236 Accepted by A. Contreras-Ramos: 26 Aug. 2013; published: 23 Sept. 2013 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2013 Magnolia Press Zootaxa 3716 (2): 236–258 www.mapress.com/zootaxa/ Article http://dx.doi.org/10.11646/zootaxa.3716.2.6 http://zoobank.org/urn:lsid:zoobank.org:pub:1D536BCD-4D3B-42B0-98B9-48D8BC31ECE5 First fossil larvae of Berothidae (Neuroptera) from Baltic amber, with notes on the biology and termitophily of the family SONJA WEDMANN 1 , VLADIMIR N. MAKARKIN 2,5 , THOMAS WEITERSCHAN 3 & THOMAS HÖRNSCHEMEYER 4 1 Senckenberg Forschungsinstitut und Naturmuseum, Forschungsstation Grube Messel, Markstrasse 35, D-64409 Messel, Germany 2 Institute of Biology and Soil Sciences, Far Eastern Branch of the Russian Academy of Sciences, Vladivostok 690022, Russia 3 Forsteler Strasse 1, 64739 Höchst Odw., Germany 4 Georg-August-Universität Göttingen, Johann-Friedrich-Blumenbach-Institut, Berliner Str. 28, 37073 Göttingen, Germany 5 Corresponding author. E-mail: [email protected] Abstract Four fossil larvae of Berothidae (Neuroptera) from Baltic amber are described in detail, and the main characters of a fifth larva are discussed briefly. Two first instars very probably belong to the Berothinae; the subfamilial affinities of three other (probably full-grown) larvae are unclear. The latter are characterized by features not found so far in extant taxa of Berothi- dae: antennae and labial palps with six to seven segments; ecdysial cleavage lines consist of only frontal and coronal su- tures (the lateral suture is absent); pronotal sclerites large and very close to each other along midline. However, these larvae belong with certainty to Berothidae as indicated by the structure of their mouthparts, and their general appearance. Morphological and biological data on the larvae of Berothidae are summarized and analyzed. It is presumed that termito- phily might have evolved during the Cretaceous (or in the early Cenozoic), and only in Berothinae (or in subfamilies close- ly related to this group). The Baltic amber berothid assemblage apparently included both termitophilous and non- termitophilous larvae. Key words: Neuroptera, Berothidae, larvae, Baltic amber Introduction The small family Berothidae (including the subfamily Rhachiberothinae) today comprises little more than one hundred species distributed mainly in the tropics and subtropics, and it is most diverse in the Ethiopian and Australian biogeographical regions (Aspöck 1986). Some berothids are remarkable for the termitophily of their larvae (e.g., Johnson & Hagen 1981). The Berothidae is an ancient taxon recorded confidently from the Middle Jurassic and assumed to be much older (Winterton et al. 2010; Makarkin et al. 2011). The majority of fossil species were described from the Cretaceous (Schlüter 1978; Whalley 1980; Klimaszewski & Kevan 1986; Martins-Neto & Vulcano 1990; Makarkin 1994; Ren & Guo 1996; Grimaldi 2000; Engel 2004; Nel et al. 2005a,b; Engel & Grimaldi 2008; McKellar & Engel 2009; Petrulevicius et al. 2010). Only a few berothids are known from the Cenozoic: one species from the Early Eocene North American Hat Creek locality (Archibald & Makarkin 2004) and several species (partly undescribed) from the Late Eocene Baltic, Rovno and English ambers (Krüger 1923; MacLeod & Adams 1968; Whalley 1983; Makarkin & Kupryjanowicz 2010). A list of all known fossil Berothidae is given in Makarkin et al. (2011). The larvae of only few extant berothid genera are known (Table 1). Additionally, three fossil larvae have been described from the Cretaceous (Whalley 1980; Engel & Grimaldi 2008). Hitherto, fossil berothid larvae have not been known from the Cenozoic. In this paper, we describe four berothid larvae from Baltic amber and mention a fifth larva, which we did not investigate in detail. Two of these larvae have characters similar to those of extant taxa. Some features not found, so far, in extant taxa of Berothidae distinguish the other three larvae. We summarize and analyze all available data on berothid biology, especially concerning the larval termitophily and its probable evolution. TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2013 Magnolia Press
Zootaxa 3716 (2): 236–258
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3716.2.6
http://zoobank.org/urn:lsid:zoobank.org:pub:1D536BCD-4D3B-42B0-98B9-48D8BC31ECE5
First fossil larvae of Berothidae (Neuroptera) from Baltic amber,
with notes on the biology and termitophily of the family
SONJA WEDMANN1, VLADIMIR N. MAKARKIN2,5, THOMAS WEITERSCHAN3 &
THOMAS HÖRNSCHEMEYER4
1Senckenberg Forschungsinstitut und Naturmuseum, Forschungsstation Grube Messel, Markstrasse 35, D-64409 Messel, Germany2Institute of Biology and Soil Sciences, Far Eastern Branch of the Russian Academy of Sciences, Vladivostok 690022, Russia3Forsteler Strasse 1, 64739 Höchst Odw., Germany4Georg-August-Universität Göttingen, Johann-Friedrich-Blumenbach-Institut, Berliner Str. 28, 37073 Göttingen, Germany5Corresponding author. E-mail: [email protected]
Abstract
Four fossil larvae of Berothidae (Neuroptera) from Baltic amber are described in detail, and the main characters of a fifth
larva are discussed briefly. Two first instars very probably belong to the Berothinae; the subfamilial affinities of three other
(probably full-grown) larvae are unclear. The latter are characterized by features not found so far in extant taxa of Berothi-
dae: antennae and labial palps with six to seven segments; ecdysial cleavage lines consist of only frontal and coronal su-
tures (the lateral suture is absent); pronotal sclerites large and very close to each other along midline. However, these
larvae belong with certainty to Berothidae as indicated by the structure of their mouthparts, and their general appearance.
Morphological and biological data on the larvae of Berothidae are summarized and analyzed. It is presumed that termito-
phily might have evolved during the Cretaceous (or in the early Cenozoic), and only in Berothinae (or in subfamilies close-
ly related to this group). The Baltic amber berothid assemblage apparently included both termitophilous and non-
termitophilous larvae.
Key words: Neuroptera, Berothidae, larvae, Baltic amber
Introduction
The small family Berothidae (including the subfamily Rhachiberothinae) today comprises little more than one hundred species distributed mainly in the tropics and subtropics, and it is most diverse in the Ethiopian and Australian biogeographical regions (Aspöck 1986). Some berothids are remarkable for the termitophily of their larvae (e.g., Johnson & Hagen 1981). The Berothidae is an ancient taxon recorded confidently from the Middle Jurassic and assumed to be much older (Winterton et al. 2010; Makarkin et al. 2011). The majority of fossil species were described from the Cretaceous (Schlüter 1978; Whalley 1980; Klimaszewski & Kevan 1986; Martins-Neto & Vulcano 1990; Makarkin 1994; Ren & Guo 1996; Grimaldi 2000; Engel 2004; Nel et al. 2005a,b; Engel & Grimaldi 2008; McKellar & Engel 2009; Petrulevicius et al. 2010). Only a few berothids are known from the Cenozoic: one species from the Early Eocene North American Hat Creek locality (Archibald & Makarkin 2004) and several species (partly undescribed) from the Late Eocene Baltic, Rovno and English ambers (Krüger 1923; MacLeod & Adams 1968; Whalley 1983; Makarkin & Kupryjanowicz 2010). A list of all known fossil Berothidae is given in Makarkin et al. (2011).
The larvae of only few extant berothid genera are known (Table 1). Additionally, three fossil larvae have been described from the Cretaceous (Whalley 1980; Engel & Grimaldi 2008). Hitherto, fossil berothid larvae have not been known from the Cenozoic. In this paper, we describe four berothid larvae from Baltic amber and mention a fifth larva, which we did not investigate in detail. Two of these larvae have characters similar to those of extant taxa. Some features not found, so far, in extant taxa of Berothidae distinguish the other three larvae. We summarize and analyze all available data on berothid biology, especially concerning the larval termitophily and its probable evolution.
236 Accepted by A. Contreras-Ramos: 26 Aug. 2013; published: 23 Sept. 2013

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
TABLE 1. List of extant species of Berothidae whose larvae are known.
Material and methods
Four inclusions in Baltic amber are investigated in detail in this study; two of the specimens are deposited in the private collection of Thomas Weiterschan in Höchst im Odenwald (larvae A and B), one in the private collection of Christel and Hans Werner Hoffeins in Hamburg, which later will be deposited in the collection of the Senckenberg Deutsches Entomologisches Institut (SDEI) (larva C) and one specimen in the collection of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt (SF) (larva D). A fifth larva was studied only in a photograph; this specimen is now held in the private collection of Thomas Schäfer (Saland, Switzerland; his collection No. 01.03.13).
Most of the photographs were taken by Sonja Wedmann and Thomas Weiterschan using a Leica MZ12.5 stereomicroscope and an attached Nikon D300 digital camera. Extension of depth of focus was achieved by stacking several photos with the program Helicon Focus, version 5.3 X64. Photos for Figs. 1 and 2 were made with a Keyence VHX-500F digital microscope, with support by Frank Wieland at the University of Hamburg. Thomas Weiterschan prepared the drawings.
Larva D was investigated additionally with synchrotron computer tomography. Synchrotron radiation tomography data were obtained at the TOMCAT beamline (Stampanoni et al. 2006), Swiss Light Source, Paul Scherrer Institut, Villigen, Switzerland (proposal ID 20120025, to SW). Data were acquired using a 4× lens and an exposure time of 160ms with continuous specimen rotation at 10keV. A total of 1601 projections were acquired over 180°. 3D-data were processed and images produced with VSG Amira 5.4. Images were subsequently processed with Adobe Photoshop CS3.
Institutional abbreviations: SF, Senckenberg Forschungsinstitut und Naturmuseum Frankfurt, Frankfurt am Main, Germany; SDEI, Senckenberg Deutsches Entomologisches Institut Müncheberg, Germany; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, USA.
General terminology follows Snodgrass (1935), MacLeod (1964) and Beutel et al. (2010); chaetotaxy terminology follows Rousset (1966) and Hoffman & Brushwein (1992).
Subfamilies Species References
Rhachiberothinae Mucroberotha vesicaria Tjeder, 1959 Minter 1990: Figs. 5–9.
Berothimerobiinae Berothimerobius reticulatus Monserrat et Deretsky, 1999
Monserrat 2006: Fig. 11.
Nyrminae Nyrma kervilea Navás, 1933 Dobosz & Górski 2008: 33 (undescribed, but the poster contained photographs of the larva).
Berothinae Lomamyia hamata (Walker, 1853) Brushwein 1987b: Fig. 2.
Lomamyia latipennis Carpenter, 1940 Toschi 1964: Fig. 3; Tauber & Tauber 1968: Figs. 1–4 [reproduced in Tauber 1991: Fig. 33.18].
Lomamyia sp. Gurney 1947: Figs. 2, 5, 7, 8, 12, 13, 18, 22.
Podallea manselli Aspöck et Aspöck, 1988
Minter 1990: Figs. 14–18 [as Podallea sp.]; Möller 2003: Fig. 70; Möller et al. 2006.
Podallea vasseana (Navás, 1910) Möller 2003: Figs. 14–9, 71–93; Möller et al. 2006: Figs. 7–48.
Spermophorella disseminata Tillyard, 1916
Tillyard 1916: Figs. 32, 33 [reproduced in Tillyard 1926: Figs. U8D; Gurney 1947: Figs. 4, 10].
Spermophorella maculatissima Tillyard, 1916
Tillyard 1916: 323.
Subfamily uncertain Berothidae gen. sp. indet. Riek 1970: Figs. 29.10G,H [reproduced in Gepp 1984: Figs. 8a,b; New 1991: Figs. 34.10A,B; Tauber et al. 2003: Figs. 6K,L].
Berothidae gen. sp. indet. Riek 1970: Fig. 29.5B [reproduced in New 1991: Fig. 34.5B].
Zootaxa 3716 (2) © 2013 Magnolia Press · 237BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Description of fossil larvae
Order Neuroptera Linnaeus, 1758
Family Berothidae Handlirsch, 1906
Berothidae indet., larva A
Figs. 1, 2
Material examined. Specimen No. 240 in the private collection of Thomas Weiterschan (Höchst Odw, Germany); a first instar larva in a relatively small piece of Baltic amber (precise collecting locality is unknown).
Description. Larva 1.83 mm long (including mouthparts), rather slender.Head capsule elongated, 0.24 mm long, 0.16 mm wide (length/width ratio 1.5). Ocular areas present laterally,
extending outwards from antennal sockets; stemmata (= convex corneae, lenses) not distinctly visible, possibly one on each side. Dorsally, ecdysial cleavage lines distinct, consisting of frontal, lateral and coronal sutures. Two anterior arms of frontal suture converging slightly for nearly two thirds of head capsule length, from inner margin of antennal tubercles to lateral sutures; posterior arms of frontal suture (after joining lateral suture) strongly convergent until they touch. Lateral sutures straight, reaching lateral margins of head (in dorsal view), inclined slightly to anterior part of frontal sutures at nearly right angle. Coronal suture rather short. Ventrally, coxopodites of maxillae (stipes and cardo) and labium clearly visible. Stipes and cardo relatively narrow; boundary between them not clearly visible. Labium composed of prementum (= prelabium) and postmentum (= postlabium). Prementum short and relatively narrow; anteriorly excised; postmentum long and broad, trapezoid in shape.
Mandibles apically pointed. Maxillary blades very broad basally (about 0.05 mm), gradually narrowing apically; closely associated with mandibles to form mandibulomaxillary stilets, 0.22 mm long.
Antennal sockets located on relatively high projections (=antennal tubercle of MacLeod (1964)). Antennae three-segmented, 0.32 mm long; basal segment slightly conical, 0.04 mm long, 0.02 mm wide; second segment long and clearly annulated, 0.12 mm long, 0.02 mm wide; third segment long and thin, 0.08 mm long, about 0.01 mm wide, bearing fine, feebly visible apical seta, 0.08 mm long.
FIGURE 1. Berothidae indet., larva A, whole larva as preserved. A, photograph. B, drawing (both dorsal view). Scale bar is 0.5 mm.
WEDMANN ET AL.238 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 2. Berothidae indet., larva A. A–D, head in dorsal (A, C) and ventral view (B, D). E, foreleg, ventral view. an, antenna; cd, cardo; cvx, cervix; cs, coronal suture; cx, coxa; em, empodium; fm, femur; fs, frontal suture; lp, labial palpus; ls, lateral suture; md, mandible; mx, maxillary blade; prm, prementum; psm, postmentum; pt, pretarsus; st, stipes; tb, tibia; tr, trochanter; ts, tarsus. Scale bars are 0.25 mm (A, B), 0.1 mm (C–E).
Zootaxa 3716 (2) © 2013 Magnolia Press · 239BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Labial palps slightly longer than mandibles, 4-segmented. Basal segment cylindrical, 0.03 mm long, 0.02 mm wide. Second segment elongate, 0.1 mm long and slightly wider than basal one, widest at 2/3 of its length; annulation clearly visible. Third segment 0.03 mm long, slender (half as wide as second). Fourth segment not clearly separated from third, 0.06 mm long and slightly thinner than third, gradually narrowing towards pointed apex.
Cervix 0.12 mm wide, retracted into prothorax; visible length 0.08 mm ventrally, not visible dorsally. Prothorax 0.18 mm long, 0.17 mm wide; two large dorsal sclerites indistinctly visible. Mesothorax 0.13 mm
long, 0.19 mm wide; metathorax 0.12 mm long, 0.19 mm wide; both with indistinctly visible paired oval dorsal sclerites.
Legs rather short, robust: coxa relatively long and narrow; trochanter relatively small; femur narrowed distally; tibia narrow, tibia and tarsus nearly equal in length; pretarsus short bearing two short claws and long, trumpet-shaped empodium. Forelegs 0.28 mm long (if stretched out).
Abdomen divided into 10 clearly separated segments, nine of which with lateral lobes. Segments 1 to 6 gradually increasing in length from 0.09 mm to 0.12 mm, their width gradually decreasing from 0.21 mm to 0.18 mm. Segments 7 to 9 gradually decreasing in length from 0.1 mm to 0.06 mm and in width from 0.18 mm to 0.11 mm. Segment 10 slightly conical (0.07 mm long, maximum 0.07 mm wide), rounded posteriorly, without lateral lobes and appendices.
Remarks. Based on its size, the larva is considered a first instar.
Berothidae indet., larva B
Figs. 3–5
FIGURE 3. Berothidae indet., larva B, whole larva as preserved. A, photograph. B, drawing (both dorsal view). Scale bar is 0.5 mm.
WEDMANN ET AL.240 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 4. Berothidae indet., larva B. A, head, dorsal view. B, head, ventral view. C, apex of abdomen, dorsal view. Scale bars are 0.1 mm.
Material examined. Specimen No. 1555 in the private collection of Thomas Weiterschan (Höchst Odw, Germany); a larva in a small piece of Baltic amber (precise collecting locality is unknown).
Description. First instar larva preserved in slightly curved position, ca. 2.25 mm long (including mouthparts), slender.
Head capsule elongate, 0.28 mm long, 0.2 mm wide (length/width ratio 1.4). Stemmata present, but not very clearly visible; probably one on each side (alternatively, a group of 2–3 very small stemmata located close to each other on each side). Dorsally, ecdysial cleavage lines clearly visible consisting of frontal, lateral and coronal sutures. Two anterior arms of frontal suture extending for nearly two thirds of head capsule length, slightly convergent, from inner margin of antennal tubercles to lateral sutures, posterior arms of frontal suture (after joining lateral suture) rather strongly convergent until they touch. Lateral sutures straight, reaching lateral margins of head
Zootaxa 3716 (2) © 2013 Magnolia Press · 241BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
(in dorsal view), inclined to anterior part of frontal sutures at obtuse angle. Coronal suture rather long (Fig. 5B). Of dorsal setae, sockets of lateral clypeal (trichobothrian) seta clearly visible, and probably S7, S5 detected. Ventrally, coxopodites of maxillae (stipes and cardo) and labium clearly distinguished. Stipes and cardo narrow; boundary between them not clearly visible. Labium composed of prementum and postmentum. Prementum short and broad (nearly quadratic, 0.06 mm long); anterior excision not detected; postmentum long and broad, nearly parallel-sided, somewhat broadened in distal portion (Fig. 5A).
Mandibles 0.24 mm long, slender, apically pointed. Shape of maxillary blades not clearly visible; these closely associated with mandibles to form mandibulomaxillary stilets.
Antennal sockets located on high projections (antennal tubercles); left antennal tubercle obliquely directed to outside as if following antennal movement. Antennae 3-segmented, longer than mouthparts. Basal segment slightly conical, 0.04 mm long and 0.03 mm wide; second segment elongate, 0.18 mm long, clearly annulated, with distinct contraction in middle and dilated in distal half; third segment very thin, 0.1 mm long, less than 0.01 mm wide, annulated; terminal filiform seta about 0.1 mm long.
FIGURE 5. Berothidae indet., larva B, drawings of body details. A, head, ventral view. B, head, dorsal view. C, right foreleg, ventral view. D, apex of abdomen, dorsal view. an, antenna; cd, cardo; cvx, cervix; cs, coronal suture; cx, coxa; fm, femur; fs, frontal suture; lcs, lateral clypeal seta; lp, labial palpus; ls, lateral suture; md, mandible; mx, maxillary blade; prm, prementum; psm, postmentum; pt, pretarsus; S5, S7, head seta; st, stipes; su, caudal sucker; tb, tibia; tr, trochanter; trb, trichobothria; ts, tarsus; VIII to X, 8th to 10th abdominal segments. Scale bars are 0.1 mm.
WEDMANN ET AL.242 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Labial palps slightly shorter than mandibulomaxillary stilet, 4-segmented. Basal segment cylindrical, 0.03 mm long; second segment elongate, widest at 2/3 of its length, 0.08 mm long (both ca. 0.02 mm maximum wide); third segment cylindrical, 0.03 mm long and half as wide as 2nd segment; fourth segment 0.07 mm long and basally slightly thinner than third segment, gradually thinned towards rather pointed apex. Second to fourth segments annulated.
Cervix 0.18 mm wide, probably partially retracted into prothorax; visible length 0.02 mm dorsally, 0.08 mm laterally and ventrally.
Prothorax 0.23 mm long, 0.23 mm wide; two large sclerites indistinct. Mesothorax and metathorax both ca. 0.14 mm long, 0.26 mm wide; both with pair of distinct oval sclerites.
Legs moderately stout; coxa rather long, stout; trochanter very distinct, rather long; femur elongate, stout; tibia short, stout; tarsus very long; pretarsus bearing two short claws and relatively short trumpet-shaped empodium (Fig. 5C).
Abdomen divided into 10 segments, nine of which have lateral lobes. Basal segments 1 to 4 short, each ca. 0.07 mm long, ca. 0.25 mm wide; distal segments 5 to 10 gradually becoming narrower, from 0.24 mm (segment 5) to 0.07 mm (segment 10); segments 6 to 8 each ca. 0.15 mm long; segment 5 somewhat shorter; segment 9 0.1 mm long; segment 10 0.08 mm long, and caudal sucker 0.05 mm long (Figs. 4C, 5D). Segment 8 with two dorsal trichobothrian sensillae (Fig. 5D).
Remarks. Judged from its size, the larva is a first instar. The terminal segments of this larva are fully extended, and its 10th segment is terminated by a caudal sucker. Hitherto, the caudal sucker of the first instar larvae in extant berothid species has never been figured (e.g., Tillyard 1916: Fig. 32; Minter 1990: Figs. 5, 14; Möller et al. 2006: Figs. 12, 13), although its presence in larvae of this age was mentioned at least in Lomamyia occidentalis (Banks in Baker, 1905) and L. hamata (Walker, 1853) (Toschi 1964; Brushwein 1987). The caudal sucker of larva B is similar to that of the 3rd instar of Podallea vasseana (Navás, 1910) (see Möller et al. 2006: Fig. 35). Usually, the caudal sucker is invaginated and externally not visible (as in the case of the larva A; see Möller 2003: Fig. 43). Interestingly, the sucker is not used when the first instar larva runs (Toschi 1964; Brushwein 1987).
Berothidae indet., larva C
Figs. 6, 7
Chrysopidae: Janzen 2002: Fig. 58.
Material examined. Specimen No. 1270-1 in the private collection of Christel and Hans Werner Hoffeins (Hamburg, Germany); a third instar larva in a piece of Baltic amber (precise collecting locality is unknown), which itself is embedded in a block of artificial resin.
Description. Larva about 5.5 mm long (including mouthparts), preserved slightly curved at its anterior and posterior end.
Head capsule 0.47 mm long, 0.2 mm wide (length/width ratio 1.6). Stemmata not visible, but ocular area apparently present; two long setae (S4, S9) in this area clearly visible. Dorsally, ecdysial cleavage lines poorly preserved; consisting of frontal and coronal sutures. Two arms of frontal suture extending for approximately two thirds of head capsule length, convergent from inner margin of antennal tubercles until they touch; lateral sutures absent; coronal suture short. Ventral view of head mostly obscured.
Mandibles and maxillary blades forming mandibulomaxillary stilets, about 0.4 mm long; broad basally, gradually narrowing and pointed apically (but not very acute).
Antennal sockets located on distinct projections (antennal tubercles). Right antenna seven-segmented, 0.42 mm long; left antenna six-segmented, 0.43 mm long. Basal segments elongate and strongly conical, basally less than 0.01 mm wide, and distally about 0.02 mm wide. The other antennal segments rectangular, about 0.02 mm wide. Length of segments from base to top: 0.06 mm, 0.1 mm, 0.05 mm, 0.05 mm, 0.09 mm, 0.08 mm (left antenna); 0.06 mm, 0.08 mm, 0.03 mm, 0.03, 0.05 mm, 0.09 mm, 0.08 mm (right antenna). No annulation visible on surface of antennal segments.
Right labial palpus visible in ventral view (Fig. 7C), consisting of at least 6 segments (additional, basal-most segment might be present). All segments (except for apical one) about 0.02 mm wide; apical segment about 0.01 mm wide (basally). Length of segments of right labial palpus from base to top: 0.05 mm, 0.09 mm, 0.03 mm, 0.08, mm, 0.08 mm; apical segment pointed apically, 0.05 mm long. No annulation visible on surface of palpomeres.
Zootaxa 3716 (2) © 2013 Magnolia Press · 243BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 6. Berothidae indet., larva C. Photograph of the larva, dorsal view. Scale bar is 0.5 mm.
FIGURE 7. Berothidae indet., larva C. A, photograph of the head, dorsal view. B, drawing of the head and thorax, dorsal view. C, labial palpus. D, right foreleg, ventral view. an, antenna; cs, coronal suture; cx, coxa; ds, prothoracic dorsal (pronotal) sclerites; em, empodium; fm, femur; fs, frontal suture; tb, tibia; tr, trochanter; ts, tarsus. Scale bars are 0.5 mm (B–D to scale).
Cervix probably strongly retracted into prothorax; in dorsal view visible only as lateral scale-like structure.Prothorax 0.52 mm long, 0.52 maximum width; with two large dorsal sclerites located anteriorly and in contact
with each other along midline. Mesothorax and metathorax both approximately 0.4 mm long and 0.6 mm wide, both with pair of small oval dorsal sclerites.
WEDMANN ET AL.244 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Legs short, robust; coxa very short; trochanter distinct, short; femur, tibia and tarsus relatively short, stout; pretarsus with two short claws, trumpet-shaped empodium.
Abdomen divided into 10 segments, terminal segments poorly visible. Segments 1 to 4 much shorter than wide, about 0.3–0.4 mm long and about 0.6–0.7 mm wide. Segments 5 to 7 about 0.5 mm long, and 0.7 mm wide. Segment 8 (and following two) gradually tapering towards end of abdomen (due to clouded preservation not measurable). Apex of abdomen obscured by cloudy, rounded vesicle, perhaps consisting of fluid leaking into resin.
Remarks. Judged from its size, this larva is a third instar (the second instar larvae of Berothidae are inactive and exhibit morphologically reduced legs and mouthparts).
This larva was schematically figured earlier (Janzen 2002: Fig. 58); the author erroneously assigned it to the family Chrysopidae.
Berothidae indet., larva D
Figs. 8–10
Material examined. Specimen Be 1297, deposited in the amber collection of SF; a third instar larva in a small piece of Baltic amber (precise collecting locality is unknown). The inclusion was bought from the collection of Marius Veta, Lithuania.
Description. Larva as preserved 3.1 mm long (from tip of cervix to abdomen); actual larval length probably about 6 mm as abdomen is distorted and posterior part with anal sucker is flapped over.
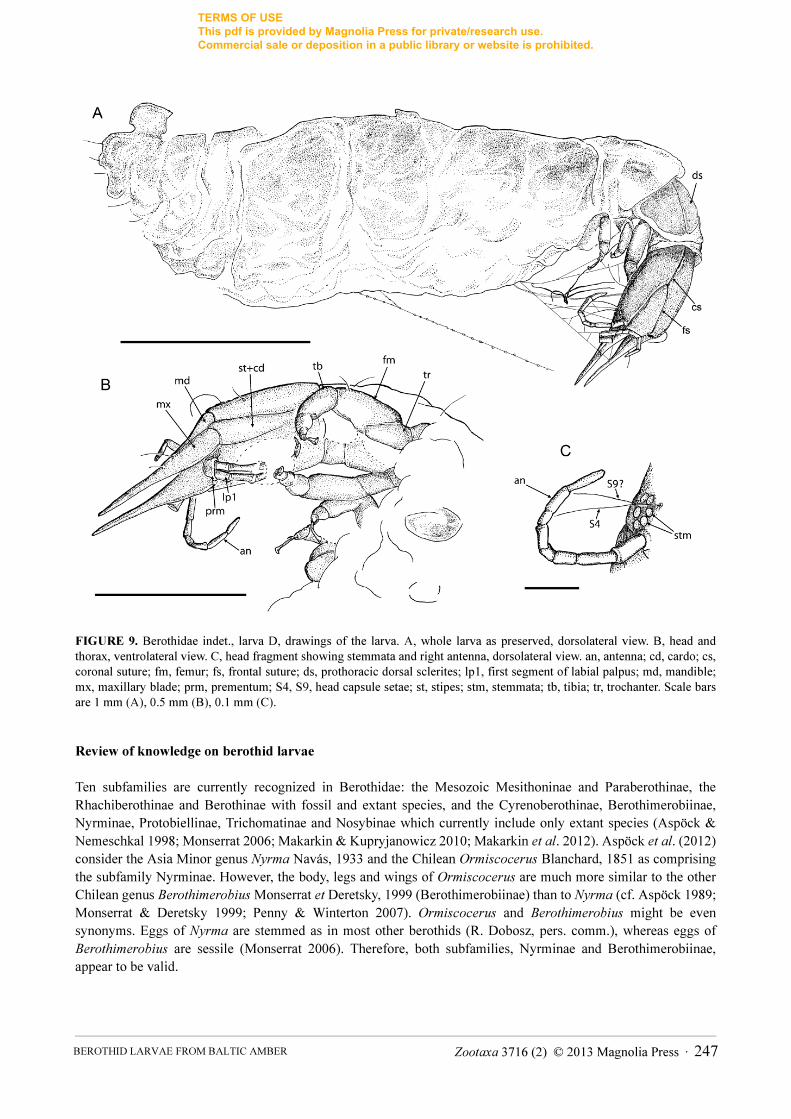
Head capsule elongated, 0.5 mm long, 0.3 mm wide (length/width ratio 1.67). Six stemmata on each side clearly visible; long setae (about 0.2 mm) ventral to stemmata (probably S9).
Dorsally, long seta (about 0.2 mm) near stemmata (S4); sockets of lateral clypeal (trichobothrian) setae on anterior part of capsule; S11 and S12 between arms of frontal suture, probably S1 just posterior to frontal suture, and some other (unidentified) setae in postero-lateral portion of head capsule clearly visible (Figs. 9C, 10A, B). Ecdysial cleavage lines very distinct; consisting of frontal and coronal sutures. Two arms of frontal suture extending for approximately half of head capsule length, convergent from inner margin of antennal tubercles until they touch. Lateral sutures absent. Coronal suture long. Ventrally, coxopodites of maxillae (stipes and cardo) rather clearly visible; stipes and cardo relatively narrow; boundary between them not distinct. Labium not clearly visible; prementum appears short (poorly preserved). Ventral setae not visible.
Mandibles 0.44 mm long, relatively broad. Maxillary blades very broad basally, gradually narrowed apically; closely associated with mandible to form mandibulomaxillary stilets, not strongly pointed apically.
Antennal sockets rather flat, without distinct antennal tubercle. Right antenna 0.4 mm long, 7-segmented; left antenna probably 6-segmented; no apical seta visible. Segment lengths of right antenna (from base to top): 0.06 mm, 0.08 mm, 0.04 mm, 0.03 mm, 0.04 mm, 0.08 mm, 0.06 mm. All segments 0.02 mm wide, except for basal segment which is 0.3 mm wide. Left antenna broken twice, but probably complete.
Labial palps poorly visible, about 0.3 mm long; probably 6-segmented; second segment longest and terminal segment very short.
Synchroton radiation computer tomography revealed remains of tissue inside of head (Fig. 10C). Oesophagus clearly visible, bifurcated towards mandibulomaxillary stilets.
Cervix probably mainly retracted into prothorax; visible length 0.02 mm dorsally, 0.1 mm laterally. Prothorax about 0.4 mm long (as preserved); with two large dorsal sclerites that touch each other at midline.
Mesothorax and metathorax crumpled, not measurable; at least mesothorax with indistinctly visible paired oval dorsal sclerites.
Legs rather short, robust. Coxa short; trochanter rather large; femur of right foreleg about 0.25 mm long, stout; tibia shorter than femur; tarsus short; pretarsus with two short claws and long, basally strongly narrowed trumpet-shaped empodium. Left midleg apparently missing; left hindleg stout (visible with Synchroton radiation computer tomography).
Synchroton radiation computer tomography revealed enigmatic, rounded structure on ventro-lateral side of thorax (dorsal of left hind coxa), ca. 0.15 mm wide, ca. 0.1 mm long; its surface appears very smooth.
Abdomen looks strongly wrinkled; its segmentation vaguely visible. Distal portion of abdomen curved forward; probably with anal sucker.
Zootaxa 3716 (2) © 2013 Magnolia Press · 245BEROTHID LARVAE FROM BALTIC AMBER
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
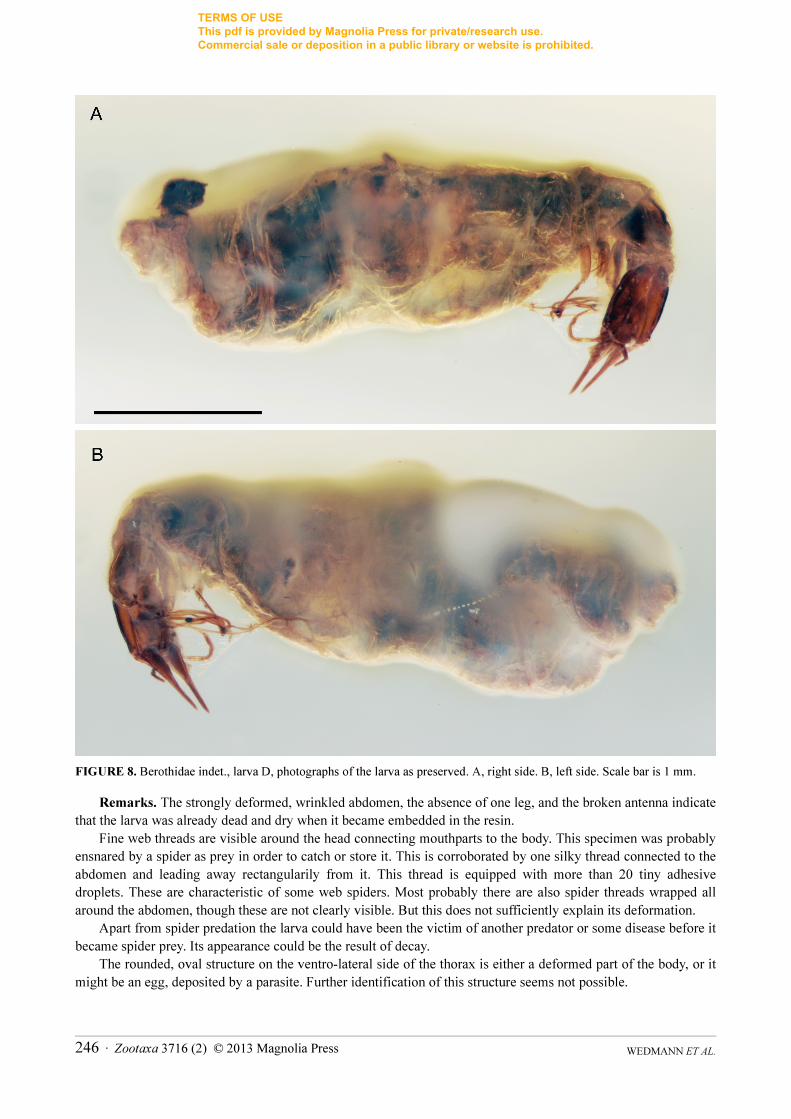
FIGURE 8. Berothidae indet., larva D, photographs of the larva as preserved. A, right side. B, left side. Scale bar is 1 mm.
Remarks. The strongly deformed, wrinkled abdomen, the absence of one leg, and the broken antenna indicate that the larva was already dead and dry when it became embedded in the resin.
Fine web threads are visible around the head connecting mouthparts to the body. This specimen was probably ensnared by a spider as prey in order to catch or store it. This is corroborated by one silky thread connected to the abdomen and leading away rectangularily from it. This thread is equipped with more than 20 tiny adhesive droplets. These are characteristic of some web spiders. Most probably there are also spider threads wrapped all around the abdomen, though these are not clearly visible. But this does not sufficiently explain its deformation.
Apart from spider predation the larva could have been the victim of another predator or some disease before it became spider prey. Its appearance could be the result of decay.
The rounded, oval structure on the ventro-lateral side of the thorax is either a deformed part of the body, or it might be an egg, deposited by a parasite. Further identification of this structure seems not possible.
WEDMANN ET AL.246 · Zootaxa 3716 (2) © 2013 Magnolia Press
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 9. Berothidae indet., larva D, drawings of the larva. A, whole larva as preserved, dorsolateral view. B, head and thorax, ventrolateral view. C, head fragment showing stemmata and right antenna, dorsolateral view. an, antenna; cd, cardo; cs, coronal suture; fm, femur; fs, frontal suture; ds, prothoracic dorsal sclerites; lp1, first segment of labial palpus; md, mandible; mx, maxillary blade; prm, prementum; S4, S9, head capsule setae; st, stipes; stm, stemmata; tb, tibia; tr, trochanter. Scale bars are 1 mm (A), 0.5 mm (B), 0.1 mm (C).
Review of knowledge on berothid larvae
Ten subfamilies are currently recognized in Berothidae: the Mesozoic Mesithoninae and Paraberothinae, the Rhachiberothinae and Berothinae with fossil and extant species, and the Cyrenoberothinae, Berothimerobiinae, Nyrminae, Protobiellinae, Trichomatinae and Nosybinae which currently include only extant species (Aspöck & Nemeschkal 1998; Monserrat 2006; Makarkin & Kupryjanowicz 2010; Makarkin et al. 2012). Aspöck et al. (2012) consider the Asia Minor genus Nyrma Navás, 1933 and the Chilean Ormiscocerus Blanchard, 1851 as comprising the subfamily Nyrminae. However, the body, legs and wings of Ormiscocerus are much more similar to the other Chilean genus Berothimerobius Monserrat et Deretsky, 1999 (Berothimerobiinae) than to Nyrma (cf. Aspöck 1989; Monserrat & Deretsky 1999; Penny & Winterton 2007). Ormiscocerus and Berothimerobius might be even synonyms. Eggs of Nyrma are stemmed as in most other berothids (R. Dobosz, pers. comm.), whereas eggs of Berothimerobius are sessile (Monserrat 2006). Therefore, both subfamilies, Nyrminae and Berothimerobiinae, appear to be valid.
Zootaxa 3716 (2) © 2013 Magnolia Press · 247BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 10. Berothidae indet., larva D, images reconstructed from synchrotron radiation computer tomography data (false color). A, B, head and prothorax. C, whole larva, partially cut open, showing esophageal tissue in the head and an artificial cavity in the body. ac, artificial cavity; et, esophageal tissue; lcs, lateral clypeal seta; S1,S4, S9, S11, S12, head setae. Scale bars are 0.2 mm (A, B), 1.0 mm (C).
WEDMANN ET AL.248 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 11. Berothidae indet., larva E. A, whole larva as preserved. B, anterior part of the larva (dorsal view). Scale bars are 1 mm (A, B).
The larvae of few species of the following subfamilies of extant berothids are described or illustrated: Rhachiberothinae, Berothimerobiinae, Nyrminae and Berothinae (Table 1). The larvae of the presumed cyrenoberothine Manselliberotha neuropterologorum Aspöck et Aspöck, 1988 are known but neither described nor illustrated (Aspöck & Aspöck 1988, p. 9). A larva from Australia not attributed to a taxon is figured by Riek (1970). Judged from its shape and from the relatively small head, it is probably fully-grown and does not belong to the Berothinae because of the position of lateral sutures, which are located distally to the frontal suture (see Riek 1970: Fig. 29.10G, H; New 1991: Fig. 34.10A, B). Such a position of the lateral sutures is similar to that found in Rhachiberothinae, and it is probably plesiomorphic compared with the condition found in Berothinae, in which the lateral sutures originate from the arms of the frontal suture. Three berothid subfamilies occur in Australia: Protobiellinae, Trichomatinae and Berothinae. The figured larva might belong to Protobiellinae, a sister group to the ‘higher’ Berothidae (i.e., Trichomatinae + Nosybinae + Berothinae) (Aspöck & Nemeschkal 1998) judged from its more ‘primitive’ character states (i.e., four-segmented antennae and palpi, position of the lateral sutures).
The most comprehensive and detailed studies on berothid larvae can be found in the unpublished thesis of Möller (2003), which is provided with numerous SEM photomicrographs and drawings of the African species Podallea vasseana, and the dissertation of MacLeod (1964), who examined the head capsule of the American species of Lomamyia Banks, 1905. The majority of other descriptions are based on the first instar larvae hatched from eggs in the cage (Tillyard 1916; Toschi 1964; Minter 1990; Monserrat 2006). Third instar larvae are known only for two genera of Berothinae (Podallea and Lomamyia) (Gurney 1947; Tauber & Tauber 1968; Möller 2003; Möller et al. 2006).
Hitherto, three fossil larvae of Berothidae have been described, all from the Cretaceous: from the Valanginian/Hauterivian of Jezzine (Jouar Es-Souss), Lebanese amber; from the earliest Cenomanian of Burmese amber (the age is after Shi et al. 2012); and from the Campanian of Grassy Lake, Canadian amber.
The fossil larva described from Lebanese amber is the oldest known berothid larva. Its family affinity as belonging to Berothidae is undoubted, judged from the structure of its mouthparts, ecdysial cleavage lines and antennae (see Whalley 1980: 161, Figs. 9–10). The larva is 6–8 mm long (i.e., probably a third instar), very hairy, and possesses a trumpet-shaped empodium; rounded head; five pairs of stemmata; four- or five-segmented labial
Zootaxa 3716 (2) © 2013 Magnolia Press · 249BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
palps; antennae with an apical seta (but the number of segments is not clear); and “a prominent epicranial suture” [=ecdysial cleavage lines] (Whalley 1980: p. 161). The structure of the latter character is very important; the photo (Whalley 1980: Fig. 10) shows that all three sutures of the ecdysial cleavage lines (frontal, coronal and lateral) are probably present. These structures of this larva are most similar to those of the larva of the extant Chilean genus Berothimerobius (although only a first instar larva of the latter species is known and the structure of the ecdysial cleavage lines is unknown, see Monserrat 2006).
The larva from the Burmese amber (about 4 mm long without mouthparts) is of rather typical berothid appearance (Engel & Grimaldi 2008: Figs. 42, 43). It possesses the following principal character states: apparently six small stemmata; palpi with eight palpomeres, with “apical most palpomere slightly broadened, with rounded apex” (however, Fig. 43 shows four palpomeres with short, acute terminal segment); antennae with 15 “articles” (but Fig. 43 shows the number of segments of antennae to be much less, 5–7 segments, with two terminal setae preserved in one antenna). The structure of the ecdysial cleavage lines is unknown.
The newly-hatched larva reported from Canadian amber as belonging to the family Chrysopidae (Engel & Grimaldi 2008: 30, Figs. 12–14) surely belongs to Berothidae. Its size (about 0.6 mm excluding mandibulomaxillary stylets, too small for chrysopid larvae) and the head morphology are typical of Berothidae: the mouthparts are straight, with maxillae which are broad basally and narrowed apically, not curved as in chrysopids; the antennae are 3-segmented, and bear a long strong terminal seta.
Main diagnostic traits of berothid larvae
A general account of the morphology of berothid larvae can be found in Tauber (1991). Here we consider some important features.
Ecdysial cleavage lines. In general, ecdysial cleavage lines on the larval head are shaped as in other insects, i.e., Y-like, consisting of two converging arms of the frontal suture anteriorly and the coronal suture posteromedially. The ecdysial cleavage lines are not included in some larval descriptions (e.g., Tillyard 1916: Figs 32, 33; Monserrat 2006: Fig. 11b), but are probably present in these species. However, in some taxa there is an additional suture running laterally from the frontal or coronal sutures. This suture is characteristic of larvae of only some Berothidae, although it may be vestigial in Psychopsidae (see MacLeod 1964: Fig. 66, very short unlabeled sutures running from the distal part of the frontal arms). We propose to name this suture as the lateral suture. In the genera of Berothinae (Lomamyia, Podallea) and Nyrminae (Nyrma) the lateral sutures originate from the arms of the frontal suture. Larvae A and B described above have a character condition as in these genera. In Rhachiberothinae (Mucroberotha) and the presumed protobielline larva (see above; New 1991: Fig. 34.10) the lateral sutures originate from the coronal suture.
Larvae C and D have no lateral suture at all, the general shape of the ecdysial cleavage is similar to that of Rhachiberothinae. The condition found in larvae C and D is probably plesiomorphic in the family.
Stemmata. The larvae of the basal groups of Berothidae have 5–6 pairs of stemmata. This condition is characteristic of Rhachiberothinae (6 in Mucroberotha), Berothimerobiinae (6 in Berothimerobius); all three Cretaceous larvae (5 or 6), and the Baltic amber larva D (6). The larvae of Berothinae (Lomamyia, Podallea) have at most two pairs of stemmata, or none. In the latter case, some component of the stemmata may be present below the cuticle (Minter 1990). Therefore, the presence of 5–6 pairs of stemmata is considered to be the plesiomorphic condition, and their reduction to 0–2 in Berothinae is considered to be an apomorphy.
Antennae. Three-segmented antennae are known in the larvae of the advanced subfamily Berothinae, i.e., in
Lomamyia, Podallea, Spermophorella (Tillyard 1916; Minter 1990; Möller et al. 2006; MacLeod 1964; Toschi 1964). Larvae A and B also have three-segmented antennae. Four-segmented antennae are present in more basal groups, i.e. in Mucroberotha, Nyrma Navás, Berothimerobius (Minter 1990; Monserrat 2006; Dobosz & Górski 2008), and the presumed prothobielline larva (see above) (New 1991: Fig. 34.10B). The latter have two very short additional segments or secondary annulations between the second and third segments. Even in berothine larvae, the third terminal segment sometimes appears to be clearly divided into two subsegments (e.g., Möller 2003: Figs. 18). All extant taxa whose larvae are known possess a terminal seta. The larvae C and D have 6 to 7 segments and lack a terminal seta (both specimens have asymmetrical antennae, six segments in one antenna, seven in the other). The antennae of the larva from Burmese amber also have numerous segments, 5–7 or 15 (see above). Altogether, the condition found in larvae C and D is probably plesiomorphic.
WEDMANN ET AL.250 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Labial palps. All extant larvae of genera whose larvae are known have four-segmented labial palps, i.e., Mucroberotha, Nyrma, Berothimerobius, Lomamyia, Podallea, Spermophorella, and the presumed prothobielline larva (New 1991: Fig. 34.10B). Larvae A and B share this character condition of the extant genera. The labial palps in the three Cretaceous larvae are presumably three- to five-segmented, but in larvae C and D these are clearly six-segmented. The latter condition is probably plesiomorphic.
Paired dorsal thoracic sclerites. All segments of the berothid larval thorax possess paired dorsal sclerites: larger ones are present on the prothorax, and smaller ones on the meso- and metathorax. These sclerites are neither described nor shown in the figures of the first instar larvae of Mucroberotha, Berothimerobius and Spermophorella
(Tillyard 1916: Fig. 32; Minter 1990: Fig. 5; Monserrat 2006: Fig.11a). The structure of the prothoracic (pronotal) sclerites is especially important. In Lomamyia, they are relatively small and widely spaced in the third larval instar (Gurney 1947: Fig. 5); generally, prothoracic sclerites are larger in the third instar than in the first instar (Gurney 1947); therefore, in the first instar these sclerites should be even smaller. Podallea possesses large prothoracic sclerites; they occupy most of the area of the pronotum, but are clearly separated along the midline (Möller 2003: Figs. 16, 32; Möller et al. 2006: Figs. 7, 31). The prothoracic sclerites of an unattributed species from Australia (the presumed prothobielline larva) are relatively small but closely spaced (New 1991: Fig. 34.10A). The structure of the prothoracic sclerites of larvae A and B is most similar to that of Podallea. The prothoracic sclerites of larvae C and D are very big and occupy most of the dorsal side of the prothorax, touching each other along the midline. Their structure is similar to that found in Osmylidae (Withycombe 1925; New 1991: Figs. 34.10E, F), and is probably plesiomorphic.
Berothidae in Baltic amber and the taxonomic affinities of the described larvae
Until now, two species of adult Berothidae have been reported from Baltic amber, i.e., Proberotha prisca Krüger, 1923 and Whalfera wiszniewskii Makarkin & Kupryjanowicz, 2010.
Proberotha prisca was described from a single specimen, which was not illustrated and now apparently lost. No other specimens have been reported subsequently. However, the original description shows that this species probably belongs to the Nevrorthidae. In particular, the strongly pectinate CuP (with nine short branches in P.
prisca), and the presence of two gradate series of crossveins in the radial to medial spaces are characteristic of Nevrorthidae, not of Berothidae. The features of several specimens of one undescribed species of Nevrorthidae are in general consistent with those of P. prisca (see e.g., Ross 1998: Fig. 133; Scheven 2004: Fig. on p. 74, left).
Therefore, the only genus of Berothidae recorded with certainty from Baltic amber is Whalfera Engel, 2004. This genus belongs with certainty to the Rhachiberothinae, and it is apparently most closely related to the extant African genus Mucroberotha Tjeder (Makarkin & Kupryjanowicz 2010).
Four other adult berothid specimens known from Baltic amber are not described. Three of these belong probably to one species or a few closely related species (see Bachofen-Echt 1949: Fig. 122; Weitschat & Wichard 1998: Figs. 55a,b; Scheven 2004: Fig. on p. 7). Their venation most closely resembles that of the extant African berothine genus Lekrugeria Navás (U. Aspöck in Scheven 2004, p. 6), but the scapus appears to be much shorter and the long hairs on the body and antennae characteristic of Lekrugeria are not visible in the photographs. A fourth specimen from the MCZ collection is not illustrated; it is only known that this is a female which “has long hypocaudae and is obviously a berothine” (MacLeod & Adams 1968, p. 258). This specimen is not found in the MCZ (P. Perkins, pers. comm. to VM, 2007). Theoretically, it could be identical with one of the above mentioned species because Lekrugeria possesses long hypocaudae (Aspöck & Aspöck 1985). Therefore, all four known adult specimens from Baltic amber could belong to few closely related species of a berothinae genus similar to Lekrugeria.
Larvae A and B belong with high certainty to the Berothinae based on all characters available (e.g., the configuration of the ecdysial cleavage lines; the structure of the thoracic sclerites; three-segmented antennae terminating in a seta; four-segmented labial palps; an ocular area without distinctly visible stemmata). However, the larvae of the subfamilies Trichomatinae and Nosybinae are as yet unknown, and so some uncertainty remains.
Larvae A and B most probably not are conspecific, but very similar. Apart from the difference in the presence/absence of the visible caudal sucker (see description), larva B has slightly longer antennae; its lateral sutures are inclined toward the anterior part of the frontal sutures at a clearly obtuse angle (not at a right angle); the legs are
Zootaxa 3716 (2) © 2013 Magnolia Press · 251BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
more slender. If these differences are not intraspecific, the two larvae might belong to closely related species of one berothine genus.
Larvae C and D possess some unique character conditions among Berothidae (e.g., antennae and labial palps are six- or seven-segmented; ecdysial cleavage lines consist of only frontal and coronal sutures (the lateral suture is absent); pronotal sclerites are large and in contact with each other along the midline). Nevertheless, we place these two larvae in Berothidae. There are six neuropteran families whose larvae have straight jaws (mandibulomaxillary stilets), and all are recorded from Baltic amber: Osmylidae, Sisyridae, Coniopterygidae, Berothidae (including Rhachiberothidae), Mantispidae and Dilaridae (Aspöck 1992). Affinities of larvae C and D to the families Sisyridae, Coniopterygidae, Mantispidae and Dilaridae may be rejected by various reasons: sisyrid larvae lack labial palps; coniopterygid larvae are very small and have a strongly different appearance; mantispid larvae at third instar are scarabaeiform. Dilarid larvae lack thoracic dorsal sclerites and visible ecdysial cleavage lines; the head is rounded; the antennae are specialized (e.g., Gurney 1947; Ghilarov 1962; Monserrat 1988). In general, they are dissimilar to our larvae C and D. Osmylid larvae resemble larvae C and D in many aspects, e.g., in possessing large pronotal sclerites that touch each other along the midline; long, slender antennae and labial palps; five to six pairs of stemmata; basally broadened maxillae (see Lestage 1921: Fig. 1; Withycombe 1925; Kawashima 1957: Pl. 2, Figs. A–C; New 1991: Figs 34.5A, 34.10E, F). However, the head of osmylid larvae is short, rounded and the mandibulomaxillary stilets are at least twice longer than the head capsule length (the head capsule of larvae C and D is elongate, and the mandibulomaxillary stilets are shorter than the head). Moreover, an osmylid larva known from Baltic amber does not differ from extant osmylid larvae in this respect (see Wichard et al. 2009: Fig. 07.04). Therefore, Berothidae is the most probable family affinity of larvae C and D. They share many features of extant berothid larvae: similar general appearance; very similar structure of head capsule, mandibulomaxillary stilets and legs; the presence of distinct ecdysial cleavage lines; general similarity in the structure of antennae and labial palps (long; slender; without specialized segments); the presence of large pronotal sclerites, and smaller ones on the meso- and metathorax.
Although larvae C and D are very similar to each other in many external characters, the ecdysial cleavage lines on the head have different proportions. Another similar larva deposited in the private collection of Thomas Schäfer (Saland, Switzerland; No. 01.03.13) is known to us only by its photograph (Larva E) (Fig. 11). This is also a complete mature larva (approximately 7.5 mm long), mostly obscured by a milky covering. Larva E is similar to larvae C and D; it shares with the latter the same structure of the head, antennae (probably seven-segmented) and pronotal sclerites. These three larvae may well belong to closely related species of the same genus; perhaps they are even conspecific.
The taxonomic affinity of larvae C and D within Berothidae is unclear. They belong to some basal subfamily, but not to Rhachiberothinae, Berothimerobiinae or Nyrminae, the larvae of which are known. Unfortunately, the larvae of the basal subfamily Cyrenoberothinae are unknown. There are also some Cretaceous taxa (including Mesithoninae and Paraberothinae) with which these larvae may be theoretically associated, but these taxa have not been recorded from Baltic amber.
Berothid biology and larval termitophily
Very little is known about the biology and life habits of larval and adult Berothidae. Adults readily come to light, and are mostly collected this way, but little is known of their ecology; for the immature stages even less is known (Aspöck 1986; New 1986).
A complete berothid life cycle was first described for Lomamyia latipennis Carpenter, 1940, by Tauber & Tauber (1968). The first and third instars of this species are active predators, but the second instar is not feeding, inactive and has reduced legs and mouthparts. It was demonstrated that Lomamyia larvae live in the subterranean galleries of Reticulitermes species (Rhinotermitidae) and prey on these termites. First and third instars of Lomamyia latipennis use a chemical substance (allomone) issued from their abdominal tip to immobilize and kill the termites (Johnson & Hagen 1981). Third instars of Lomamyia hamata were reported to insert their mouthparts into termites of the species Reticulitermes flavipes (Kollar, 1837) and inject a neurotoxin, which first immobilizes and then kills the prey (Brushwein 1987b).
The larval termitophily (that is, inhabiting termite nests and preying on termites) is a very interesting feature of the biology of Berothidae, but it is still disputed (New 1986; Aspöck et al. 2010). Termitophily was clearly
WEDMANN ET AL.252 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
documented for the American berothine genus Lomamyia (Johnson & Hagen 1981; Brushwein 1987a,b; Faulkner 1992), and is assumed for the African genus Podallea, also from Berothinae (Möller 2003, Möller et al. 2006). The biology of berothids from the other subfamilies is entirely unknown.
Direct field observations of berothid eggs, larvae and cocoons are rare, and restricted almost exclusively to Lomamyia. A cluster of eight eggs of this genus was found on an oak log (Quercus sp.) infested with Reticulitermes
hesperus Banks in Banks et Snyder, 1920 in Solano County, California (Johnson & Hagen 1981). Brushwein (1987b) observed many eggs of Lomamyia (mainly L. hamata) in the Clemson area, South Carolina, in a pine forest containing active colonies of Reticulitermes flavipes on several different substrates: dead, standing Quercus
trees; partially decomposed oak logs; decomposing pine logs; and a weathered wooden-slat fence. In South Africa eggs of Podallea have been found “in association with wood-eating termites at Mogol in Limpopo Province (M. Mansell, pers. comm.)” (Möller 2003, p. 117).
Thomas Snyder was probably the first who mentioned berothid larvae to be associated with termites: “A peculiar Neuropterous larva was found in a colony of [Reticulitermes] flavipes in a decaying branch lying on the ground near a stream at Falls Church, Virginia, on July 18. This was a fully developed first form larva of Mantispa, according to Dr. A. Böving, and is probably predaceous.” (Snyder 1920, p. 120). Berothid and dilarid larvae were not known in that time, and it is quite understandable that this larva was identified as a mantispid. Gurney (1947, p. 154) reported two Lomamyia larvae, one was found by H. G. Hubbard in 1895 in Washington, D.C., “in the fungus garden (nest) of [ant] Atta occidentalis in wooded knoll”, and another, 9.5 mm long, by R. J. Kowal in 1941 in Beltsville, Maryland, “in fallen log, in pocket containing living and dead termites”. Faulkner (1992) noted that one first instar larva of Lomamyia fulva Carpenter, 1940 “was obtained from a berlese sample in association with Reticulitermes termites.” (p. 23), and that larvae of Lomamyia occidentalis “have been obtained from wood infested with the termite, Zootermopsis angusticollis (Hagen, 1858), and have been reared to adults by Miller (pers. comm.).” (p. 34). The larvae of the Australian berothine genus Spermophorella Tillyard, 1916 occur under bark of Eucalyptus (Riek 1970).
Brushwein (1987b) found 30 cocoons of Lomamyia hamata in the pine forest in Clemson area. Most of the cocoons “were taken from pine logs and branches that also contained active Reticulitermes flavipes colonies”. (p. 678). Brushwein (1987a) reported two cocoons of Lomamyia longicollis (Walker, 1853) collected in the same pine forest in Clemson area, one from a log which “reached the red-rotten stage of decay and contained an active colony of eastern subterranean termites, Reticulitermes flavipes” (p. 151), and another from “the underside of a piece of pine bark which rested on the surface of the duff” (p. 152).
The information on prey of Berothidae obtained under laboratory conditions is summarized below. The first instar larvae of Lomamyia latipennis were offered “small living arthropods from soil samples as well as ant and scolytid larvae”, but all larvae died within ten days (Toschi 1964, p. 25). Later, the same berothid species was successfully reared when the larvae fed on the damp-wood termites Zootermopsis angusticollis (Tauber & Tauber 1968). Larvae of L. latipennis (probably, however, of a new species: Faulkner 1992, p. 29) fed on Zootermopsis
angusticollis, Z. nevadensis (Hagen, 1874) and the western subterranean termite Reticulitermes hesperus, but only those larvae that fed on R. hesperus successfully completed their development (Johnson & Hagen 1981). Lomamyia hamata was successfully reared feeding on the eastern subterranean termite Reticulitermes flavipes
(Brushwein 1987b). Larvae of several Lomamyia species have been reared: L. banksi Carpenter, 1940 on “termites (Reticulitermes flavipes) by Brushwein (pers. comm.)” (p. 16); L. flavicornis (Walker 1953) “on immature termites (Reticulitermes Holmgren, 1913) by Brushwein (pers. comm.)” (p. 19); L. hamata on the damp-wood termites of Neotermes Holmgren, 1911 [Kalotermitidae] (Faulkner 1992).
Larvae of Podallea manselli and P. vasseana fed on two species of soil-dwelling, cellulose-feeding termites from the genera Cubitermes Wasmann, 1906 and Apicotermes Holmgren, 1912 (Termitidae). Some larvae reached the third instar, and two larvae pupated and adults emerged. First instar larvae of Podallea manselli rejected dry-wood termites of the genus Cryptotermes Banks, 1906 (Kalotermitidae) and died over a period of 3–17 days (Möller 2003).
In summary, extant berothine larvae apparently are very specialized predators, they survive only when they feed on specific groups of termites. Species of two berothine genera which have successfully been reared, are associated with subterranean or damp-wood termites: the American genus Lomamyia feeds primarily on Reticulitermes (Rhinotermitidae), the most common subterranean termite genus in the USA (Su et al. 2001), and the African genus Podallea feeds on soil-feeding Termitidae.
Zootaxa 3716 (2) © 2013 Magnolia Press · 253BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Possible evolution of termitophily in berothid larvae
It is unclear when termitophily in berothid larvae appeared. A summary of all available data, however, can provide a more or less probable scenario.
One morphological character of the larvae which might be associated with their subterranean (and theoretically termitophilous) habit is the reduction of stemmata. This reduction can be detected in fossil specimens, and this feature might help to interpret the life styles of the different groups of Berothidae.
In general, termites appear later in the fossil record than berothids. The first termite fossils are from the Early Cretaceous; they appear to have risen in the Late Jurassic (Engel et al. 2009). In contrast, berothids are known from as early as the Middle Jurassic (Makarkin et al. 2011) and are assumed to be much older (Winterton et al. 2010). Therefore, the first (Triassic/Jurassic) berothids could not be associated with termites. This association might have evolved during the Cretaceous (maybe in the early Cenozoic, or even later), and only in Berothinae (or in subfamilies closely related to this group). The two extant termite families with which berothine larvae are associated, that is, Termitidae and Rhinotermitidae, are relatively young. The oldest known Rhinotermitidae are from the early Cenomanian Burmese amber (Krishna & Grimaldi 2003), where a diverse berothid assemblage is also known (Engel & Grimaldi 2008). The oldest known Termitidae are from the Eocene (Krishna & Grimaldi 2003). However, all known Cretaceous larvae (including those from Burmese amber) have an unreduced number of stemmata. Therefore, it may be tentatively concluded that these larvae were not termitophilous.
The berothid larvae C, D and E from the Baltic amber are mature, third-instar larvae. We assume that these larvae were presumably not termitophilous, as is indicated by the unreduced number of stemmata. Additionally, the probability to be caught in a spider net is much higher in corticolous larvae (the larva D) than in termitophilous ones. In extant termitophilous berothine taxa, however, the mature larva leave the termite nest to pupate, and therefore termitophily cannot be completely excluded.
The mandibulomaxillary stilets of these larvae are distinctly not so acutely pointed as those of the larvae A and B. This may also indicate a different habit of feeding or different prey. Probably the larvae C to E fed on larger prey with a stronger cuticula. The very acutely pointed mouthparts in the Berothinae larvae A and B are more specialized and presumably only useful for soft-bodied prey, like termites. Therefore, the Late Eocene Baltic amber berothid assemblage apparently includes both termitophilous and not termitophilous larvae.
Acknowledgements
We thank Dr. Roland Dobosz (Upper Silesian Museum, Bytom, Poland) for providing the poster on the biology and morphology of Nyrma kervilea and valuable information; Christel and Hans Werner Hoffeins (Hamburg) for the extended loan of fossil Neuroptera specimens; Thomas Schäfer (Saland, Switzerland) for information on the berothid larva from his collection, Frank Wieland (University of Hamburg) for his help with making photographs, and an anonymous reviewer for providing valuable comments that helped to greatly improve the manuscript. The research of TH was supported by DFG Heisenberg grant HO 2306/6-1.
References
Archibald, S.B. & Makarkin, V.N. (2004) A new genus of minute Berothidae (Neuroptera) from Early Eocene amber of British Columbia, Canada. Canadian Entomologist, 136, 61–76. http://dx.doi.org/10.4039/n03-043
Aspöck, U. (1986) The present state of knowledge of the family Berothidae (Neuropteroidea: Planipennia). In: Gepp, J., Aspöck, H. & Hölzel, H. (Eds.), Recent Research in Neuropterology. Proceedings of the 2nd International Symposium on Neuropterology. Privately printed, Graz, Austria, pp. 87–101.
Aspöck, U. (1989) Nyrma kervillea Navás – eine Berothide! (Neuropteroidea: Planipennia). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 41, 19–24.
Aspöck, U. (1992) Crucial points in the phylogeny of the Neuroptera (Insecta). In: Canard, M., Aspöck, H. & Mansell, M.W. (Eds.), Current Research in Neuropterology. Proceedings of the Fourth International Symposium on Neuropterology. Privately printed, Toulouse, pp. 63–73.
Aspöck, U. & Aspöck, H. (1985) Das Genus Lekrugeria Navás (Neuropteroidea: Planipennia: Berothidae: Berothinae). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 37, 85–98.
WEDMANN ET AL.254 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Aspöck, U. & Aspöck, H. (1988) Die Subfamilie Cyrenoberothinae – ein Gondwana-Element? Manselliberotha neuropterologorum n.g. et n. sp. aus S.W.A./Namibia (Neuropteroidea: Neuroptera: Berothidae). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 40, 1–13.
Aspöck, U., Haring, E. & Aspöck, H. (2012) The phylogeny of the Neuropterida: long lasting and current controversies and challenges (Insecta: Endopterygota). Arthropod Systematics & Phylogeny, 70 (2), 119–129.
Aspöck, U. & Nemeschkal, H.L. (1998) A cladistic analysis of the Berothidae (Neuroptera). Acta Zoologica Fennica, 209, 45–63.
Aspöck, U., Randolf, S. & Aspöck, H. (2010) The Berothidae of Madagascar (Neuropterida: Neuroptera) and Madagascar’s biological exploration by the Viennese adventuress Ida Pfeiffer (1797–1858). In: Devetak, D., Lipovšek, S. & Arnett, A.E.(Eds.), Proceedings of the Tenth International Symposium on Neuropterology. Piran, Slovenia, 2008. Maribor, Slovenia, pp. 75–82.
Bachofen-Echt, A. (1949) Der Bernstein und seine Einschlüsse. Springer Verlag, Wien, 204 pp. Baker, C.F. (1905) Notes on neuropteroid insects of the Pacific Coast of North America with descriptions of new species by
Nathan Banks. Invertebrata Pacifica, 1, 85–91. Banks, N. (1905) A revision of the Nearctic Hemerobiidae. Transactions of the American Entomological Society, 32, 21–51.Banks, N. (1906) Two new termites. Entomology News, 17, 336–337.Beutel, R.G., Friedrich, F. & Aspöck, U. (2010) The larval head of Nevrorthidae and the phylogeny of Neuroptera (Insecta).
Zoological Journal of Linnean Society, 158, 533–562. http://dx.doi.org/10.1111/j.1096-3642.2009.00560.x
Blanchard, C.É. (1851) Mirmeleonianos. In: Gay, C. (Ed.), Historia fisica y politica de Chile. Zoologia, Vol. 6. Paris, pp. 119–129.
Brushwein, J.R. (1987a) Observations on Lomamyia longicollis (Neuroptera: Berothidae) in South Carolina. Journal of the Kansas Entomological Society, 60, 150–152.
Brushwein, J.R. (1987b) Bionomics of Lomamyia hamata (Neuroptera: Berothidae). Annals of the Entomological Society of America, 80, 671–679.
Carpenter, F.M. (1940) A revision of the Nearctic Hemerobiidae, Berothidae, Sisyridae, Polystoechotidae and Dilaridae (Neuroptera). Proceedings of the American Academy of Arts and Sciences, 74, 193–280.
Dobosz, R. & Górski, G. (2008) New data on Nyrma kervilea (Neuroptera: Berothidae). In: Devetak, D. & Klenovšek, T. (Eds.), Tenth International Symposium on Neuropterology, 22–25 June 2008, Piran, Slovenia. Abstract book. Faculty of Natural Sciences and Mathematics, Maribor, pp. 33.
Engel, M.S. (2004) Thorny lacewings (Neuroptera: Rhachiberothidae) in Cretaceous Amber from Myanmar. Journal of Systematic Palaeontology, 2, 137–140. http://dx.doi.org/10.1017/s1477201904001208
Engel, M.S. & Grimaldi, D.A. (2008) Diverse Neuropterida in Cretaceous amber, with particular reference to the paleofauna of Myanmar (Insecta). Nova Supplementa Entomologica, 20, 1–86.
Engel, M.S., Grimaldi, D.A. & Krishna, K. (2009) Termites (Isoptera): their phylogeny, classification, and rise to ecological dominance. American Museum novitates, 3650, 1–27. http://dx.doi.org/10.1206/651.1
Faulkner, D.K. (1992) A revision of the genus Lomamyia Banks (Planipennia: Berothidae) with an emphasis on the western United States species. Unpublished Master thesis. California State University, Long Beach, CA, USA, xii + 119 pp.
Gepp, J. (1984) Erforschungsstand der Neuropteren-Larven der Erde (mit einem Schlüssel zur larvaldiagnose der Familien, einer Übersicht von 340 beschreibenen Larven und 600 Literaturzitaten). In: Gepp, J., Aspöck, H. & Hölzel, H. (Eds.), Progress in World's Neuropterology. Proceedings of the 1st International Symposium on Neuropterology. Graz, Austria, pp. 183–239.
Ghilarov, M.S. (1962) The larva of Dilar turcicus Hag. and the position of the family Dilaridae in the order Planipennia. Entomologicheskoe Obozrenie, 41, 402–416. [In Russian; English translation: Entomological Review, 41, 244–253]
Grimaldi, D.A. (2000) A diverse fauna of Neuropterodea in amber from the Cretaceous of New Jersey. In: Grimaldi, D.A. (Ed.), Studies on Fossil in Amber, with Particular Reference to the Cretaceous of New Jersey. Backhuys Publishers, Leiden, pp. 259–303.
Gurney, A.B. (1947) Notes on Dilaridae and Berothidae, with special reference to the immature stages of the Nearctic genera (Neuroptera). Psyche, 54, 145–169. http://dx.doi.org/10.1155/1947/78317
Hagen, H.A. (1858) Catalogue of the specimens of neuropterous insects in the collection of the British Museum. Part 1. Termitina. Printed by order of the Trustees, London, 34 pp.
Hagen, H.A. (1874) Report on the pseudo-Neuroptera and Neuroptera collected by Lieut. W. L. Carpenter in 1873 in Colorado. Annual Report of the United States Geological and Geographical Survey of the Territories, 7 (for 1873), 571–606.
Handlirsch, A. (1906–1908). Die fossilen Insekten und die Phylogenie der rezenten Formen. Ein Handbuch für Palaeontologen und Zoologen. W. Engelmann, Leipzig, ix + 1430 pp. [Issued in 1906 (pp. 1–640); 1907 (pp. 641–1140); 1908 (pp. 1120–1430)]
Hoffeins, C. & Hoffeins, H.W. (2004) Untersuchungen über die Häufigkeit von Inklusen in Baltischem und Bitterfelder Bernstein (Tertiär, Eozän) aus unselektierten Aufsammlungen unter besonderer Berucksichtigung der Ordnung Diptera. Studia dipterologica, 10 (2) (for 2003), 381–392.
Zootaxa 3716 (2) © 2013 Magnolia Press · 255BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Hoffman, K.M. & Brushwein, J.R. (1992) Descriptions of the larvae and pupae of some North American Mantispinae (Neuroptera: Mantispidae) and development of a system of larval chaetotaxy for Neuroptera. Transactions of the American Entomological Society, 118, 159–196.
Holmgren, N. (1911) Bemerkungen über einige Termiten-Arten. Zoologischer Anzeiger, 37, 545–553. Holmgren, N. (1912) Termitenstudien: 3. Systematik der Termiten. Die Familie Metatermitidae. Kungliga Svenska
Vetenskapsakademiens Handlingar, 48 (4), 1–166. Holmgren, N. (1913) Termitenstudien: 4. Versuch einer systematischen Monographie der Termiten der Orientalischen Region.
Kungliga Svenska Vetenskapsakademiens Handlingar, 50 (2), 1–276.Janzen, J.W. (2002) Arthropods in Baltic amber. Ampyx Verlag, Halle, 167 pp. Johnson, J.B. & Hagen, K.S. (1981) A neuropterous larva uses an allomone to attack termites. Nature (London), 289, 506–507.
http://dx.doi.org/10.1038/289506a0 Kawashima, K. (1957) Bionomics and earlier stages of some Japanese Neuroptera (I) Spilosmylus flavicornis (MacLachlan)
(Osmylidae). Mushi, 30, 67–70. Klimaszewski, J. & Kevan, D.K.McE. (1986) A new lacewing-fly (Neuroptera: Planipennia) from Canadian Cretaceous
Amber, with an analysis of its fore wing characters. Entomological News, 97, 124–132. Kollar, V. (1837) Naturgeschichte der schädlichen Insekten. Verhandlungen der kaiserlich-königlichen Landwirthschafts-
Gesellschaft in Wien, 5, 411–413.Krishna, K. & Grimaldi, D. (2003) The first Cretaceous Rhinotermitidae (Isoptera): a new species, genus, and subfamily in
Burmese amber. American Museum Novitates, 3390, 1–10. http://dx.doi.org/10.1206/0003-0082(2003)390<0001:tfcria>2.0.co;2
Krishna, K. & Grimaldi, D. (2009) Diverse Rhinotermitidae and Termitidae (Insecta: Isoptera) in Dominican amber. American Museum Novitates, 3640, 1–48.
Krüger, L. (1923) Neuroptera succinica baltica. Die im baltischen Bernstein eingeschlossenen Neuroptera des Westpreussischen Provinzial-Museums (heute Museum für Naturkunde und Vorgeschichte) in Danzig. Stettiner Entomologische Zeitung, 84, 68–92.
Lestage, J.A. (1921) La ponte et la larvule de l'Osmylus chrysops L. (Planipenne). Annales de Biologie Lacustre, Brussels, 10, 226–230.
Linnaeus, C. (1758) Systema naturae per regna tria naturae secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, 10th ed., Vol. 1. Salvii, Holmiae, 824 pp.
MacLeod, E.G. (1964) A comparative morphological study of the head capsule and cervix of larval Neuroptera (Insecta). Unpublished Ph.D. dissertation. Harvard University, Cambridge, MA, USA, [iii] + 528 pp.
MacLeod, E.G. & Adams, P.A. (1968) A review of the taxonomy and morphology of the Berothidae, with the description of a new subfamily from Chile (Neuroptera). Psyche, 74 (for 1967), 237–265.
Makarkin, V.N. (1994) Upper Cretaceous Neuroptera from Russia and Kazakhstan. Annales de la Societe Entomologique de France (N.S.), 30, 283–292. http://dx.doi.org/10.1080/00379271.2010.10697666
Makarkin, V.N. & Kupryjanowicz, J. (2010) A new mantispid-like species of Rhachiberothinae from Baltic amber (Neuroptera, Berothidae), with a critical review of the fossil record of the subfamily. Acta Geologica Sinica, 84, 655–664. http://dx.doi.org/10.1111/j.1755-6724.2010.00238.x
Makarkin, V.N., Ren D. & Yang Q. (2011) Two new species of Sinosmylites Hong (Neuroptera: Berothidae) from the Middle Jurassic of China, with notes on Mesoberothidae. ZooKeys, 130, 199–215. http://dx.doi.org/10.3897/zookeys.130.1418
Makarkin V.N., Yang Q., Peng, Y.Y. & Ren D. (2012) A comparative overview of the neuropteran assemblage of the Early Cretaceous Yixian Formation (China), with description of a new genus of Psychopsidae (Insecta: Neuroptera). Cretaceous Research, 35, 57–68. http://dx.doi.org/10.1016/j.cretres.2011.11.013
Martins-Neto, R.G. & Vulcano, M.A. (1990) Neurópteros (Insecta: Planipennia) da Formação Santana (Cretáceo Inferior), Bacia do Araripe, nordeste do Brasil. III. Superfamília Mantispoidea. Revista Brasileira de Entomologia, 34, 619–625.
McKellar, R.C. & Engel, M.S. (2009) A new thorny lacewing (Neuroptera: Rhachiberothidae) from Canadian Cretaceous amber. Journal of the Kansas Entomological Society, 82, 114–121. http://dx.doi.org/10.2317/jkes811.10.1
Minter, L.R. (1990) A comparison of the eggs and first-instar larvae of Mucroberotha vesicaria Tjeder with those of other species in the families Berothidae and Mantispidae (Insecta: Neuroptera). In: Mansell, M.W. & Aspöck, H. (Eds.), Advances in Neuropterology. Proceedings of the Third International Symposium on Neuropterology. South African Department of Agricultural Development, Pretoria, pp. 115–129.
Möller, A. (2003) Aspects of the larval morphology and biology of South African Podallea species (Neuropterida: Neuroptera: Berothidae). Master of Science Thesis. School of Molecular and Life Sciences in the Faculty of Sciences, Health and Agriculture, University of the North, South Africa, 123 pp.
Möller, A., Minter, L.R. & Olivier, P.A.S. (2006) Larval morphology of Podallea vasseana Navás and Podallea manselli Aspöck & Aspöck from South Africa (Neuroptera: Berothidae). African Entomology, 14, 1–12.
Monserrat, V.J. (1988) Revisión de los diláridos ibéricos (Neuropteroidea, Planipennia: Dilaridae). Eos, 64, 175–205.
WEDMANN ET AL.256 · Zootaxa 3716 (2) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Monserrat, V.J. (2006) Nuevos datos sobre algunas especies de la familia Berothidae (Insecta: Neuroptera). Heteropterus Revista de Entomología, 6, 173–207. http://dx.doi.org/10.3989/graellsia.2008.v64.i2.34
Monserrat, V.J. & Deretsky, Z. (1999) New faunistical, taxonomic and systematic data on brown lacewings (Neuroptera: Hemerobiidae). Journal of Neuropterology, 2, 45–66.
Navás, L. (1910) Hemeróbidos (Ins. Neur.) nuevos con la clave de las tribus y géneros de la familia. Brotéria (Zoológica), 9, 69–90.
Navás, L. (1933) De las cazas del Sr. Gadeau de Kerville en el Asia Menor. In: Ve Congrès International d'Entomologie, Paris, 18-24 Juillet 1932. Vol. 2. Travaux, pp. 221–225.
Nel, A., Perrichot, V., Azar, D. & Néraudeau, D. (2005a) New Rhachiberothidae (Insecta: Neuroptera) in Early Cretaceous and Early Eocene ambers from France and Lebanon. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 235, 51–85.
Nel, A., Perrichot, V., Azar, D. & Néraudeau, D (2005b) A replacement name for the neuropteran genus Eorhachiberotha Nel et al., 2005 (Neuroptera: Rhachiberothidae). Bulletin de la Société Entomologique de France, 110, 128.
New, T.R. (1986) A review of the biology of Neuroptera Planipennia. Neuroptera International (Supplemental Series), 1, 1–57.New, T.R. (1991) Neuroptera (lacewings). In: Naumann, I.D. (Ed.), The Insects of Australia, 2nd ed., Vol. 1. Melbourne
University Press, Melbourne, pp. 525–542.Penny, N.D. & Winterton, S. (2007) Rediscovery of the unusual genus Ormiscocerus (Neuroptera: Berothidae:
Cyrenoberothinae). Proceedings of the California Academy of Sciences, 4, 58, 1–6.Perkovsky, E.E., Rasnitsyn, A.P., Vlaskin, A.P. & Taraschuk, M.V. (2007) A comparative analysis of the Baltic and Rovno
amber arthropod faunas: representative samples. African Invertebrates, 48, 229–245. Petrulevicius, J.F., Azar, D. & Nel, A. (2010) A new thorny lacewing (Insecta: Neuroptera: Rhachiberothidae) from the Early
Cretaceous amber of Lebanon. Acta Geologica China, 84, 828–833. http://dx.doi.org/10.1111/j.1755-6724.2010.00242.x
Riek, E.F. (1970) Neuroptera (Lacewings). In: Naumann, I.D. (Ed.), The insects of Australia. Melbourne University Press, Melbourne, pp. 472–494.
Ren, D. & Guo, Z. (1996) On the new fossil genera and species of Neuroptera (Insecta) from the Late Jurassic of northeast China. Acta Zootaxonomica Sinica, 21, 461–479.
Ross, A. (1998) Amber: the natural time capsule. The Natural History Museum, London, 73 pp.Rousset, A. (1966) Morphologie céphalique des larves de planipennes (Insectes Névroptéroïdes). Memoires du Museum
Nationale d'Histoire Naturelle, (A), 42, 1–199. Scheven, J. (2004) Bernstein-Einschlüsse: Eine untergegangene Welt bezeugt die Schöpfung. Erinnerungen an die Welt vor der
Sintflut. Kuratorium Lebendige Vorwelt, Hofheim a.T., 160 pp. Schlüter, T. (1978) Zur Systematik und Palökologie harzkonservierter Arthropoda einer Taphozönose aus dem Cenomanium
von NW-Frankreich. Berliner Geowissenschaftliche Abhandlungen, A9, 1–150. Shi, G.H., Grimaldi, D.A., Harlow, G.E., Wang, J., Wang, J., Yang, M.C., Lei, W.Y., Li Q.L. & Li, X.H. (2012) Age constraint
on Burmese amber based on U–Pb dating of zircons. Cretaceous Research, 37, 155–163. http://dx.doi.org/10.1016/j.cretres.2012.03.014
Snodgrass, R.E. (1935) Principles of insect morphology. McGraw-Hill Book Co., New York, 667 pp.Snyder, T.E. (1920) Biology. In: Banks, N. & Snyder, T.E., A revision of the Nearctic termites, with notes on biology and
geographic distribution. Bulletin of the United States National Museum, pp. 108, 87–195. http://dx.doi.org/10.5479/si.03629236.108.i
Stampanoni, M., Groso, A., Isenegger, G., Mikuljan, Q.C., Bertrand, A., Henein, S., Betemps, R., Frommherz, U., Böhler, P.,Meister, D., Lange, M. & Abela, R. (2006) Trends in synchrotron-based tomographic imaging: The SLS experience. Proceedings of SPIE 6318: 6318M-1–6318M-14. http://dx.doi.org/10.1117/12.679497
Su, N.-Y., Scheffrahn, R.H. & Cabrera, B. (2001) Native subterranean termite, Reticulitermes flavipes (Kollar), R. virginicus (Banks), R. hageni Banks. “Featured Creatures,” University of Florida, Department of Entomology and Nematology. Website. EENY-212. Available from: http://creatures.ifas.ufl.edu/urban/termites/native_subterraneans.htm (Accessed 17 Sept. 2013)
Tauber, C.A. (1991) Order Neuroptera. In: Stehr, F.W. (Ed.), Immature Insects, Vol. 2. Kendall/Hunt Publishing Co., Dubuque, Iowa, pp. 126–143.
Tauber, C.A. & Tauber, M.J. (1968) Lomamyia latipennis (Neuroptera, Berothidae) life history and larval descriptions. Canadian Entomologist, 100, 623–629. http://dx.doi.org/10.4039/ent100623-6
Tauber, C.A., Tauber, M.J. & Albuquerque, C.S. (2003) Neuroptera (Lacewings, Antlions). In: Resh, V.H. & Carde, R.T. (Eds.), Encyclopedia of insects. Academic Press, London, pp. 785–798.
Tillyard, R.J. (1916) Studies in Australian Neuroptera. No. iv. The families Ithonidae, Hemerobiidae, Sisyridae, Berothidae, and the new family Trichomatidae; with a discussion of their characters and relationships, and descriptions of new and little-known genera and species. Proceedings of the Linnean Society of New South Wales, 41, 269–332.
Tillyard, R.J. (1926) The Insects of Australia and New Zealand. Angus and Robertson, Sydney, Australia, xi + 560 pp.
Zootaxa 3716 (2) © 2013 Magnolia Press · 257BEROTHID LARVAE FROM BALTIC AMBER

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Tjeder, B. (1959) Neuroptera-Planipennia. The Lace-wings of Southern Africa. 2. Family Berothidae. In: Hanström, B., Brinck, P. & Rudebec, G. (Eds.), South African Animal Life. Results of the Lund University Expedition in 1950–1951, Vol. 6. Almqvist & Wiksell, Stockholm, pp. 256–314.
Toschi, C.A. (1964) Observations on Lomamyia latipennis, with a description of the first instar larva. Pan-Pacific Entomologist, 40, 21–26.
Walker, F. (1853) Part II.– (Sialides – Nemopterides) In: List of the specimens of neuropterous insects in the collection of the British Museum. British Museum, London, [iii] + pp. 193–476.
Wasmann, E. (1906) Beispiele rezenter Artenbildung bei Ameisengasten und Termitengasten. Biologisches Centralblatt, 26, 565–580.
Weitschat, W. & Wichard, W. (1998) Atlas der Pflanzen und Tiere im Baltischen Bernstein. Dr. Friedrich Pfeil Verlag, München, 256 pp. [English edition: (2002) Atlas of plants and animals in Baltic amber]
Whalley, P.E.S. (1980) Neuroptera (Insecta) in amber from the Lower Cretaceous of Lebanon. Bulletin of the British Museum of Natural History (Geology), 33, 157–164.
Whalley, P.E.S. (1983) Fera venatrix gen. and sp. n. (Mantispidae) from amber in Britain. Neuroptera International, 2, 229–233.
Wichard, W., Gröhn, C. & Seredszus, F. (2009) Aquatic insects in Baltic amber. Wasserinsekten im Baltischen Bernstein. Verlag Kessel, Remagen, 336 pp.
Winterton, S.L., Hardy, N.B. & Wiegmann, B.M. (2010) On wings of lace: phylogeny and Bayesian divergence time estimates of Neuropterida (Insecta) based on morphological and molecular data. Systematic Entomology, 35, 349–378. http://dx.doi.org/10.1111/j.1365-3113.2010.00521.x
Withycombe, C.L. (1925) Some aspects of the biology and morphology of the Neuroptera. With special reference to the immature stages and their possible phylogenetic significance. Transactions of the Royal Entomological Society of London, 72 (for 1924), 303–411. http://dx.doi.org/10.1111/j.1365-2311.1925.tb03362.x
WEDMANN ET AL.258 · Zootaxa 3716 (2) © 2013 Magnolia Press
Related Documents











