FIRST KNOWN COMPLETE SKULLS OF THE SCIMITAR-TOOTHED CAT MACHAIRODUS APHANISTUS (FELIDAE, CARNIVORA) FROM THE SPANISH LATE MIOCENE SITE OF BATALLONES-1 MAURICIO ANTON 1 , MANUEL J. SALESA 2 , JORGE MORALES 1 , and ALAN TURNER 2 1 Departamento de Paleobiologı ´a, Museo Nacional de Ciencias Naturales-CSIC, Jose ´ Gutie ´ rrez Abascal, 2, 28006 Madrid, Spain, [email protected] 2 School of Biological and Earth Sciences, Liverpool John Moores University, Liverpool L3 3AF, United Kingdom ABSTRACT—We describe a sample of skulls, mandibles and isolated dentitions of Machairodus aphanistus from the Spanish late Vallesian (late Miocene) site of Batallones-1, near Madrid. This is the first time that the cranial morphology of this species has been seen in detail, and shows that late Vallesian M. aphanistus combined a relatively primitive cranial and mandibular morphology with very specialized upper canines, as flattened as in the later, cranially derived species M. giganteus. Cranio-mandibular adaptations for the canine shear-bite are incipient or non-existent, suggesting that the specialized canines of M. aphanistus were used within the context of a biting mechanism more similar to the canine bite of modern felids. Evolution of more extreme machairodont craniodental features in later members of the homotherin lineage converged with smilodontins but occurred independently of the initial development of derived saber-like canines. The high number of specimens, at least 14 individuals represented, reveals considerable variation in size within the population, with indications of sexual dimorphism in the size of the upper canines. Differences in cranial morphology suggest separation at the generic level between M. aphanistus and M. giganteus. INTRODUCTION The scimitar-tooth cat Machairodus aphanistus was a success- ful, lion-sized felid that entered Europe during the early Vall- esian as part of an immigration wave of Eastern mammal species often known as the “Hipparion datum” (Sen, 1990; Morales et al., 1999; Agustı ´ and Anto ´ n, 2002). It soon became widespread, and seems to have been the top predator in the European mam- malian faunas of its time, apparently displacing endemic carni- vores such as the large amphicyonids and barbourofelines (Agustı´ and Anto ´ n, 2002). However, despite its wide distribu- tion, M. aphanistus has traditionally been known from remark- ably fragmentary remains. This paucity of material has precluded a deeper insight into the morphology, adaptations, evolution and ecology of this carnivore. The main characteristics of M. aphanistus as defined by pre- vious authors were usefully summarized as follows by Sotnikova (1992): small lower incisors arranged in a straight row, large lower canines, a small diastema between the lower canine and p3, large premolars with a complete set of accessory cusps, a well developed metaconid-talonid complex on the lower carnassial, mandibular ramus thick and high, an undeveloped mandibular flange, an upper carnassial with a distinct protocone and pre- parastyle and all teeth probably serrated. As may be noted, this informal diagnosis is based strictly on characters of the dentition and anterior mandible, which were almost the only parts of the animal’s anatomy known at that time. However, the recently discovered site of Batallones-1, a carnivore trap of late Vallesian age near Madrid in central Spain (Morales et al., 2000), is pro- viding an unprecedented wealth of M. aphanistus fossils, includ- ing complete skulls and postcrania. The purpose of this paper is therefore twofold: first, we describe the craniodental morphol- ogy of M. aphanistus as revealed by the rich Batallones-1 sample. We then assess the functional implications of that morphology and attempt to put the adaptations of this taxon into evolution- ary perspective. We confine our discussion to cranial and dental material because these currently provide the most useful com- parisons at this stage with other late Miocene machairodont fe- lids. Postcranial remains of M. aphanistus from Batallones-1 will be considered in detail elsewhere. A particularly important aspect of comparing the craniodental anatomy of M. aphanistus with that of other felids, sabertoothed and non-sabertoothed, is the possibility of checking whether, and to what degree, the adaptations for the canine shear-bite are present. The canine shear-bite hypothesis was first proposed by Akersten (1985) in a study centered on the American Pleisto- cene genus Smilodon, but he also suggested that the same hy- pothesis could be valid, with some differences in detail, for other sabertoothed taxa. A recent study of the functional anatomy of the neck and head in the machairodont genus Homotherium (Anto ´ n and Galobart, 1999), a likely descendant of Machairodus (Turner and Anto ´ n, 1997), confirms both the presence of adap- tations for canine shear-bite in this genus and the clear differ- ences in detail. If saber-like canines are part of a set of cranio- dental adaptations for the canine shear-bite, then the specialized nature of the sabers of M. aphanistus as previously known from sites like Los Valles de Fuentiduen ˜ a (Ginsburg et al., 1981) would lead us to hypothesize that cranial features would be nearly as derived as in M. giganteus. If, on the other hand, the cranium and mandible of M. aphanistus turn out to be markedly more primitive than in M. giganteus, then we would be forced to admit that the development of derived cranial machairodont fea- tures in M. giganteus and later members of the homotherin lin- eage converged with smilodontins. Such convergences would be the result of a strong selective pressure for adaptations to a canine-shear bite, later than and independent of the original evolution of saber-like canines. The new material from Batal- lones-1 allows us to test these hypotheses for the first time. SYSTEMATIC PALEONTOLOGY Class MAMMALIA Linnaeus, 1758 Order CARNIVORA Bowdich, 1821 Family FELIDAE Fischer, 1817 Subfamily MACHAIRODONTINAE Gill, 1872 Genus MACHAIRODUS Kaup, 1833 MACHAIRODUS APHANISTUS (Kaup, 1832) Holotype—Right mandibular fragment with p3, p4 and m1 (HLMD Din 1132), Hessisches Landesmuseum Darmstadt (Ger- many). Journal of Vertebrate Paleontology 24(4):957–969, December 2004 © 2004 by the Society of Vertebrate Paleontology 957

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

FIRST KNOWN COMPLETE SKULLS OF THE SCIMITAR-TOOTHED CAT MACHAIRODUSAPHANISTUS (FELIDAE, CARNIVORA) FROM THE SPANISH LATE MIOCENE SITE
OF BATALLONES-1
MAURICIO ANTON1, MANUEL J. SALESA2, JORGE MORALES1, and ALAN TURNER2
1Departamento de Paleobiologıa, Museo Nacional de Ciencias Naturales-CSIC, Jose Gutierrez Abascal, 2,28006 Madrid, Spain, [email protected]
2School of Biological and Earth Sciences, Liverpool John Moores University, Liverpool L3 3AF, United Kingdom
ABSTRACT—We describe a sample of skulls, mandibles and isolated dentitions of Machairodus aphanistus from theSpanish late Vallesian (late Miocene) site of Batallones-1, near Madrid. This is the first time that the cranial morphologyof this species has been seen in detail, and shows that late Vallesian M. aphanistus combined a relatively primitive cranialand mandibular morphology with very specialized upper canines, as flattened as in the later, cranially derived species M.giganteus. Cranio-mandibular adaptations for the canine shear-bite are incipient or non-existent, suggesting that thespecialized canines of M. aphanistus were used within the context of a biting mechanism more similar to the canine biteof modern felids. Evolution of more extreme machairodont craniodental features in later members of the homotherinlineage converged with smilodontins but occurred independently of the initial development of derived saber-like canines.The high number of specimens, at least 14 individuals represented, reveals considerable variation in size within thepopulation, with indications of sexual dimorphism in the size of the upper canines. Differences in cranial morphologysuggest separation at the generic level between M. aphanistus and M. giganteus.
INTRODUCTION
The scimitar-tooth cat Machairodus aphanistus was a success-ful, lion-sized felid that entered Europe during the early Vall-esian as part of an immigration wave of Eastern mammal speciesoften known as the “Hipparion datum” (Sen, 1990; Morales etal., 1999; Agustı and Anton, 2002). It soon became widespread,and seems to have been the top predator in the European mam-malian faunas of its time, apparently displacing endemic carni-vores such as the large amphicyonids and barbourofelines(Agustı and Anton, 2002). However, despite its wide distribu-tion, M. aphanistus has traditionally been known from remark-ably fragmentary remains. This paucity of material has precludeda deeper insight into the morphology, adaptations, evolution andecology of this carnivore.
The main characteristics of M. aphanistus as defined by pre-vious authors were usefully summarized as follows by Sotnikova(1992): small lower incisors arranged in a straight row, largelower canines, a small diastema between the lower canine andp3, large premolars with a complete set of accessory cusps, a welldeveloped metaconid-talonid complex on the lower carnassial,mandibular ramus thick and high, an undeveloped mandibularflange, an upper carnassial with a distinct protocone and pre-parastyle and all teeth probably serrated. As may be noted, thisinformal diagnosis is based strictly on characters of the dentitionand anterior mandible, which were almost the only parts of theanimal’s anatomy known at that time. However, the recentlydiscovered site of Batallones-1, a carnivore trap of late Vallesianage near Madrid in central Spain (Morales et al., 2000), is pro-viding an unprecedented wealth of M. aphanistus fossils, includ-ing complete skulls and postcrania. The purpose of this paper istherefore twofold: first, we describe the craniodental morphol-ogy of M. aphanistus as revealed by the rich Batallones-1 sample.We then assess the functional implications of that morphologyand attempt to put the adaptations of this taxon into evolution-ary perspective. We confine our discussion to cranial and dentalmaterial because these currently provide the most useful com-parisons at this stage with other late Miocene machairodont fe-lids. Postcranial remains of M. aphanistus from Batallones-1 willbe considered in detail elsewhere.
A particularly important aspect of comparing the craniodentalanatomy of M. aphanistus with that of other felids, sabertoothedand non-sabertoothed, is the possibility of checking whether, andto what degree, the adaptations for the canine shear-bite arepresent. The canine shear-bite hypothesis was first proposed byAkersten (1985) in a study centered on the American Pleisto-cene genus Smilodon, but he also suggested that the same hy-pothesis could be valid, with some differences in detail, for othersabertoothed taxa. A recent study of the functional anatomy ofthe neck and head in the machairodont genus Homotherium(Anton and Galobart, 1999), a likely descendant of Machairodus(Turner and Anton, 1997), confirms both the presence of adap-tations for canine shear-bite in this genus and the clear differ-ences in detail. If saber-like canines are part of a set of cranio-dental adaptations for the canine shear-bite, then the specializednature of the sabers of M. aphanistus as previously known fromsites like Los Valles de Fuentiduena (Ginsburg et al., 1981)would lead us to hypothesize that cranial features would benearly as derived as in M. giganteus. If, on the other hand, thecranium and mandible of M. aphanistus turn out to be markedlymore primitive than in M. giganteus, then we would be forced toadmit that the development of derived cranial machairodont fea-tures in M. giganteus and later members of the homotherin lin-eage converged with smilodontins. Such convergences would bethe result of a strong selective pressure for adaptations to acanine-shear bite, later than and independent of the originalevolution of saber-like canines. The new material from Batal-lones-1 allows us to test these hypotheses for the first time.
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, 1758Order CARNIVORA Bowdich, 1821
Family FELIDAE Fischer, 1817Subfamily MACHAIRODONTINAE Gill, 1872
Genus MACHAIRODUS Kaup, 1833MACHAIRODUS APHANISTUS (Kaup, 1832)
Holotype—Right mandibular fragment with p3, p4 and m1(HLMD Din 1132), Hessisches Landesmuseum Darmstadt (Ger-many).
Journal of Vertebrate Paleontology 24(4):957–969, December 2004© 2004 by the Society of Vertebrate Paleontology
957

Type locality—Eppelsheim (Germany, MN 9, Upper Mio-cene).
Emended diagnosis—A lion-sized extinct felid with smalllower incisors arranged in a straight row, large lower canineswith flattened roots and an oval cross section to the crown, asmall diastema between the lower canine and p3, large lowerpremolars with a complete set of additional cusps and p3 largerelative to p4, a well-developed metaconid-talonid complex onthe lower carnassial, mandibular horizontal ramus thick andhigh, an undeveloped mandibular flange, coronoid process highand posteriorly inclined, relatively large upper incisors set in ashallow arc anterior to the upper canines, high-crowned and veryflattened upper canines, P2 variably present, P3 with a very de-veloped posterior expansion, an upper carnassial with a distinctprotocone and preparastyle and all teeth probably serrated, amoderately convex dorsal profile of the skull, a well-developedsagittal crest, skull narrow in dorsal view across the zygoma,zygomatic arch low and gently curved in side view, temporalfossa elongated, paroccipital process well-developed and project-ing inferiorly beyond the relatively small mastoid process, naso-frontal suture intermediate between pantherine (pointed) andevolved machairdontine (straight) condition, postorbital pro-cesses large but low.
Machairodus aphanistus is a sabertoothed member of the fam-ily Felidae, which is divided into four subfamilies. Living cats,including the subfamilies Felinae, Acinonychinae and Pantheri-nae (Wozencraft, 1993; McKenna and Bell, 1997), are animalswith conical upper canines. The sabertooths belong to the extinctsubfamily Machairodontinae, and their relationship to the othersis outlined in Figure 1 (Turner and Anton, 1997; Anton et al.,1998). This phylogeny compiles a number of previous studiesabout the classification of this group, in the absence of a muchneeded formal analysis which would include both extant andextinct forms.
The craniodental morphology of the living subfamilies andtheir fossil forms is conservative and resembles that of primitivefelids in most respects, while the Machairodontinae became evermore derived until their extinction at the end of the Pleistocene.The latter group, often termed simply machairodonts, are in turndivided into at least two tribes, the Smilodontini which includesrobust cats like Megantereon and Smilodon, and the Homoth-erini which includes more gracile cats of the genus Machairodusand its likely descendant Homotherium. The differences in uppercanine morphology between the crown taxa of the Smilodontiniand Homotherini have lead to coining the common terms “dirk-tooth” for the former, and “scimitar-tooth” for the latter. A third
group, the metailurines, display more moderate sabertooth ad-aptations, and have been given tribal or sub-familial rank de-pending on the authors (Crusafont and Aguirre, 1972; Beau-mont, 1978; Turner and Anton, 1997).
All felids are thought to descend from the basal genus Proai-lurus, known from the latest Oligocene and Early Miocene ofEurope, and the split between the subfamilies is assumed to havehappened in mid-Miocene times within the widespread genusPseudaelurus (Beaumont, 1978). Of course, sabertooth adapta-tions have evolved at least three times outside the family Felidae(Turner and Anton, 1997), so that when we use the word “sa-bertooths” in the text we are referring to sabertoothed mamma-lian predators in general, while the term “machairodonts” is usedhere exclusively for the sabertoothed members of the familyFelidae.
Machairodus aphanistus was the first member of the felidmachairodontine lineage to attain lion size, and it has often beenconsidered ancestral to the Turolian species M. giganteus, whichwas also lion-sized but more derived dentally. However, thisrelationship has been questioned by Ginsburg et al. (1981), whoargued that several features of M. aphanistus, such as the largelower canines, are too specialized to permit direct ancestry andwent on to propose the smaller, more primitive species M. al-berdiae as the likely ancestor of M. giganteus. Other early speciesof Machairodus are known from Asia (Schmidt Kittler, 1976;Sotnikova, 1992) but in all these primitive taxa the skull mor-phology remains virtually unknown. In contrast, complete skullsof M. giganteus are known from sites in China (Chang, 1957) andMoldavia (Riabinin, 1929), clearly showing that it had all thecranial features typical of advanced machairodonts. Those fea-tures are further developed in the Plio-Pleistocene Homothe-rium, known from complete skulls from many sites in four con-tinents (Turner and Anton, 1997).
MACHAIRODONT ADAPTATIONS AND THECANINE SHEAR-BITE
Sabertoothed predators are characterized not only by theirelongated upper canines but also by a whole set of craniodentaladaptations often termed the “machairodont traits” (Emersonand Radinsky, 1980). The functional interpretation of these fea-tures has been a matter of debate for decades (Merriam andStock, 1932; Simpson, 1941; Bohlin, 1947; Kurten, 1952; Biknev-icius et al., 1996; Turner and Anton, 1997; Anton et al., 1998;Anton and Galobart, 1999), but they obviously imply a particularway of dealing with prey, and it is clear that within the familyFelidae the presence of conical-toothed and saber-toothed spe-cies points to at least two different types of predatory behaviorand perhaps also feeding behavior.
Pantherine cats, the large conical-toothed species such as thelion, tiger, jaguar and leopard, kill large prey using either a napebite or a throat-bite. In the first case, damage to the brain orspine of prey is the main cause of death, while in the second casethe victim usually dies of strangulation (Leyhausen, 1979). Athird type of bite used by lions when killing large prey is a bite tothe muzzle, which cuts the air supply and results in suffocation(Schaller, 1972). The short, strong, and relatively blunt canines ofpantherines are well suited for these kinds of bite, but even theycan break if hard bone is encountered, so the nape bite is usuallyapplied to relatively smaller prey with more fragile bones, whilelarger animals are dispatched with a throat bite or a muzzle bite(Seidensticker and McDougal, 1993). These bite modalities relyon the exertion of enormous pressure at the tips of both upperand lower canines, which in turn requires relatively low gapes,and the canines of pantherines are relatively short and thus keepgape as low as possible. Other, generally smaller living cats usevarious versions of these killing methods as appropriate.
FIGURE 1. Phylogenetic hypothesis for the Felidae (modified fromAnton and Galobart, 1999).
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 24, NO. 4, 2004958

On the other hand, derived sabertooths are currently thoughtto have killed their prey using the “canine shear-bite” (Akersten,1985), a bite modality where the upper canines penetrate the skinand flesh of prey by means of a powerful downward thrust of thewhole skull powered by the neck musculature. In this scheme thelower canines and incisors would have first acted to provide ananchoring point for that downward thrust, after which they mayalso have penetrated the skin. A final backward pull may haveresulted in a large chunk of flesh being taken, but the main effectof this bite is likely to have been the massive loss of bloodthrough severing of major vessels combined perhaps with suffo-cation and a rapidly induced shock reaction (Turner and Anton,1997; Anton and Galobart, 1999). Throughout the bite, the mainmandible-closing muscles, the masseter and temporalis, wouldhave had a supporting function, but the main pull for penetrationof the sabers was most logically provided by neck muscles in-serting on the mastoid process and the basisphenoid.
The craniodental features interpreted by Akersten (1985) asadaptations for the canine shear-bite broadly coincide with thoserecorded by previous authors (Kurten, 1952; Emerson and Ra-dinsky, 1980; Biknevicius et al., 1996) under the generic defini-tion of sabertooth adaptations, and listed here in Figure 2.
In functional terms, some of these features can be seen asadaptations to increased gape (5, 6, 8, 9, 10, 11, 12), while othersare related to the recruitment of neck musculature for the killingbite (7, 9, 10). Involvement of neck muscles in mastication al-ready exists in living cats (Gorniak and Gans, 1980) and theaction of those muscles in the killing bite in sabertooths is seenby Bryant (1996: 297) as “an elaboration of an existing biome-chanical system that was necessitated by the low torque of thejaw adductors at the large gapes required for canine clearances”.Features of the cheek teeth (4) appear to be simply adaptationsto hypercarnivory. The projection and enlargement of the incisorbattery (features 2 and 3) was interpreted by Akersten (1985) asan adaptation to tear large chunks of flesh during the canineshear-bite and also as an aid to biting meat off the carcass duringfeeding, and more recently it has been suggested that the en-larged incisors would help to prevent breakage of the fragile
sabers by stabilizing the bitten area of prey and thus minimizinglateral stress to the upper canines (Biknevicius et al., 1996).
THE “CERRO DE LOS BATALLONES” LOCALITIES
Cerro Batallones is a site complex comprising several hectaresin Torrejon de Velasco some twenty-five kilometers to the southof Madrid, with a latitude of 4447570 and a longitude of 439220,U. T. M. coordinates (Figure 3). The sediments are composed ofclay, micaceous sands, carbonates and flint, and the fossiliferoushorizons have been exposed during commercial exploitation forsepiolites, a material used for domestic cat litter. Six discretelocalities are now known, of which only the first, Batallones-1,has so far been excavated systematically, and this has yielded thevast majority of mammal fossils, including all the specimens ofM. aphanistus described here (Morales et al., 2000). Current in-terpretations of the Batallones-1 fossil assemblage, composed of98% carnivore remains, suggest a carnivore trap (Anton andMorales, 2000).
MATERIALS AND METHODS
We studied a sample of crania, mandibles and isolated denti-tions of M. aphanistus from Batallones-1. The crania are mostlycomplete but strongly crushed owing to the compaction of theclay-rich sediments (Fig. 4). This type of preservation permitsgood observations of morphological details, but three-dimensional aspects of the skull are difficult or impossible tointerpret. Since crushing has in all cases been either dorso-ventral or lateral, or a combination of both, measurements alongthe long axis of the skull are generally reliable, but overall widthmeasurements are impossible to take in most cases. Observa-tions of general skull morphology in side view are more reliablein laterally crushed skulls, where one side has often “folded”over the other, but the dorsal outline can still be observed on theless deformed side, while in dorso-ventrally crushed skulls suchobservations are impossible. Several of the crania have man-dibles still attached to them, making observations of the palatineregion and occlusal aspects of the dentition impossible but pro-viding information about the relative dimensions of cranium andmandible in the same individual. Most crania and mandibleshave lost all their incisors, partly due to the small size and narrowroots of these teeth, so that most observations about the incisorsare based on the shape and position of the alveoli.
We offer a summary description of the main craniodental fea-tures of M. aphanistus, with emphasis on traits that may indicate
FIGURE 2. Skull of the Plio-Pleistocene scimitar-toothed cat Homothe-rium latidens showing the craniodental features typical of derived saber-tooths: 1, elongated and flattened upper canines; 2, enlarged incisorsarranged in an arc; 3, a protruding premaxillary; 4, enlarged, shearingcarnassials and reduced premolars; 5, shortened zygomatic arches andtemporal fossae; 6, a lowered glenoid; 7, an enlarged mastoid processprojecting antero-inferiorly; 8, reduced paroccipital processes; 9, largesagittal and lambdoidal crests; 10, an elevated occipital plane; 11, a re-duced coronoid process on the mandible; 12, relatively small lower ca-nines; 13, variously developed mental flanges on the mandible. FIGURE 3. Geographic situation of the Batallones-1 fossil site.
ANTON ET AL.—SKULLS OF MACHAIRODUS FROM BATALLONES-1 959
functional specialization and systematic affinities. We have fol-lowed previous authors (Merriam and Stock, 1932; Ballesio,1963; Beaumont, 1975; Emerson and Radinsky, 1980) in consid-ering the set of features listed in the introduction as indicative ofsabertooth adaptation, and the relative development of thosefeatures is compared between relevant taxa throughout the de-scription. We have also checked the presence and developmentof parietal depressions, a feature related by Duckler (1997) torelative stress at the insertion of the temporalis muscle.
For the purpose of comparison we have also studied a sampleof skulls of modern and fossil felids, including three specimens oflion, Panthera leo, three skulls of leopard, Panthera pardus, andcasts of skulls of Machairodus giganteus and Homotheriumlatidens. Data on the skull morphology in other machairodon-tines and primitive felids were taken from the literature as givenin the text, and details of muscle insertion areas on the skull aremostly based on the descriptions of modern carnivore anatomyby Barone (1967, 1989). Cranial, mandibular and dental mea-surements were recorded to the nearest 0.1 millimeter withmanual steel calipers.
DESCRIPTION AND COMPARISONS
Cranium
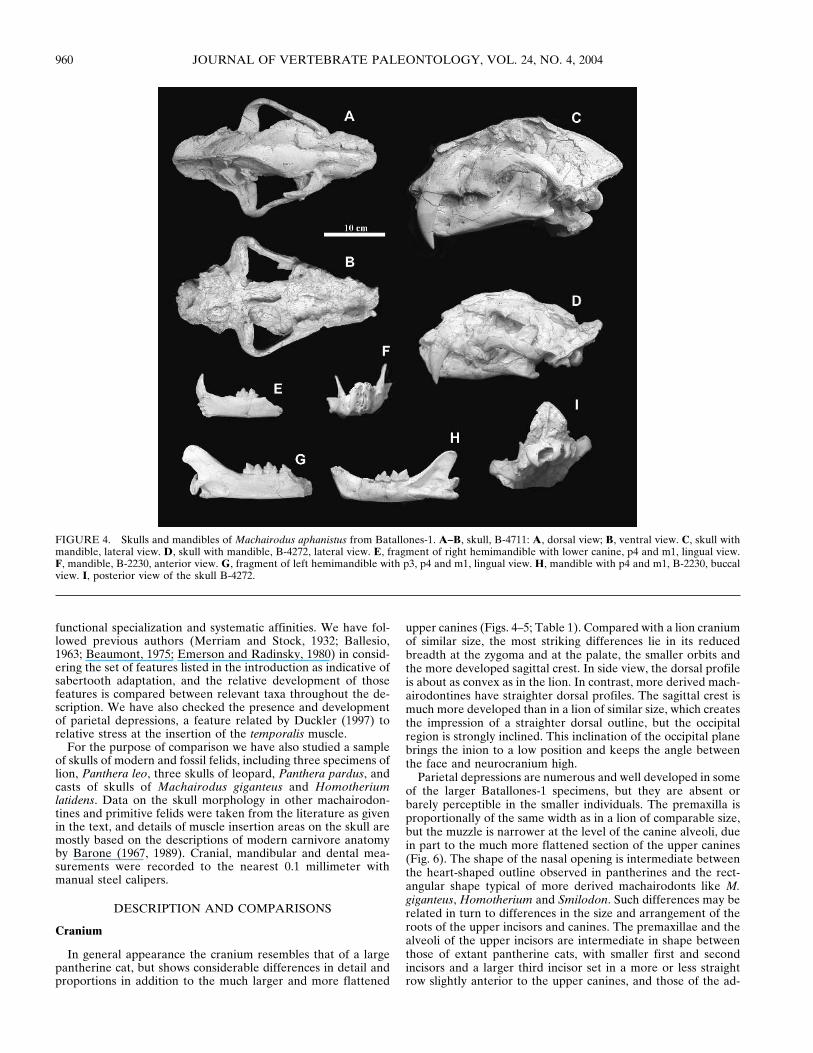
In general appearance the cranium resembles that of a largepantherine cat, but shows considerable differences in detail andproportions in addition to the much larger and more flattened
upper canines (Figs. 4–5; Table 1). Compared with a lion craniumof similar size, the most striking differences lie in its reducedbreadth at the zygoma and at the palate, the smaller orbits andthe more developed sagittal crest. In side view, the dorsal profileis about as convex as in the lion. In contrast, more derived mach-airodontines have straighter dorsal profiles. The sagittal crest ismuch more developed than in a lion of similar size, which createsthe impression of a straighter dorsal outline, but the occipitalregion is strongly inclined. This inclination of the occipital planebrings the inion to a low position and keeps the angle betweenthe face and neurocranium high.
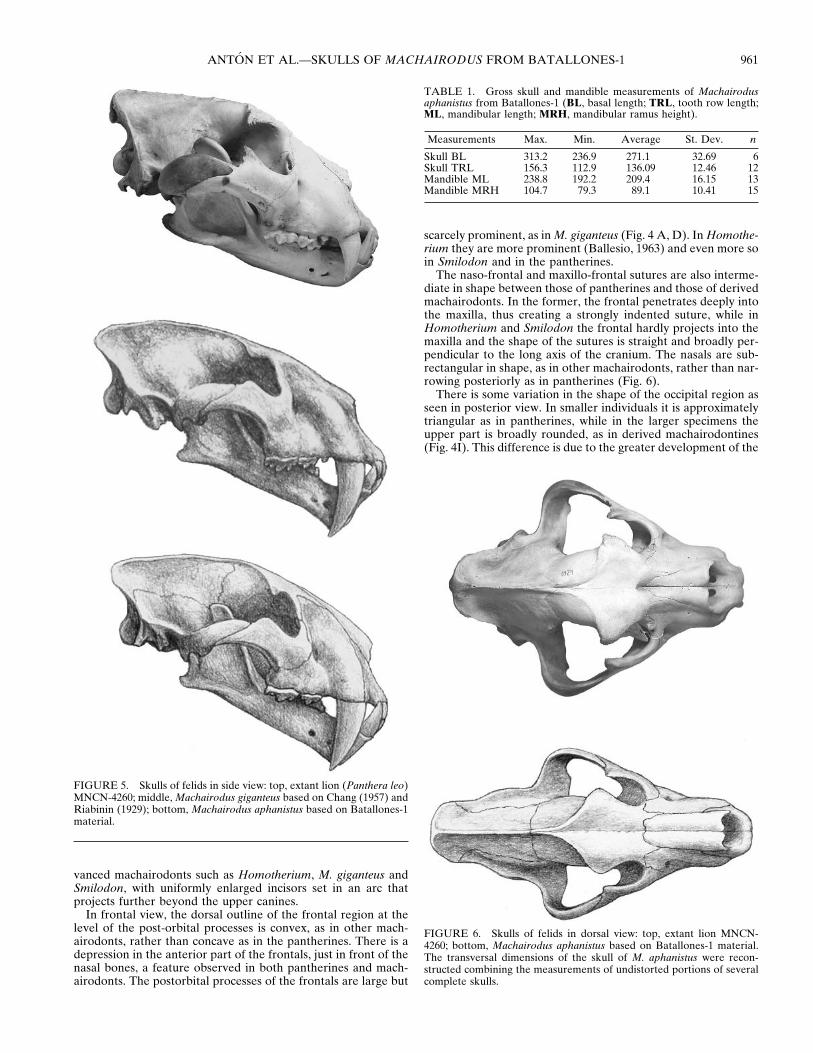
Parietal depressions are numerous and well developed in someof the larger Batallones-1 specimens, but they are absent orbarely perceptible in the smaller individuals. The premaxilla isproportionally of the same width as in a lion of comparable size,but the muzzle is narrower at the level of the canine alveoli, duein part to the much more flattened section of the upper canines(Fig. 6). The shape of the nasal opening is intermediate betweenthe heart-shaped outline observed in pantherines and the rect-angular shape typical of more derived machairodonts like M.giganteus, Homotherium and Smilodon. Such differences may berelated in turn to differences in the size and arrangement of theroots of the upper incisors and canines. The premaxillae and thealveoli of the upper incisors are intermediate in shape betweenthose of extant pantherine cats, with smaller first and secondincisors and a larger third incisor set in a more or less straightrow slightly anterior to the upper canines, and those of the ad-
FIGURE 4. Skulls and mandibles of Machairodus aphanistus from Batallones-1. A–B, skull, B-4711: A, dorsal view; B, ventral view. C, skull withmandible, lateral view. D, skull with mandible, B-4272, lateral view. E, fragment of right hemimandible with lower canine, p4 and m1, lingual view.F, mandible, B-2230, anterior view. G, fragment of left hemimandible with p3, p4 and m1, lingual view. H, mandible with p4 and m1, B-2230, buccalview. I, posterior view of the skull B-4272.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 24, NO. 4, 2004960
vanced machairodonts such as Homotherium, M. giganteus andSmilodon, with uniformly enlarged incisors set in an arc thatprojects further beyond the upper canines.
In frontal view, the dorsal outline of the frontal region at thelevel of the post-orbital processes is convex, as in other mach-airodonts, rather than concave as in the pantherines. There is adepression in the anterior part of the frontals, just in front of thenasal bones, a feature observed in both pantherines and mach-airodonts. The postorbital processes of the frontals are large but
scarcely prominent, as in M. giganteus (Fig. 4 A, D). In Homothe-rium they are more prominent (Ballesio, 1963) and even more soin Smilodon and in the pantherines.
The naso-frontal and maxillo-frontal sutures are also interme-diate in shape between those of pantherines and those of derivedmachairodonts. In the former, the frontal penetrates deeply intothe maxilla, thus creating a strongly indented suture, while inHomotherium and Smilodon the frontal hardly projects into themaxilla and the shape of the sutures is straight and broadly per-pendicular to the long axis of the cranium. The nasals are sub-rectangular in shape, as in other machairodonts, rather than nar-rowing posteriorly as in pantherines (Fig. 6).
There is some variation in the shape of the occipital region asseen in posterior view. In smaller individuals it is approximatelytriangular as in pantherines, while in the larger specimens theupper part is broadly rounded, as in derived machairodontines(Fig. 4I). This difference is due to the greater development of the
FIGURE 6. Skulls of felids in dorsal view: top, extant lion MNCN-4260; bottom, Machairodus aphanistus based on Batallones-1 material.The transversal dimensions of the skull of M. aphanistus were recon-structed combining the measurements of undistorted portions of severalcomplete skulls.
FIGURE 5. Skulls of felids in side view: top, extant lion (Panthera leo)MNCN-4260; middle, Machairodus giganteus based on Chang (1957) andRiabinin (1929); bottom, Machairodus aphanistus based on Batallones-1material.
TABLE 1. Gross skull and mandible measurements of Machairodusaphanistus from Batallones-1 (BL, basal length; TRL, tooth row length;ML, mandibular length; MRH, mandibular ramus height).
Measurements Max. Min. Average St. Dev. n
Skull BL 313.2 236.9 271.1 32.69 6Skull TRL 156.3 112.9 136.09 12.46 12Mandible ML 238.8 192.2 209.4 16.15 13Mandible MRH 104.7 79.3 89.1 10.41 15
ANTON ET AL.—SKULLS OF MACHAIRODUS FROM BATALLONES-1 961

lambdoid crests in the larger specimens. The occipital crest ismore marked than in pantherines, and it is surrounded by deepdepressions absent in the latter. The two lateral tubercles ob-served in the superior border of the foramen magnum in pan-therines are absent or barely noticeable in M. aphanistus. Theinsertions for the semispinalis capitis muscle, placed lateral to theinion, are less sculpted than in pantherines.
In ventral view, the median ridge that extends anteriorly fromthe inferior margin of the foramen magnum along the basioc-cipital is more marked than in pantherines, and it reaches thelevel of the anterior end of the tympanic bullae while in thepantherines it only extends for half that distance. At bothsides of that crest there are depressions for the insertion of therectus capitis anticus major muscle, which at least in some speci-mens are more rugose than in pantherines of comparablesize. The shape of the auditory bullae resembles that of modernpantherines, but they are relatively smaller and less inflated.The anterior edge is ahead of the tip of the mastoid processas in pantherines, while in M. giganteus and later machairo-donts that process projects forward beyond the bullae. The con-dyloid and lacerum posterior foramina occupy a common de-pression.
The morphology of the mastoid region is intermediate be-tween that of pantherines and derived machairodontines (Figs.4B-D, I, 5). The paroccipital process is well developed and pro-jects ventrally beyond the level of the tip of the mastoid process,as in pantherines and unlike derived machairodonts like Smilo-don or Homotherium. In lateral view, the paroccipital divergesfrom the mastoid process more than in pantherines, due partly toits more posterior orientation and partly to the greater inclina-tion of the mastoid plane. The mastoid process is much smallerthan in derived machairodonts. It does not project in an antero-inferior direction nearly as far as in later machairodonts, whereit tends to come ever closer to the post-glenoid process, almosttouching it in Smilodon. However, in spite of its moderate de-velopment the mastoid process resembles that of later saber-tooths in being long and low in lateral view, rather than short andtall as in pantherines, and in being delimited by a very devel-oped, laterally flaring mastoid crest, which creates a sort of“roof” over the mastoid area. The insertion of the atlanto-mastoid muscles, which is located under this crest, is thus en-larged, and it becomes oriented more inferiorly and less laterallythan in pantherines. The tip of the mastoid process, with itsmuscle insertions, faces more inferiorly and less anteriorly thanin pantherines, or in derived machairodonts. The auditory me-atus is similar to that of pantherines, while in M. giganteusand later machairodonts it is closely enveloped between thepostglenoid process and the projecting tip of the mastoid pro-cess. The temporal crest is slightly more developed than in pan-therines, forming a sort of roof over the meatus, as in later mach-airodonts.
The postglenoid process is similar to that of pantherines, butthe anterior border of the glenoid fossa is more crest-like, as inlater machairodonts. In ventral view, the fossa is elongated as inpantherines, rather than widening medially as in M. giganteus. Inlateral view, the posterior half of the zygomatic arch is long, low,and moderately curved as in pantherines, rather than short, highand strongly curved as in later machairodonts. The post-orbitalprocess is rounded and not very prominent, and the constrictionjust behind it is not strongly expressed, thus resembling othermachairodonts and differing from pantherines. The crest thatdelimits the insertion area for the masseter in the lower part ofthe zygomatic arch is closer to the lower border than in pan-therines, thus defining a narrower area, as in M. giganteus. Thecrest continues faintly towards the posterior root of M1, definingan area of varying width behind itself (Figs. 4, 5).
In the palatine region, the posterior nasal opening is shaped asa simple arch in ventral view (Fig. 4B), lacking the indentations
seen in pantherines. The palate roof has three ridges, one medialand two lateral, similar in shape and development to those ofpantherines, with a premaxillary section which is markedly crest-like, becoming more blunt in the maxillary and even more so inthe palatine.
The infraorbital foramen is similar in shape and size to that ofpantherines, and it is placed slightly more anterior relative to M1than in the lion, but more posterior than in the leopard. Theforamen opens in an anterolateral direction, as in later mach-airodonts and the lion, while in the leopard and smaller felids itopens more anteriorly, a feature that seems to be size-related.The shape and position of the lacrimal foramina are as in thelion, but in M. aphanistus there is a minute foramen just poste-rior to the lacrimal that is not seen in the former. As in latermachairodonts, and in pantherines and even the domestic cat,there is a small crest just in front of the lacrimal, which is moredeveloped in larger individuals. The sphenopalatine opening ismuch smaller than in the lion, and placed more anteriorly andcloser to the postpalatine foramen than in the latter, thus resem-bling other machairodonts. The oval foramen is placed at thelevel of the greater axis of the glenoid fossa, and is similar in sizeand shape to the same element in the lion and M. giganteus. Theorbital fossa, foramen rotundum and optic foramen cannot beobserved in detail due to the collapse of the orbital wall in allspecimens from Batallones-1.
Mandible
In ventral view, the symphysis does not extend posteriorlynearly as much as in pantherines, thus resembling M. giganteusand other derived machairodonts (Fig. 4E–H, Table 1). As in thelatter, the anterior face of the symphyseal area is more verticalthan in pantherines, and in side view it joins the lower border ofthe mandible at a marked angle, while in pantherines there is nosuch angle but rather a gentle curve. In frontal view, the anteriorface of the mandible is high and rectangular, with a verticalgroove in the midline, while in pantherines it is lower and thereis a central ridge instead of a groove. Still in frontal view, thelower outline of the symphysis is concave in M. aphanistus andderived machairodonts, while in pantherines it is convex. Thereare several paired depressions at either side of the midline, twoor three pairs of large ones in M. aphanistus and other mach-airodonts, one large pair and several smaller ones below it inpantherines. There are three mental foramina, a larger one atabout the level of the middle of the c–p3 diastema and twosmaller ones behind it, one on top of the other. This is the com-mon condition in most felids, but Smilodon may lack the upperone of the two smaller foramina. The postcanine diastema isproportionally longer than in pantherines, but shorter than in M.giganteus. Its dorsal border rises gently towards the anterior as inpantherines, while in derived machairodonts it rises moresteeply. As with the premaxilla, the size and setting of the largelymissing lower incisors is well recorded by the alveoli, which showthat these teeth were small and crowded into a more or lessstraight row between the large and oval alveoli of the lowercanines.
The coronoid process is large and posteriorly inclined as inpantherines, rather than reduced and more vertically oriented asin M. giganteus and other derived machairodonts. The massetericfossa is strongly excavated as in felids in general, and it resemblespantherines and differs from derived machairodonts in lacking astrong crest on its postero-ventral margin. The angular processprojects in a postero-dorsal direction as in M. giganteus, while inpantherines it projects more postero-ventrally and curves medi-ally. The mandibular condyle is similar to that of pantherines,while in M. giganteus and derived machairodonts there is amarked crest extending from its latero-ventral border throughthe posterior border of the mandible, marking the posterior mar-
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 24, NO. 4, 2004962
gin of the masseteric fossa. The mandibular foramen is like thatof pantherines in shape and position.
Upper Dentition
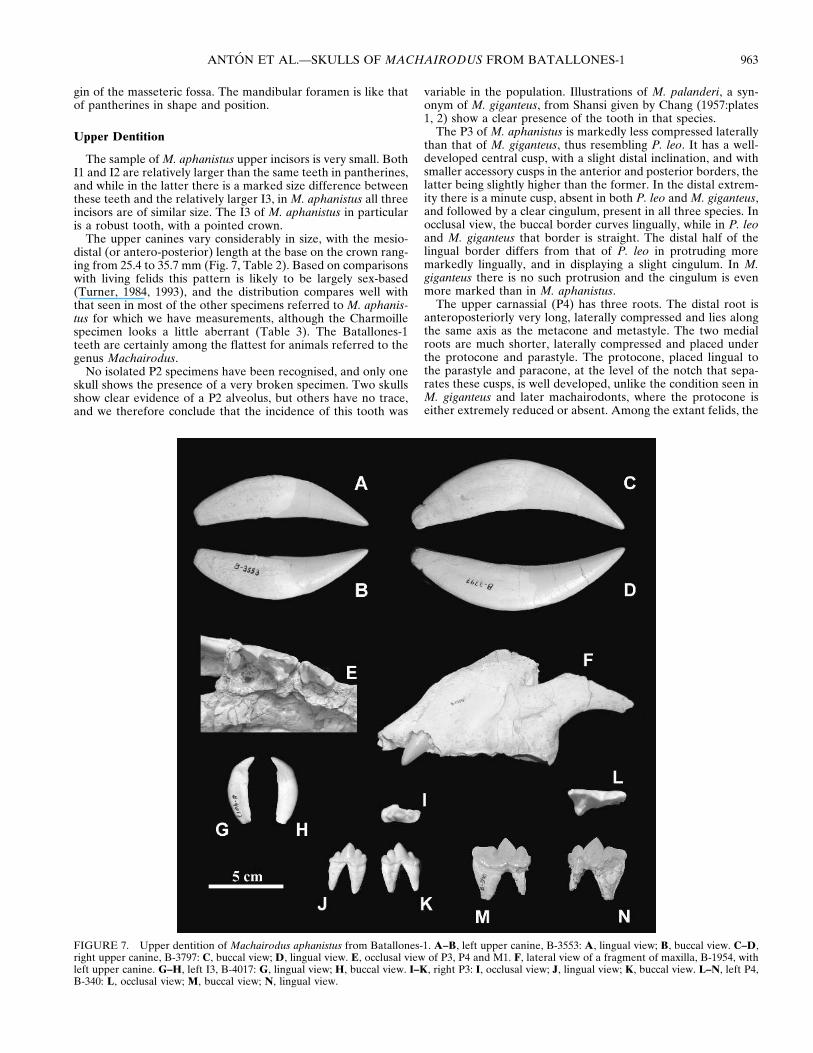
The sample of M. aphanistus upper incisors is very small. BothI1 and I2 are relatively larger than the same teeth in pantherines,and while in the latter there is a marked size difference betweenthese teeth and the relatively larger I3, in M. aphanistus all threeincisors are of similar size. The I3 of M. aphanistus in particularis a robust tooth, with a pointed crown.
The upper canines vary considerably in size, with the mesio-distal (or antero-posterior) length at the base on the crown rang-ing from 25.4 to 35.7 mm (Fig. 7, Table 2). Based on comparisonswith living felids this pattern is likely to be largely sex-based(Turner, 1984, 1993), and the distribution compares well withthat seen in most of the other specimens referred to M. aphanis-tus for which we have measurements, although the Charmoillespecimen looks a little aberrant (Table 3). The Batallones-1teeth are certainly among the flattest for animals referred to thegenus Machairodus.
No isolated P2 specimens have been recognised, and only oneskull shows the presence of a very broken specimen. Two skullsshow clear evidence of a P2 alveolus, but others have no trace,and we therefore conclude that the incidence of this tooth was
variable in the population. Illustrations of M. palanderi, a syn-onym of M. giganteus, from Shansi given by Chang (1957:plates1, 2) show a clear presence of the tooth in that species.
The P3 of M. aphanistus is markedly less compressed laterallythan that of M. giganteus, thus resembling P. leo. It has a well-developed central cusp, with a slight distal inclination, and withsmaller accessory cusps in the anterior and posterior borders, thelatter being slightly higher than the former. In the distal extrem-ity there is a minute cusp, absent in both P. leo and M. giganteus,and followed by a clear cingulum, present in all three species. Inocclusal view, the buccal border curves lingually, while in P. leoand M. giganteus that border is straight. The distal half of thelingual border differs from that of P. leo in protruding moremarkedly lingually, and in displaying a slight cingulum. In M.giganteus there is no such protrusion and the cingulum is evenmore marked than in M. aphanistus.
The upper carnassial (P4) has three roots. The distal root isanteroposteriorly very long, laterally compressed and lies alongthe same axis as the metacone and metastyle. The two medialroots are much shorter, laterally compressed and placed underthe protocone and parastyle. The protocone, placed lingual tothe parastyle and paracone, at the level of the notch that sepa-rates these cusps, is well developed, unlike the condition seen inM. giganteus and later machairodonts, where the protocone iseither extremely reduced or absent. Among the extant felids, the
FIGURE 7. Upper dentition of Machairodus aphanistus from Batallones-1. A–B, left upper canine, B-3553: A, lingual view; B, buccal view. C–D,right upper canine, B-3797: C, buccal view; D, lingual view. E, occlusal view of P3, P4 and M1. F, lateral view of a fragment of maxilla, B-1954, withleft upper canine. G–H, left I3, B-4017: G, lingual view; H, buccal view. I–K, right P3: I, occlusal view; J, lingual view; K, buccal view. L–N, left P4,B-340: L, occlusal view; M, buccal view; N, lingual view.
ANTON ET AL.—SKULLS OF MACHAIRODUS FROM BATALLONES-1 963
protocone is well developed in all species except in the cheetah,Acinonyx jubatus (Ficcarelli, 1984). As in later machairodonts,the P4 of M. aphanistus displays a well-developed ectostyle,again resembling A. jubatus and unlike all other extant felines.The combined length of metacone and metastyle is longer thanthat of the paracone, as in other machairodonts and unlike pan-therine cats. The paracone and metacone form an angle similarto that observed in pantherines, and thus greater than in latermachairodonts.
Lower Dentition
The lower incisors of M. aphanistus, although again few innumber in the sample, are relatively small, thus resembling thesame teeth in P. leo to a higher degree than in the case of theupper incisors. The width at the level of the alveolus is propor-tionally smaller than in the upper incisors, and the mesial borderof the teeth are gently curved, unlike the case of pantherines,where they are straighter.
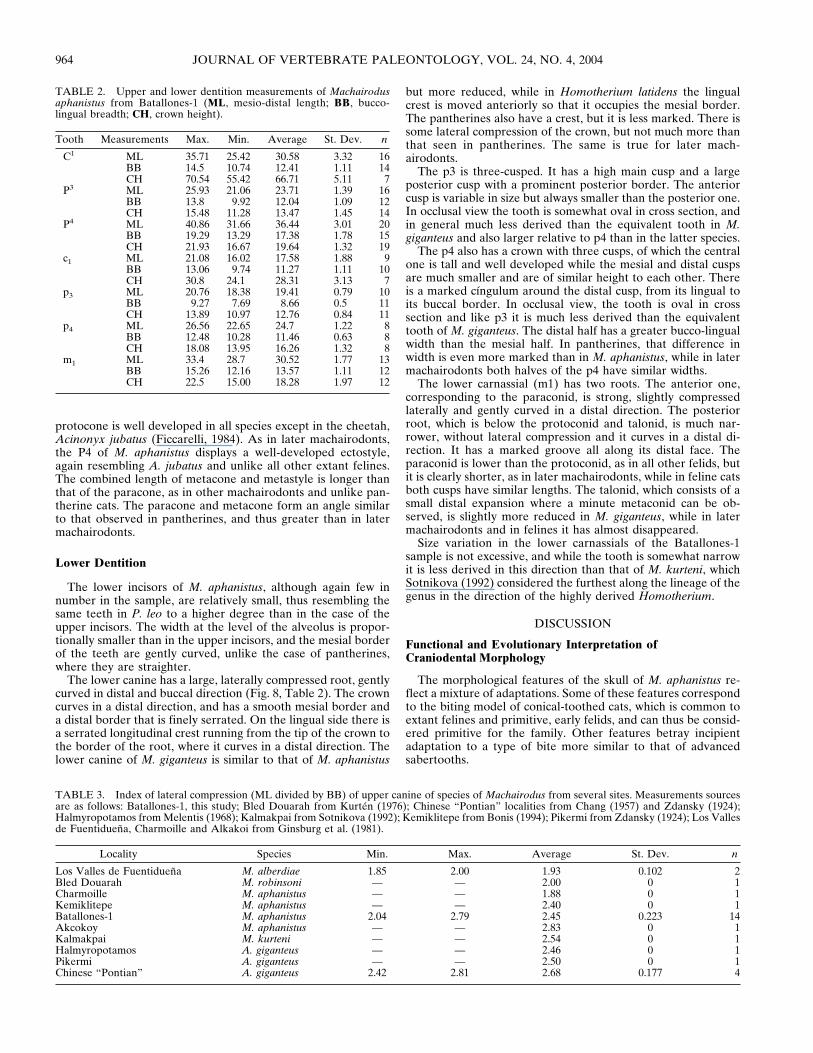
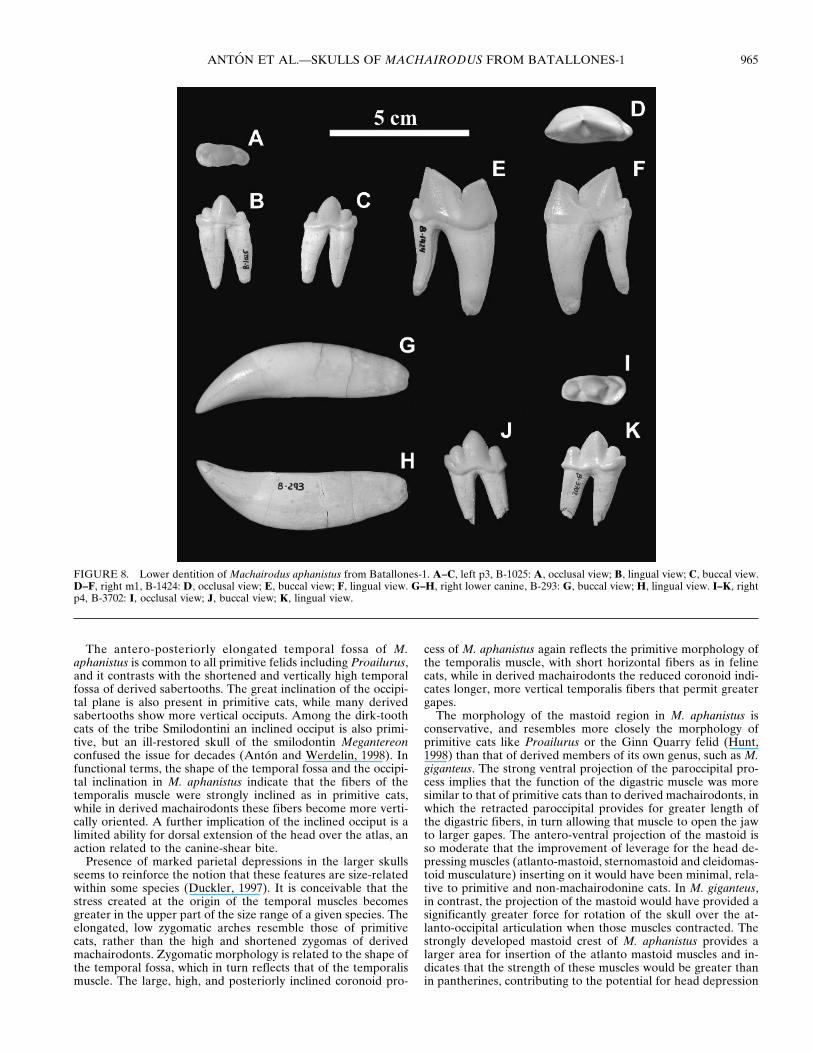
The lower canine has a large, laterally compressed root, gentlycurved in distal and buccal direction (Fig. 8, Table 2). The crowncurves in a distal direction, and has a smooth mesial border anda distal border that is finely serrated. On the lingual side there isa serrated longitudinal crest running from the tip of the crown tothe border of the root, where it curves in a distal direction. Thelower canine of M. giganteus is similar to that of M. aphanistus
but more reduced, while in Homotherium latidens the lingualcrest is moved anteriorly so that it occupies the mesial border.The pantherines also have a crest, but it is less marked. There issome lateral compression of the crown, but not much more thanthat seen in pantherines. The same is true for later mach-airodonts.
The p3 is three-cusped. It has a high main cusp and a largeposterior cusp with a prominent posterior border. The anteriorcusp is variable in size but always smaller than the posterior one.In occlusal view the tooth is somewhat oval in cross section, andin general much less derived than the equivalent tooth in M.giganteus and also larger relative to p4 than in the latter species.
The p4 also has a crown with three cusps, of which the centralone is tall and well developed while the mesial and distal cuspsare much smaller and are of similar height to each other. Thereis a marked cıngulum around the distal cusp, from its lingual toits buccal border. In occlusal view, the tooth is oval in crosssection and like p3 it is much less derived than the equivalenttooth of M. giganteus. The distal half has a greater bucco-lingualwidth than the mesial half. In pantherines, that difference inwidth is even more marked than in M. aphanistus, while in latermachairodonts both halves of the p4 have similar widths.
The lower carnassial (m1) has two roots. The anterior one,corresponding to the paraconid, is strong, slightly compressedlaterally and gently curved in a distal direction. The posteriorroot, which is below the protoconid and talonid, is much nar-rower, without lateral compression and it curves in a distal di-rection. It has a marked groove all along its distal face. Theparaconid is lower than the protoconid, as in all other felids, butit is clearly shorter, as in later machairodonts, while in feline catsboth cusps have similar lengths. The talonid, which consists of asmall distal expansion where a minute metaconid can be ob-served, is slightly more reduced in M. giganteus, while in latermachairodonts and in felines it has almost disappeared.
Size variation in the lower carnassials of the Batallones-1sample is not excessive, and while the tooth is somewhat narrowit is less derived in this direction than that of M. kurteni, whichSotnikova (1992) considered the furthest along the lineage of thegenus in the direction of the highly derived Homotherium.
DISCUSSION
Functional and Evolutionary Interpretation ofCraniodental Morphology
The morphological features of the skull of M. aphanistus re-flect a mixture of adaptations. Some of these features correspondto the biting model of conical-toothed cats, which is common toextant felines and primitive, early felids, and can thus be consid-ered primitive for the family. Other features betray incipientadaptation to a type of bite more similar to that of advancedsabertooths.
TABLE 2. Upper and lower dentition measurements of Machairodusaphanistus from Batallones-1 (ML, mesio-distal length; BB, bucco-lingual breadth; CH, crown height).
Tooth Measurements Max. Min. Average St. Dev. n
C1 ML 35.71 25.42 30.58 3.32 16BB 14.5 10.74 12.41 1.11 14CH 70.54 55.42 66.71 5.11 7
P3 ML 25.93 21.06 23.71 1.39 16BB 13.8 9.92 12.04 1.09 12CH 15.48 11.28 13.47 1.45 14
P4 ML 40.86 31.66 36.44 3.01 20BB 19.29 13.29 17.38 1.78 15CH 21.93 16.67 19.64 1.32 19
c1 ML 21.08 16.02 17.58 1.88 9BB 13.06 9.74 11.27 1.11 10CH 30.8 24.1 28.31 3.13 7
p3 ML 20.76 18.38 19.41 0.79 10BB 9.27 7.69 8.66 0.5 11CH 13.89 10.97 12.76 0.84 11
p4 ML 26.56 22.65 24.7 1.22 8BB 12.48 10.28 11.46 0.63 8CH 18.08 13.95 16.26 1.32 8
m1 ML 33.4 28.7 30.52 1.77 13BB 15.26 12.16 13.57 1.11 12CH 22.5 15.00 18.28 1.97 12
TABLE 3. Index of lateral compression (ML divided by BB) of upper canine of species of Machairodus from several sites. Measurements sourcesare as follows: Batallones-1, this study; Bled Douarah from Kurten (1976); Chinese “Pontian” localities from Chang (1957) and Zdansky (1924);Halmyropotamos from Melentis (1968); Kalmakpai from Sotnikova (1992); Kemiklitepe from Bonis (1994); Pikermi from Zdansky (1924); Los Vallesde Fuentiduena, Charmoille and Alkakoi from Ginsburg et al. (1981).
Locality Species Min. Max. Average St. Dev. n
Los Valles de Fuentiduena M. alberdiae 1.85 2.00 1.93 0.102 2Bled Douarah M. robinsoni — — 2.00 0 1Charmoille M. aphanistus — — 1.88 0 1Kemiklitepe M. aphanistus — — 2.40 0 1Batallones-1 M. aphanistus 2.04 2.79 2.45 0.223 14Akcokoy M. aphanistus — — 2.83 0 1Kalmakpai M. kurteni — — 2.54 0 1Halmyropotamos A. giganteus — — 2.46 0 1Pikermi A. giganteus — — 2.50 0 1Chinese “Pontian” A. giganteus 2.42 2.81 2.68 0.177 4
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 24, NO. 4, 2004964
The antero-posteriorly elongated temporal fossa of M.aphanistus is common to all primitive felids including Proailurus,and it contrasts with the shortened and vertically high temporalfossa of derived sabertooths. The great inclination of the occipi-tal plane is also present in primitive cats, while many derivedsabertooths show more vertical occiputs. Among the dirk-toothcats of the tribe Smilodontini an inclined occiput is also primi-tive, but an ill-restored skull of the smilodontin Megantereonconfused the issue for decades (Anton and Werdelin, 1998). Infunctional terms, the shape of the temporal fossa and the occipi-tal inclination in M. aphanistus indicate that the fibers of thetemporalis muscle were strongly inclined as in primitive cats,while in derived machairodonts these fibers become more verti-cally oriented. A further implication of the inclined occiput is alimited ability for dorsal extension of the head over the atlas, anaction related to the canine-shear bite.
Presence of marked parietal depressions in the larger skullsseems to reinforce the notion that these features are size-relatedwithin some species (Duckler, 1997). It is conceivable that thestress created at the origin of the temporal muscles becomesgreater in the upper part of the size range of a given species. Theelongated, low zygomatic arches resemble those of primitivecats, rather than the high and shortened zygomas of derivedmachairodonts. Zygomatic morphology is related to the shape ofthe temporal fossa, which in turn reflects that of the temporalismuscle. The large, high, and posteriorly inclined coronoid pro-
cess of M. aphanistus again reflects the primitive morphology ofthe temporalis muscle, with short horizontal fibers as in felinecats, while in derived machairodonts the reduced coronoid indi-cates longer, more vertical temporalis fibers that permit greatergapes.
The morphology of the mastoid region in M. aphanistus isconservative, and resembles more closely the morphology ofprimitive cats like Proailurus or the Ginn Quarry felid (Hunt,1998) than that of derived members of its own genus, such as M.giganteus. The strong ventral projection of the paroccipital pro-cess implies that the function of the digastric muscle was moresimilar to that of primitive cats than to derived machairodonts, inwhich the retracted paroccipital provides for greater length ofthe digastric fibers, in turn allowing that muscle to open the jawto larger gapes. The antero-ventral projection of the mastoid isso moderate that the improvement of leverage for the head de-pressing muscles (atlanto-mastoid, sternomastoid and cleidomas-toid musculature) inserting on it would have been minimal, rela-tive to primitive and non-machairodonine cats. In M. giganteus,in contrast, the projection of the mastoid would have provided asignificantly greater force for rotation of the skull over the at-lanto-occipital articulation when those muscles contracted. Thestrongly developed mastoid crest of M. aphanistus provides alarger area for insertion of the atlanto mastoid muscles and in-dicates that the strength of these muscles would be greater thanin pantherines, contributing to the potential for head depression
FIGURE 8. Lower dentition of Machairodus aphanistus from Batallones-1. A–C, left p3, B-1025: A, occlusal view; B, lingual view; C, buccal view.D–F, right m1, B-1424: D, occlusal view; E, buccal view; F, lingual view. G–H, right lower canine, B-293: G, buccal view; H, lingual view. I–K, rightp4, B-3702: I, occlusal view; J, buccal view; K, lingual view.
ANTON ET AL.—SKULLS OF MACHAIRODUS FROM BATALLONES-1 965

but with less efficiency than in more derived machairodonts. Inthe basicranium, the insertion of the head-depressing muscles iswell marked, with a median crest, and suggests a possible adap-tation towards the canine shear-bite.
There is no sign of a well-developed incisor arc in the premax-illa, a feature in which the Batallones-1 material is less derivedthan other machairodonts, while the incisors are relatively primi-tive, a situation that changes markedly in other machairodonts.This indicates that the action of the incisors was less independentof the maxilla and the canines, in line with the arguments putforward by Biknevicius et al. (1996). We estimate that the heightof the upper canine crowns in the sample necessitates a man-dibular gape 10° greater than the usual figure of about 70° thatis recorded for the lion (Anton et al., 1998). This is well withinthe range inferred for M. giganteus when we take account of thegreater height of the lower canines in M. aphanistus despite thefact that the adaptations that would facilitate that gape are lessdeveloped in the Batallones-1 specimens. The canines are atleast as flattened as those of M. giganteus (Table 3), and thusequally fragile and breakable (Van Valkenburgh, 1988; VanValkenburgh and Ruff, 1987). The more derived species protectthose canines by means of the greater development of the inci-sors, which stabilize the bite area, and by means of behavioralfactors that minimize the chances of hitting bone, such as theselection of larger prey and biting the throat rather than the backof the neck after immobilization, as interpreted by Turner andAnton (1997), and Anton and Galobart (1999).
Size Variation
We have remarked on the variation in size of the cranium andupper canine and the likelihood that this is in large part anindication of sexual dimorphism. We calculated the sexual di-morphism index following Van Valkenburgh and Sacco (2002).Sexual dimorphism in mammals has been associated with a rela-tively intense male competition for access to females (Short andBalaban, 1994; Weckerly, 1998). Among carnivores, it has beenshown that sexual dimorphism is mainly noted in the size of theupper canines, and it is directly associated with the breedingsystem, being higher in species in which a male defends a groupof females (Gittleman and Van Valkenburgh, 1997; Weckerly,1998). Table 4 shows the sexual dimorphism index in severalspecies of Felidae as inferred from four different measurements.It is evident that lions and leopards are the most dimorphicamong the extant felids, whereas M. aphanistus shows a highindex, similar to these species. Lions are social felids (Alderton,
1998; Bothma and Walker, 1999), but leopards are solitary ani-mals (Bailey, 1993) implying, as Van Valkenburg and Sacco(2002) have warned, that such high degree of sexual dimorphismdoes not indicate the presence of group behavior. Solitary be-havior is the rule among modern felids with lions being the onlytruly social members of the family, so it appears reasonable toconsider M. aphanistus as a solitary hunter as well. The highsexual dimorphism index would be associated with a low level oftolerance between males, as in the leopard, in which the range ofa male never overlaps with the territories of other males (Bailey,1993). Other sabertooth cats, such as Smilodon (Van Valken-burgh and Sacco, 2002) or Paramachairodus (Salesa, 2002) ex-hibit smaller sexual dimorphism indexes, which have been asso-ciated with some degree of tolerance between males, as in thecase of jaguars (Rabinowitz and Nottingham, 1986) among ex-tant felids.
Predatory Behavior and the Killing Bite inMachairodus aphanistus
The morphological features discussed above suggest a modelof predatory behavior that differs not only from that of extantpantherines but also from that hypothesized for derived mach-airodonts such as Homotherium and Smilodon, as outlined in theintroduction. On the one hand, there are several features indi-cating that M. aphanistus killed its prey by penetrating the fleshof the throat with its canines and causing massive blood loss, justas has been inferred for the derived sabertooths. These featuresinclude the high crowned, flattened and serrated upper canines,which are well adapted to penetrate the flesh of prey and wouldbe inadequate for either a crushing nape bite or a suffocatingthroat bite; the crest-like shape of the anterior border of theglenoid fossa, which allows for a slightly enlarged gape; the at-lanto-mastoid muscles inserting on the enlarged mastoid crests,which would have contributed to the downward pull of the head;the slight anteroposterior projection of the mastoid process,which would have marginally improved the head depressing ac-tion of the cephalohumeralis and of the atlanto-mastoid muscu-lature; and the well-developed insertion areas of the rectus capi-tis anticus major on the basisphenoid, which suggest that thesemuscles made a large contribution to head depression.
On the other hand there are several features indicating thatthe force applied to both upper and lower canines through con-traction of the jaw-closing musculature at moderate gapes pro-vided the main power for the bite in M. aphanistus, just as inpantherines and other modern cats. These include: insertion ar-eas of the masseter and temporalis, which show no sign of modi-fication for larger gapes; the shortness and horizontal orientationof the temporalis fibers, which provide for a strong bite at lowgapes, but would have worked poorly at larger ones; the greatheight of the crowns of the lower canines, which limited theeffective gape, but were obviously a more important part of thebite than in derived sabertooths; and the flattening of the lowercanines, which suggest an action similar and complementary tothat of the upper ones, while in derived sabertooths the lowercanines become more incisor-like and tend to act as part of thelower incisor battery.
What differences do these characters imply, in terms of preyprocurement and dispatch, between M. aphanistus and latermachairodonts, including M. giganteus? If a throat bite was themain killing technique of sabertooth cats, then the larger theprey the larger the necessary gape. The more advanced adapta-tion for gape in M. giganteus would have allowed it to bite thenecks of larger prey, such as giraffids or even rhinocerotids,while it is possible that a horse or antelope was the largest preythat M. aphanistus could kill efficiently. It is also conceivablethat, even for prey of similar size, a sabertooth cat with refinedadaptations for the canine-shear bite would have killed with
TABLE 4. Sexual dimorphism index for several species of extant andfossil felids. Based on Van Valkenburgh and Sacco (2002), and Salesa(2002) and supplemented with original data for M. aphanistus fromBatallones-1 (BL, skull basal length; ML, mesio-distal length; BB, buc-colingual breadth).
SpeciesLB
(Skull)ML
(upper C)BB
(upper C)ML(m1) Average
C. caracal 1.08 1.06 1.03 1.08 1.06L. serval 1.08 1.10 1.14 0.97 1.07S. fatalis 1.06 1.09 1.12 1.07 1.09P. onca 1.06 1.12 1.11 1.06 1.09P. concolor 1.08 1.14 1.09 1.04 1.09A. jubatus 1.10 1.15 1.11 1.03 1.10P. ogygia 1.09 1.11 1.11 1.09 1.10L. rufus 1.10 1.16 1.11 1.06 1.11F. silvestris 1.10 1.15 1.13 1.08 1.12F. chaus 1.07 1.16 1.15 1.10 1.12P. tigris 1.16 1.16 1.08 1.12 1.13M. aphanistus 1.24 1.22 1.16 1.10 1.18P. leo 1.12 1.25 1.23 1.13 1.18P. pardus 1.13 1.24 1.26 1.12 1.19
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 24, NO. 4, 2004966

more efficiency, meaning that a higher percentage of bite at-tempts would result in a kill and that prey would die faster,implying less struggling time and less opportunity for accidentsinvolving teeth breakage (Van Valkenburgh, 1988; Van Valken-burgh and Ruff, 1987) and/or prey escape.
It was recently proposed (Bakker, 1998) that saber-like teethevolved convergently in sabertoothed mammals and allosauriddinosaurs as an adaptation for predation on prey species ten ormore times heavier than the predators. On the contrary, evi-dence of limited adaptation for gape and primitive biting mecha-nism in M. aphanistus suggest that, at least in this taxon, saber-like canines originally evolved as an adaptation for killing preywithin the size range of those killed by modern felids of compa-rable size. The key advantage of the initial development of saber-like teeth would lie in the efficiency of a killing bite that causedmassive blood loss instead of suffocation, rather than in the pos-sibility of taking significantly larger prey. It is feasible, however,that more derived sabertooths with enhanced adaptations forgape, would have been able to take prey larger relative to theirbody size.
Machairodus aphanistus as a Key to UnderstandingSabertoothed Cat Evolution
The model of predatory behavior proposed above for M.aphanistus stems from a combination of morphological featuresthat has no parallel among the known species of cat or cat-likepredators living or extinct, except perhaps for the North-American species Nimravides catocopis, which may itself be asynonym of M. aphanistus according to Beaumont (1978, 1980).Among the Felidae, slight machairodont adaptations are alsodisplayed by members of the tribe Metailurini (Turner andAnton, 1997; Crusafont and Aguirre, 1972), but in these animalsthe moderate adaptations of the skull are paralleled by similarlymoderate upper canines, which have relatively much lowercrowns and are less flattened than in M. aphanistus. As a sequen-tial reconstruction of the head shows (Fig. 9) the primitive skullproportions of M. aphanistus imply a thoroughly cat-like lifeappearance, in contrast with the more straight profiles of derivedmachairodonts.
The presence of highly derived upper canines and a primitivecranio-mandibular morphology in M. aphanistus demonstratesthat while high-crowned, flattened sabers may work best as partof a complex of adaptations for the canine-shear bite, they canalso work, and did work sufficiently well, within a different, moremodern-cat like biomechanical context. It also contradicts thenotion, mentioned by Dawson et al. (1986) that the complex ofcharacters seen in all sabertoothed carnivores was under “strongpleiotropic control.” On the contrary, the evidence from Batal-lones suggests that there was no tight linkage between the actionof the genes that controlled the shape of the upper canines in M.aphanistus and those controlling the other “sabertooth charac-ters,” and it is obvious that the latter have evolved in each lin-eage of sabertoothed mammalian predators with full indepen-dence, making the precise similarity of adaptations all the moreremarkable.
In systematic terms, the new material from Batallones-1 showsvery important differences in craniomandibular morphology,which, in our view, would amply justify generic separation be-tween M. aphanistus and the Turolian species M. giganteus. Thedifferences in degree of machairodont specialization are at leastas important as those separating M. giganteus from the Plioceneand Pleistocene genus Homotherium. Since M. aphanistus is thetype species of the genus, we propose that M. giganteus be re-moved from it and placed in a genus of its own. The nameAmphimachairodus, originally coined as a generic name by Kret-zoi (1929) and proposed by Beaumont (1975) as a subgenericname for M. giganteus, is available and should in our opinion be
raised to generic status, a proposal already made by Morales(1984) on the basis of dental characters but now vastly reinforcedby craniomandibular features of M. aphanistus described in thisstudy. Keeping M. aphanistus and M. giganteus lumped in thesame genus seems unrealistic, and a clear understatement of thediversity of machairodontine taxa in the late Miocene of Eurasia.
ACKNOWLEDGMENTS
This study is part of the research project BTE2002–00410 (Di-reccion General de Investigacion-MCYT). We thank the Comu-nidad Autonoma de Madrid (Direccion General de PatrimonioHistorico) for the continuous funding support and the researchpermission. Additional support was provided by the NationalGeographic Society (Grant 6964–01).
Alan Turner thanks the British Council for assistance withcosts involved in visiting colleagues and collections in Spain. Weare grateful to Blaire Van Valkenburgh and Lars Werdelin fortheir constructive comments.
FIGURE 9. Sequential reconstruction of the head of Machairodusaphanistus in side view, based on Batallones-1 material: top, recon-structed skull; middle, restoration of the musculature, nasal and ear car-tilages, eyeball and whisker pad; bottom, reconstructed external appear-ance. Reconstruction methodology follows Anton et al. (1998).
ANTON ET AL.—SKULLS OF MACHAIRODUS FROM BATALLONES-1 967

LITERATURE CITED
Agustı, J. and M. Anton. 2002. Mammoths, Sabertooths, and Hominids.Columbia University Press, New York, 313 pp.
Akersten, W. 1985. Canine function in Smilodon (Mammalia, Felidae,Machairodontinae). Los Angeles County Museum Contributions inScience 356:1–22.
Alderton, D. 1998. Wild Cats of the World. Blandford, Hong Kong, 192pp.
Anton, M., and A. Galobart. 1999. Neck function and predatory behav-iour in the scimitar toothed cat Homotherium latidens (Owen). Jour-nal of Vertebrate Paleontology 19:771–784.
Anton, M., R. Garcıa-Perea, and A. Turner. 1998. Reconstructed facialappearance of the Sabretoothed felid Smilodon. Zoological Journalof the Linnean Society 124:369–386.
Anton, M., and J. Morales. 2000. Inferencias paleoecologicas de la aso-ciacion de carnıvoros del yacimiento de Cerro Batallones; pp.190–201 in J. Morales, M. Nieto, L. Amezua, S. Fraile, E. Gomez, E.Herraez, P. Pelaez-Campomanes, M. J. Salesa, I. M. Sanchez and D.Soria (eds.), Patrimonio Paleontologico de la Communidad deMadrid. Comunidad de Madrid, Madrid.
Anton, M., and L. Werdelin. 1998. Too well restored? The case of theMegantereon skull from Seneze. Lethaia 31:158–160.
Bailey, T. N. 1993. The African Leopard: Ecology and Behavior of aSolitary Felid. Biology and Resource Management in the TropicsSeries. Columbia University Press, New York, 429 pp.
Ballesio, R. 1963. Monographie d’un Machairodus du gisement villa-franchien de Seneze: Homotherium crenatidens Fabrini. Traveauxdu Laboratoire de Geologie de la Faculte de Sciences de Lyon 9:1–129.
Bakker, R. T. 1998. Brontosaur killers: late Jurassic allosaurids as sabre-tooth cat analogues. Gaia 15:145–158.
Barone, R. 1967. La myologie du lion (Panthera leo). Mammalia 31:459–516.
Barone, R. 1989. Anatomie Comparee des Mammiferes Domestiques.Tomes 1 et 2. Editions Vigot, Paris, 761 and 984 pp.
Beaumont, G. 1975. Recherches sur les felides (mammiferes, carnivores)du Pliocene inferieur des sables a Dinotherium des environsd’Eppelsheim (Rheinhessen). Archives des Sciences 28:369–405.
Beaumont, G. 1978. Notes complementaires sur quelques felides (carni-vores). Archives des Sciences, Geneve 31:219–227.
Beaumont, G. 1980. Contribution to the study of the genus Nimravides(Kitts) (Mammalia, Carnivora, Felidae). The species N. pedionomus(MacDonald). Archives des Sciences, Geneve 43:123–157.
Biknevicius, A. R., B. Van Valkenburgh, and J. Walker. 1996. Incisorsize and shape: implications for feeding behaviors in saber-toothed“cats.” Journal of Vertebrate Paleontology 16:510–521.
Bohlin, B. 1947. The sabre-toothed tigers once more. Bulletin of theGeological Institute of the University of Uppsala 32:11–20.
Bonis, L. de. 1994. Les gisements de mammiferes du Miocene superieurde Kemiklitepe, Turquie: 2 Carnivores. Bulletin du Museum na-tional dHistoire Naturelle, Paris 16:19–39.
Bothma, J., and C. Walker. 1999. Larger Carnivores of the African Sa-vannas. Springer-Verlag, Berlin, 274 pp.
Bowdich, T. E. 1821. An analysis of the natural classifications of Mam-malia for the use of students and travellers. J. Smith, Paris, 115 pp.
Bryant, H. N. 1996. Force generation by the jaw adductor musculature atdifferent gapes in the Pleistocene sabretoothed felid Smilodon;pp. 283–299 in K. M. Stewart and K. L. Seymour (eds.), Paleoecologyand Paleoenvironments of Late Cenozoic Mammals. University ofToronto Press, Toronto.
Chang, H. 1957. On new material of some machairodonts of Pontian agefrom Shansi. Vertebrata Palasiatica 1:193–200.
Crusafont Pairo, M., and E. Aguirre. 1972. Stenailurus, felide nouveau,du Turolien d’Espagne. Annales de Paleontologies Vertebres 58:211–223.
Dawson, M. R., R. K. Stucky, L. Krishtalka, and C. C. Black. 1986.Machaeroides simpsoni, new species, oldest known sabertooth cre-odont (Mammalia), of Lost Cabin Eocene. Contributions to Geol-ogy, University of Wyoming, Special Paper 3:177–182.
Duckler, G. L. 1997. Parietal depressions in skulls of the extinct saber-toothed felid Smilodon fatalis: evidence of mechanical strain. Jour-nal of Vertebrate Paleontology 17:600–609.
Emerson, S., and L. Radinsky. 1980. Functional analysis of sabertoothcranial morphology. Paleobiology 6:441–446.
Ficcarelli, G. 1984. The villafranchian cheetahs from Tuscany and re-marks on the dispersal and evolution of the genus Acinonyx. Palae-ontographia Italica 73:94–103.
Fischer, G. 1817. Adversaria Zoologica. Memoirs of the Society Imperialof Naturalists, Moscow 5:368–428.
Gill, T. 1872. Arrangement of the families of mammals with analyticaltables. Smithsonian Miscellaneous Collections 11:1–98.
Ginsburg, L., J. Morales, and D. Soria. 1981. Nuevos datos sobres loscarnivoros de Los Valles de Fuentiduena, Segovia. Estudios Geo-logicos 37:383–415.
Gittleman, J. L., and B. van Valkenburgh. 1997. Sexual dimorphism inthe canines and skulls of carnivores: effects of size, phylogeny, andbehavioural ecology. Journal of Zoology 242:97–117.
Gorniak, G. C., and C. Gans. 1980. Quantitative assay of electromyo-grams during mastication in domestic cats (Felis catus). Journal ofMorphology 163:253–281.
Hunt, R. M. 1998. Evolution of the aeluroid Carnivora: diversity of theearliest aeluroids from Eurasia (Quercy, Hsanda-Gol) and the ori-gin of felids. American Museum Novitates 3252:1–65.
Kaup, J. J. 1832. Vier neue Arten urweltlicher Raubthiere welche imzoologischen Museum zu Darmstadt aufbewart werden. Archivesfur Mineralogie 5:150–158.
Kaup, J. J. 1833. Description dOssemens fossiles de Mammiferes incon-nus jusqua present qui se trouvent au Museum grand ducal deDarmstadt. 2 volumes. Darmstadt.
Kretzoi, M. 1929. Materialen zur phylogenetischen klassifikation der Ae-luroideen. 10th International Zoological Congress, Budapest 1927:1293–1355.
Kurten, B. 1952. The Chinese Hipparion fauna: a quantitative surveywith comments on the ecology of the machairodonts and hyaenidsand the taxonomy of the gazelles. Commentationes Biologicae 13(4):1–82.
Kurten, B. 1976. Fossil Carnivora from the Late Tertiary of BledDouarah and Cherichira, Tunisia. Notes du Service Geologique deTunisie 42:177–214.
Leyhausen, P. 1979. Cat Behavior: the Predatory and Social Behavior ofDomestic and Wild Cats. Garland STPM Press, New York, 340 pp.
Linnaeus, C. 1758. Systema Naturae per regna tria naturae, secundumclassis, ordines, genera, species cum characteribus, differentiis, syn-onymis, locis. Laurentii Salvii, Stockholm, 824 pp.
McKenna, M. C., and S. K. Bell. 1997. Classification of Mammals abovethe Species Level. Columbia University Press, New York, 631 pp.
Melentis, J. K. 1968. Studien uber fossile vertebraten griechenlands: diePikermi fauna von Halmyropotamos (Euboa, Griechenland). 1.Odontologie und Craniologie. Annales Geologiques des Pays Hel-leniques 19:283–411.
Merrian, J. C. and Stock, C. 1932. The Felidae of Rancho La Brea.Carnegie Institution of Washington 22:1–231.
Morales, J. 1984. Venta de Moro: su macrofauna de mamiferos, ybiostratigrafia continental del Mioceno terminal Mediterraneo. Uni-versidad Complutense de Madrid, Madrid, 340 pp.
Morales, J., M. Nieto, M. Kholer, and S. Moya-Sola. 1999. Large mam-mals from the Vallesian of Spain; pp. 113–126 in J. Agusti, L. Rookand P. Andrews (eds.), The Evolution of Neogene Terrestrial Eco-systems in Europe. Cambridge University Press, Cambridge.
Morales, J., L. Alcala, L. Amezua, M. Anton, S. Fraile, E. Gomez,P. Montoya, M. Nieto, B. Perez, M. J. Salesa, and I. M. Sanchez.2000. El yacimiento del Cerro de los Batallones; pp. 179–201 in J.Morales, M. Nieto, L. Amezua, S. Fraile, E. Gomez, E. Herraez, P.Pelaez-Campomanes, M. J. Salesa, I. M. Sanchez and D. Soria (eds.),Patrimonio Paleontologico de la Comunidad de Madrid. Comuni-dad de Madrid, Madrid.
Rabinowitz, A. R., and B. G. Nottingham. 1986. Ecology and behaviourof the Jaguar (Panthera onca) in Belize, Central America. Journal ofZoology 210:149–159.
Riabinin, A. 1929. Faune de mammiferes de Taraklia. Carnivore vera,Rodentia, Subungulata. Travaux du Musee de Geologie de Lenin-grad 5.
Salesa, M. J. 2002. Estudio Anatomico, Biomecanico, Paleoecologico yFilogenetico de Paramachairodus ogygia (Kaup, 1832) Pilgrim, 1913(Felidae, Machairodontinae) del yacimiento vallesiense (Miocenosuperior) de Batallones-1 (Torrejon de Velasco, Madrid). Unpub-lished Ph.D. thesis. Departamento de Biologıa Animal I, Facultadde Ciencias Biologicas, Universidad Complutense de Madrid,Madrid, 370 pp.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 24, NO. 4, 2004968

Schaller, G. B. 1972. The Serengeti Lion. University of Chicago Press,Chicago, 480 pp.
Schmidt-Kittler, N. 1976. Raubteiere aus dem Juntertiar Kleinasiens.Palaeontographica 155:1–131.
Seidensticker, J., and C. McDougal. 1993. Tiger predatory behaviour,ecology and conservation. Symposia of the Zoological Society ofLondon 65:105–125.
Sen, S. 1990. The Hipparion datum and its chronologic evidence in theMediterranean area; pp. 498–505 in E. H. Lindsay, V. Fahlbusch andP. Mein (eds.), European Neogene Mammal Chronology. PlenumPress, New York.
Short, R. V., and E. Balaban. 1994. The Differences Between the Sexes.Cambridge University Press, Cambridge, 495 pp.
Simpson, G. G. 1941. The function of saber-like canines in carnivorousmammals. American Museum Novitates 130:1–12.
Sotnikova, M. 1992. A new species of Machairodus from the late Mio-cene Kalmakpai locality in eastern Kazakhstan (USSR). AnnalesZoologici Fennici 28:361–369.
Turner, A. 1984. Dental sex dimorphism in European lions (Panthera leoL.) of the Upper Pleistocene: palaeoecological and palaeoethologi-cal implications. Annales Zoologici Fennici 21:1–8.
Turner, A. 1993. New fossil carnivore remains; pp. 151–165 in C.K. Brain
(ed.), Swartkrans: a Cave’s Chronicle of Early Man. Transvaal Mu-seum Monograph 8. Pretoria.
Turner, A., and M. Anton. 1997. The Big Cats and their Fossil Relatives.Columbia University Press, New York, 234 pp.
Van Valkenburgh, B. 1988. Incidence of tooth breakage among large,predatory mammals. American Naturalist 131:291–300.
Van Valkenburgh, B. and C. B. Ruff. 1987. Canine tooth strength andkilling behaviour in large carnivores. Journal of Zoology 212:379–397.
Van Valkenburgh, B., and T. Sacco. 2002. Sexual dimorphism, socialbehavior, and intrasexual competition in large Pleistocene carniv-orans. Journal of Vertebrate Paleontology 22:164–169.
Weckerly, F. 1998. Sexual-size dimorphism: influence of body mass andmating systems in the most dimorphic mammals. Journal of Mam-malogy 79:33–52.
Wozencraft, W. C. 1993. Order Carnivora; pp. 286–346 in Wilson, D. E.,and D. Reeder. (eds.), Mammal Species of the World. A Taxonomicand Geographic Reference. Smithsonian Institution Press, Washing-ton.
Zdansky, O. 1924. Jungtertiare Carnivoren Chinas. PalaeontologicaSinica 2:1–149
Received 25 April 2003; accepted 8 January 2004.
ANTON ET AL.—SKULLS OF MACHAIRODUS FROM BATALLONES-1 969
Related Documents











