Annals of Botany 88: 989-997, 2001 doi:10.1006/anbo.2001.1534, available online at http://www.idealibrary.com on Flower Morphology and Pollination Mechanism in Three Sympatric Goodyerinae Orchids from Southeastern Brazil RODRIGO B. SINGER* and MARLIES SAZIMA Depto Botanica, IB, Unicamp, Caixa Postal 6109, CEP: 13083-970, Campinas, SP, Brazil Received: 6 April 2001 Returned for revision: 18 June 2001 Accepted: 27 July 2001 Published electronically: 8 October 2001 The pollination biology of Aspidogyne argentea, Aspidogyne longicornu and Erythrodes arietina was studied in south- eastern Brazil. The three species are self-compatible but are pollinator-dependent. These three orchid species offer nectar as a reward to pollinators and flower visitors. The two Aspidogyne spp. have a dorsally-adhesive viscidium, a feature which precludes pollinators other than bees. Erythrodes arietina flowers are protandrous and show a ventrally- adhesive viscidium. Aspidogyne argentea is visited by halictid bees (here ranked as probable pollinators) and Hesperiidae butterflies. Aspidogyne longicornu is pollinated by females of Euglossa (Euglossini) and visited by the hummingbird Phaethornis ruber and by the euglossine bees Eulaema seabrai (females) and Eulaema cingulata (males). The pollinarium adheres to the ventral surface of the bee labrum, a very difficult place for bees to clean. The dorsally adhesive viscidium in Aspidogyne parallels that of the Spiranthinae genera of the so-called 'Pelexia alliance'. This condition seems to be particularly adaptative under conditions of low-frequency pollinator visits. Erythrodes arietina is pollinated by bees of the genera Paratelrapedia and Osiris which carry the pollinarium on the dorsal surface of their proboscis. Occasionally, these bees remove pollinaria from their mouthparts using their forelegs. In general, in the species studied, a combination of both pollinator behaviour and morphological peculiarities promotes cross- pollination. © 2001 Annals of Botany Company Key words: Orchids, Goodyerinae, morphology, viscidium, protandry. Aspidogyne, Erythrodes, pollination, bees, hummingbirds, butterflies, INTRODUCTION Orchids of the subtribe Goodyerinae are widespread but are particularly diverse in the tropics, especially in tropical Asia (Dressier, 1993). All Brazilian Goodyerinae orchids were classified as belonging to either the genus Physurus L. C. Rich. (Hoehne, 1945) or the genus Erythrodes Blume (Pabst and Dungs, 1975). However, Garay (1977) noticed signifi- cant morphological differences among flowers of the neotropical Goodyerinae and proposed a number of generic segregates, mostly based on rostellar structure. Only a few species of Goodyera (Ackerman 1975; Kallunki, 1981; Wong and Sun, 1999) and Ludisia (Van der Pijl and Dodson, 1966) have been studied with respect to their reproductive systems or pollination biologies and, as far as we know, there are no studies of neotropical species. Studies of Goodyera species have shown that these plants are pollinated by bumble-bees (Apidae) (Ackerman, 1975; Kallunki, 1981) and that Ludisia is butterfly-pollinated (Van der Pijl and Dodson, 1966). More recently, Ackerman (1995) suggested that a few Erythrodes species from Puerto Rico and the Virgin Islands could be autogamous, owing to their relatively high fruit set. During our studies into the pollination biology of some Brazilian species of the genera Aspidogyne Garay and Erythrodes Rchb. F., we noticed significant differences in the way the orchid rostellum operates during the pollination process. The purpose of the present contribution is thus two-fold: (1) to present the first data concerning the * For correspondence. E-mail rbsingerl^yahoo.com 0305-7364/01/120989 + 09 S35.00/00 pollination biology of neotropical Goodyerinae, and (2) to highlight rostellar differences that promote different pollination mechanisms. MATERIALS AND METHODS The study was conducted in the Brazilian Atlantic rain- forest at the Reserve of Picinguaba, Ubatuba (approx. 23° 22'S, 44° 50'W), Sao Paulo State, south-eastern Brazil. Annual rainfall reaches 2600 mm and the annual average temperature is approx. 26°C (Nimer, 1977). The climate is tropical-humid (Koeppen, 1948) and without a well-defined dry-cold season (Nimer, 1977): Aspidogyne argentea (Veil.) Garay and A. longicornu (Cogn.) Garay, are syntopic in the understorey of the shrubby formation known as 'restingas', at approx. 5 m a.s.l. Erythrodes arietina (Rchb. F. & Warm.) Ames occurs in the understorey of well-defined montane forest formations at about 60 m.a.s.l. The pollination biology of these three species was studied during several field visits. Phases of anthesis and other flower characteristics were observed in situ. Flowers were observed for visitors at different times of the day, from early morning (0600 h) until late afternoon (approx. 1800 h). Pollination of A. argentea was studied by means of eight field visits: from 26-29 Sept. 1999 and from 1-30 Oct. 1999. A total of 36 h 40 min was spent in observations. The pollination biology of A. longicornu was studied during 28 field visits, from 22 Apr.-lO Jun. 1998, 30 Mar.-26 Apr. 1999 and 8-10 May 2000. A total of 120 observation hours was spent on this orchid species. Pollination of E. arietina % 2001 Annals of Botany Company by guest on June 2, 2016 http://aob.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Annals of Botany 88: 989-997, 2001doi:10.1006/anbo.2001.1534, available online at http://www.idealibrary.com on
Flower Morphology and Pollination Mechanism in Three Sympatric Goodyerinae Orchidsfrom Southeastern Brazil
RODRIGO B. SINGER* and MARLIES SAZIMA
Depto Botanica, IB, Unicamp, Caixa Postal 6109, CEP: 13083-970, Campinas, SP, Brazil
Received: 6 April 2001 Returned for revision: 18 June 2001 Accepted: 27 July 2001 Published electronically: 8 October 2001
The pollination biology of Aspidogyne argentea, Aspidogyne longicornu and Erythrodes arietina was studied in south-eastern Brazil. The three species are self-compatible but are pollinator-dependent. These three orchid species offernectar as a reward to pollinators and flower visitors. The two Aspidogyne spp. have a dorsally-adhesive viscidium, afeature which precludes pollinators other than bees. Erythrodes arietina flowers are protandrous and show a ventrally-adhesive viscidium. Aspidogyne argentea is visited by halictid bees (here ranked as probable pollinators) andHesperiidae butterflies. Aspidogyne longicornu is pollinated by females of Euglossa (Euglossini) and visited by thehummingbird Phaethornis ruber and by the euglossine bees Eulaema seabrai (females) and Eulaema cingulata (males).The pollinarium adheres to the ventral surface of the bee labrum, a very difficult place for bees to clean. The dorsallyadhesive viscidium in Aspidogyne parallels that of the Spiranthinae genera of the so-called 'Pelexia alliance'. Thiscondition seems to be particularly adaptative under conditions of low-frequency pollinator visits. Erythrodes arietinais pollinated by bees of the genera Paratelrapedia and Osiris which carry the pollinarium on the dorsal surface of theirproboscis. Occasionally, these bees remove pollinaria from their mouthparts using their forelegs. In general, in thespecies studied, a combination of both pollinator behaviour and morphological peculiarities promotes cross-pollination. © 2001 Annals of Botany Company
Key words: Orchids, Goodyerinae,morphology, viscidium, protandry.
Aspidogyne, Erythrodes, pollination, bees, hummingbirds, butterflies,
INTRODUCTION
Orchids of the subtribe Goodyerinae are widespread but areparticularly diverse in the tropics, especially in tropical Asia(Dressier, 1993). All Brazilian Goodyerinae orchids wereclassified as belonging to either the genus Physurus L. C.Rich. (Hoehne, 1945) or the genus Erythrodes Blume (Pabstand Dungs, 1975). However, Garay (1977) noticed signifi-cant morphological differences among flowers of theneotropical Goodyerinae and proposed a number of genericsegregates, mostly based on rostellar structure. Only a fewspecies of Goodyera (Ackerman 1975; Kallunki, 1981; Wongand Sun, 1999) and Ludisia (Van der Pijl and Dodson, 1966)have been studied with respect to their reproductive systemsor pollination biologies and, as far as we know, there are nostudies of neotropical species. Studies of Goodyera specieshave shown that these plants are pollinated by bumble-bees(Apidae) (Ackerman, 1975; Kallunki, 1981) and that Ludisiais butterfly-pollinated (Van der Pijl and Dodson, 1966).More recently, Ackerman (1995) suggested that a fewErythrodes species from Puerto Rico and the Virgin Islandscould be autogamous, owing to their relatively high fruit set.
During our studies into the pollination biology of someBrazilian species of the genera Aspidogyne Garay andErythrodes Rchb. F., we noticed significant differences inthe way the orchid rostellum operates during the pollinationprocess. The purpose of the present contribution is thustwo-fold: (1) to present the first data concerning the
* For correspondence. E-mail rbsingerl^yahoo.com
0305-7364/01/120989 + 09 S35.00/00
pollination biology of neotropical Goodyerinae, and (2)to highlight rostellar differences that promote differentpollination mechanisms.
MATERIALS AND METHODS
The study was conducted in the Brazilian Atlantic rain-forest at the Reserve of Picinguaba, Ubatuba (approx. 23°22'S, 44° 50'W), Sao Paulo State, south-eastern Brazil.Annual rainfall reaches 2600 mm and the annual averagetemperature is approx. 26°C (Nimer, 1977). The climate istropical-humid (Koeppen, 1948) and without a well-defineddry-cold season (Nimer, 1977): Aspidogyne argentea (Veil.)Garay and A. longicornu (Cogn.) Garay, are syntopic in theunderstorey of the shrubby formation known as 'restingas',at approx. 5 m a.s.l. Erythrodes arietina (Rchb. F. &Warm.) Ames occurs in the understorey of well-definedmontane forest formations at about 60 m.a.s.l.
The pollination biology of these three species was studiedduring several field visits. Phases of anthesis and otherflower characteristics were observed in situ. Flowers wereobserved for visitors at different times of the day, from earlymorning (0600 h) until late afternoon (approx. 1800 h).Pollination of A. argentea was studied by means of eightfield visits: from 26-29 Sept. 1999 and from 1-30 Oct. 1999.A total of 36 h 40 min was spent in observations. Thepollination biology of A. longicornu was studied during 28field visits, from 22 Apr.-lO Jun. 1998, 30 Mar.-26 Apr.1999 and 8-10 May 2000. A total of 120 observation hourswas spent on this orchid species. Pollination of E. arietina
% 2001 Annals of Botany Company
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

990 Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids
TA B L E 1. Fruit set ( %) in three Goodyerinae species following emasculation self, or cross pollination or a control treatment
N Control Emasculation Self-pollination Cross-pollination
A. argenteaA. longicornuE. arietina
1264
0 (0/43)0(0/101)0 (0/74)
0(0/34)0(0/104)0(0/21)
47-06(24/51)97-3 (36/37)48-8 (21/43)
8611 (31/36)100 (30/30)
Numbers in parentheses represent the number of fruits obtained over the number of flowers used in each treatment. N, Number of individualsper species.
X2 values (self x cross-pollination) = A. argentea: 12-57 (significant, P > 001). A. longicornu: 0-82 (non-significant, P = 0-55).
was studied during seven field visits, from 20-25 Oct. 1998and 30-31 Oct. 2000, giving a total of 33 h 30 min ofobservations. The behaviour of both pollinators and flowervisitors was recorded in the field through notes andphotographs. Some insects were captured for identificationand vouchers are deposited at the Laboratorio de Abelhasda Universidade de Sao Paulo (USP) and at the Mourecollection (Universidade Federal do Parana). Plantvouchers are deposited at the Herbarium of the Univers-idade Estadual de Campinas (UEC): voucher numbers:Aspidogyne argentea, R. B. Singer 99/06; Aspidogynelongicornu, R. B. Singer 98/42; Erythrodes arietina, R. B.Singer 98/104.
In 1999, the fruiting success (number of dehiscent fruitsdivided by the number of flowers produced) was calculatedfor the three orchid species. Seven inflorescences ofA. argentea (totalling 135 flowers), 40 of A. longicornu(totalling 1794 flowers) and seven inflorescences ofE. arietina (totalling 571 flowers) were available for study.
Breeding systems were studied in plants cultivated in aglasshouse at the Universidade Estadual de Campinas. Thenumber of plants per species and the number of flowersper treatment are summarized in Table 1. Treatmentsincluded untouched flowers (control), self-pollination,cross-pollination, and emasculation. Cross-pollinationcould not be performed in E. arietina since the fewinflorescences obtained flowered at considerably distincttimes. Treatments yielding fruits were compared by pairs,through a %2 test (a = 005, d.f. = 1). When necessary, theFischer correction was performed.
Flower features were drawn using fresh flowers, with thehelp of a stereomicroscope with a camera lucida attach-ment. Flower morphology and general orchid taxonomyfollow Dressier (1993) and generic Goodyerinae conceptsfollow Garay (1977).
RESULTS AND DISCUSSION
General flower features
The present contribution deals mainly with rostellarstructure and function. Readers interested in perianthparts, shape and measurements are referred to Hoehne(1945) and to Pabst and Dungs (1975). The plants show theterrestrial, creeping habit usually found in Goodyerinae(Dressier 1993) and the inflorescences are terminal spikes.The flowers are resupinate and more or less tubular in shape,bearing noticeable spurs (Fig. 1A, E and I), where nectaraccumulates and is offered to flower visitors. The flowers of
the species studied show an indument of hairs (Fig. 1 A, Eand I). In the species studied, the column is erect and therostellum is acute (Fig. IB, F and J). The pollinarium iscomposed of two bipartite pollinia, hard caudicles (Ras-mussen, 1986) and a well-developed viscidium (Fig. ID, Hand L). In the two Aspidogyne species, the apex of thelanceolate rostellum detaches as a viscidium, leaving a bluntrostellar remnant (Fig. 1C, and G). In Erythrodes arietina,the long viscidium detaches leaving a bifid rostellar remnant(Fig. IK). Pollinia are composed of a myriad of laminarsubunits (Fig. ID, H and L) known as 'massulae' — as isalways the case in Goodyerinae orchids (although thisfeature is not exclusive to this orchid group). Pollinariaconstructed in this way are known as 'sectile' (Rasmussen,1982; Dressier, 1993; Freudenstein and Rasmussen, 1997).The stigmatic surface is flat to slightly convex. In the twoAspidogyne species the stigmatic surface is broad and almostentire in shape (Fig. 1B, and F). In E. arietina, the stigmaticsurface is deeply bilobed (Fig. U).
Specific flower features and pollination mechanisms
Aspidogyne argentea. The flowers are whitish withnoticeable purple dots on the lateral petals and sepals(Fig. 2A), measuring about 9 mm in length. During thesunniest hours, flowers emit a sweet, musky fragrance. Theviscidium in these flowers is adhesive only on its dorsalsurface. Untouched flowers may retain their fresh appear-ance for 5-10 d.
A number of visitors to flowers of A. argentea wererecorded, but further observations are needed to confirmwhich is the legitimate pollinator. Insect visits to the flowersoccurred during the sunniest hours (1000-1400 h), coincid-ing with the emission of the strongest flower fragrances.Hesperiidae butterflies (Fig. 2A) were recorded visiting theflowers several times, probing many flowers in a singleinflorescence and visiting one-three inflorescences in thepopulation, spending up to 60-70 s in total. The butterfliesnever removed any pollinaria. The small MeliponinaePlebeia cf. droryana was occasionally sighted at the flowers,but this bee is too small to operate the flower mechanism.Medium to relatively large sweat bees (Halictidae) of thegenera Pseudoaugochlora and Augochloropsis were alsosighted at the flowers. These bees probed flowers for nectarand may well be the legitimate pollinators. The few (two)bees we captured lacked pollinaria; these were possibly lostin the net. Halictid visits to the flowers were very infrequent(one-three per day) and occurred between 1000-1400 h.
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
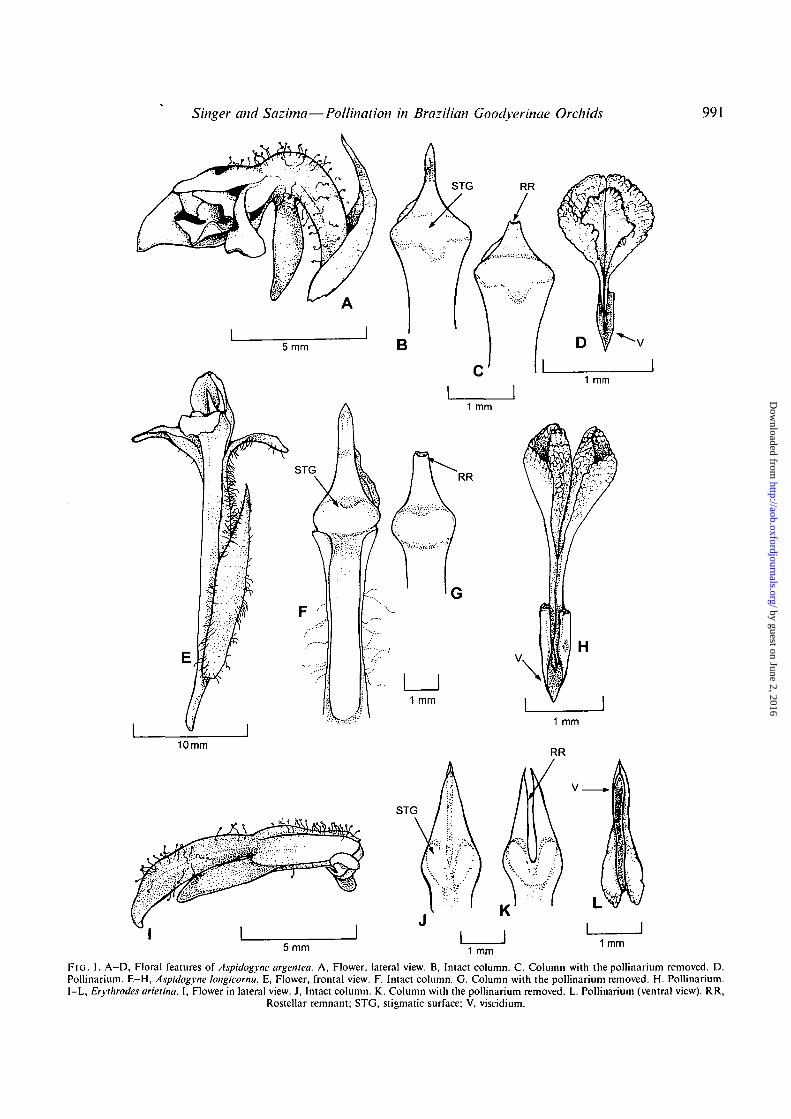
Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids 991
5 mm 1 mm1 mm
F I G . 1. A-D, Floral features of Aspidogyne argentea. A, Flower, lateral view. B, Intact column. C. Column with the pollinarium removed. D.Pollinarium. E—H, Aspidogyne longicornu. E, Flower, frontal view. F. Intact column. G. Column with the pollinarium removed. H. Pollinarium.I-L, Eryihrodes arieiina. I, Flower in lateral view. J, Intact column. K. Column with the pollinarium removed. L. Pollinarium (ventral view). RR,
Rostellar remnant; STG, stigmatic surface; V, viscidium.
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

992 Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids
F I G . 2. A, Hesperiidae butterfly probing flowers of A. argenlea. B-D, Visitors and pollinators of A. longicornu. B, Euglossacf. igniia (pollinator).C, Phaethornis ruber (visitor). D, Eulaema cingulata (visitor). E and F, Pollinators of E. arietina. E, Osiris sp. F, Paralelrapedia sp.
These bees visited a few (one-three) flowers per inflores-cence and one-two inflorescences per population, spendingin total around 5-10 s. Small sweat bees of the genusPseudoagapostemon were very frequent visitors to the
flowers, but are too small to pollinate them. As theviscidium is adhesive through its dorsal surface, it canonly be fixed when dorsally pressed. This feature precludespollinators other than bees (Singer and Sazima, 1999).
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids 993
F I G . 3. Fixation of pollinarium on the pollinators. A, Female oi Euglossa chalybeata iopoecila with a pollinarium of A. longicornu attached to theventral surface of its labrum. B, Female of Osiris sp. with a pollinarium of E. arietina attached to the dorsal surface of its proboscis.
When the bees probe this kind of flower, they extend theirmouth-parts and press the dorsal surface of the viscidiumwith the labrum. The viscidium then adheres to the ventralsurface of the labrum and the pollinarium is removed whenthe insect leaves the flower (Singer and Sazima, 1999).Other flower-visiting animals such as the Hesperiidaebutterflies, which lack a labrum, only exert pressure againstthe ventral surface of the column, without fixing andremoving pollinaria. Since the only appropriately sized beeswe noticed visiting flowers were halictids, we expect thatfurther fieldwork will confirm these bees to be legitimatepollinators of this orchid species.
In 1999, about 11% of the available flowers (15/135;n = 1 inflorescences) set fruits and the individual fruitingsuccess ranged from 0 to 33 %.
Aspidogyne longicornu. The flowers are whitish andremarkably long-spurred, about 23 mm in length (exclud-ing the spur). The spur itself measures approx. 25-26 mmin length. During the hottest hours the flowers emit amusky, somewhat unpleasant, fragrance which is betterperceived by enclosing the flowers in a vial. As inA. argentea, the viscidium is adhesive through its dorsalsurface. Flowers may keep their fresh appearance for 4-5 d.
Diverse visitors were recorded at the flowers, alwaysduring the daytime hours. Pollinators are females ofEuglossa chalybeata iopoecila and of Euglossa cf. ignita(both Apidae; Euglossini) (Fig. 2B), which were sighted atthe flowers from 0600 to 1400 h. These bees are noticeablyinfrequent visitors to the flowers, consecutive bee visitsbeing separated by 40-120 min intervals. This low polli-nator frequency may be related to both pollinator scarcityand trapliner habit (Janzen, 1971). The bees normallyvisited a few flowers (one-three) per inflorescence and
one-five inflorescences per visit to the population, spending15-60 s in total. These Euglossa bees hover facing theinflorescences, land and then extend their mouth-parts toprobe a flower. As they insert their mouth-parts they pressthe dorsal surface of the viscidium with the ventral surfaceof the labrum, thus fixing the pollinarium (Fig. 3A). Thepollinarium is removed when the bees leave the flowers.Since the labrum is bent when a bee retracts its mouth-parts, the pollinarium remains protected under the bee'shead, in much the same way as in the Spiranthinae orchidsof the 'Pelexia alliance' (Singer and Sazima, 1999). When apollinarium-bearing Euglossa visits other flowers, the distalparts of the pollinarium touch the stigmatic surface, andclumps of massulae are then deposited. The size and shapeof the fixed pollinarium, just as with the orchid species ofthe 'Pelexia alliance', hinder the fixation of a new one, thusavoiding pollinaria being wasted and enhancing the chancesof cross-pollination. Only bees carrying a single pollinariumwere observed. Since the pollinarium is friable and made upof numerous massulae, many flowers may be pollinatedwith the pollen content of a single pollinarium (Dressier,1993; Freudenstein and Rasmussen, 1997).
The flowers of A. longicornu are visited frequently by thehermit hummingbird Phaethornis ruber (Phaethorninae)(Fig. 2C). Consecutive hummingbird visits occurred at 20-60 min intervals. In all cases, the hummingbirds system-atically visited all the fresh-looking flowers, spending up to120 s per visit to the population. Hummingbirds hoverinfront of the inflorescences, inserting the tip of their bills tolick the nectar (Fig. 2C). The birds almost emptied most ofthe spurs of the flowers they visited but never removedpollinaria. Although visiting flowers in a legitimate way,P. ruber acts as a nectar thief (Buzato et al., 2001) of thisorchid species. This can be explained because of the nature
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
994 Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids
of the viscidium, which is adhesive only through its dorsalsurface. Lacking labrum-like structures suited to exert adorsal pressure against the viscidium, hummingbirds do notfix pollinaria. The hummingbirds clearly defended the plantpopulation as a foraging patch and showed territorialbehaviour against the euglossines when both kinds of flowervisitors met at the patch. Phaethorninae hummingbirds aresaid to show trapliner habit (Sazima el ai, 1996, andreferences therein), but the territorial behaviour of P. ruberat these flowers may be explained by the fact that the otherflower visitors were bees. Territoriality in P. ruber wasrecently reported by San Martin-Gajardo and Freitas(1999). Another explanation may be related to the floweringperiod of E. longicornu which flowers during a period ofresource scarcity for this hummingbird species. It seemsplausible that both the aggressive behaviour of thehummingbirds toward the euglossines and the removal ofnectar by these birds may have a deleterious effect on thereproductive success of this orchid species.
Other flower visitors to A. longicornu were females ofEulaema seabrai and, more sporadically, males of Eulaemacingulata (Apidae: Euglossini) (Fig. 2D). These bees do notremove the pollinaria since their extended mouth-parts arelonger than those of the Euglossa bees and they do not presstheir labrum against the viscidium. Phaethomis ruber alsodefended the flowers against these big bees when both kindsof animals met at the flowers. Hesperiidae butterflies wereoccasionally sighted at the flowers but, for the same reasonsnoted above for A. argentea, these insects did not removepollinaria.
In 1999, about 18% of the available flowers (323/1794;n = 40 inflorescences) set fruits and the fruiting success perinflorescence ranged from 5 to 43 %.
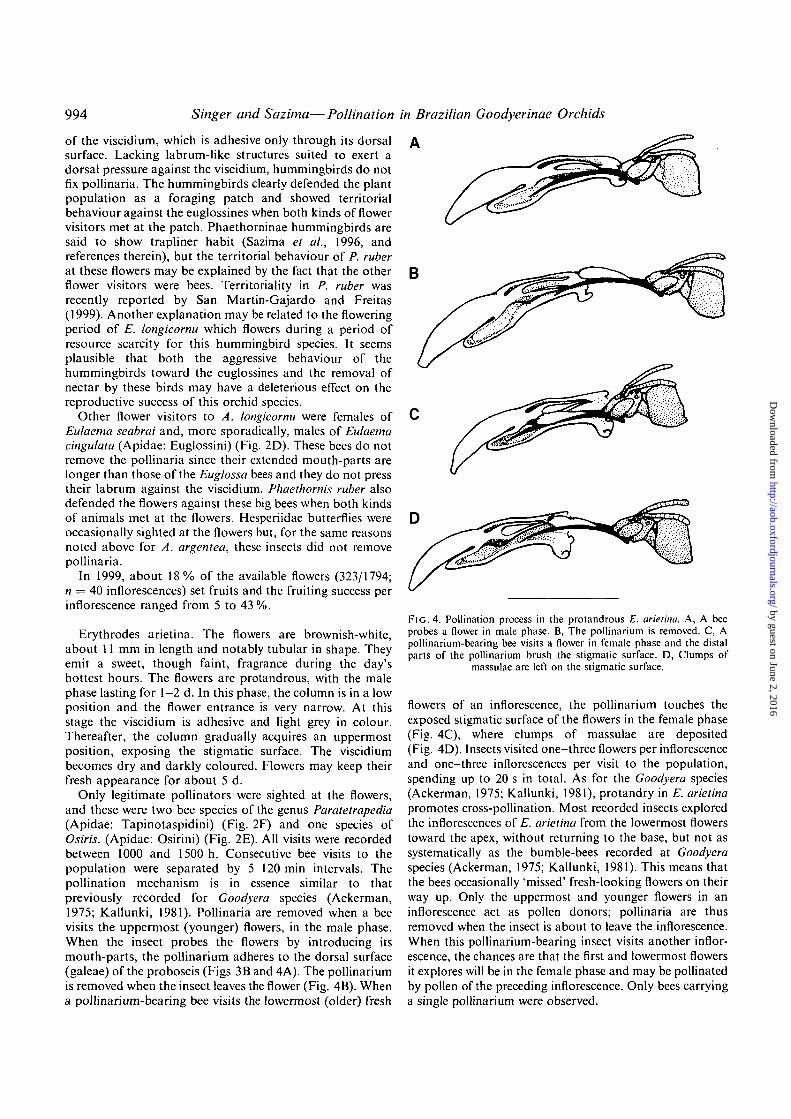
Erythrodes arietina. The flowers are brownish-white,about 11 mm in length and notably tubular in shape. Theyemit a sweet, though faint, fragrance during the day'shottest hours. The flowers are protandrous, with the malephase lasting for 1-2 d. In this phase, the column is in a lowposition and the flower entrance is very narrow. At thisstage the viscidium is adhesive and light grey in colour.Thereafter, the column gradually acquires an uppermostposition, exposing the stigmatic surface. The viscidiumbecomes dry and darkly coloured. Flowers may keep theirfresh appearance for about 5 d.
Only legitimate pollinators were sighted at the flowers,and these were two bee species of the genus Paratetrapedia(Apidae: Tapinotaspidini) (Fig. 2F) and one species ofOsiris. (Apidae: Osirini) (Fig. 2E). All visits were recordedbetween 1000 and 1500 h. Consecutive bee visits to thepopulation were separated by 5-120 min intervals. Thepollination mechanism is in essence similar to thatpreviously recorded for Goodyera species (Ackerman,1975; Kallunki, 1981). Pollinaria are removed when a beevisits the uppermost (younger) flowers, in the male phase.When the insect probes the flowers by introducing itsmouth-parts, the pollinarium adheres to the dorsal surface(galeae) of the proboscis (Figs 3B and 4A). The pollinariumis removed when the insect leaves the flower (Fig. 4B). Whena pollinarium-bearing bee visits the lowermost (older) fresh
FlG. 4. Pollination process in the protandrous E. arietina. A, A beeprobes a flower in male phase. B, The pollinarium is removed. C, Apollinarium-bearing bee visits a flower in female phase and the distalparts of the pollinarium brush the stigmatic surface. D, Clumps of
massulae are left on the stigmatic surface.
flowers of an inflorescence, the pollinarium touches theexposed stigmatic surface of the flowers in the female phase(Fig. 4C), where clumps of massulae are deposited(Fig. 4D). Insects visited one-three flowers per inflorescenceand one-three inflorescences per visit to the population,spending up to 20 s in total. As for the Goodyera species(Ackerman, 1975; Kallunki, 1981), protandry in E. arietinapromotes cross-pollination. Most recorded insects exploredthe inflorescences of E. arietina from the lowermost flowerstoward the apex, without returning to the base, but not assystematically as the bumble-bees recorded at Goodyeraspecies (Ackerman, 1975; Kallunki, 1981). This means thatthe bees occasionally 'missed' fresh-looking flowers on theirway up. Only the uppermost and younger flowers in aninflorescence act as pollen donors; pollinaria are thusremoved when the insect is about to leave the inflorescence.When this pollinarium-bearing insect visits another inflor-escence, the chances are that the first and lowermost flowersit explores will be in the female phase and may be pollinatedby pollen of the preceding inflorescence. Only bees carryinga single pollinarium were observed.
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids 995
An interesting behaviour was occasionally recorded: bothOsiris and Paratetrapedia bees carrying pollinaria landedon surrounding vegetation (e.g. a leaf) and actively removedpollinaria by extending their mouth-parts and pressingthem with their forelegs.
Osiris bees have been recorded as pollinators of somePalmorchis (Palmorchideae) species from Panama(Dressier, 1984). Remarkably, Osiris bees are parasiticand Paratetrapedia bees are suspected to be one of theirhosts (Shanks, 1986).
In 1999, about 23% of the available flowers (129/571;n = 1 inflorescences) set fruits and fruiting success perinflorescence ranged from 10 to 40%.
Breeding systems
The three species studied are self-compatible but arepollinator-dependent (Table 1). No untouched or emascu-lated flowers ever set fruits. These results comply withprevious reports involving species of the genus Goodyera(Ackerman, 1975; Kallunki, 1981; Wong and Sun, 1999). Inother words, in the three species studied, pollinators arerequired for the plants to set fruits and seeds. A. argenteashowed a noticeably higher fruit set following cross-pollination (Table 1), suggesting that inbreeding depressionmay be expressed in this orchid species; further studies arerequired to confirm this. In the three species studied thebehaviour of the flower visitors (tending to visit only afew flowers per inflorescence) plus the morphologicalpeculiarities of the flowers (namely protandry inE. arietina and the dorsally adhesive viscidium of the twoAspidogyne species) clearly promote cross-pollination.
Ackerman (1995) suggested that some Erythrodes speciesfrom Puerto Rico and the Virgin Islands could beautogamous owing to their apparently high fruiting success.Among the Brazilian Goodyerinae orchids studied here,there is at least one case suggestive of autogamy (automaticself-pollination). While studying fresh flowers of Erythrodesaustrobrasiliensis (Porsch) Pabst (Fig. 5A), we noticedsignificant rostellar modifications: in this orchid speciesthe rostellum is remarkably short (Fig. 5B) and theviscidium is completely dry and is separated from the restof the rostellum. The viscidium is thus non-adhesive andhard, surrounded by hard caudiculae (Fig. 5C). When weremoved pollinaria from the clinandrium, we noticed thatmany massulae remained at the clinandrium apparentlywetted by the abundant stigmatic secretion. It seems thatowing to reduction and narrowing of the rostellum the sidesof the pollinarium are passively wetted by the stigmaticsecretion. A similar condition was found by Catling (1987)for some autogamous forms of Stenorrhynchos lanceolatus(as Saccoila lanceolatd).
Thoughts on the ecological value of some flower features
The biological relevance of sectile pollinaria in Orchida-ceae has already been emphasized by Freudenstein andRasmussen (1996). Since the pollinaria of the Goodyerinaeorchids are sectile (composed of smaller subunits, called'massulae'), many flowers can be pollinated with the pollen
content of a single pollinarium. On the other hand, eachmassula derives from a single pollen mother cell (Freuden-stein and Rasmussen, 1997, and references therein). Thus,the pollen content of each massula represents a differentgenotype. During the pollination process, clumps ofmassulae are left on the stigmatic surface and the chancesof multiple-genotype contributions to each capsule (fruit)are then increased (Freudenstein and Rasmussen, 1997). Wewould like to add that since the stigmatic surfaces ofGoodyerinae are broad and slightly convex, the possibilityof mixed pollen loads (i.e. either from the same inflores-cence or a different one) and, consequently, of cross-pollination, is increased.
The three species studied displayed relatively low fruit set;this is in accordance with patterns suggested by Neilandand Wilcock (1998) for tropical orchids.
The presence of protandry in E. arietina is significant interms of its reproductive biology since it promotes cross-pollination (Ackerman, 1975; Catling and Catling, 1991).Among the Goodyerinae orchids, protandry has alreadybeen reported in species of the genus Goodyera (Ackerman,1975). Remarkably, the two Aspidogyne species studied arenon-protandrous, but show dorsally adhesive viscidiainstead. From an ecological point of view, it seems thatprotandry has been 'substituted' through a sum of rostellarrefinements (e.g. dorsally adhesive viscidia) and 'adequate'pollinator behaviour (e.g. trapliner habit).
Dorsally adhesive viscidia occur in the Spiranthinaeorchids of the so-called 'Pelexia alliance' (Singer andSazima, 1999, and references therein), though these viscidiaare not morphologically equivalent to those observed in theAspidogyne species. The viscidia of the 'Pelexia alliance' arewedge-like in shape, with a turgid membrane holding therostellar cement. When the viscidium is dorsally pressed (acondition only achieved in nature by a bee labrum), themembrane ruptures and liberates the rostellar cement whichfixes the pollinarium to the pollinator (Singer and Sazima,1999). The viscidium in Aspidogyne seems to be muchsimpler, consisting of the whole detachable apex of therostellum, with its dorsal surface being adhesive. Cultivatedplants of the genus Ligeophila (L. juruensis) (Fig. 5D-G)also show a dorsally adhesive viscidium. This orchid alsoshows other remarkable column features, such as anextremely flexible (but not 'triggered' as mentioned byGaray, 1977) rostellum (Fig. 5E-F) perpendicularly dis-posed to a slightly bilobed stigmatic surface. At least theAsiatic Goodyerinae genus Aenhenrya Gopalan also showsa dorsally adhesive viscidium (Kumar and Rasmussen,1997). Although dorsally adhesive viscidia apparently aroseonly once among the Spiranthinae orchids (Singer andSazima, 1999, and references therein), this does not seem tobe the case for genera of subtribe Goodyerinae, in whichdorsally adhesive viscidia may have evolved many times. Asfor the Spiranthinae, the available evidence does not allowus to ascertain how dorsally adhesive viscidia evolvedamong the Goodyerinae orchids. However, it seemsparsimonious to suppose that this kind of viscidium evolvedfrom an ancestral ventrally adhesive condition. Thisquestion can only be addressed following phylogeneticstudies of all Goodyerinae orchids.
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

996 Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids
STG
1 mm 1 mm
5 mm
1 mmF I G . 5. Floral features of other remarkable Brazilian Goodyerinae orchids. A-C, Erythrodes austrobrasiliensis. A, Flower in lateral view. B,Column in ventral view. C, Pollinarium. D-G, Ligeophila juruensis. D, Flower in lateral view. E-F, Column in lateral view (E) and ventral view
(F). G, Pollinarium. STG, Stigmatic surface; V, viscidium.
As in the Spiranthinae orchids, the advantages ofdorsally adhesive viscidia in the Goodyerinae are remark-able because: (1) the ventral surface of the labrum (wherethe pollinarium is fixed) is a difficult place for the bee toclean; (2) the labrum is flexible and bent when the insect isflying, thus the fixed pollinarium remains protected underthe insect's head; and (3) this kind of viscidium seems to beparticularly adaptive in the context of pollinator scarcity orpollinators with trapliner habit.
(COTEC 344/98, Processo SMA 41.661/98), and is part ofthe first author's PhD Thesis (in preparation). We thank thestaff of Nucleo Picinguaba, particularly its director LuizRoberto C. Numa de Oliveira for moral and logisticsupport. Glauco Machado (Unicamp) helped with statisti-cal analysis. Andre Olmos Simoes and Samantha Koehler(Unicamp) helped in many ways. Isabel A. dos Santos andF. Zanella (USP) helped with bee identification. Prof. AlainFrancois is thanked for improving English expression.
A C K N O W L E D G E M E N T S
This contribution describes the results of the project'Estudo de Biologia Reprodutiva em Orquideas Nativasno Parque Estadual da Serrado Mar (Nucleo Picinguaba)'
L ITERATURE CITED
Ackerman JD. 1975. Reproductive biology of Goodyera oblongifolia(Orchidaceae). Madrono 23: 191-198.
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Singer and Sazima—Pollination in Brazilian Goodyerinae Orchids 997
Ackerman JD. 1995. An orchid flora of Puerto Rico and The VirginIslands. Memoirs of the New York Botanical Garden. Vol 73.New York.
Buzato S, Sazima M, Sazima I. 2000. Hummingbird-pollinated florasat three Atlantic Forest sites. Biotropica 32 Supplement(4B):824-841.
Catling PM. 1987. Notes on the breeding system of Saccoila lanceolata(Aublet) Garay (Orchidaceae). Annals of the Missouri BotanicalGarden 74: 58-68.
Catling PM, Catling VR. 1991. A synopsis of breeding systems andpollination in North American orchids. Lindleyana 6: 187-210.
Dressier RL. 1984. Palmorchis in Panama, with a new species whereleast expected. Orquidea (Mex.) 9: 213-230.
Dressier RL. 1993. Phytogeny and classification of the orchid family.Portland: Dioscorides Press.
Freudenstein JV, Rassmusen FN. 1997. Sectile pollinia and relation-ships in the Orchidaceae. Plant Systemalics and Evolution 205:125-146.
Garay L. 1977. Systematics of the Physurinae (Orchidaceae) in the NewWorld. Bradea 2: 191-208.
Hoehne FC. 1945. Orchidaceas. In: Hoehne FC, ed. Flora Brasilica.Vol XII. Sao Paulo: Instituto de Botanica.
Jan/en DH. 1971. Euglossine bees as long-distance pollinators oftropical plants. Science 171: 203-205.
Kallunki JA. 1981. Reproductive biology of mixed-species populationsof Goodyera (Orchidaceae) in Northern Michigan. Brittonia 33:137-155.
Koeppen W. 1948. Climatologia: con un estudio de los climas de la tierra.Mexico: Fondo de Cultura Economica.
Kumar SC, Rassmusen FN. 1997. The reappearance of Odontochilusrotundifolius Blatter and its transfer to Aenhenrya Gopalan(Orchidaceae). Novon 7: 81-84.
Neiland MR, Wilcock CC. 1998. Fruit set, nectar reward and rarity inthe Orchidaceae. American Journal of Botany 85: 1657-1671.
Nimer E. 1977. Climatologia da regiao sudeste do Brasil. RevistaBrasileira de Geografia 34: 3-48.
Pabst G, Dungs F. 1975. Orchidaceae Brasilienses. Band I. Brucke:Hildesheim.
Rassmusen FN. 1982. The gynostemium of the neottioid orchids. OperaBotanica 65: 1-95.
Rassmusen FN. 1986. On the various contrivances by which polliniaare attached to viscidia. Lindleyana 1: 21-32.
San Martin-Gajardo I, Freitas L. 1999. Hummingbird pollination inBesleria mucronata Hoehne (Gesneriaceae) in southeastern Brazil.Biociencias 7: 13—24.
Sazima I, Buzato S, Sazima M. 1996. An assemblage of hummingbird-pollinated flowers in a montane forest in southeastern Brazil.Botanica Ada 109: 149-160.
Shanks SJ. 1986. A revision of the neotropical bee genus Osiris(Hymenoptera: Anthophoridae). The Wasmann Journal of Biology44: 1-56.
Singer RB, Sazima M. 1999. The pollination mechanism in the "Pelexiaalliance" (Orchidaceae: Spiranthinae). Botanical Journal of theLinnean Society 131: 249-262.
van der Pijl L, Dodson CH. 1966. Orchid flowers: their pollination andevolution. Coral Gables: University of Miami Press.
Wong KC, Sun M. 1999. Reproductive biology and conservationgenetics of Goodyera procera (Orchidaceae). American Journal ofBotany 86: 1406-1413.
by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

by guest on June 2, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
Related Documents











