First Hominoid From the Miocene of Ethiopia and the Evolution of the Catarrhine Elbow BRIAN G. RICHMOND, 1 * JOHN G. FLEAGLE, 2 JOHN KAPPELMAN, 3 AND CARL C. SWISHER III 4 1 Smithsonian Institution, Washington, DC 20560 2 Department of Anatomical Sciences, School of Medicine, State University of New York at Stony Brook, Stony Brook, New York 11794 3 Department of Anthropology, University of Texas at Austin, Austin, Texas 78712-1086 4 Berkeley Geochronology Center, Berkeley, California 94709 KEY WORDS fossil apes; anthropoids; postcranium; forelimb; Turkanapithecus; Pliopithecus; multivariate analysis ABSTRACT The first known fossil ape from the early-middle Miocene of Fejej, Ethiopia, is described here. The specimen, FJ-18SB-68, is a partial ulna from a locality dated by 40 Ar/ 39 Ar and paleomagnetic methods to a minimum age of 16.18 MYA. Compared to a variety of extant and fossil ulnae, FJ-18SB-68 is most similar to Turkanapithecus, Proconsul, and Pliopithecus, and appears to have been an arboreal quadruped with substantial forearm rotational mobility. Among the extant ulnae, canonical variates analysis successfully discrimi- nates platyrrhines from catarrhines and within the latter, cercopithecoids from hominoids. Basal catarrhines (e.g., Aegyptopithecus) are platyrrhine- like in their morphology. Two basic trends appear to evolve from this generalized template: one with less mobile and more habitually pronated forearms, as seen in living and fossil cercopithecoids (including Victoriapithe- cus and Paracolobus), and another with greater forearm rotational mobility in fossil and modern hominoids. Primitive Miocene apes, including Proconsul, Turkanapithecus, and FJ-18SB-68, share with extant hominoids a more laterally positioned and laterally facing radial notch and an incipient trochlear keel. This morphology, along with a large insertion area for m. brachialis, suggests a departure from the more habitually pronated hand posture of monkeys and may indicate greater climbing abilities in these arboreally quadrupedal apes. Later Miocene apes, such as Oreopithecus and Dryopithecus share additional morphological features with hominoids, indicat- ing considerable suspensory and climbing capabilities. Am J Phys Anthropol 105:257–277, 1998. r 1998 Wiley-Liss, Inc. The elbow joint of living apes and humans is one of our most distinctive anatomical regions. The unique features of the hominoid elbow are thought to be related to extensive abilities of pronation and supination pertaining to suspensory locomotor adaptations (e.g., Jenkins, 1973; Rose, 1988). Analysis of this region has long played an important role in under- standing the locomotor adaptations and phy- logenetic relationships of fossil apes (e.g., Napier and Davis, 1959; Fleagle et al., 1975; Fleagle, 1983; Rose, 1988, 1993; Larson, 1993). In this article, we describe a partial ulna of a fossil ape from the early-middle Miocene Contract grant sponsor: National Geographic Society; contract grant numbers 4393-90, 4600-91; Contract grant sponsor: NSF; contract grant number SBR-9319193; Contract grant sponsors: L.S.B. Leakey Foundation, The Wenner-Gren Foundation. *Correspondence to: Brian G. Richmond, Human Origins Program, Department of Anthropology, MRC 112, Smithsonian Institution, Washington, DC 20560. E-mail: [email protected] Received 29 May 1997; accepted 31 October 1997. AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 105:257–277 (1998) r 1998 WILEY-LISS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

First Hominoid From the Miocene of Ethiopia and the Evolutionof the Catarrhine Elbow
BRIAN G. RICHMOND,1* JOHN G. FLEAGLE,2 JOHN KAPPELMAN,3AND CARL C. SWISHER III4
1Smithsonian Institution, Washington, DC 205602Department of Anatomical Sciences, School of Medicine, State Universityof New York at Stony Brook, Stony Brook, New York 117943Department of Anthropology, University of Texas at Austin, Austin, Texas78712-10864Berkeley Geochronology Center, Berkeley, California 94709
KEY WORDS fossil apes; anthropoids; postcranium; forelimb;Turkanapithecus; Pliopithecus; multivariate analysis
ABSTRACT The first known fossil ape from the early-middle Miocene ofFejej, Ethiopia, is described here. The specimen, FJ-18SB-68, is a partial ulnafrom a locality dated by 40Ar/39Ar and paleomagnetic methods to a minimumage of 16.18 MYA. Compared to a variety of extant and fossil ulnae,FJ-18SB-68 is most similar to Turkanapithecus, Proconsul, and Pliopithecus,and appears to have been an arboreal quadruped with substantial forearmrotational mobility.
Among the extant ulnae, canonical variates analysis successfully discrimi-nates platyrrhines from catarrhines and within the latter, cercopithecoidsfrom hominoids. Basal catarrhines (e.g., Aegyptopithecus) are platyrrhine-like in their morphology. Two basic trends appear to evolve from thisgeneralized template: one with less mobile and more habitually pronatedforearms, as seen in living and fossil cercopithecoids (including Victoriapithe-cus and Paracolobus), and another with greater forearm rotational mobility infossil and modern hominoids. Primitive Miocene apes, including Proconsul,Turkanapithecus, and FJ-18SB-68, share with extant hominoids a morelaterally positioned and laterally facing radial notch and an incipienttrochlear keel. This morphology, along with a large insertion area for m.brachialis, suggests a departure from the more habitually pronated handposture of monkeys and may indicate greater climbing abilities in thesearboreally quadrupedal apes. Later Miocene apes, such as Oreopithecus andDryopithecus share additional morphological features with hominoids, indicat-ing considerable suspensory and climbing capabilities. Am J Phys Anthropol105:257–277, 1998. r 1998 Wiley-Liss, Inc.
The elbow joint of living apes andhumans is one of our most distinctiveanatomical regions. The unique featuresof the hominoid elbow are thought to berelated to extensive abilities of pronationand supination pertaining to suspensorylocomotor adaptations (e.g., Jenkins, 1973;Rose, 1988). Analysis of this region haslong played an important role in under-standing the locomotor adaptations and phy-logenetic relationships of fossil apes (e.g.,Napier and Davis, 1959; Fleagle et al., 1975;
Fleagle, 1983; Rose, 1988, 1993; Larson,1993).
In this article, we describe a partial ulnaof a fossil ape from the early-middle Miocene
Contract grant sponsor: National Geographic Society; contractgrant numbers 4393-90, 4600-91; Contract grant sponsor: NSF;contract grant number SBR-9319193; Contract grant sponsors:L.S.B. Leakey Foundation, The Wenner-Gren Foundation.
*Correspondence to: Brian G. Richmond, Human OriginsProgram, Department of Anthropology, MRC 112, SmithsonianInstitution, Washington, DC 20560.E-mail: [email protected]
Received 29 May 1997; accepted 31 October 1997.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 105:257–277 (1998)
r 1998 WILEY-LISS, INC.
of Fejej, Ethiopia (Asfaw et al., 1991; Bownet al., 1994). The limited Miocene deposits inthis area have yielded a rich flora of fossilfruits and seeds (Tiffney et al., 1994), as wellas numerous remains of large terrestrialmammals (Fleagle et al., 1994). However,the ulna is the first Miocene ape known fromEthiopia. In describing the fossil ape ulna,we compare it with other fossil apes fromboth Africa and Europe, as well as extantanthropoids. Multivariate methods are em-ployed to identify the features of the proxi-mal ulna which discriminate taxonomicgroups. These comparisons provide a broadevolutionary context for examining the adap-tive and phylogenetic evolution of the elbowjoint in catarrhine evolution.
LOCALITY AND GEOCHRONOLOGY
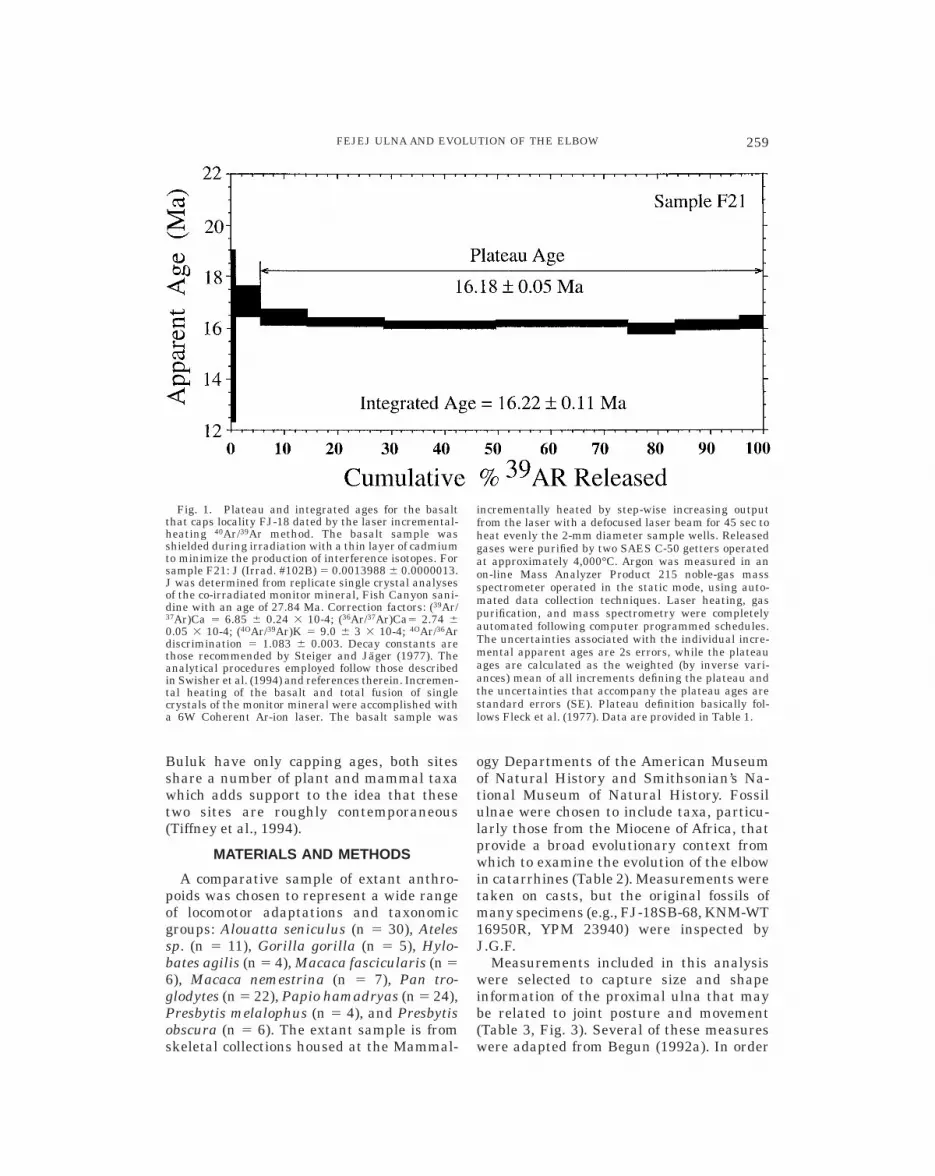
The Miocene locality of FJ-18 lies on theFejej Plain of western and southernmostEthiopia (Tiffney et al., 1994). The fossils atlocality FJ-18 include diverse plant andanimal remains from two distinct levels(Tiffney et al., 1994). Mammal fossils occurin the upper level and are found within anintraclastic conglomerate that consists ofsubangular to well-rounded granules in agray volcanic mudstone. The conglomerateranges from clast to matrix-supported andfills a minor paleovalley. The conglomerateis capped by a fine-grained porphorytic pla-gioclase and pyroxene-rich theoleiitic lava.This lava was sampled for radioisotopic dat-ing, with samples trimmed on the outcrop.The basalt has a well-defined plateau andprovides a 40Ar/39Ar age of 16.18 6 0.05 MYA(Table 1 and Fig. 1). Other volcanic lavasunderlying the fossil horizon were found tobe too altered to permit dating.
The capping basalt was also sampled forpaleomagnetic analysis. Step-wise thermaldemagnetization (Fig. 2) shows a steep de-crease in intensity to 500°C and randomdirections above 575°C. Vectors for eachsample (n 5 3) were calculated with princi-pal component analysis (Kirschvink, 1980)and reveal that the basalt is reversely mag-netized (virtual geomagnetic pole latitude 5245°). The steep positive inclination is un-usual for a site that lies this close to theequator and suggests that the remanentmagnetism may have been locked in near apolarity transition. The time scale of Candeand Kent (1995) places the date of 16.18 60.05 MYA solidly within the normal intervalof chron C5Cn.1n, while other time scales(e.g., Berggren et al., 1985) place this datenear the older end of the long reverselymagnetized interval of C5Br. The reversedpolarity and positive inclination of this ba-salt tentatively suggests that it could lienear the upper (or even lower) boundary ofC5Cn.1n but the results from this single siteare insufficient to attempt to better resolvethe timing of this polarity transition.
These data provide a minimum age of16.18 MYA for locality FJ-18 and add theFejej region to the growing number ofMiocene fossil localities in Ethiopia (Suwaet al., 1991, and references therein; Wolde-Gabriel et al., 1991). This age falls withinthe range of K-Ar ages reported for volcanicsfrom Buluk (15.7 6 0.2 – 17.2 1 0.2 MYA)located to the southeast just across theborder in Kenya that also cap fossiliferousprimate-bearing sediments (McDougall andWatkins, 1985; Leakey and Walker, 1985;Watkins, 1989). Although both FJ-18 and
TABLE 1. Laser incremental-heating 40Ar/39Ar data
Step %39Ar 40Ar/39Ar 37Ar/39Ar 36Ar/39Ar 40Ar*/39Ar %40Ar* Age SD
Sample F21/7368-01
A 0.9 44.237 4.162 0.1298 6.240 14.1 15.68 1.68B 4.7 15.472 3.322 0.0304 6.782 43.7 17.03 0.30C 8.7 10.480 2.757 0.0141 6.539 62.3 16.43 0.16D 14.3 8.297 2.377 0.0069 6.466 77.8 16.24 0.08E 20.9 7.457 2.102 0.0041 6.427 86.1 16.14 0.07F 24.9 6.823 1.985 0.0019 6.443 94.3 16.19 0.07G 8.8 6.698 3.760 0.0022 6.364 94.8 15.99 0.12H 12.3 6.728 5.992 0.0028 6.426 95.1 16.14 0.11I 4.5 6.703 6.016 0.0025 6.467 96.1 16.25 0.14
258 B.G. RICHMOND ET AL.
Buluk have only capping ages, both sitesshare a number of plant and mammal taxawhich adds support to the idea that thesetwo sites are roughly contemporaneous(Tiffney et al., 1994).
MATERIALS AND METHODS
A comparative sample of extant anthro-poids was chosen to represent a wide rangeof locomotor adaptations and taxonomicgroups: Alouatta seniculus (n 5 30), Atelessp. (n 5 11), Gorilla gorilla (n 5 5), Hylo-bates agilis (n 5 4), Macaca fascicularis (n 56), Macaca nemestrina (n 5 7), Pan tro-glodytes (n 5 22), Papio hamadryas (n 5 24),Presbytis melalophus (n 5 4), and Presbytisobscura (n 5 6). The extant sample is fromskeletal collections housed at the Mammal-
ogy Departments of the American Museumof Natural History and Smithsonian’s Na-tional Museum of Natural History. Fossilulnae were chosen to include taxa, particu-larly those from the Miocene of Africa, thatprovide a broad evolutionary context fromwhich to examine the evolution of the elbowin catarrhines (Table 2). Measurements weretaken on casts, but the original fossils ofmany specimens (e.g., FJ-18SB-68, KNM-WT16950R, YPM 23940) were inspected byJ.G.F.
Measurements included in this analysiswere selected to capture size and shapeinformation of the proximal ulna that maybe related to joint posture and movement(Table 3, Fig. 3). Several of these measureswere adapted from Begun (1992a). In order
Fig. 1. Plateau and integrated ages for the basaltthat caps locality FJ-18 dated by the laser incremental-heating 40Ar/39Ar method. The basalt sample wasshielded during irradiation with a thin layer of cadmiumto minimize the production of interference isotopes. Forsample F21: J (Irrad. #102B) 5 0.0013988 6 0.0000013.J was determined from replicate single crystal analysesof the co-irradiated monitor mineral, Fish Canyon sani-dine with an age of 27.84 Ma. Correction factors: (39Ar/37Ar)Ca 5 6.85 6 0.24 3 10-4; (36Ar/37Ar)Ca5 2.74 60.05 3 10-4; (4OAr/39Ar)K 5 9.0 6 3 3 10-4; 4OAr/36Ardiscrimination 5 1.083 6 0.003. Decay constants arethose recommended by Steiger and Jager (1977). Theanalytical procedures employed follow those describedin Swisher et al. (1994) and references therein. Incremen-tal heating of the basalt and total fusion of singlecrystals of the monitor mineral were accomplished witha 6W Coherent Ar-ion laser. The basalt sample was
incrementally heated by step-wise increasing outputfrom the laser with a defocused laser beam for 45 sec toheat evenly the 2-mm diameter sample wells. Releasedgases were purified by two SAES C-50 getters operatedat approximately 4,000°C. Argon was measured in anon-line Mass Analyzer Product 215 noble-gas massspectrometer operated in the static mode, using auto-mated data collection techniques. Laser heating, gaspurification, and mass spectrometry were completelyautomated following computer programmed schedules.The uncertainties associated with the individual incre-mental apparent ages are 2s errors, while the plateauages are calculated as the weighted (by inverse vari-ances) mean of all increments defining the plateau andthe uncertainties that accompany the plateau ages arestandard errors (SE). Plateau definition basically fol-lows Fleck et al. (1977). Data are provided in Table 1.
259FEJEJ ULNA AND EVOLUTION OF THE ELBOW

to measure the angle of the radial notch, theulna was placed horizontally in a paddedvicegrip with the sigmoid notch and coro-noid process facing superiorly. For mostextant samples, the humerus was positionedvertically on the superiorly directed sigmoidnotch in order to help orient the ulna. Withthe ulna firmly in position, one arm of agoniometer was placed flush against thecenter of the radial notch in a transverseplane. The other arm fell vertically underthe influence of gravity, and the angle of theradial notch was recorded. Angles are diffi-cult to measure, in this case primarily due tothe reliance on accurate, consistent orienta-tion of the ulna. To test the reliability of thismeasurement, an Alouatta ulna (conserva-tively chosen for its small radial notch) waspositioned and measured ten times over atwo-month period and all measures fellwithin four degrees of each other (mean 545.5, s.d. 5 1.18, range: 43–47).
Statistical analyses were performed onboth raw and size-adjusted shape data. Thesize of the ulna is taken as the geometricmean (GM) of the subset of measurementsthat capture the width, depth, and length ofthe proximal ulna, calculated as the nth rootof the product of n measurements (Mosi-mann, 1970; Darroch and Mosimann, 1985;
Fig. 2. Results from demagnetization treatments ofthe dated basalt that caps FJ-18. Sample F21c ischaracteristic of the other samples from the basalt anddemonstrates a stable reversed declination and positiveinclination from 200°–500°C and random directionsabove 575°C. The basalt is reversely magnetized. Sym-bols: solid squares equal declination, hollow squaresequal inclination, crosses are lower hemisphere, hollowcircles are upper hemisphere, hollow cross is the meanof the three samples (declination equals 183.0°, inclina-tion equals 59.6°, R equals 2.97) calculated with Fisherstatistics (Fisher, 1953). Samples were measured with a2G SCT superconducting magnetometer in a magneti-cally shielded room at the University of Texas at Austin.Step-wise thermal demagnetization was conducted witha Schonstedt Thermal Demagnetizer (TSD-1).
TABLE 2. Fossil primate ulnae included in this analysis
Specimen Locality Age (MYA) Reference Taxon
FJ-18SB-68 Fejej, Ethiopia .16.18 this manuscript indet.KNM-WK16950R Kalodirr, Kenya 16–18 Leakey et al., 1988 Turkanapithecus kala-
kolensisKNM-RU2036 Rusinga Island, Keyna e. Miocene Walker & Pickford,
1983Proconsul heseloni
KNM-RU1786 Rusinga Island, Kenya e. Miocene Szalay & Delson, 1979 Proconsul nyanzaeKNM-RU2099E Rusinga Island, Kenya e. Miocene LeGros Clark &
Thomas, 1951Dendropithecus macin-
nesiKNM-FT3381 Fort Ternan, Kenya 13–14 Andrews & Walker,
1976hominoid?
KNM-BC3D Lake Baringo, Kenya m. Pliocene Birchette, 1982 Paracolobus chemeroniKNM-MB32 Maboko Island, Kenya 15–16 von Koenigswald, 1969 Victoriapithecus macin-
nesiYPM23940 Fayum, Egypt .33 Fleagle et al., 1975 Aegyptopithecus zeuxisRUD22 Rudabanya, Hungary 11 Kretzoi, 1975; Morbeck,
1983Dryopithecus brancoi
Klein Hadersdorf Klein Hadersdorf,Austria
13–14 Ehrensberg, 1938;Begun, 1992a
Austriacopithecus wein-furti
Ind I Neudorf an der Marche, m-l. Miocene Zapfe, 1958 Pliopithecus vindo-bonensis
Ind II Czech Republic m-l. Miocene Zapfe, 1958 Pliopithecus vindo-bonensis
IGF2011V Baccinello, Italy l. Miocene Hurzeler, 1958 Oreopithecus bambolii
260 B.G. RICHMOND ET AL.

Richmond and Jungers, 1995). Ulnar shapeis computed here as the ratio of each vari-able to the GM (Darroch and Mosimann,1985; Jungers et al., 1995).
Multivariate summary statistics are usedhere to assess the morphological affinities ofthe extant and fossil ulnae. Because of thevariability in preservation of the fossil sam-ple, canonical variates analyses, or CVAs,are calculated for two subsets of the raw andsize-adjusted data. In the first, the entiresuite of measurements (except the angularmeasure, RNA) and only the most complete
fossils are included. The second CVA isbased on a subset of the measurements toallow the inclusion of the full fossil sample(Table 3). The fossil specimens were in-cluded in the CVA a priori to allow them tocontribute to the canonical shape space (Ox-nard, 1972; Corruccini, 1978). SAS and SYS-TAT statistical packages were employed tocalculate and plot the CVAs. The variableswith the greatest discriminatory poweramong the extant and fossil taxa (i.e., thosewith the highest between-group canonicalstructure coefficients; Klecka, 1980) providea focus for the subsequent investigation intothe structural and functional differences
Fig. 3. Line drawing of a gorilla proximal ulna inanterior, lateral, and proximal views, illustrating mostof the measurements used in this analysis (MAH, notshown, is measured on the medial side opposite LAH).RNA is shown on a line drawing of the proximal view of aPapio ulna. Measurement abbreviations are describedin Table 3.
TABLE 3. Measurements included in this analysis(see also Figure 3)
Abbreviation Description
PAB*† Proximal articular mediolateralbreadth of radial notch and trochlea
TAB* Trochlear articular mediolateralbreadth
PSML*† Proximal shaft mediolateral width atdistal margin of radial notch
PAP*† Proximal shaft anteroposterior thick-ness at distal margin of radial notch
RAP* Radial notch anteroposterior thicknessRPD* Radial notch proximodistal lengthPAAP*† Proximal articular anteroposterior
thickness at coronoid processSND*† Sigmoid notch depth; minimum antero-
posterior thickness of sigmoid notchSML*† Sigmoid notch mediolateral widthPAH Proximal articular anteroposterior
thickness at anconeal processOPAP Olecranon process anteroposterior
thicknessOPL Olecranon process length from anconeal
processNPD Sigmoid notch proximodistal length
from proximal to distal beaksOPML Olecranon process mediolateral breadthSHMN Minimum mediolateral width of shaft
anterior to brachialis insertionSHMX Maximum mediolateral width of shaft
including brachialis insertionMAH* Medial anteroposterior articular thick-
ness from posterior border of medialarticular surface to posterior borderof shaft
LAH* Lateral anteroposterior articular thick-ness from posterior border of lateralarticular surface to posterior borderof shaft
RNA Radial notch angle (in degrees) relativeto anteroposterior plane
* Subset of measurements used in analyses involving the entirefossil sample.† Subset of measurements used in calculation of geometric meanfor full fossil sample.Many of these measures are adapted from Begun (1992a).Measurements are shown in mm unless otherwise indicated.
261FEJEJ ULNA AND EVOLUTION OF THE ELBOW
between these groups (McHenry and Corruc-cini, 1975; Oxnard, 1975).
Description
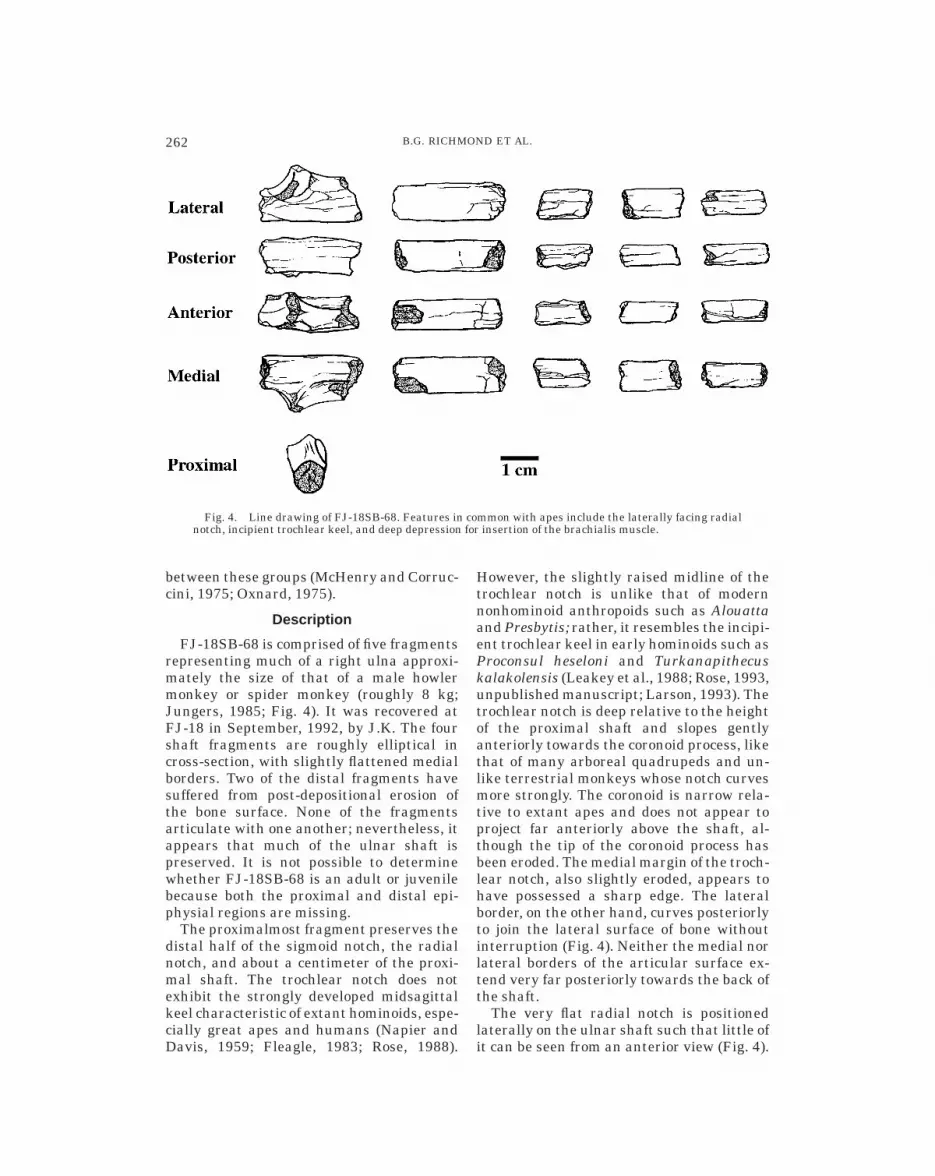
FJ-18SB-68 is comprised of five fragmentsrepresenting much of a right ulna approxi-mately the size of that of a male howlermonkey or spider monkey (roughly 8 kg;Jungers, 1985; Fig. 4). It was recovered atFJ-18 in September, 1992, by J.K. The fourshaft fragments are roughly elliptical incross-section, with slightly flattened medialborders. Two of the distal fragments havesuffered from post-depositional erosion ofthe bone surface. None of the fragmentsarticulate with one another; nevertheless, itappears that much of the ulnar shaft ispreserved. It is not possible to determinewhether FJ-18SB-68 is an adult or juvenilebecause both the proximal and distal epi-physial regions are missing.
The proximalmost fragment preserves thedistal half of the sigmoid notch, the radialnotch, and about a centimeter of the proxi-mal shaft. The trochlear notch does notexhibit the strongly developed midsagittalkeel characteristic of extant hominoids, espe-cially great apes and humans (Napier andDavis, 1959; Fleagle, 1983; Rose, 1988).
However, the slightly raised midline of thetrochlear notch is unlike that of modernnonhominoid anthropoids such as Alouattaand Presbytis; rather, it resembles the incipi-ent trochlear keel in early hominoids such asProconsul heseloni and Turkanapithecuskalakolensis (Leakey et al., 1988; Rose, 1993,unpublished manuscript; Larson, 1993). Thetrochlear notch is deep relative to the heightof the proximal shaft and slopes gentlyanteriorly towards the coronoid process, likethat of many arboreal quadrupeds and un-like terrestrial monkeys whose notch curvesmore strongly. The coronoid is narrow rela-tive to extant apes and does not appear toproject far anteriorly above the shaft, al-though the tip of the coronoid process hasbeen eroded. The medial margin of the troch-lear notch, also slightly eroded, appears tohave possessed a sharp edge. The lateralborder, on the other hand, curves posteriorlyto join the lateral surface of bone withoutinterruption (Fig. 4). Neither the medial norlateral borders of the articular surface ex-tend very far posteriorly towards the back ofthe shaft.
The very flat radial notch is positionedlaterally on the ulnar shaft such that little ofit can be seen from an anterior view (Fig. 4).
Fig. 4. Line drawing of FJ-18SB-68. Features in common with apes include the laterally facing radialnotch, incipient trochlear keel, and deep depression for insertion of the brachialis muscle.
262 B.G. RICHMOND ET AL.
Article ID # 911 (disk)@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_911z13 dawn

In this regard, FJ-18SB-68 deviates fromthe morphology seen in most quadrupedalanthropoids, in which the radial notch facesantero-laterally and covers about half of thewidth of the ulnar shaft (Fig. 5c). The latterradio-ulnar configuration is seen in Aegypto-pithecus and appears to be primitive forcatarrhines. The edges of the articular sur-face do not project laterally from the shaft,thereby providing little or no support poste-riorly for the head of the radius.
Immediately distal to the coronoid andradial notch, the shaft possesses a deepgroove on the medial side for the insertion ofm. brachialis (Fig. 4). The supinator crest isdistinct, as in Turkanapithecus (Rose, 1997),but not as well developed as in living greatapes, Dryopithecus, and Austriacopithecus(Klein Hadersdorf; Table 2). In general, FJ-18SB-68 most closely resembles T. kalakolen-sis and P. heseloni, but differs in its slightlymore laterally facing and less rugose radialnotch and its more extensively excavatedbrachialis depression.
RESULTS
We initially investigate complete proxi-mal ulnae of living and some fossil anthro-poids.An analysis of complete ulnar morphol-ogy reveals how well the measurementsused here discriminate among extant taxain terms of function and phylogeny. It pro-vides the context for the following multivari-ate analysis based on more fragmentary re-mains, which permits consideration of a morediverse fossil sample, including the Fejej speci-men, but suffers in discriminatory power.
Complete measurement set
The CVA of shape variables (ratios ofvariables to GM) separates extant anthro-poids into three major clusters: hominoids,cercopithecoids, and platyrrhines (Fig. 6).CAN1 discriminates well between apes andall other nonhominoid anthropoids, whileCAN2 separates platyrrhines (Ateles andAouatta) from catarrhines. This is not areflection of size differences. The position ofthe monkey-sized gibbons in the cluster ofthe much larger great apes underscores thisfact. Correlations between the group cen-troids for each shape variable and the canoni-cal axes are shown in Table 4.
The third canonical axis (not shown), rep-resenting 11.6% of the canonical variation(CAN4, 3.3%), distinguishes gibbons from allother anthropoids, based on their relativelylarge radial notches (RPD, RAP), shallowdepth posterior to the sigmoid notch (SND),and shallow (AP) olecranon process (OPAP).
The shape of the proximal ulna is distinctin each of the three major groups, suggest-ing fundamental differences in elbow func-tion. The platyrrhine ulnae examined hereare characterized by being relatively narrow(mediolaterally, or ML) proximally in boththe shaft and articular surface, with antero-posteriorly (AP) short radial notches andlong (proximodistally, or PD), narrow olecra-non processes (Fig. 6 and Table 4). Thecercopithecoid ulna is like that of platyr-rhines in many of these features (i.e., cerco-pithecoids fall on the platyrrhine side ofCAN1) but are generally more extreme ineach variable. The olecranon process is longand deep, the sigmoid notch is exceptionallynarrow and deep, the coronoid process doesnot project markedly, and the radial notch isAP elongated (see also Bown et al., 1982;Harrison, 1987; Rose, 1988; Ciochon, 1993).These features are uniquely combined incercopithecoids with the broad, deep shaftsand articular surfaces and the relativelyshort sigmoid notch length (NPD) that char-acterize modern catarrhines. Hominoids alsopossess these catarrhine features but aredistinguished from all other anthropoids inhaving a relatively large trochlear surface,broad sigmoid notch that is well buttressedposteriorly, broad, but otherwise small olec-ranon process, and projecting coronoid pro-cess (see also Harrison, 1987; Fleagle andKay, 1987; Rose, 1988).
The fossil ulnae are encompassed by thetotal ulnar shape variation displayed by theextant taxa (Fig. 6). Aegyptopithecus andPliopithecus most closely resemble the liv-ing New World monkeys in this sample andappear to lack the specializations of eitherthe extant cercopithecoids or hominoids(Fleagle et al., 1975; Fleagle, 1983, 1988).The Miocene hominoids Turkanapithecusand Proconsul nyanzae clearly cluster withthe quadrupeds along CAN1 and the catar-rhines on CAN2, but do not overlap with the
263FEJEJ ULNA AND EVOLUTION OF THE ELBOW
Article ID # 911 (disk)@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_911z13 dawn
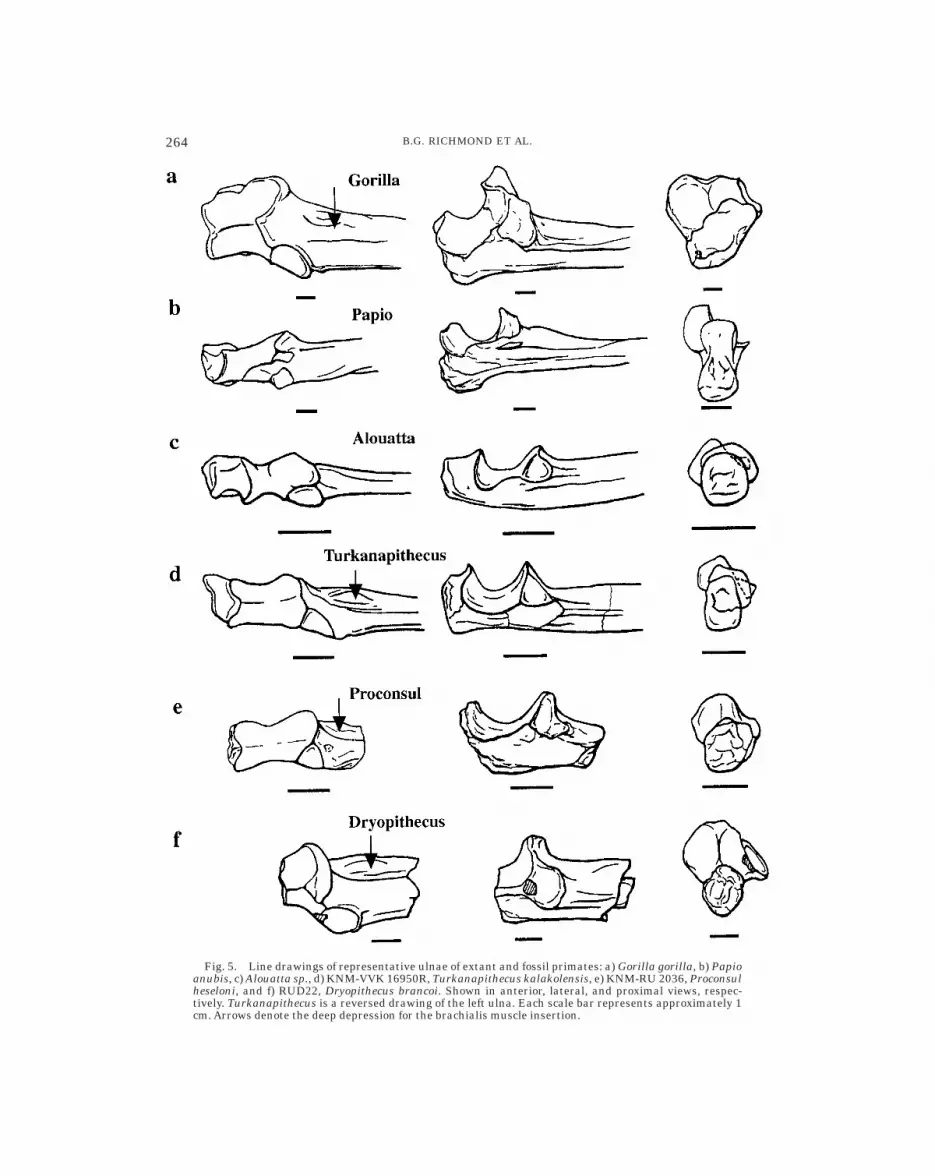
Fig. 5. Line drawings of representative ulnae of extant and fossil primates: a) Gorilla gorilla, b) Papioanubis, c) Alouatta sp., d) KNM-VVK 16950R, Turkanapithecus kalakolensis, e) KNM-RU 2036, Proconsulheseloni, and f) RUD22, Dryopithecus brancoi. Shown in anterior, lateral, and proximal views, respec-tively. Turkanapithecus is a reversed drawing of the left ulna. Each scale bar represents approximately 1cm. Arrows denote the deep depression for the brachialis muscle insertion.
264 B.G. RICHMOND ET AL.
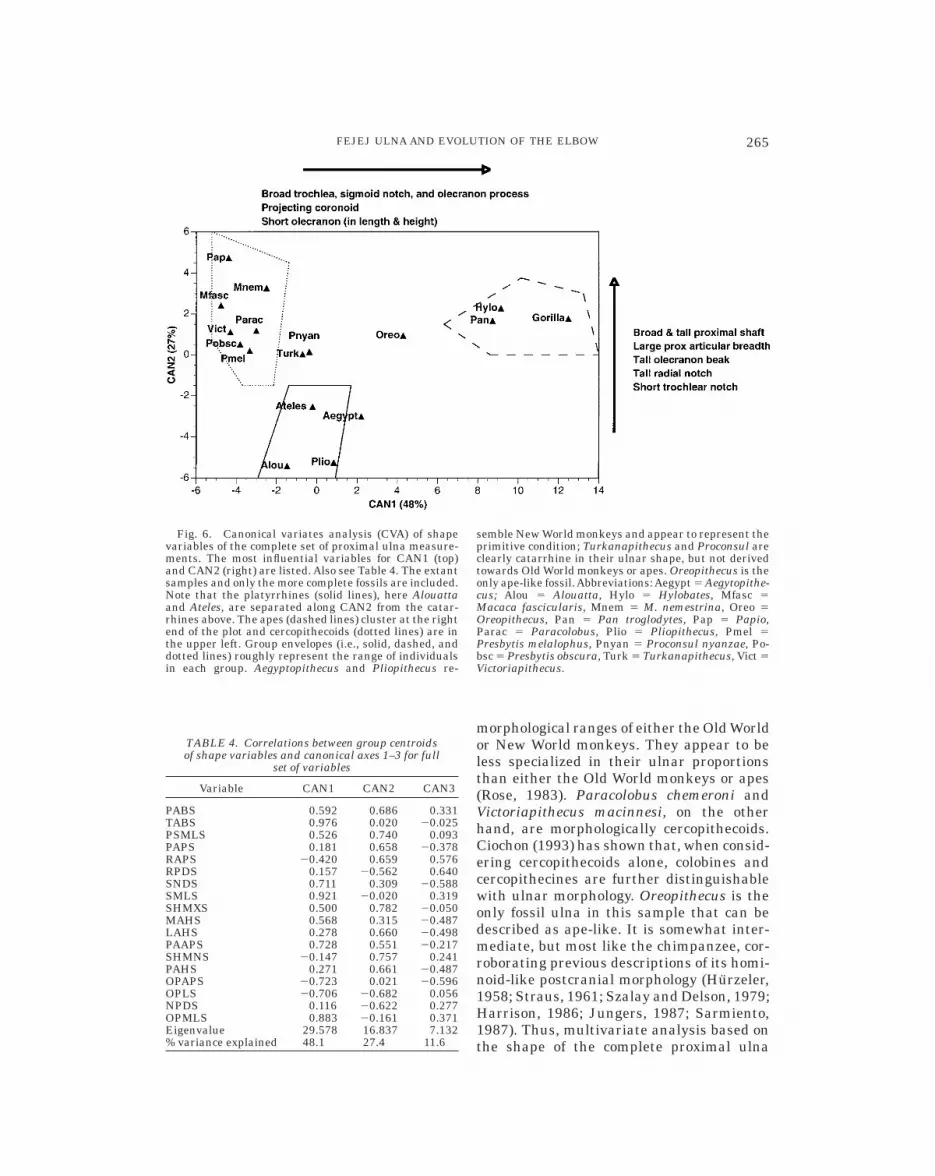
morphological ranges of either the Old Worldor New World monkeys. They appear to beless specialized in their ulnar proportionsthan either the Old World monkeys or apes(Rose, 1983). Paracolobus chemeroni andVictoriapithecus macinnesi, on the otherhand, are morphologically cercopithecoids.Ciochon (1993) has shown that, when consid-ering cercopithecoids alone, colobines andcercopithecines are further distinguishablewith ulnar morphology. Oreopithecus is theonly fossil ulna in this sample that can bedescribed as ape-like. It is somewhat inter-mediate, but most like the chimpanzee, cor-roborating previous descriptions of its homi-noid-like postcranial morphology (Hurzeler,1958; Straus, 1961; Szalay and Delson, 1979;Harrison, 1986; Jungers, 1987; Sarmiento,1987). Thus, multivariate analysis based onthe shape of the complete proximal ulna
Fig. 6. Canonical variates analysis (CVA) of shapevariables of the complete set of proximal ulna measure-ments. The most influential variables for CAN1 (top)and CAN2 (right) are listed. Also see Table 4. The extantsamples and only the more complete fossils are included.Note that the platyrrhines (solid lines), here Alouattaand Ateles, are separated along CAN2 from the catar-rhines above. The apes (dashed lines) cluster at the rightend of the plot and cercopithecoids (dotted lines) are inthe upper left. Group envelopes (i.e., solid, dashed, anddotted lines) roughly represent the range of individualsin each group. Aegyptopithecus and Pliopithecus re-
semble New World monkeys and appear to represent theprimitive condition; Turkanapithecus and Proconsul areclearly catarrhine in their ulnar shape, but not derivedtowards Old World monkeys or apes. Oreopithecus is theonly ape-like fossil.Abbreviations:Aegypt 5 Aegytopithe-cus; Alou 5 Alouatta, Hylo 5 Hylobates, Mfasc 5Macaca fascicularis, Mnem 5 M. nemestrina, Oreo 5Oreopithecus, Pan 5 Pan troglodytes, Pap 5 Papio,Parac 5 Paracolobus, Plio 5 Pliopithecus, Pmel 5Presbytis melalophus, Pnyan 5 Proconsul nyanzae, Po-bsc 5 Presbytis obscura, Turk 5 Turkanapithecus, Vict 5Victoriapithecus.
TABLE 4. Correlations between group centroidsof shape variables and canonical axes 1–3 for full
set of variables
Variable CAN1 CAN2 CAN3
PABS 0.592 0.686 0.331TABS 0.976 0.020 20.025PSMLS 0.526 0.740 0.093PAPS 0.181 0.658 20.378RAPS 20.420 0.659 0.576RPDS 0.157 20.562 0.640SNDS 0.711 0.309 20.588SMLS 0.921 20.020 0.319SHMXS 0.500 0.782 20.050MAHS 0.568 0.315 20.487LAHS 0.278 0.660 20.498PAAPS 0.728 0.551 20.217SHMNS 20.147 0.757 0.241PAHS 0.271 0.661 20.487OPAPS 20.723 0.021 20.596OPLS 20.706 20.682 0.056NPDS 0.116 20.622 0.277OPMLS 0.883 20.161 0.371Eigenvalue 29.578 16.837 7.132% variance explained 48.1 27.4 11.6
265FEJEJ ULNA AND EVOLUTION OF THE ELBOW
discriminates phylogenetically among an-thropoids.
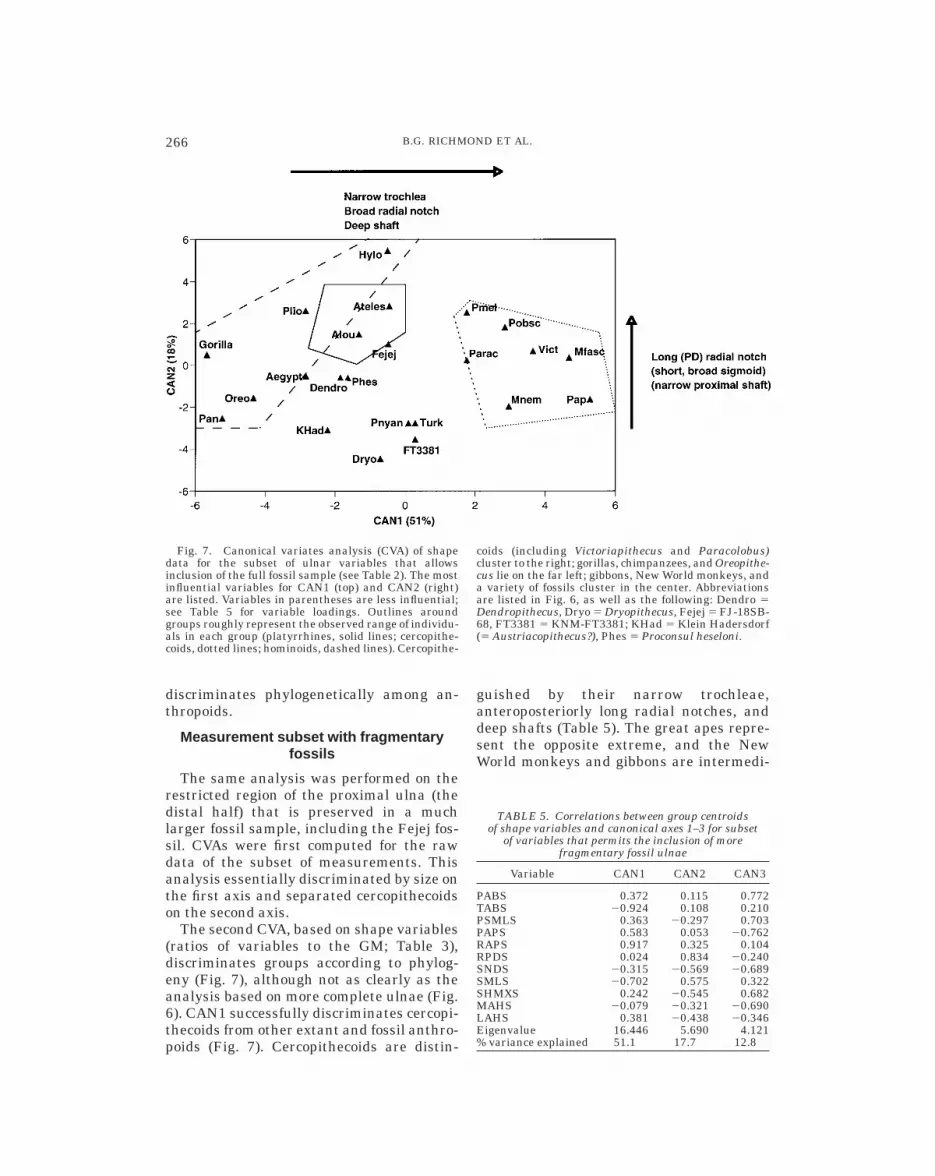
Measurement subset with fragmentaryfossils
The same analysis was performed on therestricted region of the proximal ulna (thedistal half) that is preserved in a muchlarger fossil sample, including the Fejej fos-sil. CVAs were first computed for the rawdata of the subset of measurements. Thisanalysis essentially discriminated by size onthe first axis and separated cercopithecoidson the second axis.
The second CVA, based on shape variables(ratios of variables to the GM; Table 3),discriminates groups according to phylog-eny (Fig. 7), although not as clearly as theanalysis based on more complete ulnae (Fig.6). CAN1 successfully discriminates cercopi-thecoids from other extant and fossil anthro-poids (Fig. 7). Cercopithecoids are distin-
guished by their narrow trochleae,anteroposteriorly long radial notches, anddeep shafts (Table 5). The great apes repre-sent the opposite extreme, and the NewWorld monkeys and gibbons are intermedi-
Fig. 7. Canonical variates analysis (CVA) of shapedata for the subset of ulnar variables that allowsinclusion of the full fossil sample (see Table 2). The mostinfluential variables for CAN1 (top) and CAN2 (right)are listed. Variables in parentheses are less influential;see Table 5 for variable loadings. Outlines aroundgroups roughly represent the observed range of individu-als in each group (platyrrhines, solid lines; cercopithe-coids, dotted lines; hominoids, dashed lines). Cercopithe-
coids (including Victoriapithecus and Paracolobus)cluster to the right; gorillas, chimpanzees, and Oreopithe-cus lie on the far left; gibbons, New World monkeys, anda variety of fossils cluster in the center. Abbreviationsare listed in Fig. 6, as well as the following: Dendro 5Dendropithecus, Dryo 5 Dryopithecus, Fejej 5 FJ-18SB-68, FT3381 5 KNM-FT3381; KHad 5 Klein Hadersdorf(5 Austriacopithecus?), Phes 5 Proconsul heseloni.
TABLE 5. Correlations between group centroidsof shape variables and canonical axes 1–3 for subset
of variables that permits the inclusion of morefragmentary fossil ulnae
Variable CAN1 CAN2 CAN3
PABS 0.372 0.115 0.772TABS 20.924 0.108 0.210PSMLS 0.363 20.297 0.703PAPS 0.583 0.053 20.762RAPS 0.917 0.325 0.104RPDS 0.024 0.834 20.240SNDS 20.315 20.569 20.689SMLS 20.702 0.575 0.322SHMXS 0.242 20.545 0.682MAHS 20.079 20.321 20.690LAHS 0.381 20.438 20.346Eigenvalue 16.446 5.690 4.121% variance explained 51.1 17.7 12.8
266 B.G. RICHMOND ET AL.

ate (Fig. 7). Although fossils appear to formtheir own group in the lower center of theplot, the CVA is overwhelmingly influencedby the variation in the extant data. A CVAperformed exclusively on the extant sample(not shown) is virtually identical to the oneshown in Figure 7 that includes the fossils,indicating that the variation in shape in theextant taxa largely encompasses that in thefossil ulnae (cf. Oxnard, 1972).
It appears that the portion of the ulnapreserved by the Fejej specimen is notenough to discriminate clearly among thefossil taxa. However, the CVA allows us toconclude that FJ-18SB-68 lacks the derivedproportions of living cercopithecoids and Af-rican apes. It clusters near primitive fossilapes (e.g., P. heseloni and Dendropithecus)and platyrrhines (here, Alouatta and At-eles), suggesting arboreal quadrupedalismwith some forelimb mobility. The functionaland phyletic affinities will be discussed inmore detail below with respect to specificmorphological traits.
DISCUSSION
The results presented here reinforce thoseof previous studies that the elbow joint isone of the most distinctive regions of thepostcranium in anthropoids (McHenry andCorruccini, 1975; Fleagle, 1983; Rose, 1988).The shape of the proximal ulna alone distin-guishes platyrrhines from catarrhines and,within the latter group, cercopithecoids fromhominoids. The distinctive ulnar morphol-ogy of each group is presumably related tofundamental differences in elbow functionin each of these groups. The functional sig-nificance of certain ulnar characteristics arediscussed below. The phyletic affinities ofFJ-18SB-68 and other fossil ulnae are thenexamined in light of derived features thatcharacterize extant groups.
Catarrhines
Few characteristics of the elbow havebeen identified as derived for catarrhines.They include the loss of the entepicondylarforamen (Fleagle, 1983; Harrison, 1987) anda non-translatory humeroulnar joint (Rose,1988). This analysis contributes at least twoadditional features in which modern catar-rhines are derived relative to platyrrhines
and primitive fossil catarrhines (Aegyptopi-thecus and Pliopithecus; see Fig. 6): catar-rhine ulnae 1) are more robust, with broadand deep shafts and articular surfaces, and2) have relatively short (PD) sigmoid notches(Fig. 5). The functional significances of thesefeatures are not clear.
Another non-metric feature of the ulna offunctional and phylogenetic import is themorphology of the proximal radioulnar joint.The position of the radius relative to theulna varies widely in anthropoid primates(Sarmiento, 1985; Rose, 1988). New Worldmonkeys like Alouatta, and fossils like Ae-gyptopithecus (Fleagle et al., 1975), repre-sent the primitive form in which the radialhead lies anteriolaterally to the proximalulna and anteriorly overlaps with roughlyone half of the width of the ulnar shaft (Fig.5c, anterior view). Therefore, the radius ispartially buttressed by the ulnar shaft medi-ally and posteriorly. The oval, tilted radialhead in these taxa fits in the radial notchmost tightly when the forearm is fully pro-nated; this ‘‘close-packed’’ fit is the positionof maximum stability (Rose, 1983, 1988).Cercopithecoids and hominoids (especiallygreat apes) are both derived in their radialnotch morphology (Rose, 1988, 1993).
Cercopithecoids
The elbow joint of cercopithecoids displaysa suite of derived features (Rose, 1983, 1988;Harrison, 1987; Ciochon, 1993). Those spe-cific to the ulna include: an unevenly curvedradial notch; limited, laterally facing area ofthe trochlear notch for articulation withlateral side of humeral trochlea; anteriorlyfacing radial notch (Rose, 1988); wide radialnotch articular surface; radial notch posi-tioned more posteriorly; and a narrow troch-lear articular surface (Ciochon, 1993). Themultivariate analysis here is consistent withthese studies. The cercopithecoid ulna isderived in its exceptionally narrow distaltrochlea and sigmoid notch (see Fig. 5a–c).Multivariate results also point to the nar-row, deep olecranon process, and a deep (aswell as narrow) sigmoid notch. Note thatdiscussing character ‘‘states’’ resulting fromthe multivariate analyses often requires im-posing categorical boundaries to continuoustraits (Ciochon, 1993). While few of these
267FEJEJ ULNA AND EVOLUTION OF THE ELBOW
Article ID # 911 (disk)@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_911z13 dawn

traits are discrete, they are nonethelessimportant in characterizing ulnar shape.
The functional significance of olecranonprocess morphology has been discussed insome detail (Oxnard, 1963; Bown et al.,1982; Fleagle, 1988; Harrison, 1989; Hilde-brand, 1995). In anthropoids, an elongatedolecranon is indicative of quadrupedalism,with arboreal quadrupeds possessing thelongest olecranon and terrestrial quadru-peds possessing a posteriorly expanded (and,therefore, deep) olecranon process, in bothproviding effective leverage for the m. tri-ceps brachii.
Rose (1983, 1988) noted that the narrowtrochlear articular surface of cercopithe-coids is partially convergent on that of morecursorially adapted mammals (Jenkins,1973; Hildebrand, 1995; see also Conroy,1976). Reduction in trochlear size may berelated to increased radial head size, inwhich force transmission is shifted from theulno-humeral to the radio-humeral joint(Conroy, 1976; Hildebrand, 1995). Trochlearbreadth shape is plotted in Figure 8, andcercopithecoids exhibit the narrowest troch-leae in most cases. Generally, the moreterrestrial taxa have narrower ulnae, but M.fascicularis appears to have the narrowesttrochlea in this sample (Fig. 8). This ispartly due to the flaring coronoids in M.
nemestrina and Papio that influenced mea-sures of trochlear width taken here. Thesetaxa have very narrow trochleae near thebase of the trochlear notch and the ex-panded coronoid may be allometrically re-lated to weight support in these larger-bodied monkeys.
Recent experimental data shed new lighton the functional significance of the cercopi-thecoid trochlear articular surface. In OldWorld monkeys, the trochlear articular sur-face faces medially at the coronoid beak,spirals anteriorly to the base of the notch,and wraps laterally onto the olecranon pro-cess (see Papio in Fig. 5). Rose (1983) notesthat the humeral articulation of cercopithe-coids is designed to withstand primarilymedial forces tending to adduct the ulna (seealso Jenkins, 1973). All cercopithecoid taxaexamined here exhibit this morphology, butthe semi-terrestrial baboon ulna appearsmost specialized in the extent of articularsurface dedicated to resisting this mediallydirected force on the forearm. Experimentaldata corroborate this interpretation. Schmitt(1994) documented medial ground reactionforces in the baboon while walking on theground and Demes et al. (in press) recordedmediolateral bending as the dominant stressregime in vivo in the macaque ulna duringnormal terrestrial walking. Therefore, thenarrow ulna of cercopithecoids is apparentlydesigned as a stable hinge joint for anteropos-terior movements that are accompanied by aregular medially directed force during thestance phase of locomotion. Because of thismedial ‘‘adducting’’ ground reaction force,the coronoid portion of the ulnar trochlea isbuttressed against the medial lip of thehumeral trochlea and the laterally facingolecranon part of the ulnar articulation con-tacts the heavily developed lateral surface ofthe humeral olecranon fossa. In effect, thetrochlear notch pivots around the proximo-distal axis of the distal humerus duringmovement about the mediolateral axis. Thebuttressing that resists this regular medialforce may contribute to humeroulnar stabil-ity during running. An exaggerated form ofthis construction can be seen in many curso-rial mammals (Hildebrand, 1995).
A number of the characteristic features ofthe cercopithecoid ulna involve the radial
Fig. 8. Trochlear breadth shape reflects the width ofthe coronoid process relative to the size (GM) of the ulna.Living apes and more derived fossil apes have muchbroader coronoid processes, and trochleae in general,than other anthropoids.
268 B.G. RICHMOND ET AL.
Article ID # 911 (disk)@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_911z13 dawn
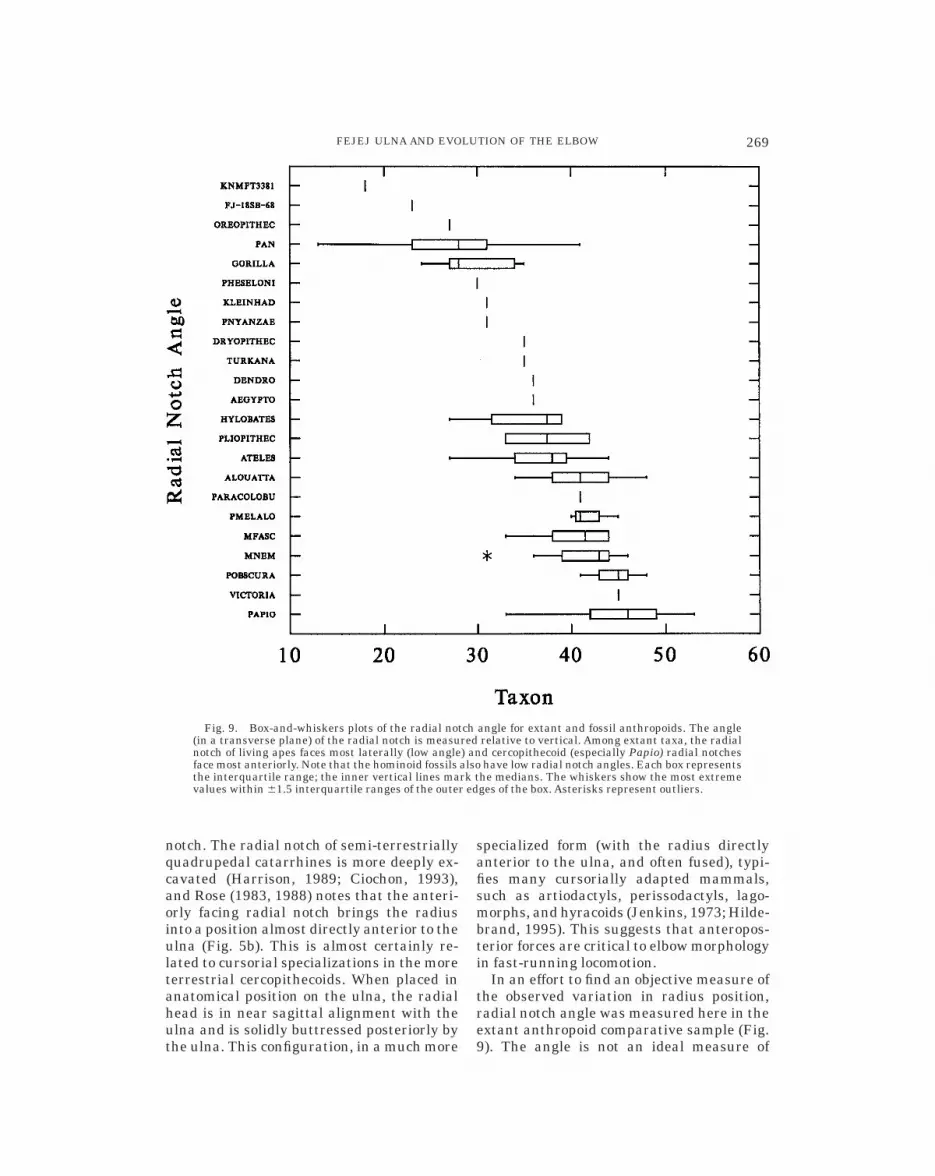
notch. The radial notch of semi-terrestriallyquadrupedal catarrhines is more deeply ex-cavated (Harrison, 1989; Ciochon, 1993),and Rose (1983, 1988) notes that the anteri-orly facing radial notch brings the radiusinto a position almost directly anterior to theulna (Fig. 5b). This is almost certainly re-lated to cursorial specializations in the moreterrestrial cercopithecoids. When placed inanatomical position on the ulna, the radialhead is in near sagittal alignment with theulna and is solidly buttressed posteriorly bythe ulna. This configuration, in a much more
specialized form (with the radius directlyanterior to the ulna, and often fused), typi-fies many cursorially adapted mammals,such as artiodactyls, perissodactyls, lago-morphs, and hyracoids (Jenkins, 1973; Hilde-brand, 1995). This suggests that anteropos-terior forces are critical to elbow morphologyin fast-running locomotion.
In an effort to find an objective measure ofthe observed variation in radius position,radial notch angle was measured here in theextant anthropoid comparative sample (Fig.9). The angle is not an ideal measure of
Fig. 9. Box-and-whiskers plots of the radial notch angle for extant and fossil anthropoids. The angle(in a transverse plane) of the radial notch is measured relative to vertical. Among extant taxa, the radialnotch of living apes faces most laterally (low angle) and cercopithecoid (especially Papio) radial notchesface most anteriorly. Note that the hominoid fossils also have low radial notch angles. Each box representsthe interquartile range; the inner vertical lines mark the medians. The whiskers show the most extremevalues within 61.5 interquartile ranges of the outer edges of the box. Asterisks represent outliers.
269FEJEJ ULNA AND EVOLUTION OF THE ELBOW
Article ID # 911 (disk)@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_911z13 dawn

radial position (i.e., lateral, anterolateral,and anterior) because of the influence ofother structures, such as the height of thecoronoid process in some taxa (e.g., Papio,see Fig. 5b). There is a great deal of varia-tion in the angle of the radial notch, butcercopithecoids do possess significantlyhigher angles (t-test, P , 0.001) than othergroups (Fig. 9). Platyrrhines examined herehave intermediate radial notch angles.Among cercopithecoids, the terrestrial quad-rupeds possess the largest radial angles(Fig. 9). The radial notches of the mostterrestrial cercopithecoids possess two dis-tinct (e.g., Papio) or partially connected (M.nemestrina) articular facets—one facing al-most directly laterally on the lateral surfaceof the coronoid beak and another facingvirtually anteriorly (nearly 90° to the sagit-tal plane) on the anterior surface of theulnar shaft. Morphology suggests that taxawith high (more anterior) radial angles arecharacterized by habitually pronated fore-arm use (Rose, 1983, 1988; Hildebrand, 1995).This is consistent with the higher radial angles(Fig. 9) and more flattened radial heads (Rose,1988) found in terrestrial quadrupeds.
In sum, the morphology specific to cercopi-thecoids appears to reflect adaptations to-wards terrestriality not seen in other ulnaeexamined here (see also Harrison, 1989;Ciochon, 1993).
Hominoids
Ulnar morphology in hominoids has re-ceived a considerable amount of attention,resulting in a long list of features identifiedas derived for hominoids. These include alaterally facing radial notch, relatively shortolecranon beak, distally wide trochlear notch,marked median ridge in trochlear notch,surface for articulation with the medial hu-meral trochlear keel that occupies the wholelength of the trochlear notch, extensive areafor articulation with humeral lateral keeland surface in the olecranon fossa, reducedolecranon process, strongly developed supi-nator ridge, relatively low and broad troch-lear notch, and a more robust, less bilater-ally compressed shaft (Morbeck, 1975;Fleagle et al., 1975; Fleagle and Kay, 1987;Sarmiento, 1985; Harrison, 1987; Rose, 1988,1997). Despite the numerous studies on thehominoid elbow, the proximal ulna of homi-
noids has not been investigated previouslyusing multivariate methods (of which we areaware). The CVA here identifies featureswhich hominoids share with cercopithecoidsas modern catarrhine traits, including thebroad and deep shaft and articular surfacesand a relatively short sigmoid notch. Homi-noids are distinguished here by their excep-tionally wide trochlea and sigmoid notch,depth of bone posterior to the sigmoid notch,and broad but otherwise small olecranonprocess (see Fig. 6).
Relative trochlear breadth (ratio of troch-lear breadth to GM) distinguishes all extantand some fossil apes from other anthropoidtaxa (Fig. 8). The broad trochlea is undoubt-edly related to the presence of a trochlearkeel. The trochlear keel has been argued tobe an adaptation for increasing mediolateraland/or rotational stability of humeroulnarjoint for climbing/suspensory locomotion inapes (Jenkins, 1973; Sarmiento, 1985; Rose,1988). This is consistent with the enlargedmusculature for pronation and supination,including the ulnar head of m. pronator terestypically found in living apes but not inprosimians and monkeys (Howell andStraus, 1933).
Despite substantial variation, extanthominoids have significantly lower radialnotch angles (i.e., laterally facing radialnotches) than other anthropoids (Fig. 9).These results are broadly similar to measure-ments taken on hominoid ulnae bySarmiento (1985); however, our measure-ments are consistently higher, indicatingthat slightly different methods may havebeen used and preclude direct comparisons.Just as the anteriorly facing radial notch isargued to be an adaptation for stability andpronated forearm posture in cercopithe-coids, the laterally positioned radius in homi-noids is probably associated with forearmrotational mobility (Fig. 5; Jenkins, 1973;Rose, 1983; Sarmiento, 1985). This interpre-tation is supported by the fact that Ateles, aplatyrrhine with unusual forearm mobility(Larson, in press), has a laterally facingradial notch comparable to gibbons. A num-ber of possible advantages of forearm rota-tional mobility have been put forth, includ-ing increased proficiency in climbing (Prost,1965; Stern, 1971; Stern and Oxnard, 1973;Fleagle, 1976; Fleagle et al., 1981; Sar-
270 B.G. RICHMOND ET AL.
Article ID # 911 (disk)@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_911z13 dawn

miento, 1987), brachiation (Jenkins, 1981),and below-branch suspension (Grand, 1972).Range of rotational motion in cadaveric pri-mates is empirically higher in taxa withmore laterally facing radial notches(O’Connor and Rarey, 1979; Sarmiento,1985). The lateral radial position also sug-gests that anteroposterior forces either playa lesser role in the hominoid radius or thatforce is somehow transferred to the highlystabilized humero-ulnar joint. Thus, it ap-pears that the hominoid ulna dedicates arelatively greater proportion of its articularsurface to the humerus than to the radiuscompared to other anthropoids, presumablyrelated to increased mobility (and lessweight-transmission?) of the radius.
The abbreviated length of the olecranonprocess in hominoids has been noted bymany (e.g., Oxnard, 1963; Morbeck, 1975;Fleagle et al., 1975; Fleagle, 1983, 1988;Sarmiento, 1985; Harrison, 1987; Rose,1988). Reduced length (and depth?) of theolecranon process may be necessary to allowthe full elbow extension important to suspen-sory behaviors (Sarmiento, 1987). Althoughshort, the olecranon is broad in apes, prob-ably to maintain adequate surface area forthe insertion of mm. triceps brachii.
Evolution of the elbow
The multivariate analysis (Fig. 6) pre-sented here supports previous observationsthat the ancestral catarrhine ulna moreclosely resembles platyrrhines than eithergroup of extant catarrhines (Fleagle et al.,1975; Fleagle, 1983; Rose, 1983). This is trueof Aegyptopithecus, Pliopithecus, and Den-dropithecus, which are all undisputable cat-arrhines based on derived craniodental mor-phology (Fleagle et al., 1975; Harrison, 1987;Fleagle, 1988; Simons, 1995). From the fairlygeneralized morphology observed in Aegypto-pithecus (Fig. 10), two basic trends follow:one with specializations for terrestriality,including an anteriorly facing and centrallypositioned radial notch and a narrow troch-lea seen in cercopithecoids, and anotherwith various degrees of specializations forincreased forearm rotational mobility, includ-ing a laterally facing and positioned radialnotch and a wide trochlear notch.
In Victoriapithecus and Paracolobus, thederived morphology of extant cercopithe-
coids is already present (von Koenigswald,1969; Szalay and Delson, 1979; Harrison,1987; McCrossin and Benefit, 1992, 1994).Both fossils possess narrow trochleae (Fig.8), anteriorly facing radial notches (Fig. 9),and long, narrow olecranon processes. Victo-riapithecus more closely resembles the cerco-pithecines, with its more deeply excavatedradial notch that is partially separated intotwo subfacets (Fig. 10). Furthermore, thetrochlear surface indicates considerable re-sistance to the medial adducting force ob-served in terrestrial but not arboreal locomo-tion (Schmitt, 1994). This and other features,such as the posteriorly expanded olecranonprocess, are consistent with interpretationsof a semi-terrestrial positional behavior withrestricted forearm rotational mobility forVictoriapithecus (Fleagle, 1988; Harrison,1989). The cercopithecine-like morphologyin this early cercopithecoid leads us to agreewith Ciochon (1993, p. 212) that the ‘‘locomo-tor adaptations of the earliest cercopithe-coids must have tended towards cursorial-ity,’’ and that some of the arborealadaptations of living colobines may repre-sent secondary adaptations to an arboreallifestyle (see also Ripley, 1979; Andrews,1982; Strasser, 1988).
It is clear from the multivariate results(Fig. 7) that the Fejej ulna is not a cercopithe-coid, as it lacks a narrow trochlea, broadradial notch, and deep (AP) shaft. With thelimited morphology preserved, the multivar-iate analysis failed to discriminate the otherextant and fossil anthropoids reliably. Phe-netically, the Fejej ulna is most similar toPliopithecus, Turkanapithecus, Aegyptopithe-cus, and Dendropithecus—a UPGMAcluster-ing analysis (not shown) links FJ-18SB-68with these taxa to the exclusion of all others,including extant taxa. However, it differsfrom Pliopithecus and Dendropithecus, whichdo not possess the prominent depression (or‘‘medial buttressing,’’ Rose, 1997) for theinsertion of brachialis exhibited by FJ-18SB-68 (Fig. 4). Furthermore, Pliopithecusis not known from Africa, unless the distalhumerus from Moghara (Simons, 1994) isconsidered to be Pliopithecus. The largebrachialis depression is present in Aegytopi-thecus and primitively retained in fossil andextant apes, including proconsulids (see P.nyanzae and P. heseloni) and Turkanapithe-
271FEJEJ ULNA AND EVOLUTION OF THE ELBOW
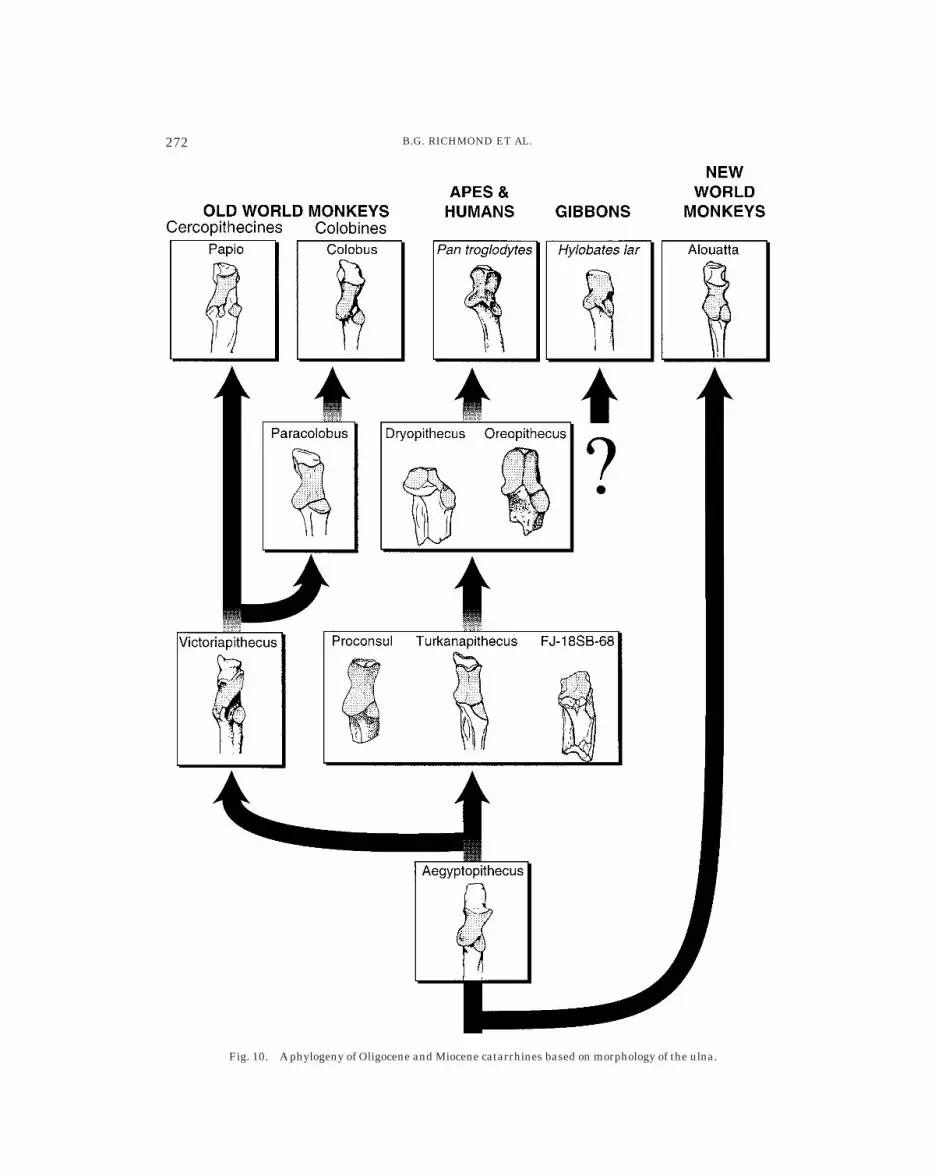
Fig. 10. A phylogeny of Oligocene and Miocene catarrhines based on morphology of the ulna.
272 B.G. RICHMOND ET AL.

cus (Figs. 5 and 9; Rose, 1983, 1997), andsuggests the presence of a large brachialismuscle for powerful forearm flexion in thesetaxa. The trochlea of the Fejej ulna is notwide, as in extant apes, but resembles otherMiocene forms such as Turkanapithecus andDendropithecus (Fig. 8). The radial notch ofFJ-18SB-68 is extreme in its lateral orienta-tion and is one of the only features linking itwith extant hominoids (Fig. 9). Like Turka-napithecus and Proconsul, the midline of thetrochlear notch in the Fejej ulna is slightlyraised and may be considered an incipienttrochlear keel. Of known fossil ulnae, that ofTurkanapithecus is most similar to FJ-18SB-68, and not unreasonable biogeographically.
Locality FJ-18 may be contemporaneouswith the nearby site of Buluk in northernKenya (Leakey and Walker, 1985; Tiffney etal., 1994). The primates at Buluk includeProhylobates sp., Simiolus enjiessi, Afropithe-cus turkanensis (Leakey et al., 1988), andpossibly Limnopithecus evansi (Rose et al.,1992). Both sites are geographically andtemporally close to Kalodirr, a 16–18 My-oldlocality west of Lake Turkana, Kenya, thathas yielded Turkanapithecus remains as wellas Afropithecus and Simiolus (Leakey andLeakey, 1986). The morphological similari-ties and approximate contemporaneity ofFejej with Kalodirr suggest that FJ-18SB-68may represent Turkanapithecus, but thefragmentary nature of the fossil ulna doesnot allow us to rule out other possibilities.
The morphology of the Fejej ulna appearsmuch like other primitive Miocene homi-noids (i.e., Proconsul, Turkanapithecus, ‘‘Aus-triacopithecus’’) and functional interpreta-tions are similar. In overall shape, the Fejejulna resembles arboreal quadrupeds (Fig.7), without the narrow trochlea and anteri-orly facing radial notch of terrestrial quadru-peds nor the wide trochlea and well-devel-oped median keel of living apes. Rose (1983)points out that the functional morphology ofthe primitive Miocene apes cannot be consid-ered ape-like or monkey-like (see also Rose,1993, 1994; Ward et al., 1991, 1993). Rose(1988, 1993) and Ward et al. (1993) agreethat Turkanapithecus and Proconsul are ar-boreal quadrupeds, but do not consider ex-tant monkeys as appropriate behavioral ana-logs (contra Andrews and Pilbeam, 1996).
They reconstruct slow, deliberate climbingas important components of their locomotorrepertoire, and this seems to be the case inthe Fejej ulna and ‘‘Austriacopithecus’’ (Be-gun, 1992a) as well. This is consistent withthe lateral orientation and position of theirradial notches (Figs. 4, 5, 10), moderatelybroad trochleae (Fig. 8), poorly developedtrochlear keel (Figs. 4, 5), and large brachia-lis insertions for strong elbow flexion.
The development of an ulnar trochlearkeel and spool-shaped humeral trochlea maybe related to biomechanical consequences oftail loss in these large arboreal quadrupeds(Richmond, unpublished ms.; Kelley, 1995).Forearm pronation and supination may playan important role in maintaining balanceabove a branch, especially without the helpof a tail (Larson and Stern, in prep). If this isthe case, then features related to forearmrotational mobility (e.g., laterally facing ra-dial notch, enlarged rotational musculature,including an ulnar head of m. pronator teres)and humero-ulnar stability (widened spool-shaped trochlea and a trochlear keel) thatare typically associated with climbing andsuspension may have evolved in arborealquadrupedal ape ancestors as an adaptationfor above-branch balance.
The trend towards powerful forearm flex-ion, broad trochleae, and displacement ofthe radius from an anterior position to amore lateral one is most strongly expressedin extant great apes. Of the fossil ulnaeconsidered here, only Dryopithecus, Oreopi-thecus, and KNM-FT 3381 have trochlea aswide as living apes (Fig. 8). The wide troch-lea is undoubtedly related to the presence ofa trochlear keel and Dryopithecus and Oreo-pithecus are the only fossil apes examinedhere that possess a well-developed trochlearkeel. KNM-FT 3381 is unusual in that itdoes not possess a trochlear keel, but has anexceptionally broad coronoid. This enig-matic specimen has been considered as ‘‘pos-sibly hominoid’’ (Rose, 1983, p. 190), possiblyoreopithecid (Harrison, 1992), and attrib-uted to Kenyapithecus wickeri (Kelley andPilbeam, 1986) and Hyainailouros sulzeri orMegistotherium (Pickford, cited in Senut,1989); it does appears to be a primate (Kitkoand Richmond, 1997). If it does repre-
273FEJEJ ULNA AND EVOLUTION OF THE ELBOW

sent Kenyapithecus, then it exhibits astrange mixture of features (and is too largeto match with the distal humerus, KNM-FT2751). The laterally facing radial notch (Fig.9), wide trochlea (Fig. 8), and deep buttress-ing posterior to the sigmoid notch are consis-tent with the ape-like descriptions of otherKenyapithecus forelimb remains fromMaboko Island (McCrossin and Benefit,1997a). On the other hand, the narrow baseof the sigmoid notch, lack of a median troch-lear keel, narrow and deep shaft, and ex-panded coronoid resemble the condition inquadrupeds (terrestrial in some features),morphology that is consistent with a suite ofclear terrestrial features (e.g., retroflexedhumeral curvature, posteriorly directed me-dial epicondyle, long and retroflected olecra-non process, dorsal transverse ridge on meta-carpal, and relatively straight phalanges;Richmond, 1995; McCrossin and Benefit,1997b). Additional work on this fossil apewill hopefully clarify the functional andphyletic position of Kenyapithecus.
The phyletic position of Oreopithecus haslong been debated because of the combina-tion of primitive craniodental features andunusual dentition (Simons, 1960; DelsonandAndrews, 1975; Szalay and Delson, 1979;Harrison, 1986) and the apparently derivedpostcranial similarities with living homi-noids (Hurzeler, 1958; Szalay and Delson,1979; Harrison, 1986; Jungers, 1987;Sarmiento, 1987; Harrison and Rook, 1997).In this multivariate analysis, Oreopithecusdoes not fall within the range of extanthominoids examined here (Fig. 6); however,it is the most ape-like in this sample, particu-larly when derived hominoid features areconsidered (Fig. 10). These are a well-developed trochlear keel, wide trochlea, lat-erally facing radial notch, and short, broadolecranon, among others (Rose, 1997; Harri-son and Rook, 1997).
Whatever the phylogenetic affinities, Oreo-pithecus was undoubtedly very ape-like interms of locomotor behavior, with a greatdeal of below-branch suspension and clam-bering (see also Jungers, 1987; Fleagle,1988).
Unlike Oreopithecus, Dryopithecus is gen-erally allied with great apes based on cra-nial and postcranial morphology (Andrews
and Martin, 1987; Begun, 1992a,b, 1993;Moya-Sola and Kohler, 1996; Begun andKordos, 1997; but see McCrossin and Ben-efit, 1994). In the limited ulnar proportionsavailable, it does not clearly ally with thegreat apes (Fig. 7), but other characteristicslead us to agree with its hominoid status.Namely, it shares with extant apes the well-developed trochlear keel, laterally posi-tioned and oriented radial notch, broad troch-lea (Fig. 8), and numerous features fromother parts of the skeleton (Moya-Sola andKohler, 1996). Dryopithecus, too, most likelyspent a considerable amount of time inclimbing and suspensory behaviors (see alsoBegun, 1988, 1993; Moya-Sola and Kohler,1996).
SUMMARY
New fossil ulnar remains represent thefirst Miocene ape from Ethiopia. Multivari-ate analyses presented here support previ-ous observations that the ancestral condi-tion for the elbow region of catarrhines isbasically platyrrhine-like, rather than likethat of either extant catarrhine group. Fromthis fairly generalized template, two basictrends have developed: one toward restrictedforearm rotational mobility and more habitu-ally pronated forearms, as seen in living andfossil cercopithecoids, and another towardgreater forearm rotational mobility leadingto modern hominoids. Fossil taxa like Victo-riapithecus and Paracolobus already pos-sess the derived ulnar morphology that char-acterizes extant cercopithecoids. Thismorphology includes a very narrow andshort trochlear notch, more anteriorly facing(and more centrally positioned) radial notch,long and deep olecranon process, and rela-tively robust proximal shaft.
Fossil ulnae of the Miocene, includingthose of Turkanapithecus, Proconsul, andFJ-18SB-68 share with extant hominoids alaterally positioned and oriented radial notchand slightly more of a trochlear keel thanseen in most extant monkeys. These charac-teristics, along with the retention of a large,medial depression for the insertion of m.brachialis, suggest increased rotational mo-bility and use of the forearm in nonpronatedhand postures. This functional anatomy is
274 B.G. RICHMOND ET AL.

consistent with an increased role of climbingin an otherwise arboreal quadrupedal life-style and represents what may be a grade oflocomotor adaptation in these early-middleMiocene primitive apes.
Oreopithecus and Dryopithecus share addi-tional features with living hominoids, includ-ing a true trochlear keel and the accompany-ing increase in trochlear breadth, projectingcoronoid beak, and markedly reduced olecra-non process. These features are presumablyrelated to elbow joint stability in a widerange of supinated–pronated hand posturesand full elbow extension and stability insuspensory postures, especially in associa-tion with relatively large body size. Thisanalysis supports the suggestion that manyof the suspensory and, especially, climbingfeatures of extant hominoids are furtherdevelopments of earlier catarrhine forelimbadaptations.
ACKNOWLEDGMENTS
Casts of KNM-WK16950R and KNM-WK17171I-M were kindly loaned by Dr.M.D. Rose, and a cast of RUD22 was loanedby Dr. D.R. Begun. We thank Dr. R.W.Thorington, Jr., Curator of Mammals at theSmithsonian Institution’s National Museumof Natural History, Dr. Ross MacPhee, Cura-tor of Mammals at the American Museum ofNatural History, and Dr. Meave Leakey,Director of Palaeontology at the NationalMuseum of Kenya, for permission to exam-ine the specimens in their care. We areespecially grateful to T. Terfa and B. Asfawfor permitting this fieldwork in 1990, andDrs. K. Begashaw and C. Tesfatsion forpermission in 1992. T. White (University ofCalifornia, Berkeley) and G. Suwa (Univer-sity of Tokyo) suggested that J.G.F. work inthe Fejej area and provided invaluable ad-vice and assistance. T. Bown (U.S. Geologi-cal Survey), T. Rassmusen (Washington Uni-versity, St. Louis), Z. Assefa (SUNY, StonyBrook), S. Yirga (Addis Ababa University), T.Hagos, A. Nagesha, C. Tilahun, and M.Fisseha (Ministry of Culture and SportsAffairs, Ethiopia) contributed greatly in thesuccess of the fieldwork. The artwork inFigures 3, 4, and 5 was prepared by Williamand Barry E. Yee and B.G.R.; Luci Betti-Nash prepared Figure 10. We also thank Bill
Jungers, John Polk, and Mike Lague forhelpful discussions and statistical advice.Earlier drafts of this paper were improvedby comments from Laura MacLatchy, JohnPolk, Terry Harrison, and two anonymousreviewers.
LITERATURE CITED
Andrews PJ (1982) Ecological polarity in primate evolu-tion. Zool. J. Linn. Soc. 74:233–244.
Andrews PJ, and Martin LB (1987) Cladistic relation-ships of extant and fossil hominoids. J. Hum. Evol.16:101–118.
Andrews PJ, and Pilbeam DR (1996) The nature of theevidence. Nature 379:123–124.
Andrews PJ, and Walker AC (1976) The primate andother fauna from Fort Teman, Kenya. In GL Isaac andER McCown (eds.): Human Origins: Louis Leakey andthe East African Evidence. Menlo Park, NJ: W.A.Benjamin, pp. 279–304.
Asfaw B, Beyene Y, Semaw S, Suwa G, White T, andWoldeGabriel G (1991) Fejej:Anew paleoanthropologi-cal research area in Ethiopia. J. Hum. Evol. 21:137–143.
Begun DR (1988) Catarrhine phalanges from the LateMiocene (Vallesian) of Rudabanya, Hungary. J. Hum.Evol. 17:413–438.
Begun DR (1992a) Phyletic diversity and locomotion inprimitive European hominids. Am. J. Phys. Anthro-pol. 87:311–340.
Begun DR (1992b) Miocene fossil hominids and thechimp–human clade. Science 257:1929–1933.
Begun DR (1993) New catarrhine phalanges from Ruda-banya (northeastern Hungary) and the problem ofparallelism and convergence in the hominoid postcra-nial morphology. J. Hum. Evol. 24:373–402.
Begun DR, and Kordos L (1997) Phyletic affinities andfunctional convergence in Dryopithecus and otherMiocene and living hominoids. In DR Begun, CVWard, and MD Rose (eds): Function, Phylogeny, andFossils: Miocene Hominoid Evolution and Adapta-tions. New York: Plenum Press, pp. 291–316.
Berggren WA, Kent DV, Flynn JJ, and Van Couvering JA(1985) Cenozoic geochronology. Geol. Soc. Am. Bull.96:1407–1418.
Birchette MG (1982) The Postcranial Skeleton of Para-colobus chemeroni. Ph.D. dissertation, Harvard Uni-versity.
Bown TM, Kraus MJ, Wing SL, Fleagle JG, Tiffney BH,Simons EL, and Vondra CF (1982) The Fayum Pri-mate Forest revisited. J. Hum. Evol. 11:603–632.
Bown TM, Fleagle JG, Kappelman J, Swisher CC, andYirga S (1994) Age of Pliocene and Miocene primatesfrom Fejej, Ethiopia (abstract). Am. J. Phys. Anthro-pol. 18:57–58.
Cande SC, and Kent DV (1995) Revised calibration ofthe geomagnetic polarity time scale for the LateCretaceous and Cenozoic. J. Geophys. Res. 100:6093–6095.
Ciochon RL (1993) Evolution of the Cercopithecoidforelimb: Phylogenetic and functional implicationsfrom morphometric analyses. Univ. Calif. Publ. Geol.Sci. 138:1–251.
Conroy GC (1976) Primate postcranial remains from theOligocene of Egypt. Contr. Primatol. 8:1–123.
Corruccini RS (1978) Morphometric analysis: Uses andabuses. Yrbk. Phys. Anthropol. 21:134–150.
Darroch JN, and Mosimann JE (1985) Canonical andprincipal components of shape. Biometrika 72:241–252.
275FEJEJ ULNA AND EVOLUTION OF THE ELBOW

Delson E, and Andrews P (1975) Evolution and interrela-tionships of the Catarrhine primates. In WP Luckettand FS Szalay (eds): Phylogeny of the Primates. NewYork: Plenum, pp. 405–446.
Demes B, Stern JT, Hausman MR, Larson SG, McLeodKJ, Rubin CT (in press) Patterns of strain in themacaque ulna during functional activity. Am. J. Phys.Anthropol.
Ehrensberg K (1938) Austriacopithecus, ein neuer men-schen-affenartiger Primate aus dem Miozan von Klein-Hadersdorf bei Poysdorf in Niederosteffeich (Nieder-Donau). S. Per. Akad. Wiss. Wies., Math-nat., abstr.1147:71–100.
Fisher RA (1953) Dispersion on a sphere. Proc. Roy. Soc.217:295–305.
Fleagle JG (1976) Locomotion and posture in the Ma-layan siamang and implications for hominoid evolu-tion. Folia primatol. 26:245–269.
Fleagle JG (1983) Locomotor adaptations of Oligoceneand Miocene hominoids and their phyletic implica-tions. In RS Corruccini and RL Ciochon (eds): NewInterpretations of Ape and Human Ancestry. NewYork: Academic Press, pp. 301–324.
Fleagle JG (1988) Primate Adaptation and Evolution.New York: Academic Press.
Fleagle JG, and Kay RF (1987) The phyletic position ofthe Parapithecidae. J. Hum. Evol. 16:483–532.
Fleagle JG, Simons EL, and Conroy GC (1975) Ape limbbone from the Oligocene of Egypt. Science 189:135–137.
Fleagle JG, Stern JT, Jungers WL, Susman RL, VangorAK, and Wells JP (1981) Climbing: A biomechanicallink with brachiation and bipedalism. Symp. Zool.Soc. Lond. 48:359–373.
Fleagle JG, Yirga S, Bown TM, Rasmussen DT, Assefa Z,Hagos T, and Negash A (1994) New paleontologicaldiscoveries from Fejej, southern Omo, Ethiopia. InZewde B, Pankhurst R, and Beyene T (eds): Proceed-ings of the Eleventh International Conference ofEthiopian Studies. Addis Ababa: Addis Ababa Univer-sity, pp. 15–22.
Fleck RJ, Sutter JF, and Elliott DH (1977) Interpreta-tion of discordant 4OAr/39Ar age spectra of Mesozoictheoleiites from Antarctica. Geochim. Cosmochim.Acta 41:15–32.
Grand TI (1972) A mechanical interpretation of terminalbranch feeding. J. Mammal. 53:198–201.
Harrison T (1986) A reassessment of the phylogeneticrelationships of Oreopithecus bambolii Gervais. J.Hum. Evol. 15:541–583.
Harrison T (1987) The phylogenetic relationships of theearly catarrhine primates: A review of the currentevidence. J. Hum. Evol. 16:41–80.
Harrison T (1989) New postcranial remains of Victoriap-ithecus from the middle Miocene of Kenya. J. Hum.Evol. 18:3–54.
Harrison T (1992) A reassessment of the taxonomic andphylogenetic affinities of the fossil catarrhines fromFort Teman, Kenya. Primates 33:501–522.
Harrison T, and Rook L (1997) Enigmatic anthropoid ormisunderstood ape? The phylogenetic status of Oreopi-thecus bambolii reconsidered. In DR Begun, CV Ward,and MD Rose (eds): Function, Phylogeny, and Fossils:Miocene Hominoid Evolution and Adaptations. NewYork: Plenum Press, pp. 327–362.
Hildebrand M (1995) Analysis of Vertebrate Structure.New York: John Wiley and Sons.
Howell AB, and Straus WL Jr (1933) The muscularsystem. In CG Hartman and WL Straus Jr (eds.): TheAnatomy of the Rhesus Monkey. New York: Hafner,pp. 89–175.
Hurzeler J (1958) Oreopithecus bambolii Gervais: Apreliminary report. Verh. Naturf. Gesell. Basel 69:1–48.
Jenkins FA Jr (1973) The functional anatomy andevolution of the mammalian humero-ulnar articula-tion. Am. J. Anat. 137:281–298.
Jenkins FA Jr (1981) Wrist rotation in primates: Acritical adaptation for brachiators. Symp. Zool. Soc.Lond. 48:429–451.
Jungers WL (1985) Body size and scaling of limb propor-tions in primates. In WL Jungers (ed): Size andScaling in Primate Biology. New York: Plenum Press,pp. 345–381.
Jungers WL (1987) Morphometric affinities of the appen-dicular skeleton of Oreopithecus bambolii (IGF 11778).J. Hum. Evol. 16:445–456.
Jungers WL, Falsetti AB, and Wall CE (1995) Shape,relative size, and size-adjustments in morphometrics.Yrbk. Phys. Anthropol. 38:137–161.
Kelley J (1995) A functional interpretive framework forthe early hominoid postcranium (abstract). Am. J.Phys. Anthropol. 20:125.
Kelley J, and Pilbeam DR (1986) The Dryopithecines:Taxonomy, comparative anatomy, and phylogeny ofMiocene large hominoids. In DR Swindler and JErwin (eds): Comparative Primate Biology, Vol. 1:Systematics, Evolution, and Anatomy. New York: AlanR. Liss, pp. 361–411.
Kirschvink JL (1980) The least-squares line and planeand the analysis of palaeomagnetic data. Geophys.Jour. Royal Astro. Soc. 62:699–718.
Kitko R, and Richmond BG (1997) The functional andtaxonomic affinities of KNM-FT3381 (abstract). Am.J. Phys. Anthropol. 24:144.
Klecka WR (1980) DiscriminantAnalysis. Beverley Hills,CA: Sage Publications.
Kretzoi M (1975) New ramapithecines and Pliopithecusfrom the lower Pliocene of Rudabanya in northeasternHungary. Nature 257:578–581.
Larson SG (1993) Phylogeny. In BF Monrey (ed): TheElbow and Its Disorders, 2nd Edition. Philadelphia:WB Saunders, pp. 6–15.
Larson SG (in press) Parallel evolution in the hominoidtrunk and forelimb. Evol. Anthropol.
Leakey REF, and Leakey MG (1986) A new Miocenehominoid from Kenya. Nature 324:143–148.
Leakey REF, and Walker AC (1985) New higher pri-mates from the early Miocene of Buluk, Kenya. Na-ture 318:173–175.
Leakey REF, Leakey MG, and Walker AC (1988) Mor-phology of Turkanapithecus kalakolensis from Kenya.Am. J. Phys. Anthropol. 76:277–288.
LeGros Clark WE, and Thomas DP (1951) Associatedjaws and limb bones of Limnopithecus macinnesi. Fos.Mamm. Africa (BMNH) 3:1–27.
McCrossin ML, and Benefit BR (1992) Comparativeassessment of the ischial morphology of Victoriapithe-cus macinnesi. Am. J. Phys. Anthropol. 87:277–290.
McCrossin ML, and Benefit BR (1994) Maboko Islandand the evolutionary history of Old World monkeysand apes. In RS Corruccini and RL Ciochon (eds):Integrative Paths to the Past. Englewood Cliffs, NJ:Prentice Hall, pp. 95–122.
McCrossin ML, and Benefit BR (1997a) New postcranialremains of Kenyapithecus and their implications forunderstanding the origins of hominoid terrestriality(abstract). Am. J. Phys. Anthropol. 24:164.
McCrossin ML, and Benefit BR (1997b) On the relation-ships and adaptations of Kenyapithecus, a large-bodied hominoid from the Middle Miocene of easternAfrica. In DR Begun, CV Ward, and MD Rose (eds):Function, Phylogeny, and Fossils: Miocene HominoidEvolution and Adaptations. New York: Plenum Press,pp. 241–267
McDougall I, and Watkins RT (1985) Age of hominoid-bearing sequence at Buluk, northern Kenya. Nature318:175–178.
276 B.G. RICHMOND ET AL.

McHenry HM, and Corruccini RS (1975) Distal humerusin hominoid evolution. Folia Primatol. 23:227–244.
Morbeck ME (1975) Dryopithecus africanus forelimb. J.Hum. Evol. 4:39–46.
Morbeck ME (1983) Miocene hominoid discoveries fromRudabanya: Implications from the postcranial skel-eton. In RS Corruccini and RL Ciochon (eds): NewInterpretations of Ape and Human Ancestry. NewYork: Academic Press, pp. 369–404.
Mosimann JE (1970) Size allometry: Size and shapevariables with characteristics of the lognormal andgeneralized gamma distributions. J. Am. Stat. Assoc.665:930–945.
Moya-Sola S, and Kohler M (1996) A Dryopithecusskeleton and the origins of great-ape locomotion.Nature 379:156–159.
Napier JR, and Davis P (1959) The forelimb skeletonand associated remains of Proconsul africanus. Foss.Mamm. Afr. 16:1–70.
O’Connor BL, and Rarey KE (1979) Normal amplitudesof pronation and supination in several genera ofanthropoid primates. Am. J. Phys. Anthropol. 51:39–44.
Oxnard CE (1963) Locomotor adaptations in the pri-mate forelimb. Symp. Zool. Soc. Lond. 10:165–182.
Oxnard CE (1972) Some African fossil foot bones: A noteon the interpolation of fossils into a matrix of extantspecies. Am. J. Phys. Anthropol. 37:3–12.
Oxnard CE (1975) Uniqueness and Diversity in HumanEvolution: Morphometric Studies of Australopithe-cines. Chicago: University of Chicago Press.
Prost JH (1965) A definitional system for the classifica-tion of primate locomotion. Am. Anthropol. 67:1198–1214.
Richmond BG (1995) Diaphyseal curvature in living andfossil anthropoid humeri (abstract). Am. J. Phys.Anthropol. 20:181.
Richmond BG, and Jungers WL (1995) Size variationand sexual dimorphism in Australopithecus afarensisand living hominoids. J. Hum. Evol. 29:229–245.
Ripley S (1979) Environmental grain, niche diversifica-tion, and positional behavior in Neogene primates: Anevolutionary hypothesis. In ME Morbeck, H Preus-choft, and N Gomberg (eds.): Environment, Behavior,and Morphology: Dynamic Interactions in Primates.New York: Gustav Fisher, pp. 37–74.
Rose MD (1983) Miocene hominoid postcranial morphol-ogy: Monkey-like, ape-like, neither, or both? In RLCiochon and JG Fleagle (eds): Primate Evolution andHuman Origins. New York: Aldine de Gruyter, pp.189–193.
Rose MD (1988) Another look at the anthropoid elbow. J.Hum. Evol. 17:193–224.
Rose MD (1993) Locomotor anatomy of Miocene homi-noids. In DL Gebo (ed): Postcranial Adaptation inNonhuman Primates. DeKalb, IL: Northern IllinoisUniversity Press, pp. 252–272.
Rose MD (1994) Quadrupedalism in some Miocenecatarrhines. J. Hum. Evol. 26:387–411.
Rose MD (1997) Functional and phylogenetic features ofthe forelimb in Miocene hominoids. In DR Begun, CVWard, and MD Rose (eds): Function, Phylogeny, andFossils: Miocene Hominoid Evolution and Adapta-tions. New York: Plenum Press, pp. 79–100.
Rose MD, Leakey MG, Lzakey REF, and Walker AC(1992) Postcranial specimens of Simiolus enjiessi andother primitive catarrhines from the early Miocene ofLake Turkana, Kenya. J. Hum. Evol. 22:171–237.
Sarmiento EE (1985) Functional Differences in theSkeleton of Wild and Captive Orang-utans and theirAdaptive Significance. PhD dissertation, New YorkUniversity.
Sarmiento EE (1987) The phyletic position of Oreopithe-cus and its significance in the origin of the Hominoi-dea. Am. Mus. Nov. 2881:1–44.
Schmitt D (1994) Forelimb mechanics as a function ofsubstrate type during quadrupedalism in two anthro-poid primates. J. Hum. Evol. 26:441–457.
Senut B (1989) Le coude des Primates Hominoides:Anatomie, fonction, taxonomie, evolution. Cahiers depaleoanthropologie, CNRS.
Simons EL (1960) Apidium and Oreopithecus. Nature186:824–826.
Simons EL (1994) New monkeys (Prohylobates) and anape humerus from the Miocene Moghara Formation ofNorthern Egypt. Proc. XIV Int. Primatol. Conf. Stras-sbourg, France, 247–253.
Simons EL (1995) Egyptian Oligocene primates: A re-view. Yrbk. Phys. Anthropol. 38:199–238.
Steiger RH, and Jager E (1977) Subcommission ongeochronology: Convention in the use of decay con-stants in geo- and cosmochronology. Earth Plant. Sci.Lett. 36:359–361.
Stern JT Jr (1971) Functional myology of the hip andthigh of cebid monkeys and its implications for theevolution of erect posture. Biblio. Primat., No. 14.Basel: Karger, pp. 1–318.
Stern JT Jr, and Oxnard CE (1973) Primate Locomotion:Some Links with Evolution and Morphology. Basel:Karger. Primatologia 4.
Strasser E (1988) Pedal evidence for the origin anddiversification of cercopithecoid clades. J. Hum. Evol.16:81–99.
Straus WL (1961) Primate taxonomy and Oreopithecus.Science 133:760–761.
Suwa G, White T, Asfaw B, WoldeGabriel G, and YemaneT (1991) Miocene faunal remains from the Burji-Soyama area, Amaro Horst, Southern Sector of theMain Ethiopian Rift. Palaeont. Afr. 28:23–28.
Swisher CC, Curtis GH, Jacob T, Getty AG, Suprijo A,and Widiasmoro BL (1994) Age of the earliest knownhominids in Java, Indonesia. Science 263:1118–1121.
Szalay FS, and Delson E (1979) Evolutionary History ofthe Primates. New York: Academic Press.
Tiffney BH, Fleagle JG, and Bown TM (1994) Early toMiddle Miocene angiosperm fruits and seeds fromFejej, Ethiopia. Tert. Res. 15:25–42.
von Koenigswald GHR (1969) Miocene Cercopithecoideaand Oreopithecoidea from the Miocene of East Africa.In Leakey LSB (ed): Fossil Vertebrates of Africa (1).New York: Academic Press, pp. 39–52.
Walker AC, and Pickford M (1983) New postcranialfossils of Proconsul africanus and Proconsul nyanzae.In RL Ciochon and RS Corruccini (eds): New Interpre-tations of Ape and Human Ancestry. New York: Ple-num Press, pp. 325–351.
Ward CV, Walker AC, and Teaford MF (1991) Proconsuldid not have a tail. J. Hum. Evol. 21:215–221.
Ward CV, Walker AC, Teaford MF, and Odhiambo I(1993) Partial skeleton of Proconsul nyanzae fromMfangano Island, Kenya. Am. J. Phys. Anthropol.90:77–111.
Watkins RT (1989) The Buluk Member, a fossil hominoid-bearing sedimentary sequence of Miocene age fromnorthern Kenya. J. Afr. Earth Sci. 8:107–112.
WoldeGabriel G, Yemane T, Suwa G, White T, and AsfawB (1991) Age of volcanism and rifting in the Burji-Soyoma area, Amaro Horst, southern Main EthiopianRift: Geo- and biochronologic data. J. Afr. Earth Sci.13:437–447.
Zapfe H (1958) The skeleton of Pliopithecus (Epipliopi-thecus) vindobonensis Zapfe and Hurzeler. Am. J.Phys. Anthropol. 16:441–458.
277FEJEJ ULNA AND EVOLUTION OF THE ELBOW
Related Documents











