504 Transactions of the American Fisheries Society 133:504–514, 2004 q Copyright by the American Fisheries Society 2004 Fish Size and Prey Availability Affect Growth, Survival, Prey Selection, and Foraging Behavior of Larval Yellow Perch BRIAN D. S. GRAEB* 1 AND JOHN M. DETTMERS Center for Aquatic Ecology, Illinois Natural History Survey; and Department of Natural Resources and Environmental Sciences, University of Illinois, 607 East Peabody Drive, Champaign, Illinois 61820, USA DAVID H. WAHL Center for Aquatic Ecology, Illinois Natural History Survey; and School of Integrative Biology, University of Illinois, 505 South Goodwin Avenue, Urbana, Illinois 61821, USA CARLA E. CA ´ CERES School of Integrative Biology, University of Illinois, 505 South Goodwin Avenue, Urbana, Illinois 61821, USA Abstract.—Zooplankton availability is strongly linked with the growth, survival, and ultimately recruitment of fish during their early life history. We examined how different types of zooplankton affected the growth, survival, and prey selection of larval yellow perch Perca flavescens in a series of laboratory experiments. The growth and survival of newly hatched (5–7-mm) to 12-mm yellow perch larvae was greatest when feeding on adult copepods and copepod nauplii. The growth and survival of yellow perch larvae longer than 12 mm was greatest when feeding on adult copepods and small cladocerans. Prey selection patterns closely followed trends in growth and survival; smaller larvae positively selected adult and naupliar copepods, whereas larger larvae selected adult copepods and small cladocerans. Foraging behavior experiments conducted with larvae longer than 12 mm revealed that these fish derived similar energetic gains when feeding solely on adult copepods and small cladocerans. The pathway to this energetic benefit differed substantially be- tween prey types. Adult copepods yielded substantial energy to larval yellow perch because of the minimal handling time involved, despite the lower capture efficiency than with cladocerans. Conversely, yellow perch larvae realized high energetic gain from small cladocerans because of high capture efficiency, despite the higher handling times than with adult copepods. These results illustrate the importance of experimentally quantifying the feeding behavior of fish larvae to gain insight into how larval fish behavior and food type interact to shape larval fish growth patterns. Understanding the mechanisms influencing re- cruitment has long been a goal in fisheries ecology. Of interest from a management perspective is the ability to predict year-class strength and set har- vest regulations accordingly. Furthermore, iden- tifying recruitment mechanisms will improve our understanding of community structure and species distributions (Bremigan and Stein 1994). Recruit- ment success in fish is often determined during their early life history (Rice et al. 1987) because most fish experience very high mortality during this life stage (Houde 1994). Although numerous mortality sources act on the early life stages of fish, predation and starvation are generally viewed * Corresponding author: brianp[email protected] 1 Present address: Department of Wildlife and Fish- eries Sciences, South Dakota State University, Brook- ings, South Dakota 57007, USA. Received February 27, 2003; accepted August 31, 2003 as the two most ubiquitous ones (Houde 1987; Miller et al. 1988). Both predation and starvation are size dependent (Miller et al. 1988) and strongly interact with growth rate, such that higher growth rates may reduce starvation and predation (Houde 1987; Miller et al. 1990) whereas lower growth rates increase mortality from these sources. Thus, understanding the factors influencing growth dur- ing the early life history of fish is important to understanding recruitment and ultimately to achieving better fishery management. Growth in larval fishes is strongly influenced by the availability of appropriate food items (Welker et al. 1994). Many larval fish rely on small zoo- plankton as prey due to limited gape width (Bre- migan and Stein 1994; DeVries et al. 1998) and reduced visual acuity (Wahl et al. 1993). However, different species of zooplankton differ markedly in morphology and behavior. For example, cla- docerans and copepods (two groups commonly ob-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

504
Transactions of the American Fisheries Society 133:504–514, 2004q Copyright by the American Fisheries Society 2004
Fish Size and Prey Availability Affect Growth, Survival, PreySelection, and Foraging Behavior of Larval Yellow Perch
BRIAN D. S. GRAEB*1 AND JOHN M. DETTMERS
Center for Aquatic Ecology, Illinois Natural History Survey;and Department of Natural Resources and Environmental Sciences,
University of Illinois, 607 East Peabody Drive, Champaign, Illinois 61820, USA
DAVID H. WAHL
Center for Aquatic Ecology, Illinois Natural History Survey;and School of Integrative Biology, University of Illinois,
505 South Goodwin Avenue, Urbana, Illinois 61821, USA
CARLA E. CACERES
School of Integrative Biology, University of Illinois,505 South Goodwin Avenue, Urbana, Illinois 61821, USA
Abstract.—Zooplankton availability is strongly linked with the growth, survival, and ultimatelyrecruitment of fish during their early life history. We examined how different types of zooplanktonaffected the growth, survival, and prey selection of larval yellow perch Perca flavescens in a seriesof laboratory experiments. The growth and survival of newly hatched (5–7-mm) to 12-mm yellowperch larvae was greatest when feeding on adult copepods and copepod nauplii. The growth andsurvival of yellow perch larvae longer than 12 mm was greatest when feeding on adult copepodsand small cladocerans. Prey selection patterns closely followed trends in growth and survival;smaller larvae positively selected adult and naupliar copepods, whereas larger larvae selected adultcopepods and small cladocerans. Foraging behavior experiments conducted with larvae longerthan 12 mm revealed that these fish derived similar energetic gains when feeding solely on adultcopepods and small cladocerans. The pathway to this energetic benefit differed substantially be-tween prey types. Adult copepods yielded substantial energy to larval yellow perch because ofthe minimal handling time involved, despite the lower capture efficiency than with cladocerans.Conversely, yellow perch larvae realized high energetic gain from small cladocerans because ofhigh capture efficiency, despite the higher handling times than with adult copepods. These resultsillustrate the importance of experimentally quantifying the feeding behavior of fish larvae to gaininsight into how larval fish behavior and food type interact to shape larval fish growth patterns.
Understanding the mechanisms influencing re-cruitment has long been a goal in fisheries ecology.Of interest from a management perspective is theability to predict year-class strength and set har-vest regulations accordingly. Furthermore, iden-tifying recruitment mechanisms will improve ourunderstanding of community structure and speciesdistributions (Bremigan and Stein 1994). Recruit-ment success in fish is often determined duringtheir early life history (Rice et al. 1987) becausemost fish experience very high mortality duringthis life stage (Houde 1994). Although numerousmortality sources act on the early life stages offish, predation and starvation are generally viewed
* Corresponding author: [email protected] Present address: Department of Wildlife and Fish-
eries Sciences, South Dakota State University, Brook-ings, South Dakota 57007, USA.
Received February 27, 2003; accepted August 31, 2003
as the two most ubiquitous ones (Houde 1987;Miller et al. 1988). Both predation and starvationare size dependent (Miller et al. 1988) and stronglyinteract with growth rate, such that higher growthrates may reduce starvation and predation (Houde1987; Miller et al. 1990) whereas lower growthrates increase mortality from these sources. Thus,understanding the factors influencing growth dur-ing the early life history of fish is important tounderstanding recruitment and ultimately toachieving better fishery management.
Growth in larval fishes is strongly influenced bythe availability of appropriate food items (Welkeret al. 1994). Many larval fish rely on small zoo-plankton as prey due to limited gape width (Bre-migan and Stein 1994; DeVries et al. 1998) andreduced visual acuity (Wahl et al. 1993). However,different species of zooplankton differ markedlyin morphology and behavior. For example, cla-docerans and copepods (two groups commonly ob-
Carol.Jacobson
Text Box
383-F

505LARVAL YELLOW PERCH GROWTH AND FORAGING
served in the diets of larval fish) have differentswimming patterns and morphology. Copepods arecylindrical with a fairly uniform body width andswim along a smooth path (calanoid copepods) orin a ‘‘hop-and-sink’’ manner (cyclopoid copepods)(Kerfoot et al. 1980). Both of these copepods swimfaster and are more evasive than the sphericallyshaped cladocerans, which move by small jumpsthrough the water column. Furthermore, capturesuccess is generally higher for fish feeding on cla-docerans than for fish feeding on copepods (Dren-ner et al. 1978; Mills et al. 1984). Thus, zooplank-ton of similar lengths may provide variable en-ergetic benefits to larval fish based on the effi-ciency with which the fish can capture them.
Despite the importance of larval fish growth andsurvival in determining eventual recruitment,many studies that have experimentally quantifiedthe relationship between food availability and thegrowth, survival, prey selection, and feeding ecol-ogy of young freshwater fishes have focused onlarger juvenile stages (i.e., .20 mm; but see Mayerand Wahl 1997). Although studies of juvenile fishprovide important information, they may not beapplicable to larvae because of the large morpho-logical and physiological differences between lar-vae and later life stages (e.g., the former have lessgape width, visual acuity, and swimming ability).For example, laboratory experiments and fieldstudies indicated that juvenile yellow perch Percaflavescens and bluegills Lepomis macrochirus pos-itively selected large cladocerans (Werner and Hall1974; Mills et al. 1984; Bremigan and Stein 1994).Yellow perch and bluegills also experienced highgrowth and survival when these prey were abun-dant, and foraging experiments revealed that cla-docerans were energetically more favorable thanother prey items. Conversely, most fish larvae havea much smaller gape width and consume smallerzooplankton prey (such as copepods) in the field,suggesting that the role of a particular species ofzooplankton is very different for larval fish thanfor juveniles (Keast 1980; Schael et al. 1991; Fish-er and Willis 1997). However, the relationship be-tween zooplankton taxonomic composition and thegrowth, survival, prey selection, and feeding ecol-ogy of most larval fish has not been experimentallyquantified. Thus, laboratory experiments thatmechanistically examine the role of zooplanktongroup in influencing the growth, survival, and ul-timately recruitment of larval fish are needed tounderstand recruitment during this critical stage.
One reason for the paucity of studies that mech-anistically examine the role food availability plays
in larval fish growth and survival is the difficultyof conducting experiments on such a small lifestage. Larval fish experience very high naturalmortality (Houde 1994), and maintaining appro-priate prey levels requires much time and effort(Chick and Van Den Avyle 1999). Further, ob-serving larval fish during foraging experiments isdifficult because of their small size and the smallsize of their prey items (Mayer and Wahl 1997).Despite these difficulties, we investigated howzooplankton group affects the growth, survival,and prey selection of larval yellow perch amongseveral discreet size-classes of larvae.
We chose to work with larval yellow perch be-cause food availability may be a factor in the re-duced recruitment of yellow perch in Lake Mich-igan since 1989. Changes in the zooplankton as-semblage, including reduced zooplankton densityand a shift in taxonomic composition, may haveacted to decrease the growth and survival of larvalyellow perch (Francis et al. 1996). The recruitmentof larval yellow perch is thought to be affected bylower survival at low zooplankton densities (Dett-mers et al. 2003). Furthermore, yellow perch lon-ger than 20 mm positively select cladocerans,growing best when the latter are abundant (Hansenand Wahl 1981; Mills and Forney 1981; Mills etal. 1989). Little is known, however, about howzooplankton taxonomic composition affects thegrowth and survival of yellow perch shorter than20 mm. For this reason, we conducted a series oflaboratory experiments that quantified the role ofzooplankton availability in structuring larval yel-low perch growth and survival.
We began by quantifying the growth and sur-vival of four size-classes of larval yellow perchfeeding on different types of zooplankton (i.e., cla-docerans, copepods, and rotifers). We then con-ducted prey selection experiments to determinewhether larval yellow perch chose prey items thatconferred the best growth. Finally, we quantifiedthe foraging efficiencies of yellow perch larvae todetermine the energetic costs of different zoo-plankton groups and to help explain trends ingrowth, survival, and prey selection.
Methods
We conducted experiments on four size-classesof larval yellow perch—newly hatched (5–7 mm),small (7–12 mm), medium (12–16 mm), and large(.16 mm)—to account for important ontogeneticchanges that occur during early life history, suchas first feeding and swim bladder inflation. Be-cause mortality for these sizes can be high, pre-

506 GRAEB ET AL.
liminary work suggested that the best approachwould be to conduct a series of independent ex-periments for each size-class of larvae. This designallowed us to characterize the ontogenetic stagesin which the preferred zooplankton prey are im-portant to growth and survival.
Yellow perch egg skeins collected from LakeMichigan during late May and early June 2001were hatched and larvae reared on a diet of brineshrimp Artemia spp. and mixed zooplankton in lab-oratory facilities at the Lake Michigan BiologicalStation, Zion, Illinois. All experiments were con-ducted under controlled laboratory conditions,with a photoperiod of 12 h light: 12 h dark and awater temperature of 18.9 6 0.18C (mean 6 SE).The zooplankton used in our experiments were cul-tured on site but supplemented with zooplanktonfrom Lake Michigan as needed. Rotifers Brachion-us spp. were cultured separately. To establish zoo-plankton treatments, we separated cladocerans andadult copepods from copepod nauplii using 153-mm-mesh sieves.
Growth and survival.—To determine the effectof zooplankton group on larval yellow perchgrowth and survival, we conducted experimentsusing the following treatments replicated fivetimes each: cladocerans, adult copepods, copepodnauplii, rotifers, and a foodless control. Commontaxa in these groups included rotifers Brachionusspp., cladocerans Ceriodaphnia and Bosmina spp.,and cyclopoid and calanoid copepods (both adultsand nauplii). Lengths for these groups were as fol-lows: cladocerans, 0.55 6 0.02 mm (n 5 110);copepods, 0.98 6 0.02 mm (n 5 109); copepodnauplii, 0.18 6 0.004 mm (n 5 98); and rotifers,0.21 6 0.01 mm (n 5 30). Rotifers were includedonly in experiments with newly hatched larvae be-cause such prey were never observed in the dietsof larvae during a pilot study of yellow perch preyselection (Graeb, unpublished data). In contrast,rotifers have been observed in the diets of smalllarval yellow perch in the field (Whiteside et al.1985). Thus, 25 aquaria (our experimental unit)were used for the newly hatched yellow perch size-class, and 20 aquaria were used for the small, me-dium, and large size-classes.
Yellow perch from each size-class (n 5 150–200 newly hatched larvae, 75 small larvae, 5 me-dium larvae, and 1 large larva) were held in 38-L aquaria with randomly assigned zooplanktontreatments. The number of larvae used in each rep-licate changed according to the size-class involved(smaller larvae had higher mortality, requiringmore larvae per replicate) and availability (normal
mortality resulted in fewer fish being available atlarger size-classes). The treatment densities of thezooplankton were maintained at or above 75 in-dividuals/L; we considered this food level to bead libitum for larval yellow perch based on theasymptote of a functional response for larval wall-eyes Sander vitreus (formerly Stizostedion vitreum)consuming zooplankton (Johnston and Mathias1994). Treatment densities were estimated every1–3 d using a polyvinyl chloride tube sampler withan inner diameter of 47 mm. Zooplankton wereadded every 1–2 d to ensure that zooplankton den-sities remained at or above 75/L (zooplankton den-sities were rarely below 75/L).
To ensure that newly hatched larvae experiencedtheir first feeding during the experiment, we di-vided six fertilized egg skeins into approximatelyequal portions of 175 eggs and allowed them tohatch in randomly assigned aquaria. Aquaria wereinoculated with zooplankton treatments after lar-vae hatched. The experiment started at 2 d post-hatch to allow for partial (but not complete) yolksac absorption. Prior observations showed thatnewly hatched larvae begin exogenous feedingshortly before the yolk sac is completely absorbed.Initial sizes were determined by either sacrificinglarvae at 2 d posthatch (newly hatched larvae) orby sub-sampling 50 individuals at the start of eachexperiment (larger larvae). Up to five larvae weresacrificed daily to determine growth in experi-ments with newly hatched and small larvae be-cause these size-classes experienced high mortal-ity. The duration of the experiment was 6 d forthe newly hatched size-class, 9 d for the small size-class, and 10 d for both the medium and large size-classes. Analysis of variance (ANOVA) was usedto analyze average growth rate (mm/d), and Tu-key’s honestly significant difference test was usedto separate treatment means.
The survival of yellow perch larvae was mea-sured concomitantly with growth. Surviving lar-vae were counted every morning 2 h after the startof the light period using a small, narrow-beamflashlight. Life tables were generated from thesedata. Survival functions were then fitted to eachsize-class and treatment combination (fish that weremoved were censored). Cumulative survivorshipwas analyzed among treatments by means of Wil-coxon chi-square tests. If at least one treatmentdiffered from the others, we separated the survivalcurves (cumulative survivorship) using the co-variance matrix from the Wilcoxon statistics tocalculate Z-scores for individual pairwise com-

507LARVAL YELLOW PERCH GROWTH AND FORAGING
parison (Fox 1993). Pairwise comparisons wereconducted at an overall significance level of 0.05.
Prey selection.—Yellow perch were starved forat least 12 h and then introduced into 38-L glassaquaria that were inoculated with equal densities(50/L each) of cladocerans, adult copepods, co-pepod nauplii, and rotifers (the latter for newlyhatched larvae only). Black plastic was attachedto three sides of all aquaria; fluorescent lightingwas mounted above the aquaria to increase preycontrast. Larvae were allowed to feed for 1 h andthen euthanatized and preserved in a 95% solutionof ethanol. Equal prey densities were chosen togive each yellow perch an equal opportunity (bynumber) to consume a given prey item. Up to fivelarvae were used per trial for the newly hatchedand small size-classes because of the prevalenceof empty stomachs among these size-classes; onelarva was present during each trial with mediumand large larvae. We conducted 10 feeding trialsfor newly hatched larvae, 72 for small larvae, 8for medium larvae, and 17 for large larvae. Thenumber of replicate trials varied across size-classesbased on the availability of appropriately sizedyellow perch. More trials were conducted onsmaller larvae to overcome the high occurrence ofempty stomachs that we observed in larvae lessthan 12 mm. Digestive tracts were removed forenumeration and measurement of prey items witha dissecting microscope and digitizing tablet. Preyselectivity was estimated by calculating Chesson’s(1983) coefficient of selectivity,
r /ni ia 5 ,m
(r /n )O i ii51
where ri is the proportion of food type i in thepredator diet, ni is the proportion of food type i inthe environment, and m is the number of prey typesavailable. Selection coefficients were calculatedfor each fish, but mean values were pooled for eachtrial. For each size-class, mean selection coeffi-cients and 95% confidence intervals were com-pared with random feeding (1/m) to determine preyselectivity. We concluded that there was positiveselectivity if the 95% confidence intervals wereabove the random-feeding line, neutral selectivityif the 95% confidence intervals overlapped the ran-dom-feeding line, and negative selectivity if theywere below the random-feeding line.
Feeding behavior.—We quantified capture effi-ciency and handling time for medium and largelarvae feeding on cladocerans and copepods. At-
tempts to observe smaller larvae and smaller preyitems such as copepod nauplii were unsuccessfulbecause we could not discern successful captures.A single yellow perch was placed in a 4-L rect-angular feeding arena blackened on three sides.After an acclimation time of 1.5 h, 10 prey items(copepods or cladocerans) were introduced into thearena. Strikes, captures, and handling times werethen observed for 30 min. Five replicate trials wereconducted on each zooplankton group and size-class combination, for a total of 20 trials. Captureefficiency (the number of captures per strike) wasaveraged for each trial. Handling time (the timerequired to begin active searching after a captureevent) was recorded for the first capture event onlyto avoid interactions between handling times andgut fullness. Capture efficiency was analyzed witha two-factor (larva size and zooplankton group)ANOVA; handling time was analyzed with theWilcoxon rank sums test because these data werenonnormal.
To quantify more explicitly the energetic benefitof each zooplankton group to perch larvae, wedetermined the relative benefits (calories) andcosts (handling time and capture efficiency) forthe medium and large larval yellow perch feedingon cladocerans and copepods. We first determinedthe average length (6SE) of the zooplankton groupchosen by the two size-classes of larvae from theselection experiments (n 5 approximately 50 preyitems from the diets of randomly chosen fish). Forthe medium larvae, cladoceran length was 0.73 60.05 mm and copepod length 0.61 6 0.08 mm; forthe large larvae, cladoceran length was 0.94 60.06 mm and copepod length 0.59 6 0.03 mm.Then we converted these lengths into biomassbased on length–weight regressions from Bottrellet al. (1976). Using our estimated biomass, wedetermined the number of calories per individualzooplankton based on caloric densities from Cum-mins and Wuycheck (1970). These caloric valueswere combined with the average capture efficiencyand handling times to estimate the number of cal-ories gained per strike and the number of caloriesgained per second of handling time by mediumand large yellow perch larvae feeding on cladoc-erans and copepods. Finally, we compared the es-timates of net energetic gain using a two-factor(size and prey type) ANOVA. Treatments wereseparated using least-square means and Bonferroniadjustments when needed.
ResultsGrowth
Zooplankton group influenced the growth of yel-low perch larvae across all size-classes. Newly
508 GRAEB ET AL.
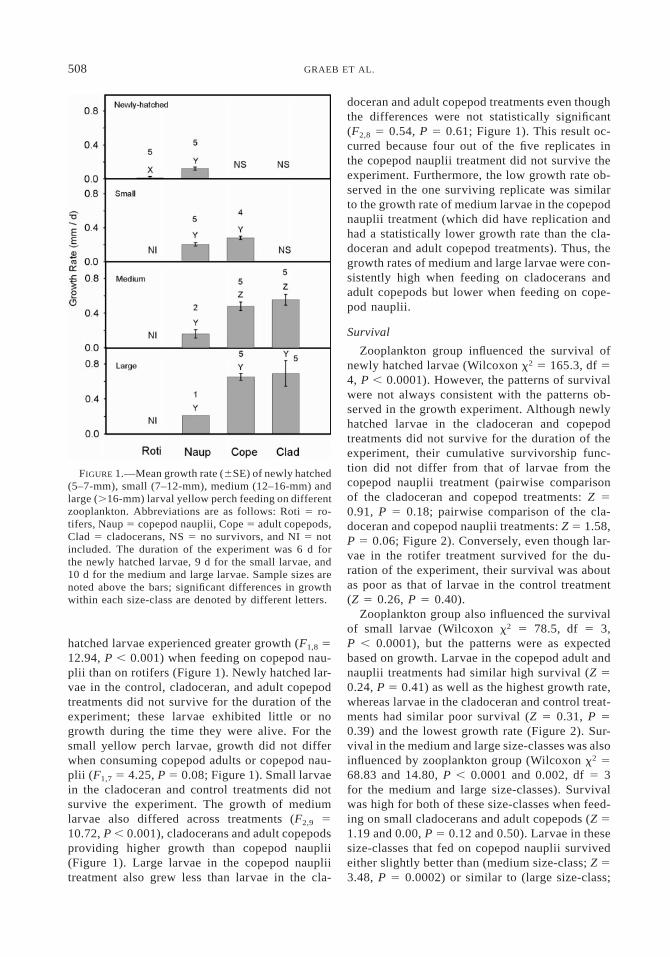
FIGURE 1.—Mean growth rate (6SE) of newly hatched(5–7-mm), small (7–12-mm), medium (12–16-mm) andlarge (.16-mm) larval yellow perch feeding on differentzooplankton. Abbreviations are as follows: Roti 5 ro-tifers, Naup 5 copepod nauplii, Cope 5 adult copepods,Clad 5 cladocerans, NS 5 no survivors, and NI 5 notincluded. The duration of the experiment was 6 d forthe newly hatched larvae, 9 d for the small larvae, and10 d for the medium and large larvae. Sample sizes arenoted above the bars; significant differences in growthwithin each size-class are denoted by different letters.
hatched larvae experienced greater growth (F1,8 512.94, P , 0.001) when feeding on copepod nau-plii than on rotifers (Figure 1). Newly hatched lar-vae in the control, cladoceran, and adult copepodtreatments did not survive for the duration of theexperiment; these larvae exhibited little or nogrowth during the time they were alive. For thesmall yellow perch larvae, growth did not differwhen consuming copepod adults or copepod nau-plii (F1,7 5 4.25, P 5 0.08; Figure 1). Small larvaein the cladoceran and control treatments did notsurvive the experiment. The growth of mediumlarvae also differed across treatments (F2,9 510.72, P , 0.001), cladocerans and adult copepodsproviding higher growth than copepod nauplii(Figure 1). Large larvae in the copepod naupliitreatment also grew less than larvae in the cla-
doceran and adult copepod treatments even thoughthe differences were not statistically significant(F2,8 5 0.54, P 5 0.61; Figure 1). This result oc-curred because four out of the five replicates inthe copepod nauplii treatment did not survive theexperiment. Furthermore, the low growth rate ob-served in the one surviving replicate was similarto the growth rate of medium larvae in the copepodnauplii treatment (which did have replication andhad a statistically lower growth rate than the cla-doceran and adult copepod treatments). Thus, thegrowth rates of medium and large larvae were con-sistently high when feeding on cladocerans andadult copepods but lower when feeding on cope-pod nauplii.
Survival
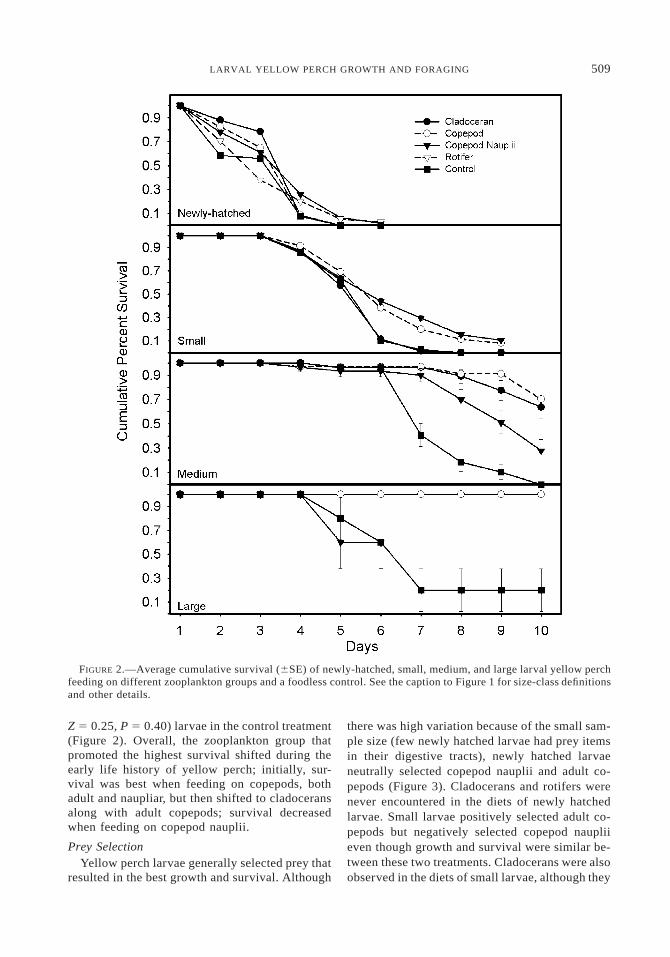
Zooplankton group influenced the survival ofnewly hatched larvae (Wilcoxon x2 5 165.3, df 54, P , 0.0001). However, the patterns of survivalwere not always consistent with the patterns ob-served in the growth experiment. Although newlyhatched larvae in the cladoceran and copepodtreatments did not survive for the duration of theexperiment, their cumulative survivorship func-tion did not differ from that of larvae from thecopepod nauplii treatment (pairwise comparisonof the cladoceran and copepod treatments: Z 50.91, P 5 0.18; pairwise comparison of the cla-doceran and copepod nauplii treatments: Z 5 1.58,P 5 0.06; Figure 2). Conversely, even though lar-vae in the rotifer treatment survived for the du-ration of the experiment, their survival was aboutas poor as that of larvae in the control treatment(Z 5 0.26, P 5 0.40).
Zooplankton group also influenced the survivalof small larvae (Wilcoxon x2 5 78.5, df 5 3,P , 0.0001), but the patterns were as expectedbased on growth. Larvae in the copepod adult andnauplii treatments had similar high survival (Z 50.24, P 5 0.41) as well as the highest growth rate,whereas larvae in the cladoceran and control treat-ments had similar poor survival (Z 5 0.31, P 50.39) and the lowest growth rate (Figure 2). Sur-vival in the medium and large size-classes was alsoinfluenced by zooplankton group (Wilcoxon x2 568.83 and 14.80, P , 0.0001 and 0.002, df 5 3for the medium and large size-classes). Survivalwas high for both of these size-classes when feed-ing on small cladocerans and adult copepods (Z 51.19 and 0.00, P 5 0.12 and 0.50). Larvae in thesesize-classes that fed on copepod nauplii survivedeither slightly better than (medium size-class; Z 53.48, P 5 0.0002) or similar to (large size-class;
509LARVAL YELLOW PERCH GROWTH AND FORAGING
FIGURE 2.—Average cumulative survival (6SE) of newly-hatched, small, medium, and large larval yellow perchfeeding on different zooplankton groups and a foodless control. See the caption to Figure 1 for size-class definitionsand other details.
Z 5 0.25, P 5 0.40) larvae in the control treatment(Figure 2). Overall, the zooplankton group thatpromoted the highest survival shifted during theearly life history of yellow perch; initially, sur-vival was best when feeding on copepods, bothadult and naupliar, but then shifted to cladoceransalong with adult copepods; survival decreasedwhen feeding on copepod nauplii.
Prey SelectionYellow perch larvae generally selected prey that
resulted in the best growth and survival. Although
there was high variation because of the small sam-ple size (few newly hatched larvae had prey itemsin their digestive tracts), newly hatched larvaeneutrally selected copepod nauplii and adult co-pepods (Figure 3). Cladocerans and rotifers werenever encountered in the diets of newly hatchedlarvae. Small larvae positively selected adult co-pepods but negatively selected copepod naupliieven though growth and survival were similar be-tween these two treatments. Cladocerans were alsoobserved in the diets of small larvae, although they

510 GRAEB ET AL.
FIGURE 3.—Mean prey selection by larval yellowperch in different size-classes as determined by Ches-son’s alpha (see text for definition). When the 95% con-fidence interval (vertical line) lies above the random-feeding (dashed) line, there is positive selection for thatprey; when it lies below the line there is negative se-lection; and when it crosses the line there is neutralselection. The random-feeding line for the newlyhatched larvae is different from that for the other larvalsize-classes because four prey groups were available fornewly hatched larvae but only three for the others. Sam-ple sizes (i.e., the number of larvae with diet items inthe stomach) are noted above the size-classes.
FIGURE 4.—Handling time (i.e., the time from suc-cessful capture of the prey to the resumption of search-ing) and percent capture efficiency (number of capturesper strike 3 100) for medium (12-mm) and large (16-mm) yellow perch larvae foraging on cladocerans andcopepods. Significant differences are denoted by differ-ent letters.
were negatively selected. Medium larvae exhibitedneutral selection for both adult copepods and cla-docerans but negatively selected copepod nauplii.Large larvae positively selected adult copepodsand neutrally selected cladocerans. Overall, adultcopepods were neutrally or positively selectedacross all yellow perch sizes. Selection for cope-pod nauplii decreased with increased larval fishsize, whereas selection for small cladocerans in-creased.
Feeding Behavior
Capture efficiency by medium and large larvaewas higher for cladocerans than for copepods (F1,16
5 17.97, P , 0.01; Figure 4) but did not differbetween the two size-classes (F1,16 5 1.76, P 50.20). Handling time for cladocerans by both themedium (4.7 s) and large size-classes (4.4 s) wasconsistently higher than for copepods (,1 s formedium larvae and undetectable (recorded as 0 s)for large larvae; F1,16 5 30.76, P , 0.01; Figure4). Thus, the higher capture efficiency and higher
handling time for cladocerans than for copepodssuggest that these two prey items provide similarenergetic benefits. The net energetic gain (caloriesconsumed per unit of cost) also supported the ideathat cladocerans and copepods provided similarenergetic benefits. The net energy gained during asingle capture event was similar for cladoceransand copepods when both capture efficiency andhandling time were considered. Calories gainedper second of handling time was higher for co-pepods than for cladocerans (F1,16 5 14.41, P 50.02; Figure 5). These differences were similaracross size-classes (main effect of larva size: F1,16
5 3.17, P 5 0.09). Calories gained per strike wassignificantly affected by the interaction of larvasize and prey type (F1,16 5 27.69, P , 0.0001).Yellow perch larvae in the medium size-class feed-ing on cladocerans gained more calories per strikethan large larvae feeding on cladocerans and bothsize-classes feeding on copepods (t 5 4.63–6.84,df 5 16, P 5 ,0.001–0.0017; Figure 5). Largelarvae feeding on cladocerans gained similar num-bers of calories per strike as medium (t 5 1.6,

511LARVAL YELLOW PERCH GROWTH AND FORAGING
FIGURE 5.—Net energetic gain for medium (12-mm)and large (16-mm) yellow perch larvae feeding on cla-docerans and copepods. Mean calories per second ofhandling time was calculated by estimating the averagebiomass and caloric density of each zooplankton taxonand dividing by the average handling time. Mean cal-ories per strike was calculated by dividing the averagenumber of calories in cladocerans and copepods by theaverage number of strikes. Significant differences withineach panel are denoted by different letters; N 5 5 forall replicates.
df 5 16, P 5 0.77) and large (t 5 2.21, df 5 16,P 5 0.25) larvae feeding on copepods. Taken to-gether, the higher net energetic gain per unit han-dling time for copepods was generally balancedout by higher net energetic gain per strike whilefeeding on cladocerans. This tradeoff resulted insimilar net energetic gains for both prey types.
Discussion
Our results demonstrate that zooplankton groupinfluences the growth and survival of larval yellowperch. Small larvae (,12 mm) positively selectedcopepods (both adult and naupliar) and experi-enced the highest growth when feeding on thisgroup. A diet shift occurred around 12 mm, whenlarvae began consuming cladocerans along withadult copepods but avoided copepod nauplii.These larger larvae also experienced enhanced
growth and survival when feeding on cladoceransand adult copepods than on copepod nauplii. Thesimilarities in the growth and survival of larvae12 mm or longer when feeding on cladocerans andadult copepods, and their willingness to consumeboth of these groups, was explained by the similarnet energetic gains derived from these prey types.Behaviorally, the high capture efficiency and highhandling time for larvae consuming cladoceranscontrasted with the low capture efficiency and lowhandling time when consuming copepods yet re-sulted in similar net energetic gains between thesetwo prey types.
Growth is one of the most important processesdetermining recruitment during the early life his-tory of fish (e.g., Crowder et al. 1987; Rice et al.1987). Furthermore, fish tend to select prey itemsthat optimize their growth and survival (Wernerand Hall 1974). Therefore, the patterns we ob-served during our experiments provide insight intohow the availability of zooplankton influences therecruitment of yellow perch during the larval pe-riod. The growth of larval yellow perch less than12 mm long should be high when copepods (bothadult and naupliar) are abundant. This finding alsoagrees with patterns observed in the field, wherecopepods dominated the diet of small yellow perch(Keast 1980; Whiteside et al. 1985; Fisher andWillis 1997) and were positively selected through-out the larval period (Schael et al. 1991). Yellowperch larvae up to12 mm did not grow or survivewell when feeding on rotifers, nor did they posi-tively select them. Similarly, rotifers were avoidedby larval yellow perch from Green Bay, LakeMichigan (Bremigan et al. 2003). However, roti-fers are a diverse group of freshwater inverte-brates; the poor growth and avoidance that we ob-served for Brachionus spp. may not represent lar-val yellow perch preference for all rotifer genera.Cladocerans were also less important than cope-pods for yellow perch larvae smaller than 12 mm,probably because the foraging costs of feeding oncladocerans were high relative to those for cope-pods. Handling times were always much greaterfor cladocerans than for copepods in the case oflarvae exceeding 12 mm; these costs were offsetby the higher capture efficiency for cladocerans.The cost of high handling time may not be offsetfor larvae smaller than 12 mm because reducedvisual acuity decreases capture efficiency (Wahl etal. 1993). In addition to the greater foraging costsassociated with cladocerans, their digestibilitymay be lower than that of copepods (Confer andLake 1987). Thus, the high foraging costs of cla-

512 GRAEB ET AL.
docerans and the total avoidance of rotifers resultin poor growth and survival for larval yellow perchless than12 mm that feed on these groups. Becauseyellow perch larvae up to 12 mm grew and sur-vived best when consuming adult and naupliar co-pepods, we predict that the recruitment of smalllarvae will be best when copepods are abundant.
When larval yellow perch were approximately12 mm long, the zooplankton groups that conferredthe highest growth and survival shifted from co-pepod nauplii to cladocerans and adult copepods.Cladocerans and adult copepods remained impor-tant to yellow perch as they grew to 20 mm. Thistrend probably continues into the early juvenileperiod, when the growth of yellow perch is bestif large cladocerans are abundant (Mills et al.1989). Although the shift to cladoceran taxa is welldocumented for yellow perch, our results indicatethat cladocerans may become important at smallerfish sizes than previously reported if present atsufficiently high densities. Previous research in-dicates that larger yellow perch experience an on-togenetic diet shift from copepods to cladoceransaround 20–30 mm in systems in which the dom-inant cladocerans are large-bodied (1.0–2.5-mm)Daphnia spp. (Mills et al. 1987; Schael et al.1991). As a result, growth and recruitment remainhigh when large cladocerans are present (Mills etal. 1984). We chose to use the smaller Bosminaand Ceriodaphnia spp. (mean size 5 0.55 mm)because they are important in many zooplanktonassemblages and were small enough to be availableto all sizes of yellow perch larvae (i.e., the larvaewere not gape limited). When smaller cladocerantaxa are available, medium and large yellow perchlarvae should experience similar growth and sur-vival when feeding on copepods and cladocerans.Although we do not dispute that Daphnia are fa-vorable for the recruitment of larger juvenile yel-low perch, cladocerans can be as important as co-pepods for sustained yellow perch growth and sur-vival at smaller sizes (i.e., 12–20 mm) in systemswith smaller cladoceran taxa.
The dietary ontogenetic shift from immature co-pepods to adult copepods and eventually cladoc-erans during the larval period of yellow perch issimilar to that of other larval fishes. For example,black crappie Pomoxis nigromaculatus, freshwaterdrum Aplodinotus grunniens, and bluegill all feedon copepod nauplii at small sizes (,8 mm) andthen progress to adult copepods and eventuallycladocerans with increasing total length and gapewidth (Keast 1980; Schael et al. 1991). Like yel-low perch, these larvae have a transition period at
approximately 12–20 mm where both cladoceransand copepods are positively selected. However,larval gizzard shad Dorosoma cepedianum contin-ue to select small zooplankton at sizes greater than13 mm (DeVries and Stein 1992), probably be-cause of the small mouth gape these larvae have(Bremigan and Stein 1994). Although we have notyet quantified it, we would expect most larval fish-es to have size-dependent patterns of selectivityand energetic gain that are similar to those wedocumented for yellow perch feeding on cladoc-erans and copepods.
Much of the focus has been placed on the roleof large-bodied cladocerans (particularly Daphnia)when examining the role of zooplankton taxa onthe growth and recruitment of young yellow perch(e.g., Mills and Forney 1981; Mills et al. 1989).This focus is based on field and laboratory workwith yellow perch exceeding 25 mm; the situationappears to be very different for larval yellowperch. Our results suggest that the availability ofcopepods and small cladocerans is an importantfactor in determining the growth and survival oflarval yellow perch. These differences in the im-portance of zooplankton group between larval andjuvenile yellow perch are probably driven by mor-phological differences in both the fish and the zoo-plankton that affect foraging efficiency. For ex-ample, although larvae longer than 16 mm are ca-pable of consuming smaller Daphnia (the gapewidth of 16-mm yellow perch is approximately 1mm, which is the size of D. pulicaria; Schael etal. 1991), the results from our foraging experi-ments suggest that the cost in terms of handlingtime would be high. As such, the large-bodied cla-docerans preferred by yellow perch longer than 20mm are probably a poor food source for smallerlarvae. Taken together, the availability of the ap-propriate size and group of zooplankton is criticalto recruitment during the entire early life historyof yellow perch, from first-feeding larvae throughthe juvenile stage.
Our findings suggest that the current composi-tion of zooplankton in Lake Michigan (particularlythe dominance of adult and naupliar copepods) isfavorable for the growth and survival of larvalyellow perch. Copepods were important to all sizesof larvae during our experiment, and we believethat the lack of small cladocerans in the currentLake Michigan assemblage (Pientka et al. 2002)will be offset by the presence of adult copepodsfor larvae longer than 12 mm. The reality in LakeMichigan, however, is a continued pattern of pooryellow perch recruitment (Shroyer and McComish

513LARVAL YELLOW PERCH GROWTH AND FORAGING
2000). Because the survival of age-0 yellow perchto fall is related to the density of zooplanktonavailable to first-feeding larvae and recent (1996–1998) densities in Lake Michigan have been 5–30/L (compared with 50–500/L during 1988–1990;Dettmers et al. 2003), the current low density isprobably a more important determinant of thegrowth and survival of larval yellow perch thanthe taxonomic composition.
Given the importance of the larval stage in fishand the general lack of knowledge about growth,survival, and foraging relationships for larval fish,we encourage more laboratory experiments thatmechanistically examine the role of food avail-ability in larval fish recruitment. Our approach ofquantifying growth, survival, and prey selectionover a range of larval fish sizes was successful inovercoming the difficulties (like high mortality)associated with conducting experiments on larvalfish. Further, our foraging behavior experimentsidentified mechanisms influencing the trends ingrowth, survival, and prey selection for larval yel-low perch. These results were markedly differentfrom those of previous studies conducted on ju-venile yellow perch, and we caution about extrap-olating the results from experiments on juvenilefish to larval life stages. Further studies that mech-anistically examine the role of zooplankton duringearly life stages will greatly increase our under-standing of fish recruitment.
Acknowledgments
This project was greatly enhanced by the sup-port of M. Simpson, A. Arce, and S. Czesny. Wethank the staff at the Lake Michigan BiologicalStation and the Kaskaskia Biological Station, Il-linois Natural History Survey, for their technicaladvice and assistance. Funding for this project wasprovided by the Federal Aid in Sport Fish Res-toration Program, Project F-123-R, administeredby the Illinois Department of Natural Resources,Illinois–Indiana Sea Grant through grant numberNA86RG0048, and the Illinois Natural HistorySurvey.
References
Bottrell, H. H., A. Duncan, Z. M. Gliwicz, E. Grygierek,A. Herzig, A. Hillbricht-Ilkowska, H. Kurasawa, P.Larsson, and T. Weglenska. 1976. A review of someof the problems in zooplankton production studies.Norwegian Journal of Zoology 24:419–456.
Bremigan, M. T., J. M. Dettmers, and A. L. Mahan. 2003.Zooplankton selectivity by larval yellow perch inGreen Bay, Lake Michigan. Journal of Great LakesResearch. 29:501–510.
Bremigan, M. T., and R. A. Stein. 1994. Gape-dependentlarval foraging and zooplankton size: implicationsfor fish recruitment across systems. Canadian Jour-nal of Fisheries and Aquatic Sciences 51:913–922.
Chesson, J. 1983. The estimation and analysis of pref-erences and its relationship to foraging models.Ecology 70:1227–1235.
Chick, J. H., and M. J. Van Den Avyle. 1999. Effectsof zooplankton spatial variation on growth of larvalstriped bass: an experimental approach. Transac-tions of the American Fisheries Society 128:339–351.
Confer, J. L., and G. J. Lake. 1987. Influence of preytype on growth of young yellow perch (Perca fla-vescens). Canadian Journal of Fisheries and AquaticSciences 44:2028–2033.
Crowder, L. B., M. E. McDonald, and J. A. Rice. 1987.Understanding recruitment of Lake Michigan fishes:the importance of size-based interactions betweenfish and zooplankton. Canadian Journal of Fisheriesand Aquatic Sciences 44(Supplement 2):141–147.
Cummins, K. W., and J. C. Wuycheck. 1970. Caloricequivalents for investigations in ecological ener-getics. Mitteilungen der Internationale Vereinigungfur Theoretische und Angewandte Limnologie 18:1–151.
Dettmers, J. M., M. J. Raffenberg, and A. K. Weis. 2003.Exploring zooplankton changes in southern LakeMichigan: implications for yellow perch recruit-ment. Journal of Great Lakes Research 29:355–364.
DeVries, D. R., M. T. Bremigan, and R. A. Stein. 1998.Prey selection by larval fishes as influenced byavailable zooplankton and gape limitation. Trans-actions of the American Fisheries Society 127:1040–1050.
DeVries, D. R., and R. A. Stein. 1992. Complex inter-actions between fish and zooplankton: quantifyingthe role of an open-water planktivore. CanadianJournal of Fisheries and Aquatic Sciences 49:1216–1227.
Drenner, R. W., J. R. Strickler, and W. J. O’Brien. 1978.Capture probability: the role of zooplankter escapein the selective feeding of planktivorous fish. Jour-nal of the Fisheries Research Board of Canada 35:1370–1373.
Fisher, S. J., and D. W. Willis. 1997. Early life historyof yellow perch in two South Dakota glacial lakes.Journal of Freshwater Ecology 12:421–429.
Fox, G. A. 1993. Failure-time analysis: emergence,flowering, survivorship, and other waiting times.Pages 253–389 in S. M. Scheiner and J. Gurevitch,editors. Design and analysis of ecological experi-ments. Chapman and Hall, New York.
Francis, J. T., S. R. Robillard, and J. E. Marsden. 1996.Essay—yellow perch management in Lake Michi-gan: a multi-jurisdictional challenge. Fisheries21(2):18–23.
Hansen, M. J., and D. H. Wahl. 1981. Selection of smallDaphnia pulex by yellow perch in Oneida Lake,New York. Transactions of the American FisheriesSociety 110:64–71.
Houde, E. D. 1987. Fish early life dynamics and re-

514 GRAEB ET AL.
cruitment variability. Pages 17–29 in R. D. Hoyt,editor. Tenth annual larval fish conference. Amer-ican Fisheries Society, Symposium 2, Bethesda,Maryland.
Houde, E. D. 1994. Differences between marine andfreshwater fish larvae: implications for recruitment.ICES Journal of Marine Science 51:91–97.
Johnston, T. A., and J. A. Mathias. 1994. Feeding ecol-ogy of walleye, Stizostedion vitreum, larvae: Effectsof body size, zooplankton abundance, and zoo-plankton community composition. Canadian Jour-nal of Fisheries and Aquatic Sciences 51:2077–2089.
Keast, A. 1980. Food and feeding relationships of youngfish in the first few weeks after beginning of ex-ogenous feeding in Lake Opinicon, Ontario. Envi-ronmental Biology of Fishes 5:305–314.
Kerfoot, W. C., D. L. Kellogg, Jr., and J. R. Strickler.1980. Visual observations of live zooplankters:evasion, escape, and chemical defenses. Pages 10–27 in W. C. Kerfoot, editor. Evolution and ecologyof zooplankton communities. University Press ofNew England, Hanover, New Hampshire.
Mayer, C. M., and D. H. Wahl. 1997. The relationshipbetween prey selectivity and growth and survivalin a larval fish. Canadian Journal of Fisheries andAquatic Sciences 54:1504–1512.
Miller, T. J., L. B. Crowder, and F. P. Binkowski. 1990.Effects of changes in the zooplankton assemblageon growth of bloater and implications for recruit-ment success. Transactions of the American Fish-eries Society 119:483–491.
Miller, T. J., L. B. Crowder, J. A. Rice, and E. A. Mar-schall. 1988. Larval size and recruitment mecha-nisms: toward a conceptual framework. CanadianJournal of Fisheries and Aquatic Sciences 45:1657–1670.
Mills, E. L., J. L. Confer, and R. C. Ready. 1984. Preyselection by young yellow perch: the influence ofcapture success, visual acuity, and prey choice.Transactions of the American Fisheries Society 113:579–587.
Mills, E. L., and J. L. Forney. 1981. Energetics, foodconsumption, and growth of young yellow perch inOneida Lake, New York. Transactions of the Amer-ican Fisheries Society 110:479–488.
Mills, E. L., M. V. Pol, R. E. Sherman, and T. B. Culver.1989. Interrelationships between prey body sizeand growth of age-0 yellow perch. Transactions ofthe American Fisheries Society 118:1–10.
Mills, E. L., D. V. Widzowski, and S. R. Jones. 1987.Food conditioning and prey selection by young yel-low perch (Perca flavescens). Canadian Journal ofFisheries and Aquatic Sciences 44:549–555.
Pientka, B., B. D. S. Graeb, and J. M. Dettmers. 2002.Yellow perch population assessment in Southwest-ern Lake Michigan, including the identification offactors that determine yellow perch year-classstrength. Illinois Natural History Survey, AquaticEcology Technical Report 02/06, Champaign.
Rice, J. A., L. B. Crowder, and M. E. Holey. 1987.Exploration of mechanisms regulating larval sur-vival in Lake Michigan bloater: a recruitment anal-ysis based on characteristics of individual larvae.Transactions of the American Fisheries Society 116:703–718.
Schael, D. M., L. G. Rudstam, and J. R. Post. 1991.Gape limitation and prey selection in larval yellowperch (Perca flavescens), freshwater drum (Aplodi-notus grunniens), and black crappie (Pomoxis ni-gromaculatus). Canadian Journal of Fisheries andAquatic Sciences 48:1919–1925.
Shroyer, S. R., and T. S. McComish. 2000. Relationshipbetween alewife abundance and yellow perch re-cruitment in southern Lake Michigan. North Amer-ican Journal of Fisheries Management 20:220–225.
Wahl, C. M., E. L. Mills, W. N. McFarland, and J. S.DeGisi. 1993. Ontogenetic changes in prey selec-tion and visual acuity of yellow perch, Perca fla-vescens. Canadian Journal of Fisheries and AquaticSciences 50:743–749.
Welker, M. T., C. L. Pierce, and D. W. Wahl. 1994.Growth and survival of larval fishes: roles of com-petition and zooplankton abundance. Transactionsof the American Fisheries Society 123:703–717.
Werner, E. E., and D. J. Hall. 1974. Optimal foragingand the size selection of prey by bluegill sunfish(Lepomis macrochirus). Ecology 55:1042–1052.
Whiteside, M. C., C. M. Swindoll, and W. L. Doolittle.1985. Factors affecting the early life history of yel-low perch, Perca flavescens. Environmental Biology12:47–56.
Related Documents











