Vol. 35, No. 1 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 1991, p. 117-123 0066-4804/91/010117-07$02.00/0 Copyright © 1991, American Society for Microbiology First-Exposure Adaptive Resistance to Aminoglycoside Antibiotics In Vivo with Meaning for Optimal Clinical Use GEORGE L. DAIKOS,1 VALENTINA T. LOLANS,2 AND GEORGE GEE JACKSON3* Department of Medicine, University of Miami, Miami, Florida 331241; Department of Pediatrics, Rush-Presbyterian- St. Luke's School of Medicine, Chicago, Illinois 606122; and Division of Infectious Diseases, Department of Medicine, University of Utah School of Medicine, 50 North Medical Drive, Salt Lake City, Utah 841323 Received 6 June 1990/Accepted 12 October 1990 The first exposure of gram-negative bacilli to an aminoglycoside antibiotic in vitro induces a biphasic bactericidal response and adaptive drug resistance (G. L. Daikos, G. G. Jackson, V. T. Lolans, and D. M. Livermore, J. Infect. Dis. 162:414-420, 1990; G. G. Jackson, G. L. Daikos, and V. T. Lolans, J. Infect. Dis. 162:408-413, 1990). The therapeutic implications were examined in netilmicin treatment of a Pseudomonas aeruginosa infection of normal and neutropenic mice. For 2 h after the first dose, the bactericidal rates were rapid, 0.75, 1.0, and 1.5 loglo CFU/h with doses of 10, 30, and 60 mg/kg, respectively. Each twofold increase in dosage reduced the number of surviving bacteria fivefold. Between 2 and 6 h, the second-phase bactericidal rate was slow, c0.3 loglo CFU/h, regardless of the dose. In a multiple-dose regimen, the same amount of netilmicin given in one dose was 70 and 90% more effective than two or three doses, respectively. Doses calculated to keep the drug level in plasma above the MIC were less effective than regimens giving first exposure to a high drug concentration. Adaptive resistance occurred when doses were given more than 2 h after the start of treatment. Temporary survival of bacteremic neutropenic mice was 60 to 70% greater with a second dose at 2 h than after a longer interval. In a thigh infection of neutropenic mice treated every 2 h, doses 4, 6, and 8 h after the first one showed no bactericidal effect. A drug-free interval of 8 h (20 times the drug half-life) renewed bacterial susceptibility to drug action. The results in vivo confirm the biphasic bactericidal action and induction of adaptive resistance that characterize first exposure of gram-negative bacilli to aminoglycoside antibiotics. The phenomena have meaning for the optimum clinical use of aminoglycosides. The rate of the bactericidal action of an aminoglycoside antibiotic on Pseudomonas aeruginosa and other aerobic gram-negative bacilli has been observed to be biphasic in vitro (23). An initial phase of rapid bacterial killing is induced by passive ionic binding of the drug to bacterial lipopolysac- charide (16, 23, 24, 30). The killing rate is directly related to the initial drug concentration. A second phase of slower bacterial killing is associated with decreased energy-depen- dent uptake of the aminoglycoside, and the rate is indepen- dent of the initial or persistent drug level (6, 14, 23, 28). Bacteria surviving the first exposure develop adaptive resis- tance, which is mediated by impermeability to all aminogly- cosides (7, 22b). Adaptive resistance is unstable and is reversed during growth in drug-free media. If the same interactions between aminoglycosides and bacteria apply in the treatment of bacterial infections, the data are relevant to selection of the most effective dosing regimen for therapeutic use of these antibiotics. The inves- tigations reported here were designed to determine whether the biphasic bactericidal action of aminoglycoside antibiotics observed in vitro also applies in the treatment of an experi- mental infection and whether the first-exposure effect in- cludes the development of adaptive resistance. MATERIALS AND METHODS Experimental infection. Infection was produced in the thighs of normal or neutropenic female ICR mice weighing 23 to 28 g (Sprague Dawley Laboratory, Madison, Wis.) by injection of 0.1 ml of broth containing approximately 107 CFU of bacteria in the log phase of growth per ml. Experi- * Corresponding author. ments were performed with three strains of P. aeruginosa: the laboratory strain, ATCC 27853, and two clinical isolates, PA-Dl and PA-D13. The MIC and MBC of netilmicin were 0.5 and 2.0 ,ug/ml, respectively, for all three strains. The number of viable organisms was measured at specified times by skinning the thigh and excising the soft tissues from the bone of animals killed by anesthesia. The muscles were homogenized with 1 ml of sterile 0.85% saline with a hand-operated ground-glass tissue homogenizer. The ho- mogenate was kept at 4°C while quantitative cultures were done. Serial dilutions of the homogenate were cultured on Mueller-Hinton agar (MHA) plates, and the CFU were counted after overnight incubation at 37°C. Neutropenia was produced in mice by intraperitoneal injection of 150 and 100 mg of cyclophosphamide per kg 3 and 1 days, respectively, before the experiment. On the days of the experiments, 16 uninfected mice were exsanguinated to confirm the neutropenia. A fatal bacteremic infection was initiated in groups of neutropenic mice by intraperitoneal injection of 0.1 ml of broth containing approximately 108 CFU of bacteria. The survival record of the animals and the treatment schedule included observations after 2, 4, 6, 8, 12, and 24 h. Antibiotic treatment. Netilmicin was made up fresh from a powder standard in a stock solution of 15 mg/ml. For use in treatment, dilutions were prepared from this stock solution in which the prescribed dose was present in 0.1 ml. Antibi- otic treatment was given by intraperitoneal injection of the dose at specified times. The level in plasma was determined by using an agar-well diffusion microbiologic assay. Blood was pooled from three to five uninfected mice exsanguinated 15, 60, and 120 min after the administration of drug. As therapy, the first dose was given after initiation of the 117 on June 30, 2018 by guest http://aac.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 35, No. 1ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 1991, p. 117-1230066-4804/91/010117-07$02.00/0Copyright © 1991, American Society for Microbiology
First-Exposure Adaptive Resistance to Aminoglycoside AntibioticsIn Vivo with Meaning for Optimal Clinical Use
GEORGE L. DAIKOS,1 VALENTINA T. LOLANS,2 AND GEORGE GEE JACKSON3*Department of Medicine, University of Miami, Miami, Florida 331241; Department of Pediatrics, Rush-Presbyterian-St. Luke's School of Medicine, Chicago, Illinois 606122; and Division of Infectious Diseases, Department of Medicine,
University of Utah School of Medicine, 50 North Medical Drive, Salt Lake City, Utah 841323
Received 6 June 1990/Accepted 12 October 1990
The first exposure of gram-negative bacilli to an aminoglycoside antibiotic in vitro induces a biphasicbactericidal response and adaptive drug resistance (G. L. Daikos, G. G. Jackson, V. T. Lolans, and D. M.Livermore, J. Infect. Dis. 162:414-420, 1990; G. G. Jackson, G. L. Daikos, and V. T. Lolans, J. Infect. Dis.162:408-413, 1990). The therapeutic implications were examined in netilmicin treatment of a Pseudomonasaeruginosa infection of normal and neutropenic mice. For 2 h after the first dose, the bactericidal rates wererapid, 0.75, 1.0, and 1.5 loglo CFU/h with doses of 10, 30, and 60 mg/kg, respectively. Each twofold increasein dosage reduced the number of surviving bacteria fivefold. Between 2 and 6 h, the second-phase bactericidalrate was slow, c0.3 loglo CFU/h, regardless of the dose. In a multiple-dose regimen, the same amount ofnetilmicin given in one dose was 70 and 90% more effective than two or three doses, respectively. Dosescalculated to keep the drug level in plasma above the MIC were less effective than regimens giving first exposureto a high drug concentration. Adaptive resistance occurred when doses were given more than 2 h after the startof treatment. Temporary survival of bacteremic neutropenic mice was 60 to 70% greater with a second doseat 2 h than after a longer interval. In a thigh infection of neutropenic mice treated every 2 h, doses 4, 6, and8 h after the first one showed no bactericidal effect. A drug-free interval of 8 h (20 times the drug half-life)renewed bacterial susceptibility to drug action. The results in vivo confirm the biphasic bactericidal action andinduction of adaptive resistance that characterize first exposure of gram-negative bacilli to aminoglycosideantibiotics. The phenomena have meaning for the optimum clinical use of aminoglycosides.
The rate of the bactericidal action of an aminoglycosideantibiotic on Pseudomonas aeruginosa and other aerobicgram-negative bacilli has been observed to be biphasic invitro (23). An initial phase of rapid bacterial killing is inducedby passive ionic binding of the drug to bacterial lipopolysac-charide (16, 23, 24, 30). The killing rate is directly related tothe initial drug concentration. A second phase of slowerbacterial killing is associated with decreased energy-depen-dent uptake of the aminoglycoside, and the rate is indepen-dent of the initial or persistent drug level (6, 14, 23, 28).Bacteria surviving the first exposure develop adaptive resis-tance, which is mediated by impermeability to all aminogly-cosides (7, 22b). Adaptive resistance is unstable and isreversed during growth in drug-free media.
If the same interactions between aminoglycosides andbacteria apply in the treatment of bacterial infections, thedata are relevant to selection of the most effective dosingregimen for therapeutic use of these antibiotics. The inves-tigations reported here were designed to determine whetherthe biphasic bactericidal action of aminoglycoside antibioticsobserved in vitro also applies in the treatment of an experi-mental infection and whether the first-exposure effect in-cludes the development of adaptive resistance.
MATERIALS AND METHODSExperimental infection. Infection was produced in the
thighs of normal or neutropenic female ICR mice weighing23 to 28 g (Sprague Dawley Laboratory, Madison, Wis.) byinjection of 0.1 ml of broth containing approximately 107CFU of bacteria in the log phase of growth per ml. Experi-
* Corresponding author.
ments were performed with three strains of P. aeruginosa:the laboratory strain, ATCC 27853, and two clinical isolates,PA-Dl and PA-D13. The MIC and MBC of netilmicin were0.5 and 2.0 ,ug/ml, respectively, for all three strains. Thenumber of viable organisms was measured at specified timesby skinning the thigh and excising the soft tissues from thebone of animals killed by anesthesia. The muscles werehomogenized with 1 ml of sterile 0.85% saline with ahand-operated ground-glass tissue homogenizer. The ho-mogenate was kept at 4°C while quantitative cultures weredone. Serial dilutions of the homogenate were cultured onMueller-Hinton agar (MHA) plates, and the CFU werecounted after overnight incubation at 37°C.Neutropenia was produced in mice by intraperitoneal
injection of 150 and 100 mg of cyclophosphamide per kg 3and 1 days, respectively, before the experiment. On the daysof the experiments, 16 uninfected mice were exsanguinatedto confirm the neutropenia. A fatal bacteremic infection wasinitiated in groups of neutropenic mice by intraperitonealinjection of 0.1 ml of broth containing approximately 108CFU of bacteria. The survival record of the animals and thetreatment schedule included observations after 2, 4, 6, 8, 12,and 24 h.
Antibiotic treatment. Netilmicin was made up fresh from apowder standard in a stock solution of 15 mg/ml. For use intreatment, dilutions were prepared from this stock solutionin which the prescribed dose was present in 0.1 ml. Antibi-otic treatment was given by intraperitoneal injection of thedose at specified times. The level in plasma was determinedby using an agar-well diffusion microbiologic assay. Bloodwas pooled from three to five uninfected mice exsanguinated15, 60, and 120 min after the administration of drug. Astherapy, the first dose was given after initiation of the
117
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ANTIMICROB. AGENTS CHEMOTHER.
64'
32.Ia. 16'
c 8o0
E 4-2; 2
1'
0.5
1
60Time post Dose (min)
120
FIG. 1. Serum netilmicin levels at intervals during the first 2 hafter intraperitoneal injection of 15, 30, or 60 mg/kg into miceweighing approximately 25 g. The serum from three to five animalswas pooled to obtain the average level.
infection in the mouse thigh. Retreatment of different groups
of mice was varied between only the initial dose and a doseevery 2 h for five doses. The total amount of drug given per
mouse varied between 10 and 130 mg/kg. The statisticalsignificance of the variance among groups was determinedby the t test, and the differences in survival were calculatedby chi square.
RESULTSLevels of netilmicin in serum during the 2 h following a
dose of 15, 30, or 60 mg/kg are shown in Fig. 1. The level 15min after the 60-mg/kg dose was 50 ,ug/ml. In the first 2 h, thedrug level declined to 3 ,ug/ml, indicating a drug half-life ('1,2;for mice) of s25 min. Doses of 30 and 15 mg/kg producedproportionately lower levels in serum. All doses producedlevels in plasma above the MIC for the test strain for at least2 h.The bactericidal effect of three regimens that delivered the
same total dose of netilmicin with one, two, or six doses was
used to evaluate the relative effects of a high peak level anda continuous inhibitory concentration in groups of miceinfected with one of two different strains of P. aeruginosa.An initial dose of 60, 30, or 10 mg/kg was given immediatelyafter infection and was followed by no, one, or five doses ofthe respective amount at intervals of 3 h or hourly from 0 to5 h. Results of the treatment regimens after 2 and 6 h are
shown in Fig. 2. In untreated mice, the number of CFU inthe thigh muscles 6 h after infection with 3 x 106 organismsof strain PA-D13 increased five times over the input number;an inoculum of 6 x 106 organisms of strain ATCC 27853increased 1.6 times. The resultant level of the untreatedinfection was 7.2 and 7.0 log1o CFU per mouse thigh,respectively, for the two strains.The bactericidal effect of netilmicin in any treatment
regimen was biphasic. During the first 2 h, the largest dose,60 mg/kg, with the highest first-exposure drug level, killedthe inoculum at a rate of 1.5 log1o CFU/h. With a doseone-half as large (30 mg/kg), the bactericidal rate was one-
third lower (1.0 log1o CFU/h) and the number of survivingCFU was five times greater. One-third the dose, 20 mg/kg,given in two doses at 0 and 1 h had a bactericidal rate
one-half as fast, 0.75 log1o CFU/h, and more than 25 timesthe number of surviving CFU. The results were comparablewith the two strains.Between 2 and 6 h of treatment, the bactericidal rates were
reduced and similar for the different dosage regimens, asshown by the parallel slope of the lines in Fig. 2. In animalsinfected with strain PA-D13, the second-phase bactericidalrate was 0.3 log1o CFU/h regardless of the dosage regimen.For strain ATCC 27835, the second-phase bactericidal rateswere 0.1, 0.16, and 0.31 log1o CFU/h with regimens of one,two, and six doses. The net bactericidal effect after 6 hamong mice given the same cumulative dose of netilmicin ondifferent schedules was significantly different. The singledose with a high initial level reduced surviving CFU 6 and 12times more than the same amount of drug given in two or sixdoses, respectively. Two cycles of treatment with an inter-mediate-size dose reduced surviving CFU more effectivelythan six doses that were calculated to keep the drug levelabove the MIC. Thus, the height of the level and not thepressure of a constant MIC was the most bactericidalregimen. The results with the two strains were comparable,and the differences were statistically significant.
In a similar experiment, but with extension of the treat-ment period to 12 h, three groups of mice were treated witha total dose of 30 mg of netilmicin per kg given as one, two,or three doses at 0, 0, and 6 or at 0, 6, and 12 h after initiationof infection. The numbers of surviving bacteria with eachregimen were compared after 6 h (one dose on each regimen)and 24 h when the amount of drug received was the same(Table 1). The kinetics of the bactericidal action were thesame: early killing at a faster rate, 1.4 to 0.8 log1o CFU/h,was directly related to the size of the first dose. After 6 h, adose of 30 mg/kg was 80% (five times fewer CFU) and 87%(eight times fewer CFU) more effective in reducing thenumber of surviving bacteria than smaller first doses of 15mg (one-half) and 10 mg (one-third), respectively.
In the second phase, from 6 to 24 h, the rates of decline inviable bacteria were slower and comparable for each of thetreatment schedules. The net CFU in the latter periodincluded the postantibiotic effect and ability of normal miceto contain the treated local infection. After 24 h, when allanimals had received the same cumulative dose of netilmicinin 12 h, first exposure to a high drug level from a single dosereduced surviving CFU by 50 to 80% and 63 to 90% morethan regimens that delivered the drug in two or three divideddoses, respectively. The differences are statistically signifi-cant.The 6-h dosing interval used was approximately every
15th half-life of the drug in mice. From the observed levels inserum in Fig. 1, the aggregate times that the drug level inserum was above the MIC for the infecting strain wereapproximately 3, 4, and 5 h for the one-, two-, and three-dose regimens, respectively. Thus, the time that the level inserum was above the MIC was inversely rather than directlyrelated to the final bactericidal effect, and the concentrationof the drug upon first exposure correlated directly withbacterial reduction.The bactericidal effects of the first and second doses were
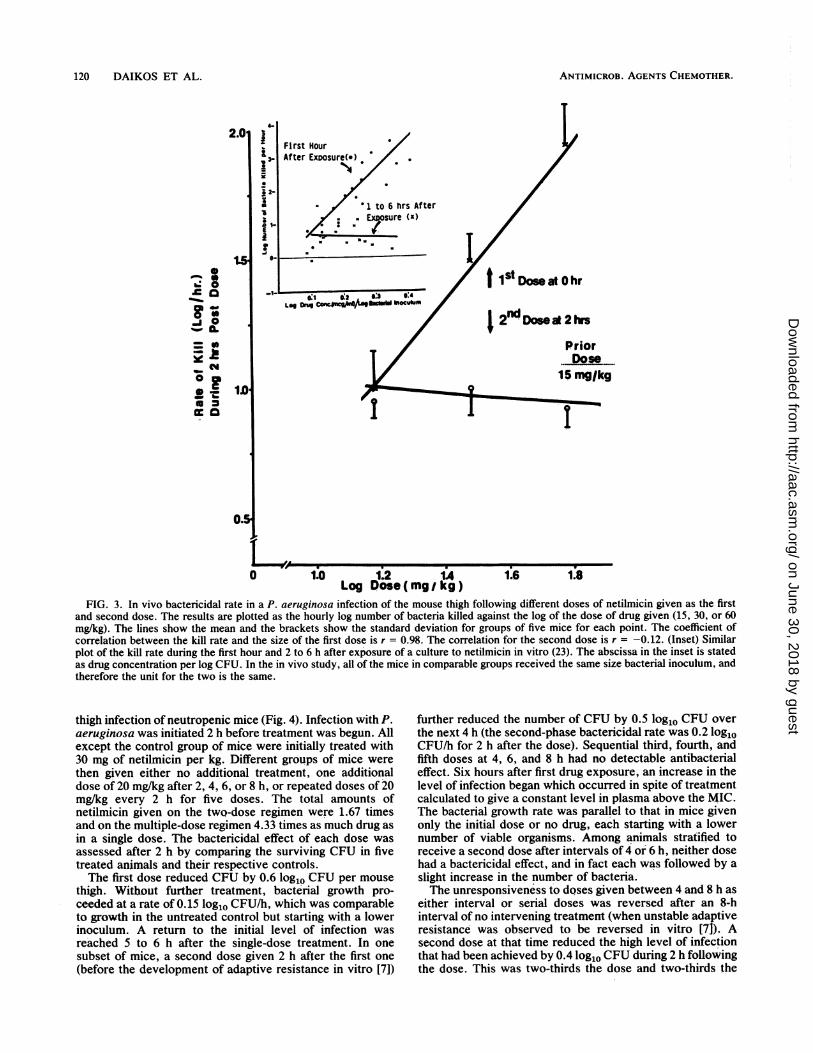
compared directly in the treatment of a pseudomonal thighinfection with strain ATCC 27853 (Fig. 3). Treatment wasinitiated with a first dose of either 15, 30, or 60 mg/kg; 2 hlater, the bactericidal rates were observed to be 1.0 ± 0.27(standard deviation), 1.5 ± 0.2, and 1.9 + 0.3 log1o CFU/h,respectively. Mice initially given the lowest dose, 15 mg/kg,were reassigned to one of three new subgroups and retreatedwith a second dose of either 15, 30, or 60 mg/kg. Two hours
118 DAIKOS ET AL.
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ADAPTIVE RESISTANCE TO AMINOGLYCOSIDES IN VIVO
cO0.01
Time (hours)
6
P; 0.01
e aeru gino s a ATCC 27853
0 2 4
Time (hours)
6
FIG. 2. Biphasic bactericidal action of the first-exposure effect of netilmicin in reducing the number of surviving bacteria in the thighmuscle of mice experimentally infected with P. aeruginosa. The right- and left-hand panels show the results for two different strains of P.aeruginosa. Each line shows the mean results, and the brackets show the standard deviation, for groups of five mice given a different initialdose in a treatment regimen delivering the same total amount of netilmicin in six doses (O to 5 h) after initiation of the infection. The rapidlybactericidal first phase and slowly bactericidal second phase applied to all dosage regimens, producing statistically significant differences inthe number of surviving bacteria. Control animals were given no treatment and had an increase in the number of bacteria in the infected thighmuscle.
later, the CFU per thigh were again enumerated. The bac-tericidal rates during 2 h after the second dose were 0.9, 1.0,and 0.9 log1o CFU/h for the 15-, 30-, and 60-mg/kg doses,respectively. Thus, the bactericidal rate following the sec-ond dose was the same and was not determined by the sizeof the dose. The inset in Fig. 3 compares the kinetics ofsimilar observations made in vitro with another strain of P.aeruginosa (23).To further evaluate the importance of the first-exposure
effect noted above and the in vivo expression of adaptive
resistance, mice were made neutropenic to remove thataspect of host immunity. On the day of the experiments, theleukocyte count of 16 leukopenic mice was 700 140/mm3.No more than two granulocytes were found on each bloodsmear. In eight normal mice, the leukocyte count was 3,400
850/mm3, and granulocytes predominated in the bloodfilm.The bactericidal effects of a single dose and different
schedules of repetitive dosing with evaluation over an ex-tended treatment period of 12 h were compared in the mouse
TABLE 1. Effect of dosing regimen of netilmicin in reducing the number of surviving bacteria during P. aeruginosainfection in neutropenic micea
Strain PA-D13 Strain ATCC 27853Treatment regimen LgoCUtih-- DLgoCUtih--S(total given) No. of Log10 CFU/thigh ± SD No. of Log10 CFU/thigh ± SD
mice 6h 24 h mice 6 h 24 h
No treatment 10 6.8 ± 0.10 7.2 ± 0.25 5 6.9 ± 0.20 7.5 ± 0.30(a) 10 mg/kg every 6 h (30 mg/kg) 7 4.9 ± 0.30 3.8 + 0.37 5 5.7 ± 0.78 4.3 ± 0.45(b) 15 mg/kg every 3 h (30 mg/kg) 10 4.7 ± 0.40 3.7 ± 0.80 5 5.5 ± 0.73 4.2 ± 0.60(c) 30 mg/kg at 0 h 10 4.0 + 0.40 2.8 + 0.50 5 4.8 + 0.16 3.5 ± 0.55
a Significance: a versus c, P = 0.01 at 6 and 24 h for PA-D13, P = 0.05 at 6 and 24 h for ATCC 27853; b versus c, P = 0.01 at 6 h and 0.05 at 24 h for strainPA-D13, not significant at either time for ATCC 27853.
8
a,
N
IL
0or0-J
VOL. 35, 1991 119
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
ANTIMICROB. AGENTS CHEMOTHER.
First HourAfter ExDosure(s) .
N
I1 to 6 hrs AfteroEsure (x)
*
-is
IStDoseatOhr
2nodDoseat 2hrs
PriorDose*.*
15 mglkg
I
l1. 1.2 IALog Dose(mgj kg)
1.6 1.8
FIG. 3. In vivo bactericidal rate in a P. aeruginosa infection of the mouse thigh following different doses of netilmicin given as the firstand second dose. The results are plotted as the hourly log number of bacteria killed against the log of the dose of drug given (15, 30, or 60mg/kg). The lines show the mean and the brackets show the standard deviation for groups of five mice for each point. The coefficient of
correlation between the kill rate and the size of the first dose is r = 0.98. The correlation for the second dose is r = -0.12. (Inset) Similarplot of the kill rate during the first hour and 2 to 6 h after exposure of a culture to netilmicin in vitro (23). The abscissa in the inset is statedas drug concentration per log CFU. In the in vivo study, all of the mice in comparable groups received the same size bacterial inoculum, andtherefore the unit for the two is the same.
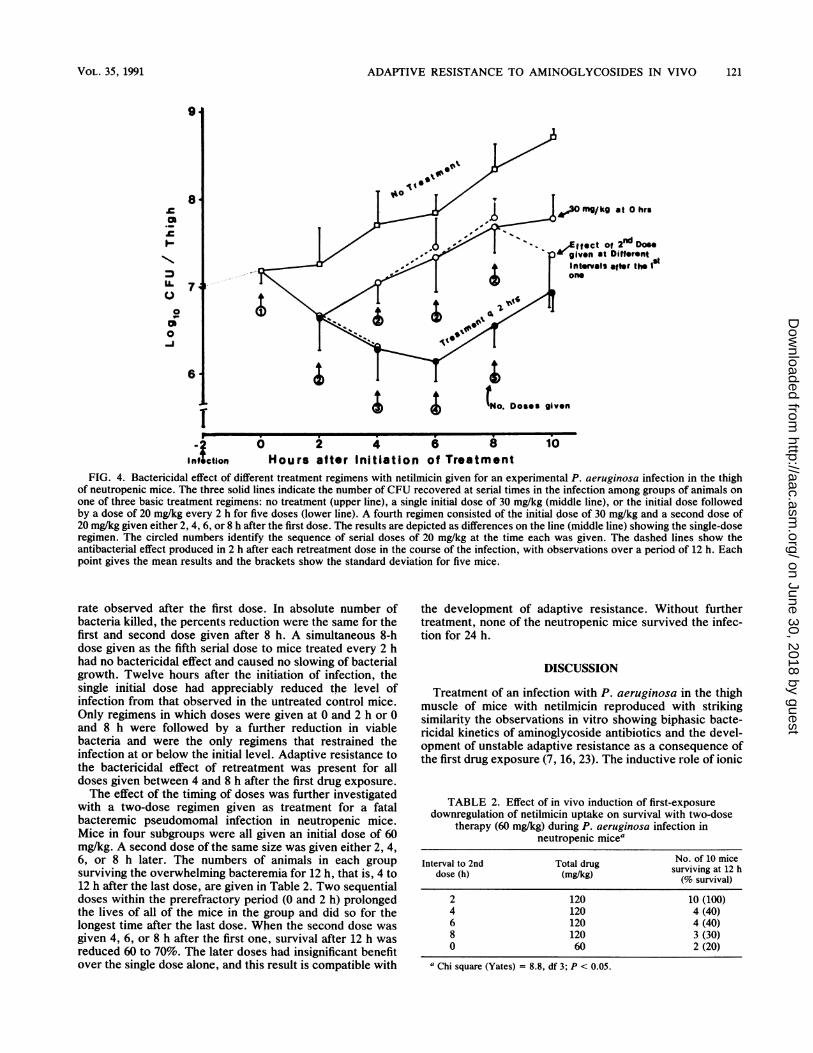
thigh infection of neutropenic mice (Fig. 4). Infection with P.aeruginosa was initiated 2 h before treatment was begun. Allexcept the control group of mice were initially treated with30 mg of netilmicin per kg. Different groups of mice werethen given either no additional treatment, one additionaldose of 20 mg/kg after 2, 4, 6, or 8 h, or repeated doses of 20mg/kg every 2 h for five doses. The total amounts ofnetilmicin given on the two-dose regimen were 1.67 timesand on the multiple-dose regimen 4.33 times as much drug as
in a single dose. The bactericidal effect of each dose wasassessed after 2 h by comparing the surviving CFU in fivetreated animals and their respective controls.The first dose reduced CFU by 0.6 log1o CFU per mouse
thigh. Without further treatment, bacterial growth pro-ceeded at a rate of 0.15 log1o CFU/h, which was comparableto growth in the untreated control but starting with a lowerinoculum. A return to the initial level of infection wasreached 5 to 6 h after the single-dose treatment. In onesubset of mice, a second dose given 2 h after the first one
(before the development of adaptive resistance in vitro [7])
further reduced the number of CFU by 0.5 log1o CFU over
the next 4 h (the second-phase bactericidal rate was 0.2 log1oCFU/h for 2 h after the dose). Sequential third, fourth, andfifth doses at 4, 6, and 8 h had no detectable antibacterialeffect. Six hours after first drug exposure, an increase in thelevel of infection began which occurred in spite of treatmentcalculated to give a constant level in plasma above the MIC.The bacterial growth rate was parallel to that in mice givenonly the initial dose or no drug, each starting with a lowernumber of viable organisms. Among animals stratified toreceive a second dose after intervals of 4 or 6 h, neither dosehad a bactericidal effect, and in fact each was followed by a
slight increase in the number of bacteria.The unresponsiveness to doses given between 4 and 8 h as
either interval or serial doses was reversed after an 8-hinterval of no intervening treatment (when unstable adaptiveresistance was observed to be reversed in vitro [71). Asecond dose at that time reduced the high level of infectionthat had been achieved by 0.4 log1o CFU during 2 h followingthe dose. This was two-thirds the dose and two-thirds the
2.01
0.= *_ zoj 0
= lD
ws- C#40 ?D
I
X 2-
a1-E
.j0-
10
p
120 DAIKOS ET AL.
4.-
o.s
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
ADAPTIVE RESISTANCE TO AMINOGLYCOSIDES IN VIVO
N."
CN
0
0
-1
6
030 mg/ kg at O hrs
Effect of 2nd Doe.given at Different SIntervals after theone
I o.Doses given
I fSftion
0 2 4 6 8Hours after Initiation of Treatment
10
FIG. 4. Bactericidal effect of different treatment regimens with netilmicin given for an experimental P. aeruginosa infection in the thighof neutropenic mice. The three solid lines indicate the number of CFU recovered at serial times in the infection among groups of animals onone of three basic treatment regimens: no treatment (upper line), a single initial dose of 30 mg/kg (middle line), or the initial dose followedby a dose of 20 mg/kg every 2 h for five doses (lower line). A fourth regimen consisted of the initial dose of 30 mg/kg and a second dose of20 mg/kg given either 2, 4, 6, or 8 h after the first dose. The results are depicted as differences on the line (middle line) showing the single-doseregimen. The circled numbers identify the sequence of serial doses of 20 mg/kg at the time each was given. The dashed lines show theantibacterial effect produced in 2 h after each retreatment dose in the course of the infection, with observations over a period of 12 h. Eachpoint gives the mean results and the brackets show the standard deviation for five mice.
rate observed after the first dose. In absolute number ofbacteria killed, the percents reduction were the same for thefirst and second dose given after 8 h. A simultaneous 8-hdose given as the fifth serial dose to mice treated every 2 hhad no bactericidal effect and caused no slowing of bacterialgrowth. Twelve hours after the initiation of infection, thesingle initial dose had appreciably reduced the level ofinfection from that observed in the untreated control mice.Only regimens in which doses were given at 0 and 2 h or 0and 8 h were followed by a further reduction in viablebacteria and were the only regimens that restrained theinfection at or below the initial level. Adaptive resistance tothe bactericidal effect of retreatment was present for alldoses given between 4 and 8 h after the first drug exposure.The effect of the timing of doses was further investigated
with a two-dose regimen given as treatment for a fatalbacteremic pseudomomal infection in neutropenic mice.Mice in four subgroups were all given an initial dose of 60mg/kg. A second dose of the same size was given either 2, 4,6, or 8 h later. The numbers of animals in each groupsurviving the overwhelming bacteremia for 12 h, that is, 4 to12 h after the last dose, are given in Table 2. Two sequentialdoses within the prerefractory period (0 and 2 h) prolongedthe lives of all of the mice in the group and did so for thelongest time after the last dose. When the second dose wasgiven 4, 6, or 8 h after the first one, survival after 12 h wasreduced 60 to 70%. The later doses had insignificant benefitover the single dose alone, and this result is compatible with
the development of adaptive resistance. Without furthertreatment, none of the neutropenic mice survived the infec-tion for 24 h.
DISCUSSION
Treatment of an infection with P. aeruginosa in the thighmuscle of mice with netilmicin reproduced with strikingsimilarity the observations in vitro showing biphasic bacte-ricidal kinetics of aminoglycoside antibiotics and the devel-opment of unstable adaptive resistance as a consequence ofthe first drug exposure (7, 16, 23). The inductive role of ionic
TABLE 2. Effect of in vivo induction of first-exposuredownregulation of netilmicin uptake on survival with two-dose
therapy (60 mg/kg) during P. aeruginosa infection inneutropenic mice'
Interval to 2nd Total drug No. of 10 micedose (h) (mg/kg) surviving at 12 h
2 120 10 (100)4 120 4 (40)6 120 4 (40)8 120 3 (30)0 60 2 (20)
a Chi square (Yates) = 8.8, df 3; P < 0.05.
VOL. 35, 1991 121
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ANTIMICROB. AGENTS CHEMOTHER.
binding initiating the early phase of rapid bactericidal actionis passive and entirely dependent on the drug concentration.The accelerated unidirectional bacterial transport of thebound drug was shown by the direct relationship betweendose and the high rate of bactericidal action immediatelyafter the first dose. The second phase of antibacterial action,in which drug uptake is rate limited, independent of the dose,and has a slow fixed bactericidal rate, was also observed.Together they represent the unique first-exposure effect ofaminoglycosides on gram-negative bacteria (16, 23).
Adaptive resistance appeared before 4 h after the firstdose. It was revealed by the failure of doses between 4 and8 h after the first one to produce any appreciable bactericidalaction while the number of bacteria was increasing. That theresistance is adaptive and not the result of selection ofresistant mutants is shown by the requirement for an adap-tive period for development of drug refractoriness andreversal to susceptibility after an interval without drugexposure. Further evidence from other studies has shownadaptive resistance to be inducible by either one-fourth orfour times the MIC with the same kinetics and that the effectinduced by one member of the class produced uniformadaptive cross-over impermeability resistance to other ami-noglycosides (7, 22b). None of these effects suggest that theobservations are the result of drug-selected resistant mu-tants. The mechanism of adaptive resistance was shown tobe mediated by failure to transport drug into the bacterialcell (7). This ability of aminoglycosides to alter the bacterialprocesses necessary for their own uptake has been observedin various ways (1, 13, 14, 22a, 24).
Netilmicin and P. aeruginosa were chosen for investiga-tion, but other gram-negative bacilli and aminoglycosidesinteract in a similar fashion. However, the evolution of thephase, completeness of the refractory state, and its reversalmay vary among different species and requires additionalinvestigation. Biphasic bactericidal kinetics and unstableadaptive resistance were demonstrated by Kapusnic andcoworkers treating experimental pneumonia in nonneutro-penic guinea pigs with tobramycin (18). Similar results maybe obsprved in the results of other investigators usingdifferent experimental models of infection (4, 9-12, 18, 19,21, 31).The unique drug-reactive phenomena, including the devel-
opment of adaptive resistance, give importance to the ob-servations in developing optimum therapeutic regimens forclinical use of aminoglycoside antibiotics. It has been sug-gested in other reports that the area under the time-concen-tration curve (AUC) is a reliable pharmacokinetic determi-nant for aminoglycoside efficacy (22, 35). Although thiscorrelation can be verified by the present work, its rational-ization lies in the effect of the height of the initial drug leveland its reflection in the AUC; and the correlation with AUCapplies only to the first and not serial doses if the interval isshort. The proportional amount of time that the drug level inplasma was above the MIC for the infecting strain wasunimportant. This irrationality in comparison with beta-lactam antibiotics, for which maintenance of an inhibitorydrug level is thp major pharmacokinetic determinant ofefficacy, can be explained by the differences in site andmechanisms of action; the self-promoted internalization ofbound aminoglycoside, the concentration-independent sec-ond phase of aminoglycoside action, and adaptive resistanceare all involved.Some clinical experience supports the importance of the
first dose and the height of the peak plasma drug levelsachieved (25-27, 29). In the treatment of experimental infec-
tion in mice, a second dose during the prerefractory periodalso increased the antibacterial effect of the first dose. Invitro and in neutropenic mice, evidence of increasing drugrefractoriness is apparent within 3 to 4 h after the first dose.Both in vitro and in vivo experience indicates that after thistime, the continued presence of drug in the medium in-creases and prolongs adaptive resistance. The common useof low-dose, short-interval administration of aminoglyco-sides and the aim of maintaining the level above the MICcould both augment adaptive resistance and minimize thebactericidal potential (2, 5, 7, 17). Both effects work to thedetriment of patients.
Reversal from adaptive resistance to full susceptibility,which was clearly demonstrable in vitro, was also observedin vivo. However, it is more difficult to show in vivo becauseof the intervening importance of host status. In some exper-iments, independent recovery of normal animals or failure toprevent death of neutropenic mice with bacteremia occurredbefore the time and bacterial growth required for observa-tion of reemergence of susceptibility were fulfilled. Whetherdrug-free periods of adequate length to permit return of drugsusceptibility can be recommended clinically must dependon the immunologic competence of the host and the antibac-terial effectiveness of the high first dose and cyclic levels.Once-daily dosing approaches the principle and appears tobe at least as effective as multiple daily dosing (20, 33, 34).
Toxicity has always been a major concern in the use ofaminoglycoside antibiotics. Implications of the first-expo-sure effect and development of adaptive resistance suggestthe uselessness of doses given during the unrecognizedrefractory period. These doses could double or quadruplethe amount of drug, increasing toxicity without a parallelbenefit from increased antibacterial activity. Increasingly,experience and opinion indicate that severe oto- and neph-rotoxicities are primarily caused by duration of treatmentwith a persistent drug level and not by brief periods of highdrug levels (3, 8, 31, 32, 36). Transient levels of amikacin andtobramycin higher than 300 and 40 ,ug/ml, respectively, inpatients caused no evidence of neuromuscular blockade,nephrotoxicity, or ototoxicity (15, 31). The margin withnetilmicin is greater. Ensuring an adequate initial dose toobtain the early bactericidal effect is critical, and avoidingrepeated doses in the period required for complete renalclearance of the drug before the next dose would be wise.Acceptance of such a regimen could accomplish the twingoals of avoiding severe toxicity and minimizing adaptiveresistance, leading to a more favorable toxicity-efficacyratio. Furthermore, the practice would permit dosing on thebasis of known pharmacokinetics and renal function, withelimination of the expense of serial monitoring of the druglevel. Thus, not only the toxicity-efficacy but also thecost-benefit ratio could be improved.The experimental data presented give a rationale for
high-dosage, long-interval administration of aminoglycosideantibiotics. Understanding the unique bactericidal kineticsrelated to the first-exposure effect of aminoglycoside antibi-otics, including adaptive resistance, should improve oppor-tunities to use them more effectively and safely in thetreatment of severe infections caused by the common aero-bic gram-negative bacilli.
ACKNOWLEDGMENTS
The technical assistance of Sue Lyza and Ria Chiu in the conductof the animal experiments is gratefully acknowledged.
122 DAIKOS ET AL.
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ADAPTIVE RESISTANCE TO AMINOGLYCOSIDES IN VIVO
REFERENCES1. Allen, N. E., W. E. Alborn, Jr., A. Kirst, and J. E. Toth. 1987.
Comparison of aminoglycoside antibiotics with respect to up-take and lethal activity in Escherichia coli. J. Med. Chem.30:333-340.
2. Anderson, E. T., L. S. Young, and W. L. Hewitt. 1976. Simul-taneous antibiotic levels in "breakthrough" gram-negative rodbacteremia. Am. J. Med. 61:493-497.
3. Bennett, W. M., C. E. Plamp, D. N. Gilbert, R. A. Parker, andG. A. Porter. 1979. The influence of dosage regimen on exper-imental gentamicin nephrotoxicity: dissociation of peak serumlevel from renal failure. J. Infect. Dis. 140:576-580.
4. Blaser, J., B. B. Stone, M. C. Groner, and S. H. Zinner. 1987.Comparative study with enoxacin and netilmicin in a pharma-codynamic model to determine importance of ratio of antibioticpeak concentration to MIC for bactericidal activity and emer-gence of resistance. Antimicrob. Agents Chemother. 31:1054-1060.
5. Bodey, G. P., E. Middleman, T. Umsawasid, and V. Rodriguez.1971. Intravenous gentamicin therapy for infections in patientswith cancer. J. Infect. Dis. 124:S174-S179.
6. Bryan, L. E., and S. Kwan. 1983. Roles of ribosomal binding,membrane potential, and electron transport in bacterial uptakeof streptomycin and gentamicin. Antimicrob. Agents Chemo-ther. 23:835-845.
7. Daikos, G. L., G. G. Jackson, V. T. Lolans, and D. M. Liver-more. 1990. Adaptive resistance to aminoglycoside antibioticsinduced by first exposure down regulation. J. Infect. Dis.162:414-420.
8. Frame, P. T., J. T. Phair, C. Watanakunakorn, and T. W. P.Bannister. 1977. Pharmacologic factors associated with genta-micin nephrotoxicity in rabbits. J. Infect. Dis. 135:952-956.
9. Gerber, A. U., H. P. Brugger, C. Geller, T. Stritzko, and B.Stalder. 1986. Antibiotic therapy of infections due to P. aerug-inosa in normal and granulocytopenic mice: comparison ofmurine and human pharmacokinetics. J. Infect. Dis. 153:90-97.
10. Gerber, A. U., W. A. Craig, H. P. Brugger, C. Geller, A. P.Vastola, and J. Brandel. 1983. Impact of dosing intervals onactivity of gentamicin and ticarcillin against P. aeruginosa ingranulocytopenic mice. J. Infect. Dis. 147:910-917.
11. Gerber, A. U., A. P. Vastola, J. Brandel, and W. A. Craig. 1982.Selection of aminoglycoside-resistant variants of Pseudomonasaeruginosa in an in vivo model. J. Infect. Dis. 146:691-697.
12. Gerber, A. U., P. Wiprachtiger, U. Stettler-Spichiger, and G.Lebek. 1982. Constant infusions versus intermittent doses ofgentamicin against P. aeruginosa in vitro. J. Infect. Dis. 145:554-560.
13. Gilleland, Jr. 1988. Adaptive alterations in the outer membraneof gram-negative bacteria during human infection. Can. J.Microbiol. 34:499-502.
14. Hancock, R. E. W., V. Y. Raffle, and T. I. Nicas. 1981.Involvement of the outer membrane in gentamicin and strepto-mycin uptake and killing in Pseudomonas aeruginosa. Antimi-crob. Agents Chemother. 19:777-785.
15. Ho, P. W. L., F. D. Pieu, and N. Kiminani. 1979. Massiveamikacin "overdose." Ann. Intern. Med. 91:227-228.
16. Jackson, G. G., G. L. Daikos, and V. T. Lolans. 1990. Theinductive role of ionic binding in the bactericidal action and postantibiotic effects of aminoglycoside antibiotics. J. Infect. Dis.162:408-413.
17. Jackson, G. G., and L. J. Riff. 1971. Pseudomonas bacteremia:pharmacologic and other bases for failure of treatment withgentamicin. J. Infect. Dis. 124(Suppl.):S185-S191.
18. Kapusnik, J. E., C. J. Hackbarth, H. F. Chambers, T. Carpen-ter, and M. A. Sande. 1988. Single large daily dosing versusintermittent dosing of tobramycin for treating experimentalPseudomonas pneumonia. J. Infect. Dis. 158:7-12.
19. Kelly, N. M., E. G. Rawling, and R. E. W. Hancock. 1989.Determinants of the efficacy of tobramycin therapy againstisogenic nonmucoid and mucoid derivatives of Pseudomonasaeruginosa PA01 growing in peritoneal chambers in mice.Antimicrob. Agents Chemother. 33:1207-1211.
20. Labovitz, E., M. Levison, and D. Kaye. 1974. Single-dose daily
gentamicin therapy in urinary tract infection. Antibicrob.Agents Chemother. 6:465-470.
21. Ledergerber, B., J. Blaser, and R. Luthy. 1985. Computer-controlled in-vitro simulation of multiple dosing regimens. J.Antimicrob. Chemother. 15(Suppl. A):169-173.
22. Legget, J. E., B. Fantin, S. Ebert, K. Totsuka, B. Vogelman, W.Calame, H. Mattie, and W. A. Craig. 1989. Comparative antibi-otic dose-effect relations at several dosing intervals in murinepneumonitis and thigh-infection models. J. Infect. Dis. 159:281-292.
22a.Leviton, M. A., H. S. Fraimow, N. Carrasco, and M. H. Miller.1989. Program Abstr. 29th Intersci. Conf. Antimicrob. AgentsChemother., abstr. 1311.
22b.Lolans, V. T., A. Said, G. G. Jackson, G. L. Daikos, and D. M.Livermore. 1988. Program Abstr. 28th Intersci. Conf. Antimi-crob. Agents Chemother., abstr. 1501.
23. MacArthur, R. F., V. T. Lolans, F. A. Zar, and G. G. Jackson.1984. Biphasic, concentration-dependent and rate-limited, con-centration-independent bacterial killing by an aminoglycosideantibiotic. J. Infect. Dis. 150:778-779.
24. Moore, R. A., N. C. Bates, and R. E. W. Hancock. 1986.Interaction of polycationic antibiotics with Pseudomonasaeruginosa lipopolysaccharide and lipid A studied by usingdansyl-polymyxin. Antimicrob. Agents Chemother. 29:498-500.
25. Moore, R. D., P. S. Lietman, and C. R. Smith. 1987. Clinicalresponse to aminoglycoside therapy: importance of the ratio ofpeak concentration to minimal inhibitory concentration. J. In-fect. Dis. 155:93-99.
26. Moore, R. D., C. R. Smith, and P. S. Lietman. 1984. Theassociation of aminoglycoside plasma levels with mortality inpatients with gram-negative bacteremia. J. Infect. Dis. 149:443-448.
27. Moore, R. D., C. R. Smith, and P. S. Lietman. 1984. Associationof aminoglycoside plasma levels with therapeutic outcome ingram-negative pneumonia. Am. J. Med. 77:657-662.
28. Nikaido, H., and R. E. W. Hancock. 1885. Outer membranepermeability of Pseudomonas aeruginosa, p. 145-193. In J. R.Sokatch (ed.), The bacteria: a treatise on structure and function,vol. 10. Academic Press, Inc., New York.
29. Noone, P., T. M. C. Parsons, J. R. Pattison, R. C. B. Slack, D.Garfield-Davis, and K. Hughes. 1974. Experience in monitoringgentamicin therapy during treatment of serious gram-negativesepsis. Br. Med. J. 1:477-481.
30. Peterson, A. A., R. E. W. Hancock, and E. J. McGroarty. 1985.Binding of polycationic antibiotics and polyamines to li-popolysacharrarides of Pseudomonas aeruginosa. J. Bacteriol.164:1256-1261.
31. Powell, S. H., W. L. Thompson, M. A. Luthe, R. C. Stern, D. A.Grossinklaus, D. D. Bloxham, D. L. Groden, M. R. E. Jacoabs,A. 0. DiScenna, H. A. Cash, and J. D. Klinger. 1983. Once dailyversus continuous aninoglycoside dosing: efficacy and toxicityin animal and clinical studies of gentamicin, netilmicin andtobramycin. J. Infect. Dis. 147:918-932.
32. Reiner, N. E., D. D. Bloxham, and W. L. Thomson. 1978.Nephrotoxicity of gentamicin and tobramycin given once dailyor continuously in dogs. J. Antimicrob. Chemother. 4(Suppl.):85-101.
33. Schwartz, A. 1988. Overview of the efficacy and safety ofnetilmicin in once daily versus multiple daily dosage regimens.J. Drug Dev. 1(Suppl. 3):147-154.
34. Sturm, A. W. 1989. Netilmicin in the treatment of gram-negativebacteremia: single daily versus multiple daily dosage. J. Infect.Dis. 159:931-937.
35. Vogelman, B., S. Gudmundsson, J. Leggett, J. Turnidge, S.Ebert, and W. A. Craig. 1988. Correlation of antimicrobialpharmacokinetic parameters with therapeutic efficacy in ananimal model. J. Infect. Dis. 158:831-847.
36. Wood, C. A., D. R. Norton, S. J. Kohlhepp, P. W. Kohnem,G. A. Porter, D. C. Houghton, R. E. Brumett, W. M. Bennett,and D. N. Gilbert. 1988. The influence of tobramycin dosageregimens on nephrotoxicity, ototoxicity and antimicrobial effi-cacy in a rat model of subcutaneous abscess. J. Infect. Dis.158:13-22.
VOL. 35, 1991 123
on June 30, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
Related Documents