Flora 203 (2008) 449–468 Flower morphology, phenology and visitor patterns in an alpine community on Mt Olympos, Greece $ Nikolaos Makrodimos a , George J. Blionis a , Nikolaos Krigas b , Despoina Vokou a, a Department of Ecology, School of Biology, Aristotle University, GR-54124 Thessaloniki, Greece b Department of Botany, School of Biology, Aristotle University, GR-54124 Thessaloniki, Greece Received 10 April 2007; accepted 25 July 2007 Abstract Alpine vegetation, restricted to the top of high mountains, is among the vegetation types most endangered by global warming, currently predicted to raise temperatures from 1.1 to 6.4 1C, by the end of the century. Nevertheless, background information allowing evaluation of impacts is rather scarce for some geographic zones. Our study of an alpine community on the Plateau of Muses (2600–2750 m a.s.l.) of Mt Olympos, the highest mountain of Greece, conducted in 1993–1994, can provide such background information for the Mediterranean region. We studied features relating to phenology of flowering, floral morphology, distribution and abundance, and flower visitors of plant species that exhibit a biotic pollination syndrome. We identified dominant patterns and we further (i) explored the relative contribution of the plant features and abiotic factors studied in explaining the activity patterns of flower visitors, (ii) examined if flower and visitor traits of the alpine community match each other according to the classical pollination syndromes, and (iii) investigated whether the responses of individual plant species to the yearly climatic variability result into phenological patterns that characterize the whole community. The common strategy of the alpine community was for early flowering and long flower life span; consistently early flowering species were twice as many as late flowering ones, whereas floral longevity (estimated for 36 species) averaged 5.2 days. Duration of flowering (estimated for 57 species) averaged 18.2 days. Climatic variability affected onset of flowering; all late flowering species delayed their flowering during the year characterized by a humid and cold summer. Duration of flowering and floral longevity did not change in a consistent way. Hymenoptera (Aculeates) were the dominant flower visitors. They accounted for 43.3% of the visits recorded, with bumblebees making a little less than half. Diptera followed making 37.5% of the visits (most made by syrphid flies). There was a mismatch between flower-morphology and flower visitor traits; the alpine community had predominantly non-specialized, pale-colour flowers, which are traits assumed to correspond to Diptera dominance and absence of social bees. Visitation was influenced by flower abundance and duration of flowering; proportionately more Diptera, and proportionately less Hymenoptera visited species with short flowering periods and few flowers present in the field. In a number of cases, the phenological and flower visitor patterns of the community of Mt Olympos deviated from those observed in other alpine environments suggesting a mediterranean influence even at high altitudes. r 2008 Elsevier GmbH. All rights reserved. Keywords: Flowering; Insects; Mediterranean; Mountain; Pollination; Strategy ARTICLE IN PRESS www.elsevier.de/flora 0367-2530/$ - see front matter r 2008 Elsevier GmbH. All rights reserved. doi:10.1016/j.flora.2007.07.003 $ The paper is dedicated to the memory of M. Tziolas, our colleague and friend, who worked with us on this project. Corresponding author. Tel.: +30 2310 998323; fax: +30 2310 998379. E-mail address: [email protected] (D. Vokou).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
0367-2530/$ - se
doi:10.1016/j.flo
$The paper�CorrespondE-mail addr
Flora 203 (2008) 449–468
www.elsevier.de/flora
Flower morphology, phenology and visitor patterns in an alpine community
on Mt Olympos, Greece$
Nikolaos Makrodimosa, George J. Blionisa, Nikolaos Krigasb, Despoina Vokoua,�
aDepartment of Ecology, School of Biology, Aristotle University, GR-54124 Thessaloniki, GreecebDepartment of Botany, School of Biology, Aristotle University, GR-54124 Thessaloniki, Greece
Received 10 April 2007; accepted 25 July 2007
Abstract
Alpine vegetation, restricted to the top of high mountains, is among the vegetation types most endangered by globalwarming, currently predicted to raise temperatures from 1.1 to 6.4 1C, by the end of the century. Nevertheless,background information allowing evaluation of impacts is rather scarce for some geographic zones. Our study of analpine community on the Plateau of Muses (2600–2750ma.s.l.) of Mt Olympos, the highest mountain of Greece,conducted in 1993–1994, can provide such background information for the Mediterranean region. We studied featuresrelating to phenology of flowering, floral morphology, distribution and abundance, and flower visitors of plant speciesthat exhibit a biotic pollination syndrome. We identified dominant patterns and we further (i) explored the relativecontribution of the plant features and abiotic factors studied in explaining the activity patterns of flower visitors, (ii)examined if flower and visitor traits of the alpine community match each other according to the classical pollinationsyndromes, and (iii) investigated whether the responses of individual plant species to the yearly climatic variability resultinto phenological patterns that characterize the whole community. The common strategy of the alpine community wasfor early flowering and long flower life span; consistently early flowering species were twice as many as late floweringones, whereas floral longevity (estimated for 36 species) averaged 5.2 days. Duration of flowering (estimated for 57species) averaged 18.2 days. Climatic variability affected onset of flowering; all late flowering species delayed theirflowering during the year characterized by a humid and cold summer. Duration of flowering and floral longevity did notchange in a consistent way. Hymenoptera (Aculeates) were the dominant flower visitors. They accounted for 43.3% ofthe visits recorded, with bumblebees making a little less than half. Diptera followed making 37.5% of the visits (mostmade by syrphid flies). There was a mismatch between flower-morphology and flower visitor traits; the alpine communityhad predominantly non-specialized, pale-colour flowers, which are traits assumed to correspond to Diptera dominanceand absence of social bees. Visitation was influenced by flower abundance and duration of flowering; proportionatelymore Diptera, and proportionately less Hymenoptera visited species with short flowering periods and few flowers presentin the field. In a number of cases, the phenological and flower visitor patterns of the community of Mt Olympos deviatedfrom those observed in other alpine environments suggesting a mediterranean influence even at high altitudes.r 2008 Elsevier GmbH. All rights reserved.
Keywords: Flowering; Insects; Mediterranean; Mountain; Pollination; Strategy
e front matter r 2008 Elsevier GmbH. All rights reserved.
ra.2007.07.003
is dedicated to the memory of M. Tziolas, our colleague and friend, who worked with us on this project.
ing author. Tel.: +302310 998323; fax: +30 2310 998379.
ess: [email protected] (D. Vokou).

ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468450
Introduction
Flower and flower-visitor traits of alpine plants aredescribed from several mountains around the world[Chile (Arroyo et al., 1982, 1985), Japan (Yumoto,1986), Australia (Inouye and Pyke, 1988), Norway(Totland, 1993), Canary Islands (Dupont et al., 2003),USA (McCall and Primack, 1992; Moldenke, 1976;Warren et al., 1988)]. Insect diversity, abundance andactivity are often reported to decrease with elevation(Arroyo et al., 1985; Bingham and Orthner, 1998;Blionis and Vokou, 2001; Medan, 2002), whereascomposition of the pollinating fauna to shift fromHymenoptera to Diptera, i.e. from a more specialized toa more generalized type of pollinators (Inouye andPyke, 1988; Korner, 1999; McCall and Primack, 1992;Totland, 1993, 1994; Warren et al., 1988). Nevertheless,the often reported reduction in pollinator availability athigh altitudes does not necessarily restrict pollination;several mechanisms can compensate for it, like longerflower life-span (Arroyo et al., 1981; Blionis et al., 2001;Fabbro and Korner, 2004; Primack, 1985) or stigmaticreceptivity, or a shift to more efficient pollinators, likebumblebees (Bingham and Orthner, 1998; Korner,1999).
Regarding flower morphology, a feature repeatedlyreported is the open-flower dominated flora of alpinecommunities (Collins et al., 1983; Totland, 1993;Warren et al., 1988); open flowers allowing visits ofgeneralists rather than specialists are interpreted as anadaptation to unpredictable pollination conditions(Rathcke, 1988). In contrast, coloration has been amuch debated topic. According to Korner (1999), thehigh fragmentation of alpine habitats, combined with ashort flowering season and often windy climate shouldselect for showy flowers, but this is not the case for therich alpine flora of New Zealand, which (with fewexceptions) is a ‘white’ flora. Similar is the situation inAustralia, where Pickering and Stock (2004) found 127of 204 alpine species to have large showy flowers, butmost of them to be white (53.5%) or yellow (21.3%), asreported previously for this flora by Inouye and Pyke(1988). Pickering and Stock (2004) attributed thepredominance of plant taxa with pale flowers inthe alpine floras of Australia and New Zealand to theprevalence of flies that prefer pale colours and to theabsence of other types of potential pollinators that areattracted to bright colours, such as social bees and birds.
Alpine vegetation, restricted to the top of highmountains, is among the most endangered vegetationtypes by the predicted global warming of 1.1–6.4 1C, bythe end of the century (IPCC, 2007). It is thereforeimportant to have background information for alpinecommunities, which will permit future comparisons andevaluation of impacts. Among the traits that are ofparticular importance are those related to flower
phenology, because they could serve as indicators ofthe ongoing climatic change. Nevertheless, backgroundinformation is rather scarce for some geographic zoneslike the Mediterranean.
Greece is a mountainous country with more than 130peaks exceeding 2000m (Strid, 1986). Because of itsrelief, a sizeable part of its surface is climatically non-mediterranean, with alpine communities making aconsiderable part of its natural systems; similar is thesituation in other Mediterranean countries. However,the dominant features of their alpine communities arenot known. Neither is it known whether there is anyMediterranean influence in the shaping of these features.
To address these issues, we studied phenology offlowering, floral morphology, distribution and abun-dance, and flower visitors of plant species that exhibit abiotic pollination syndrome in an alpine community onMt Olympos, the highest mountain of Greece. Afteridentifying the dominant patterns, we (i) explored therelative contribution of the plant features and abioticfactors studied in explaining the activity patterns offlower visitors, (ii) investigated whether the flower andvisitor traits match each other according to the classicalpollination syndromes (Faegri and van der Pijl, 1979),and (iii) investigated whether the responses of individualplant species to the yearly climatic variability result intophenological patterns that characterize the whole com-munity. We hypothesized (a) the dominant strategy ofthe alpine plant community to be for early flowering, asis manifested in short season environments (Billings andMooney, 1968; Bliss, 1971; Inouye and Pyke, 1988;Totland, 1993) and for long flower life span, as ismanifested under low pollinator availability and withincreasing elevation (Arroyo et al., 1981, 1985; Ashmanand Schoen, 1994; Bingham and Orthner, 1998; Blionisand Vokou, 2001; Blionis et al., 2001; Evanhoe andGalloway, 2002; Primack, 1985; Rathcke, 2003), and (b)onset of flowering to be strongly affected by the yearlyclimatic variability, given its sensitivity to temperaturechanges, as many studies with simulated warming haveshown (Alatalo and Totland, 1997; Henry and Molau,1997; Price and Waser, 1998; Stenstrom et al., 1997;Suzuki and Kudo, 1997).
Materials and methods
The present study was conducted on the ‘Plateau ofMuses’ of Mt Olympos (Greece) (40140N, 221270E), atabout 2600–2750ma.s.l., during the snow-free periodsof 2 consecutive years (1993, 1994). The Plateau, locatednext to the mountain top, covers approximately 2.2 km2
and can be separated into two major habitat types: thescree and the meadow. On a daily basis during the studyperiod, we recorded at 08:00 a.m. maximum and

ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468 451
minimum temperatures, relative air humidity, andheight of precipitation (rainfall or snowfall).
All plant species encountered on the Plateau thatexhibit a biotic pollination syndrome (Faegri and vander Pijl, 1979) were recorded and documented byherbarium specimens; species not exhibiting a bioticpollination syndrome were excluded from the study.Taxonomic identification of specimens and nomencla-ture follows Mountain Flora of Greece (Strid, 1986;Strid and Tan, 1991). The full names and the traitsexamined per species are given in Appendix A.
For each species, we recorded in situ flower colourand shape. Each capitulum of a member of theAsteraceae was treated as a single flower. Based on theshape of the corolla, flowers were described as eithertubular or open; within the latter group, we furtherdistinguished the subgroup of capitate flowers. Wedistinguished two major classes with respect to corollacolour: pale-colour flowers (predominantly white andyellow) and bright-colour ones (predominantly violet-blue, purple, red and pink).
To have an estimate of species abundance anddistribution on the Plateau, we used 36 sampling units,18 for each of the two habitats (scree and meadow).They consisted of 5� 5m quadrats randomly selected.Within each sampling unit, we estimated local speciesabundance by means of 12 randomly selected smallquadrats (1� 1m); therefore, local abundance of aspecies equals its average density over the 12 smallquadrats of a sampling unit. Abundance of a species onthe Plateau (or at each of the two habitat types) equalsthe mean value of its local abundances over all samplingunits (or over the sampling units for the habitat). Thesum of sampling units, where species were encountered,equals their distribution on the Plateau.
Flowering was monitored on a regular basis, every 3days, in the most densely populated sites; 49 species werestudied in both years and another 8 species in 1 year.First and last day of anthesis were recorded as calendardays (day 1 equals January 1st). Duration of floweringwas defined as the difference between first and last dayof flowering. For some species, onset of floweringpreceded the beginning of our observations. This wasdue to snow melting progressing faster in someareas, and hence allowing early flowering species locatedthere to blossom, before the Plateau itself becameaccessible. Species were ranked according to the orderof anthesis, using first day of flowering as the primarycriterion and duration of flowering as a secondary one.For instance, species with same first day of floweringand short duration were ranked first, whereas those withsame first day but long duration of flowering wereranked second. Significance of differences in the speciesranking in the 2 years of study was assessed by use of thenon-parametric Wilcoxon’s Matched-Pairs Signed-Ranks test.
We estimated the flower life span of 36 species, 10 ofwhich were studied in both 1993 and 1994. For this,a total number of 388 flowers, 5–30 for each species,were marked and were observed daily from bud towilting.
Observations of flower visitors took place during thesecond year of study, in two 5� 5m plots, in thescree–meadow interface, selected the first year becauseof their relatively high species richness. As a measure ofa species floral display, we counted the number of itsflowers, within the experimental plots, around the peakof its flowering. Each observation of flower visitorslasted 15min. The sampling protocol comprised threeobservations per day, unless climatic conditions (rainfallor strong winds) did not allow visitor activity at thespecified hours, in consecutive hours between 8.00 and17.00. Flower visitors were identified in situ at thelevel of order, and in a few cases at lower levels(bumblebees and syrphid flies). We kept records of theplant species visited and of date and time. After the firstweek of observations, we also took measurements ofwind velocity (with a portable digital AM-4201 LU-TRON anemometer), solar light intensity (with aportable digital LX-102 LUTRON light meter), airtemperature and relative air humidity. We conducted119 observations of insect activity, which amount to1785min (or 30 h), in total; meteorological data areavailable for 95 of these observations. After each 15-minobservation, we allowed another 15min for capturinginsect visitors. Captured specimens were sent to insecttaxonomists for further identification (see Acknowl-edgements).
We used Regression Tree Analysis (RTA, also knownas classification and regression trees or CART) todetermine the morphological and phenological traitsthat can explain the size and composition of the flowervisiting entomofauna (Breiman et al., 1984; De’ath andFabricius, 2000). For this analysis, we used the S-Plus(Statistical Sciences, 1999) statistical software package.To study the effect of environmental parameters onflower visitors’ activity, we used the K-Mean Clustermethod to group the 15-min observation periods.Environmental data pertaining to each cluster wereanalysed by ANOVA to check for differences amongclusters. For all other analyses, the SPSS statisticalpackage was used.
Results
The flowering period of the entomophilous alpinecommunity approximates the snow free period; it lastedfor about 3 months in both years. In 1994, July wascolder than in 1993 (mean temperature7S.E.:9.270.44 1C for 1994, 10.970.58 1C for 1993, paired t-

ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468452
test: t ¼ 2.63, p ¼ 0.014, n ¼ 27), whereas Septemberwas hotter (11.870.45 1C for 1994, 9.770.74 1C for1993, paired t-test: t ¼ �3.25, p ¼ 0.0034, n ¼ 25); therewas no difference in August (12.270.47 1C for 1994,11.270.47 1C for 1993, paired t-test: t ¼ �1.12, p ¼
0.273, n ¼ 31). Total precipitation during the 3-monthsummer period was 61mm in 1993 (0mm in July, 31mmin August, and 30mm in September) and 265mm in1994 (205mm in July, 30mm in August, and 30mm inSeptember). As a result of the exceptionally wet July1994, precipitations in summer 1994 were almost fourtimes higher than in 1993.
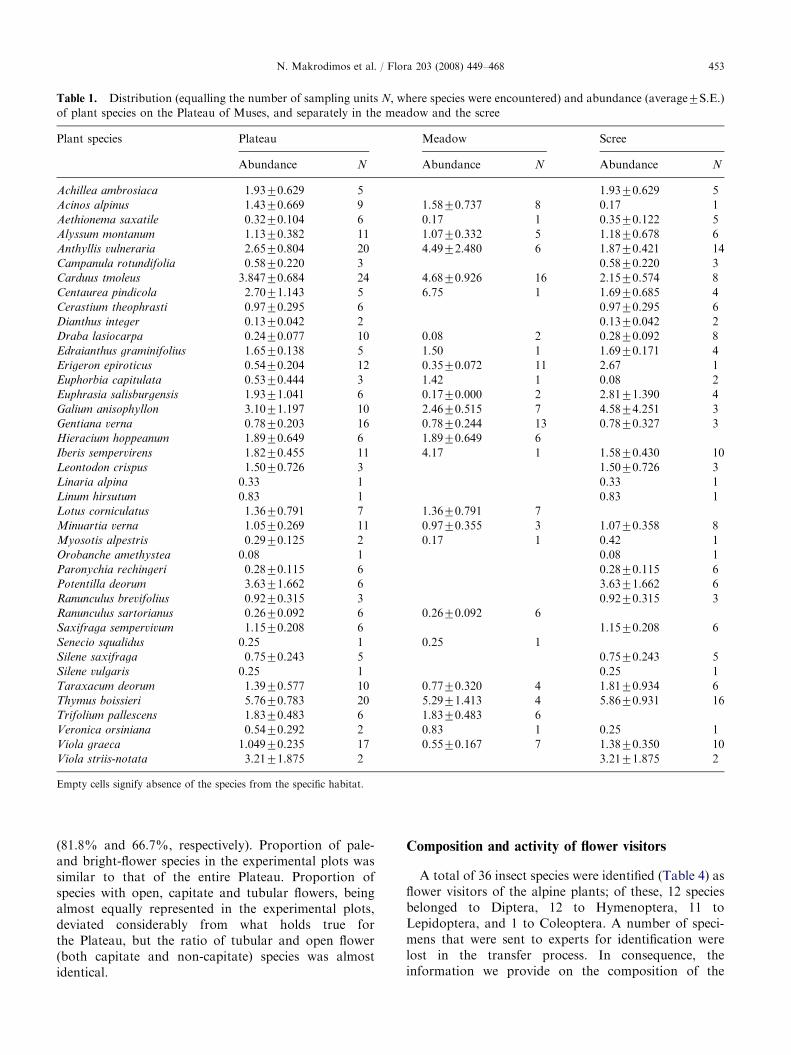
Species distribution and abundance
We recorded 61 angiosperm species displaying a bioticpollination syndrome (Appendix A). Of these, 41 have awide geographic distribution, whereas 20 are narrowlydistributed. In particular, 6 are single mountainendemics (occurring only on Mt Olympos), 2 are Greekendemics, and another 12 are Balkan endemics.Asteraceae (with 11 species) and Caryophyllaceae(with 8 species) are the most abundantly representedfamilies. We studied the distribution and abundance of40 species on the Plateau; of these, 15 were found only inthe scree and 5 only in the meadow. The large majorityof species on the Plateau (87.5%) were narrowlydistributed (found in less than 50% of the samplingunits). Carduus tmoleus, Thymus boissieri and Anthyllis
vulneraria were the most widespread (Table 1). Themost locally abundant species in both habitats wasT. boissieri; it was followed by C. tmoleus andA. vulneraria, in the meadow, and by Galium anisophyl-
lon and Potentilla deorum, in the scree. Though locallyabundant, Viola striis-notata had a limited distributionon the Plateau.
Flower morphology and phenology
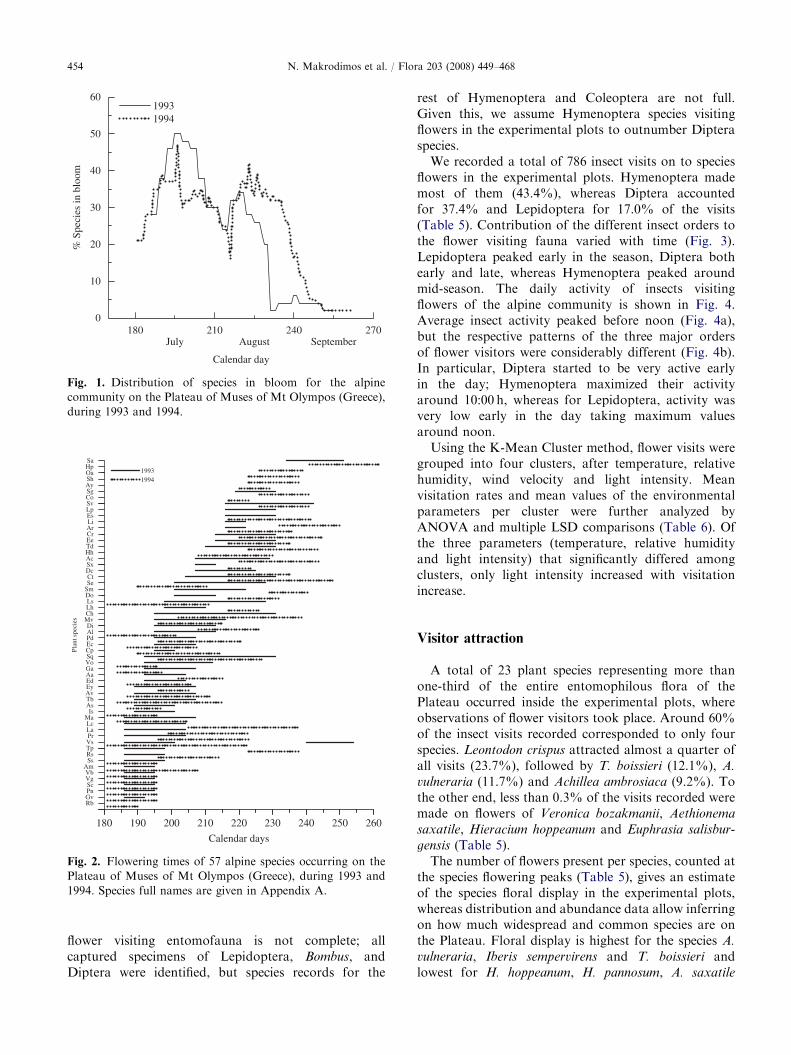
The distribution of species in bloom is bimodal inboth years of study (Fig. 1). This indicates existence oftwo groups, of the early and the late flowering species.Early flowering species outnumbered the late floweringones in 1994; the two groups were more equallyrepresented in 1993. Time and duration of flowering ofthe different species are shown in Fig. 2. The temporalsequence of species in bloom did not remain constantduring the 2 years of study (Wilcoxon’s Matched-PairsSigned-Ranks test, z ¼ �2.820, p ¼ 0.005). There wereshifts of the onset of flowering (32, in total) both earlierand later in the season, but the dominant pattern was fordelayed flowering in 1994. Because of the uncertaintyregarding the onset of flowering for a number of species(for the reasons explained in Materials and methods),we could not quantify all shifts observed. Of the 28
species exhibiting delayed flowering, shifts were quanti-fiable for 23 and were on average equal to 9.571.4 days.Of the 12 species exhibiting earlier flowering, shifts werequantifiable for 9 species equaling 6.071.0 days, onaverage (Fig. 2). V. striis-notata presented a peculiardouble flowering period in 1993.
To examine whether there is any pattern in floweringshifts, we classified species into three categories: earlyflowering (July), late flowering (August), and very lateflowering (September) species. For a species to beassigned to a certain category, at least 60% of thelength of its flowering period should fall within the limitsspecified for the category in both years. From a total of49 species that were studied in both years, 15 were quitelabile. Consistently early flowering species (23) made thelargest group. Consistently late flowering speciesamounted to 10, whereas only Sedum album floweredvery late in both years; all the latter 11 species, from nowon referred to as late flowering species, delayed theirflowering in 1994 (Fig. 2, Table 2).
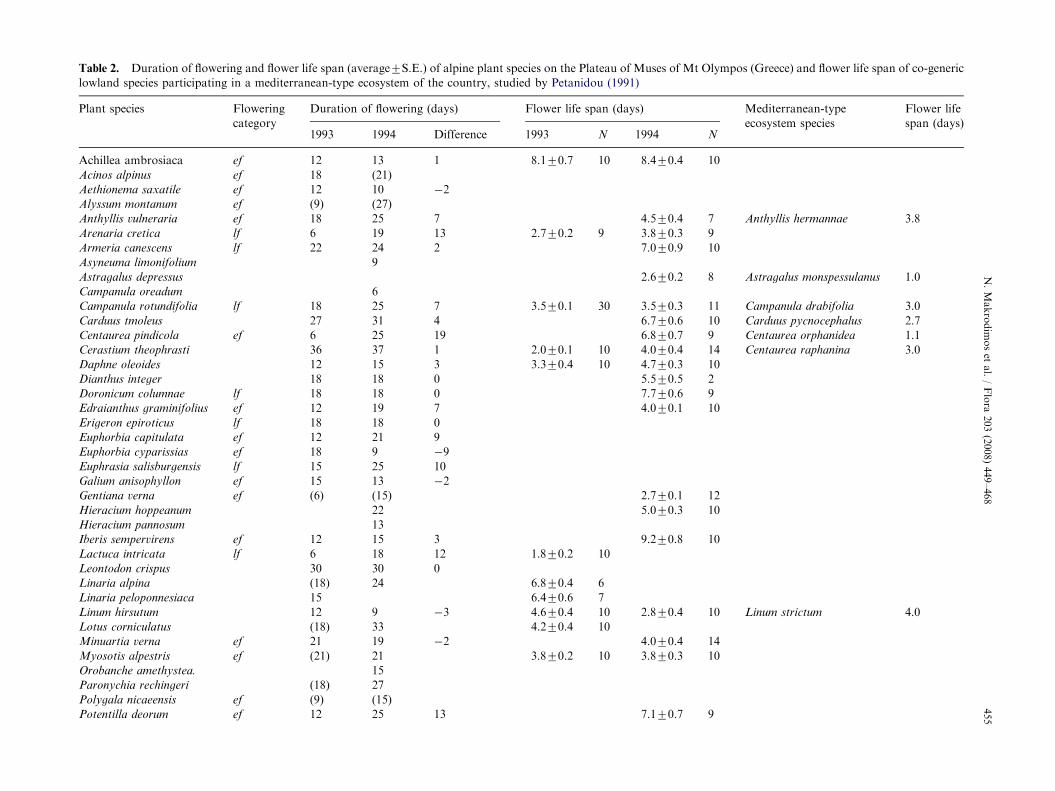
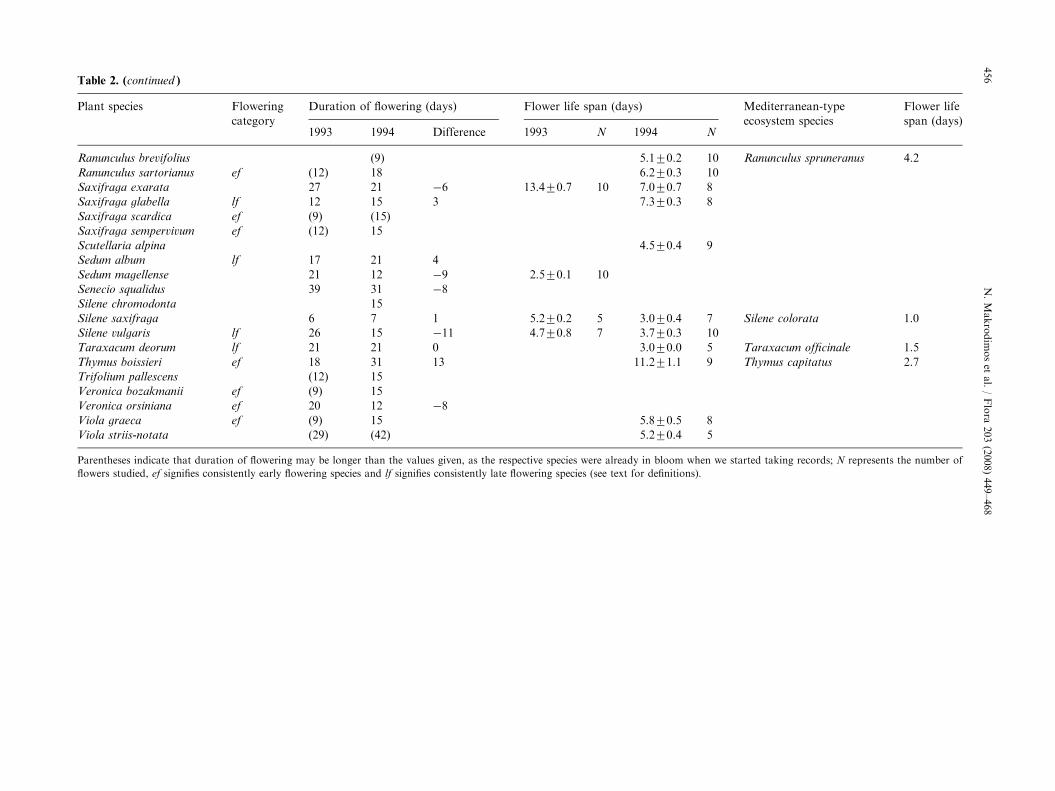
Duration of flowering of the different species rangedfrom 6 to 42 days at least, averaging 18.370.8days (Table 3); this value was estimated after bothyears of study and after only those species for whichwe knew the exact day of the onset of flowering.V. striis-notata had the longest duration. On average,species flowering lasted 2.171.2 days more in 1994,but the difference was not significant (paired t ¼ �1.71,p ¼ 0.097). Of the 34 species that could be compared(Table 2), duration in 1994 increased for 19 species(by 6.971.2 days), decreased for 10 species (by 6.071.1 days), and remained the same for five species.We did not find significant differences either, whenwe compared early and late flowering species, sepa-rately.
Flower life span ranged from 1.8 days in Lactuca
intricata to 13.4 days in Saxifraga exarata (Table 2)averaging 5.270.35 days. Between-year comparisonsthat could be made for 10 species did not revealany specific pattern of change; floral longevity waslonger for three species, shorter for four species, andnot differing for the remaining three; consequently,paired t-test analysis showed no significant difference(t ¼ 1.42, p ¼ 0.19) between years. Comparisonbetween the groups of early and late flowering speciesdid not reveal significant differences either (t ¼ �1.83,p ¼ 0.08).
Distribution of species found on the whole Plateauand in the experimental plots into different flowercolours and shapes is shown in Table 3. The alpine florahad predominantly open and pale-colour flowers; non-capitate, open-flower species accounted for half of thealpine flora displaying a biotic pollination syndrome,whereas bright-flower species accounted for only 36.1%.Open-flower species (both capitate and non-capitate)had a higher share of pale- than of bright-colour flowers
ARTICLE IN PRESS
Table 1. Distribution (equalling the number of sampling units N, where species were encountered) and abundance (average7S.E.)
of plant species on the Plateau of Muses, and separately in the meadow and the scree
Plant species Plateau Meadow Scree
Abundance N Abundance N Abundance N
Achillea ambrosiaca 1.9370.629 5 1.9370.629 5
Acinos alpinus 1.4370.669 9 1.5870.737 8 0.17 1
Aethionema saxatile 0.3270.104 6 0.17 1 0.3570.122 5
Alyssum montanum 1.1370.382 11 1.0770.332 5 1.1870.678 6
Anthyllis vulneraria 2.6570.804 20 4.4972.480 6 1.8770.421 14
Campanula rotundifolia 0.5870.220 3 0.5870.220 3
Carduus tmoleus 3.84770.684 24 4.6870.926 16 2.1570.574 8
Centaurea pindicola 2.7071.143 5 6.75 1 1.6970.685 4
Cerastium theophrasti 0.9770.295 6 0.9770.295 6
Dianthus integer 0.1370.042 2 0.1370.042 2
Draba lasiocarpa 0.2470.077 10 0.08 2 0.2870.092 8
Edraianthus graminifolius 1.6570.138 5 1.50 1 1.6970.171 4
Erigeron epiroticus 0.5470.204 12 0.3570.072 11 2.67 1
Euphorbia capitulata 0.5370.444 3 1.42 1 0.08 2
Euphrasia salisburgensis 1.9371.041 6 0.1770.000 2 2.8171.390 4
Galium anisophyllon 3.1071.197 10 2.4670.515 7 4.5874.251 3
Gentiana verna 0.7870.203 16 0.7870.244 13 0.7870.327 3
Hieracium hoppeanum 1.8970.649 6 1.8970.649 6
Iberis sempervirens 1.8270.455 11 4.17 1 1.5870.430 10
Leontodon crispus 1.5070.726 3 1.5070.726 3
Linaria alpina 0.33 1 0.33 1
Linum hirsutum 0.83 1 0.83 1
Lotus corniculatus 1.3670.791 7 1.3670.791 7
Minuartia verna 1.0570.269 11 0.9770.355 3 1.0770.358 8
Myosotis alpestris 0.2970.125 2 0.17 1 0.42 1
Orobanche amethystea 0.08 1 0.08 1
Paronychia rechingeri 0.2870.115 6 0.2870.115 6
Potentilla deorum 3.6371.662 6 3.6371.662 6
Ranunculus brevifolius 0.9270.315 3 0.9270.315 3
Ranunculus sartorianus 0.2670.092 6 0.2670.092 6
Saxifraga sempervivum 1.1570.208 6 1.1570.208 6
Senecio squalidus 0.25 1 0.25 1
Silene saxifraga 0.7570.243 5 0.7570.243 5
Silene vulgaris 0.25 1 0.25 1
Taraxacum deorum 1.3970.577 10 0.7770.320 4 1.8170.934 6
Thymus boissieri 5.7670.783 20 5.2971.413 4 5.8670.931 16
Trifolium pallescens 1.8370.483 6 1.8370.483 6
Veronica orsiniana 0.5470.292 2 0.83 1 0.25 1
Viola graeca 1.04970.235 17 0.5570.167 7 1.3870.350 10
Viola striis-notata 3.2171.875 2 3.2171.875 2
Empty cells signify absence of the species from the specific habitat.
N. Makrodimos et al. / Flora 203 (2008) 449–468 453
(81.8% and 66.7%, respectively). Proportion of pale-and bright-flower species in the experimental plots wassimilar to that of the entire Plateau. Proportion ofspecies with open, capitate and tubular flowers, beingalmost equally represented in the experimental plots,deviated considerably from what holds true forthe Plateau, but the ratio of tubular and open flower(both capitate and non-capitate) species was almostidentical.
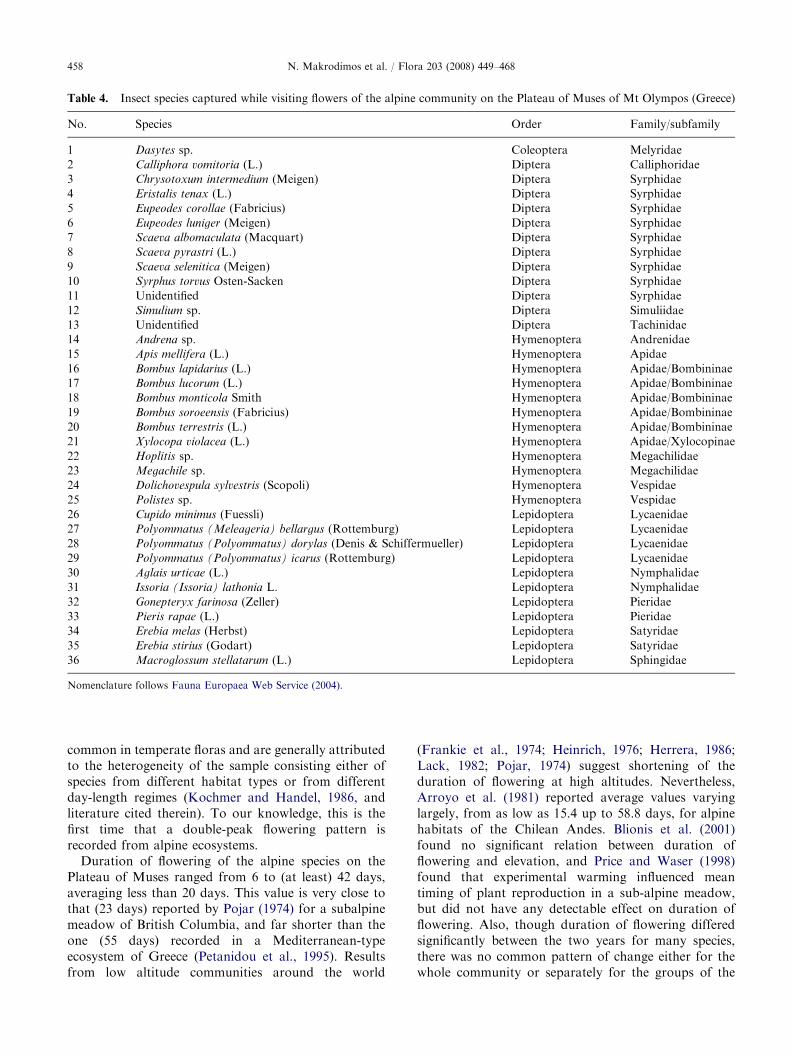
Composition and activity of flower visitors
A total of 36 insect species were identified (Table 4) asflower visitors of the alpine plants; of these, 12 speciesbelonged to Diptera, 12 to Hymenoptera, 11 toLepidoptera, and 1 to Coleoptera. A number of speci-mens that were sent to experts for identification werelost in the transfer process. In consequence, theinformation we provide on the composition of the
ARTICLE IN PRESS
1800
10
20
30
40
50
60
SeptemberAugustJuly
19931994
% S
peci
es in
blo
om
Calendar day
210 240 270
Fig. 1. Distribution of species in bloom for the alpine
community on the Plateau of Muses of Mt Olympos (Greece),
during 1993 and 1994.
RbGvPnScVgVb
AmSsRsTpVsPrLaLc
MaIs
AsTbAvEyEdAaGaVoSqCpEcPdAlDi
MvChLhLsDoSmSeCtDcSxAcHhTdEeCrArLiEsLpSvCoSgAyShOaHpSa
180
Calendar days
Plan
t spe
cies
19931994
190 200 210 220 230 240 250 260
Fig. 2. Flowering times of 57 alpine species occurring on the
Plateau of Muses of Mt Olympos (Greece), during 1993 and
1994. Species full names are given in Appendix A.
N. Makrodimos et al. / Flora 203 (2008) 449–468454
flower visiting entomofauna is not complete; allcaptured specimens of Lepidoptera, Bombus, andDiptera were identified, but species records for the
rest of Hymenoptera and Coleoptera are not full.Given this, we assume Hymenoptera species visitingflowers in the experimental plots to outnumber Dipteraspecies.
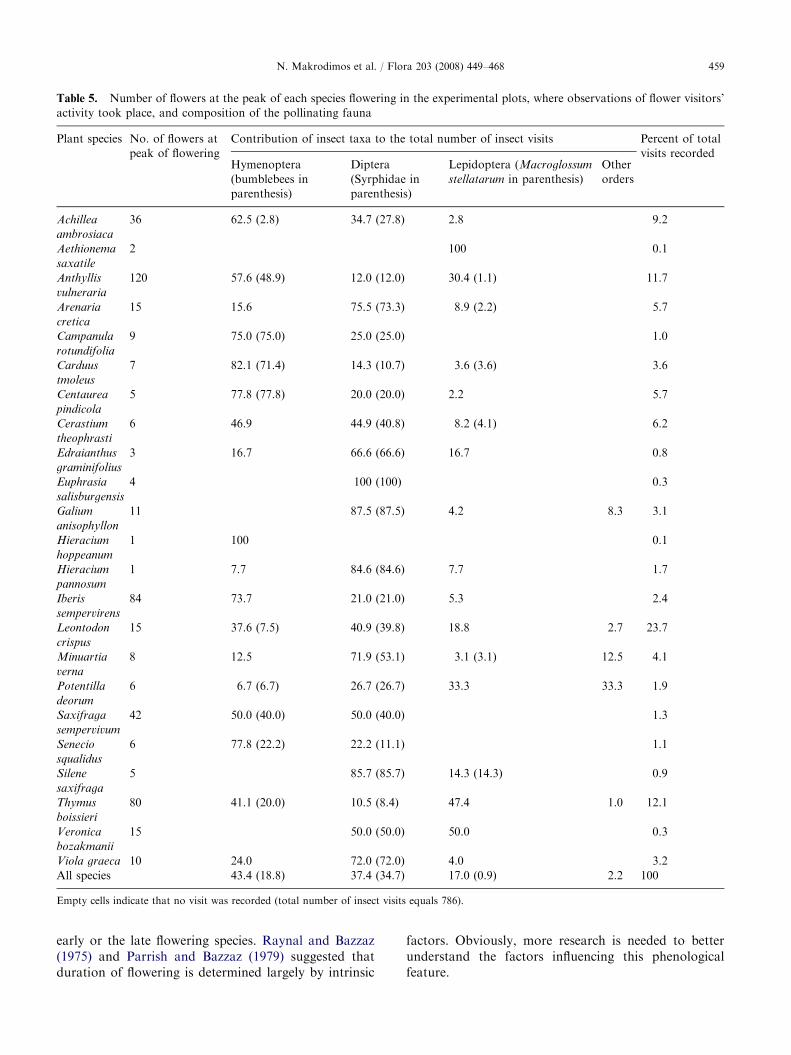
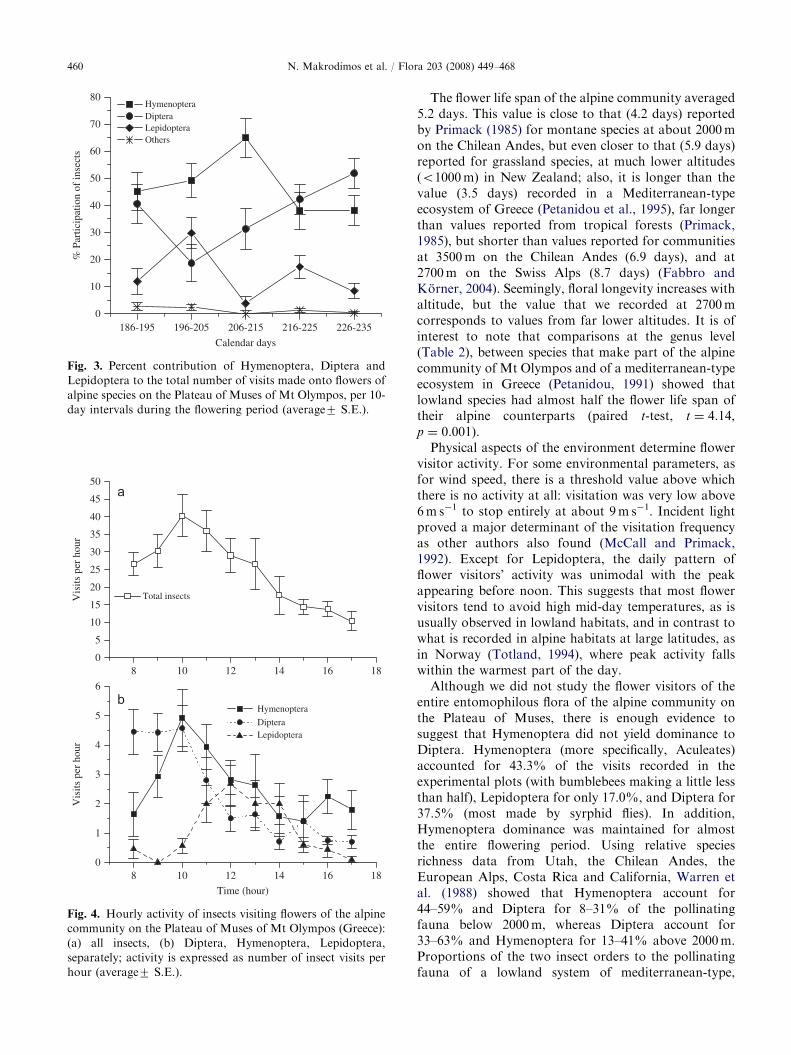
We recorded a total of 786 insect visits on to speciesflowers in the experimental plots. Hymenoptera mademost of them (43.4%), whereas Diptera accountedfor 37.4% and Lepidoptera for 17.0% of the visits(Table 5). Contribution of the different insect orders tothe flower visiting fauna varied with time (Fig. 3).Lepidoptera peaked early in the season, Diptera bothearly and late, whereas Hymenoptera peaked aroundmid-season. The daily activity of insects visitingflowers of the alpine community is shown in Fig. 4.Average insect activity peaked before noon (Fig. 4a),but the respective patterns of the three major ordersof flower visitors were considerably different (Fig. 4b).In particular, Diptera started to be very active earlyin the day; Hymenoptera maximized their activityaround 10:00 h, whereas for Lepidoptera, activity wasvery low early in the day taking maximum valuesaround noon.
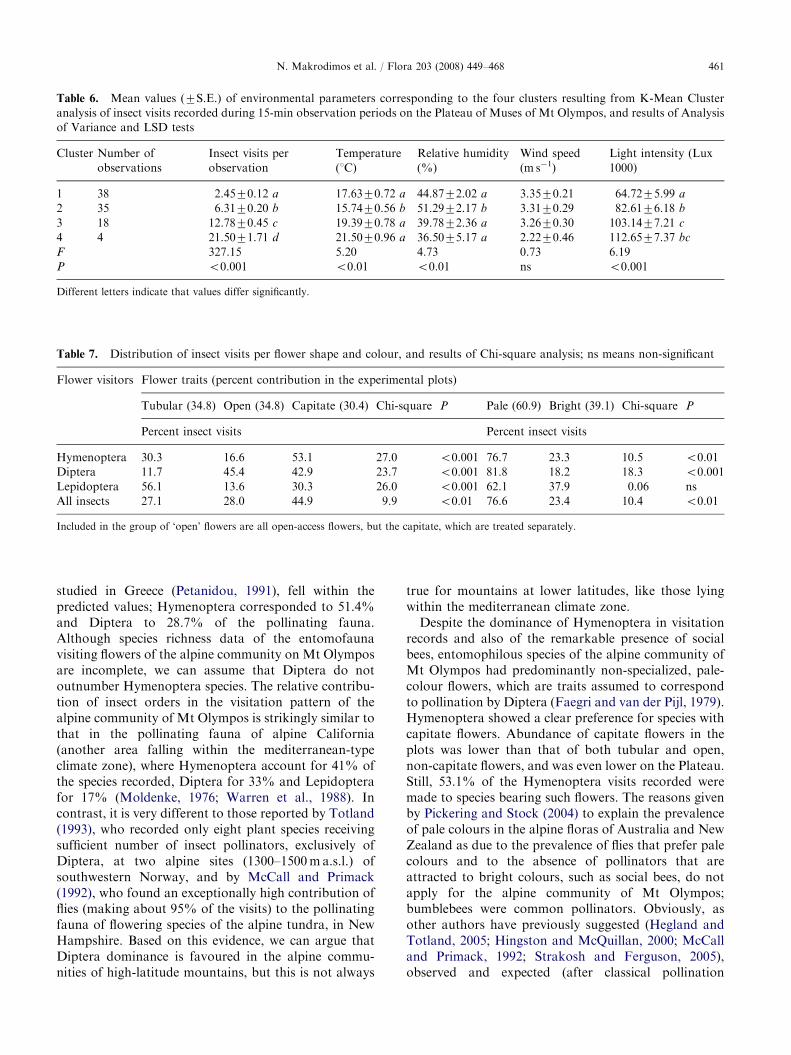
Using the K-Mean Cluster method, flower visits weregrouped into four clusters, after temperature, relativehumidity, wind velocity and light intensity. Meanvisitation rates and mean values of the environmentalparameters per cluster were further analyzed byANOVA and multiple LSD comparisons (Table 6). Ofthe three parameters (temperature, relative humidityand light intensity) that significantly differed amongclusters, only light intensity increased with visitationincrease.
Visitor attraction
A total of 23 plant species representing more thanone-third of the entire entomophilous flora of thePlateau occurred inside the experimental plots, whereobservations of flower visitors took place. Around 60%of the insect visits recorded corresponded to only fourspecies. Leontodon crispus attracted almost a quarter ofall visits (23.7%), followed by T. boissieri (12.1%), A.
vulneraria (11.7%) and Achillea ambrosiaca (9.2%). Tothe other end, less than 0.3% of the visits recorded weremade on flowers of Veronica bozakmanii, Aethionema
saxatile, Hieracium hoppeanum and Euphrasia salisbur-
gensis (Table 5).The number of flowers present per species, counted at
the species flowering peaks (Table 5), gives an estimateof the species floral display in the experimental plots,whereas distribution and abundance data allow inferringon how much widespread and common species are onthe Plateau. Floral display is highest for the species A.
vulneraria, Iberis sempervirens and T. boissieri andlowest for H. hoppeanum, H. pannosum, A. saxatile
ARTIC
LEIN
PRES
STable 2. Duration of flowering and flower life span (average7S.E.) of alpine plant species on the Plateau of Muses of Mt Olympos (Greece) and flower life span of co-generic
lowland species participating in a mediterranean-type ecosystem of the country, studied by Petanidou (1991)
Plant species Flowering
category
Duration of flowering (days) Flower life span (days) Mediterranean-type
ecosystem species
Flower life
span (days)1993 1994 Difference 1993 N 1994 N
Achillea ambrosiaca ef 12 13 1 8.170.7 10 8.470.4 10
Acinos alpinus ef 18 (21)
Aethionema saxatile ef 12 10 �2
Alyssum montanum ef (9) (27)
Anthyllis vulneraria ef 18 25 7 4.570.4 7 Anthyllis hermannae 3.8
Arenaria cretica lf 6 19 13 2.770.2 9 3.870.3 9
Armeria canescens lf 22 24 2 7.070.9 10
Asyneuma limonifolium 9
Astragalus depressus 2.670.2 8 Astragalus monspessulanus 1.0
Campanula oreadum 6
Campanula rotundifolia lf 18 25 7 3.570.1 30 3.570.3 11 Campanula drabifolia 3.0
Carduus tmoleus 27 31 4 6.770.6 10 Carduus pycnocephalus 2.7
Centaurea pindicola ef 6 25 19 6.870.7 9 Centaurea orphanidea 1.1
Cerastium theophrasti 36 37 1 2.070.1 10 4.070.4 14 Centaurea raphanina 3.0
Daphne oleoides 12 15 3 3.370.4 10 4.770.3 10
Dianthus integer 18 18 0 5.570.5 2
Doronicum columnae lf 18 18 0 7.770.6 9
Edraianthus graminifolius ef 12 19 7 4.070.1 10
Erigeron epiroticus lf 18 18 0
Euphorbia capitulata ef 12 21 9
Euphorbia cyparissias ef 18 9 �9
Euphrasia salisburgensis lf 15 25 10
Galium anisophyllon ef 15 13 �2
Gentiana verna ef (6) (15) 2.770.1 12
Hieracium hoppeanum 22 5.070.3 10
Hieracium pannosum 13
Iberis sempervirens ef 12 15 3 9.270.8 10
Lactuca intricata lf 6 18 12 1.870.2 10
Leontodon crispus 30 30 0
Linaria alpina (18) 24 6.870.4 6
Linaria peloponnesiaca 15 6.470.6 7
Linum hirsutum 12 9 �3 4.670.4 10 2.870.4 10 Linum strictum 4.0
Lotus corniculatus (18) 33 4.270.4 10
Minuartia verna ef 21 19 �2 4.070.4 14
Myosotis alpestris ef (21) 21 3.870.2 10 3.870.3 10
Orobanche amethystea. 15
Paronychia rechingeri (18) 27
Polygala nicaeensis ef (9) (15)
Potentilla deorum ef 12 25 13 7.170.7 9
N.Makrodim
oset
al./Flora
203(2008)449–468
455
ARTIC
LEIN
PRES
S
Table 2. (continued )
Plant species Flowering
category
Duration of flowering (days) Flower life span (days) Mediterranean-type
ecosystem species
Flower life
span (days)1993 1994 Difference 1993 N 1994 N
Ranunculus brevifolius (9) 5.170.2 10 Ranunculus spruneranus 4.2
Ranunculus sartorianus ef (12) 18 6.270.3 10
Saxifraga exarata 27 21 �6 13.470.7 10 7.070.7 8
Saxifraga glabella lf 12 15 3 7.370.3 8
Saxifraga scardica ef (9) (15)
Saxifraga sempervivum ef (12) 15
Scutellaria alpina 4.570.4 9
Sedum album lf 17 21 4
Sedum magellense 21 12 �9 2.570.1 10
Senecio squalidus 39 31 �8
Silene chromodonta 15
Silene saxifraga 6 7 1 5.270.2 5 3.070.4 7 Silene colorata 1.0
Silene vulgaris lf 26 15 �11 4.770.8 7 3.770.3 10
Taraxacum deorum lf 21 21 0 3.070.0 5 Taraxacum officinale 1.5
Thymus boissieri ef 18 31 13 11.271.1 9 Thymus capitatus 2.7
Trifolium pallescens (12) 15
Veronica bozakmanii ef (9) 15
Veronica orsiniana ef 20 12 �8
Viola graeca ef (9) 15 5.870.5 8
Viola striis-notata (29) (42) 5.270.4 5
Parentheses indicate that duration of flowering may be longer than the values given, as the respective species were already in bloom when we started taking records; N represents the number of
flowers studied, ef signifies consistently early flowering species and lf signifies consistently late flowering species (see text for definitions).
N.Makrodim
oset
al./Flora
203(2008)449–468
456

ARTICLE IN PRESS
Table 3. Distribution of plant species on the Plateau of
Muses and in the experimental plots, where observations of
flower visitors’ activity took place, into different types of
flower colour and shape
Plateau of Muses Experimental plots
Flower
shape
Proportion (%) Flower
shape
Proportion (%)
Flower
colour
Flower colour
Pale Bright Pale Bright
Open 49.2 66.7 33.3 Open 34.8 75.0 25.0
Capitate 18.0 81.8 18.2 Capitate 30.4 85.7 14.3
Tubular 32.8 50.0 50.0 Tubular 34.8 25.0 75.0
Any type 100.0 63.9 36.1 Any type 100.0 60.9 39.1
Given also are the corresponding percentages of pale and bright
coloured flowers per shape type. Included in the group of ‘open’
flowers are all open-access flowers but the capitate, which are
presented separately.
N. Makrodimos et al. / Flora 203 (2008) 449–468 457
and Edraianthus graminifolius. Highly visited species areusually, but not always, those with abundant flowers.For instance, the most preferred species, L. crispus, hadonly intermediate floral display in the experimental plotsat the peak of anthesis and intermediate abundance onthe Plateau.
The majority of flowers visited by Hymenoptera(53.1%) were capitate, whereas those by Lepidopterawere tubular (Table 7). Almost 90% of Diptera visitswere made to open-access flowers, equally to capitateand non-capitate. Both Diptera and Hymenopteravisitors preferred pale-flower species. Given the discre-pancy between expected and actual preferences and inorder to identify the features that best explain therelative contribution of Diptera, Hymenoptera andLepidoptera to the visitation of alpine species, weapplied RTA. The variables examined per species weredistribution and abundance, duration of flowering, peakof flowering, date of maximum flower visitor activity,flower colour and shape and floral display. V. bozakma-
nii, A. saxatile, H. hoppeanum and E. salisburgensis, forwhich the total number of insect visits recorded was lessthan three, were excluded from the analysis. Thevariance explained by RTA was satisfactory forHymenoptera and Diptera (39.6% and 67.0%, respec-tively), but low for Lepidoptera (21.5%). For bothHymenoptera and Diptera, duration of floweringfollowed by floral abundance were the most explanatoryvariables (Fig. 5a and b), but with opposite trends.Duration of flowering longer than 22 days wasassociated with the highest proportion of Hymenoptera(55.8%) and the lowest of Diptera (24.1%). Forduration of flowering shorter than 22 days, when flower
abundance was less than 10.5 flowers, proportion ofHymenoptera minimized (12.2%), whereas proportionof Diptera maximized (76.2%). For Lepidoptera, themain explanatory variable was corolla colour. However,due to the low variance explained, Lepidoptera were notfurther analysed. We also tested for linear relationshipsbetween the relative visitation of either Hymenoptera orDiptera and the length of the flowering period. Theywere both significant: %Hy ¼ 3.41+1.72*DF, r ¼
0.48, p ¼ 0.038; %Di ¼ 93.01�2.21*DF, r ¼ �0.65, p ¼
0.002, where %Hy corresponds to percent visitation ofHymenoptera, %Di to percent visitation of Diptera, andDF to flowering period length.
Discussion
The 61 angiosperm species that we found on thePlateau of Muses make 46% of the species displaying abiotic pollination syndrome and occurring above 2400mon Mt Olympos (Strid, 1980). This means that the studysite represents fairly well the entomophilous alpine floraof the mountain. Early flowering is the favouritestrategy of the alpine community on Mt Olympos;consistently early flowering species were twice as manythose consistently flowering late. Limited time availablefor seed maturation seems to be the reason for earlyflowering selection in short-season environments (Tot-land, 1993).
Flowering varied between years. Shifts in start offlowering were large enough to move some species fromthe group of early flowering to the group of lateflowering ones, and vice versa. Trifolium pallescens andSedum magellense were the strongest shifters. Never-theless, at least 60% of the species remained in the samegroup in both years. Shifts in flowering time seem torespond to the yearly variation of climatic parameters.While several of the consistently early flowering speciesflowered earlier, all consistently late flowering speciesdelayed flowering in 1994. Many authors reportedearlier flowering of alpine plants with simulated climatewarming treatments (Alatalo and Totland, 1997; Henryand Molau, 1997; Price and Waser, 1998; Stenstrom etal., 1997; Suzuki and Kudo, 1997), Blionis et al. (2001)found delays of the onset of flowering of Campanula
taxa on Mt Olympos by 2–3 days for every 100m ofelevation increase (and concomitant temperature de-crease), and Petanidou et al. (1995) found that earlierflowering in a Mediterranean community is associatedwith warmer and drier conditions. Given these, delayedflowering of all late flowering species can be consideredas a consequence of the climatic character of July, whichwas colder and by far wetter in 1994 compared to 1993.
Flowering of the alpine community on Mt Olymposwas characterized by bimodality. Bimodal patterns are
ARTICLE IN PRESS
Table 4. Insect species captured while visiting flowers of the alpine community on the Plateau of Muses of Mt Olympos (Greece)
No. Species Order Family/subfamily
1 Dasytes sp. Coleoptera Melyridae
2 Calliphora vomitoria (L.) Diptera Calliphoridae
3 Chrysotoxum intermedium (Meigen) Diptera Syrphidae
4 Eristalis tenax (L.) Diptera Syrphidae
5 Eupeodes corollae (Fabricius) Diptera Syrphidae
6 Eupeodes luniger (Meigen) Diptera Syrphidae
7 Scaeva albomaculata (Macquart) Diptera Syrphidae
8 Scaeva pyrastri (L.) Diptera Syrphidae
9 Scaeva selenitica (Meigen) Diptera Syrphidae
10 Syrphus torvus Osten-Sacken Diptera Syrphidae
11 Unidentified Diptera Syrphidae
12 Simulium sp. Diptera Simuliidae
13 Unidentified Diptera Tachinidae
14 Andrena sp. Hymenoptera Andrenidae
15 Apis mellifera (L.) Hymenoptera Apidae
16 Bombus lapidarius (L.) Hymenoptera Apidae/Bombininae
17 Bombus lucorum (L.) Hymenoptera Apidae/Bombininae
18 Bombus monticola Smith Hymenoptera Apidae/Bombininae
19 Bombus soroeensis (Fabricius) Hymenoptera Apidae/Bombininae
20 Bombus terrestris (L.) Hymenoptera Apidae/Bombininae
21 Xylocopa violacea (L.) Hymenoptera Apidae/Xylocopinae
22 Hoplitis sp. Hymenoptera Megachilidae
23 Megachile sp. Hymenoptera Megachilidae
24 Dolichovespula sylvestris (Scopoli) Hymenoptera Vespidae
25 Polistes sp. Hymenoptera Vespidae
26 Cupido minimus (Fuessli) Lepidoptera Lycaenidae
27 Polyommatus (Meleageria) bellargus (Rottemburg) Lepidoptera Lycaenidae
28 Polyommatus (Polyommatus) dorylas (Denis & Schiffermueller) Lepidoptera Lycaenidae
29 Polyommatus (Polyommatus) icarus (Rottemburg) Lepidoptera Lycaenidae
30 Aglais urticae (L.) Lepidoptera Nymphalidae
31 Issoria (Issoria) lathonia L. Lepidoptera Nymphalidae
32 Gonepteryx farinosa (Zeller) Lepidoptera Pieridae
33 Pieris rapae (L.) Lepidoptera Pieridae
34 Erebia melas (Herbst) Lepidoptera Satyridae
35 Erebia stirius (Godart) Lepidoptera Satyridae
36 Macroglossum stellatarum (L.) Lepidoptera Sphingidae
Nomenclature follows Fauna Europaea Web Service (2004).
N. Makrodimos et al. / Flora 203 (2008) 449–468458
common in temperate floras and are generally attributedto the heterogeneity of the sample consisting either ofspecies from different habitat types or from differentday-length regimes (Kochmer and Handel, 1986, andliterature cited therein). To our knowledge, this is thefirst time that a double-peak flowering pattern isrecorded from alpine ecosystems.
Duration of flowering of the alpine species on thePlateau of Muses ranged from 6 to (at least) 42 days,averaging less than 20 days. This value is very close tothat (23 days) reported by Pojar (1974) for a subalpinemeadow of British Columbia, and far shorter than theone (55 days) recorded in a Mediterranean-typeecosystem of Greece (Petanidou et al., 1995). Resultsfrom low altitude communities around the world
(Frankie et al., 1974; Heinrich, 1976; Herrera, 1986;Lack, 1982; Pojar, 1974) suggest shortening of theduration of flowering at high altitudes. Nevertheless,Arroyo et al. (1981) reported average values varyinglargely, from as low as 15.4 up to 58.8 days, for alpinehabitats of the Chilean Andes. Blionis et al. (2001)found no significant relation between duration offlowering and elevation, and Price and Waser (1998)found that experimental warming influenced meantiming of plant reproduction in a sub-alpine meadow,but did not have any detectable effect on duration offlowering. Also, though duration of flowering differedsignificantly between the two years for many species,there was no common pattern of change either for thewhole community or separately for the groups of the
ARTICLE IN PRESS
Table 5. Number of flowers at the peak of each species flowering in the experimental plots, where observations of flower visitors’
activity took place, and composition of the pollinating fauna
Plant species No. of flowers at
peak of flowering
Contribution of insect taxa to the total number of insect visits Percent of total
visits recordedHymenoptera
(bumblebees in
parenthesis)
Diptera
(Syrphidae in
parenthesis)
Lepidoptera (Macroglossum
stellatarum in parenthesis)
Other
orders
Achillea
ambrosiaca
36 62.5 (2.8) 34.7 (27.8) 2.8 9.2
Aethionema
saxatile
2 100 0.1
Anthyllis
vulneraria
120 57.6 (48.9) 12.0 (12.0) 30.4 (1.1) 11.7
Arenaria
cretica
15 15.6 75.5 (73.3) 8.9 (2.2) 5.7
Campanula
rotundifolia
9 75.0 (75.0) 25.0 (25.0) 1.0
Carduus
tmoleus
7 82.1 (71.4) 14.3 (10.7) 3.6 (3.6) 3.6
Centaurea
pindicola
5 77.8 (77.8) 20.0 (20.0) 2.2 5.7
Cerastium
theophrasti
6 46.9 44.9 (40.8) 8.2 (4.1) 6.2
Edraianthus
graminifolius
3 16.7 66.6 (66.6) 16.7 0.8
Euphrasia
salisburgensis
4 100 (100) 0.3
Galium
anisophyllon
11 87.5 (87.5) 4.2 8.3 3.1
Hieracium
hoppeanum
1 100 0.1
Hieracium
pannosum
1 7.7 84.6 (84.6) 7.7 1.7
Iberis
sempervirens
84 73.7 21.0 (21.0) 5.3 2.4
Leontodon
crispus
15 37.6 (7.5) 40.9 (39.8) 18.8 2.7 23.7
Minuartia
verna
8 12.5 71.9 (53.1) 3.1 (3.1) 12.5 4.1
Potentilla
deorum
6 6.7 (6.7) 26.7 (26.7) 33.3 33.3 1.9
Saxifraga
sempervivum
42 50.0 (40.0) 50.0 (40.0) 1.3
Senecio
squalidus
6 77.8 (22.2) 22.2 (11.1) 1.1
Silene
saxifraga
5 85.7 (85.7) 14.3 (14.3) 0.9
Thymus
boissieri
80 41.1 (20.0) 10.5 (8.4) 47.4 1.0 12.1
Veronica
bozakmanii
15 50.0 (50.0) 50.0 0.3
Viola graeca 10 24.0 72.0 (72.0) 4.0 3.2
All species 43.4 (18.8) 37.4 (34.7) 17.0 (0.9) 2.2 100
Empty cells indicate that no visit was recorded (total number of insect visits equals 786).
N. Makrodimos et al. / Flora 203 (2008) 449–468 459
early or the late flowering species. Raynal and Bazzaz(1975) and Parrish and Bazzaz (1979) suggested thatduration of flowering is determined largely by intrinsic
factors. Obviously, more research is needed to betterunderstand the factors influencing this phenologicalfeature.
ARTICLE IN PRESS
186-1950
10
20
30
40
50
60
70
80HymenopteraDipteraLepidopteraOthers
% P
artic
ipat
ion
of in
sect
s
Calendar days
196-205 206-215 216-225 226-235
Fig. 3. Percent contribution of Hymenoptera, Diptera and
Lepidoptera to the total number of visits made onto flowers of
alpine species on the Plateau of Muses of Mt Olympos, per 10-
day intervals during the flowering period (average7 S.E.).
0
5
10
15
20
25
30
35
40
45
50
Total insectsVis
its p
er h
our
80
1
2
3
4
5
6
Hymenoptera
DipteraLepidoptera
Vis
its p
er h
our
Time (hour)
10 12 14 16 18
8 10 12 14 16 18
Fig. 4. Hourly activity of insects visiting flowers of the alpine
community on the Plateau of Muses of Mt Olympos (Greece):
(a) all insects, (b) Diptera, Hymenoptera, Lepidoptera,
separately; activity is expressed as number of insect visits per
hour (average7 S.E.).
N. Makrodimos et al. / Flora 203 (2008) 449–468460
The flower life span of the alpine community averaged5.2 days. This value is close to that (4.2 days) reportedby Primack (1985) for montane species at about 2000mon the Chilean Andes, but even closer to that (5.9 days)reported for grassland species, at much lower altitudes(o1000m) in New Zealand; also, it is longer than thevalue (3.5 days) recorded in a Mediterranean-typeecosystem of Greece (Petanidou et al., 1995), far longerthan values reported from tropical forests (Primack,1985), but shorter than values reported for communitiesat 3500m on the Chilean Andes (6.9 days), and at2700m on the Swiss Alps (8.7 days) (Fabbro andKorner, 2004). Seemingly, floral longevity increases withaltitude, but the value that we recorded at 2700mcorresponds to values from far lower altitudes. It is ofinterest to note that comparisons at the genus level(Table 2), between species that make part of the alpinecommunity of Mt Olympos and of a mediterranean-typeecosystem in Greece (Petanidou, 1991) showed thatlowland species had almost half the flower life span oftheir alpine counterparts (paired t-test, t ¼ 4.14,p ¼ 0.001).
Physical aspects of the environment determine flowervisitor activity. For some environmental parameters, asfor wind speed, there is a threshold value above whichthere is no activity at all: visitation was very low above6m s�1 to stop entirely at about 9m s�1. Incident lightproved a major determinant of the visitation frequencyas other authors also found (McCall and Primack,1992). Except for Lepidoptera, the daily pattern offlower visitors’ activity was unimodal with the peakappearing before noon. This suggests that most flowervisitors tend to avoid high mid-day temperatures, as isusually observed in lowland habitats, and in contrast towhat is recorded in alpine habitats at large latitudes, asin Norway (Totland, 1994), where peak activity fallswithin the warmest part of the day.
Although we did not study the flower visitors of theentire entomophilous flora of the alpine community onthe Plateau of Muses, there is enough evidence tosuggest that Hymenoptera did not yield dominance toDiptera. Hymenoptera (more specifically, Aculeates)accounted for 43.3% of the visits recorded in theexperimental plots (with bumblebees making a little lessthan half), Lepidoptera for only 17.0%, and Diptera for37.5% (most made by syrphid flies). In addition,Hymenoptera dominance was maintained for almostthe entire flowering period. Using relative speciesrichness data from Utah, the Chilean Andes, theEuropean Alps, Costa Rica and California, Warren etal. (1988) showed that Hymenoptera account for44–59% and Diptera for 8–31% of the pollinatingfauna below 2000m, whereas Diptera account for33–63% and Hymenoptera for 13–41% above 2000m.Proportions of the two insect orders to the pollinatingfauna of a lowland system of mediterranean-type,
ARTICLE IN PRESS
Table 6. Mean values (7S.E.) of environmental parameters corresponding to the four clusters resulting from K-Mean Cluster
analysis of insect visits recorded during 15-min observation periods on the Plateau of Muses of Mt Olympos, and results of Analysis
of Variance and LSD tests
Cluster Number of
observations
Insect visits per
observation
Temperature
(1C)
Relative humidity
(%)
Wind speed
(m s�1)
Light intensity (Lux
1000)
1 38 2.4570.12 a 17.6370.72 a 44.8772.02 a 3.3570.21 64.7275.99 a
2 35 6.3170.20 b 15.7470.56 b 51.2972.17 b 3.3170.29 82.6176.18 b
3 18 12.7870.45 c 19.3970.78 a 39.7872.36 a 3.2670.30 103.1477.21 c
4 4 21.5071.71 d 21.5070.96 a 36.5075.17 a 2.2270.46 112.6577.37 bc
F 327.15 5.20 4.73 0.73 6.19
P o0.001 o0.01 o0.01 ns o0.001
Different letters indicate that values differ significantly.
Table 7. Distribution of insect visits per flower shape and colour, and results of Chi-square analysis; ns means non-significant
Flower visitors Flower traits (percent contribution in the experimental plots)
Tubular (34.8) Open (34.8) Capitate (30.4) Chi-square P Pale (60.9) Bright (39.1) Chi-square P
Percent insect visits Percent insect visits
Hymenoptera 30.3 16.6 53.1 27.0 o0.001 76.7 23.3 10.5 o0.01
Diptera 11.7 45.4 42.9 23.7 o0.001 81.8 18.2 18.3 o0.001
Lepidoptera 56.1 13.6 30.3 26.0 o0.001 62.1 37.9 0.06 ns
All insects 27.1 28.0 44.9 9.9 o0.01 76.6 23.4 10.4 o0.01
Included in the group of ‘open’ flowers are all open-access flowers, but the capitate, which are treated separately.
N. Makrodimos et al. / Flora 203 (2008) 449–468 461
studied in Greece (Petanidou, 1991), fell within thepredicted values; Hymenoptera corresponded to 51.4%and Diptera to 28.7% of the pollinating fauna.Although species richness data of the entomofaunavisiting flowers of the alpine community on Mt Olymposare incomplete, we can assume that Diptera do notoutnumber Hymenoptera species. The relative contribu-tion of insect orders in the visitation pattern of thealpine community of Mt Olympos is strikingly similar tothat in the pollinating fauna of alpine California(another area falling within the mediterranean-typeclimate zone), where Hymenoptera account for 41% ofthe species recorded, Diptera for 33% and Lepidopterafor 17% (Moldenke, 1976; Warren et al., 1988). Incontrast, it is very different to those reported by Totland(1993), who recorded only eight plant species receivingsufficient number of insect pollinators, exclusively ofDiptera, at two alpine sites (1300–1500ma.s.l.) ofsouthwestern Norway, and by McCall and Primack(1992), who found an exceptionally high contribution offlies (making about 95% of the visits) to the pollinatingfauna of flowering species of the alpine tundra, in NewHampshire. Based on this evidence, we can argue thatDiptera dominance is favoured in the alpine commu-nities of high-latitude mountains, but this is not always
true for mountains at lower latitudes, like those lyingwithin the mediterranean climate zone.
Despite the dominance of Hymenoptera in visitationrecords and also of the remarkable presence of socialbees, entomophilous species of the alpine community ofMt Olympos had predominantly non-specialized, pale-colour flowers, which are traits assumed to correspondto pollination by Diptera (Faegri and van der Pijl, 1979).Hymenoptera showed a clear preference for species withcapitate flowers. Abundance of capitate flowers in theplots was lower than that of both tubular and open,non-capitate flowers, and was even lower on the Plateau.Still, 53.1% of the Hymenoptera visits recorded weremade to species bearing such flowers. The reasons givenby Pickering and Stock (2004) to explain the prevalenceof pale colours in the alpine floras of Australia and NewZealand as due to the prevalence of flies that prefer palecolours and to the absence of pollinators that areattracted to bright colours, such as social bees, do notapply for the alpine community of Mt Olympos;bumblebees were common pollinators. Obviously, asother authors have previously suggested (Hegland andTotland, 2005; Hingston and McQuillan, 2000; McCalland Primack, 1992; Strakosh and Ferguson, 2005),observed and expected (after classical pollination

ARTICLE IN PRESS
Duration of flowering: <22 days
Number of flowers: <10.5
24.05
53.7776.17
55.84
40.3512.17
Number of flowers: <10.5
Duration of flowering: <22 days
Fig. 5. Regression Tree Analysis for (a) Hymenoptera and (b)
Diptera visiting flowers of the alpine community on the
Plateau of Muses of Mt Olympos (Greece).
N. Makrodimos et al. / Flora 203 (2008) 449–468462
syndromes) flower visitors do not always match. There-fore, different approaches are needed to explain deviat-ing behaviours.
RTA showed that visitation of Hymenoptera andDiptera is determined by duration of flowering andflower abundance. Whereas proportionately more Dip-tera visit species with short durations of flowering andfew flowers present in the field, Hymenoptera areassociated with plant species having longer durationsof flowering and producing more flowers. These findingsare consistent with pollination theory: the relativeabundance of a species and also the duration of itsflowering period are two factors of great importance forthe development of insect fidelity (Faegri and van derPijl, 1979). These findings also support the concept of‘ecological pollination syndromes’, developed afterreports of poor support of data to the expectationsderived from the classical pollination syndromes.Describing the foraging behaviour of two pollinatortaxa (bumblebees and syrphid flies) of alpine plants,Yumoto (1986) argued that there exist ‘ecologicalpollination syndromes’, which are associated with planttraits related to the flowering behaviour of individualplants, the spatial distribution of plant populations, andthe species flowering phenology. Our results showing
that duration of flowering and flower abundance aremore important than morphological features in explain-ing flower visitation provide additional evidence of thevalue of ‘ecological pollination syndromes’.
Our initial hypotheses regarding selection for earlyflowering and long flower life span, and also sensitivityof onset of flowering of the alpine flora to yearly climaticvariability were supported by the experimental data. Incontrast, we did not find the flower-morphology traits tomatch the flower-visitors traits, as expected after theclassical pollination syndromes; ecological featuresseemed to better explain visitation patterns. We mustalso remark that in a number of cases, the phenologicaland visitation patterns of the alpine community of MtOlympos deviated from those observed in other alpineenvironments. Such deviations were manifested (i) in thedistribution of species in bloom, characterized by twopeaks, as is usually the case in temperate environments,(ii) in floral longevity, which is shorter than that of otheralpine communities and (iii) in the relative contributionof Hymenoptera and Diptera in flower visitation, withHymenoptera making most of the visits. Given thesimilarities with alpine communities of other mountainsfalling within mediterranean-climate zones, a Mediter-ranean influence is likely to exist even at high altitudesmoderating the climatic character and lessening ex-tremes.
Acknowledgements
This work was funded by the Secretariat of Researchand Technology of Greece (Projects 91ED 906 and 97EL-6). Insect specimens were identified by P.A. Langleyand H.A. Dawah (University of Wales, Cardiff), J.C.Deeming (National Museum of Wales, Cardiff), W.N.Ellis (University of Amsterdam) and E. Obrecht(University of Bern). The Forest Service of PieriaPrefecture facilitated our work in Mt Olympos NationalPark. A. Prokovas and his family, tenants of S.E.O.Refuge during the time of our study, offered theirhospitality and help.
Appendix A
Plant taxa (species and subspecies) with a bioticpollination syndrome on the Plateau of Muses of MtOlympos (Greece), ranked by alphabetical order offamily, genus and species. Given are their geographicdistribution (Strid 1980), their flower colour and shape,and the traits studied per species. P ¼ pale colouredflowers (including white, whitish, and yellow),B ¼ bright coloured flowers (including violet-blue,purple, red and pink).
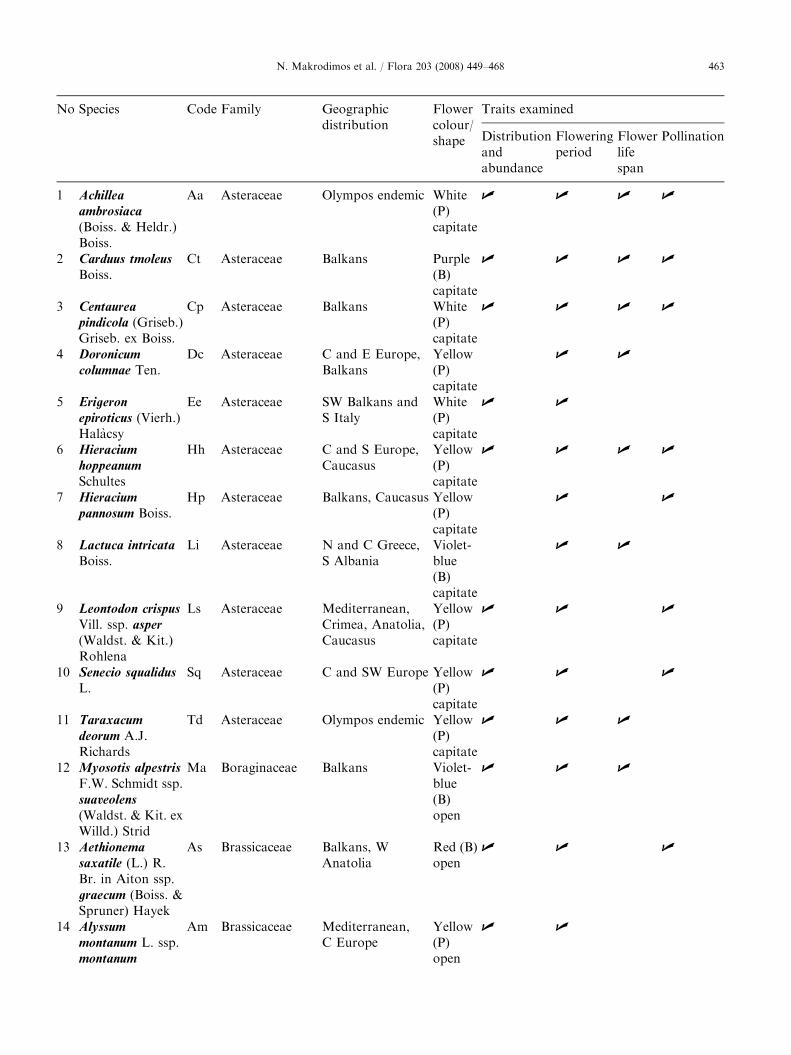
ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468 463
No
Species Code Family GeographicdistributionFlowercolour/shape
Traits examined
Distributionandabundance
Fp
loweringeriodFlowerlifespan
Pollination
1
Achilleaambrosiaca(Boiss. & Heldr.)Boiss.Aa
Asteraceae Olympos endemic White(P)capitate| |
| |2
Carduus tmoleusBoiss.Ct
Asteraceae Balkans Purple(B)capitate| |
| |3
Centaureapindicola (Griseb.)Griseb. ex Boiss.Cp
Asteraceae Balkans White(P)capitate| |
| |4
Doronicumcolumnae Ten.Dc
Asteraceae C and E Europe,BalkansYellow(P)capitate
|
|5
Erigeronepiroticus (Vierh.)HalacsyEe
Asteraceae SW Balkans andS ItalyWhite(P)capitate
| |
6
HieraciumhoppeanumSchultesHh
Asteraceae C and S Europe,CaucasusYellow(P)capitate
| |
| |7
Hieraciumpannosum Boiss.Hp
Asteraceae Balkans, Caucasus Yellow(P)capitate|
|8
Lactuca intricataBoiss.Li
Asteraceae N and C Greece,S AlbaniaViolet-blue(B)capitate
|
|9
Leontodon crispusVill. ssp. asper(Waldst. & Kit.)RohlenaLs
Asteraceae Mediterranean,Crimea, Anatolia,CaucasusYellow(P)capitate
| |
|10
Senecio squalidusL.Sq
Asteraceae C and SW Europe Yellow(P)capitate| |
|11
Taraxacumdeorum A.J.RichardsTd
Asteraceae Olympos endemic Yellow(P)capitate| |
|12
Myosotis alpestrisF.W. Schmidt ssp.suaveolens(Waldst. & Kit. exWilld.) StridMa
Boraginaceae Balkans Violet-blue(B)open| |
|13
Aethionemasaxatile (L.) R.Br. in Aiton ssp.graecum (Boiss. &Spruner) HayekAs
Brassicaceae Balkans, WAnatoliaRed (B)open
| |
|14
Alyssummontanum L. ssp.montanumAm
Brassicaceae Mediterranean,C EuropeYellow(P)open
| |
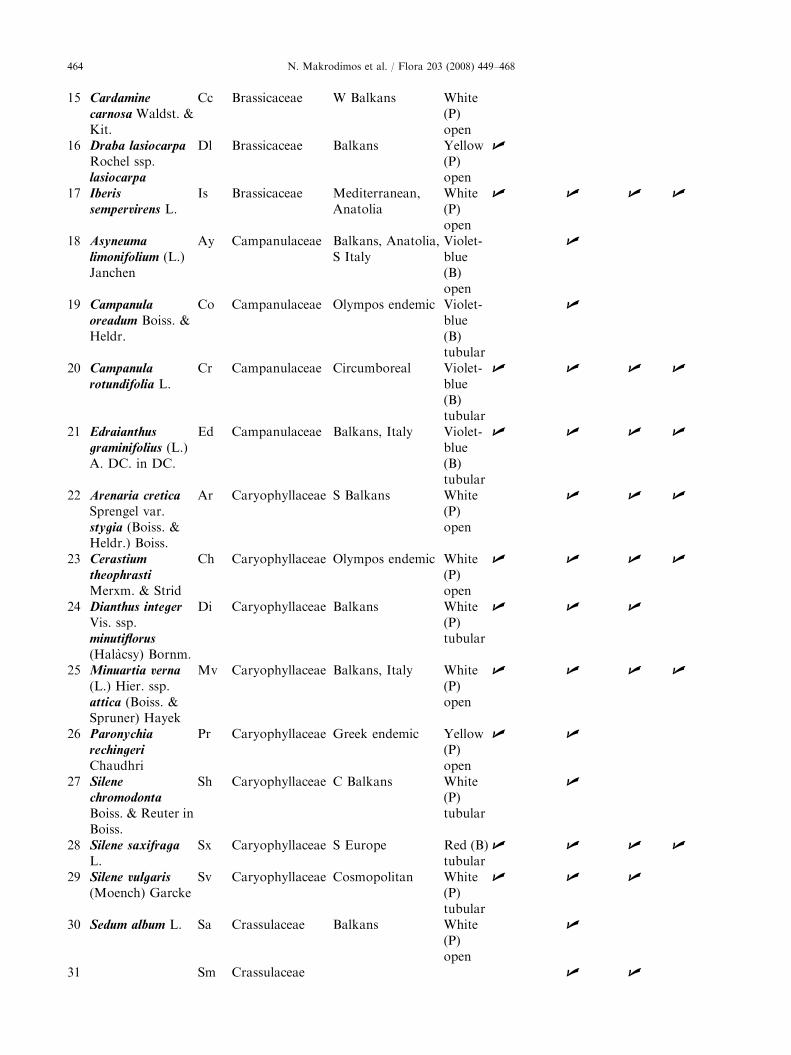
ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468464
15
Cardaminecarnosa Waldst. &Kit.C
c Brassicaceae W Balkans White(P)open16
Draba lasiocarpaRochel ssp.lasiocarpaD
l Brassicaceae Balkans Yellow(P)open|
17
Iberissempervirens L.I
s Brassicaceae Mediterranean,AnatoliaWhite(P)open
| |
| |18
Asyneumalimonifolium (L.)JanchenA
y Campanulaceae Balkans, Anatolia,S ItalyViolet-blue(B)open
|
19
Campanulaoreadum Boiss. &Heldr.C
o Campanulaceae Olympos endemic Violet-blue(B)tubular|
20
Campanularotundifolia L.C
r Campanulaceae Circumboreal Violet-blue(B)tubular| |
| |21
Edraianthusgraminifolius (L.)A. DC. in DC.E
d Campanulaceae Balkans, Italy Violet-blue(B)tubular| |
| |22
Arenaria creticaSprengel var.stygia (Boiss. &Heldr.) Boiss.A
r Caryophyllaceae S Balkans White(P)open|
| |23
CerastiumtheophrastiMerxm. & StridC
h Caryophyllaceae Olympos endemic White(P)open| |
| |24
Dianthus integerVis. ssp.minutiflorus(Halacsy) Bornm.D
i Caryophyllaceae Balkans White(P)tubular| |
|25
Minuartia verna(L.) Hier. ssp.attica (Boiss. &Spruner) HayekM
v Caryophyllaceae Balkans, Italy White(P)open| |
| |26
ParonychiarechingeriChaudhriP
r Caryophyllaceae Greek endemic Yellow(P)open| |
27
SilenechromodontaBoiss. & Reuter inBoiss.S
h Caryophyllaceae C Balkans White(P)tubular|
28
Silene saxifragaL.S
x Caryophyllaceae S Europe Red (B)tubular| |
| |29
Silene vulgaris(Moench) GarckeS
v Caryophyllaceae Cosmopolitan White(P)tubular| |
|30
Sedum album L. S a Crassulaceae Balkans White(P)open|
31
Sm Crassulaceae | |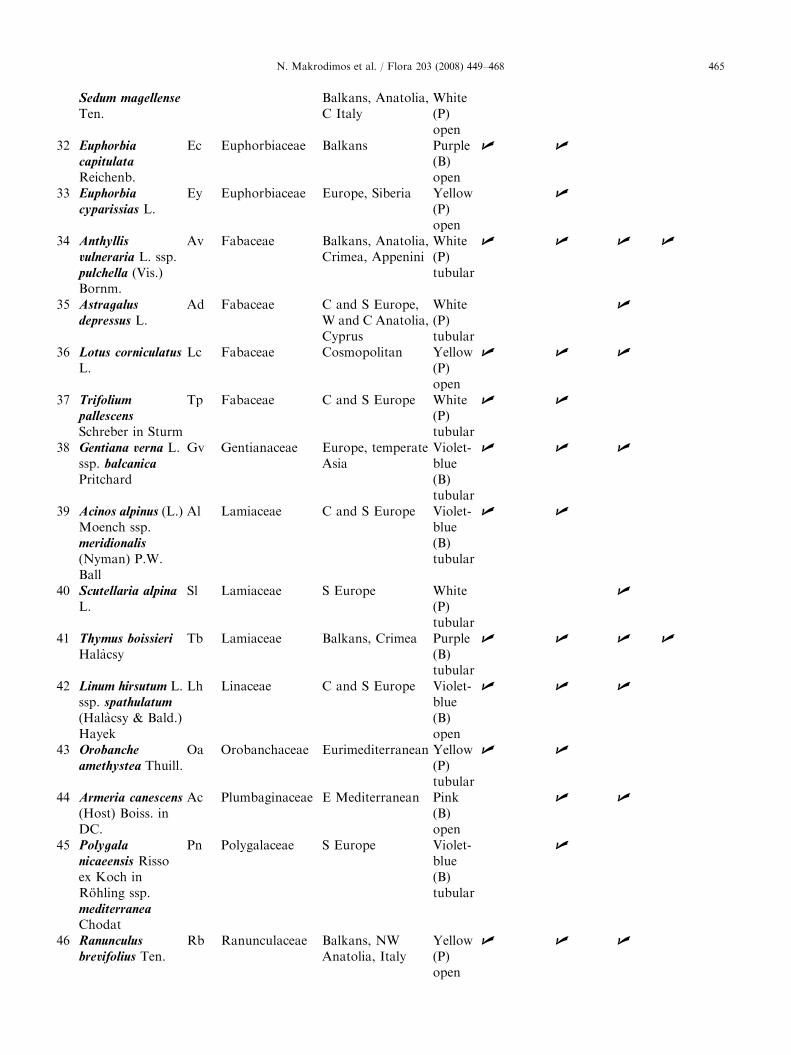
ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468 465
Sedum magellenseTen.
Balkans, Anatolia,C Italy
White(P)open
32
EuphorbiacapitulataReichenb.Ec
Euphorbiaceae Balkans Purple(B)open| |
33
Euphorbiacyparissias L.Ey
Euphorbiaceae Europe, Siberia Yellow(P)open|
34
Anthyllisvulneraria L. ssp.pulchella (Vis.)Bornm.Av
Fabaceae Balkans, Anatolia,Crimea, AppeniniWhite(P)tubular
| |
| |35
Astragalusdepressus L.Ad
Fabaceae C and S Europe,W and C Anatolia,CyprusWhite(P)tubular
|
36
Lotus corniculatusL.Lc
Fabaceae Cosmopolitan Yellow(P)open| |
|37
TrifoliumpallescensSchreber in SturmTp
Fabaceae C and S Europe White(P)tubular| |
38
Gentiana verna L.ssp. balcanicaPritchardGv
Gentianaceae Europe, temperateAsiaViolet-blue(B)tubular
| |
|39
Acinos alpinus (L.)Moench ssp.meridionalis(Nyman) P.W.BallAl
Lamiaceae C and S Europe Violet-blue(B)tubular| |
40
Scutellaria alpinaL.Sl
Lamiaceae S Europe White(P)tubular|
41
Thymus boissieriHalacsyTb
Lamiaceae Balkans, Crimea Purple(B)tubular| |
| |42
Linum hirsutum L.ssp. spathulatum(Halacsy & Bald.)HayekLh
Linaceae C and S Europe Violet-blue(B)open| |
|43
Orobancheamethystea Thuill.Oa
Orobanchaceae Eurimediterranean Yellow(P)tubular| |
44
Armeria canescens(Host) Boiss. inDC.Ac
Plumbaginaceae E Mediterranean Pink(B)open|
|45
Polygalanicaeensis Rissoex Koch inRohling ssp.mediterraneaChodatPn
Polygalaceae S Europe Violet-blue(B)tubular|
46
Ranunculusbrevifolius Ten.Rb
Ranunculaceae Balkans, NWAnatolia, ItalyYellow(P)open
| |
|
ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468466
47
Ranunculussartorianus Boiss.& Heldr. in Boiss.R
s Ranunculaceae Balkans, Italy Yellow(P)open| |
|48
Potentilla deorumBoiss. & Heldr. inBoiss.P
d Rosaceae Olympos endemic Yellow(P)open| |
| |49
Galiumanisophyllon Vill.G
a Rubiaceae C and S Europe White(P)open| |
|50
Saxifraga exarataVill.S
e Saxifragaceae C and S Europe toC AsiaWhite(P)open
|
|51
Saxifraga glabellaBertol.S
g Saxifragaceae Balkans, S Italy White(P)open|
|52
Saxifraga scardicaGriseb.S
c Saxifragaceae Balkans White(P)open|
53
Saxifragasempervivum C.KochS
s Saxifragaceae S Balkans, NWAnatoliaPurple(B)tubular
| |
|54
EuphrasiasalisburgensisFunck ex HoppeE
s Scrophulariaceae C and S Europe White(P)tubular| |
|55
Linaria alpina (L.)MillerL
a Scrophulariaceae C and S Europe Violet-blue(B)open| |
|56
LinariapeloponnesiacaBoiss. & Heldr. inBoiss. var.parnassica(Boiss.& Heldr.)HalacsyL
p Scrophulariaceae SW Balkans Yellow(P)open|
|57
Veronicabozakmanii M.A.FischerV
b Scrophulariaceae Greece toAfghanistanViolet-blue(B)tubular
|
|58
Veronica orsinianaTen. ssp.teucrioides (Boiss.& Heldr.) M.A.FischerV
o Scrophulariaceae Greek endemic Violet-blue(B)open| |
59
Daphne oleoidesSchreberD
o Thymelaeaceae S Europe, N andC AfricaWhite(P)tubular
|
|60
Viola graeca (W.Becker) HalacsyV
g Violaceae SW Balkans, SEItalyPurple(B)open
| |
| |61
Viola striis-notata(J. Wagner)Merxm. & LippertV
s Violaceae Olympos endemic Violet-blue(B)open| |
|
ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468 467
References
Alatalo, J.M., Totland, Ø., 1997. Response to simulated
climatic change in an alpine and subarctic pollen-risk
strategist, Silene acaulis. Global Change Biol. 3, 74–79.
Arroyo, M.T.K., Armesto, J.J., Villagran, C., 1981. Plant
phenological patterns in the high Andean cordillera of
central Chile. J. Ecol. 69, 205–223.
Arroyo, M.T.K., Primack, R.B., Armesto, J.J., 1982. Com-
munity studies in pollination ecology in the high temperate
Andes of Central Chile I. Pollination mechanisms and
altitudinal variation. Am. J. Bot. 69, 82–97.
Arroyo, M.T.K., Armesto, J.J., Primack, R.B., 1985. Com-
munity studies in pollination ecology in the high temperate
Andes of Central Chile II. Effects of temperature on
visitation rates and pollination possibilities. Plant Syst.
Evol. 149, 187–203.
Ashman, T.L., Schoen, D.J., 1994. How long should flowers
live? Nature 371, 788–790.
Billings, W.D., Mooney, H.A., 1968. The ecology of arctic and
alpine plants. Biol. Rev. 43, 481–529.
Bingham, R.A., Orthner, A.R., 1998. Efficient pollination of
alpine plants. Nature 391, 238–239.
Blionis, G.J., Vokou, D., 2001. Pollination ecology of
Campanula species on Mt Olympos, Greece. Ecography
24, 287–297.
Blionis, G.J., Halley, J.M., Vokou, D., 2001. Flowering
phenology of Campanula on Mt Olympos, Greece. Eco-
graphy 24, 696–706.
Bliss, L.C., 1971. Arctic and alpine plant life cycles. Ann. Rev.
Ecol. Syst. 2, 405–438.
Breiman, L., Friedman, J.H., Olshen, R.A., Stone, C.J., 1984.
Classification and regression-trees. Wadsworth, Belmont.
Collins, P.D., Harper, K.T., Pendleton, B.K., 1983. Compara-
tive life history and floral characteristics of desert and
mountain floras in Utah. Great Basin Nat. 43, 385–393.
De’ath, G., Fabricius, K.E., 2000. Classification and regres-
sion trees: a powerful yet simple technique for ecological
data analysis. Ecology 81, 3178–3192.
Dupont, Y.L., Hansen, D.M., Olesen, J.M., 2003. Structure of
a plant – flower-visitor network in the high-altitude sub-
alpine desert of Tenerife, Canary Islands. Ecography 26,
301–310.
Evanhoe, L., Galloway, L.F., 2002. Floral longevity in Campanula
americana (Campanulaceae): a comparison of morphological
and functional gender phases. Am. J. Bot. 89, 587–591.
Fabbro, T., Korner, C., 2004. Altitudinal differences in flower
traits and reproductive allocation. Flora 199, 70–81.
Faegri, K., van der Pijl, L., 1979. The Principles of Pollination
Ecology. Pergamon Press, Oxford.
Fauna Europaea Web Service, 2004. Fauna Europaea version
1.1. /http://www.faunaeur.orgS.
Frankie, G.W., Baker, H.G., Opler, P.A., 1974. Comparative
phenological studies of trees in tropical wet and dry forests
in the lowlands of Costa Rica. J. Ecol. 62, 881–913.
Hegland, S.J., Totland, Ø., 2005. Relationships between
species’ floral traits and pollinator visitation in a temperate
grassland. Oecologia 145, 586–594.
Heinrich, B., 1976. Flowering phenologies: bog, woodland,
disturbed habitats. Ecology 57, 890–899.
Henry, G.H.R., Molau, U., 1997. Tundra plants and climate
change: the International Tundra Experiment (ITEX).
Global Change Biol. 3, 1–9.
Herrera, J., 1986. Flowering and fruiting phenology in the coastal
shrublands of Donana, South Spain. Vegetatio 68, 91–98.
Hingston, A.B., McQuillan, P.B., 2000. Are pollination
syndromes useful predictors of floral visitors in Tasmania?
Aust. Ecol. 25, 600–609.
Inouye, D.W., Pyke, G.H., 1988. Pollination biology in the
Snowy Mountains of Australia: comparisons with montane
Colorado, USA. Aust. J. Ecol. 13, 191–210.
IPCC (Intergovernmental Panel on Climate Change), 2007.
Climate Change 2007: The Physical Science Basis. Sum-
mary for Policymakers. WMO, UNEP, Geneva.
Kochmer, J.P., Handel, S.N., 1986. Constraints and competi-
tion in the evolution of flowering phenology. Ecol. Monogr.
56, 303–325.
Korner, C., 1999. Alpine Plant Life. Functional Plant Ecology
of High Mountain Ecosystems. Springer, Berlin.
Lack, A.J., 1982. The ecology of flowers of chalk grassland
and their insect pollinators. J. Ecol. 70, 773–790.
McCall, C., Primack, R.B., 1992. Influence of flower character-
istics, weather, time of day, and season on insect visitation
rates in three plant communities. Am. J. Bot. 79, 434–442.
Medan, D., 2002. Reproductive biology of the Andean shrub
Discaria nana (Rhamnaceae). Plant Biol. 5, 94–101.
Moldenke, A.R., 1976. California pollination ecology and
vegetation types. Phytologia 34, 305–361.
Parrish, J.A.D., Bazzaz, F.A., 1979. Difference in pollination
niche relationships in early and late successional plant
communities. Ecology 60, 597–610.
Petanidou, T., 1991. Pollination ecology in a phryganic
ecosystem. Ph.D. Dissertation, Aristotle University of
Thessaloniki, Greece (in Greek, with an English summary).
Petanidou, T., Ellis, W.N., Margaris, N.S., Vokou, D., 1995.
Constraints on flowering phenology in a phryganic (East
Mediterranean shrub) community. Am. J. Bot. 82, 607–620.
Pickering, C.M., Stock, M., 2004. Insect colour preference
compared to flower colours in the Australian Alps. Nord. J.
Bot. 23, 217–223.
Pojar, J., 1974. Reproductive dynamics of four plant communities
of southwestern British Columbia. Can. J. Bot. 52, 1819–1834.
Price, M.V., Waser, N.M., 1998. Effects of experimental
warming on plant reproductive phenology in a subalpine
meadow. Ecology 79, 1261–1271.
Primack, R., 1985. Longevity of individual flowers. Ann. Rev.
Ecol. Syst. 16, 15–37.
Rathcke, B., 1988. Flowering phenology in a shrub commu-
nity: competition and constraints. J. Ecol. 76, 975–994.
Rathcke, B., 2003. Floral longevity and reproductive assur-
ance: seasonal patterns and an experimental test with
Kalmia latifolia (Ericaceae). Am. J. Bot. 90, 1328–1332.
Raynal, D.J., Bazzaz, F.A., 1975. Interference of winter
annuals with Ambrosia artemisifolia in early successional
fields. J. Ecol. 56, 37–49.
Statistical Sciences, 1999. S-PLUS, version 2000 for Windows.
Mathsoft Inc., Seattle.
Stenstrom, M., Gugerli, F., Henry, H.B.R., 1997. Response of
Saxifraga oppositifolia L. to simulated climate change at
three contrasting latitudes. Global Change Biol. 3, 44–54.

ARTICLE IN PRESSN. Makrodimos et al. / Flora 203 (2008) 449–468468
Strakosh, S.C., Ferguson, C.J., 2005. Pollination biology of
four south-western species of Phlox (Polemoniaceae): insect
visitation in relation to corolla tube length. Southwest. Nat.
50, 291–301.
Strid, A., 1980. Wild Flowers of Mount Olympus. Goulandris
Natural History Museum, Athens.
Strid, A. (Ed.), 1986. Mountain Flora of Greece, vol. 1.
Cambridge University Press, Cambridge.
Strid, A., Tan, K. (Eds.), 1991. Mountain Flora of Greece, vol.
2. Edinburgh University Press, Edinburgh.
Suzuki, S., Kudo, G., 1997. Short-term effects of simulated
environmental change on phenology, leaf traits, and shoot
growth of alpine plants on a temperate mountain, northern
Japan. Global Change Biol. 3, 108–115.
Totland, Ø., 1993. Pollination in alpine Norway: flowering
phenology, insect visitors, and visitation rates in two plant
communities. Can. J. Bot. 71, 1072–1079.
Totland, Ø., 1994. Influence of climate, time of day and
season, and flower density on insect flower visitation in
alpine Norway. Arct. Alp. Res. 26, 66–71.
Warren, S.D., Harper, K.T., Booth, G.M., 1988. Elevational
distribution of insect pollinators. Am. Midl. Nat. 120, 325–330.
Yumoto, T., 1986. The ecological pollination syndromes of insect-
pollinated plants in an alpine meadow. Ecol. Res. 1, 83–95.
Related Documents











