JMBA2 - Biodiversity Records Published on-line First observation of a double tentacle bifurcation in cephalopods Ángel F. González* and Ángel Guerra ECOBIOMAR, Instituto de Investigaciones Marinas, CSIC, Eduardo Cabello nº6, 36208,Vigo,Spain. *Corresponding author, e-mail: [email protected] A female with the two tentacles bifurcated was found within a sample of 54 specimens of Moroteuthis ingens collected by a Spanish trawler fishing in the South Atlantic around the Falklands Islands dur - ing the austral spring of 2004. This is the first case of double tentacle bifurcation reported for this oegopsid squid and also for cephalopods. In spite of the lack of normal tentacles, the specimen does not show signs of malnutrition. Potential causes for this anomaly, implications to the feeding behav- iour of the animal and the regeneration process are discussed. INTRODUCTION Anatomical anomalies occasionally occur in several animal groups, including cephalopods. Howev- er, information regarding abnormalities in these marine molluscs is scarce. Among these abnormali- ties, the bifurcation of the arms is not a rare event. The first branched arm was described by Ikeda (1891), and it was later illustrated by Sasaki (1929) in Octopus vulgaris inhabiting Japanese waters. Verrill (1882) reported on the regeneration of suckers in the giant squid, Architeuthis harveyi, and Parona (1900) described and figured bifurcation in an arm of the musky octopus Eledone moschata, an additional arm in the lesser octopus E. aldovrandi (now E. cirrhosa) and a bifurcate arm in Octopus vulgaris from the Mediterranean Sea. A similar case in one specimen of the common octopus from Japanese waters was described by Smith (1907). Robson’s monograph on octopodinae (1929a) in- cludes a review of the structural abnormalities found until that time, mainly consisting in octopuses with seven or less arms. Afterwards, other cases were reported for the common octopus from the Japanese and adjacent waters (Okada, 1935, 1965a,b; Gleadal, 1989), and the anatomy of octopuses with seven arms were studied in details (Gleadal, 1989), with special reference to the arrangement of brachial nerves. Kumpf (1960) described a male specimen of Octopus briareus with the third left arm divided into two branches: it showed a well-developed web between these divisions as found at the bases of the normal arms in the species. The first case of true hexapody among the Octopoda, resulting from bilateral agenesis of one arm pair, was described by Toll & Binger (1991) for a male Pteroctopus tetracirrhus; the same authors also reported the first case of decapody in octopuses, for a male Octopus briareus and concluded that both conditions apparently result from developmental anomalies of the embryonic arm anlagen. After a survey on the anomalous conditions relating to arm development and regeneration in cephalopods, Toll & Binger (1991) suggested a possible re- lationship of polyfurcation of arm tips in the Octopoda with regenerative processes in amphibian limbs. Cases of bilateral hectocotylization were described by Robson (1929b), Palacio (1973) and Jereb et al. (1989). These, however, seem to be rare events compared to the branching of not modified arms, considering the low number of available references. Also, the phenomenon of dichotomy in the arms of Decabrachia seems to be rarer than in Octobrachia: as far as we know, there is only one case of a decapod cephalopod (Sepia esculenta) referenced to date (Okada, 1937), while four cases of anomalies in the copulatory arms in the Sepiolinae were observed by Jereb et al. (1997). To date, the only reported case on anomalies in the tentacles is that by Adam (1932), of a 92 mm mantle length (ML) female of Alloteuthis subulata caught in the Belgium waters (north-eastern Atlan- tic) in which the right and left tentacular club showed clear differences in suckers number, size and disposition. The second case is reported in the present paper, for a specimen of Moroteuthis ingens (Smith, 1881), and we think it is particularly remarkable because the anomaly (i.e. the bifurcation) affects both tentacles. Moroteuthis ingens is a large-bodied onychoteuthid, commonly growing up to 500 mm ML. Clarke (1966), based on the beaks found in the stomach contents of marine mammals, estimates a pos- sible maximum ML of 940 mm.The species is circumpolar in sub-Antarctic waters and recently was placed within the genus Onykia Lesueur, 1821 by Vecchione et al. (2003). Although that taxonomic decision could be correct we preferred to retain its previous status for reasons of comparisons with the literature. Several population units exist in the extensive area of the species distribution and there is some evidence of genetic mixing among these populations. Since adult migration of the required magni-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JMBA2 - Biodiversity RecordsPublished on-line
First observation of a double tentacle bifurcation in cephalopods
Ángel F. González* and Ángel GuerraECOBIOMAR, Instituto de Investigaciones Marinas, CSIC, Eduardo Cabello nº6, 36208, Vigo,Spain.
*Corresponding author, e-mail: [email protected]
A female with the two tentacles bifurcated was found within a sample of 54 specimens of Moroteuthis ingens collected by a Spanish trawler fishing in the South Atlantic around the Falklands Islands dur-ing the austral spring of 2004. This is the first case of double tentacle bifurcation reported for this oegopsid squid and also for cephalopods. In spite of the lack of normal tentacles, the specimen does not show signs of malnutrition. Potential causes for this anomaly, implications to the feeding behav-iour of the animal and the regeneration process are discussed.
INTRODUCTIONAnatomical anomalies occasionally occur in several animal groups, including cephalopods. Howev-
er, information regarding abnormalities in these marine molluscs is scarce. Among these abnormali-ties, the bifurcation of the arms is not a rare event. The first branched arm was described by Ikeda (1891), and it was later illustrated by Sasaki (1929) in Octopus vulgaris inhabiting Japanese waters. Verrill (1882) reported on the regeneration of suckers in the giant squid, Architeuthis harveyi, and Parona (1900) described and figured bifurcation in an arm of the musky octopus Eledone moschata, an additional arm in the lesser octopus E. aldovrandi (now E. cirrhosa) and a bifurcate arm in Octopus vulgaris from the Mediterranean Sea. A similar case in one specimen of the common octopus from Japanese waters was described by Smith (1907). Robson’s monograph on octopodinae (1929a) in-cludes a review of the structural abnormalities found until that time, mainly consisting in octopuses with seven or less arms. Afterwards, other cases were reported for the common octopus from the Japanese and adjacent waters (Okada, 1935, 1965a,b; Gleadal, 1989), and the anatomy of octopuses with seven arms were studied in details (Gleadal, 1989), with special reference to the arrangement of brachial nerves. Kumpf (1960) described a male specimen of Octopus briareus with the third left arm divided into two branches: it showed a well-developed web between these divisions as found at the bases of the normal arms in the species. The first case of true hexapody among the Octopoda, resulting from bilateral agenesis of one arm pair, was described by Toll & Binger (1991) for a male Pteroctopus tetracirrhus; the same authors also reported the first case of decapody in octopuses, for a male Octopus briareus and concluded that both conditions apparently result from developmental anomalies of the embryonic arm anlagen. After a survey on the anomalous conditions relating to arm development and regeneration in cephalopods, Toll & Binger (1991) suggested a possible re-lationship of polyfurcation of arm tips in the Octopoda with regenerative processes in amphibian limbs.
Cases of bilateral hectocotylization were described by Robson (1929b), Palacio (1973) and Jereb et al. (1989). These, however, seem to be rare events compared to the branching of not modified arms, considering the low number of available references. Also, the phenomenon of dichotomy in the arms of Decabrachia seems to be rarer than in Octobrachia: as far as we know, there is only one case of a decapod cephalopod (Sepia esculenta) referenced to date (Okada, 1937), while four cases of anomalies in the copulatory arms in the Sepiolinae were observed by Jereb et al. (1997).
To date, the only reported case on anomalies in the tentacles is that by Adam (1932), of a 92 mm mantle length (ML) female of Alloteuthis subulata caught in the Belgium waters (north-eastern Atlan-tic) in which the right and left tentacular club showed clear differences in suckers number, size and disposition. The second case is reported in the present paper, for a specimen of Moroteuthis ingens (Smith, 1881), and we think it is particularly remarkable because the anomaly (i.e. the bifurcation) affects both tentacles.
Moroteuthis ingens is a large-bodied onychoteuthid, commonly growing up to 500 mm ML. Clarke (1966), based on the beaks found in the stomach contents of marine mammals, estimates a pos-sible maximum ML of 940 mm. The species is circumpolar in sub-Antarctic waters and recently was placed within the genus Onykia Lesueur, 1821 by Vecchione et al. (2003). Although that taxonomic decision could be correct we preferred to retain its previous status for reasons of comparisons with the literature.
Several population units exist in the extensive area of the species distribution and there is some evidence of genetic mixing among these populations. Since adult migration of the required magni-
Á.F. González and Á. Guerra Double tentacle bifurcation in cephalopods
JMBA2 - Biodiversity RecordsPublished on-line
2
tude is unlikely, the most plausible explanation for this is eggs and /or larval dispersal via jet-stream currents (Sands et al., 2003). The species plays a significant role in pelagic and demersal ecosystems, being a voracious predator and prey of conspecifics, and prey of several mammals, bird and fish spe-cies (Jackson et al., 1998a). During the last decade many studies focusing on growth (Jackson, 1993, 1997), reproduction (Jackson & Mladenov, 1994; Jackson, 2001; Jackson & Jackson, 2004), distribu-tion (Jackson et al., 1998a, 2000), diet (Phillips et al., 2001, 2003; Jackson et al., 1998b) and prey status (Jackson et al., 2000a) have clarified important aspects of its biology and ecology. Adults inhabit sub-Antarctic waters, mainly the shelf (Jackson et al., 2000), from 95 to 420 m in the South Atlantic (Jackson et al., 1998a) and depths greater than 1500 m in the South Pacific (Jackson, 1997; Jackson et al., 2000). The Falkland Island population appears to be closely associated with the continental shelf bottom (100–300 m), moving offshore to deeper waters at the onset of the breeding season (Jackson et al., 1998b).
The aim of this paper is to present the first case of double tentacle bifurcation in cephalopods, and to discuss its potential causes and the implications to the feeding behaviour of the oegopsid squid Moroteuthis ingens.
MATERIALS AND METHODSA female Moroteuthis ingens with the two tentacles bifurcated was found among a sample of 54
specimens caught by a Spanish trawler fishing in the southern Atlantic around the Falklands Islands during the Austral spring of 2004. It was frozen on board and defrosted in the laboratory, at room temperature. For each individual beaks and statoliths were removed and preserved in ethanol for further analysis. Mantle length (ML), absolute length (AL=mantle length+arm length), total length (TL) and total body weight (BW) were taken. Terminal organ length and testis weight were taken for males, and ovary weight and nidamental gland length were taken for females. Maturity was de-termined by the presence of spermatophores in the terminal organ and/or in the spermatophore sac of males or the presence of mature oocytes in females. The four-levels maturity stage scale by Jackson (1997) was used to quickly assess individ-ual maturity. Stomach fullness was determined on a 6-point subjective scale (after Zuev et al., 1985 in Jackson et al., 1998b). Stomach contents were removed and weighted. Prey items were sorted and identified under dissecting microscope.
RESULTSThe main measurements of this female were:
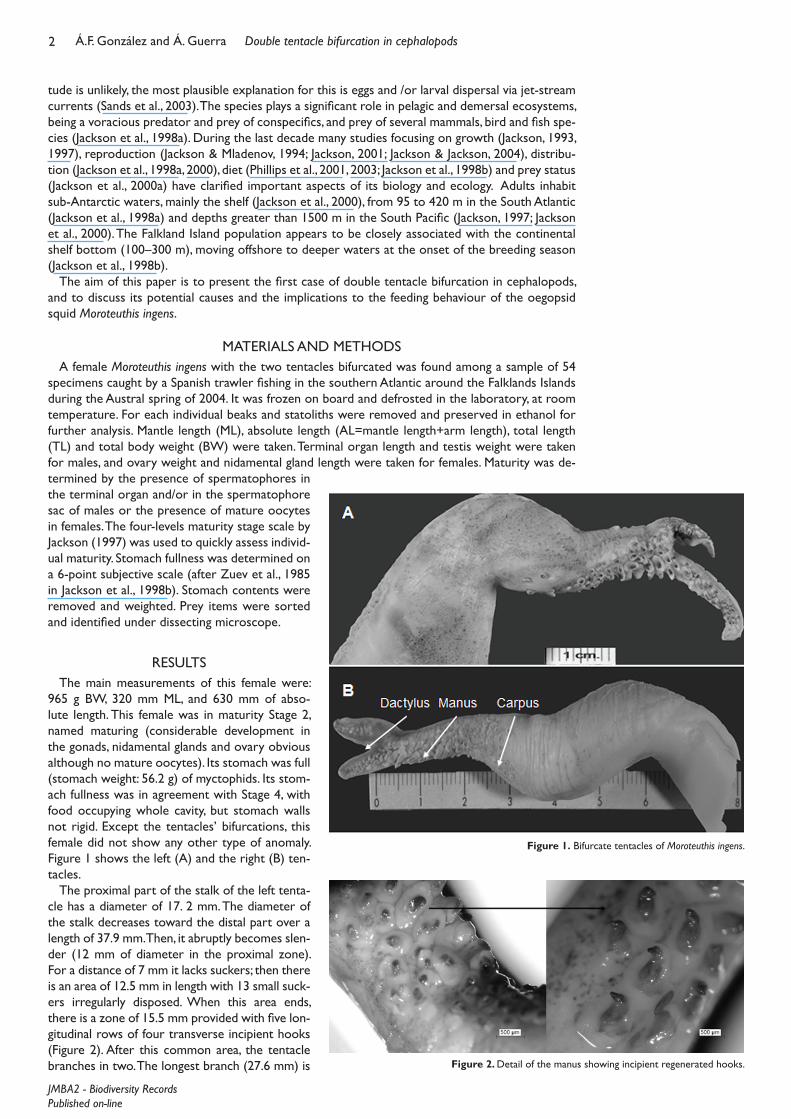
965 g BW, 320 mm ML, and 630 mm of abso-lute length. This female was in maturity Stage 2, named maturing (considerable development in the gonads, nidamental glands and ovary obvious although no mature oocytes). Its stomach was full (stomach weight: 56.2 g) of myctophids. Its stom-ach fullness was in agreement with Stage 4, with food occupying whole cavity, but stomach walls not rigid. Except the tentacles’ bifurcations, this female did not show any other type of anomaly. Figure 1 shows the left (A) and the right (B) ten-tacles.
The proximal part of the stalk of the left tenta-cle has a diameter of 17. 2 mm. The diameter of the stalk decreases toward the distal part over a length of 37.9 mm. Then, it abruptly becomes slen-der (12 mm of diameter in the proximal zone). For a distance of 7 mm it lacks suckers; then there is an area of 12.5 mm in length with 13 small suck-ers irregularly disposed. When this area ends, there is a zone of 15.5 mm provided with five lon-gitudinal rows of four transverse incipient hooks (Figure 2). After this common area, the tentacle branches in two. The longest branch (27.6 mm) is
Figure 1. Bifurcate tentacles of Moroteuthis ingens.
Figure 2. Detail of the manus showing incipient regenerated hooks.
Double tentacle bifurcation in cephalopods Á.F. González and Á. Guerra
JMBA2 - Biodiversity RecordsPublished on-line
3
provided with 15 longitudinal rows of incipi-ent hooks disposed in four transverse rows. The size and development of these hooks de-crease towards the apex. The shorter (10.4 mm) branch is provided with 12 rows of longitudinal incipient hooks disposed in two transverse rows. The size and development of these hooks also decrease towards the apex. Both branches show a relatively well devel-oped keel (Figure 1).
The proximal part of the stalk of the right tentacle shows a diameter of 16.3 mm. The diameter of the stalk decreases toward the distal part over a length of 51.1 mm. Then, it abruptly becomes slender (10.3 mm of di-ameter in the proximal zone), from a zone where a deep split is detectable. The stalk lacks suckers for an area of 10 mm length; a zone of about 10 mm in length follows, which is provided with 26–28 small suckers irregularly disposed. When this area ends, there is a zone of 21.3 mm provided with six longitudinal rows of two transverse in-cipient hooks. After this common area, the tentacle branches in two. The longest branch (12.6 mm) shows a zone of 5.6 mm without any arm structure, followed by an area with nine longitudinal rows of incipient hooks dis-posed in two transverse rows. The size and development of these hooks decrease towards the apex. The shorter branch (10.2 mm) is provided with 16 rows of longitudinal incipient hooks disposed in two transverse rows along all its length. The size and development of these hooks also decrease towards the apex. Both branches show a relatively well developed keel.
On the whole, the total length of the abnormal left tentacle is 90.0 mm, and the total length of the right tentacle is 95.0 mm. Tentacles length represent 28.1 and 29.7% respectively of the speci-men ML (320 mm). Based on the 37 specimens in the sample (ML between 125 and 420 mm) the percentage of the normal tentacles was estimated to range between 164.7 and 220.2 with respect to ML, depending on the sex and maturation stage.
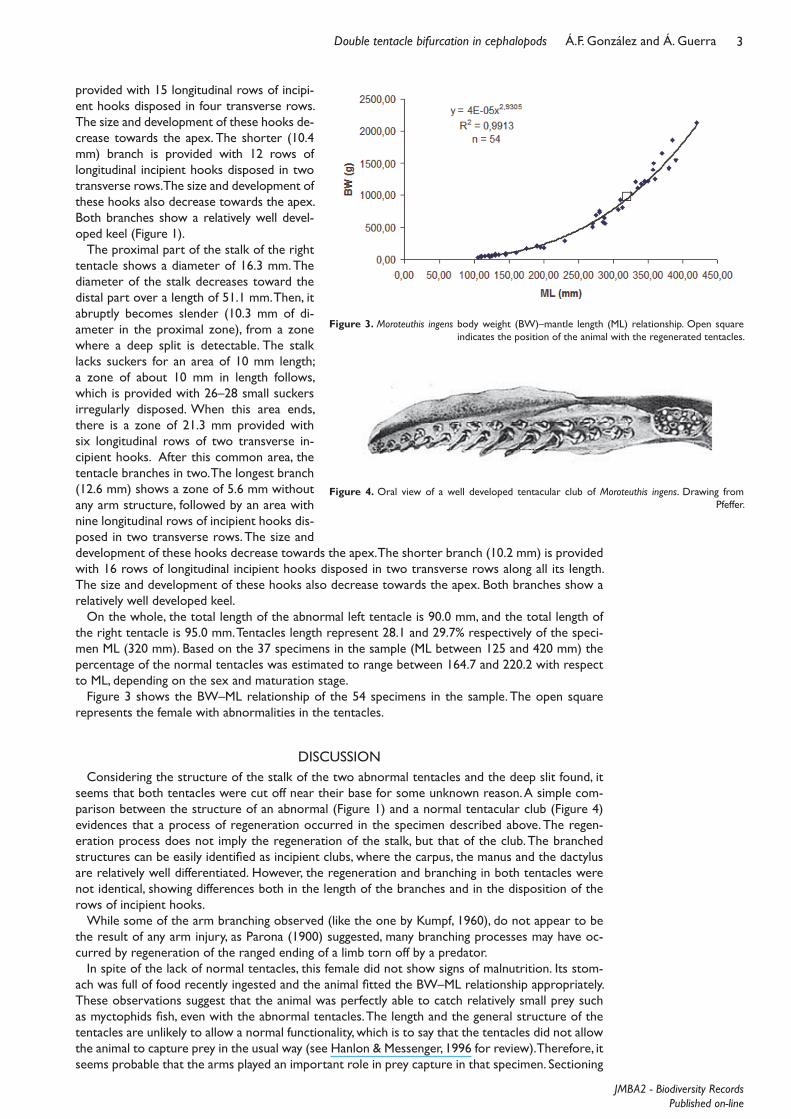
Figure 3 shows the BW–ML relationship of the 54 specimens in the sample. The open square represents the female with abnormalities in the tentacles.
DISCUSSIONConsidering the structure of the stalk of the two abnormal tentacles and the deep slit found, it
seems that both tentacles were cut off near their base for some unknown reason. A simple com-parison between the structure of an abnormal (Figure 1) and a normal tentacular club (Figure 4) evidences that a process of regeneration occurred in the specimen described above. The regen-eration process does not imply the regeneration of the stalk, but that of the club. The branched structures can be easily identified as incipient clubs, where the carpus, the manus and the dactylus are relatively well differentiated. However, the regeneration and branching in both tentacles were not identical, showing differences both in the length of the branches and in the disposition of the rows of incipient hooks.
While some of the arm branching observed (like the one by Kumpf, 1960), do not appear to be the result of any arm injury, as Parona (1900) suggested, many branching processes may have oc-curred by regeneration of the ranged ending of a limb torn off by a predator.
In spite of the lack of normal tentacles, this female did not show signs of malnutrition. Its stom-ach was full of food recently ingested and the animal fitted the BW–ML relationship appropriately. These observations suggest that the animal was perfectly able to catch relatively small prey such as myctophids fish, even with the abnormal tentacles. The length and the general structure of the tentacles are unlikely to allow a normal functionality, which is to say that the tentacles did not allow the animal to capture prey in the usual way (see Hanlon & Messenger, 1996 for review). Therefore, it seems probable that the arms played an important role in prey capture in that specimen. Sectioning
Figure 3. Moroteuthis ingens body weight (BW)–mantle length (ML) relationship. Open square indicates the position of the animal with the regenerated tentacles.
Figure 4. Oral view of a well developed tentacular club of Moroteuthis ingens. Drawing from Pfeffer.

Á.F. González and Á. Guerra Double tentacle bifurcation in cephalopods
JMBA2 - Biodiversity RecordsPublished on-line
4
of both tentacles in an early stage of development is likely to quickly induce two processes in paral-lel: (a) the adaptation to a new situation, i.e. the development of a new hunting behaviour (different from the usual one which involve the use of tentacles to collect prey) that allowed the animal not only to survive but to feed appropriately; and (b) the regeneration of the tentacles.
Although the information on the development of behaviour in cephalopods is still scarce, there is experimental evidence that some octopuses, the common cuttlefish and also squid species are able to learn; and, as pointed out by Hanlon & Messenger (1996), ‘it is surely unlikely that animals that are so good at learning in an aquarium would not put this ability to good use in the sea, and the inference must be that learning is extremely important to octopuses in their natural habitat’. To date, no studies on the behaviour or the nervous system have been undertaken on Moroteuthis ingens. However, the receptors and central nervous system of Onychoteuthis banksi, a M. ingens re-lated species, are well developed, the optic lobes representing 234% of the total brain volume, and the central nervous system closely resembles that of squid Loligo (Nixon & Young, 2003). On this basis, there is no reason to reject the possibility that M. ingens is able to learn and also that it could develop individual behaviour.
A large number of organisms are able to regenerate body parts (Bryant et al., 2002). Post-trau-matic regeneration of arms and tentacles involves a process well studied in cephalopods (Féral, 1978, 1979; Bello, 1995). For example, the regenerative process of a sectioned arm in Sepia officinalis is very complex and was differentiated in six phases (Féral, 1978, 1979); the arm is regenerated com-pletely after three months, it is structurally identical to the other arms and absolutely functional. Similar processes were observed in sepiolids and octopods (Féral, 1978, 1979). The case we present in this paper, however, is different, since the regeneration concerned only the tentacular clubs, not the whole tentacles.
The most comprehensive study regarding the regeneration process in a cephalopod was made by Toll & Binger (1991). These authors suggested a possible relationship of the process of arm and tentacle tips polyfurcation in octopods with the regenerative processes in amphibian limbs. How-ever, to date, there is a complete lack of studies concerning this issue in cephalopods except the previously cited and those of Féral (1978, 1979) in Sepia officinalis, where the process was studied only from morphological, histological and cytological points of view. From those perspectives, the regeneration process in cephalopods, overall, practically coincides with the ones observed in other invertebrates and vertebrates, such as urodels (Lodish et al., 2003).
The regeneration process always starts with the epidermis covering the stump within 24 hours, forming a structure called the Apical Epidermal Crest (AEC). This results in the formation of a blast-em. At the same time, pattern formation genes—homeobox genes such as Hox A and Hox D—are activated, the same way they were when the limb was formed in the embryo. From the blastema distal structures form first. Intermediate layers are filled in afterwards. Motor neurons grow with the regenerated limb, and innervate the same target muscles they originally controlled. The entire process takes around three months in the adult and then the limb becomes fully functional (Lodish et al., 2003).
Recent studies have examined the developmental expression pattern for eight out of nine Hox genes in the Hawaiian bobtail squid Euprymna scolopes, by whole-mount in situ hybridization. The results show that Hox genes correctly expressed have been repeatedly recruited and in different ways in the origin of the new cephalopod structures. It was demonstrated that the structure of the brachial crown, funnel, stellate ganglia of the mantle and the neurons that regulate their behaviour, including the ganglia and the lobes of the nervous system, are the expression of several Hox genes interacting co-ordinately during the embryonic development of that species (Lee et al., 2003).
We suggest that both abnormalities observed in our M. ingens specimen, the bifurcation and the regeneration of the tentacular clubs only, could be due to Hox genes mutation. It was demonstrated that Hox genes determine where limbs and other body segments will grow in a developing foetus or larva. Also, that mutations in any one of these genes can lead to the growth of extra, typically non-functional, body parts in invertebrates (Lodish et al. 2003). Although mutation in vertebrate Hox genes usually results in spontaneous abortion, mutations to homeobox genes in invertebrates can produce visible phenotypic changes (Lodish et al., 2003).
The new tools and strategies available to study the regeneration process in planarians and other worms (Saló & Baguñá, 2002), as well as in vertebrates (Bryant et al., 2002) should be taken into consideration when programming future and necessary researches on the regeneration process in cephalopods.
We would like to express our thanks to Mr Julio Maroto for providing the sample, Dr Ann Pulsford for the improvement of the paper, and Dr Francisco Rocha and Dr Chingis Nigmatullin for their valuable collabora-tion, Mr Manuel Enrique Garcia (‘Garci’) for the photographs and the other components of the ECOBIOMAR group for technical assistance.

Double tentacle bifurcation in cephalopods Á.F. González and Á. Guerra
JMBA2 - Biodiversity RecordsPublished on-line
5
REFERENCESAdam, W., 1932. Notes sur les Céphalopodes: 1. Anomalie de la massue tentaculaire chez Alloteuthis subulata
Lamarck. Bulletin of the Museum Royal d’ Histoire Naturelle Belgique, 8, 1–3.Bello, G., 1995. Hectocotylus regeneraltion in wild-caught sepiolid squid. Journal of the Marine Biological Association
of the United Kingdom, 75, 491–494.Bryant, S.V., Endo, T. & Gardiner, D.M., 2002. Vertebrate limb regeneration and the origin of limb stem cells.
International Journal of Developmental Biology, 46, 887–896.Clarke, M.R., 1966. A review of the systematics and ecology of oceanic squids. Advances in Marine Biology, 4,
91–300.Féral, J.P., 1978. La régénération des bras de la seiche Sepia officinalis L. (Cephalopoda, Sepioidea). I. Étude mor-
phologique. Cahiers de Biologie Marine, 19, 355–361.Féral, J.P., 1979. La régénération des bras de la seiche Sepia officinalis L. (Cephalopoda, Sepioidea). II. Étude his-
tologique et cytologique. Cahiers de Biologie Marine, 20, 29–42.Gleadall, I.G., 1989. An octopus with only seven arms: anatomical details. Journal of Molluscan Studies, 55,
479–487.Hanlon, R.T. & Messenger, J.B., 1996. Cephalopod behaviour. Cambridge: Cambridge University Press.Ikeda, S., 1891. A list of Japanese Cephalopoda in the Zoological Institute of the Imperial University. Zoology
Magazine, 2, 479–482. [In Japanese.]Jackson, G.D., 1993. Growth zones within the statolith microstructure of the deepwater squid Moroteuthis
ingens (Cephalopoda: Onychoteuthidae): evidence for a habitat shift? Canadian Journal of Fisheries and Aquatic Sciences, 50, 2366–2374.
Jackson, G.D., 1997. Age, growth and maturation of the deepwater squid Moroteuthis ingens (Cephalopoda: Onychoteuthidae) in New Zealand waters. Polar Biology, 17, 268–274.
Jackson, G.D., 2001. Confirmation of winter spawning of Moroteuthis ingens (Cephalopoda: Onychoteuthidae) in the Chatham Rise of New Zealand. Polar Biology, 24, 97–100.
Jackson, G.D. & Jackson C.H., 2004. Mating and spermatophore placement in the onychoteuthid squid Moro-teuthis ingens. Journal of the Marine Biological Association of the United Kingdom, 84, 783–784.
Jackson, G.D, George M.J.A. & Buxton N.G., 1998a. Distribution and abundance of the squid Moroteuthis in-gens (Cephalopoda: Onychoteuthidae) in the Falkland Islands region of the South Atlantic. Polar Biology, 20, 161–169.
Jackson, G.D, Mckinnon, J.F, Lalas, C., Ardern, R. & Buxton, N.G., 1998b. Food spectrum of the deepwater squid Moroteuthis ingens (Cephalopoda: Onychoteuthidae) in New Zealand Waters. Polar Biology, 20, 56–65.
Jackson, G.D. & Mladenov, P.V., 1994. Terminal spawning in the deepwater squid Moroteuthis ingens (Cephalo-poda: Onychoteuthidae). Journal of Zoology, 234, 189–201.
Jackson, G.D, Shaw, A.G.P. & Lalas, C., 2000. Distribution and biomass of two squid species off southern New Zealand: Nototodarus sloanii and Moroteuthis ingens. Polar Biology, 23, 699–705.
Jereb, P,, Ragonese, S. & Murone, R., 1989. Sue due esemplari anomali di Scaergus unicirrhus (Mollusca, Cephalo-poda). Oebalia, 15, 807–809.
Jereb, P., Stefano, M. di & Mazzola, A., 1997. Anomalies in Sepiolinae hectocotylization Journal of the Marine Bio-logical Association of the United Kingdom, 77, 1239–1242.
Kumph, H.E., 1960. Arm abnormality in Octopus. Nature, London, 185, 334–335.Lee, P.N., Callaerts, P., Couet, H.G. de & Martindale, M.Q., 2003. Cephalopod Hox genes and the origin of mor-
phological novelties. Nature, London, 424, 1061–1065.Lodish, H., Berk, A., Zipursky, S., Matsudaira, P., Baltimore, D., Darnell, J. & Zipursky, L., 2003. Molecular cell biology
5th edn. W.H. Freeman & Co.Nixon, M. & Young, J.Z., 2003. The brains and lives of cephalopods. Oxford: Oxford University Press.Okada, Y.K., 1935. An octopus with branched arms and mode of branching. Annotated Zoology of Japan, 15, 5–23.Okada, Y.K., 1937. An occurrence of branched arms in the decapod cephalopod, Sepia esculenta Hoyle. Anno-
tated Zoology of Japan, 17, 93–94.Okada, Y.K., 1965a. On Japanese octopuses with branched arms, with special reference to their captures from
1884 to 1964. Proceedings of the Japanese Academy, 41, 618–623.Okada, Y.K., 1965b. Rule of arm-branching in Japanese octopuses with branched arms. Proceedings of the Japa-
nese Academy, 41, 624–629.Palacio, F.J., 1973. On the double hectocotylisation of octopods. Nautilus, 87, 99–102.Parona, C., 1900. Sulla dicotomía delle braccia nei cephalopodi. Atti della Società di Scienze e Geografia di Genova,
9, 224–230.Pfeffer, G., 1912. Results of the Plankton Expedition of the Humboldt Foundation. Vol. 2. The Cephalopoda of the
Plankton Expedition. Washington, D.C.: Smithsonian Institution Libraries and the National Science Foundation. [English Translation.]
Phillips, K.L., Jackson, G.D. & Nichols, P.D., 2001. Predation on myctophids by the squid Moroteuthis ingens around Macquarie and Heard Islands: stomach contents and fatty acid analysis. Marine Ecology Progress Series, 215, 179–189.
Phillips, K.L., Nichols, P.D. & Jackson, G.D., 2003. Size-related dietary changes in the squid Moroteuthis ingens at the Falkland Islands: stomach contents and fatty-acid analyses. Polar Biology, 26, 474–485.
Robson, G.C., 1929a. A monograph of the recent Cephalopoda. I. Octopodinae. London: British Museum (Natural History).
Robson, G.C., 1929b. On a case of bilateral hectocotylization in Octopus rugosus. Proceedings of the Zoological Society of London, 7, 95–97.

Á.F. González and Á. Guerra Double tentacle bifurcation in cephalopods
JMBA2 - Biodiversity RecordsPublished on-line
6
Sasaki, M., 1929. A monograph of the dibranchiate cephalopods of the Japanese and adjacent waters. Journal of the Faculty of Agriculture, Hokkaido Imperial University, 20, 1–357.
Saló, E. & Baguñá, J., 2002. Regeneration in Planarians and other worms: new findings, new tools, and new per-spectives. Journal of Experimental Zoology, 292, 528–539.
Sands, C.J., Jarman, S.N. & Jackson, G.D., 2003. Genetic differentiation in the squid Moroteuthis ingens inferred from RAPD analysis. Polar Biology, 26, 166–170.
Smith, E.A., 1907. Notes on an “octopus” with branching arms. Annual Magazine of Natural History Series, 7, 407–411.
Toll, R.B. & Binger, L.C., 1991. Arm anomalies: cases of supernumerary development and bilateral agenesis of arm pairs in Octopoda (Mollusca, Cephalopoda). Zoomorphology, 110, 313–316.
Vecchione, M., Young, R.E. & Tsuchiya, K., 2003. Tree of life web project. http://tolweb.org/tree?group=Onykia_ingensandcontgroup=Onykia.
Verrill, A.E., 1882. The Cephalopoda of the north-eastern coast of America. Transactions of the Connecticut Acad-emy Science, V, Pt.1.
Submitted 19 July 2006. Accepted 18 October 2006.
Related Documents











