pres- ence of aesculin positive and mannitol negative Listeria spp., after enrichment, is often indicated by a brown-black colour.
Composition (grams)
Special peptone (Oxoid) Yeast extract powder Lab lemco (Oxoid) Peptonized milk (Oxoid) Sodium chloride Mannitol Aesculin Iron (III) ammonium citrate Phenol red Polymyxin B * (i.u.) Acriflavine HC1 * Lithium chloride Ceftazidime * Egg yolk emulsion * (ml) Distilled or deionized water
23.0 5.0 5.0 5.0 5.0 5.0 0.8 0.5 0.08
100 000.0 0.005
10.0 0.03
25.0 1000.0

400
The egg yolk emulsion may be omitted.
Preparation
Suspend the above ingredients except those marked * in 925 ml water and bring to the boil to dissolve completely. Adjust pH to 7.2 + 0.1. Sterilize by autoclaving at 121~ for 15 min. Cool to 47~ and aseptically add the egg yolk emulsion if required and the following filter-sterilized solutions: (i) 10 ml of 0.1% polymyxin B sulphate (Pfizer), (ii) 10 ml of 0.05% ethanolic acriflavine (Sigma) and (iii) 30 ml of 0.116% sodium ceftazidime pentahydrate (Glaxo). Mix gently and distribute in 90 ml or 225 ml volumes in sterile flasks.
Physical properties
Appearance Pink-purple, opaque. pH 7.2 + 0.1
Shelf life
Ready to use medium 2 days at 4 + 2~
Inoculation method for samples
Use macerates of foods, faeces, biological or environmental samples and inoculate broth in proportion 1" 10.
Incubation method
At 30~ for 24 to 48 h, in air.
Reading of results and interpretation
Often change of colour of the medium to brown-black provides presumptive evidence for the presence of Listeria spp. However if only small numbers of Listeria spp. are present this colour change may not be produced and it is therefore essential to subculture all broths onto PALCAM agar (q.v.).
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152 /
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria ivanovii serovar 5 50095

401
Inoculation method Dilution to extinction.
Criteria Recovery in L - P A L C A M Y should be within one titre unit of the recovery in Columbia broth after 48 h at 30~
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 11938)
Inoculation method Dilution to extinction.
Criteria Recovery in L - P A L C A M Y should be less than four titre units of the recovery in Columbia broth after 48 h at 30~
Comments
Selective propert ies of acriflavine may vary from lot to lot and manufac turer to manufacturer . Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the opt imum concentrat ion for use, with regard to the efficiency of selectivity and absence of inhibition of Listeria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.
References
Anon. (1987) Testing methods for use in quality assurance of culture media. Int. J. Food Microbiol. 5, 291-296.
van Netten, P., Perales, I., Curtis, G.D.W. and Mossel, D.A.A. (1989) Liquid and solid selective differential media for the detection and enumeration of L. monocytogenes and other Listeria spp. Int. J. Food Microbiol. 8, 299-316.

402 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Polymyxin Pyruvate Egg Yolk Mannitol Bromothymol blue agar (PEMBA)
Description and history
A diagnostic selective medium formulated by Holbrook and Anderson (1980) for the enumeration of Bacillus cereus in foods. Selectivity is attained with polymyxin and a critical concentration of nutrients. Cycloheximide may be used to inhibit the growth of moulds. Bacillus cereus is identified by colony form, colour and egg yolk hydrolysis and rapidly confirmed by cell and spore morphology.
Composition (grams)
Peptone Mannitol Magnesium sulphate. 7 H 2 0 Sodium chloride Di-sodium hydrogen orthophosphate Potassium di-hydrogen orthophosphate Bromothymol blue (water soluble) Egg yolk emulsion 20% (ml) Polymyxin (i.u.) Sodium pyruvate Cycloheximide (Actidione) Agar Distilled or deionized water
1.0 10.0
0.1 2.0 2.5 0.25 0.12
50.0 100000.0
10.0 0.04
15.0 1000.0
Commercially available egg yolk emulsion may be used or it may be prepared by Billing and Luckhurst's method (1957).
Preparation
Basal agar
Add components except egg yolk and antibiotics to the water, soak then steam to dissolve. Adjust pH, dispense in 90 ml amounts and autoclave at 121~ for 15 min.

403
Complete medium
Melt and cool base to 50~ Aseptically add egg yolk emulsion 5 ml; 100,000 i.u./ml polymyxin, 1 ml; 20% w / v sodium pyruvate, 5 ml and 0.4% w / v cyclohex- imide, 1 ml. Mix and pour in approximately 12 ml amounts into 9 cm diameter Petri dishes.
Physical properties
Appearance pH
Apple green, turbid almost opaque. 7.2 _+ 0.2
Shelf life
Prepared basal medium 3 months at 20~ Commercial media containing pyruvate may have shorter shelf life.
Complete medium 4 days at 4 + 2~ or 1 day at 20~
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per 9 cm Petri dish.
Incubation method
At 37~ for 18-24 h in air, then if necessary at room temperature until next day.
Reading of results and interpretation
Bacillus cereus grow as crenate, fimbriate or slightly rhizoid colonies up to 5 mm diameter (at 24 h) turquoise to peacock blue in colour with flat ground glass surface and surrounded by a precipitate from hydrolysed egg yolk. At 48 h colonies are peacock blue with raised greyish centre. Occasionally weak or negative egg yolk reacting strains may be isolated. Bacillus cereus is indistinguishable from Bacillus thuringiensis. Bacillus megaterium and Bacillus coagulans are completely inhibited. Bacillus subtilis and Bacillus licheniformis are readily differentiated from Bacillus cereus by colony form and colour. Confirmation of identity should be carried out.
Quality assessment
(i) Productivity Test strain Bacillus cereus 50014
Inoculation method Modified Miles-Misra or ecometry.

404
(ii) Selectivity T e s t s t ra ins Escherichia coli 50034
Micrococcus luteus 50063
I n o c u l a t i o n M o d i f i e d M i l e s - M i s r a or e c o m e t r y .
(iii) Characteristic appearance of colonies see above .
References
Holbrook, R. and Anderson, J.M. (1980) An improved selective and diagnostic medium for the isolation and enumeration of Bacillus cereus in foods. Can. J. Microbiol. 26, 753-759.
Billing, E. and Luckhurst, E.R. (1957) A simplified method for the preparation of egg yolk media. J. Appl. Bacteriol. 20, 90.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
405
Preston campylobacter selective agar
Description and history
This medium was formulated by Bolton and Robertson (1982) to facilitate the isolation of thermophilic Campylobacter spp. from all types of specimens (human, animal, and environmental (Bolton et al., 1983)). The selective inhibitory compo- nents are polymyxin, rifampicin, trimethoprim, and cycloheximide (actidione). The majority of unwanted contaminating organisms are suppressed whilst most ther- mophilic Campylobacter spp. produce typical moist, grey, flat spreading colonies.
Composition (grams)
Beef extract 10.0 Peptone 10.0 Sodium chloride 5.0 Polymyxin B (i.u.) 5000.0 Rifampicin 0.01 Trimethoprim lactate 0.01 C~cloheximide (Actidione) 0.1 Agar 15.0 Distilled or deionized water 1000.0 Lysed horse blood (ml) 50.0
Preparation
Suspend the beef extract, peptone, sodium chloride and agar in 1 litre of water and bring to the boil to dissolve completely. Sterilize by autoclaving at 121~ for 15 min. Cool to 50~ Add aseptically 50 ml of lysed horse blood and the selective ingredients to give final concentrations stated.
Physical properties
Appearance Red, translucent. pH 7.4 _+ 0.2

406
Shelf life
Ready to use medium 10 days at 4 + 2~
Inoculation method for samples
Plates should not be overdried but preferably left overnight at room temperature. This medium can be inoculated using 0.5 ml per 9 cm diameter plate for whole surface spreading methods or by conventional plating to produce discrete colonies. As with other media containing rifampicin caution should be taken when using this medium where stressed organisms may be encountered.
Incubation method
At 42~ for 48 h, microaerobically in a jar with an atmosphere containing approximately 5% 0 2, 10% CO 2 and 85% N 2 or H 2.
Reading of results and interpretation
Thermophilic Campylobacter spp. tend to produce moist, grey, flat spreading growth, which tends to coalesce. A pink or green hue may be exhibited by some strains.
Quality assessment
(i) Productivity Test strains Campylobacter jejuni 50091
Campylobacter coli 50092
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Escherichia coli 50034
Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 19938)
Inoculation method Modified Miles-Misra or streaking/ecometry.
(iii) Characteristic appearance of colonies
Occasionally some contaminating organisms may grow on this medium but they are usually restricted to the area of the primary inoculum. These include" Pseudomonas spp., more resistant coliforms, Streptococcus spp. and yeasts.

407
References
Bolton, F.J. and Robertson, L. (1982) A selective medium for isolating Campylobacter jejuni/coli. J. Clin. Pathol. 35, 462-467.
Bolton, F.J., Coates, D., Hinchliffe P.M. and Robertson, L. (1983) Comparison of selective media for isolation of Campylobacter jejuni/coli J. Clin. Pathol, 36, 78-83.

408 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Preston enrichment broth
Description and history
This medium was formulated to improve the recovery of thermophilic campylobac- ters when present in small numbers in specimens containing large numbers of contaminants (Bolton and Robertson, 1982). It has been used successfully for the isolation of these organisms from human, animal, avian, food, milk and water samples (Bolton et al., 1982, 1983; Korhonen and Martikainen, 1990).
Composition (grams)
Beef extract Peptone Sodium chloride Iron (II) sulphate * Sodium metabisulphite * Sodium pyruvate * Polymyxin B (i.u.) Rifampicin Trimethoprim lactate Cycloheximide (Actidione) Distilled or deionized water Lysed horse blood (ml)
10.0 10.0 5.0 0.25 0.25 0.25
5000.0 0.01 0.01 0.1
1000.0 50.0
* FBP supplement (George et al., 1978)
Preparation
Dissolve the beef extract, peptone and sodium chloride in the water and sterilize by autoclaving at 121~ for 15 min. Cool to 50~ Add aseptically 50 ml of lysed horse blood and the selective agents. The FBP supplement is added to the cooled medium to give the final concentrations stated from sterile stock solutions of the individual ingredients.
Physical properties
Appearance Dark red, translucent. pH 7.4 _+ 0.2

409
Shelf life
Ready to use med ium 7 days at 4 + 2~
Inoculation method for samples
Food samples are usually added to the broth in a ratio of 1 in 4 i.e. 10 g of food plus 30 ml of broth, etc. As with o ther media containing rifampicin caution should be exercised when using this med ium where stressed organisms may be encoun- tered.
Incubation method
Broth dispensed in screw capped containers with the min imum of air space can be incubated aerobically at 42~ for 24-48 h. Alternat ively the broth can be incubated microaerobical ly in an a tmosphere containing approximately 5% 0 2, 10% CO 2 and 85% N 2 or H 2 which may improve the sensitivity of the m e d i u m with some types of samples.
Broths are subcul tured to plates of Pres ton agar (q.v.) (Bolton and Rober tson, 1982) or modif ied CCD agar (Hutchinson and Bolton, 1984).
Quality assessment
(i) Productiuity Test strains
Inoculat ion me thod
(ii) Selectivity Test strains
Inoculat ion me thod
Campylobacter jejuni 50091 Campylobacter coli 50092
Dilut ion to extinction.
Escherichia coli 50034 Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C 19938)
Dilut ion to extinction.
References
Bolton, F.J., Coates, D., Hinchliffe, P.M. and Robertson, L. (1982) A most probable number method for estimating small numbers of campylobacter in water. J. Hyg. 89, 185-190.
Bolton, F.J., Coates, D., Hinchliffe P.M. and Robertson, L. (1983) Comparison of selective media for isolation of Campylobacter jejuni/coli. J. Clin. Pathol. 36, 78-83.
George, H.A., Hoffman, P.S., Krieg N.R. and Smimbert, R.M. (1978) Improved media for growth and aerotolerance of Campylobacter fetus. J. Clin. Microbiol. 8, 36-41.
Bolton, F.J. and Robertson, L. (1982) A selective medium for isolating Campylobacter jejuni/coli. J. Clin. Pathol. 35, 462-467.
Hutchinson, D.N. and Bolton, F.J. (1984) An improved blood-free selective medium for the isolation of Campylobacter jejuni from faeces. J. Clin. Pathol. 37, 956-957.
Korhonen, L.K. and Martikainen, P.J. (1990) Comparison of some enrichment broths and growth media for the isolation of thermophilic campylobacters from surface water samples. J. Appl. Bacteriol. 68, 593-599.

410 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Rambach agar (propylene glycol deoxycholate neutral red agar)
Description and history
This agar medium for the differentiation of Salmonella spp. from other members of the family Enterobacteriaceae was described by Rambach (1990). It exploits a novel phenotypic characteristic of Salmonella spp.: the formation of acid from propylene glycol. This characteristic is used in combination with a chromogenic indicator of/3-galactosidase to differentiate Salmonella spp. from Proteus spp. and from other members of the Enterobacteriaceae. Deoxycholate is included in the medium as an inhibitor of Gram-positive organisms. Salmonellae other than Salmonella typhi, yield distinct, bright red colonies on the medium, allowing easy identification and unambiguous differentiation from Proteus spp.
Composition (grams)
Propylene glycol Peptone Yeast extract Sodium deoxycholate Neutral red 5-bromo-4-chloro-3-indolyl/3-D-galactopyranoside Agar Distilled or deionized water
10.0 5.0 2.0 1.0 0.03 0.1
15.0 1000.0
Preparation
Suspend the ingredients in the water. Mix and heat very carefully to boiling, with frequent agitation, to dissolve soluble material. Cool to 45-50~ Adjust pH to 7.4. Pour 15-20 ml quantities into 9 cm diameter Petri dishes.
Physical properties
Appearance Pink, semi-opaque. pH 7.4 _+ 0.2

411
Shelf life
Prepared plates 14 days at 4 + 2~
Inoculation method for samples
Streak pre-dried plates from selective enrichment broths to obtain well-isolated colonies.
Incubation method
At 37 _+ 1~ for 18-24 h in air.
Reading of results and interpretation
Typical colonies of salmonellae (other than Salmonella typhi) i.e. those that produce acid from propylene glycol, are bright red in colour. Colonies positive for /3-galactosidase e.g. Escherichia coli are blue in colour; colonies producing both acid from propylene glycol and/3-galactosidase e.g. Citrobacter freundii, are violet. Colonies producing neither acid from propylene glycol nor /3-galactosidase e.g. Salmonella typhi and Proteus mirabilis, are colourless. Pseudomonas spp. may mimic the appearance of salmonellae from which they may be distinguished by an oxidase test.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Escherichia coli 50034
Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938)
Inoculation method Modified Miles-Misra or streaking/ecometry.
Criteria Escherichia coli is not inhibited but should produce blue colonies clearly distinguishable from Salmonella spp.. Proteus mirabilis may be partially inhibited, produce colourless or orange to light brown colonies and should not swarm.

412
(iii) Characteristic appearance of colonies See above .
Reference
Rambach, A. (1990) New plate medium for facilitated differentiation of Salmonella spp. from Proteus spp. and other enteric bacteria. Appl. Environ. Microbiol. 56, 301-303.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
413
Rapid Perfringens Medium (RPM)
Description and h&tory
This medium was developed by Erickson and Deibel (1978) for the detection and enumeration of Clostridium perfringens in foods. RPM is a liquid medium with a litmus milk base and is prepared in tubes. Selectivity is provided by the antibiotics polymyxin B sulphate and neomycin sulphate, coupled with an incubation tempera- ture of 46~ Detection of Clostridium perfringens is based on production of a stormy fermentation within 24 h. Gelatin hydrolysis, a characteristic common to strains of Clostridium perfringens, can also be detected in RPM (De Boer and Boot, 1983). A 3-tube most probable number (MPN) procedure can be used to enumerate Clostridium perfringens in food samples and other products (Smith and Mood, 1983; Smith, 1985).
Composition (grams)
Solution A Litmus milk powder (Difco) Neomycin sulphate Polymyxin B sulphate Distilled or deionised water
Solution B Thioglycollate medium, fluid (Difco) Gelatin Peptone Glucose Di-potassium hydrogen or thophosphate Yeast extract Sodium chloride Iron (II) sulphate Distilled or deionized water
140.0 0.150 0.025
1000.0
60.0 120.0
10.0 10.0 10.0
6.0 3.0 1.0
1000.0
Preparation
Prepare solution A by suspending the litmus milk powder in the water and bring to the boil. Adjust pH to 6.8. Sterilize by autoclaving at 121~ for 5 min. Cool to 50~

414
and aseptically add neomycin sulphate and polymyxin B sulphate. Prepare solution B by suspending the ingredients in the water and bring to the boil. Adjust pH to 7.1. Distribute 5 ml amounts in sterile screw-capped glass tubes. Sterilize by autoclaving at 121~ for 5 min. Cool to 50~
Prepare the final medium by aseptically adding 5 ml of solution A to each tube of solution B. Cap the tubes tightly and invert several times to facilitate mixing. Store at 4~ Before use liquify the medium by placing the tubes in a waterbath at 45-50~ for 30 min.
Physical properties
Appearance Light brown, opaque. pH 7.0 + 0.2
Shelf life
Ready to use medium 4 weeks at 4 + 2~
Inoculation method for samples
Inoculate RPM tubes with 1 ml of macerates of foods and mix well.
Incubation method
At 46~ for 18-20 h in air.
Reading of results and interpretation
Detection of Clostridium perfringens is based on production of a stormy fermenta- tion reaction. As stormy fermentation is a presumptive test, the identity of isolates should be confirmed by additional tests.
Quality assessment
(i) Productivity Test strains Clostridium perfringens (haemolytic) 50027
Clostridium perfringens (non haemolytic) 50028
Inoculation method Inoculate medium with 100-1000 cells of the test strains.
Criteria Stormy fermentation within 24 h at 46~

415
(ii) Selectivity Tes t s t ra ins Clostridium bifermentans 50026
Proteus mirabilis ( A T C C 2 9 9 0 6 / C E C T 4 1 6 8 / N C T C
11938)
I n o c u l a t i o n m e t h o d D i lu t i on to ext inct ion.
Cr i t e r i a D i f f e r ence in g rowth shou ld be equa l to or less t h a n
5 t i t re uni ts of the g rowth in t r y p t o n e soya b ro th . No
s to rmy f e r m e n t a t i o n wi th in 24 h at 46~
References
De Boer, E. and Boot, E. (1983) Comparison of methods for the isolation and confirmation of Clostridium perfringens from spices and herbs. J. Food Protect. 46, 533-536.
Erickson, J.E. and Deibel, R.H. (1978) New medium for rapid screening and enumeration of Clostrid- ium perfringens in foods. Appl. Environ. Microbiol. 36, 567-571.
Smith, M. and Mood, T.J. (1983) Direct testing of gelatin hydrolysis in Rapid Perfringens Medium. J. Assoc. Off. Anal. Chem. 66, 1045-1046.
Smith, M. (1985) Comparison of Rapid Perfringens Medium and Lactose Sulfite Medium for detection of Clostridium perfringens. J. Assoc. Off. Anal. Chem. 68, 807-808.

416 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Rappaport-Vassiliadis (RV) broth
Description and history
The efficiency of Rappaport-Vassiliadis enrichment medium for salmonellae is based on the following: (a) the ability of Salmonella spp. to multiply at relatively high osmotic pressures (concentration of hexahydrate magnesium chloride of 28.6 g/1 in the final medium), at relatively low pH values, at a high temperature and with modest nutritional requirements, and (b) the suppression of the toxic effect of malachite green towards salmonellae by the presence of magnesium chloride.
The medium was first proposed by Rappaport et al. (1956) for the enrichment of selected Salmonella serotypes and was later modified by Vassiliadis et al. (1976; 1983), reducing the concentration of malachite green to one third. Owing to the unusual method of preparation there has been some confusion about the concen- tration of magnesium chloride - 35 g per litre of medium final concentration is too high (Peterz et al. 1984; see Busse, this volume). The ratio of inoculum to broth is normally 1 : 100 and incubation temperature 41-42~ because some strains particu- larly Samonella dublin do not grow at 43~ (Peterz et al. 1984).
Composition (grams)
Soya peptone Sodium chloride Potassium di-hydrogen or thophosphate Magnesium chloride- 6H 20 Malachite green oxalate Distilled or deionized water
4.5 7.2 1.44
28.6 0.036
1000.0
Preparation
Three solutions are required:
Solution A - To 1000 ml of distilled water are added: soya peptone (Difco) 5 g; sodium chloride (AR) 8 g; potassium di-hydrogen orthophosphate (AR) 1.6 g. This solution is made on the day of the preparation of the RV medium. It is heated to about 80~ until the ingredients are completely dissolved.
Solution B - Contains 400 g of MgC12 �9 6H20 (AR) and 1000 ml of distilled water (31.8% w/v). As this salt is very hygroscopic it is advisable to

417
dissolve the entire contents of a newly opened container in distilled water. Solution B can be stored unsterilized in dark bottles at room temperature for at least 2 years.
Solution C - Contains 0.4 g of analytically pure malachite green oxalate (Merck No. 1398) and 100 ml of distilled water. Solution C can be kept for at least 8 months at room temperature in a dark bottle.
The final medium is prepared by adding to 1000 ml of solution A, 100 ml of solution B and 10 ml of solution C (final volume 1110 ml). It is then distributed in test tubes in 10 ml quantities or in three screw-capped bottles of 500 ml capacity each, and sterilized at 115~ for 15 min.
Physical properties
Appearance pH
Shelf life
Ready to use medium
Green-blue, transparent. 5.2 + 0.2
6 months in screw-capped bottles held at 4 + 2~ transferring 10 ml to test tubes when needed (Vassiliadis et al., 1985).
Inoculation method for samples
A pre-enrichment of the sample should precede the enrichment in RV medium. For this purpose 25 g of the sample (food, water or other environmental sample) is added to 225 ml of a nutrient broth without sugars or, preferably, to buffered peptone water and incubated at 37~ for 16 to 24 h in air. From the pre-enrich- ment medium 0.1 ml is inoculated into 10 ml of Rappaport-Vassiliadis medium.
Incubation method
At 42~ for 24 and 48 h in air.
Reading of results and interpretation
Appearance of turbidity; after 24 h and 48 h incubation of the RV medium, subcultures are made on selective plating media which are incubated for 24 h at 37~
Quality assessment
(i) Productivity Test strains
Inoculation method
Criteria
Salmonella virchow 50077 Salmonella enteritidis 50073 Salmonella typhimurium (NCTC 12190)
Dilution to extinction.
Recovery in RV broth should be within 2 titre units of recovery in tryptone soya broth after 48 h at 42~

418
(ii) Selectivity Test strains Enterobacter cloacae (ATCC 23355)
Pseudomonas aeruginosa 50067
Inoculation method Dilution to extinction.
Criteria Recovery in RV broth should be less than 5 titre units of the recovery in tryptone soya broth after 48 h at 42~
References
Peterz, M., Wiberg, C. and Norberg, P. (1989) The effect of incubation temperature and magnesium chloride concentration on growth of salmonella in home-made and in commercially available dehydrated Rappaport-Vassiliadis broths. J. Appl. Bacteriol. 66, 523-528.
Rappaport, F., Konforti, N. and Navon, B. (1956) A new enrichment medium for certain salmonellae, J. Clin. Pathol. 9, 261-266.
Vassiliadis, P. (1983) The Rappaport-Vassiliadis (RV) enrichment medium for the isolation of salmonellas: An overview. J. Appl. Bacteriol. 54, 69-76.
Vassiliadis, P., Mavrommati, Ch., Efstratiou M. and Chronas, G. (1985) A note on the stability of Rappaport-Vassiliadis enrichment medium. J. Appl. Bacteriol., 59, 143-145.
Vassiliadis, P., Paternaki, E., Papaiconomou, N., Papadakis, J.A. and Trichopoulos, D. (1976) Nouveau proc~d~ d'enrichessement de salmonella. Ann. Microbiol. Inst. Pasteur, 127 B, 195-200.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
419
Rappaport (SR) medium-semisolid modification
Description and history
Goossens et al. (1984) first described this semisolid selective-motility medium for the isolation of Salmonella spp. from faeces. Perales and Audicana (1989) and Perales and Erkiaga (1991) reported on its use in the examination of foodstuffs. It relies on the migration of salmonellae across the surface of a highly selective medium. It is based on Rappaport broth (q.v.) diluted to 60% and made semisolid by the addition of agar.
Composi t ion (grams)
Tryp tone Sodium chloride Potass ium di-hydrogen o r thophospha te Magnes ium ch lor ide . 6 H 2 0 Malachi te green oxalate Agar (Difco 0140) Disti l led or deionized water
3.25 4.34 0.87
17.25 0.065 3.2
1000.0
Preparation
Four solutions are required:
Solut ion A - To 1000 ml of distilled water are added: tryptone 6 g; sodium chloride (AR) 8 g; potassium di-hydrogen orthophosphate (AR) 1.6 g. This solution is made on the day of preparation of the SR medium. It is heated to about 80~ until the ingredients are com- pletely dissolved.
Solution B - Contains 400 g of MgCI2" 6H20 (AR) in 1000 ml of distilled water or 31.8%, w/v. As this salt is very hygroscopic it is advisable to dissolve the entire contents of a newly opened container in distilled water. Solution B can be stored unsterilized in dark bottles at room temperature for at least 2 years.
Solution C - Contains 2 g of analytically pure malachite green oxalate (Merck No. 1398) in 100 ml of distilled water. Solution C can be kept for at least 8 months at room temperature in a dark bottle.

420
Solution D - Contains 3.2 g of agar in 400 ml of distilled water and it is boiled until completely dissolved. Sterilize at 121~ for 15 min.
The Rappaport broth is prepared by adding to 1000 ml of solution A, 100 ml of solution B and 6 ml of solution C. It is then sterilized at 115~ for 15 rain. After cooling to 50~ 600 ml of the Rappaport broth are aseptically mixed with 400 ml of solution D at the same temperature and poured into 9 cm diameter Petri dishes. Plates should be dried before inoculation.
Physical properties
Appearance Blue, transparent. pH 5.2 +_ 0.2
Shelf life
Ready to use medium 5 days at room temperature.
Inoculation method for samples
Three drops (approx. 0.1 ml) of incubated pre-enrichment broth are inoculated in separate spots on the surface of one plate of SR medium and air dried. Enrich- ment cultures are treated similarly.
Incubation method
At 35-37~ for 18-24 h in air.
Reading of results and interpretation
After incubation the plates are examined for motile bacteria. A loopful of culture from the edge of migration is checked for purity and biochemical and serological identifications are made directly with culture from the edge of migration.
Quality assessment
(i) Productivity Test strains Salmonella uirchow 50077
Salmonella enteritidis 50073 Salmonella typhimurium (NCTC 12190)
Inoculation method Dilution to extinction with inoculation of each dilu- tion step.

421
(ii) Selectivity T e s t s t ra ins Enterobacter cloacae ( A T C C 23355)
Pseudomonas aeruginosa 50067
I n o c u l a t i o n m e t h o d D i l u t i o n to ex t inc t i on wi th i n o c u l a t i o n of e a c h d i lu -
t ion s tep .
References
Goossens, H., Wauters, G., DeBoeck, M., Janssens, M. and Butzler, J.P. (1984) Semisolid selective- motility enrichment medium for isolation of salmonellae from faecal specimens. J. Clin. Microbiol. 19, 940-941.
Perales, I. and Audicana, A. (1989) Evaluation of semisolid Rappaport medium for detection of salmonellae in meat products. J. Food Protect. 52, 316-319.
Perales, I. and Erkiaga, E. (1991) Comparison of semisolid Rappaport and modified semi-solid Rappaport-Vassiliadis media for the isolation of Salmonella spp. from foods and feeds. Int. J. Food. Microbiol. 14, 51-58.

422 Culture Media for Food Microbiology, J.E.L. Corrv et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Rappaport-Vassiliadis (MSRV) Medium-Semisolid modification
Description and history
The medium is a semisolid modification of Rappaport-Vassiliadis enrichment broth and is used as a motility enrichment in Petri dishes (De Smedt et al., 1986). It is a rapid and sensitive medium for the isolation of Salmonella spp. from food products and can be seeded directly after pre-enrichment or after 8 h enrichment in selective broth (De Smedt and Bolderdijk, 1987). Its efficiency is due to the ability of salmonellae to move through the highly selective medium.
Composition (grams)
Tryptose Casein hydrolysate (acid) Sodium chloride Potassium di-hydrogen or thophosphate Magnesium chloride- 6H 20 Malachite green oxalate Novobiocin Agar, technical (Oxoid L13) Distilled or deionized water
4.59 4.59 7.34 1.47
23.34 0.037 0.02 2.7
1000.0
Preparation
Five solutions are required"
Solution A - To 600 ml of distilled water are added: tryptose 5 g; casein hydro- lysate (acid) 5 g; sodium chloride (AR) 8 g; potassium di-hydrogen orthophosphate (AR) 1.6 g. This solution is made on the day of the preparation of the MSRV medium. It is heated to about 80~ until the ingredients are completely dissolved.

423
Solution B -
Solution C -
Solution D -
Solution E -
Contains 400 g of MgC12 �9 6H20 (AR) and 1000 ml of distilled water (31.8% w/v). As this salt is very hygroscopic it is advisable to dissolve the entire contents of a newly opened container in distilled water. Solution B can be stored unsterilized in dark bottles at room temperature for at least 2 years. Contains 0.4 g of analytically pure malachite green oxalate (Merck No. 1398) and 100 ml of distilled water. Solution C can be kept for at least 8 months at room temperature in a dark bottle. Contains 3 g of agar in 400 ml of distilled water. It is boiled until completely dissolved, then sterilized at 121~ for 15 min. Dissolve 100 mg of novobiocin in 5 ml of distilled water. Sterilize by filtration.
The medium is prepared by adding to 600 ml of solution A, 80 ml of solution B and 10 ml of solution C. It is then sterilized at 115~ for 15 rain. After cooling to 50~ 400 ml of solution D at the same temperature and 1.1 ml of solution E are aseptically added. After mixing, the medium is poured into Petri dishes. As the gel of different lots of agar can vary, each new lot must be tested and if necessary the agar concentration adjusted. A gel stability between 5 and 7 g / c m 2 is optimal (De Smedt et al., 1987).
Physical properties
Appearance Green-blue, transparent. pH 5.2 + 0.2
Shel f life
Ready to use medium 5 days at room temperature.
Inoculation me thod for samples
Three drops (approx. 0.1 ml) of incubated pre-enrichment broth are inoculated in separate spots on the surface of one plate of MSRV medium and air dried. Enrichment cultures are treated similarly.
Incubation me thod
At 42~ for 16-24 h in air.
Reading o f results and interpretation
After incubation the plates are examined for motile bacteria. A loopful of culture from the edge of migration is checked for purity and biochemical and serological identifications are made directly with culture from the edge of migration.

424
Quality assessment
(i) Productivity Test strains Salmonella virchow 50077
Salmonella enteritidis 50073
Supplementary strain Salmonella saintpaul 50075
Inoculation method Dilution to extinction with inoculation of each dilution step.
Criteria Recovery on M S R V should be within 4 titre units of the recovery in tryptone soya broth after 24 h at 42~
(ii) Selectivity Test strains Proteus mirabilis (ATCC 2 9 9 0 6 / C E C T 4 1 6 8 /
NCTC 11938) Pseudomonas aeruginosa 50067
Supplementary strain Citrobacter freundii 50025
Inoculation method Dilution to extinction with inoculation of each dilution step.
Criteria Recovery on M S R V should be less than 6 titre units of the recovery on tryptone soya broth after 24 h at 42~
References
De Smedt, J.M., Bolderdijk, R.F., Rappold, H. and Lautenschlaeger, D. (1986) Rapid Salmonella detection in foods by motility enrichment on a modified semi-solid Rappaport-Vassiliadis medium. J.Food Protect. 49, 510-514.
De Smedt, J.M. and Bolderdijk, R.F. (1987) Dynamics of Salmonella isolation with modified semi-solid Rappaport-Vassiliadis medium. J. Food Protect. 50, 658-661.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
425
Rogosa Agar
Description and history
Rogosa Agar (Rogosa et al., 1951) is a selective medium for the cultivation of lactobacilli (genus Lactobacillus) from various sources. High acetate concentration and low pH effectively suppress other bacteria while allowing lactobacilli to flourish.
Composition (grams)
Peptone (Tryptic digest of casein) Yeast extract Glucose Sorbitan monooleate (Tween 80) Potassium di-hydrogen orthophosphate Ammonium citrate Sodium acetate Magnesium sulphate �9 7 H 2 0 Manganese (II) sulphate. 4 H 2 0 Iron (II) sulphate. 7 H 2 0 Agar Acetic acid glacial (ml) Distilled or deionized water
10.0 5.0
20.0 1.0 6.0 2.0
15.0 0.575 0.14 0.034
15.0 1.32
1000.0
Preparation
Suspend all the ingredients except the acetic acid in the water and heat to boiling to dissolve completely. Add the glacial acetic acid and mix thoroughly. Adjust pH to 5.5 at about 50~ Heat to 95~ for 3 min. Do not autoclave. Cool to 50~ and distribute into 9 cm diameter Petri dishes.
Physical properties
Appearance pH
Light amber, clear to slightly opalescent. 5.5 _+ 0.1 at 50~

426
Shelf life
Ready to use medium 5-7 days at 4 _+ 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
This depends on the particular habitat of the organisms to be cultivated. Dairy strains should be incubated at 30~ for 2 days followed by 1 day at 22~ meat strains at 25~ for 3 days, intestinal or yoghurt strains for 2 days at either 37 or 42~ All incubations should be performed under anaerobic or microaerobic (5% O2: 10% CO 2 in N 2) conditions.
Reading of results and interpretation
The results should be read after a defined incubation time (see above). Lactobacilli show good growth with the exception of some strains (Lactobacillus sake, Lacto- bacillus curvatus=atypical streptobacteria), which produce smaller colonies. Carnobacterium spp. do not grow.
Quality assessment
(i) Productivity Test strains Lactobacillus gassed 50040
Lactobacillus sake 50056
Supplementary strains Lactobacillus brevis 50043 Lactobacillus plantarum 50054
Inoculation method Modified Miles-Misra or streaking/ecometry.
Criteria Recovery on Rogosa agar should be within 1.0 lOg l0 of recovery on MRS agar.
(ii) Selectivity Test strains Lactococcus lactis ssp. lactis 50058
Enterococcus hirae 50031
Supplementary strain Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or streaking/ecometry.

427
Cri ter ia Recovery on Rogosa agar should be 5.0 log10 below the recovery on M R S agar.
(iii) Characteristic appearance of colonies
Small greyish-white colonies, flat or raised, smooth or rough or in te rmedia te .
Reference
Rogosa, M., Mitchell J.A., and Wiseman, R.F. (1951) A selective medium for the isolation of oral and faecal lactobacilli. J. Bacteriol. 62, 132-133.

428 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Rogosa agar modified (pH 6.2)
Description and history
Rogosa agar in the original version is primarily a selective medium for the cultivation of lactobacilli (genus Lactobacillus). High acetate concentration and low pH effectively suppress other bacteria but also many strains of other lactic acid bacteria. The modification of the pH to 6.2 instead of 5.5 alters the selectivity of the medium for the whole group of lactic acid bacteria (ISO, 1984; Reuter, 1985).
Composition (grams)
Peptone (tryptic digest of casein) Yeast extract Glucose Sorbitan monooleate (Tween 80) Potassium di-hydrogen orthophosphate Ammonium citrate Sodium acetate Magnesium sulphate . 7 H 2 0 Manganese (II) sulphate �9 4 H 2 0 Iron (II) sulphate �9 7 H 2 0 Agar Acetic acid glacial (ml) Distilled or deionized water
10.0 5.0
20.0 1.0 6.0 2.0
15.0 0.575 0.14 0.034
15.0 1.32
1000.0
Preparation
To rehydrate, suspend all the ingredients except the acetic acid in cold water and heat to boiling to dissolve completely. Add the glacial acetic acid and mix thoroughly. Adjust pH to 6.2 at about 50~ Heat to 95~ for 3 min. Do not autoclave. Cool to 45~ and distribute into 9 cm diameter Petri dishes.
Physical properties
Appearance pH
Light amber, clear to slightly opalescent. 6.2 + 0.1 at 50~

429
Shelf life
Ready to use medium 5-7 days at 4 + 2~
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
This depends on the particular habitat of the organisms to be cultivated. Dairy strains should be incubated at 30~ for 2 days followed by 1 day at 22~ meat strains at 25~ for 3 days, intestinal or yoghurt strains for 2 days at either 37 or 42~ All incubations should be performed under anaerobic or microaerobic (5% O2: 10% CO 2 in N 2) conditions.
Reading of results and interpretation
Reading the results should be performed after a defined incubation time (see above). All well grown colonies may be considered as lactic acid bacteria. Entero- cocci and some pediococci show a reduced growth rate, some psychrotrophic leuconostocs from meat show slime production at 25~ Some unwanted strains e.g. Bacillus spp., micrococci and yeasts may occur. Carnobacterium piscicola does not grow. Confirmation of identity must be done by Gram stain and catalase test.
Quality assessment
(i) Productivity Test strains Lactobacillus gasseri 50040
Lactobacillus sake 50056
Supplementary strains Leuconostoc mesenteroides 50060 Lactobacillus plantarum 50054
Inoculation method Modified Miles-Misra or streaking/ecometry.
Criteria Recovery on Rogosa agar (pH 6.2) should be within 1.0 loga0 of recovery on MRS agar.
(ii) Selectivity Test strains Staphylococcus aureus 50080
Bacillus cereus 50014

430
S u p p l e m e n t a r y strain Escherichia coli 50034
Inocu la t ion m e t h o d Modi f i ed Miles-Misra or s t r e a k i n g / e c o m e t r y .
Cr i ter ia Recovery on Rogosa agar (pH 6.2) should be 5.0
log10 below the recovery on M R S agar.
(iii) Characteristic appearance of colonies
Small greyish-white colonies, flat or raised, smooth , rough or i n t e rmed ia t e . U n w a n t e d strains usually a t ta in a d i a m e t e r > 2.5 mm.
References
ISO/TC 34/SC 6/WG 15, No. 3 and No. 5 (1984) Draft reports: Enumeration of Lactobacteriaceae in meat and meat products.
Reuter, G. (1985) Elective and selective media for lactic acid bacteria. Int. J. Food Microbiol. 2, 55-68. Rogosa, J., Mitchell J.A. and Wiseman, R.F. (1951) A selective medium for the isolation and
enumeration of oral and fecal lactobacilli. J. Bacteriol. 62, 132-133.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
431
Rose Bengal Chloramphenicol (RBC) agar
Description and history
A selective medium for the enumeration of moulds and yeasts in foods, developed by Jarvis (1973) and containing chlortetracycline. It is now more commonly used with chloramphenicol. The inclusion of rose bengal not only restricts the size and height of mould colonies but assists enumeration in that the colour is taken up by the fungi. Chloramphenicol is used as the selective agent to suppress most bacteria. RBC agar is suitable for use with proteinaceous foods and where higher than normal incubation temperatures (35~ are required. It neither selects for specific groups of fungi, nor is diagnostic for detecting specific mycotoxin produc- ers.
Composition (grams)
Mycological peptone Glucose Di-potassium hydrogen orthophosphate Magnesium sulphate- 7H 2 O Rose Bengal Chloramphenicol Agar Distilled or deionized water
5.0 10.0
1.0 0.5 0.05 0.1
15.0 1000.0
Preparation
Suspend the basal ingredients in water and bring to the boil to dissolve completely. Add 10 ml of a 1% ethanolic solution of chloramphenicol, mix and sterilize by autoclaving at 121~ for 15 min. Cool to below 50~ and dispense approximately 15 ml amounts into 9 cm diameter Petri dishes. Use immediately or store at 4 _+ 2~ in the dark until required.
Physical properties
Appearance pH
Deep pink, without any significant precipitate. 7.2 _+ 0.2 at 25~

432
Shelf life
Ready to use medium 7 days at 4 + 2~ in the dark.
Inoculation method for samples
Surface spreading over whole plate using 0.1 or 0.2 ml per 9 cm diameter plate.
Incubation
At 25 _+ 0.5~ in the dark for 5 days. Where identification is required, prolong incubation until characteristic colonies are formed.
Reading of results and interpretation
Where separate counts of moulds and yeasts are required, identify by morphologi- cal appearance and, where necessary, microscopic examination of the two groups of microorganisms. Colonies of yeasts and bacteria can be confused and should be checked microscopically.
Quality assessment
Moulds Use stab inoculation procedure. Growth rate should be within 30% of the figure given.
Test strains CMI ATCC
Rhizopus stolonifer Aspergillus flauus Penicillium cyclopium
Growth rate (mm per day
at 25~
61269 13.2 91856ii 8.0 19759 16025 4.6
Yeasts Use stab inoculation procedure. Growth should be within 30% of figure given.
Test strain
Saccharomyces cereuisiae
Growth rate (mm per day at 25~
50105 1.8
Bacteria Use streaking procedure (Appendix I, Method B) with a loopful (10/xl) from a 24 h broth culture. No growth should be evident on sectors C, D and E of the streaked plate after incubation at 25~ for 5 days.

Test strain
433
Bacillus subtilis 50018
Reference
Jarvis, B. (1973) Comparison of an improved rose-bengal-chlortetracycline agar with other media for the selective isolation and enumeration of moulds and yeasts in food. J. Appl. Bacteriol. 36, 723-727.

434 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1993 Elsevier Science B.V. All rights reserved
Salmonella Shigella Deoxycholate Calcium (SSDC) agar
Description and history
This medium, first described by Wauters (1973), is formulated for detection of pathogenic Yersinia enterolitica, especially biotype 4 serotype 0:3, and to some extent biotype 2 serotypes 0:9 and 0:5, 27. It may be less suitable or not suitable at all for other biotypes or other Yersinia species. The selectivity relies on the basic components of SS-agar supplemented by sodium deoxycholate, yeast extract and calcium chloride. As in SS-agar the indicative principle is the absence of lactose fermentation leading to colourless colonies, lactose fermenting colonies being pink. The medium is particularly suitable for subculturing from ITC (q.v.) enrichment (Wauters et al., 1988).
Composition (grams)
Beef extract 5.0 Peptone from meat or tryptose 5.0 Yeast extract 5.0 Lactose 10.0 Ox bile dried 8.5 Sodium deoxycholate 10.0 Sodium citrate 10.0 Sodium thiosulphate 8.5 Iron (III) citrate 1.0 Brilliant green 0.0003 Neutral red 0.025 Calcium chloride 1.0 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients in the water. Bring to the boil to dissolve completely. Avoid prolonged heating. Pour into 9 cm diameter Petri dishes.

435
Physical properties
Appearance pH
Pale yellowish-brownish, clear. 7.4 + 0.1
Shelf life
Ready to use medium 7 days at room temperature in the dark. Do not store at 4~
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml on a 9 cm diameter pre-dried plate. Best results are achieved after ITC (q.v.) broth enrichment, by streaking one loopful on the surface of a whole plate.
Incubation method
At 30~176 for 24 h in air.
Reading of results and interpretation
Yersinia enterocolitica appears as round colourless colonies, about 1 mm in diame- ter. By transillumination under a binocular (10 • ) lens they are finely granular. Confirmatory tests are required.
Quality assessment
(i) Productivity Test strain Yersinia enterocolitica biotype 4, serotype 0:3 50087
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(ii) Selectivity Test strains Escherichia coli 50034
Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(iii) Characteristic appearance of colonies
Round, colourless about 1 mm in diameter.

436
References
Wauters, G. (1973) Improved methods for the isolation and the recognition of Yersinia enterocolitica. Contrib. Microbiol. Immunol. 2, 68-70.
Wauters, G., Goossens, V., Janssens, M. and Vandepitte, J. (1988) New enrichment method for isolation of pathogenic Yersinia enterocolitica serogroup 0:3 from pork. Appl. Environ. Microbiol. 54, 851-854.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
437
Selenite cystine broth
Description and history
This is a selective enrichment medium used in procedures for the detection and isolation of salmonellae. Selectivity is conferred by the inclusion of sodium selenite in the medium, shown by Leifson (1936) to be much more inhibitory to other members of the Enterobacteriaceae than to many strains of Salmonella, in particu- lar Salmonella typhi and Salmonella paratyphi. Selenite cystine broth is a modifica- tion (North and Bartram, 1953) of the original Leifson (1936) formula, differing only in the addition of L-cystine which is considered to enhance Salmonella growth by reduction of toxicity.
Selenite cystine broth is an internationally accepted medium (see ICMSF, 1978; Anon., 1981; 1985; Speck, 1984).
Composition (grams)
Tryptone Lactose Di-sodium hydrogen or thophosphate �9 12H20 Sodium hydrogen selenite * L-Cystine Distilled or deionized water
5.0 4.0
10.0 4.0 0.01
1000.0
Preparation
Base
Dissolve the tryptone, lactose and di-sodium hydrogen orthophosphate in water by boiling for 5 min. After cooling add the sodium hydrogen selenite. Adjust the pH to 7.0 + 0.1 at 20~ Store at 4~
m
L-Cystine solution
Add 0.1 g cystine to 15 ml N/1 NaOH. Dilute to 100 ml with sterile water in a sterile flask. Do not heat.
* WARNING: Extreme care should be taken with the laboratory use of selenite solutions because of their potentially toxic effect.

438
Complete medium
Cool base and add L-cystine solution at the rate of 0.1 ml per 10 ml of base. Adjust the pH to 7.0 _+ 0.1 at 20~ Transfer the complete medium in appropriate quantities e.g. 10 ml, 100 ml, etc. to sterile containers, to give a depth of 50-60 mm.
Physical properties
Appearance
pH
Pale straw coloured; clear or may have a slight precipi- tate. A brick red precipitate indicates overheating. 7.0 + 0.1 at 20~
m
Shelf life
Ready to use medium Use on day of preparation.
Inoculation method for samples
1. Where damaged cells are sought transfer 10 ml (or 1 ml) of sample-inoculated and incubated pre-enrichment broth into 100 ml (or 10 ml) Selenite cystine broth
or 2. Add comminuted (food) sample to Selenite cystine broth in the ratio 1 part
sample to 10 parts medium.
Incubation method
At 37~ for 24 h.
Reading of results and interpretation
Inoculated and incubated medium is subcultured after 24 h onto selective diagnos- tic agar media in such a way as to obtain well-isolated colonies. Suspect colonies are subcultured and their identity confirmed by biochemical and serological tests.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077 Salmonella typhimurium (NCTC 12190)

439
Inocula t ion me thod Dilut ion to extinction.
Cri ter ia Recovery in seleni te cystine b ro th should be within one t i tre unit of the recovery in t ryp tone soya bro th after 24 h at 37~
(ii) Selectivity Test strains Enterobacter cloacae (ATCC 23355)
Escherichia coli 50034
Inocula t ion m e t h o d Dilut ion to extinction.
Cri ter ia Recovery in seleni te cystine bro th should be less than 2 t i tre units of the recovery in t ryp tone soya bro th after 24 h at 37~
Comments
This m e d i u m may be markedly inf luenced by the following:
1. Source of ingredients or dehydra t ed comple te m e d i u m 2. The p resence of food or o the r organic mater ia l 3. The physiological state of the organisms sought 4. The compet ing microflora 5. Possibly by combinat ions of the above
Successful pe r fo rmance of the m e d i u m tes ted in one set of s t andard condit ions may not be achieved if any of these condit ions are al tered.
References
Anon. (1981) International Organisation for Standardisation. Microbiology - General guidance on methods for the detection of Salmonella. ISO, 6579-1981.
Anon. (1985) International Organisation for Standardisation. Milk and milk products - Detection of Salmonella. ISO 6785-2985(E).
ICMSF. (1978) Microorganisms in foods, 1. Their significance and methods of enumeration. 2nd ed. University of Toronto Press.
Leifson, E. (1936) New selenite enrichment media for the isolation of typhoid and paratyphoid (Salmonella) bacilli. Amer. J. Hyg. 24, 423-432.
North, W.R. and Bartram, M.T. (1953) The efficiency of selenite broth of different compositions in the isolation of Salmonella. Appl. Microbiol. 1, 130-124.
Speck, M.L. (1984) Compendium of methods for the microbiological examination of foods. 2nd ed. American Public Health Association.

440 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Skirrow Campylobacter selective agar
Description and history
This medium was developed for the selective isolation of Campylobacter jejuni and Campylobacter coli from human faeces (Skirrow, 1977). It replaced the more cumbersome method of selective filtration through 0.65 Izm pore size membranes. The medium relies on the extreme degree of resistance of campylobacters to trimethoprim and moderate resistance to vancomycin and polymyxin. It is neces- sary to include lysed horse blood in order to neutralise trimethoprim antagonists; failure of trimethoprim activity will result in growth of Proteus spp. which are among the few groups of Gram negative bacteria that are resistant to polymyxins.
Composition (grams)
Proteose peptone 15.0 Liver digest 2.5 Yeast extract 5.0 Sodium chloride 5.0 Vancomycin 0.01 Polymyxin B (i.u.) 2500.0 Trimethoprim 0.005 Agar 15.0 Distilled or deionized water 1000.0 Lysed horse blood 50 ml
Preparation
Prepare base or Columbia agar without antibiotics and autoclave 121~ for 15 min. Cool to 50~ add antibiotics and lysed horse blood.
Physical properties
Appearance Dark red, clear. pH 7.4 +_ 0.2

441
Shelf life
Ready to use med ium 7 days at 4 _+ 2~
Inoculation method for samples
Suspend mater ia l in saline or broth or subculture from an enr ichment broth. Inoculate section of agar and spread for discrete colonies.
Incubation method
At 42~ for 48 h microaerobical ly in a jar with an a tmosphere containing approxi-
mately 5% O 2, 10% CO 2 and 85% N 2 or H 2.
Reading of results and interpretation
Typical colonies are flat, glossy and effuse, with a tendency to form a spreading film if the agar is moist. Ma tu re colonies are low convex, often tan coloured.
Quality assessment
(i) Productivity Test strains Campylobacter jejuni 50091
Campylobacter coli 50092
Inoculat ion me thod Modif ied Miles-Misra or s t r e a k i n g / e c o m e t r y .
(ii) Selectivity Test strain Escherichia coli 50034
Inoculat ion me thod Modif ied Miles-Misra or s t r e a k i n g / e c o m e t r y .
(iii) Characteristic appearance of colonies See above.
References
Skirrow, M.B. (1977) Campylobacter enteritis: a 'new' disease. Brit. Med. J. 2, 9-11. Skirrow, M.B., Benjamin, J., Razi, M.H.H. and Waterman, S. (1982) Isolation, cultivation and identifi-
cation of Campylobacter jejuni and C. coli. In: Isolation and identification methods for food poisoning organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner, SAB Technical Series No. 17, Academic Press, London, pp. 313-328.

442 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Streptomycin Thallous Acetate Actidione (STAA) agar
Description and history
This medium, developed by Gardner (1966), is used for the quantitative enumera- tion of Brochothrix thermosphacta in meat and meat products (Gardner, 1981). The combination of antibiotics aims to exclude all other organisms.
Composition (grams)
Peptone (Oxoid L37) Yeast extract Glycerol Di-potassium hydrogen orthophosphate Magnesium sulphate . 7H 2 O Streptomycin sulphate * (as streptomycin) Cycloheximide (Actidione) * Thallium (I) acetate * Agar Distilled or deionized water
20.0 2.0
15.0 1.0 1.0 0.5 0.05 0.05
13.0 1000.0
Preparation
All the ingredients except those marked * are dissolved in the water, pH adjusted to 7.0 and sterilized in measured amounts by autoclaving at 121~ for 15 min. The selective agents dissolved in distilled water are added to the cooled (45~ molten base.
Physical properties
Appearance pH
Pale straw and slightly opaque. 7.0 _+ 0.2

443
Shelf life
Prepared plates Additive solutions Bottled basal medium
7 days at 4 + 2~ 1 month at 4 + 2~ 1 month at ambient.
These estimations are known to be useful but no definitive data exist on the maximum shelf life of the components.
Inoculation method for samples
Surface spreading over whole plate or modified Miles-Misra.
Incubation method
At 22~ for 48 h in air.
Reading of results and interpretation
Occasional yeast and pseudomonad colonies may be found. The former are easily recognised and the latter also have a different appearance and may be confirmed by flooding the plate with Kovac's oxidase reagent: 1% (w/v) tetramethyl-p-phen- ylenediamine dihydrochloride in water; blue colonies should be subtracted from the total.
Quality assessment
(i) Productivity Test strain Brochothrix thermosphacta 50019
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(ii) Selectivity Test strains Lactobacillus sake 50056
Enterococcus faecium 50032
Supplementary strains Enterococcus faecalis 50030 Microbacterium lacticum 50062
Inoculation method Modified Miles-Misra or s t reaking/ecometry.
(iii) Characteristic appearance of colonies
White or semi-transparent convex, with or without an irregular margin and may exhibit a structure similar to masses of woven threads.

444
References
Gardner, G.A. (1966) A selective medium for the enumeration of Microbacterium thermosphactum in meat and meat products. J. Appl. Bacteriol. 29, 455-460.
Gardner, G.A. (1981) Brochothrix thermosphacta (Microbacterium thermosphactum) in the spoilage of meats: a review. In: Psychrotrophic microorganisms in spoilage and pathogenicity, edited by T.A. Roberts, G. Hobbs, J.H.B. Christian and N. Skovgaard. Academic Press, London, pp. 139-173.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
445
Sulphite Cycloserine Azide (SCA) agar
Description and history
SCA agar is a selective medium for the enumeration of sulphite-reducing anaer- obes (essentially mesophilic sulphite-reducing clostridia). The medium utilises the selective inhibitory properties of D-cycloserine and an indicator system involving sulphite and iron. Most unwanted species are suppressed, while Clostridium perfringens and related species will reduce the sulphite and form black colonies due to the production of ferrous sulphide. Originally the medium was prepared with higher amounts of D-cycloserine, sulphite and iron (Hauschild and Hilsheimer, 1974). Modifications and the addition of sodium azide as a further selective component were first recommended by Eisgruber (1986). The standardized recipe was published by Eisgruber and Reuter (1991).
Composition (grams)
Tryptose 15.0 Peptone from soya meal 5.0 Meat extract 5.0 Yeast extract 5.0 Glucose 2.0 Di-sodium disulphite * 0.5 Iron (III) ammonium citrate * 0.5 D-Cycloserine (97%) * 0.3 Sodium azide * 0.05 Agar 14.0 Distilled or deionized water 900.0
Preparation
Suspend ingredients except those marked * in the water and heat to boiling to dissolve completely. Sterilize by autoclaving at 121~ for 10 min then cool to 45-50~ Add 100.0 ml of a filter-sterilized aqueous solution containing 0.5 g di-sodium disulphite, 0.5 g iron (III) ammonium citrate and 1.0 ml of a stock solution of D-cycloserine/sodium azide.

446
The stock solution is prepared by suspending 1.5 g D-cycloserine and 0.25 g sodium azide in 50.0 ml distilled water, and sterilized by filtration.
pH adjustment should be performed after addition of the solutions at 45~
Physical properties
Appearance Pale amber. pH 7.4 +_ 0.2 at 45~
Shelf life
Prepared plates Additive solution Botted basal medium
2 weeks at 4 _+ 2~ several months at 4 + 2~ 2 weeks in dark at 4-2~
Inoculation method for samples
Pour plate method, using 1 ml suspension of sample-dilution per 9 cm diameter plate and mixing with about 15 ml SCA medium about 45-50~
Incubation method
At 37~ for 48 h anaerobically.
Reading of results and interpretation
Discrete blackened colonies are considered to be presumptive sulphite reducing mesophilic clostridia. An easy confirmation test is Gram staining of at least 10 colonies from one plate to be considered for count evaluation.
Quality assessment
(i) Productivity Test strains Clostridium perfringens 50027
Clostridium sporogenes 50099
Inoculation method As above.
Criteria Recovery on SCA should be within 0.5 log10 of the recovery on a non-selective medium after 48 h at 37~ anaerobically.

447
(ii) Selectivity Test strains
Inoculation method
Bacillus subtilis 50018 Escherichia coli 50034 Staphylococcus aureus 50080 Lactobacillus delbrueckii ssp. bulgaricus 50050 As above.
Criteria The recovery of Lactobacillus delbrueckii ssp. bulgaricus on SCA should be less than 2 log 10 of the recovery on a non-selective medium after 48 h at 37~ anaerobically without blackening of colonies. Other test Strains should show no growth at all.
(iii) Characteristic appearance of colonies
Blackened colonies of 1-5 mm diameter.
References
Eisgruber, H. (1986) Priifung von Verfahren zur Kultivierung und Schnellidentifizierung von Clostri- dien aus frischem Fleisch sowie aus anderen Lebensmitteln. Vet. Med. Diss. FU Berlin.
Eisgruber, H. and Reuter, G. (1991) SCA - ein Selektivniihrmedium zum Nachweis mesophiler sulfitreduzierender Clostridien in Lebensmitteln, speziell fiir Fleisch und Fleischerzeugnisse. Arch. Lebensmittelhyg. 42, 125-129.
Hauschild, A.H.W. and Hilsheimer, R. (1974) Enumeration of food-borne Clostridium perfringens in egg-yolk free tryptose-sulphite cycloserine agar. Appl. Microbiol. 27, 521-527.

448 Culture Media for Food Microbiology, J.E.L. Corl~' et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Thallous Acetate Tetrazolium Glucose (T1TG) agar
Description and history
The medium incorporates thallous acetate as a selective inhibitory agent. Differen- tiation between Enterococcus faecalis and Enterococcus faecium depends upon the reduction of tetrazolium to the maroon formazan at pH 6.0. Selectivity of the medium can be increased by incubation at 44-45~
Composition (grams)
Proteose peptone Beef extract powder Glucose Thall ium (I) acetate 2,3,5-Triphenyltetrazolium chloride Agar Distilled or deionized water
10.0 8.0
10.0 1.0 0.1
14.0 1000.0
Preparation
The peptone and beef extract powder are dissolved by boiling in half the required volume of water, making allowance for additions (a)-(c) below. The pH value is adjusted to 6.0-6.1. The agar is dissolved separately and the two solutions are mixed and distributed into bottles (or final containers) in 92 ml amounts for autoclaving at 121~ for 15 min. After cooling to 50-52~ the following solutions are added to each 92 ml of molten basal medium:
a) 5 ml of 20% (w/v)filter-sterilized glucose b) 2 ml of 5% (w/v) thallous acetate, autoclaved at 115~ for 15 min. c) 1 ml of 1% triphenyltetrazolium chloride (filter-sterilized).
Physical properties
Appearance Pale straw. pH 6.0-6.1

449
Shelf life
Ready to use basal medium Poured plates
At least one month at 20 + 2~ m
7 days at 4 + 2~
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per 9 cm diameter pre-dried plate.
Incubation method
At 37~ for 24 h.
Reading of results and interpretation
Colonies are round and about 1 mm in diameter. Those which are characteristic of Enterococcus faecalis have a deep red centre with a narrow white periphery, whilst colonies of Enterococcus faecium are white or pink. Since certain streptococci can also grow on this medium at 37~ usually forming very small red or white colonies, confirmatory tests may be necessary.
Quality assessment
(i) Productivity Test strains Enterococcus faecalis 50030
Enterococcus faecium 50032
Inoculat ion method Modified Miles-Misra or s t r eak ing /ecomet ry .
(ii) Selectivity Test strains Lactococcus lactis ssp. lactis 50058
Escherichia coli 50034
Inoculation method Modified Miles-Misra or s t r eak ing /ecomet ry .
(iii) Characteristic appearance of colonies
Round w h i t e / p a l e pink colonies (Enterococcus faecium) or colonies with a deep red centre and narrow white per iphery (Enterococcus faecalis) each about 1 mm diameter.
Reference
Barnes, E.M. (1956) Methods for the isolation of faecal streptococci (Lancefield group D) from bacon factories. J. Appl. Bacteriol. 19, 193-203.

450 Culture Media for Food Microbiology, J.E.L. Corry et ai. (Eds.) �9 1994 Elsevier Science B.V. All rights reserved
Thiosulphate Citrate Bile-salt Sucrose (TCBS) agar
Description and history
This medium was originally developed by Kobayashi et al. (1963) for the isolation of Vibrio parahaemolyticus but is also suitable for the isolation of Vibrio cholerae and most other pathogenic Vibrio spp. from clinical specimens. The selective components of the medium are bile-salts, thiosulphate, citrate and a relatively high pH of 8.6. Differentiation is based on sucrose fermentation detected by the pH indicators and H zS production from thiosulphate detected by the production of black iron sulphide from ferric citrate. Vibrio species do not produce H zS but depending on the species may ferment sucrose. Variations in the performance of different commercial preparations of this formula were noted by Nicholls et al. (1976) and West et al. (1982) who also addressed problems related to quality control of the medium.
Composition (grams)
Yeast extract 5.0 Peptone 10.0 Sodium thiosulphate 10.0 Sodium citrate 10.0 Ox bile 8.0 Sucrose 20.0 Sodium chloride 10.0 Iron (III) citrate 1.0 Bromothymol blue 0.04 Thymol blue 0.04 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Prepare in a flask at least three times larger than the volume of medium being made. Suspend the ingredients in the water and slowly heat with constant stirring.

451
On boiling remove from the heat immediately, cool to 50~ and pour into Petri dishes. Do not autoclave.
Physical properties
Appearance pH
Shelf life
Poured plates
Inoculation method
Dark blue-green or green, clear. 8.6 _+0.2
14 days at 4 _+ 2~
Surface spreading over whole plate. If material is sufficiently dilute 0.1 ml amounts of food suspensions may be spread on 9 cm diameter plates.
Incubation method
At 30-37~ for 18-24 h in air. Longer incubation reduces the selectivity of the medium.
Reading of results and interpretation
Strains of different Vibrio spp. produce yellow or green colonies of various sizes usually with a minimum diameter of 0.5 mm. Vibrio cholerae usually has yellow (sucrose fermenting) colonies of 1-3 mm diameter and Vibrio parahaemolyticus green colonies of 2-5 mm. Such colonies are considered to be presumptive Vibrio spp.
Quality assessment
(i) Productivity Test strains
Inoculation method
(ii) Selectivity Test strains
Vibrio cholerae (non 01)(NCTC 11348) Vibrio parahaemolyticus (NCTC 11344) Vibrio fluvialis 50084
Modified Miles-Misra or streaking/ecometry.
Escherichia coli 50034 Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938) Pseudomonas aeruginosa 50067
Inoculation method Modified Miles-Misra or streaking/ecometry.
(iii) Characteristic appearance of colonies
Round yellow (Vibrio cholerae, Vibrio fluvialis) or green (Vibrio parahaemolyti- cus) colonies without any evidence of blackening due to HzS production.

452
References
Kobayashi, T., Enomoto, S., Sakazaki R. and Kuwahara, S. (1963) A new selective medium for pathogenic Vibrios TCBS agar (modified Nakanishi's agar). Jap. J. Bacteriol. 18, 387-391.
Nicholls, K.M., Lee, J.V. and Donovan, T.J. (1976) An evaluation of commercial thiosulphate citrate bile salt sucrose agar (TCBS). J. Appl. Bacteriol. 41, 265-269.
West, P.A., Russek, E., Brayton, P.R. and Colwell, R.R. (1982) Statistical evaluation of a quality control method for isolation of pathogenic Vibrio species on selected thiosulphate-citrate-bile salts-sucrose agars. J. Clin. Microbiol. 16, 1110-1116.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
453
Tryptone Bile Agar (TBA)
Description and history
This medium forms part of a direct plating method for the rapid enumeration of Escherichia coli in food materials (Anderson and Baird-Parker, 1975; Holbrook and Anderson, 1982). Initially resuscitation of inocula, spread over cellulose acetate membranes overlaid on Minerals Modified Glutamate agar, allows recov- ery of injured cells and also elimination of substances inhibitory to indole forma- tion. The membrane is then transferred to TBA. Selectivity of TBA is achieved by a combination of bile salts and incubation at 44~ to suppress the majority of unwanted organisms. Specific identification of Escherichia coli relies on the demonstration of indole production by colonies formed on the membrane.
Composition (grams)
Tryp tone 20.0 Bile Salts No. 3 (Oxoid) 1.5 Agar 15.0 Disti l led or deionized water 1000.0
Preparation
Add the ingredients to the water, soak and steam to dissolve agar. Adjust pH. Dispense in 100 ml amounts and sterilize by autoclaving at 121~ for 15 min.
Physical properties
Appearance Clear straw. pH 7.2 _+ 0.2
Shelf life
Ready to use medium 4 days at 4~ or 1 day at 20~
Inoculation method for samples
Molten cooled media is poured in 12-15 ml amounts into 9 cm diameter Petri dishes. Surface moisture is removed by drying at 50~ immediately prior to use.

454
The inoculum (0.1-1.0 ml) adsorbed into the cellulose acetate membrane (pore size 450-1200 nm) overlayed on appropriately dried Minerals Modified Glutamate Agar is incubated at 37~ for 4 h. The membrane is then transferred to the TBA; avoid trapping air bubbles, do not use a spreader.
Incubation method
At 44 _+ 0.5~ for 18-24 h in air with membrane surface uppermost. Dishes in stacks of 3 or less.
Reading of results and interpretation
Membranes are removed from the agar surface into a Petri dish lid, containing 2 ml of Vracko and Sherris indole reagent, so that the whole of the lower surface is wetted. Remove excess reagent after 5 min and develop reaction in bright sunlight or under U.V. lamp. Count pink indole positive colonies.
Quality assessment
(i) Productivity Test strain Escherichia coli 50035
Inoculation method Dilution to extinction. Decimal dilutions in 0.1% peptone water from overnight culture in nutrient broth are prepared and 0.1 ml spread over membrane surface on MMGA, then to TBA as above.
Criteria Following the resuscitation procedure this fastidious strain of Escherichia coli should give counts within 0.7 log10 of counts on Heart Infusion agar incubated at 37~ for 24 h. Colonies should be well coloured by the indole reaction.
(ii) Selectivity Test strains Klebsiella oxytoca 50038
Pseudomonas aeruginosa 50067 Staphylococcus aureus 50080
Inoculation method As above to give c. 200 colonies on membrane.
Criteria Following resuscitation procedure, inhibition of growth to give a negative indole reaction. The Kleb- siella oxytoca strain may form small colonies at 44~ It will not produce indole.

455
(iii) Characteristic appearance o f colonies
Escherichia coli - f rom 1 to 3 m m d i a m e t e r , d e p e n d i n g on densi ty , en t i r e
edge , p ink s t a ined on m e m b r a n e by indo le r eac t ion .
O t h e r E n t e r o b a c t e r i a c e a e - co lour le s s to pa le b rown, 0.5 to 2 m m d i a m e t e r ,
or c o m p l e t e l y inh ib i ted .
References
Anderson, J.M. and Baird-Parker, A.C. (1975) A rapid direct plate method for enumerating Escherichia coli biotype I in foods. J. Appl. Bacteriol. 39, 111-117.
Holbrook, R. and Anderson, J.M. (1982) The rapid enumeration of Escherichia coli in foods by using a direct plating method. Society for Applied Bacteriology Technical Series No. 17, edited by J.E.L. Corry. D. Roberts and F.A. Skinner Academic Press, London.
Vracko, R. and Sherris, J.C. (1963) Indole spot test in bacteriology. Amer. J. Clin. Pathol. 39, 429-432.

456 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Tryptone Soya Broth with 10% NaC1 & 1% Sodium Pyruvate (PTSBS)
Description and history
Tryptone soya broth with 10% sodium chloride and 1% sodium pyruvate (PTSBS) was developed by Lancette and Lanier (1972) for the recovery of low numbers of stressed Staphylococcus aureus in foods with a large population of competing bacteria. The addition of pyruvate to the medium allows for the growth and recovery of Staphylococcus aureus cells injured or stressed by sublethal heating, freezing or drying. PTSBS can be used for the qualitative, or quantitative (MPN) detection of low numbers of Staphylococcus aureus in foods (Lancette and Lanies, 1987). For more than 100 cfu/g of Staphylococcus aureus in foods, it is easier to count the bacteria by direct plating on Baird-Parker agar.
Composition (grams)
Tryptone soya broth Sodium chloride Sodium pyruvate Distilled or deionized water
30.0 95.0 10.0
1000.0
Preparation
Dissolve all the ingredients, mix and adjust the pH to 7.3 if necessary. Dispense 10 ml to 16 • 150 mm tubes and sterilize the tubes for 15 min at 121~
Physical properties
Appearance Light yellow broth. pH 7.3 + 0.2
Shelf life
Ready to use medium 30 days at 4 _+ 2~

457
Inoculation method for samples
Homogenize 25 g of food in 225 ml of Butterfield's phosphate buffer (potassium di-hydrogen phosphate 26.22 g, sodium carbonate 7.78 g/L) . Alternatively use 50 g of foods in 450 ml of buffer. For qualitative detection, add 1 ml of the blended food suspension to one tube of PTSBS broth. For quantitative MPN determina- tion, first make serial decimal dilutions of the blended food suspension from 10 -2 to 10 -6 and then add 1 ml of each dilution to 3 or 5 replicate tubes of PTSBS broth.
Incubation method
At 35~ for 48 h in air.
Reading of results and interpretation
Observe each PTSBS tube for signs of bacterial growth. To confirm the presence of Staphylococcus aureus mix the contents of the positive tubes and streak a loopful of the growth on Baird-Parker agar. Only tubes from which confirmed Staphylococcus aureus are recovered are considered positive. From the numbers of positive and negative tubes in each dilution, determine the MPN of Staphylococcus aureus in 1 g of food using MPN tables.
Quality assessment
(i) Productivity Test strain Staphylococcus aureus 50080
Inoculation method Dilution to extinction.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Escherichia coli 50034
Supplementary strain Listeria monocytogenes serovar 1/2a (ATCC 35152/ NCTC 7973)
Inoculation method Dilution to extinction.
References
Lancette, G.A. (1986) Current resuscitation methods for recovery of stressed Staphylococcus aureus cells from foods. J. Food. Prot. 49, 477-481.
Lancette, G.A. and Lanier, J. (1987) Most probable number method for isolation and enumeration of Staphylococcus aureus in foods: Collaborative study. J. Assoc. Off. Anal. Chem. 70, 35-39.

458 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Tryptose Sulphite Cycoloserine (TSC) (without egg yolk)
agar
Description and history
The medium utilizes the selective inhibitory properties of D-cycloserine and an indicator system involving sulphite and ferric iron. Most unwanted organisms are suppressed, while Clostridium perfringens and related species will reduce the sulphite and form black colonies due to the production of ferrous sulphide. Originally the medium was used with the addition of egg yolk (Harmon et al., 1971) but the egg yolk-free modification is more convenient (Hauschild and Hilsheimer, 1974a, b).
Composition (grams)
Tryptose 15.0 Soytone 5.0 Yeast extract 5.0 Sodium metabisulphite 1.0 Iron (III) ammonium citrate 1.0 D-Cycloserine 0.4 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend ingredients other than D-cycloserine in 960 ml distilled water. Bring to the boil to dissolve completely. Dispense 96 ml amounts into bottles (or final containers) and sterilize by autoclaving at 121~ for 10 min. After cooling to 50-52~ add 4 ml of 1% filter-sterilized D-cycloserine.
Physical properties
Appearance Pale straw. pH 7.6 _+ 0.2

459
Shelf life
Ready to use medium Day of preparation.
Inoculation method for samples
Pour plate procedure, using 1 ml per 9 cm diameter plate. When set, plates are overlaid with sterile medium.
Incubation method
At 37~ for 18-24 h anaerobically.
Reading of results and interpretation
Discrete black colonies of 1-5 mm diameter are considered to be presumptive Clostridium perfringens.
Quality assessment
(i) Productiuity Test strains Clostridium perfringens (haemolytic) 50027
Clostridium perfringens (non-haemolytic) 50028
Inoculation method Modified Miles-Misra or streaking method using surface inoculation of pre-poured plates with subse- quent overlay.
Criteria Recovery on TSC should be within 0.7 log10 of the recovery on Reinforced Clostridial Medium after 18-24 h at 37~ in an anaerobic jar.
(ii) Selectivity Test strains Proteus mirabilis (ATCC 29906 /CECT 4 1 6 8 / N C T C
11938) Enterococcus faecium 50032
Inoculation method As (i) above
Criteria Recovery on TSC (cf. above) should be less than 5 lOg l0 of the recovery on Reinforced Clostridial Medium after 18-24 h at 37~ in an anaerobic jar.
(iii) Characteristic appearance of colonies
Discrete black colonies of 1-5 mm diameter.

460
References
Harmon, S.M., Kautter, D.A. and Peeler, J.T. (1971) Improved medium for enumeration of Clostridium perfringens. Appl. Microbiol. 22, 688-692.
Hauschild, A.H.W. and Hilsheimer, R. (1974a) Evaluation and modification of media for enumeration of Clostridium perfringens. Appl. Microbiol. 27, 78-82.
Hauschild, A.H.W. and Hilsheimer, R. (1974b) Enumeration of food-borne Clostridium perfringens in egg yolk-free tryptose-sulphite-cycloserine agar. Appl. Microbiol. 27, 521-526.
Mead, G.C., Adams, B.W., Roberts T.A. and Smart, J.L. (1982) Isolation and enumeration of Clostridium perfringens. In: Isolation and identification methods for food poisoning organisms, edited by J.E.L. Corry, D. Roberts and F.A. Skinner. SAB Technical Series No. 17 Academic Press, London, pp. 99-110.
Mossel, D.A.A. and Pouw, H. (1973) Studies on the suitability of sulphite cycloserine agar for the enumeration of Clostridium perfringens in food and water. Zbl. Bakteriol. Hyg. I. Abt. Orig. A 223, 559-561.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
461
University of Vermont (UVM) broths I & II
Description and history
UVM (University of Vermont) medium was originally reported by Donnelly and Baigent (1986) as a modification of the Listeria Enrichment Broth of Dominguez- Rodriguez et al. (1984). This medium was more recently modified to its present form (McClain and Lee, 1988) for use in the official United States Department of Agriculture Food Safety Inspection Service (FSIS) method for meats and poultry. The difference between UVM broths I and II is simply the inclusion of a higher concentration of acriflavine in the latter (McClain and Lee, 1988). Further work has shown the effectiveness of the substitution of UVM II with modified Fraser broth (q.v.) in the modified USDA-FSIS enrichment method (McClain and Lee, 1989; Warburton et al., 1991).
Composition (grams)
Proteose peptone (Difco) Tryptone (Difco) Yeast Extract Lab Lemco Powder (Oxoid) Sodium chloride Di-sodium hydrogen orthophosphate �9 2 H 2 0 Potassium di-hydrogen orthophosphate Aesculin (Sigma) Nalidixic Acid (Sigma) Acriflavine HCI (Sigma) Distilled or deionized water
5.0 5.0 5.0 5.0
20.0 12.0
1.35 1.0 0.02 0.012 a
960.0 or 0.025 b
a UVM I b UVM II
Preparation
Base
All ingredients except aesculin, nalidixic acid, and acriflavine HC1 are combined with 960 ml of distilled water and autoclaved at 121~ for 15 min.

462
Additional solutions
Prepare a 0.25% stock solution of acriflavine HC1 (0.25 g acriflavine HC1 in 100 ml distilled water) and a 5% stock solution of aesculin (5 g aesculin in 100 ml distilled water) and sterilize both solutions by autoclaving at 121~ for 15 min. Prepare a 0.4% nalidixic acid solution (0.4 g nalidixic acid in 100 ml of 0.1 N NaOH and sterilize by filtration through a 0.22/zm filter.
Complete medium
Add 4.8 ml (0.012 g) of the acriflavine solution plus 5.2 ml of sterile distilled water (UVM I) or 10 ml of the acriflavine solution (UVM II) to the cooled base to give final concentrations of 0.012 g/1 (UVM I) or 0.025 g/1 (UVM II), respectively. Add 20 ml (1.0 g) of the sterile aesculin solution to the sterile base to give a final concentration of 1 g/1 and 10 ml of the filtered nalidixic acid solution to give a final concentration of 0.04 g/1.
Physical properties
Appearance Yellowish, translucent. pH 7.2 +_ 0.2
Shelf life
Ready to use medium 5-7 days at 4 +_ 2~
Inoculation method for samples
Primary enrichment is accomplished by mixing 25 g of food sample with 225 ml of UVM I and blending or mixing for 2 min. The mixture is then incubated at 30~ After 4 h incubation, 0.2 ml of the UVM I enrichment broth is spread on LPM agar to determine the presence of presumptive Listeria monocytogenes colonies. After 24 h incubation UVM I is again spread on LPM agar. Isolated colonies are then recovered by streaking onto the uninoculated half of the LPM plate (McClain and Lee, 1988). Also after 24 h incubation, 0.1 ml of UVM I is added to 10 ml UVM II and incubated for 24 h before the sample is again swabbed onto LPM as with UVM I.
Incubation method
30~ for 24h for each enrichment step.

463
Quality assessment
(i) Productivity Test strains Listeria monocytogenes serovar 1/2a (ATCC 35152/
NCTC 7973) Listeria monocytogenes serovar 4b (ATCC 13932/ NCTC 10527) Listeria ivanouii serovar 5 50095
Inoculation method Dilution to extinction.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938)
Inoculation method Dilution to extinction.
Comments
Selective properties of acriflavine may vary from lot to lot and manufacturer to manufacturer. Each new batch must be assayed in combination with other selective agents to be used in the medium to determine the optimum concentration for use, with regard to the efficiency of selectivity and absence of inhibition of Listeria spp. Storage of stock solutions of acriflavine for periods of more than one month is not recommended.
References
Dominguez-Rodriguez, L., Suarez-Fernandez, G., Fernandez-Garayzabal, J.F. and Rodriguez-Ferri, E. (1984) New methodology for the isolation of Listeria microorganisms from heavily contaminated environments. Appl. Environ. Microbiol. 47, 1188-1190.
Donnelly, C.W. and Baigent, G.K. (1986) Method for flow cytometric detection of Listeria monocyto- genes in milk. Appl. Environ. Microbiol. 52, 689-695.
McClain, D. and Lee, W.H. (1988) Development of USDA-FSIS method for isolation of Listeria monocytogenes from raw meat and poultry. J. Assoc. Off. Anal. Chem. 71,660-664.
McClain, D. and Lee, W.H. (1989) FSIS method for the isolation and identification of Listeria monocytogenes from processed meat and poultry products. Laboratory Communication No. 57 (revised). USDA-FSIS. Beltsville MD.
Warburton, D.W., Farber, J.M., Armstrong, A., Caldeira, R., Tiwari, N.P., Babiuck, T., Lacasse, P. and Read, S. (1991) A Canadian comparative study of modified versions of the 'FDA' and 'USDA' methods for the detection of Listeria monocytogenes. J. Food Protect. 54, 669-676.

464 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
Violet Red Bile Glucose (VRBG) agar
Description and history
This medium was designed for the enumeration of Enterobacteriaceae (Mossel et al., 1978). It relies on the use of the selective inhibitory components crystal violet and bile salts and the indicator system glucose and neutral red. Thus growth of many unwanted organisms is suppressed, while sought bacteria will dissimilate glucose and produce purple zones around the colonies. Some other related Gram negative bacteria may grow but may be suppressed by the overlay procedure. The selectivity of the medium can also be increased by incubation under anaerobic conditions and/or at elevated temperature, i.e. equal to or above 42~ (Mossel et al., 1979; 1986).
Composition (grams)
Peptone 7.0 Yeast extract 3.0 Sodium chloride 5.0 Glucose 10.0 Bile salts No. 3 1.5 Crystal violet 0.002 Neutral red 0.03 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients in the water. Bring to the boil to dissolve completely. Further heating is neither necessary nor desirable. Mix well and dispense into bottles or tubes.
Physical properties
Appearance Light purple-violet. pH 7.4 _+ 0.2

465
Shelf life
Ready to use plated medium 5 days at 4 + 2~
Inoculation method for samples
Spread drop plate or poured plate procedure with or without overlay. Medium for the pour plate method should be freshly prepared, tempered to 47~ and used within 3 h. The medium can also be used as an overlayer for spread-drop plates of a rich primary plating medium, used for allowing resuscitation of sublethally stressed populations of Enterobacteriaceae. Stab inoculation procedures may also be carried out using this medium.
Incubation method
To detect mesophilic members of the Enterobacteriaceae incubate at 30~ for 24 h. Psychrotrophic Enterobacteriaceae can be detected by incubation at 4~ for 10 days. Increased selectivity may be obtained by incubation at 42~
Reading of results and interpretation
Dark purple colonies, 1-2 mm diameter surrounded by purple haloes are included in the presumptive count. Confirmation may be obtained by the use of 'Gram- negative diagnostic tubes' (Mossel et al., 1977).
Quality assessment
(i) Productivity Test strains Escherichia coli 50034
Hafnia alvei 50037
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or steaking/ecometry.
(iii) Characteristic appearance of colonies
Round, purple, 1-2 mm diameter surrounded by purple haloes.

466
References
Mossel, D.A.A., Eelderink, I. and Sutherland, J.P. (1977) Development and use of single, "polytropic" diagnostic tubes for the approximate taxonomic grouping of bacteria, isolated from foods, water and medicinal preparations. Zbl. Bakt. Hyg., I, Orig. A, 278, 66-79.
Mossel, D.A.A., Eelderink, I., Koopmans, M. and van Rossem, F. (1978) Optimalisation of a Mac- Conkey-type medium for the enumeration of Enterobacteriaceae. Lab. Pract. 27, 1049-1050.
Mossel, D.A.A., Eelderink, I., Koopmans M. and van Rossem, F. (1979) Influence of carbon source, bile salts and incubation temperature on the recovery of Enterobacteriaceae from foods using MacConkey type agars. J. Food Protect. 42, 470-475.
Mossel, D.A.A., van der Zee, H., Hardon A.P. and van Netten, P. (1986) The enumeration of thermotropic types amongst the Enterobacteriaceae colonizing perishable foods. J. Appl. Bacteriol. 60, 289-295.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
467
Violet Red Bile (VRB) agar (syn. violet red bile lactose agar)
Description and history
This medium, designed for the enumeration of bacteria of the coli-aerogenes group is derived from MacConkey's original formula (1905). It relies on the use of the selective inhibitory components crystal violet and bile salts and the indicator system lactose and neutral red. Thus the growth of many unwanted organisms is suppressed, while tentative identification of sought bacteria can be made. Organ- isms which rapidly attack lactose produce purple colonies surrounded by purple haloes. Non-fermenters or late lactose fermenters produce pale colonies with greenish zones. Some other related Gram-negative bacteria may grow but may be suppressed by the overlay procedure. Selectivity can also be increased by incuba- tion under anaerobic conditions and at elevated temperatures, i.e. > 42~ (Mosse! and Vega, 1973; Mossel et al., 1979; 1986).
Composition (grams)
Peptone 7.0 Yeast extract 3.0 Sodium chloride 5.0 Lactose 10.0 Bile salts No. 3 1.5 Crystal violet 0.002 Neutral red 0.03 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients in distilled water. Bring to the boil to dissolve completely. Further heating is neither necessary nor desirable. Mix well and distribute into bottles or tubes.

468
Physical properties
Appearance pH
Light purple-violet, transparent. 7.4 +0.2
Shelf life
Ready to use medium 5 days at 4 _+ 2~
Inoculation method
Spread drop plate or poured plate procedure with or without overlay. Medium for the pour plate method should be freshly prepared, tempered to 47~ and used within 3 h. The medium can also be poured as an overlayer for spread drop plates of a rich primary plating medium used for allowing-resuscitation of sublethally injured populations of Enterobacteriaceae, though discriminating between lactose + ve and lactose - v e types must then be done with care.
Incubation method
To detect members of the coli-aerogenes group count typical colonies after incubation at 35 ~ or 37~ for 24 h. Where it is desired to count the full range of coliform bacteria, incubate at 30~ for 24 h and count all colonies present. Psychrotrophic coliforms can be detected by incubation at 4~ for 10 days. For Escherichia coli a temperature of 44 + 0.1~ is specifically recommended.
Reading of results and interpretation
Dark purple colonies, 1-2 mm diameter surrounded by purple haloes and pale colonies with greenish zones are included in presumptive counts. Confirmation may be obtained by the use of 'Gram-negative diagnostic tubes' (Mossel et al., 1977).
Quality assessment
(i) Productivity Test strain Escherichia coli 50034
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Staphylococcus aureus 50080
Inoculation method Modified Miles-Misra or streaking/ecometry.

469
(iii) Characteristic appearance of colonies
R o u n d , p u r p l e 1 - 2 m m d i a m e t e r , s u r r o u n d e d by p u r p l e h a l o e s ( l ac tose + ve
types) , or p a l e wi th g r e e n i s h z o n e s ( l ac tose - v e types) .
References
MacConkey, A. (1905) Lactose fermenting bacteria in faeces. J. Hyg. 5, 333-379. Mossel, D.A.A., and Vega, C.L. (1973) The direct enumeration of Escherichia coli in water using
MacConkey's agar at 44~ in plastic pouches. Health Labor. Sci. 11, 303-307. Mossel, D.A.A., Eelderink, I. and Sutherland, J.P. (1977) Development and use of single, "polytropic"
diagnostic tubes for the approximate taxonomic grouping of bacteria, isolated from foods, water and medicinal preparations. Zbl. Bakt. Hyg. I., Orig., A, 278, 66-79.
Mossel, D.A.A., Eelderlink, I., Koopmans, M. and van Rossem, F. (1979) Influence of carbon source, bile salts and incubation temperature on the recovery of Enterobacteriaecae from foods using MacConkey type agars. J. Food Protect. 42, 470-475.
Mossel, D.A.A., van der Zee, H., Hardon, A.P. and van Netten, P. (1986) The enumeration of thermotropic types amongst the Enterobacteriaceae colonizing perishable foods. J. Appl. Bacteriol. 60, 289-295.

470 Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1993 Elsevier Science B.V. All rights reserved
Xylose Lysine Deoxycholate (XLD) agar
Description and history
The medium was introduced by Taylor (1965) for the isolation of salmonellae and shigellae (Taylor and Schelhart, 1971). It relies on the use of the selective inhibitory component sodium deoxycholate and three indicator systems, i.e. xylose, lactose and sucrose combined with phenol red; lysine HC1 and once more phenol red; sodium thiosulphate and iron. Thus the growth of many unwanted organisms is suppressed, while sought bacteria can be tentatively grouped by reading the net effect of carbohydrate dissimilation and lysine decarboxylation and the formation of hydrogen sulphide from thiosulphate, leading to black colonies.
Composition (grams)
Yeast extract 3.0 Sodium chloride 5.0 Xylose 3.75 Lactose 7.5 Sucrose 7.5 L-Lysine hydrochloride 5.0 Sodium thiosulphate 6.8 Iron (III) ammonium citrate 0.8 Phenol red 0.08 Sodium deoxycholate 1.0 Agar 15.0 Distilled or deionized water 1000.0
Preparation
Suspend the ingredients in the water. Heat, with frequent agitation until the medium starts boiling. Avoid overheating. Transfer immediately to a water bath tempered at about 50~ continue to agitate until the medium has cooled to approximately 50~ and pour into plates. Preparing large volumes, requiring prolonged heating is to be avoided.

471
Physical properties
Appearance Light rose, transparent. pH 7.4 _+ 0.2
Shelf life
Ready to use medium 5 days at 4 _+ 2~ in the dark.
Inoculation method for samples
Surface spreading over whole plate using 0.1 ml per 9 cm diameter plate and 0.5 ml per 12 cm diameter plate.
Incubation method
At 37 or 43~ for 18-24 h in air, dependent on degree of selectivity to be attained.
Reading of results and interpretation
A tentative grouping of isolates can be made from colonial appearance on XLD. Roughly three groups of Enterobacteriaceae can be discriminated: (i) colonies with red zones and black centre: Salmonella, Arizona, and Edwardsiella spp.; (ii) colonies with red zones and red centre: Shigella, Providencia spp. and hydrogen sulphide negative salmonellae; (iii) colonies with yellow haloes and yellow centres: the genera Escherichia, Enterobacter, Citrobacter, Kluyvera, Klebsiella, Hafnia, Serratia and Proteus, and the species Yersinia enterocolitica. For confirmation of this presumptive evidence the usual biochemical tests must be carried out.
Quality assessment
(i) Productivity Test strains Salmonella enteritidis 50073
Salmonella virchow 50077 Shigella flexneri 50079 Shigella sonnei (ATCC 29930)
Supplementary strain Salmonella typhimurium 50100
Inoculation method Modified Miles-Misra or streaking/ecometry.
(ii) Selectivity Test strains Enterococcus faecalis 50030
Proteus mirabilis (ATCC 29906/CECT 4168 /NCTC 11938) - qualitative test for control of swarming

472
S u p p l e m e n t a r y s t ra ins Bacillus cereus 50014
Hafnia alvei 50037 Lactococcus lactis ssp. lactis 50058
Micrococcus luteus 50063
Staphylococcus aureus 50080
I n o c u l a t i o n m e t h o d Mod i f i ed Mi l e s -Mis ra or s t r e a k i n g / e c o m e t r y .
(iii) Characteristic appearance of colonies See above.
References
Taylor, W.I. (1965) Isolation of shigellae. I. Xylose lysine agars: new media for isolation of enteric pathogens. Amer. J. Clin. Pathol. 44, 471-475.
Taylor, W.I. and Schelhart, D. (1971) Isolation of shigellae. VIII. Comparison of xylose lysine deoxycholate agar, Hektoen enteric agar, Salmonella-Shigella-agar and eosin methylene blue agar with stool specimens. Appl. Microbiol. 21, 32-37.

Culture Media for Food Microbiology, J.E.L. Corrv et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
473
Appendix I
Testing methods for use in quality assurance of culture media
Introduction
Six testing methods are outlined in the following section. Familiarity with general microbiological techniques is assumed and therefore the methods are not given in exhaustive detail. Suitable test organisms are listed in the sections on individual media. The reference medium for bacteria is tryptone soya agar or broth unless another medium is specified in the monograph. The use of stationary phase cultures is suggested but the worker is at liberty to employ log phase cultures if a more exacting test is required and thought justified.
General criteria for assessment of bacteriological media are given in the Notes on the use of the monographs. Where these are inappropriate, specific criteria are given in individual monographs.
Definition of productivity ratios (PRs)
Where the method employed entails total counts these are expressed as N s / N o where N o = count on reference medium and Ns = count on test medium.
Absolute and relative growth indices (AGI and RGI)
When using the ecometric streaking method, the AGI and RGI may be calculated as described by Mossel et al. (1983). Strains growing on all five streaks have an AGI of 5, those growing only on the first three streaks have an AGI of 3, and so on. The RGI for a given strain is the ratio of the AGI on the test medium to the AGI on the reference medium. The AGI and RGI are used when testing solid media by semi-quantitative techniques (see method B).

474
Methods
Plated media
A. Modified Miles-Misra method
This is a general method suitable for most bacteriological agars.
1. Using an aseptic technique prepare decimal dilutions of a freshly raised and well mixed stationary phase culture of the test organism in saline peptone diluent.
2. Prepare plates according to the recommended formula with a minimum depth of agar of 4 mm and an adequately dried surface. Plates of the reference medium should be similarly prepared.
3. Using the same dropper for all the dilutions of one organism and commencing with the highest dilution to be tested, dispense four drops from each appropri- ate dilution onto the surface of four test and four reference plates. (Other dropping systems or a spiral plater may be used, see Corry, 1982.)
4. Spread each drop over 1 /4 of the plate using a sterile spreader for each plate and starting at the highest dilution.
5. Incubate plates under conditions specified in individual monographs. 6. Assess results visually for numbers and appearance.
B. Streaking method (Ecometry)
This may be a suitable alternative to method A when quantitative results are not required.
Experience may show the need for initial dilution of the stationary phase culture for method (ii) below. This may be performed according to ISO 6887 (1983).
1. Take a freshly raised stationary phase culture in liquid medium. 2. Prepare plates as in A2. 3. Either (i) Using a standard loop streak plates of test and reference media
according to the ecometric method of Mossel et al. (1983) or: (ii) Deposit a
Fig. 1.

475
standard loopful (5 /z l) of culture over sector A of the agar surface (Fig. 1.) Draw four parallel lines approximately 5 mm apart across the area of the spread culture in sector A and across sector B. Repeat for sectors C and D. Finally, draw a single line across sector D at right angles to the lines of inoculation and continue streaking into sector E. It is essential to flame and cool the loop in between each sector in order to obtain well isolated colonies. Repeat on reference medium.
4. Incubate plates under conditions specified in individual medium protocols. 5. Assess results visually for numbers and appearance of colonies.
(a) where the ecometric method has been used the absolute and relative growth indices may be calculated as described by Mossel et al. (1983).
(b) a similar method of assessment can be employed for the streaking method or each sector may be scored separately with a minimum score of 1 for well spaced discrete colonies and a maximum of 4 for confluent growth along the lines of inoculation. The sum of the score for each sector may then be calculated and test and reference agars compared.
C. Stab inoculation method
This technique is suitable for assessing the performance of mycological media.
1. Prepare a heavy spore suspension from a 7-10 day old culture of the test organism by mixing a loopful of growth in 0.5 ml of 0.25% agar.
2. Prepare plates according to the recommended formula with a minimum depth of agar of 4 mm and an adequately dried surface.
3. Take three test plates and stab inoculate the medium in each plate from below to a depth of 1-2 mm with the wire loaded with spore suspension. The point of inoculation should be approximately midway between the rim and the centre of the plate. Sterilize the wire and reload with suspension for each inoculation.
4. Repeat operation 3 with three more test organisms, stab inoculating the same plates on the remaining three quadrants.
5. If required, use a further three plates and inoculate in a similar manner for other test organisms.
6. Incubate plates, inverted, under conditions specified in individual monographs and measure the diameter of each colony at daily intervals.
7. Plot the average colony diameter against time for each test organism and calculate the growth rate. Alternatively, determine the mean colony diameter on one occasion after a suitable incubation period.
Liquid media
The interactions leading to the successful isolation of organisms from liquid selective media are complex (Mackey, 1985; Beckers et al., 1987), hence devising quality control methods is less straightforward than with solid media.
Often, as in the isolation of salmonellae, selective enrichment in liquid medium is only one step in a complex isolation procedure. The performance of the entire system can be monitored by testing for the recovery of low numbers of a marked

476
strain inoculated into appropriate sample material (Beckers et al., 1985). However it may also be desirable to test the performance of individual media. For this purpose the following methods using pure cultures of standard test strains are proposed.
D. Assessment of productivity and selectivity of liquid selective media by dilution to extinction. (Mossel et al., 1974; Official French dilution method, Richard, 1982).
1. Prepare stationary phase cultures of test strains. 2. Dilute 10 -1 to 10 -12 in tryptone soya broth. 3. Add 1 ml volumes to 9 ml selective enrichment medium and a non-selective
broth. (Other volumes may be found to be more suitable.) 4. Incubate at the appropriate temperature for 18-24 h. 5. Record highest dilutions showing turbidity.
E. Assessment of the performance of liquid selective media with mixed cultures of wanted and unwanted organisms
This method was devised for testing enrichment media for salmonellae. When used for this purpose the nalidixic acid resistant strain of Salmonella typhimurium (NCTC 12190) should be used as the wanted organism and plates of tryptone soya agar or other non-selective agar containing 20 ~ g / m l nalidixic acid prepared for use as described in sections 4 and 7 below.
1. Prepare stationary phase cultures of test strains (suitable unwanted strains for salmonella enrichment broths are Citrobacter freundii 50025, Escherichia coli 50034, Proteus mirabilis (ATCC 29906/CECT 4168/NCTC 11938), Pseu- domonas aeruginosa 50067).
2. Mix together 1 ml volumes of the unwanted organisms and dilute to 10 -1 in peptone saline.
3. Dilute the wanted organism 10 -1 to 10 -9 in peptone saline. 4. Estimate the viable count of the wanted organism by surface plating on tryptone
soya agar (with and without nalidixic acid). 5. To determine the ability of the enrichment broth to recover small numbers of
the. wanted organism, inoculate 1 ml from each peptone saline dilution into 9 ml enrichment broth, incubate at the appropriate temperature for 18-24 h then streak onto plates of tryptone soya agar.
6. Examine tubes and plates from 4 and 5 above and, from the viable count (steps 3 and 4), calculate the minimum number of organisms required to initiate growth in the enrichment broth. Check that growth of the nalidixic acid resistant strain is not inhibited on the nalidixic acid containing medium.
7. To determine the ability of enrichment broth to recover small numbers of the wanted organism from mixed cultures inoculate 1 ml from each peptone saline dilution (step 3) into a separate tube containing 10 ml enrichment broth and 0.02 ml of the 10-1 dilution of unwanted organisms (step 2). Incubate at the

477
appropriate temperature for 18-24 h then streak onto plates of tryptone soya agar with and without nalidixic acid and, if required, other selective agars (e.g. SS, XLD).
8. Examine plates from 7 above for typical colonies of the wanted strain. Record the highest dilution from which the wanted strain could be detected.
9. Interpretation of results. i) Productivity. Liquid selective media should support the growth of a pure
culture of the wanted organism from an inoculum of 20 or less. By plating on selective and non-selective agars any interaction between liquid and solid selective media may also be detected.
ii) Selectivity. Growth of the wanted strain in enrichment broth containing competing (unwanted) organisms should be detectable within one tenfold dilution of the highest dilution supporting growth of the pure culture.
F. Method of assessing growth rate in broth
This is intended as a general method for use with non-selective or selective (enrichment) liquid media. In the latter case it might also be necessary to add sample material comparable to that to be tested.
1. Prepare stationary phase culture, mix well and dilute in saline peptone water at room temperature to an approximate concentration of 105 cfu/ml.
2. Distribute 10 ml quantities of reference and test broths (see individual mono- graphs) into 25-30 ml screw-capped containers and sterilize. Equilibrate broths to incubation temperature.
3. Inoculate a pair of broths for each test organism with 0.1 ml of the 105 c fu /ml suspension and mix.
4. Remove 0.1 ml of the broth to a non-inhibitory agar plate and spread over half of the surface. Repeat on the other half of the plate with 0.1 ml from the reference broth.
5. Incubate broths and inoculated plates under required conditions. 6. At suitable time intervals remove samples from each broth with a standard loop
or pipette. Dilute if necessary and spread over a quarter of a plate of the same medium as used in 4.
7. Results may be assessed visually and may be plotted as counts vs. time.
References
Beckers, H.J., van Leusden, F.M., Meijssen, M.J.M. and Kampelmacher, E.H. (1985) Reference material for the evaluation of a standard method for the detection of salmonellas in food and feeding stuffs. J. Appl. Bacteriol. 59, 507-512.
Beckers, H.J., Heide, J.V.D., Fenigsen-Narucka, U. and Peters, R. (1987) Fate of salmonellas and competing flora in meat sample enrichments in buffered peptone water and in Muller-Kauffmann's tetrathionate medium. J. Appl. Bacteriol. 62, 97-104.

478
Corry, J.E.L. (1982) Quality assessment of culture media by the Miles-Misra method. In: Quality assurance and quality control of microbiological culture media, edited by J.E.L. Corry, G.I.T.-Verlag, Darmstadt, pp 21-37.
ISO 6887 (1983) Microbiology - General guidance for the preparation of dilutions for microbiological examinations, International Standards Organisation.
Mackey, B.M. (1985) Quality control monitoring of liquid selective enrichment media used for isolating salmonellae. Int. J. Food Microbiol. 2, 41-48.
Mossel, D.A.A., Harrewijn, G.A. and Nesselrooy-van Zadelhoff, C.F.M. (1974) Standardisation of the selective inhibitory effect of surface active compounds used in media for the detection of Enterobac- teriaceae in foods and water. Health Lab. Sci. 11, 260-267.
Mossel, D.A.A., Bonants-van Laarhoven, T.M.G., Ligtenberg-Merkus A.M.T. and Werdler, M.E.B. (1983) Quality assurance of selective culture media for bacteria, moulds and yeasts: an attempt at standardisation on the international level. J. Appl. Bacteriol. 54, 313-327.
Richard, N. (1982) Monitoring the quality of selective liquid media using the official French dilution technique for the bacteriological examination of food. In: Quality assurance and quality control of microbiological culture media, edited by J.E.L. Corry, G.I.T.-Verlag, Darmstadt, pp 51-57.

Culture Media for Food Microbiology, J.E.L. Corry et al. (Eds.) �9 1995 Elsevier Science B.V. All rights reserved
479
Appendix II
Test strains
Where a strain is known to be in more than one collection, all known reference numbers are given. Absence of a reference number for a particular collection does not necessarily imply absence of the strain from that collection. Organisms appear- ing in these monographs only as supplementary strains are marked with an asterisk (*).
Abbreviation key:
ATCC
BUCSAV
CBS CCM
CCUG
CECT
CIP CMI
DSM
HNCMB
IAM
IFO JCM NCDO/NCFB
American Type Culture Collection, 12301 Parklawn Drive, Rockville, Maryland, U.S.A. Biologicky Ustav, Ceskoslovenska Akademie Ved (Institute of Biology, Czechoslovak Academy of Sciences), Prague, Czech Republic. Centraalbureau voor Schimmelcultures, Baarn, Netherlands. Czechoslovak Collection of Microorganisms, Masaryk University, Tvrd6ho 14, Brno, Czech Republic. Culture Collection, University of Goteborg, Guldhedsg 10, S-413 46, Goteborg, Sweden. Coleccion Espanola de Cultivos Tipo, Departemento de Micro- biologia, Universidad de Valencia, Burjasot, Valencia, Spain. Collection de l'Institute Pasteur, Paris, France. Commonwealth Mycological Institute, Ferry Lane, Kew, Surrey, TW9 3AF, U.K. Deutsche Sammlung von Mikroorganismen, (German Collection of Microorganisms), D-38124 Braunschweig, Germany. Hungarian National Collection of Medical Bacteria, OK1, Gyali ut 2-6, H-1966 Budapest, Hungary. Institute of Applied Microbiology, University of Tokyo, Tokyo, Japan. Institute of Fermentation, Osaka, Japan. Japanese Collection of Microorganisms, Saitama, Japan. National Collection of Food Bacteria (formerly National Collec- tion of Dairy Organisms). This collection has now been merged with NCIMB.

480
NCIMB
NCTC
NCYC
SLCC
National Collections of Industrial and Marine Bacteria Ltd., 23, St. Machar Drive, Aberdeen, AB24 3RY, U.K. (includes National Collection of Industrial Bacteria). National Collection of Type Cultures, Central Public Health Lab- oratory, Colindale Avenue, London, NW9 5HT, U.K. National Collection of Yeast Cultures, Food Research Institute, Colney Lane, Norwich NR4 7UA, U.K. Prof. Dr. H.ER. Seeliger Listeria Culture Collection, Institute of Hygiene, Medical Faculty, University of Wtirzburg, Wtirzburg, Germany.
Bacteria NCIMB QC Collection
ATCC CCM CECT CIP DSM NCDO / N C F B
NCIMB NCTC Other
Acinetobacter sp. Aerococcus viridans * Bacillus cereus Bacillus licheniformis * Bacillus subtilis Brochothrix therrnosphacta Campylobacter coli Campylobacter jejuni Carnobacterium divergens Carnobacterium piscicola Carnobacterium piscicola Citrobacter freundii Clostridium bifermentans Clostridium perfringens
(haem) Clostridium perfringens
(non-haem) Clostridium sporogenes Enterobacter cloacae Enterococcus faecalis Enterococcus hirae Enterococcus faecium Escherichia coli Escherichia coli Hafnia alvei Klebsiella oxytoca Lactobacillus acidophilus Lactobacillus brevis Lactobacillus casei
ssp. rhamnosus *
Lactobacillus delbrueckii ssp. bulgaricus
Lactobacillus gasseri Lactobacillus plantarum
50011 50012 50014 50016 50018 50019 50092 50091 50021 50020 50024 50025 50026
50027
50028 50099
50030 50031 50032 50034 50035 50037 50038 50101 50043
50045
50050 50040 50054
10764 11563 11778 14580
11509
35586
13124
12916 19404 23355
8043
11775
13337
4356 14869
7469
11842 19992 14917
1914 869
2145
2423
5172
1825
978 193 20
847
4016
4020
550
376
4176 279
4102 515
158
748
7 080
79.03
54.8
A157
20 340 345
13
20171
20623 20624
756
1664
20160 2918
30083
20054
20021
20081 20077 20174
2763
2762
997
1989
1749
243
1752
11775 8012 9375
13061 10018
11952 13093 2 264 8645
506
6125
13079 532
8 260 8123
12672 11943 13059 11999 12819 8690
11973
8010
11778 13081 11974
8251 10320 10341 5 398
10822 11366 11168
6272 506
8237
8238 532
8213
9001 10090 8105
12712
HNCMB 86001; IFO 12219 HNCMB 100003 IFO 12200; JCM 2505
CCUG 11283 CCUG 6 824
JCM 1666
JCM 1059
BUCSAV 227; IAM 1118; IFO 3 425
IAM 12 477; JCM 1149

Bacteria NCIMB QC Collection
ATCC CCM CECT CIP DSM NCDO / N C F B
NCIMB NCTC Other
Lactobacillus sake Lactobacillus viridescens Lactococcus lactis
ssp. lactis Leuconostoc mesenteroides * Listeria innocua sv 6a * Listeria ivanovii sv 5 Listeria monocytogenes
sv 1 /2a Listeria monocytogenes
sv 4a * Listeria monocytogenes
sv 4b Listeria seeligeri *
sv 1 /2 Listeria welshimeri * Microbacterium lacticum * Micrococcus luteus Morganella morganii * Pediococcus damnosus * Photobacterium leiognathi Proteus mirabilis Pseudomonas aeruginosa Pseudomonas fluorescens Pseudomonas fragi Pseudomonas putida * Salmonella dublin * Salmonella enteritidis Salmonella saintpaul Salmonella typhimurium Salmonella typhimurium Salmonella virchow
50056 50057
50O58 50060 50094 50095
50000
50001 50062 50063 50064 50065 50103
50067 50068 50069 50070 50072 50073 50075 50100
50077
15521 12 706
19435 9135
33090 19119 35152
19114
13932
35967
35897 8180 4698
25 830 29358 25521 29906 25 668 13525 4973
12633
13311
1877 1851
169
2115 1974
906 56
185
910 913
934
935
917
51 173 793
4168 118 378 446 324 4152 4155 4153 443
4154
283
20481 20241
100100
81.49T
A 270 A 231
665
69.13 55.4 52.191
20017 20410
20650 20172 20030 30164 20331
50090 3456
291
2714 1655
604 522
747
1832
1524 752
13 090 8965
6681 6 992
8540 9278
235 12010
2193
13063 9046 8542 9494
6681
11288 11846 7973
5214
10527
11856
11857
2665 235
11938 10662 10038 10689 10936 9676 5188 6022
74 12190 5742
JCM 1 174
BUCSAV 302; HNCMB 80146 BUCSAV 312 CCUG 15 531; SLCC 3 379 CCUG 15 528; SLCC 2379
SLCC 3 954
SLCC 5 334
IAM 1056; IFO 3 333; JCM 1464 IFO 3 848
IAM 12 022; IFO 14160 IFO 3 458 IFO 14164
Bacteria NCIMB ATCC QC Collection
CCM CECT CIP DSM NCDO / N C F B
NCIMB NCTC Other
Shigella flexneri 50079 29 903 Shigella sonnei 29 930 Staphylococcus aureus 50080 6 538P Staphylococcus aureus 50 081 Staphylococcus warneri 50 082 Streptococcus boris Streptococcus salivarius ssp.
thermophilus 50 083 19 258 Vibrio cholerae non 0:1 Vibrio fluvialis 50 084 33 809 Vibrio parahaemolyticus Vibrio vulnifl'cus 50104 27 562 Yersinia enterocolitica
biotype 1 serotype 0 :8 50085 9610 biotype 4 serotype 0:3 50087
213
240
986
4782
53.156 346
20480 597
573
8625 12820 13078
8510
2249
2 046
7 447
10518 8177
11348 11327 11344
CCUG 11291 CCUG 4 586
Fungi NCIMB ATCC CMI NCYC Others
Eurotium amstelodami 16018 17 455 CECT 2 586 Aspergillus flavus 91856 ii Penicillium cyclopium 16 025 19 759 Rhizopus stolonifer 61269 Saccharomyces cerevisiae 50105 7 754 79 CBS 1368 Zygosaccharomyces rouxii 50106 1522

This Page Intentionally Left Blank

485
Subject Index
No attempt has been made to cite in the Index all
the media mentioned in Part 1 ( Review Sect ion) .
The media cited are mostly those covered in the
monographs and are to be found in Part 2 ( pp.
251-472). Other media mentioned in the reviews
are either of mainly historical interest or new, and
as yet, untested media and hence are not cited
separately in the Index.
Absolute and relative growth indices, 473
Absolute growth index, 13,473
Acriflavine selective properties, 312, 318, 322, 346,
383,397,401,463
Aeromonas spp:
Review article, 111-127
General characteristics, 111 - 112
Isolation and enumeration, 113-120
Enrichment procedures, 119
Identification, 119-120
Incubation temperature, 119
Plating media for food and environmental
samples, 114-119
Recovery of sub-lethally injured cells, 118-
119, 124 Starch ampicillin agar, 111, 114-115, 117
AFPA - see Aspergillusflavus and parasiticus agar
AGI - see Absolute and relative growth indices
Alkaline peptone water, 203,206-209,215
All Purpose Tween agar - modified (for H202 detec-
tion), 97, 99-100, 103,251
APT - see All Purpose Tween agar - modified (for
H202 detection)
APW - see Alkaline peptone water
Aspergillusflavus and parasiticus agar, 235,254
Bacillus cereus :
Review article, 35-49
Enumeration in foods, 45-47
Inoculation and culture conditions, 43-45
Media for isolation and enumeration, 36-40
Mode of action of diagnostic systems in iso-
lation media, 41-43
Mode of action of selective systems in isola- tion media, 40-41
MPN technique, 36
Rationale for screening foods for B.cereus,
35-36
Resuscitation of sub-lethally injured orga-
nisms, 47
Bacillus cereus, recommended media
Mannitol egg yolk polymyxin agar, 367
Polymyxin egg yolk mannitol bromothymol blue
agar, 402
Baird-Parker agar, 77, 79-84, 257
Baird-Parker liquid medium, 79-80, 84, 261
BGB - see Brilliant Green Bile Broth
Bile Oxalate Sorbose broth, 219, 221-222, 225-226,
263
Bismuth sulphite agar, 188-190, 266
BOS - see Bile Oxalate Sorbose broth
Briggs agar, 90, 99-100, 103,269
Brilliant Green Bile Broth, 166, 173-175, 180-181,
272
Brilliant green specification, 394
Brochothrix spp., 93-94
Brochothrix thermosphacta, recommended medium
Streptomycin thallous acetate actidione agar, 442
Campylobacters:
Review article, 129-162
Classification, 130
Antibiotic activity, 139
Atmosphere during incubation of enrich-
ment media, 149
Basal media, 135
Blood ( reason for incorporation), 135
Choice of antibiotics, 138
Comparisons of media, 149-152
Effect of damage on cell viability, 147-149
Enrichment methods for isolation from food,
145-147
FBP and other bloodless supplements, 135-
138
Isolation from foods, 153-155
Membrane filtration method, 135,145, 151-
152

486 Subject lndex
MPN technique, 144, 146, 153
pH, 138
Recovery of sub-lethally injured cells, 147-
149
Campylobacter spp., recommended media
Charcoal cefoperazone deoxycholate agar, 284
Charcoal cefoperazone deoxycholate broth, 287
Preston campylobacter selective agar, 405
Preston enrichment broth, 408
Skirrow campylobacter selective agar, 440
Carnobacterium spp., 97-99
Carnobacterium spp., recommended media
Cresol red thallium acetate sucrose agar, 293
CATC - see Citrate Azide Tween Carbonate agar
CCD - see Charcoal Cefoperazone Deoxycholate
agar- modified and Charcoal Cefoperazone
Deoxycholate broth
Cefsulodin Irgasan Novobiocin agar, 219, 222-223,
225-226, 275
Cellobiose Polymyxin B Colistin agar, 210-213, 215,
278
Cephaloridine Fucidin Cetrimide agar, 111,123-124,
281
CFC - see Cephaloridine Fucidin Cetrimide agar
Charcoal Cefoperazone Deoxycholate agar - modi-
fied, 134,137, 139, 143-145, 149, 151-155,
284
Charcoal Cefoperazone Deoxycholate broth, 134,
136, 139, 143-145, 154, 287
CIN - see Cefsulodin Irgasan Novobiocin agar
Citrate Azide Tween Carbonate agar, 51,54-59, 290
Clostridia spp:
Review article, 25-33
Contaminants in food, 25-26
Factors affecting isolation, 26-27
MPN technique, 27-30
Recovery of sub-lethally injured cells, 25, 32
Sample preparation and conditions of
anaerobiosis, 31-32
Types of isolation media 27-31
Clostridium pelfringens, recommended media
Differential reinforced clostridial agar (modi-
fied), 306
Iron sulphite agar, 332
Oleandomycin polymyxin sulphadiazine perfrin-
gens agar, 378
Rapid perfringens medium, 413
Sulphite cycloserine azide agar, 445
Tryptose sulphite cycloserine agar, 458
Coli-aerogenes group ( see also Enterobacteriaceae),
recommended media
Brilliant green bile broth, 272
Lauryl tryptose broth, 343
Coliforms:
Review article - see Enterobacteriaceae, 163-185
CPC - see Cellobiose Polymyxin B Colistin agar
Cresol red Thallium Acetate Sucrose agar, 98-99, 293
CTAS - see Cresol red Thallium Acetate Sucrose agar
Culture collections, 479-483
DG 18 - see Dichloran Glycerol agar
Diagnostic Salmonella Selective Semisolid Medium,
296
DIASALM - see Diagnostic Salmonella Selective
Semisolid Medium
Dichloran Glycerol agar, 229, 233,235,237-238, 300
Dichloran Rose Bengal Chloramphenicol agar, 229,
232, 303
Differential Reinforced Clostridial agar - Modified,
28-29, 306
Dilution to extinction technique - see Methods
Dominguez Rodriguez LSAMm agar, 68-69, 71,309
DRBC - see Dichloran Rose Bengal Chlorampheni-
col agar
DRCM - see Differential Reinforced Clostridial
agar ( Modified )
Ecometry - see Methods
EE - see Enterobacteriaceae Enrichment broth
Electivity, 3
Enterobacteriaceae Enrichment broth, 167-168, 174,
180-181,314
Enterobacteriaceae:
Review article, 163-185
Classification, 163-164 Comparative studies of media, 173-177
Filtration methods 169-172
Incubation conditions, 172-173
Incubation temperature, 165, 166-167, 172-
173, 175-176, 178, 182
Liquid media, 165-168
MacConkey agar, 165
Minerals Modified Glutamate (MMG) media,
165, 167, 170, 173-174, 181
MPN technique, 172-177, 179-181
Recovery of sub-lethally injured organisms,
165, 174-176, 182
Solid media, 168- 1 6 9
Enterobacteriaceae, recommended media
Enterobacteriaceae enrichment broth, 314
Violet red bile glucose agar, 464
Violet red bile agar, 467

Subject Index 487
Enterococci:
Review article, 51-61
Distribution of enterococci and streptococci,
51-52
Ecological and serological grouping, 52-55
Recovery of sub-lethally damaged cells, 60
Requirements and composition of selective
isolation media, 55-56
Selective media for enterococcci from food,
56-60
Selective procedures, 60
Enterococcus spp., recommeded media
Citrate azide tween carbonate agar, 290
Kanamycin aesculin azide agar, 335
M- enterococcus agar, 372
Enteropathogenic E.coli, 177-179
HC agar, 179
Sorbitol MacConkey agar, 178-179
Error associated with colony counts, 6-9
Escherichia coli :
Review article - see Enterobacteriaceae, 163-185
Escherichia coli, recommended media
Brilliant green bile broth, 272
Enterobacteriaceae enrichment broth, 314
Lauryl tryptose broth, 343
Tryptone bile agar, 453
Violet red bile glucose agar, 464
Violet red bile agar, 467
European Community Bureau of Reference, 2
FDA Listeria enrichment broth, 64-67, 70-71,317
Fraser broth - modified, 64-67, 70-71,320
GCB - see Giolitti and Cantoni Broth
Giolitti and Cantoni Broth, 79-80, 84, 323
Glucose Salt Teepol Broth, 203,208-209
Gram-positive, non-sporulating food spoilage bacte-
ria:
Review article, 89-109
Aerococcus, 97
Brevibacterium, 92-93
Brochothrix, 93-94
Carnobacterium, 97-99
Commercial media 90-92
Kurthia, 94-95
Lactobacillus, Leuconostoc and Pediococ-
cus, 99-105
Microbacterium, 95
Miclwcoccus, 95-96
Propionibacterium, 96-97
Spoilage association 89-91
Growth assessment in broth - see Methods
GSTB - see Glucose Salt Teepol Broth
HE - see Hektoen Enteric agar
Hektoen Enteric agar, 188-189, 326
IBB - see Inositol Brilliant green Bile salts agar
Inositol Brilliant green Bile salts agar, 111,121
Irgasan Ticarcillin Chlorate broth, 219,221-222,225-
226,329
Iron sulphite agar, 332
ITC - see Irgasan Ticarcillin Chlorate broth
KAA - see Kanamycin Aesculin Azide agar
Kanamycin Aesculin Azide agar, 51, 57-59, 335
Karmali agar, 134, 136-137, 144, 151-152
KRANEP- see Kalium- Rhodanid Actidione Natriu-
mazid Eigelb Pyruvat
Kalium- Rhodanid Actidione Natriumazid Eigelb
Pyruvat agar, 80-82,337
Lactic acid bacteria, recommended media
All purpose tween agar-modified, 251
Briggs agar, 269
L-S differential agar, 354
M- 17 agar, 360
de Man, Rogosa and Sharpe agar, 362
de Man, Rogosa and Sharpe agar with sorbic acid
( pH 6.2), 364
Lactobacilli ( see also Lactic acid bacteria), recom-
mended media
Briggs agar, 269
Lactobacillus sorbic acid agar, 340
de Man, Rogosa and Sharpe agar, 362
Rogosa agar, 425
Lactobacillus spp., 99-105
Lactobacillus Sorbic acid agar, 99, 101-102,340
Lactococcus lactis ssp. lactis, recommended media
Briggs agar, 269
de Man, Rogosa and Sharpe agar, 362
de Man, Rogosa and Sharpe agar with sorbic
acid, 364
Rogosa agar ( pH 6.2), 428
LaS - see Lactobacillus Sorbic acid agar
Lauryl tryptose broth (syn. lauryl sulphate broth),
167-168, 172-175, 180-181,343
LBP - see Baird-Parker liquid medium
LCT - see Lithium chloride Ceftazidime Tween 80
agar
Leuconostoc spp., 99-105 and see Lactic acid bacteria

488 Subject Index
Levinthal broth with acriflavine and nalidixic acid,
64-65,345
LICNR - see Lysine-Iron-Cystine-Neutral Red broth
Listeria monocytogenes :
Review article, 63-75
Enumeration in foods by direct plating, 71-
72
Listeria enrichment broths, 64-66
Listeria selective agars, 67-70
MPN technique, 71
Outbreaks of human listeriosis, 63-64
Procedures for isolation of L.monocytoge-
nes, 70-71
Recovery of sub-lethally injured Listeria
monocytogenes, 66-67
Listeria spp., recommended media
Dominguez Rodriguez LSAMm agar, 309
FDA listeria enrichment broth, 317
Fraser broth, 320
Levinthal broth with acriflavine and nalidixic
acid, 345
Lithium chloride ceftazidime tween 80 agar, 347
Lithium chloride phenylethanol moxalactam
agar, 350
Oxford agar, 381
Oxford agar-modified, 385
Polymyxin acriflavine lithium chloride ceftazidi-
me aesculin mannitol agar, 395
Polymyxin acriflavine lithium chloride ceftazidi-
me aesculin mannitol egg yolk broth, 399
University of Vermont broths I and II, 461
Lithium chloride Ceftazidime Tween 80 agar, 347
Lithium chloride Phenylethanol Moxalactam agar,
68-71,350
L-PALCAMY - see Polymyxin Acriflavine Lithium
Chloride Ceftazidime Aesculin Mannitol
Egg Yolk broth
LPM - see Lithium chloride Phenylethanol Moxalac-
tam agar
LSAMm - see Dominguez Rodriguez agar
LSD - see L-S Differential agar
L-S Differential agar, 100, 354
LST - see Lauryl tryptose broth
Lysine-Iron-Cystine-Neutral Red broth, 357
M 17 agar, 100, 360
de Man, Rogosa and Sharpe agar, 89-90,97, 99-
100,102-103,362
de Man, Rogosa and Sharpe agar with sorbic acid, 99-
100, 102, 104, 364
Mannitol Egg Yolk Polymyxin agar, 38-47, 367
Mannitol Lysine Crystal violet Brilliant green agar,
189-190, 369
MC - see Oxford agar-Modified
mDRCM - see Differential Reinforced Clostridial
agar - Modified
ME - see M-Enterococcus agar
M- Enterococcus agar, 51, 57-59, 372
Methods
Liquid media
Dilution to extinction ( serial dilution techni-
que), 14-19,476
Growth assessment in broth, 477
Most Probable Number ( MPN ), 15-18
Performance assessment with mixed cultu-
res, 476
Plating media
Ecometry, 4-6, 12-13,474
Modified Miles Misra, 3-4, 6, 474
Stab inoculation, 5,475
Streaking method (Ecometry), 474
MEYP - see Mannitol Egg Yolk Polymyxin agar
Miles- Misra modified technique - see Methods
MLCB -see Mannitol Lysine Crystal violet Brilliant
green agar
Modified brilliant green agar see - Phenol red brilliant
green agar
Monographs, 251- 472
Notes on the use of, 243
Most Probable Number technique - see Methods
Moulds and yeasts :
Review article, 229-242
Culture film method, 232
Detection and enumeration in food, 230
Diluent, 231
General purpose culture media, 232-233
Heat-resistant moulds, 234
Incubation, 238
Interpretation of data, 238
Membrane filtration technique, 232
Mycogenic moulds, 234
Plating technique, 237
Precautions, 239
Preparation of samples, 230-231
Resuscitation of sub-lethally injured cells,
231
Sampling procedure, 230
Selective media for yeasts, 236-237
Xerophilic fungi, 233
Moulds, recommended media
Aspergillusflavus and parasiticus agar, 254
Dichloran glycerol (DG 18) agar, 300

Subject Index 489
Dichloran rose bengal chloramphenicol agar, 303
Oxytetracycline glucose yeast extract agar, 388
MRS - see de Man, Rogosa and Sharpe agar
MRS-S - see de Man, Rogosa and Sharpe agar with
sorbic acid
MSRV - see Rappaport-Vassiliadis Medium - Semi-
solid modification
Muller Kauffmann tetrathionate broth, 191-193,374
OGY -see Oxytetracycline Glucose Yeast extract agar
Oleandomycin Polymyxin Sulphadiazine Perfringens
Agar, 378
OPSPA - see Oleandomycin Polymyxin Sulphadiazi-
ne Perfringens Agar
Oxford agar, 68-71,381
Oxford agar-Modified, 68-71,385
Oxytetracycline Glucose Yeast extract agar, 229, 232,
236, 388
PALCAM - see Polymyxin Acriflavine Lithium
Chloride Ceftazidime Aesculin Mannitol
agar
Park and Sanders broth, 134, 148, 153-154
Pediococcus spp., 99-105
Pediococci, recommended media
Briggs agar, 269
de Man, Rogosa and Sharpe agar, 362
de Man, Rogosa and Sharpe agar with sorbic
acid, 364
Rogosa agar ( pH 6.2), 428
PEMBA - see Polymyxin Egg Yolk Mannitol Bromo-
thymol blue agar
Performance assessment with mixed cultures - see
Methods
Pharmacopoeia of Culture Media, 243
Phenol red brilliant green agar, 391
PL - see Plesiomonas agar
Plesiomonas agar, 111, 121
Plesiomonas shigelloides :
Review article, 120-122
Enrichment procedures, 122
Incubation temperature, 121-122
Recovery of sub-lethally injured cells, 121
Polymyxin Acriflavine Lithium Chloride Ceftazidi-
me Aesculin Mannitol agar, 68-71,395
Polymyxin Acriflavine Lithium Chloride Ceftazidi-
me Aesculin Mannitol Egg Yolk broth, 64-
66, 70, 399
Polymyxin Egg Yolk Mannitol Bromothymol blue
agar, 38-47,402
Pour plate technique, 3
Preston campylobacter selective agar, 134-136, 138-
139, 145, 147, 150-155,405
Preston enrichment broth, 134-136, 139, 145, 149,
155,408
Productivity ratios, definition of, 473
Propylene glycol deoxycholate neutral red agar - see
Rambach agar
PRs - see Productivity ratios, definition of
Pseudomonas spp:
Review article, 122-124
Pseudomonas spp., recommended media
Cephaloridine fucidin cetrimide agar
PTSBS - see Tryptone Soya Broth with 10% NaC1
and 1% Sodium Pyruvate
QC methods for liquid media, 14-19
Statistics of methods for QC of liquid media, 15 -
19
Comparison of methods used for QC of liquid
media, 19
QC methods for solid media, 2-14
Statistics of methods for QC of solid media, 6-13
Comparison of methods used for QC of solid
media, 13-14
Quality assessment of culture media, 1-23
- see also under Methods
Rambach agar, 189, 410
Rapid Perfringens Medium, 413
Rappaport medium-semisolid modification, 191 - 192,
195-196, 419
Rappaport-Vassiliadis broth, 187, 189, 191-193, 195,
416
Rappaport-Vassiliadis Medium-Semisolid modifica-
tion, 195-196,422
RBC - see Rose Bengal Chloramphenicol agar
Relative growth index - see Absolute and relative
growth indices
Resuscitation of sub-lethally injured cells, 231,244
RGI - see Absolute and relative growth indices
Rogosa agar, 89, 92, 97, 99-100, 103-104, 425
Rogosa agar modified ( pH 6.2), 99-100, 428
Rose Bengal Chloramphenicol agar, 431
RPM - see Rapid Perfringens Medium
RV - see Rappaport-Vassiliadis broth
SAA - see Starch Ampicillin Agar
Salmonella:
Review article, 187-201
Bile salt media, 188-189
Brilliant green media, 189-190
Bismuth sulphite agar, 190-191

490 Subject lndex
Magnesium content of Rappaport-Vassilia-
dis broth, 193-194
Rappaport-Vassiliadis medium and modifi-
cations, 192-196
Selective enrichment and competitive bacte-
ria, 196-198
Salmonella Shigella agar, 188-189
Salmonella Shigella Deoxycholate Calcium agar,
219, 222, 225-226, 434
Salmonella spp. ( see also Enterobacteriaceae),
recommended media
Bismuth sulphite agar, 266
Diagnostic salmonella selective semisolid medi-
um, 296
Hektoen enteric agar, 326
Lysine iron cystine neutral red broth, 357
Mannitol lysine crystal violet brilliant green
agar, 369
Muller Kauffmann tetrathionate broth, 374
Phenol red brilliant green agar, 391
Rambach agar, 410
Rappaport Vassiliadis broth, 416
Rappaport Vassiliadis medium -semisolid modi-
fication, 422
Selenite cystine broth, 437
Xylose lysine deoxycholate agar, 470
Salt polymyxin broth, 203,209
SCA - see Sulphite Cycloserine Azide agar
Selenite cystine broth, 191-192,437
Serial dilution technique - see Methods
Shigella spp. (see also Enterobacteriaceae), recom-
mended media
Violet red bile agar, 467
Xylose lysine deoxycholate agar, 470
Skirrow Campylobacter selective agar, 134, 136, 139,
143, 147, 150-152, 154-155,440
Sorbic acid agar base - see Lactobacillus Sorbic acid
agar
SPB - see Salt polymyxin broth
Spiral plate technique, 3-4, 6
Spread plate technique, 3, 6
SPS - see Sulphite polymyxin sulphadiazine agar
SR - see Rappaport medium-semisolid modification
SS - see Salmonella Shigella agar
SSDC - see Salmonella Shigella Deoxycholate Calci-
um agar
STAA - see Streptomycin Thallous Acetate Actidione
agar
Stab inoculation technique - see Methods
Standardized reference strains, 2
Staphylococcus aureus :
Review article, 77-87
Comparative studies of media, 80-83
Detection of low numbers, 83-84
MPN technique, 77, 80, 83-84
Rapid identification methods, 84
Recovery of sub-lethally injured cells, 77,
79-84
Selective culture media for isolation and
enumeration, 79-83
Staphylococcal foodborne disease, 77-79
Staphylococcal enterotoxins, 77-79
Staphylococcus aureus, recommended media
Baird-Parker agar, 257
Baird-Parker liquid medium, 261
Giolitti and Cantoni broth, 323
KRANEP agar, 337
Tryptone soya broth with 10% NaC1 and 1%
sodium pyruvate, 456
Storage of media, 60, 138
Streaking method - see Methods
Starch Ampicillin Agar, 111, 114, 116-117
Streptococci, Group D:
Review article - see Enterococci, 51-61
Streptococcus boris, recommended media
Kanamycin aesculin azide agar, 335
M-enterococcus agar, 372
Streptococcus thermophilus, recommended media
Briggs agar, 269
Streptomycin Thallous Acetate Actidione agar, 93-
94, 442
Sulphite Cycloserine Azide agar, 445
Sulphite polymyxin sulphadiazine agar, 29-30
Summary of organisms and recommended media,
247
Surface plating techniques, 6, 13
TBA - see Tryptone Bile Agar
TCBS - see Thiosulphate Citrate Bile-salt Sucrose
agar
Test strains, 479
Testing methods for use in quality assurance of culture
media, 473
Thallous Acetate Tetrazolium Glucose agar, 51, 57-
60, 448
Thiosulphate Citrate Bile-salt Sucrose agar, 203,209-
213,215,450
TITG - see Thallous Acetate Tetrazolium Glucose
agar
Tryptone BileAgar, 169,171-175, 178, 181,453
Tryptone Soya Broth with 10% NaCI and 1% Sodium
Pyruvate, 77, 80, 83-84, 456

Subject Index 491
Tryptose Sulphite Cycloserine agar (without egg
yolk), 458 TSC - see Tryptose Sulphite Cycloserine agar (with-
out egg yolk)
University of Vermont broths I and II, 65-67, 70-71,
461
UVM - see University of Vermont broths 1 and II
Vibrio : Review article, 203-217
Classification, 204
Enrichment media for V.cholerae, 206-208
Enrichment media for V.parahaemolyticus,
208-209
Enrichment media for V.vulnificus, 209
Mode of action of selective systems in culture
media, 214-215
MPN technique, 206
Plating media for V.cholerae, 209-211
Plating media for V.parahaemolyticus, 211-
213
Plating media for V. vulnificus, 213-214
Rationale for screening foods, 204-206
Ecology, 204
Mode of infection, 205
Methods of detection and control, 206
Recommended culture media, 215
Resuscitation of sub-lethally injured orga-
nisms, 206
Vibrio spp., recommended media
Cellobiose polymyxin B colistin agar, 278
Thiosulphate citrate bile salts agar, 450
Violet Red Bile agar, 171, 173, 175-176, 180-181, 467
Violet Red Bile Glucose agar, 169, 171, 173, 175,
180-181,464
Violet red bile lactose agar - see Violet Red Bile agar
VRB - see Violet Red Bile agar
VRBG - see Violet Red Bile Glucose agar
XLD - see Xylose Lysine Deoxycholate agar
Xylose Lysine Deoxycholate agar, 188-189, 470
Yeasts and moulds:
Review article, 229-242
Yersinia enterocolitica :
Review ariicle, 219-227
Classification, 219-220
Comparative studies, 225
Enrichment, 220-221
Identification, 223-225
Plating media, 221-223
Potassium hydroxide treatment, 221,225
Resuscitation of sub-lethally injured cells,
221
Serogroups associated with human disease,
220-221,223-224, 226
Yersinia spp. (see also Enterobacteriaceae), recom-
mended media
Bile oxalate sorbose broth, 263
Cefsulodin irgasan novobiocin agar, 275
Irgasan ticarcillin chlorate broth, 329
Salmonella shigella deoxycholate calcium agar,
434
Wilson and Blair medium - see Bismuth sulphite agar
Related Documents











