First assessment of the Caulerpa racemosa (Caulerpales, Chlorophyta) invasion along the French Mediterranean coast Sandrine Ruitton a, * , Fabrice Javel b , Jean-Michel Culioli c , Alexandre Meinesz b , Ge ´rard Pergent d , Marc Verlaque a a UMR 6540 CNRS, Centre d’Oce ´anologie de Marseille, Campus de Luminy, Case 901, 13288 Marseille Cedex 09, France b EA 3156, Gestion de la biodiversite ´, Laboratoire Environnement Marin Littoral, Universite ´ de Nice-Sophia Antipolis, Parc Valrose, 06108 Nice Cedex 02, France c Re ´serve Naturelle des Bouches-de-Bonifacio, Office de l’Environnement de la Corse, Avenue Jean Nicoli, 20250 Corte, France d Equipe E ´ cosyste `mes Littoraux, Universite ´ de Corte, BP 52, 20250 Corte, France Abstract The introduced green alga Caulerpa racemosa var. cylindracea has been rapidly spreading in the Mediterranean Sea since 1990. It was first observed in France in 1997 (Marseilles). In early 2004, the stretch of the French Mediterranean coastline and the surface area affected by the invasion were estimated at about 83 km and 4014 ha, respectively. The depth range of colonized areas was usu- ally 10–35 m depth. Shallow (0–10 m) and deep (down to 40 m) dense meadows were rarely observed. In contrast to the dead matte of Posidonia oceanica, which constituted the most widely colonized substratum, dense P. oceanica meadows and fine sand with large ripple-marks were not invaded. Few rocky areas were colonized and coarse sand bottoms were usually colonized below 20 m depth. All the colonized areas were exposed to human activities and more than 40% were fishing areas. Mild climate, suitable substrata, presence of vectors of dispersal and absence of efficient biological control make the French Mediterranean coast particularly vulnerable to the further spread of the alga. Ó 2005 Elsevier Ltd. All rights reserved. Keywords: Introduced species; Biological invasion; Caulerpa racemosa var. cylindracea; France; Mediterranean Sea; Distribution 1. Introduction Caulerpa racemosa var. cylindracea (Sonder) Verl- aque, Huisman et Boudouresque, a south-western Aus- tralian variety newly introduced into Europe, is currently achieving dramatic and continuous expansion throughout most of the Mediterranean Sea and the Atlantic (Verlaque et al., 2000, 2003, 2004). This inva- sive variety differs from the two other varieties already present in the Mediterranean Sea [C. racemosa var. tur- binate-uvifera, an intermediate variety between var. turbinata (J. Agardh) Eubank and var. uvifera (C. Agardh) J. Agardh, and C. racemosa var. lamourouxii f. requienii (Montagne) Weber-van Bosse] mainly by its upright axes with clavate, more or less inflated branch- lets, which are uncrowded and radially to distichously disposed and its thin rhizoids (Verlaque et al., 2000, 2003). Observed for the first time off the coast of Libya (Nizamuddin, 1991), the introduced C. racemosa var. cylindracea has so far been reported from at least eleven Mediterranean countries (Albania, Croatia, Cyprus, France, Greece, Italy, Libya, Malta, Spain, Tunisia and Turkey), all the major Mediterranean islands (Bale- aric Islands, Corsica, Crete, Cyprus, Sardinia and Sicily) and from the neighbouring areas of the Atlantic (Canary 0025-326X/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2005.04.009 * Corresponding author. Tel.: +33 4 91 829136; fax: +33 4 91 411265. E-mail address: [email protected] (S. Ruitton). www.elsevier.com/locate/marpolbul Marine Pollution Bulletin 50 (2005) 1061–1068

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/marpolbul
Marine Pollution Bulletin 50 (2005) 1061–1068
First assessment of the Caulerpa racemosa (Caulerpales,Chlorophyta) invasion along the French Mediterranean coast
Sandrine Ruitton a,*, Fabrice Javel b, Jean-Michel Culioli c, Alexandre Meinesz b,Gerard Pergent d, Marc Verlaque a
a UMR 6540 CNRS, Centre d’Oceanologie de Marseille, Campus de Luminy, Case 901, 13288 Marseille Cedex 09, Franceb EA 3156, Gestion de la biodiversite, Laboratoire Environnement Marin Littoral, Universite de Nice-Sophia Antipolis, Parc Valrose,
06108 Nice Cedex 02, Francec Reserve Naturelle des Bouches-de-Bonifacio, Office de l’Environnement de la Corse, Avenue Jean Nicoli, 20250 Corte, France
d Equipe Ecosystemes Littoraux, Universite de Corte, BP 52, 20250 Corte, France
Abstract
The introduced green alga Caulerpa racemosa var. cylindracea has been rapidly spreading in the Mediterranean Sea since 1990. It
was first observed in France in 1997 (Marseilles). In early 2004, the stretch of the French Mediterranean coastline and the surface
area affected by the invasion were estimated at about 83 km and 4014 ha, respectively. The depth range of colonized areas was usu-
ally 10–35 m depth. Shallow (0–10 m) and deep (down to 40 m) dense meadows were rarely observed. In contrast to the dead matte
of Posidonia oceanica, which constituted the most widely colonized substratum, dense P. oceanica meadows and fine sand with large
ripple-marks were not invaded. Few rocky areas were colonized and coarse sand bottoms were usually colonized below 20 m depth.
All the colonized areas were exposed to human activities and more than 40% were fishing areas. Mild climate, suitable substrata,
presence of vectors of dispersal and absence of efficient biological control make the French Mediterranean coast particularly
vulnerable to the further spread of the alga.
� 2005 Elsevier Ltd. All rights reserved.
Keywords: Introduced species; Biological invasion; Caulerpa racemosa var. cylindracea; France; Mediterranean Sea; Distribution
1. Introduction
Caulerpa racemosa var. cylindracea (Sonder) Verl-
aque, Huisman et Boudouresque, a south-western Aus-
tralian variety newly introduced into Europe, iscurrently achieving dramatic and continuous expansion
throughout most of the Mediterranean Sea and the
Atlantic (Verlaque et al., 2000, 2003, 2004). This inva-
sive variety differs from the two other varieties already
present in the Mediterranean Sea [C. racemosa var. tur-
0025-326X/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.marpolbul.2005.04.009
* Corresponding author. Tel.: +33 4 91 829136; fax: +33 4 91 411265.
E-mail address: [email protected] (S. Ruitton).
binate-uvifera, an intermediate variety between var.
turbinata (J. Agardh) Eubank and var. uvifera (C.
Agardh) J. Agardh, and C. racemosa var. lamourouxii
f. requienii (Montagne) Weber-van Bosse] mainly by its
upright axes with clavate, more or less inflated branch-lets, which are uncrowded and radially to distichously
disposed and its thin rhizoids (Verlaque et al., 2000,
2003). Observed for the first time off the coast of Libya
(Nizamuddin, 1991), the introduced C. racemosa var.
cylindracea has so far been reported from at least eleven
Mediterranean countries (Albania, Croatia, Cyprus,
France, Greece, Italy, Libya, Malta, Spain, Tunisia
and Turkey), all the major Mediterranean islands (Bale-aric Islands, Corsica, Crete, Cyprus, Sardinia and Sicily)
and from the neighbouring areas of the Atlantic (Canary

1062 S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068
Islands) (Verlaque et al., 2003, 2004; Piazzi et al., in
press).
In France, C. racemosa var. cylindracea was first ob-
served in 1997 in the Gulf of Marseilles (Provence,
France) where a large population was already thriving
in the north of the gulf. The probable year of introduc-tion was estimated to be 1993 or 1994 (Verlaque et al.,
2000) and, by the end of 2004, C. racemosa var. cylindr-
acea had become common in the gulf. Since 1999, other
colonized areas of the French Mediterranean coast have
been discovered in eastern Provence, the French Riviera
and, recently, in Corsica. A similar rapid spread has al-
ready been reported in Italy (Piazzi et al., 1997, 1999,
2001b,c; Buia et al., 1998; Piazzi and Cinelli, 1999; Cec-cherelli et al., 2000; Ceccherelli and Piazzi, 2001).
The spread of C. racemosa var. cylindracea along the
French Mediterranean coast has been monitored since
1997 and the colonized areas inventoried. At a smaller
scale, the colonization of the Gulf of Marseilles has been
studied. The aims of the present study are (1) to provide
a first survey of the colonization of the French Mediter-
ranean coast; (2) to describe the ecological characteris-tics of areas colonized by C. racemosa var. cylindracea;
and (3) to discuss the possible outcome of the invasion.
2. Materials and methods
2.1. The French Mediterranean coast
For the monitoring campaign for the spread of Caul-
erpa racemosa var. cylindracea along the French Medi-
terranean coast, two networks of observers carried out
several hundred SCUBA dives: the ‘‘Observatoire sur
l’expansion des Caulerpes envahisantes en Mediterranee’’
(Provence and French Riviera) and the ‘‘Caulerpa Net-
work’’ (Corsica). The observations were taken all year
round. Date of observation, geographical position,depth (0–10, 11–20, 21–30, 31–40 and 41–50 m), length
of coastline and surface area affected, substratum [rocky
substratum, Posidonia oceanica (L.) Delile meadows,
dead matte of P. oceanica, fine sand or coarse sand
and co-occurrence with the other invasive Caulerpa,
C. taxifolia (Vahl) C. Agardh were noted. The coloniza-
tion levels were estimated according to the standardized
procedure proposed for C. taxifolia by Vaugelas et al.(1999) and Meinesz et al. (2001):
• Level I: area where one or several small colonies
cover a total surface area of less than 1000 m2.
• Level II: area where several colonies cover a total sur-
face greater than 1000 m2 with small colonies dis-
persed over a surface area of less than 10 ha.
• Level III: area with several large colonies greater than1000 m2 dispersed over a surface area greater than
10 ha.
Field data were input into a Geographic Information
System (ArcView microcomputer program, Version 8.3,
Esri�) and the Caulerpa online web database (http://
www.caulerpa.org, Meinesz et al., 2004).C. racemosa var. cylindracea spreads by vegetative
multiplication (cuttings, vegetative propagules) (Cec-
cherelli et al., 2000; Ceccherelli and Piazzi, 2001; Renon-
court and Meinesz, 2002) and sexual reproduction
(Panayotidis and Zuljevic, 2001). Human activities such
as fishing, anchoring and ballast water unloading are
probably the main vectors of remote dispersal of propa-
gules. Thus, in each colonized area, fishing, diving,anchorages and harbour vicinity were noted.
The data were analyzed per area and for the whole
French Mediterranean coast.
2.2. Gulf of Marseilles
The spread of C. racemosa var. cylindracea has been
monitored in the gulf of Marseilles since 1998. Two fieldsurvey techniques are used in SCUBA diving, either pin-
point observation (punctual observation) or transects.
The nature of the substratum, depth and density of C.
racemosa var. cylindracea meadows (0: uncolonized; 1:
sparse individuals with only few creeping horizontal
axes; 2: small patches less than 1 m in diameter; 3:
patches more than 1m in diameter; 4: continuous mead-
ows with only one layer of stolons and 5: continuousmeadows with a thick layer of stolons) are noted. The
estimation of the colonized areas in 1998 and 2004
was performed using kriging as gridding method. The
kriging method uses spherical functions as a variogram
model (Surfer microcomputer program, Version 8,
Golden Software�).
3. Results
3.1. Distribution of C. racemosa var. cylindracea
meadows along the French Mediterranean coast
In early 2004, C. racemosa var. cylindracea colonized
26 distinct areas in Provence and the French Riviera and
3 in Corsica (Fig. 1 and Table 1). The number of colo-nized areas has increased, in particular since 2002 (Table
1). Field data and detailed maps are available on the
Caulerpa online web site. The increase in the length of
coastline and surface area affected by the invasion of
C. racemosa var. cylindracea is given in Table 2. In early
2004, the length of coastline and surface area affected by
C. racemosa var. cylindracea reached 82.9 km and
4014 ha respectively (Table 2). C. racemosa var. cylindr-acea co-occurred with C. taxifolia in three large bays:
Villefranche-sur-Mer, Hyeres and Toulon.
In western Provence (Marseilles), C. racemosa var.
cylindracea presented a strongly marked annual cycle

Fig. 1. Map of Caulerpa racemosa colonized areas along the French Mediterranean coast. Numbers correspond to areas colonized by Caulerpa
racemosa according to Table 1. Level of colonization according to Vaugelas et al.�s classification (1999).
S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068 1063
of development with a growth period from late spring to
early autumn, a decay period in late autumn and early
winter, and a resting period in late winter and early
spring during which only some very inconspicuous small
fragments scattered on the substrate were present. Con-
versely, in eastern Provence and the French Riviera, theseasonal decline of C. racemosa var. cylindracea mead-
ows was limited to a more or less complete fall of erect
axes without disappearance of the stolons. Field data in
Corsica were insufficient to establish a seasonal cycle.
3.2. Characteristics of the colonized areas
Although observed down to 50 m depth, the usualdepth range of C. racemosa var. cylindracea was 10–
35 m depth. Shallow (0–10 m) and deep (down to
40 m) dense meadows were rare (Table 1, Fig. 2). How-
ever in the vicinity of large and dense colonized areas, C.
racemosa var. cylindracea was usually present, although
very discrete in shallow waters.
The dead matte of P. oceanica was the most fre-
quently colonized substratum (Table 1, Fig. 2). OnlyP. oceanica meadows with low density of shoots were
colonized. Dense P. oceanica meadows and the soft sub-
strata under strong currents (fine sand with large ripple-
marks) were not colonized. A few rocky areas, above all
those constituted of boulders surrounded by dead matte
of P. oceanica or sand, were colonized. Development on
vertical rocky substratum was rarely observed. Coarse
sand areas were usually colonized below 20 m depth
(Table 1).
Most of the colonized areas were exposed to human
activities and more than 40% were fishing areas (Table
1, Fig. 2). Moreover, most of the first settlements were
found in the vicinity of fishing areas, anchoring areasor harbours (Table 1).
3.3. The gulf of Marseilles
As in other French Mediterranean areas, at Mar-
seilles Caulerpa racemosa var. cylindracea mainly colo-
nized the P. oceanica dead matte between 5 and 30 m
depth, and coarse sand below 20 m depth. The cumu-lated field data gave a gross estimation of colonized sur-
face area in the Gulf of Marseilles and vicinity of
1120 ha in early 2004. By the kriging method, the esti-
mation of colonized surface area in 1998 and 2004 was
900 ha and 3180 ha, respectively, corresponding to an
increase of more than 350% in six years (Fig. 3).
4. Discussion
4.1. Distribution and depth range along the French
Mediterranean coast
The spread of C. racemosa var. cylindracea along
the French Mediterranean coast was faster and more

Table 1
Characteristics of areas colonized by Caulerpa racemosa on French Mediterranean coast (from east to west)
No. Areas Date of first
observation
Coastline (m) Depth Substratum Level Human
activities
1 St-Jean-Cap-Ferrat, south of Pointe
de Crau de Nao
Sep-2003 2 6 DM/FS I F/A/D
2 St-Jean-Cap-Ferrat, bay of
Villefranche-sur-Mer
Sep-1999 6500 1–45 RS/PO/DM/ FS/CS III F/A/D/H
3 Nice, Coco Beach May-2003 2 3 RS I –
4 St-Tropez, west of the harbour Nov-2002 170 12–14 DM/FS I F/A/H
5 La Londe, Hyeres, north of
the bay of Hyeres
Sep-2003 4300 18–25 PO/DM III F/A/H
6 Hyeres-les-Palmiers, south-east
of the Gapeau mouth
Jun-2003 200 13–16 PO/DM I F
7 Hyeres-les-Palmiers, centre of
the bay of Hyeres
Sep-2002 2600 20–32 PO/DM/CS III F/A/H
8 Port-Cros island, Pointe du Vaisseau Jul-2001 15 20–28 PO/DM I F/D
9 Port-Cros island, west of the bay of Port-Cros Oct-2002 6 18 DM I F
10 Port-Cros island, Gabiniere islet md md 26–36 RS md D
11 Porquerolles island, south-west
of Cap des Medes
Feb-1999 15 12 DM I D
12 Porquerolles island, Pointes du Gros
Baou and Galeasson
Jul-2001 40 20–25 PO/FS I F
13 Porquerolles island, south-west of the
Pointe du Galeasson
Jan-2001 20 12 DM I F/A
14 Porquerolles island, Langoustier Shoal Aug-2003 100 20–35 CS II F/D
15 West of Giens peninsula, Fourmigue islet Jul-2002 20 35 CS I F/D
16 Hyeres-les-Palmiers, gulf of Giens Nov-2003 1700 28–35 DM/FS III F/A
17 From Carqueiranne (Salettes harbour)
to the bay of Toulon
May-2000 22,200 1–40 RS/PO/DM/FS/CS III F/D/A/H
18 Marseilles, south of the Calanque de l�Escu Nov-2003 30 48 CS I F
19 Marseilles, north of Plane island Oct-2003 30 27 DM/CS I F/D
20 Marseilles, Plateau des Chevres Oct-2003 30 30 CS I F
21 Marseilles, Planier island Aug-2003 1 28 RS I F/D
22 Marseilles, from Callelongue to
Pointe Rouge harbour
Jan-1998 8500 3–50 RS/PO/DM/CS III F/A/D/H
23 Marseilles, bay of Prado Jan-1999 6200 4–42 DM III F/A/H
24 Marseilles, Estaque Nov-1997 500 12–22 RS/DM/FS II F/A/H
25 Marseilles, Pointe of Corbiere Dec-1998 150 5–15 DM II F/H
26 Le Rove, from Cap Ragnon
to Niolon harbour
Jan-1998 1600 7–20 RS/DM/FS II F/D/A
27 Bastia, from Golo mouth to Pietra Nera 2002 25,000 32–45 DM/CS III F/A
28 Bonifacio, gulf of Sant�Amanza Nov-2002 3000 11–41 PO/DM/FS III A
29 Propriano, gulf of Valinco Jun-2004 500 15–25 DM/FS II F/H
Numbers (No.) correspond to Fig. 1. Level of colonization according to Vaugelas et al.�s classification (1999); coastline affected; md = missing data.
Substratum: RS: rocky substratum; PO: Posidonia oceanica meadow with low shoot density; DM: dead matte of P. oceanica; FS: fine sand; CS:
coarse sand. Human activities in the area: F: Fishing; D: Diving; A: Anchorage; H: Harbour vicinity. All the detailed information about the
colonized zone is available on the web site www.caulerpa.org (Meinesz et al., 2004).
1064 S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068
widespread than that of C. taxifolia, which has not yet
been sighted at Marseilles or in Corsica (Meinesz
et al., 2004). Since the first sighting in 1984, the coastline
affected by the C. taxifolia invasion had reached 22.0 km
in 1994, 77.3 km in 2001 and 122.7 km in 2004 (Meinesz
et al., 2004). In comparison, the coastline affected by the
C. racemosa var. cylindracea invasion reached 82.9 km
after only 10 years of colonization (if we consider itsprobable introduction at Marseilles in 1994; Verlaque
et al., 2000). At Marseilles, the colonized area increased
by 350% in six years, which is less than in Italy where a
similar increase was observed in only one year (Piazzi
et al., 2001c). However, this difference could be related
to the mapping scale.
Depth range distribution of C. racemosa var. cylindr-
acea appears to be more restricted along the continental
French Mediterranean coast than in the southern and
eastern Mediterranean where the species was observed
down to 70 m (Southern Italy and Cyprus) (Piazzi
et al., in press). In the present study, although C. race-
mosa var. cylindracea was observed down to 50 m depth
(Marseilles), the areas colonized were usually at between
10 and 35 m depth. The poor development of C. race-
mosa var. cylindracea close to the sea surface is probably
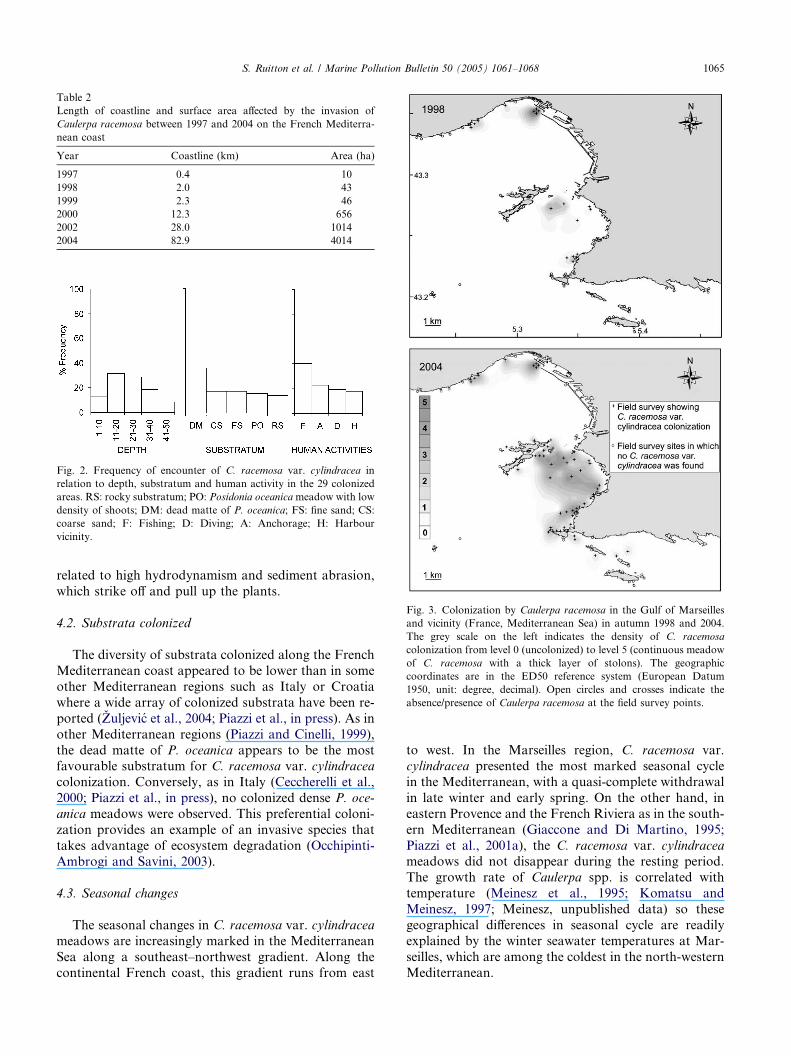
Table 2
Length of coastline and surface area affected by the invasion of
Caulerpa racemosa between 1997 and 2004 on the French Mediterra-
nean coast
Year Coastline (km) Area (ha)
1997 0.4 10
1998 2.0 43
1999 2.3 46
2000 12.3 656
2002 28.0 1014
2004 82.9 4014
Fig. 2. Frequency of encounter of C. racemosa var. cylindracea in
relation to depth, substratum and human activity in the 29 colonized
areas. RS: rocky substratum; PO: Posidonia oceanicameadow with low
density of shoots; DM: dead matte of P. oceanica; FS: fine sand; CS:
coarse sand; F: Fishing; D: Diving; A: Anchorage; H: Harbour
vicinity.
Fig. 3. Colonization by Caulerpa racemosa in the Gulf of Marseilles
and vicinity (France, Mediterranean Sea) in autumn 1998 and 2004.
The grey scale on the left indicates the density of C. racemosa
colonization from level 0 (uncolonized) to level 5 (continuous meadow
of C. racemosa with a thick layer of stolons). The geographic
coordinates are in the ED50 reference system (European Datum
1950, unit: degree, decimal). Open circles and crosses indicate the
absence/presence of Caulerpa racemosa at the field survey points.
S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068 1065
related to high hydrodynamism and sediment abrasion,
which strike off and pull up the plants.
4.2. Substrata colonized
The diversity of substrata colonized along the French
Mediterranean coast appeared to be lower than in some
other Mediterranean regions such as Italy or Croatia
where a wide array of colonized substrata have been re-
ported (Zuljevic et al., 2004; Piazzi et al., in press). As in
other Mediterranean regions (Piazzi and Cinelli, 1999),
the dead matte of P. oceanica appears to be the mostfavourable substratum for C. racemosa var. cylindracea
colonization. Conversely, as in Italy (Ceccherelli et al.,
2000; Piazzi et al., in press), no colonized dense P. oce-
anica meadows were observed. This preferential coloni-
zation provides an example of an invasive species that
takes advantage of ecosystem degradation (Occhipinti-
Ambrogi and Savini, 2003).
4.3. Seasonal changes
The seasonal changes in C. racemosa var. cylindracea
meadows are increasingly marked in the Mediterranean
Sea along a southeast–northwest gradient. Along the
continental French coast, this gradient runs from east
to west. In the Marseilles region, C. racemosa var.
cylindracea presented the most marked seasonal cycle
in the Mediterranean, with a quasi-complete withdrawal
in late winter and early spring. On the other hand, in
eastern Provence and the French Riviera as in the south-
ern Mediterranean (Giaccone and Di Martino, 1995;
Piazzi et al., 2001a), the C. racemosa var. cylindraceameadows did not disappear during the resting period.
The growth rate of Caulerpa spp. is correlated with
temperature (Meinesz et al., 1995; Komatsu and
Meinesz, 1997; Meinesz, unpublished data) so these
geographical differences in seasonal cycle are readily
explained by the winter seawater temperatures at Mar-
seilles, which are among the coldest in the north-western
Mediterranean.

1066 S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068
4.4. Competition with Caulerpa taxifolia
At Toulon and Hyeres, the dense meadows have a
different depth range distribution: C. taxifolia meadows
thrive between 2 and 15 m depth and C. racemosa var.
cylindracea meadows between 20 and 30 m depth (Bel-sher et al., 2003). At Villefranche-sur-Mer (France), at
16–20 m in depth, C. taxifolia almost completely elimi-
nated C. racemosa var. cylindracea over a wide area
(ca. 1 ha) in only one year (Meinesz, unpublished data).
Conversely, near Livorno (Italy), in mixed beds (Piazzi
et al., 2003) and through manipulative experiments
(Piazzi and Ceccherelli, 2002), C. racemosa var. cylindr-
acea showed higher competitive ability than C. taxifolia.In the Mediterranean Sea, C. taxifolia is taller than C.
racemosa var. cylindracea (Meinesz and Hesse, 1991)
and has a less marked seasonal cycle, a higher stolon
growth rate and a higher annual cumulated growth
(Komatsu and Meinesz, 1997; Ruitton et al., in press-
b; Capiomont et al., 2005). However, C. racemosa var.
cylindracea could have a higher capacity to occupy the
substratum than C. taxifolia, because the length of sto-lons and number of apices per square meter are higher
in C. racemosa var. cylindracea meadows (Meinesz and
Hesse, 1991; Meinesz et al., 1995; Ruitton et al., in
press-b; Capiomont et al., 2005). More investigations
are needed for a better understanding of the competition
between the two Caulerpa.
4.5. Human activities and spread vectors
Several potential natural and anthropic vectors of
dispersal of C. racemosa var. cylindracea have been
clearly identified in the Mediterranean Sea. They give
rise to two distance scales of spread:
(1) Long to medium distance (>1 km): zygotes trans-
ported by currents or ballast waters; cuttings andpropagules carried with ballast waters, ship
anchors and fishing equipment (nets, fish pots
and trawls) (Magri et al., 2001; Zuljevic et al.,
2004).
(2) Short distance (<1 km): zygotes, cuttings and
propagules transported by water movements, mac-
rophytes and benthic invertebrates or disseminated
by ship anchors and fishing equipment (Ceccherelliand Piazzi, 2001; Ceccherelli et al., 2001; Renon-
court and Meinesz, 2002; Zuljevic et al., 2004).
Like C. taxifolia, cuttings of C. racemosa var. cylindr-
acea could probably survive for several days in cold, wet
anchor chain lockers and among fishing equipment, so
transport by ships might generate long or medium spread
depending on vessel movements. These vectors are high-lighted by the fact that more than 40% of colonized areas
along the French Mediterranean coast were fishing areas
and most of first settlements were found in the vicinity of
fishing areas, anchoring areas or harbours.
4.6. Accuracy of the mapping
The present study showed that the standardizedmethod used to describe theC. taxifolia invasion can also
be used to monitor the spread of C. racemosa var.
cylindracea. However, the estimation of colonized sur-
face area from the cumulated field data gave a much
lower value than those obtained by kriging method (Gulf
of Marseilles). Consequently, the total gross estimation
for the whole French coastline (4014 ha) probably under-
estimates the true colonized area. In fact, the mapping ofC. racemosa var. cylindracea meadows is more difficult
than C. taxifolia. Conversely to C. taxifolia which has
a mainly contagious pattern of colonization from vegeta-
tive fragments fallen to the bottom (Hill and Coquillard,
1999), C. racemosa var. cylindracea has a diffuse pattern
of colonization that is very difficult to map with the cur-
rent methods (meadows with complex limits). The kri-
ging method, which is probably a good method formapping the diffuse populations could be improved in
the future for example by mapping the dead matte of
P. oceanica, and by increasing the sampling effort.
5. Conclusion
The rapid spread of C. racemosa var. cylindracea inthe Mediterranean Sea has few equivalents in other
introduced marine macrophytes (Verlaque et al., 2004).
Since its discovery at Marseilles, C. racemosa var.
cylindracea quickly spread along the French Mediterra-
nean coast where it found suitable conditions for devel-
opment. Despite a greater winter withdrawal than in
southern areas, probably related to the colder winter
seawater temperatures of the north western basin, largeC. racemosa var. cylindracea meadows already thrive in
several major bays of the littoral. Moreover, to date, all
maps probably underestimate the true colonized area.
In conclusion, the French Mediterranean coast fea-
tures a wide range of favourable characteristics for the
development of C. racemosa var. cylindracea a climate
similar to its native region (south-western Australia;
Verlaque et al., 2003), large areas of dead matte of P.oceanica, numerous vectors of spread in particular
human activities and no efficient biological enemy
(Ruitton et al., in press-a). Consequently, the further
spread of C. racemosa var. cylindracea is to be expected.
Acknowledgements
This work was supported by Rhone—Mediterra-
nee—Corse Water Agency, the Provence-Alpes-Cotes

S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068 1067
d�Azur Region, the DIREN PACA (Direction Regio-
nale de l�Environnement Provence-Alpes-Cote d�Azur),
the Environment Office of Corsica and ALIENS (‘‘Algal
Introductions to European Shores’’), a 5th Framework
Programme of the European Union.The authors are in-
debted to Michael Paul for correcting the English. Theauthors also thank Roland Graille, Frederique Zuberer,
Bernard De Ligondes and Eric Pironneau for field
assistance.
References
Belsher, T., Lunven, M., Le Gall, E., Caisey, X., Dugornay, O.,
Mingant, C., 2003. Observations concerning the expansion of
Caulerpa taxifolia and Caulerpa racemosa in Rade d�Hyeres and
Rade of Toulon (France). Oceanologica Acta 26, 161–166.
Buia, M.C., Petrocelli, A., Saracino, O.D., 1998. Caulerpa racemosa
spread in the Mediterranean Sea. First record in the Gulf of
Taranto. Biologia Marina Mediterranea 5, 527–529.
Capiomont, A., Breugnot, E., Den Haan, M., Meinesz, A., 2005.
Phenology of a deep-water population of Caulerpa racemosa var.
cylindracea in the North-western Mediterranean Sea. Botanica
Marina 48, 80–83.
Ceccherelli, G., Piazzi, L., 2001. Caulerpa racemosa e Caulerpa
taxifolia in Mediterraneo: effetto della complessita dei popolamenti
macroalgali autoctoni e dei loro rapporti competitivi. 32� Con-
gresso SIBM, 45.
Ceccherelli, G., Piazzi, L., Cinelli, F., 2000. Response of non-
indigenous Caulerpa racemosa (Forsskal) J. Agardh to the native
seagrass Posidonia oceanica (L.) Delile: effect of density of shoots
and orientation of edges of meadows. Journal of Experimental
Marine Biology and Ecology 243, 227–240.
Ceccherelli, G., Campo, D., Piazzi, L., 2001. Some ecological aspects
of the introduced alga Caulerpa racemosa in the Mediterranean:
way of dispersal and impact on native species. Biologia Marina
Mediterranea 8, 94–99.
Giaccone, G., Di Martino, V., 1995. La vegetazione a Caulerpa
racemosa (Forsskal) J. Agardh nella Baia di S. Panagia (Sicilia Sud-
Orientale). Bollettino Accademia Gioenia Scienze Naturali Catania
28 (349), 59–73.
Hill, D., Coquillard, P., 1999. Modelling and simulating Caulerpa
taxifolia (Vahl) C. Agardh in the north-western Mediterranean Sea:
results and perspectivesProceedings of the Workshop on Invasive
Caulerpa in the Mediterranean, Heraklion, Greece. UNEP pub-
lisher, Athens, Greece, pp. 159–174.
Komatsu, T., Meinesz, A., 1997. Temperature and light responses of
alga Caulerpa taxifolia introduced into the Mediterranean Sea.
Marine Ecology Progress Series 146, 145–153.
Magri, M., Piazzi, L., Serena, F., 2001. La presence de Caulerpa
racemosa le long des cotes septentrionales de la Toscane et les
consequences possibles sur l�activite de peche. In: Gravez, V.,
Ruitton, S., Boudouresque, C.F., Le Direac�h, L., Meinesz, A.,
Scabbia, G., Verlaque, M. (Eds.), Fourth International Work-
shop on Caulerpa Taxifolia. GIS Posidonie publ., Marseilles,
France, pp. 338–344.
Meinesz, A., Hesse, B., 1991. Introduction et invasion de l�alguetropicale Caulerpa taxifolia en Mediterranee nord-occidentale.
Oceanologica Acta 14, 415–426.
Meinesz, A., Benichou, L., Blachier, J., Komatsu, T., Lemee, R.,
Molenaar, H., Mari, X., 1995. Variations in the structure,
morphology and biomass of Caulerpa taxifolia in the Mediterra-
nean Sea. Botanica marina 38, 499–508.
Meinesz, A., Belsher, T., Thibaut, T., Antolic, B., Ben Mustapha, K.,
Boudouresque, C.F., Chiaverini, D., Cinelli, F., Cottalorda, J.M.,
Djellouli, A., El Abed, A., Orestano, C., Grau, A.M., Ivesa, L.,
Jaklin, A., Langar, H., Massuti-Pascual, E., Peirano, A., Tunesi,
L., Vaugelas, J. de, Zavodnik, N., Zuljevic, A., 2001. The
introduced green alga Caulerpa taxifolia continues to spread in
the Mediterranean. Biological Invasions 3, 201–210.
Meinesz, A., Javel, F., Cottalorda, J.M., Garcia, D., 2004. Caulerpa
online. World-wide electronic publication. Laboratoire Environn-
ement Marin Littoral, Universite de Nice Sophia-Antipolis. http://
www.caulerpa.org (10.xi.2004).
Nizamuddin, M., 1991. The Green Marine Algae of Libya. Elga, Bern.
Occhipinti-Ambrogi, A., Savini, D., 2003. Biological invasions as a
component of global change in stressed marine ecosystems. Marine
Pollution Bulletin 46, 542–551.
Panayotidis, P., Zuljevic, A., 2001. Sexual reproduction of the invasive
green alga Caulerpa racemosa var. occidentalis in the Mediterra-
nean sea. Oceanologica acta 24, 199–203.
Piazzi, L., Ceccherelli, G., 2002. Effects of competition between two
introduced Caulerpa. Marine Ecology Progress Series 225, 189–
195.
Piazzi, L., Cinelli, F., 1999. Developpement et dynamique saisonniere
d�un peuplement mediterraneen de l�algue tropicale Caulerpa
racemosa (Forsskal) J. Agardh. Cryptogamie, Algologie 20, 295–
300.
Piazzi, L., Balestri, E., Magri, M., Cinelli, F., 1997. Expansion de
l�algue tropicale Caulerpa racemosa (Forsskal) J. Agardh (Bryop-
sidophyceae, Chlorophyta) le long de la cote Toscane (Italie).
Cryptogamie, Algologie 18, 343–350.
Piazzi, L., Balata, D., Cinelli, F., 1999. Mapping to monitor the spread
of introduced green alga Caulerpa racemosa in the north-western
Mediterranean. In: Fifth International Conference on Ecology of
Invasive Alien Plants, 93.
Piazzi, L., Ceccherelli, G., Cinelli, F., 2001a. Expansion de Caulerpa
taxifolia et de Caulerpa racemosa le long des cotes toscanes (Italie),
situation en 1998. In: Gravez, V., Ruitton, S., Boudouresque, C.F.,
Le Direach, L., Meinesz, A., Scabbia, G., Verlaque, M. (Eds.), 4th
International Workshop on Caulerpa Taxifolia, Lerici, 1–2 Febru-
ary 1999. GIS Posidonie Publisher, pp. 71–77.
Piazzi, L., Ceccherelli, G., Balata, D., Cinelli, F., 2001b. Description
des peuplements a Caulerpa taxifolia et a Caulerpa racemosa a
Calafuria (Livorno, Italie). In: Gravez, V., Ruitton, S.,
Boudouresque, C.F., Le Direac�h, L., Meinesz, A., Scabbia, G.,
Verlaque, M. (Eds.), Fourth International Workshop on
Caulerpa Taxifolia. GIS Posidonie publ., Marseilles, France, pp.
345–348.
Piazzi, L., Balata, D., Ceccherelli, G., Cinelli, F., 2001c. Comparative
study of the growth of the two co-occuring introduced green algae
Caulerpa taxifolia and Caulerpa racemosa along the Tuscan coast
(Italy, western Mediterranean). Cryptogamie, Algologie 22, 459–
466.
Piazzi, L., Balata, D., Cecchi, E., Cinelli, F., 2003. Co-occurrence of
Caulerpa taxifolia and C. racemosa in the Mediterranean Sea:
interspecific interactions and influence on native macroalgal
assemblages. Cryptogamie, Algologie 24, 233–243.
Piazzi, L., Meinesz, A., Verlaque, M., Akcali, B., Antolic, B., Argyrou,
M, Balata, D., Ballesteros, E., Calvo, S., Cinelli, F., Cirik, S.,
Cossu, A., D�Archino, R., Djellouli, A.S., Javel, J., Lanfranco, E.,
Mifsud, C., Pala, D., Panayotidis, P., Peirano, A., Pergent, G.,
Petrocelli, A., Ruitton, S., Zuljevic, A., Ceccherelli, G. in press.
Invasion of Caulerpa racemosa var. cylindracea (Caulerpales,
Chlorophyta) in the Mediterranean Sea: an assessment of the
spread. Cryptogamie, Algologie.
Renoncourt, L., Meinesz, A., 2002. Formation of propagules on an
invasive strain of Caulerpa racemosa (Chlorophyta) in the Medi-
terranean sea. Phycologia 41, 533–535.

1068 S. Ruitton et al. / Marine Pollution Bulletin 50 (2005) 1061–1068
Ruitton S., Verlaque M., Aubin G., Boudouresque C.F. in
press-a. Grazing of the introduced Caulerpa racemosa var.
cylindracea (Caulerpales, Chlorophyta) in Mediterranean sea by
herbivorous fish and sea urchins. Vie et Milie—Life &
Environment.
Ruitton, S., Verlaque, M., Boudouresque, C.F. in press-b. Seasonal
changes of the introduced Caulerpa racemosa var. cylindracea
(Caulerpales, Chlorophyta) at the northwest limit of its Mediter-
ranean range. Aquatic Botany.
Vaugelas, J. de, Meinesz, A., Antolic, B., Ballesteros, E., Belsher, T.,
Cassar, N., Ceccherelli, G., Cinelli, F., Cottalorda, J.M., Frada
Orestano, C., Grau, A.M., Jaklin, A., Morucci, C., Relini, M.,
Sandulli, R., Span, A., Tripaldi, G., Van Klaveren, P., Zavodnik,
N., Zuljevic, A., 1999. Standardization proposal for the mapping of
Caulerpa taxifolia expansion in the Mediterranean Sea. Oceano-
logica Acta 22, 85–94.
Verlaque, M., Boudouresque, C.F., Meinesz, A., Gravez, V., 2000.
TheCaulerpa racemosa complex (Caulerpales, Ulvophyceae) in the
Mediterranean Sea. Botanica marina 43, 49–68.
Verlaque, M., Durand, C., Huisman, J.M., Boudouresque, C.F., Le
Parco, Y., 2003. On the identity and origin of the Mediterranean
invasive Caulerpa racemosa (Caulerpales, Chlorophyta). European
Journal of Phycology 38, 325–339.
Verlaque, M., Afonso-Carrillo, J., Gil-Rodriguez, M.C., Durand, C.,
Boudouresque, C.F., Le Parco, Y., 2004. Blitzkrieg in a marine
invasion: Caulerpa racemosa var. cylindracea (Bryopsidales, Chlo-
rophyta) reaches the Canary Islands (NE Atlantic). Biological
Invasions 6, 269–281.
Zuljevic, A., Antolic, B., Despalatovic, M., Onofri, V., 2004. The
spread of the invasive variety of Caulerpa racemosa in the Adriatic
Sea. Rapport Commission Internationale Exploration Scientifique
Mer Mediterranee 37, 466.
Related Documents











