Flavonoids: a colorful model for the regulation and evolution of biochemical pathways Ronald Koes, Walter Verweij and Francesca Quattrocchio Department of Genetics, Institute for Molecular Biological Sciences, Vrije Universiteit, de Boelelaan 1085, 1081HV Amsterdam, The Netherlands For more than a century, the biosynthesis of flavonoid pigments has been a favorite of scientists to study a wide variety of biological processes, such as inheritance and transposition, and has become one of the best- studied pathways in nature. The analysis of pigmenta- tion continues to provide insights into new areas, such as the channeling and intracellular transport of metab- olites, regulation of gene expression and RNA inter- ference. Moreover, because pigmentation is studied in a variety of species, it provides unique molecular insights into the evolution of biochemical pathways and regu- latory networks. Synthesis and function of flavonoid pigments The pigments that color most flowers, fruits and seeds are flavonoid secondary metabolites. Flavonoids are syn- thesized by a branched pathway that yields both color- less compounds (e.g. flavonols) and colored pigments (e.g. anthocyanins and the polymeric phlobaphenes and proanthocyanidins) (Figure 1). The different flavonoids have a multitude of biological functions, including protection against UV-light and phytopathogens (e.g. phytoalexins in legumes), signaling during nodulation, male fertility, and auxin transport [1,2]. It has long been suspected that the coloration of flowers is a visual signal to attract pollinating animals. Recent field studies with nearly isogenic Mimulus lines have shown that a (carotenoid-based) color change can indeed lead to a switch from bumblebee to hummingbird- pollination, which can result in genetic isolation and ultimately speciation [3]. However, whether pigmentation (or other floral traits) plays an equally important role in other species is debatable [4]. Analysis of the flower pigmentation gene PhAN2 in petunia species with colored and white flowers showed that the color change as a result of the an2 mutation happened after their genetic isolation and thus was, at least in this case, not the prime cause of speciation [5]. The pigmentation pathways provide a natural reporter gene system that has been used to study a wide variety of processes since the works of Gregor Mendel. Here we review more recent work on the molecular analysis of pigmentation genes, providing new insights into the intra- cellular transport of metabolites and the regulation of biochemical pathways. Curiously, pigmentation is inti- mately linked with the control of several other processes, including cell morphogenesis (hair formation) and the biogenesis and physiology of vacuoles. Biosynthesis and transport Most of the flavonoid enzymes are recovered in the ‘soluble’ cell fractions; immuno-localization experiments suggest that they are loosely bound to the endoplasmic reticulum, possibly in a multi-enzyme complex [6,7], whereas the pigments themselves accumulate in the vacuole (anthocyanins and proanthocyanidins) or the cell wall (phlobaphenes) [8]. In maize, the vacuolar sequestration of anthocyanins requires an MRP-type (multidrug resistance associated protein) of transporter on the tonoplast membrane, the expression of which is co-regulated with the structural anthocyanin genes [9]. MRPs are often referred to as glutathione S-X (GS-X) pumps because they transport a variety of glutathione conjugates. Indeed, glutathione S-transferase (GST)-like proteins have been shown to be required for vacuolar sequestration of pigments in maize (ZmBz2), petunia (PhAN9) and Arabidopsis (AtTT19) [10–12]. Because anthocyanin–glutathione conjugate(s) have not been found, it was proposed that these GSTs might deliver their flavonoid substrate directly, without glutathionation, to the transporter [13]. AtTT12 of Arabidopsis encodes a protein of the MATE (multidrug and toxic compound extrusion) group of transporters that is required for vacuolar localization of proanthocyanidins in the testa of the seed [14]. Although MATE transporters are not known to require glutathion- ated substrates, the vacuolar localization of proantho- cyanidins does require the GST-like protein AtTT19 [12]. Proanthocyanidin precursors in tt19 mutants appear to accumulate in vesicle-like structures rather than freely in the cytoplasm, suggesting that AtTT19 does not act in membrane transport as such but in subsequent transport of the vesicles to the large vacuole [12]. In tomato, an AtTT12-like membrane transporter and a PhAN9/ AtTT19-like GST have been identified that are co- regulated with other structural anthocyanin genes and Corresponding author: Koes, R. ([email protected]). Available online 26 March 2005 Review TRENDS in Plant Science Vol.10 No.5 May 2005 www.sciencedirect.com 1360-1385/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.tplants.2005.03.002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Flavonoids: a colorful model for theregulation and evolution ofbiochemical pathwaysRonald Koes, Walter Verweij and Francesca Quattrocchio
Department of Genetics, Institute for Molecular Biological Sciences, Vrije Universiteit, de Boelelaan 1085, 1081HV Amsterdam,
The Netherlands
For more than a century, the biosynthesis of flavonoid
pigments has been a favorite of scientists to study a
wide variety of biological processes, such as inheritance
and transposition, and has become one of the best-
studied pathways in nature. The analysis of pigmenta-
tion continues to provide insights into new areas, such
as the channeling and intracellular transport of metab-
olites, regulation of gene expression and RNA inter-
ference. Moreover, because pigmentation is studied in a
variety of species, it provides unique molecular insights
into the evolution of biochemical pathways and regu-
latory networks.
Synthesis and function of flavonoid pigments
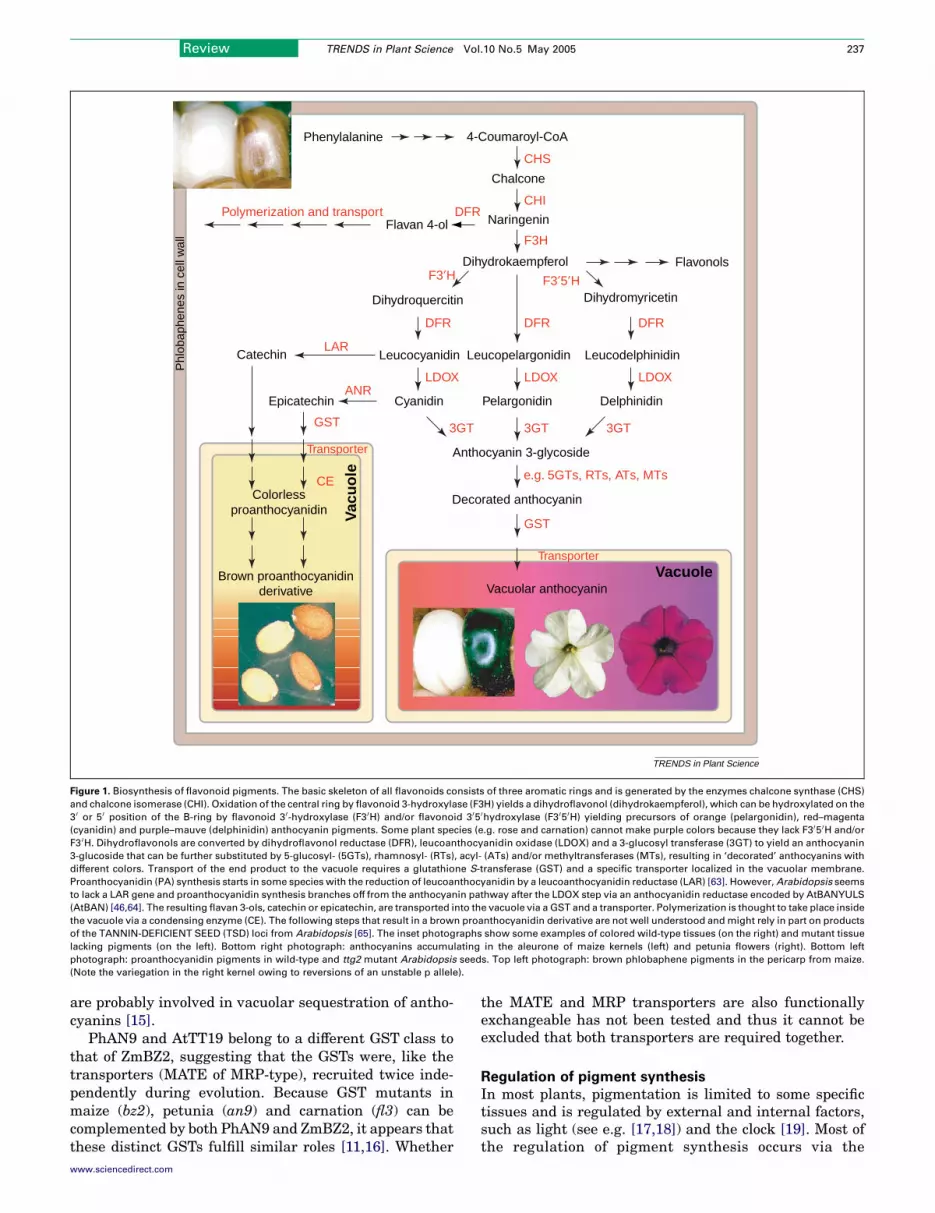
The pigments that color most flowers, fruits and seeds areflavonoid secondary metabolites. Flavonoids are syn-thesized by a branched pathway that yields both color-less compounds (e.g. flavonols) and colored pigments(e.g. anthocyanins and the polymeric phlobaphenes andproanthocyanidins) (Figure 1).
The different flavonoids have a multitude of biologicalfunctions, including protection against UV-light andphytopathogens (e.g. phytoalexins in legumes), signalingduring nodulation, male fertility, and auxin transport[1,2]. It has long been suspected that the coloration offlowers is a visual signal to attract pollinating animals.Recent field studies with nearly isogenic Mimulus lineshave shown that a (carotenoid-based) color change canindeed lead to a switch from bumblebee to hummingbird-pollination, which can result in genetic isolation andultimately speciation [3]. However, whether pigmentation(or other floral traits) plays an equally important role inother species is debatable [4]. Analysis of the flowerpigmentation gene PhAN2 in petunia species with coloredand white flowers showed that the color change as a resultof the an2mutation happened after their genetic isolationand thus was, at least in this case, not the prime cause ofspeciation [5].
The pigmentation pathways provide a natural reportergene system that has been used to study a wide variety ofprocesses since the works of Gregor Mendel. Here wereview more recent work on the molecular analysis of
Corresponding author: Koes, R. ([email protected]).Available online 26 March 2005
www.sciencedirect.com 1360-1385/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
pigmentation genes, providing new insights into the intra-cellular transport of metabolites and the regulation ofbiochemical pathways. Curiously, pigmentation is inti-mately linked with the control of several other processes,including cell morphogenesis (hair formation) and thebiogenesis and physiology of vacuoles.
Biosynthesis and transport
Most of the flavonoid enzymes are recovered in the‘soluble’ cell fractions; immuno-localization experimentssuggest that they are loosely bound to the endoplasmicreticulum, possibly in a multi-enzyme complex [6,7],whereas the pigments themselves accumulate in thevacuole (anthocyanins and proanthocyanidins) or the cellwall (phlobaphenes) [8].
In maize, the vacuolar sequestration of anthocyaninsrequires an MRP-type (multidrug resistance associatedprotein) of transporter on the tonoplast membrane, theexpression of which is co-regulated with the structuralanthocyanin genes [9]. MRPs are often referred to asglutathione S-X (GS-X) pumps because they transport avariety of glutathione conjugates. Indeed, glutathioneS-transferase (GST)-like proteins have been shown to berequired for vacuolar sequestration of pigments in maize(ZmBz2), petunia (PhAN9) and Arabidopsis (AtTT19)[10–12]. Because anthocyanin–glutathione conjugate(s)have not been found, it was proposed that these GSTsmight deliver their flavonoid substrate directly, withoutglutathionation, to the transporter [13].
AtTT12 of Arabidopsis encodes a protein of the MATE(multidrug and toxic compound extrusion) group oftransporters that is required for vacuolar localization ofproanthocyanidins in the testa of the seed [14]. AlthoughMATE transporters are not known to require glutathion-ated substrates, the vacuolar localization of proantho-cyanidins does require the GST-like protein AtTT19 [12].Proanthocyanidin precursors in tt19 mutants appear toaccumulate in vesicle-like structures rather than freely inthe cytoplasm, suggesting that AtTT19 does not act inmembrane transport as such but in subsequent transportof the vesicles to the large vacuole [12]. In tomato, anAtTT12-like membrane transporter and a PhAN9/AtTT19-like GST have been identified that are co-regulated with other structural anthocyanin genes and
Review TRENDS in Plant Science Vol.10 No.5 May 2005
. doi:10.1016/j.tplants.2005.03.002
TRENDS in Plant Science
Phenylalanine 4-Coumaroyl-CoA
CHS
CHI
F3H
Chalcone
Naringenin
Dihydrokaempferol
Dihydromyricetin
DelphinidinPelargonidin
Dihydroquercitin
Cyanidin
Vacuolar anthocyanin
Leucocyanidin LeucodelphinidinLeucopelargonidin
Flavan 4-ol
LARCatechin
Polymerization and transport
e.g. 5GTs, RTs, ATs, MTs
Decorated anthocyanin
DFRDFR
DFR
DFR
LDOX LDOXLDOX
GST
Flavonols
Transporter
EpicatechinANR
GST
Colorless proanthocyanidin
CE
F3′H F3′5′H
Transporter
Vacuole
Vac
uo
le
3GT 3GT3GT
Anthocyanin 3-glycoside
Phl
obap
hene
s in
cel
l wal
l
Brown proanthocyanidinderivative
Figure 1. Biosynthesis of flavonoid pigments. The basic skeleton of all flavonoids consists of three aromatic rings and is generated by the enzymes chalcone synthase (CHS)
and chalcone isomerase (CHI). Oxidation of the central ring by flavonoid 3-hydroxylase (F3H) yields a dihydroflavonol (dihydrokaempferol), which can be hydroxylated on the
3 0 or 5 0 position of the B-ring by flavonoid 3 0-hydroxylase (F3 0H) and/or flavonoid 3 05 0hydroxylase (F3 05 0H) yielding precursors of orange (pelargonidin), red–magenta
(cyanidin) and purple–mauve (delphinidin) anthocyanin pigments. Some plant species (e.g. rose and carnation) cannot make purple colors because they lack F3 05 0H and/or
F3 0H. Dihydroflavonols are converted by dihydroflavonol reductase (DFR), leucoanthocyanidin oxidase (LDOX) and a 3-glucosyl transferase (3GT) to yield an anthocyanin
3-glucoside that can be further substituted by 5-glucosyl- (5GTs), rhamnosyl- (RTs), acyl- (ATs) and/or methyltransferases (MTs), resulting in ‘decorated’ anthocyanins with
different colors. Transport of the end product to the vacuole requires a glutathione S-transferase (GST) and a specific transporter localized in the vacuolar membrane.
Proanthocyanidin (PA) synthesis starts in some species with the reduction of leucoanthocyanidin by a leucoanthocyanidin reductase (LAR) [63]. However,Arabidopsis seems
to lack a LAR gene and proanthocyanidin synthesis branches off from the anthocyanin pathway after the LDOX step via an anthocyanidin reductase encoded by AtBANYULS
(AtBAN) [46,64]. The resulting flavan 3-ols, catechin or epicatechin, are transported into the vacuole via a GST and a transporter. Polymerization is thought to take place inside
the vacuole via a condensing enzyme (CE). The following steps that result in a brown proanthocyanidin derivative are not well understood andmight rely in part on products
of the TANNIN-DEFICIENT SEED (TSD) loci from Arabidopsis [65]. The inset photographs show some examples of colored wild-type tissues (on the right) and mutant tissue
lacking pigments (on the left). Bottom right photograph: anthocyanins accumulating in the aleurone of maize kernels (left) and petunia flowers (right). Bottom left
photograph: proanthocyanidin pigments in wild-type and ttg2 mutant Arabidopsis seeds. Top left photograph: brown phlobaphene pigments in the pericarp from maize.
(Note the variegation in the right kernel owing to reversions of an unstable p allele).
Review TRENDS in Plant Science Vol.10 No.5 May 2005 237
are probably involved in vacuolar sequestration of antho-cyanins [15].
PhAN9 and AtTT19 belong to a different GST class tothat of ZmBZ2, suggesting that the GSTs were, like thetransporters (MATE of MRP-type), recruited twice inde-pendently during evolution. Because GST mutants inmaize (bz2), petunia (an9) and carnation (fl3) can becomplemented by both PhAN9 and ZmBZ2, it appears thatthese distinct GSTs fulfill similar roles [11,16]. Whether
www.sciencedirect.com
the MATE and MRP transporters are also functionallyexchangeable has not been tested and thus it cannot beexcluded that both transporters are required together.
Regulation of pigment synthesis
In most plants, pigmentation is limited to some specifictissues and is regulated by external and internal factors,such as light (see e.g. [17,18]) and the clock [19]. Most ofthe regulation of pigment synthesis occurs via the

TRENDS in Plant Science
TTG1
GL3/EGL3
TT8
TT2 GL1/WER/
MYB23
MYB61
AN11
AN1
JAF13
AN2/AN4 PH4
PAC
PA Anthocyanin Hairs Mucilage PA Anthocyanin pH Seedcoat Anthocyanin Phlobaphene
B/R
C1/PL P
TTG2 TTG2
TT1
TT16
Arabidopsis (At) Petunia (Ph) Maize (Zm)
MYBx
PAC
PAP1 or 2
CPC/TRY
WD40
HLH1
HLH2
R2R3-MYB
WRKY
Zn-finger
MADS
R3-MYB
Figure 2.Regulatory genes controlling pigment synthesis inArabidopsis, petunia andmaize. The diagrams depict the involvement of various types of transcription regulators
(indicated on the left) in the synthesis of anthocyanin, proanthocyanidin (PA) and phlobaphene pigments. The bars indicate the genes that encode these factors in
Arabidopsis, petunia and maize. The pigmentation regulates several additional processes: hair development and seed mucilage production in Arabidopsis and in petunia
vacuolar acidification (pH) andmorphogenesis of epidermal cells in the seed coat. If the function of a regulator is inferred froma loss-of-function phenotype it is indicated by a
colored bar, otherwise it is indicated by a colorless bar. Note that the color coding corresponds to that in Figure 3. Abbreviations of Arabidopsis (At) gene names and
references: AtTTG1 [49] and AtTTG2 [43], TRANSPARENT TESTA GLABRA1 and 2; AtTT2, TRANSPARENT TESTA2; AtPAP1 or AtPAP2 [52], PRODUCTION ANTHCYANIN
PIGMENT1 or 2; AtGL1, GLABROUS1; AtMYB23 [66]; AtWER, WEREWOLF; AtTT8 [34]; GL3 and EGL3, ENHANCER OF GLABROUS3 [29]; AtTT16 [42]; AtTT1 [41]; AtCPC,
CAPRICE; AtTRY, TRIPTYCHON [67]. Abbreviations of petunia (Ph) gene names and references: PhAN11, ANTHOCYANIN11 [27]; PhAN2, ANTHOCYANIN2 [5]; PhAN1 [21];
PhJAF13 [36]; PhPH4; PhMYBx [62] [A. Kroon (2004) PhD thesis, Vrije Universiteit, Amsterdam]. Abbreviations of maize (Zm) gene names and references: ZmPAC, PALE
ALEURONE COLOR [50]; C1, COLOURLESS1; ZmPL, PURPLE LEAF; ZmR, RED; ZmB, BOOSTER; P, PERICARP COLOR [1].
TRENDS in Plant Science
R2R3-MYB
WD40
HLH
HLH
HIPDFR
?
R3-MYB
Figure 3. Model depicting the role of MYB, HLH and WD40 regulators in
transcription activation of a structural pigmentation gene (DFR). The various
proteins (circles) are color coded as in Figure 2. Genes, including upstream
sequences are indicated by rectangles. (Some) of the MYBs control transcription of
(some) of the HLH factors, and subsequently form a complex that also involves the
WD40 protein. Whether the WD40 protein is also present in the transcription
complex on the promoter of the structural gene (DFR) is unclear, indicated by the
questionmark. TheMYB factors contact the DNA directly, whereas the HLH proteins
probably bind indirectly via a hypothetical HLH INTERACTING PROTEIN (HIP) [25].
The small R3-MYB acts as an inhibitor, probably by sequestration of the HLH
protein into an inactive complex.
Review TRENDS in Plant Science Vol.10 No.5 May 2005238
coordinated transcriptional control of the structuralgenes [1]. In addition, some genes are subject to post-transcriptional control [20], but this has not been properlyinvestigated for most genes. Via mutation analysis,numerous regulators have been identified that controltranscription of the structural genes inArabidopsis, maizeand petunia (Figure 2).
Role of MYB, HLH and WD40 proteins in transcription
regulation
In all species analyzed to date, the common denominatorsin the regulation of structural anthocyanin and proantho-cyanidin genes are transcription factors with MYB orhelix–loop–helix (HLH) domains and a WD40 protein(Figure 2). Yeast two hybrid assays indicate that theseproteins can mutually interact, indicating that they arepart of one transcription activation pathway (Figure 3).Experiments in which the activity of these regulatorscould be controlled post-translationally – by fusing themto the ligand-binding domain of the glucocorticoid receptor– indicated that they activate transcription of structuralgenes directly, without synthesis of ‘intermediate’ regu-lators [21,22]. Although the MYB ZmC1 can bind toregulatory elements in the promoters of structuralanthocyanin genes, it is insufficient for transcriptionactivation, and even when the DNA-binding activity ofZmCI is enhanced, its HLH partner (ZmR or ZmB)remains essential [23–26]. The HLH regulators have notbeen shown to bind DNA but might activate transcriptionby recruiting an unknown factor that binds to a second cis-element promoter of the anthocyanin gene (Figure 3) andby freeing ZmC1 from an inhibitory protein [25].
The WD40 regulators are highly conserved, even inorganisms that cannot synthesize anthocyanins (i.e. algae,fungi and animals); for example, the human homolog,HsAN11, can at least in part replace PhAN11 in functional
www.sciencedirect.com
assays [27]. However, the precise molecular function ofthese WD40s is still unclear. Arabidopsis ttg1 and petuniaan11 mutants can be (partially) rescued by highexpression of their HLH or MYB partners, suggesting

Review TRENDS in Plant Science Vol.10 No.5 May 2005 239
that the WD40 somehow activates the downstream MYBand HLH protein complex post-translationally [27–29]. Inpetunia petals, most of the PhAN11 protein is in thecytosol [27] and the Perilla homolog PfWD is also localizedto the cytosol when expressed in onion cells. However,when co-expressed with its HLH partner (PfMYC), somePfWD protein enters the nucleus, which suggests that itcould be directly involved in transcription activation [30].When expressed in yeast, AtTTG1 boosts transcriptiondriven by a MYB (AtTT2) and HLH protein (AtTT8). Theaddition of a transcription activation domain to AtTTG1enhanced its effect, suggesting that AtTTG1 is present inthe transcription complex [22]. However, other explana-tions cannot be ruled out completely because localizationand folding were not analyzed because of the effect onprotein stability. By contrast, human HsAN11 was foundin a protein kinase complex that controls the activity ofglycogen synthase by phosphorylation, suggesting a rolein signal transduction [31].
Based on the observations summarized above, simplemodels have been proposed for the activation of structuralpigmentation genes (Figure 3); similar models have beenput forward for MYB, HLH and WD proteins controllinghair formation in Arabidopsis [32,33] (Figure 2). Thesemodels are perhaps better seen as working hypothesesand are likely to undergo many modifications in the nearfuture. One potential problem is that Figure 3 assumes a1:1:1 stoichiometry for MYB, HLH and WD40 proteins. Inyeast, the HLH proteins can form dimers [29], not throughthe HLH domain as one might expect but via a regionjust downstream [A. Kroon (2004) PhD thesis, VrijeUniversiteit, Amsterdam], and some MYBs (AtTT2) canform homodimers [22]. Thus, in vivo the transcriptioncomplex might include multiple HLH and MYB proteins.
Based on their sequences, the HLH-type regulatorsgroup into multiple phylogenetic clades; one includesPhJAF13, AmDELILA, ZmLC and ZmR, whereas PhAN1and AtTT8 are in a distinct clade [21,34]. The pigmenta-tion MYBs might also constitute two distinct clades, repre-sented by the pairs AtPAP1–PhAN2 and AtTT2–ZmC1,respectively [35]. In functional assays, MYBs and HLHs ofdistinct clades are often exchangeable [21,36,37], but thehigh expression levels that are generally used can concealsubstantial differences (see e.g. Refs [38,39]). Nevertheless,some MYBs and HLHs display clear functional differences,evenwhen overexpressed. For example: (i)Arabidopsis ttg1mutants can be complemented by (over)expression of theHLH factors ZmR [28], AtGL3 and AtEGL3 [29] butnot AtTT8 [35]; (ii) the HLH PhAN1 can boost the activityof ZmP, a MYB regulator of phlobaphene synthesis inpetunia cells, but ZmR and PhJAF13 cannot [21].Furthermore, the phenotype of an1 mutants shows thatPhJAF13 cannot replace PhAN1 in vivo, even though bothare co-expressed [21,36].
These findings can be explained in two ways that arenot mutually exclusive. One possibility is that the in vivotranscription complex includes multiple MYB and HLHproteins that differ in function and evolutionary origin.The other possibility is that the transcription complexcontains one member of each class and that duringevolution multiple MYBs and HLHs were recruited
www.sciencedirect.com
independently to fulfill the same function. It is noteworthythat a single amino acid change can turn the HLH factorAtATR2, a regulator of tryptophan synthesis, into aninducer of pigmentation [40].
Other activators
The synthesis of proanthocyanidins in Arabidopsis seedsrequires the induction of DFR, ANR (encoded by AtBAN)and the transporter AtTT12 by the MYB AtTT2 and theHLH AtTT8 [34,35]. By analyzing Arabidopsis mutants,several more regulatory proteins have been identified thatcontrol proanthocyanidin synthesis, including transcrip-tion factors with a MADS box (AtTT16), a Zn-finger(AtTT1), or a WRKY domain (AtTTG2) [41–43] (Figure 2).At present, it is unclear if and how these factors cooperatewith the MYB, HLH and WD proteins. AtTT16 andAtTT1 are required for proanthocyanidin synthesis in aparticular region of the testa only but do not controlpigmentation in other tissues [41,42,44]. Moreover, intt1 mutants, AtBAN expression is down regulated butAtDFR and the earlier structural genes remain normallyexpressed [41]. Thus, if TT1 directly cooperates withMYB, HLH and WD40 proteins, it does so only for theactivation of AtBAN and not AtDFR. The action ofAtTT16 and TTG2 also seems confined to genes that actlate in proanthocyanidin synthesis, after the split fromthe anthocyanin pathway, but detailed molecular dataare currently lacking [42,44].
Differential activation of distinct pigmentation
pathways
In spite of the plethora of regulatory genes andmutants identified, it is still largely unclear how acell determines which class of pigment it synthesizes.In part, this happens by the competition of distinctpathways for a common substrate. For example, loss ofANR activity and proanthocyanidin synthesis enhancesanthocyanin accumulation in Arabidopsis seeds [45,46].Differential transcription of structural genes in distinctbranches of the pathway is likely to play a major role,but this has only been studied in maize, not in petuniaor Arabidopsis.
In the aleurone of maize seeds, the structuralanthocyanin genes are activated by a MYB (ZmC1),WD40 (ZmPAC) and HLH (ZmR or ZmB) triad, whereasphlobaphenes are synthesized in the pericarp, and thechaff of the maize cob under the control of the MYBZmP (Figure 2). Probably, ZmP cooperates with a WD40partner, encoded by ZmPAC and/or a paralogous gene[47], but no HLH partner is known. ZmP and ZmC1bind to the same sites in the DFR (ZmA1) promoter butZmC1 needs an HLH partner, ZmR or ZmB, for tran-scription activation, whereas ZmP does not [23,25,26,48].However, the 3GT gene (ZmBZ1), which is required foranthocyanin synthesis only, cannot be activated byZmP because it lacks ZmP binding sites but it stillresponds to the combination of ZmC1 and ZmR or ZmB[25]. Thus, the specificity of ZmC1 and ZmP isdetermined by their capability to interact with HLHpartners in combination with cis-elements in specifictarget promoters [25].

Review TRENDS in Plant Science Vol.10 No.5 May 2005240
Regulation of the regulators
In general, the WD40 regulators are expressed more orless ubiquitously [27,49,50], whereas the expression of theMYB and HLH factors and consequently the structuralgenes is limited to pigmented tissues [1]. To date, only afew of the factors that control the expression of theregulatory genes have been identified and a comprehen-sive view is still lacking.
The transcription factor AtFUSCA3 (AtFUS3) regu-lates a variety of processes during Arabidopsis embryo-genesis, in part by limiting the expression domain ofAtTTG1 [51], but it is unclear how (in)direct thisregulation is.
Several observations indicate that (some of) the identi-fied MYBs have a dual function: (i) as a direct activator ofstructural genes, together with a HLH partner; and (ii) asan activator of the gene(s) encoding the HLH regulator(Figure 1). This explains why PhAN2, AtTT2, AtPAP1 orAtPAP2 alone are sufficient for ectopic activation ofstructural genes in transgenic plants [35,36,52], whereasin transient assays – which measure in the short timespan only relatively direct interactions – an HLH factormust be co-expressed from a separate gene construct[36,37]. Constitutive expression of PhAN2 and AtTT2 intransgenic plants activates mRNA expression of theirHLH partners (PhAN1 and AtTT8), but because an2 andtt2 loss-of-function mutations do not alter HLH geneexpression, the biological relevance of this finding wasunclear [21,35].
In petunia, PhAN4 is required for expression of PhAN1,but not PhJAF13, in anthers [21]. Because an4 mutantscan be fully complemented by ectopic expression ofPhAN2, PhAN4 was thought to encode a similar MYBprotein [21,36]. Recent experiments have shown thatPhAN4 does indeed encode a MYB protein with highsimilarity to PhAN2, providing conclusive evidence for arole of (some) of the MYBs as regulators of the HLH genes[A.Kroon (2004)PhDthesis,VrijeUniversiteit,Amsterdam,see http://www.bio.vu.nl/vakgroepen/genetica/Research_projects/Projects_Francesca.htm).
PhMYBx of petunia encodes a single repeat MYBprotein that can bind the HLHs PhAN1 and PhJAF13 inyeast cells. It is structurally similar to AtCPC and AtTRYand acts as an inhibitor because constitutive expression ofPhMYBx down-regulated anthocyanin synthesis appar-ently by sequestering PhAN1 into inactive complexes[A. Kroon (2004) PhD thesis, Vrije Universiteit, Amster-dam], similar to the action of some dominant negativemutant alleles of ZmC1 [53]. Because PhMYBx mRNAexpression is partially down regulated in an1 and an11petals, it might be part of an auto-regulatory loop thatmodulates PhAN1 activity. However, why PhAN1 activityneeds to be modulated is unclear because overexpressionof PhAN1 does not result in any obvious alterations [21].
In maize, distinct alleles of ZmP are found thatcondition pigmentation of either the pericarp (P-rw), thechaff of the cob (P-wr) or both (P-rr). The introduction inmaize plants of P-rr and P-wr (hybrid) promoter-cDNAconstructs resulted in all cases in a P-rr phenotype (redpericarp and red cob). In subsequent generations, thesetransgenics switched at a low frequency from P-rr to a
www.sciencedirect.com
P-wr phenotype, which correlated with increased methyl-ation of the transgene [54]. Thus, P-wr is controlled atleast in part by an epigenetic mechanism, which might besimilar to that inactivating ZmPL in some (epi)alleles [55].Whereas P-rr contains a single MYB gene, the P-wr allelecontains six tandemly repeated gene copies, suggestingthat this complex structure is required for epigeneticsilencing in the pericarp [56,57].
Integration into the regulatory web
We previously proposed that the anthocyanin-specificbranch of flavonoid metabolism originates from the birthof new structural genes that were placed under the controlof pre-existing regulators [36,58]. The strong conservationof the WD40 regulator even in mammals showed that(some of) the regulators are much older than the pathwayitself, and therefore might co-regulate other (older)processes. It is now well established that besides pigmen-tation the WD40 and HLH factors also regulate severalother seemingly unrelated processes by associating withMYB proteins with a more specific function. Curiously, theadditional processes that have been identified to dateseem limited to small groups of species and therefore seemto represent relatively new ‘acquisitions’ during evolution.
In Arabidopsis, in addition to controlling pigmentsynthesis, TTG1, TTG2, Gl3 and EGL3 also control theformation of mucilage in the seed and the development ofhairs on the aerial plant body. In addition, ttg1 and ttg2mutants and the gl3 egl3 double mutant form ectopic roothairs, indicating that in roots the wild-type genes promotea non-hair (atrichoblast) cell fate [33]. For the specifica-tion of (non)-hair cell fate, TTG1, GL3 and EGL3 interactwith specific MYB transcription activators, WEREWOLF(AtWER) and AtGL1, in a manner that closely resemblesthe activation of pigmentation genes [29,33,59,60]. Theformation of seed mucilage requires yet another MYBprotein (AtMYB61), but it is not fully (dis)proven that itfunctions in the AtTTG1–AtGL3–AtEGL3 pathway [61].Strikingly, in petunia or maize, neither KO mutations norectopic expression of any of the pigmentation regulatorsaffects trichome formation in their host [27,50], eventhough expression of ZmPAC or ZmR readily rescues hairdefects in Arabidopsis ttg1 mutants [28,50].
In petunia, PhAN1 and PhAN11 affect multipleprocesses in the seed coat epidermis [39]. After fertiliza-tion, ovules develop into seeds and increase several-fold insize. During expansion of the seed coat, the epidermal cellsdo not divide but increase several-fold in size and acquirea brown color, presumably because of proanthocyanidinaccumulation. In an1 and an11 seeds, the epidermal cellsare yellow (blocked proanthocyanidin synthesis); insteadof growing as large as wild-type epidermal cells, theydivide one or two times. Whether the primary effect hereof PhAN1 and PhAN11 is to suppress cell division, tostimulate cell growth, or both, is unclear. In petals,PhAN1, PhAN2 and PhAN11 activate, besides anthocya-nin synthesis, yet another pathway that leads to acidi-fication of the vacuole [39]. Some mutations in PhAN1down-regulate acidification but have little or no effect onanthocyanin synthesis, suggesting that PhAN1 activatesboth pathways by different mechanisms [39]. The PhPH4

Review TRENDS in Plant Science Vol.10 No.5 May 2005 241
gene is required for vacuolar acidification but not foranthocyanin synthesis [1,62], and encodes a MYB proteinthat interacts with PhAN1 and PhJAF13 [A. Kroon (2004)PhD thesis, Vrije Universiteit, Amsterdam]. Thus, inpetunia also, the WD40 and HLH regulators seem tocontrol distinct processes by interacting with different,more specific, MYB proteins.
Concluding remarks and future directions
The identification of a large plethora of regulatory genesshows that the regulation of a simple biological featuresuch as pigment synthesis is regulated by an unexpectedlycomplex network that is intimately linked with the controlof seemingly unrelated pathways (such as hair develop-ment and vacuolar acidification). In general, the WD40and HLH regulators seem the most pleiotropic in effectand involved in multiple processes; they hook up withdifferent MYB proteins that act more specifically (lesspleiotropic) to activate specific pathways.
Given the many possible interactions and the highfunctional similarity of the many transcription regulatorsidentified, it will be difficult to unravel the precise roles ofeach protein using the genetic methods applied to date.The composition of (mixtures of) transcription complexesdepends on protein concentrations and mutual affinitiesand these are difficult to reconcile by expressing thesefactors in yeast or plant cells but rather require thebiochemical analysis of complexes formed in vivo. Arabi-dopsis seeds or hairs are not easily amenable to suchanalyses but alternative systems such as petunia petals orvarious maize tissues provide an excellent alternative.
In general, applying knowledge from model plants inother (crop) species requires some understanding of theevolution (and thereby diversification) of the processesthat are being addressed. The ongoing analysis of theregulation of pigmentation in diverse species is likely toprovide new molecular insights into the evolution ofregulatory networks. It is particularly intriguing howthe regulation of pigmentation became linked to suchvaried processes as hair development and vacuolaracidification in distinct species, and the extent to whichthese processes are similar at the molecular level.
AcknowledgementsResearch in our laboratory was supported by grants from the NetherlandsTechnology Foundation (STW) and the Netherlands Organization forScientific Research (NWO).
References
1 Mol, J. et al. (1998) How genes paint flowers and seeds. Trends PlantSci. 3, 212–217
2 Winkel-Shirley, B. (2002) Biosynthesis of flavonoids and effects ofstress. Curr. Opin. Plant Biol. 5, 218–223
3 Bradshaw, H.D. and Schemske, D.W. (2003) Allele substitution at aflower colour locus produces a pollinator shift in monkeyflowers.Nature 426, 176–178
4 Brown, K. (2002) Floral evolution. A compromise on floral traits.Science 298, 45–46
5 Quattrocchio, F. et al. (1999) Molecular analysis of the anthocyanin2gene of Petunia and its role in the evolution of flower color. Plant Cell11, 1433–1444
6 Saslowsky, D. and Winkel-Shirley, B. (2001) Localization of flavonoidenzymes in Arabidopsis roots. Plant J. 27, 37–48
www.sciencedirect.com
7 Winkel-Shirley, B. (2001) Flavonoid biosynthesis. A colorful model forgenetics, biochemistry, cell biology, and biotechnology. Plant Physiol.126, 485–493
8 Grotewold, E. (2004) The challenges of moving chemicals within andout of cells: insights into the transport of plant natural products.Planta 219, 906–909
9 Goodman, C.D. et al. (2004) A multidrug resistance-associatedprotein involved in anthocyanin transport in Zea mays. Plant Cell
16, 1812–182610 Marrs, K.A. et al. (1995) A glutathione S-transferase involved in
vacuolar transfer encoded by the maize gene Bronze-2. Nature 375,397–400
11 Alfenito, M.R. et al. (1998) Functional complementation of antho-cyanin sequestration in the vacuole by widely divergent glutathioneS-transferases. Plant Cell 10, 1135–1149
12 Kitamura, S. et al. (2004) TRANSPARENT TESTA 19 is involved inthe accumulation of both anthocyanins and proanthocyanidins inArabidopsis. Plant J. 37, 104–114
13 Mueller, L.A. et al. (2000) AN9, a petunia glutathione S-transferaserequired for anthocyanin sequestration, is a flavonoid-bindingprotein. Plant Physiol. 123, 1561–1570
14 Debeaujon, I. et al. (2001) The TRANSPARENT TESTA12 gene ofArabidopsis encodes a multidrug secondary transporter-like proteinrequired for flavonoid sequestration in vacuoles of the seed coatendothelium. Plant Cell 13, 853–871
15 Mathews, H. et al. (2003) Activation tagging in tomato identifies atranscriptional regulator of anthocyanin biosynthesis, modification,and transport. Plant Cell 15, 1689–1703
16 Larsen, E.S. et al. (2003) A carnation anthocyanin mutant iscomplemented by the glutathione S-transferases encoded by maizeBz2 and petunia An9. Plant Cell Rep. 21, 900–904
17 Piazza, P. et al. (2002) Members of the c1/pl1 regulatory gene familymediate the response of maize aleurone and mesocotyl to differentlight qualities and cytokinins. Plant Physiol. 128, 1077–1086
18 Procissi, A. et al. (1997) Light-dependent spatial and temporalexpression of pigment regulatory genes in developing maize seeds.Plant Cell 9, 1547–1557
19 Harmer, S.L. et al. (2000) Orchestrated transcription of key pathwaysin Arabidopsis by the circadian clock. Science 290, 2110–2113
20 Pairoba, C.F. and Walbot, V. (2003) Post-transcriptional regulationof expression of the Bronze2 gene of Zea mays L. Plant Mol. Biol.
53, 75–8621 Spelt, C. et al. (2000) anthocyanin1 of petunia encodes a basic-helix
loop helix protein that directly activates structural anthocyaningenes. Plant Cell 12, 1619–1631
22 Baudry, A. et al. (2004) TT2, TT8, and TTG1 synergistically specify theexpression of BANYULS and proanthocyanidin biosynthesis inArabidopsis thaliana. Plant J. 39, 366–380
23 Sainz, M. et al. (1997) Evidence for direct activation on ananthocyanin promoter by the maize C1 protein and comparison ofDNA binding by related Myb-domain proteins. Plant Cell 9, 611–625
24 Lesnick, M.L. and Chandler, V.L. (1998) Activation of the maizeanthocyanin gene a2 is mediated by an element conserved in manyanthocyanin promoters. Plant Physiol. 117, 437–445
25 Hernandez, J.M. et al. (2004) Different mechanisms participate in theR-dependent activity of the R2R3 MYB transcription factor C1.J. Biol. Chem. 279, 48205–48213
26 Grotewold, E. et al. (2000) Identification of the residues in the Mybdomain of maize C1 that specify the interaction with the bHLHcofactor R. Proc. Natl. Acad. Sci. U. S. A. 97, 13579–13584
27 de Vetten, N. et al. (1997) The an11 locus controlling flowerpigmentation in petunia encodes a novel WD-repeat protein conservedin yeast, plants and animals. Genes Dev. 11, 1422–1434
28 Lloyd, A.M. et al. (1992) Arabidopsis and Nicotiana anthocyaninproduction activated by maize regulators R and C1. Science 258,1773–1775
29 Zhang, F. et al. (2003) A network of redundant bHLH proteinsfunctions in all TTG1-dependent pathways of Arabidopsis. Develop-ment 130, 4859–4869
30 Sompornpailin, K. et al. (2002) A WD-repeat-containing putativeregulatory protein in anthocyanin biosynthesis in Perilla frutescens.Plant Mol. Biol. 50, 485–495

Review TRENDS in Plant Science Vol.10 No.5 May 2005242
31 Skurat, A.V. and Dietrich, A.D. (2004) Phosphorylation of Ser640 inmuscle glycogen synthase by DYRK family protein kinases. J. Biol.Chem. 279, 2490–2498
32 Szymanski, D.B. et al. (2000) Progress in the molecular geneticanalysis of trichome initiation and morphogenesis in Arabidopsis.Trends Plant Sci. 5, 214–219
33 Pesch, M. and Hulskamp, M. (2004) Creating a two-dimensionalpattern de novo during Arabidopsis trichome and root hair initiation.Curr. Opin. Genet. Dev. 14, 422–427
34 Nesi, N. et al. (2000) The TT8 gene encodes a basic helix–loop–helixdomain protein required for expression of DFR and BAN genes inArabidopsis siliques. Plant Cell 12, 1863–1878
35 Nesi, N. et al. (2001) TheArabidopsis TT2 gene encodes an R2R3MYBdomain protein that acts as a key determinant for proanthocyanidinaccumulation in developing seed. Plant Cell 13, 2099–2114
36 Quattrocchio, F. et al. (1998) Analysis of bHLH and MYB-domainproteins: species-specific regulatory differences are caused by diver-gent evolution of target anthocyanin genes. Plant J. 13, 475–488
37 Zimmermann, I.M. et al. (2004) Comprehensive identification ofArabidopsis thaliana MYB transcription factors interacting withR/B-like BHLH proteins. Plant J. 40, 22–34
38 Liu, Y. et al. (1998) Molecular consequences of Ds insertion into andexcision from the helix–loop–helix domain of the maize R gene.Genetics 150, 1639–1648
39 Spelt, C. et al. (2002) ANTHOCYANIN1 of petunia controls pigmentsynthesis, vacuolar pH, and seed coat development by geneticallydistinct mechanisms. Plant Cell 14, 2121–2135
40 Smolen, G.A. et al. (2002) Dominant alleles of the basic helix–loop–helix transcription factor ATR2 activate stress-responsive genes inArabidopsis. Genetics 161, 1235–1246
41 Sagasser, M. et al. (2002) A. thaliana TRANSPARENT TESTA 1 isinvolved in seed coat development and defines the WIP subfamily ofplant zinc finger proteins. Genes Dev. 16, 138–149
42 Nesi, N. et al. (2002) The TRANSPARENT TESTA16 locus encodesthe ARABIDOPSIS BSISTER MADS domain protein and is requiredfor proper development and pigmentation of the seed coat. Plant Cell14, 2463–2479
43 Johnson, C.S. et al. (2002) TRANSPARENT TESTA GLABRA2, atrichome and seed coat development gene of Arabidopsis, encodes aWRKY transcription factor. Plant Cell 14, 1359–1375
44 Debeaujon, I. et al. (2003) Proanthocyanidin-accumulating cells inArabidopsis testa: regulation of differentiation and role in seeddevelopment. Plant Cell 15, 2514–2531
45 Albert, S. et al. (1997)BANYULS, a novel negative regulator offlavonoidbiosynthesis in the Arabidopsis seed coat. Plant J. 11, 289–299
46 Xie, D.Y. et al. (2003) Role of anthocyanidin reductase, encoded byBANYULS in plant flavonoid biosynthesis. Science 299, 396–399
47 Selinger, D.A. and Chandler, V.L. (1999) A mutation in the palealeurone color1 gene identifies a novel regulator of the maizeanthocyanin pathway. Plant Cell 11, 5–14
48 Pooma, W. et al. (2002) Transposon insertions in the promoter of theZea mays a1 gene differentially affect transcription by the Myb factorsP and C1. Genetics 161, 793–801
49 Walker, A.R. et al. (1999) The TRANSPARENT TESTA GLABRA1
Reproduction of material
Interested in reproducing part or all of an article published by ElGlobal Rights Department with details of how and where the req
on-line, plea
http://www.elsevier.com/wps/find/obtainpermi
Alternatively, ple
ElseviGlobal Rights D
PO BoxOxford OX5
Phone: (+44) 18Fax: (+44) 186
permissions@e
www.sciencedirect.com
locus, which regulates trichome differentiation and anthocyaninbiosynthesis in Arabidopsis, encodes a WD40 repeat protein. PlantCell 11, 1337–1350
50 Carey, C.C. et al. (2004) Mutations in the pale aleurone color1regulatory gene of the Zea mays anthocyanin pathway have distinctphenotypes relative to the functionally similar TRANSPARENTTESTA GLABRA1 gene in Arabidopsis thaliana. Plant Cell 16,450–464
51 Tsuchiya, Y. et al. (2004) The FUS3 transcription factor functionsthrough the epidermal regulator TTG1 during embryogenesis inArabidopsis. Plant J. 37, 73–81
52 Borevitz, J.O. et al. (2000) Activation tagging identifies a con-served MYB regulator of phenylpropanoid biosynthesis. Plant Cell12, 2383–2394
53 Chen, B. et al. (2004) Ectopic expression of a c1-I allele from maizeinhibits pigment formation in the flower of transgenic tobacco. Mol.Biotechnol. 26, 187–192
54 Cocciolone, S.M. et al. (2001) Tissue-specific patterns of a maize Mybtranscription factor are epigenetically regulated. Plant J. 27, 467–478
55 Hoekenga, O.A. et al. (2000) Developmental patterns of chromatinstructure and DNA methylation responsible for epigenetic expressionof a maize regulatory gene. Genetics 155, 1889–1902
56 Chopra, S. et al. (1998) A maize Myb homolog is encoded by amulticopy gene complex. Mol. Gen. Genet. 260, 372–380
57 Chopra, S. et al. (2003) The maize Unstable factor for orange1 is adominant epigenetic modifier of a tissue specifically silent allele ofpericarp color1. Genetics 163, 1135–1146
58 Koes, R.E. et al. (1994) The flavonoid biosynthetic pathway in plants:function and evolution. BioEssays 16, 123–132
59 Payne, C.T. et al. (2000) GL3 encodes a bHLH protein that regulatestrichome development in Arabidopsis through interaction with GL1and TTG1. Genetics 156, 1349–1362
60 Bernhardt, C. et al. (2003) The bHLH genes GLABRA3 (GL3) andENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in theArabidopsis root. Development 130, 6431–6439
61 Penfield, S. et al. (2001) MYB61 is required for mucilage depositionand extrusion in the Arabidopsis seed coat. Plant Cell 13, 2777–2791
62 van Houwelingen, A. et al. (1998) Analysis of flower pigmentationmutants generated by random transposon mutagenesis in Petuniahybrida. Plant J. 13, 39–50
63 Tanner, G.J. et al. (2003) Proanthocyanidin biosynthesis in plants.Purification of legume leucoanthocyanidin reductase and molecularcloning of its cDNA. J. Biol. Chem. 278, 31647–31656
64 Abrahams, S. et al. (2003) The Arabidopsis TDS4 gene encodesleucoanthocyanidin dioxygenase (LDOX) and is essential forproanthocyanidin synthesis and vacuole development. Plant J. 35,624–636
65 Abrahams, S. et al. (2002) Identification and biochemical character-ization of mutants in the proanthocyanidin pathway in Arabidopsis.Plant Physiol. 130, 561–576
66 Kirik, V. et al. (2001) Ectopic expression of the Arabidopsis AtMYB23gene induces differentiation of trichome cells. Dev. Biol. 235, 366–377
67 Schellmann, S. et al. (2002) TRIPTYCHON and CAPRICE mediatelateral inhibition during trichome and root hair patterning inArabidopsis. EMBO J. 21, 5036–5046
from Elsevier articles
sevier, or one of our article figures? If so, please contact ouruested material will be used. To submit a permission requestse visit:
ssionform.cws_home/obtainpermissionform
ase contact:
erepartment800,1DX, UK.65-8438305-853333lsevier.com
Related Documents











