Final Report “Genetic enhancement of Dolichos bean through integration of conventional breeding and molecular approaches” Sponsored by The Kirkhouse Trust, United Kingdom Project Leader Dr. M. Byre Gowda Sr. Scientist (Plant Breeding) AICRP on Pigeonpea, GKVK, Bangalore – 560065. University of Agricultural Sciences GKVK Campus, Bangalore – 560065

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Final Report
“Genetic enhancement of Dolichos bean through integration of conventional
breeding and molecular approaches”
Sponsored by The Kirkhouse Trust,
United Kingdom
Project Leader Dr. M. Byre Gowda
Sr. Scientist (Plant Breeding) AICRP on Pigeonpea,
GKVK, Bangalore – 560065.
University of Agricultural Sciences
GKVK Campus, Bangalore – 560065

Final Report
Dolichos bean, Lablab purpureus L. Sweet (2n = 22) belongs to the family Fabaceae and is one of
the most ancient crops among cultivated plants. It is presently grown through out the tropical regions in Asia
and Africa. It is cultivated either as a pure crop or mixed with other crops, such as finger millet, groundnut,
castor, corn, bajra or sorghum in tropical region. It is a multipurpose crop grown for pulse, vegetable and
forage. The crop is grown for its green pods, while dry seeds are used in various vegetable food
preparations. It is also grown in home gardens as annual crop or on fences as perennial crop. It is one of the
major sources of protein in the diets in southern states of India. The consumer preference varies with pod
size, shape, colour and aroma (pod fragrance).
Within India, Lablab as a field crop mostly confined to the peninsular region and cultivated to a large
extent in Karnataka and adjoining districts of Tamil Nadu, Andhra Pradesh and Maharashtra. Karnataka
contributes a major share, accounting for nearly 90 per cent in terms of both area and production in the
country. Karnataka state records production of about 18,000 tonnes from an area of 85,000 hectares. Outside
India, the crop is cultivated in East Africa, with similar uses, and in Australia as a fodder crop.
The efforts of improving the crop utilizing indigenous and exotic germplasm have been useful in
breaking the yield barriers to a small extent resulting in a compact plant type, reduced duration and photo-
insensitive types. Despite the many good attributes, the crop has remained unexploited owing to low
productivity, long duration, photosensitivity and indeterminate growth habit. Hence comprehensive
germplasm collection and evaluation together with development of suitable genotypes with desirable traits
for a pure crop and investigation of its value as an inter-crop with other food and forage crops are essential.
To exploit these resources, a combination of genetic and phenotypic data is required to assist conventional
breeding programmes to achieve the targeted aims efficiently. With all this background information, the
following objectives are formulated with time-linked outcomes.
1. Collection of germplasm and phenotypic characterization of morpho-agronomic traits with emphasis
on resistance to pod borers and bruchids.
2. To develop high yielding genotypes having suitable plant type for mono and intercropping systems.
3. Molecular characterization and identification of marker/trait associations for marker assisted selection
to develop varieties for desirable traits.
4. Testing and validation of advanced breeding lines for improved traits.
I. Recruitment of staff
The following persons were worked against the sanctioned posts. The details are as follows.
Sl No
Name Job Title Salary Date of Joining
Date of leaving
1 Dr. G. Girish Res. Associate Rs.18, 000/- 01/03/2006 12-03-2009 2 Mr. Ganapathy SRF Rs. 14,000/- 01/02/2008 30-06-2008 3 Miss Nagrathnamma SRF Rs. 14,000/- 19/07/2008 31-01-2009 4 Mr. Suresha Field Assistant Rs. 5000/- 01/03/2006 31-01-2009 5 Mr. Subramanya Lab Assistant Rs 5000/- 01/09/2006 28-02-2009 6 Mr. Lingaraju Driver Rs. 5000/- 23/05/2006 28-02-2009
II. Purchase of Vehicle
A New vehicle was purchased on 3rd May 2006 for Rs. 7, 05,067.
The vehicle was used for collection programmes; conduct of multilocation
trails at different locations and in farmers’ fields, purchase of inputs and for
training programmes.
III. Construction of Seed Storage building
Opening ceremony of the seed storage building was held on 09.01.2008. Building was inaugurated
by Sir Edwin Southern and Madam Dr. Sonia Morgan and Presided by Dr. P .G. Chengappa, Vice
chancellor, UAS, Bangalore. The ceremony was attended by Dr. H. S. Sharathchandra, Emeritus Professor,
Human Genome Project, IISC, Bangalore, Dr. T. K. Prabhakara Setty, Director of Research, Dr. T. K. S.
Gowda, Director of Post graduate studies and Dr. R. Govindan, Director of Instruction (Agri) and other
dignitaries, students and staff of the university.
Two Freezers (-20) were purchased and placed in the building for storing of seeds. We are storing
seeds kept in seed boxes at – 20 temperatures.

c. Visitors during the project period
1. Sir Edwin Southern
2. Dr. Sonia Morgan
3. Dr. Robert Kobner
4. Dr. H. S. Sharathchandra
5. Dr. Noel Ellis
6. Dr. Maggie Knox
7. Dr. Brigette Mass
8. Dr. Bruce Pengelly
9. Dr. Miriam Kinyua
III. Technical Progress:
1. Collection and Maintenance of germplasm lines
Germplasm collections maintained No.1 Local collections 3742 Exotic types 113 Collections received from UAS, Dharwad 554 Kenya collections 45 Total 485
The following are the exotic germplasm lines maintained after rejuvenation.
Sl. Gl lines Sl. Gl lines 1 CPI 52544 7 Rangoi 2 CPI 36903 8 CPI 76977 3 CPI 13700 9 CPI 51564 4 CPI 31113 10 CPI 60126 5 CPI 52508 11 High Worth 6 CPI 24973

2. Development of Descriptor list of Dolichos The descriptor list developed at Bangalore was used for cataloguing of germplasm.
During the first year of the project, characterization of germplasm was done for various
morphological characters as per the Dolichos descriptor list developed at Bangalore based on AVRDC data and local knowledge. This was also shared with Dr. Ravikumar, UAS, Dharwad and Dr. Miriam Kinyua, Kenya. The descriptor developed at Bangalore is presented below.







3. Evaluation of germplasm for qualitative and quantitative traits.
All the collected accessions during the previous years were evaluated during 2007 and 2008. For
evaluation purpose, germplasm was grouped as indeterminates, semi determinates, pendal types and wild
types and were grown separately. All the accessions were characterized with respect to qualitative and
evaluated for quantitative traits.
a. Variability for qualitative characters in the germplasm accessions
Sl. Character Type No. of Germplasm White ( 1) 80 Green ( 2) 322
1 Emerging cotyledon colour
Purple ( 3) 0 Green ( 1) 402 2. Hypocotyl colour Purple (2) 0 No pigmentation (0) 314 Localized to node (3) 47 Extensive (5) 29
3. Stem pigmentation
Almost solid ( 7) 12 Green (1) 363 4. Main colour Purple (2) 39 Absent ( 0) 402 5. Leaf anthocyanin Present ( 2) 0 Pale green (1) 171 Green (3) 213 Dark green (5) 189 Purple (7) 0
6. Leaf colour
Dark Purple (0) 0 Glabrous (0) 158 Low pubescent (3) 230 Moderately pubescent ( 5) 14
7 Leaf hairiness (on inner surface )
Highly pubescent (7) 0 Round (1) 383 Ovate (3) 9 Ovate Lanceolate (5) 10 Lanceolate (7) 0
8 Leaf shape
Linear lanceolate (9) 0 Few leaves remaining (3) 361 9 Leaf persistence Intermediate (5) 41

Table continued..
Sl. Character Type No. of Germplasm Determinate (1) 16 10 Growth habit Semi determinate (2) 16
Indeterminate (3) 370 Short and erect lateral branches (3) 22 Branches tending to be perpendicular to main stem, medium in length ( 5)
306 11 Branch orientation
First lateral branches long and spreading over ground (7)
74
White (1) 26 Cream (2) 301 Light yellow (3) 10 Pink (4) 52
12 Flower bud colour
Purple (5) 13 White (1) 312 Cream (2) 25 Light yellow (3) 0 Pink (4) 52
13 Standard petal colour
Purple (5) 13 White (1) 318 Cream (2) 19 Light yellow (3) 0 Pink (4) 53
14 Wing petal colour
Purple (5) 13 White (1) 316 Cream (2) 21 Light yellow (3) 0 Pink (4) 51
15 Keel petal colour
Purple (5) 10 Within foliage (3) 32 Intermediate (5) 314
16 Raceme position
Emerging from leaf canopy 56 Straight (0) 172 Slightly curved (3) 133
17 Pod curvature
Curved (5) 97 Glabrous (0) 65 Moderately pubescent (3) 322
18 Pod pubescence
Pubescent (5) 15 Short beak (1) 79 Medium length (2) 27 Long beak (3) 285
19 Pod beak
Thick beak (4) 13
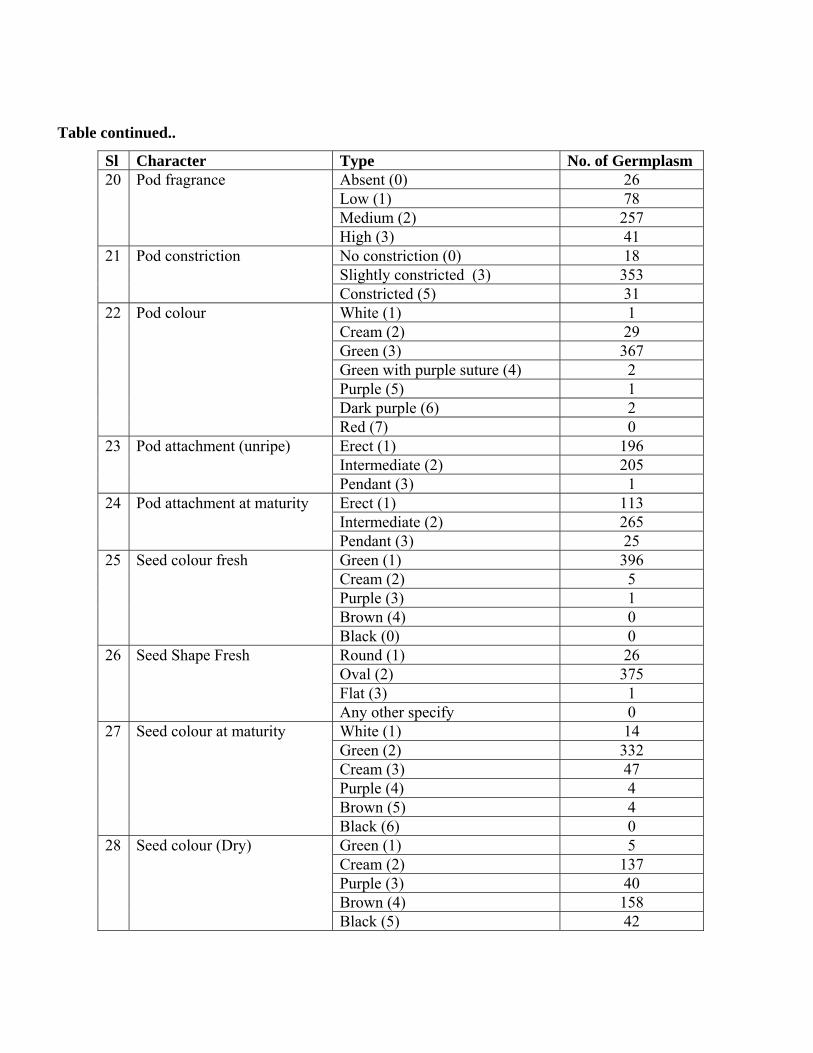
Table continued..
Sl Character Type No. of Germplasm Absent (0) 26 Low (1) 78 Medium (2) 257
20 Pod fragrance
High (3) 41 No constriction (0) 18 Slightly constricted (3) 353
21 Pod constriction
Constricted (5) 31 White (1) 1 Cream (2) 29 Green (3) 367 Green with purple suture (4) 2 Purple (5) 1 Dark purple (6) 2
22 Pod colour
Red (7) 0 Erect (1) 196 Intermediate (2) 205
23 Pod attachment (unripe)
Pendant (3) 1 Erect (1) 113 Intermediate (2) 265
24 Pod attachment at maturity
Pendant (3) 25 Green (1) 396 Cream (2) 5 Purple (3) 1 Brown (4) 0
25 Seed colour fresh
Black (0) 0 Round (1) 26 Oval (2) 375 Flat (3) 1
26 Seed Shape Fresh
Any other specify 0 White (1) 14 Green (2) 332 Cream (3) 47 Purple (4) 4 Brown (5) 4
27 Seed colour at maturity
Black (6) 0 Green (1) 5 Cream (2) 137 Purple (3) 40 Brown (4) 158
28 Seed colour (Dry)
Black (5) 42

Table continued..
Sl. Character Type No. of Germplasm Round (1) 25 Oval (2) 255 Flat (3) 5
29 Seed shape (Dry)
Any other specify (4) 0 Smooth (3) 400 Moderately ridged 2
30 Seed texture at maturity
Markedly ridged 0
b. Variability for quantitative characters in the germplasm accessions
Sl. Characters Range Mean Max Value Min Value
11.1 2.5 1. Leaf length (cm) D-72 GL-56
4.9
10 2.3 2. Leaf width (cm) D-64 GL-530
4.7
21.4 6.2 3 Leaf let length (cm) GL-201 GL-532
12
11 1 4 Primary branches CPI 31113 D-62
2.9
23 2 5 Secondary branches CPI 31113 D-16
5.8
17 8.6 6 Flower bud length (mm) D-51 GL-122
11.9
12 4 7 Flower bud width (mm) GL-413 D-35
9.5
36 5 8 Number of flower buds/ raceme GL-29 GL-69
18.3
25 2 9 No. Raceme/plant GL-513 GL-404
5.9
43 2.5 10 Raceme length (cm) D-27 GL-527
14.7
0.26 0.1 11 Peduncle length (cm) GL-18 CPI 76977
0.2
13 2 12 No. nodes/ raceme GL-100 GL-69
7
6 1 13 No. of buds/ node GL- 118 D-51
3.9
113 43 14 Days to 50% flowering GL-548 D-44
78.9
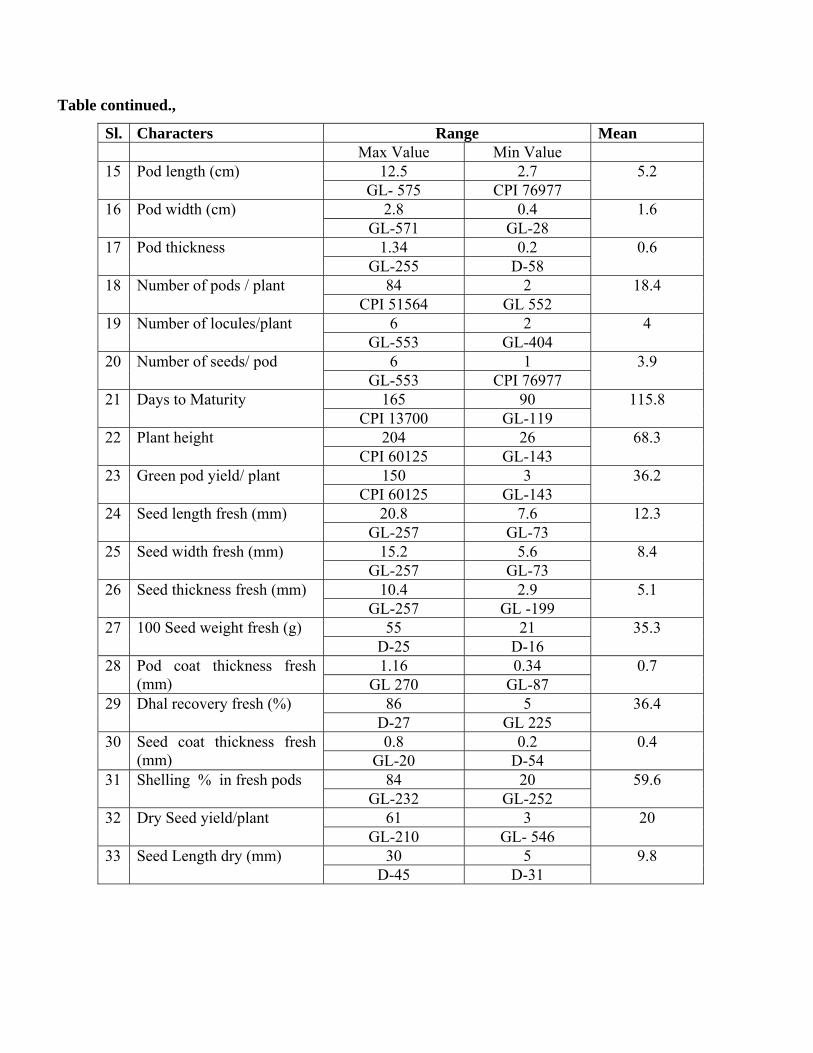
Table continued.,
Sl. Characters Range Mean Max Value Min Value
12.5 2.7 15 Pod length (cm) GL- 575 CPI 76977
5.2
2.8 0.4 16 Pod width (cm) GL-571 GL-28
1.6
1.34 0.2 17 Pod thickness GL-255 D-58
0.6
84 2 18 Number of pods / plant CPI 51564 GL 552
18.4
6 2 19 Number of locules/plant GL-553 GL-404
4
6 1 20 Number of seeds/ pod GL-553 CPI 76977
3.9
165 90 21 Days to Maturity CPI 13700 GL-119
115.8
204 26 22 Plant height CPI 60125 GL-143
68.3
150 3 23 Green pod yield/ plant CPI 60125 GL-143
36.2
20.8 7.6 24 Seed length fresh (mm) GL-257 GL-73
12.3
15.2 5.6 25 Seed width fresh (mm) GL-257 GL-73
8.4
10.4 2.9 26 Seed thickness fresh (mm) GL-257 GL -199
5.1
55 21 27 100 Seed weight fresh (g) D-25 D-16
35.3
1.16 0.34 28 Pod coat thickness fresh (mm) GL 270 GL-87
0.7
86 5 29 Dhal recovery fresh (%) D-27 GL 225
36.4
0.8 0.2 30 Seed coat thickness fresh (mm) GL-20 D-54
0.4
84 20 31 Shelling % in fresh pods GL-232 GL-252
59.6
61 3 32 Dry Seed yield/plant GL-210 GL- 546
20
30 5 33 Seed Length dry (mm) D-45 D-31
9.8
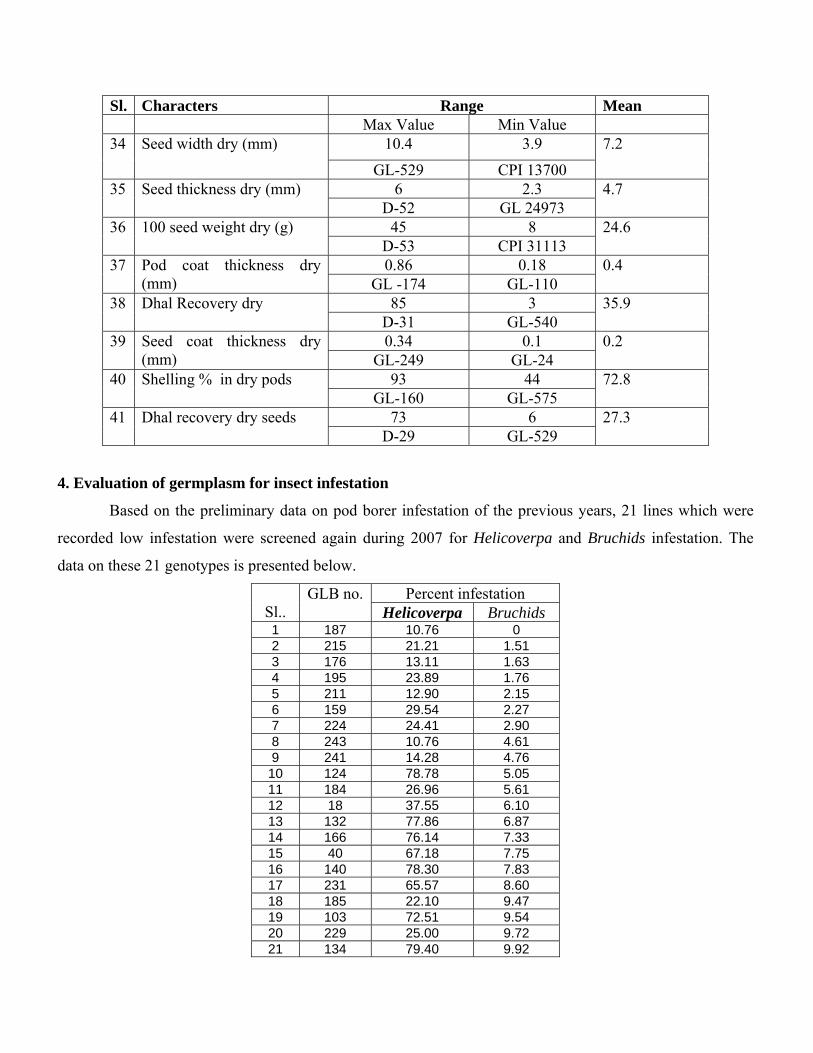
Sl. Characters Range Mean Max Value Min Value
10.4 3.9 34 Seed width dry (mm)
GL-529 CPI 13700
7.2
6 2.3 35 Seed thickness dry (mm) D-52 GL 24973
4.7
45 8 36 100 seed weight dry (g) D-53 CPI 31113
24.6
0.86 0.18 37 Pod coat thickness dry (mm) GL -174 GL-110
0.4
85 3 38 Dhal Recovery dry D-31 GL-540
35.9
0.34 0.1 39 Seed coat thickness dry (mm) GL-249 GL-24
0.2
93 44 40 Shelling % in dry pods GL-160 GL-575
72.8
73 6 41 Dhal recovery dry seeds D-29 GL-529
27.3
4. Evaluation of germplasm for insect infestation
Based on the preliminary data on pod borer infestation of the previous years, 21 lines which were
recorded low infestation were screened again during 2007 for Helicoverpa and Bruchids infestation. The
data on these 21 genotypes is presented below.
Percent infestation Sl..
GLB no.Helicoverpa Bruchids
1 187 10.76 0 2 215 21.21 1.51 3 176 13.11 1.63 4 195 23.89 1.76 5 211 12.90 2.15 6 159 29.54 2.27 7 224 24.41 2.90 8 243 10.76 4.61 9 241 14.28 4.76 10 124 78.78 5.05 11 184 26.96 5.61 12 18 37.55 6.10 13 132 77.86 6.87 14 166 76.14 7.33 15 40 67.18 7.75 16 140 78.30 7.83 17 231 65.57 8.60 18 185 22.10 9.47 19 103 72.51 9.54 20 229 25.00 9.72 21 134 79.40 9.92

Among the 21 genotypes screened, pod borer and bruchid infestation was low in the genotypes GL
187, GL 243, GL 211, GL176 and GL 241.
Again during 2008, totally 132 germplasm lines which including the above mentioned 21 lines were
evaluated for insect infestation. Pod damage due to Helicoverpa ranges from 1.8(GL 187) to 36.8 per cent
(GL 9), whereas Adisura infestation varies from 0.5(GL 55) to 7.9 per cent (GL 14) and Bruchids infestation
ranges from 0.6(GL 187) to 40 percent (GL 37). The average infestation was 18 % with Helicoverpa
followed by 12.1 % with bruchids and 3.1 percent with Adisura. Percent infestation, number of germplasm
lines with different levels of infestation and lines showing less than 10 % infestation due to Helicoverpa,
Adisura and Bruchids is presented below.
Per cent pod infestation of Helicoverpa, Adisura and Bruchids in Dolichos bean
Total Range (%) Helicoverpa Adisura Bruchids Min. 18 0.5 0.6 Max. 36.8 7.9 40
145
Average 18 3.1 12.1
Number of germplasm lines with different level of infestation
Scale (%)
Helicoverpa Adisura Bruchids
0-10 13 132 42 11-20 70 - 83 21-30 46 - 7 31-50 3 - - >50% - - - Total 132 132 132
Helicoverpa Adisura Bruchid
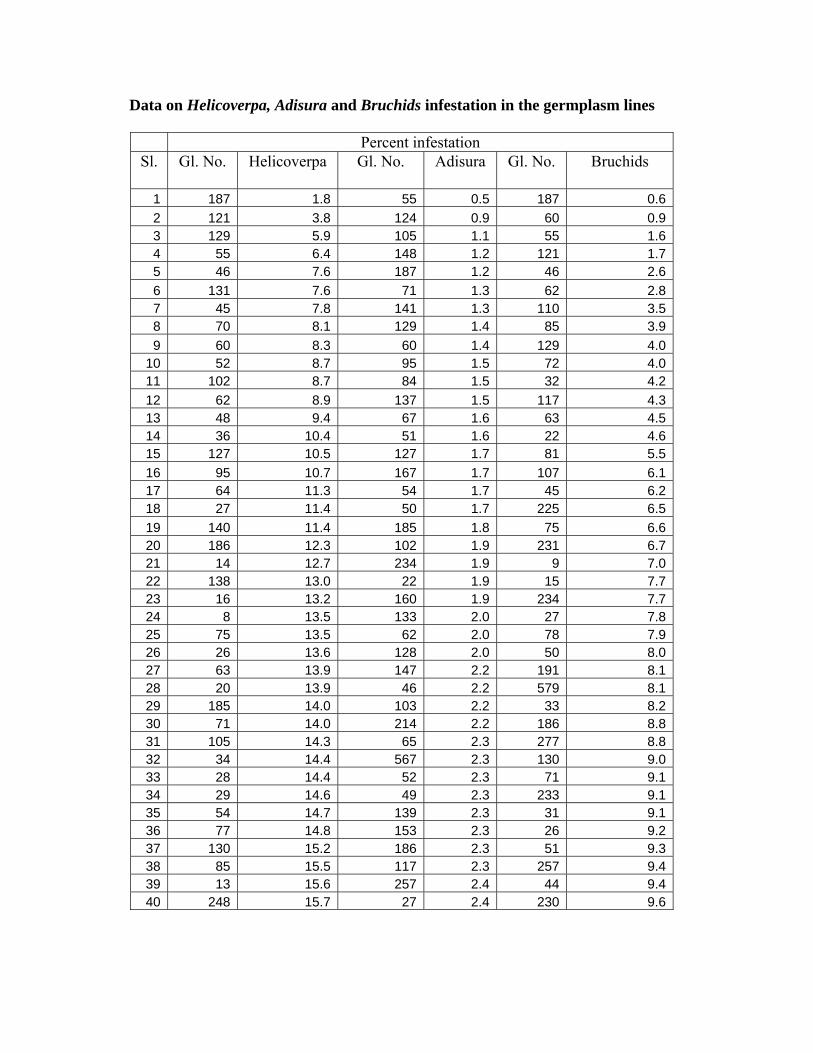
Data on Helicoverpa, Adisura and Bruchids infestation in the germplasm lines
Percent infestation Sl. Gl. No. Helicoverpa Gl. No. Adisura Gl. No. Bruchids
1 187 1.8 55 0.5 187 0.62 121 3.8 124 0.9 60 0.93 129 5.9 105 1.1 55 1.64 55 6.4 148 1.2 121 1.75 46 7.6 187 1.2 46 2.66 131 7.6 71 1.3 62 2.87 45 7.8 141 1.3 110 3.58 70 8.1 129 1.4 85 3.99 60 8.3 60 1.4 129 4.0
10 52 8.7 95 1.5 72 4.011 102 8.7 84 1.5 32 4.212 62 8.9 137 1.5 117 4.313 48 9.4 67 1.6 63 4.514 36 10.4 51 1.6 22 4.615 127 10.5 127 1.7 81 5.516 95 10.7 167 1.7 107 6.117 64 11.3 54 1.7 45 6.218 27 11.4 50 1.7 225 6.519 140 11.4 185 1.8 75 6.620 186 12.3 102 1.9 231 6.721 14 12.7 234 1.9 9 7.022 138 13.0 22 1.9 15 7.723 16 13.2 160 1.9 234 7.724 8 13.5 133 2.0 27 7.825 75 13.5 62 2.0 78 7.926 26 13.6 128 2.0 50 8.027 63 13.9 147 2.2 191 8.128 20 13.9 46 2.2 579 8.129 185 14.0 103 2.2 33 8.230 71 14.0 214 2.2 186 8.831 105 14.3 65 2.3 277 8.832 34 14.4 567 2.3 130 9.033 28 14.4 52 2.3 71 9.134 29 14.6 49 2.3 233 9.135 54 14.7 139 2.3 31 9.136 77 14.8 153 2.3 26 9.237 130 15.2 186 2.3 51 9.338 85 15.5 117 2.3 257 9.439 13 15.6 257 2.4 44 9.440 248 15.7 27 2.4 230 9.6

Percent infestation Sl. Gl. No. Helicoverpa Gl. No. Adisura Gl. No. Bruchids
41 128 15.8 85 2.4 77 9.642 110 15.9 142 2.4 228 9.743 132 15.9 248 2.5 36 10.044 141 16.0 37 2.5 52 10.245 139 16.7 236 2.5 25 10.346 226 16.7 121 2.5 185 10.447 230 16.7 43 2.5 13 10.548 101 16.8 199 2.5 195 10.549 11 16.8 76 2.5 232 10.550 81 17.0 520 2.6 161 10.751 43 17.0 114 2.6 48 10.852 108 17.1 106 2.6 218 10.953 72 17.2 195 2.6 411 11.354 113 17.3 232 2.6 139 11.455 567 17.4 220 2.7 219 11.456 218 17.6 28 2.7 28 11.557 103 17.8 166 2.7 131 11.558 148 17.8 176 2.7 212 11.659 83 18.1 191 2.7 229 11.660 106 18.2 33 2.7 147 11.761 50 18.2 113 2.7 128 11.762 133 18.2 83 2.8 127 11.863 176 18.2 45 2.9 132 11.864 147 18.3 226 2.9 141 12.065 119 18.6 140 2.9 220 12.066 32 18.6 159 2.9 176 12.267 80 18.7 209 2.9 30 12.568 117 18.7 277 2.9 209 12.569 10 18.9 138 3.0 14 12.670 25 19.0 233 3.0 102 12.771 142 19.0 101 3.0 10 12.772 78 19.1 119 3.1 113 12.773 33 19.1 151 3.1 134 12.874 124 19.1 75 3.1 67 12.875 211 19.2 40 3.1 165 12.876 22 19.2 17 3.1 567 12.977 137 19.4 63 3.2 65 13.078 76 19.5 134 3.2 224 13.079 238 19.7 228 3.2 226 13.080 51 19.7 132 3.3 74 13.1
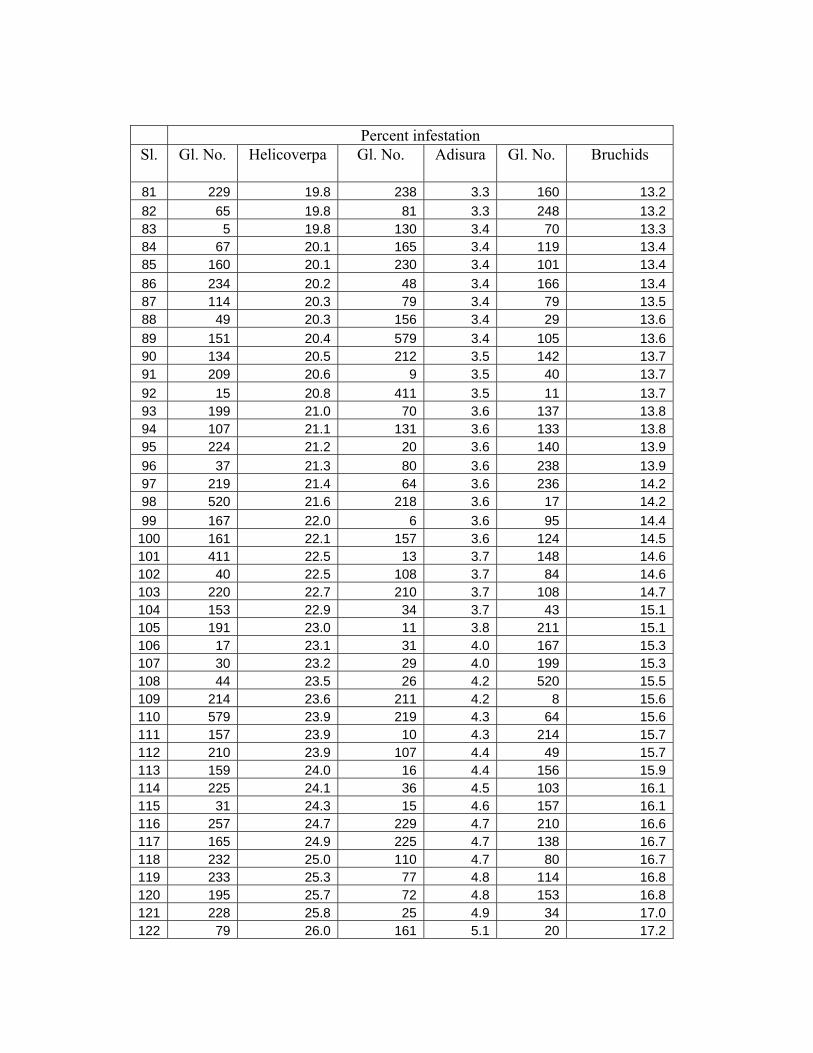
Percent infestation Sl. Gl. No. Helicoverpa Gl. No. Adisura Gl. No. Bruchids
81 229 19.8 238 3.3 160 13.282 65 19.8 81 3.3 248 13.283 5 19.8 130 3.4 70 13.384 67 20.1 165 3.4 119 13.485 160 20.1 230 3.4 101 13.486 234 20.2 48 3.4 166 13.487 114 20.3 79 3.4 79 13.588 49 20.3 156 3.4 29 13.689 151 20.4 579 3.4 105 13.690 134 20.5 212 3.5 142 13.791 209 20.6 9 3.5 40 13.792 15 20.8 411 3.5 11 13.793 199 21.0 70 3.6 137 13.894 107 21.1 131 3.6 133 13.895 224 21.2 20 3.6 140 13.996 37 21.3 80 3.6 238 13.997 219 21.4 64 3.6 236 14.298 520 21.6 218 3.6 17 14.299 167 22.0 6 3.6 95 14.4100 161 22.1 157 3.6 124 14.5101 411 22.5 13 3.7 148 14.6102 40 22.5 108 3.7 84 14.6103 220 22.7 210 3.7 108 14.7104 153 22.9 34 3.7 43 15.1105 191 23.0 11 3.8 211 15.1106 17 23.1 31 4.0 167 15.3107 30 23.2 29 4.0 199 15.3108 44 23.5 26 4.2 520 15.5109 214 23.6 211 4.2 8 15.6110 579 23.9 219 4.3 64 15.6111 157 23.9 10 4.3 214 15.7112 210 23.9 107 4.4 49 15.7113 159 24.0 16 4.4 156 15.9114 225 24.1 36 4.5 103 16.1115 31 24.3 15 4.6 157 16.1116 257 24.7 229 4.7 210 16.6117 165 24.9 225 4.7 138 16.7118 232 25.0 110 4.7 80 16.7119 233 25.3 77 4.8 114 16.8120 195 25.7 72 4.8 153 16.8121 228 25.8 25 4.9 34 17.0122 79 26.0 161 5.1 20 17.2

Percent infestation Sl. Gl. No. Helicoverpa Gl. No. Adisura Gl. No. Bruchids
123 84 26.4 74 5.3 76 17.6124 277 26.5 224 5.5 16 18.5125 231 26.7 32 5.5 159 18.6126 236 26.7 30 5.6 54 21.4127 212 26.7 5 5.6 5 22.1128 156 27.0 231 5.7 106 26.4129 166 28.2 8 6.3 6 27.3130 74 32.0 78 7.0 151 27.3131 6 32.7 44 7.7 83 32.7132 9 36.8 14 7.9 37 40.0
Amongst the germplasm lines evaluated over two years, GL 187 showed resistant reaction to
Helicovrapa, Adisura and Bruchids infestation.
5. Breeding for high yield with desirable traits
In order to develop high yielding genotypes, different crosses were affected involving the newly
identified variety HA 4 and germplasm lines. Different cross combinations made over the years is presented
below.
sl Cross combinations 1 HA 4 X GLB 16 2 HA 4 X GLB 91 3 HA 4 X GLB 121 4 HA 4 X GLB 124 5 HA 4 X GLB 153 6 HA 4 X GLB 122 7 HA 4 X GLB 43
During the subsequent years, different generations of segregating materials (F1 generation to
F4 generation) were raised and selections were made in the segregating populations.

F2 generation
In the F2 generation, these populations showed segregation for many traits viz., growth habit,
plant type, flowering, maturity and flower color, pod characters and seed color. We had imposed selection in
these populations for desirable traits which are having high yield, determinate growth habit, and high
number of branches, good pod fragrance and constricted pods.
Sl. Crosses No. of selections Type of selections made
1 HA 4 X GLB 16 Early types -40 Med duration-82
2 HA 4 X GLB 91 Early types –68 Med duration-18
3 HA 4 X GLB 121 Early types –6 Med duration-24
4 HA 4 X GLB 124 Early types –14 Med duration-36
5 HA 4 X GLB 153 Early types -46 Med duration-144
Growth habit, plant type, pod shape, pod constriction, pod fragrance, pod number, branch number, long pod bearing branches, seed type, high shelling percentage
6 HA 4 X GLB 122 Early types –8 Med duration-32
Growth habit, pod bearing at nodes, pod shape, pod constriction, pod fragrance, pod number, branch number, long pod bearing branches, high shelling percentage
7 HA 4 X GLB 43 Early types –16 Med duration-38
Growth habit, pod shape, pod constriction, pod fragrance, pod number, branch number, long pod bearing branches, seed type
8 Sel 246 X GLB 122 Early types –11 Med duration-31
Growth habit, pod bearing at nodes, pod constriction, pod fragrance, pod number, branch number
Crossing HA 4 GLB 122
F2 generation

F3 generation
All the early selections from the above crosses were forwarded for F3 generation. These
selections were raised in plant to progeny rows. Progeny rows exhibiting uniformity for the desirable
characters and few high yielding single plants from other rows were selected. These selections will be
forwarded to F4 family confirmation.
F4 generation
The selections made in F4 were forwarded to F5 generation. From these progenies, progenies
selections were made based on earliness, branch number, pod type, pod number, pod fragrance and growth
habit (both determinate and semi determinate types).
F5 generation
The selections made in F4 generation during the first part of the season of 2007 were sown were
grown again during March 2008 at Hebbal. Few high yielding lines were also included in the evaluation
trail. The crop is at flowering, pod maturity and harvesting stage.
F6 generation
The progeny selections made in F5 generation of the following crosses during the previous year
(2007) were sown in March 2008. From these populations, high yielding progenies (20) were selected based
on their performance on yield, duration, growth habit, pod number, pod shape, pod fragrance and named
them as HA selections.
In addition to above, we have made other crosses also and they are in different generations. Selections were made from these crosses also for desirable traits. The progress on these populations will be reported in the next years report.
Sl. Crosses No. of F5 progenies row sown
No. of selections Made in
1 HA 4 X GLB 16 10 03 2 HA 4 X GLB 91 03 01 3 HA 4 X GLB 121 08 02 4 HA 4 X GLB 124 18 03 5 HA 4 X GLB 153 18 05 6 HA 4 X GLB 43 11 03 7 HA 4 X GLB 122 13 01 8 Sel 246 X GLB 122 15 02 20

6. Evaluation of breeding material in the Multilocation trial:
All the 20 advanced breeding lines were subjected to post harvest observations for seed weight, seed
colour, seed shape and seed lustrous ness. From these 20, finally 12 lines based on desirable characters were
selected viz., HA 7-1, HA 7-2, HA 7-3, HA 7-4, HA 7-5, HA 7 -6 HA 7-7, HA 8-1, HA 8-2, HA 8-3, HA 8-
4 and HA 8-5. All these 12 lines along with check HA 4 were evaluated during September-December 2008
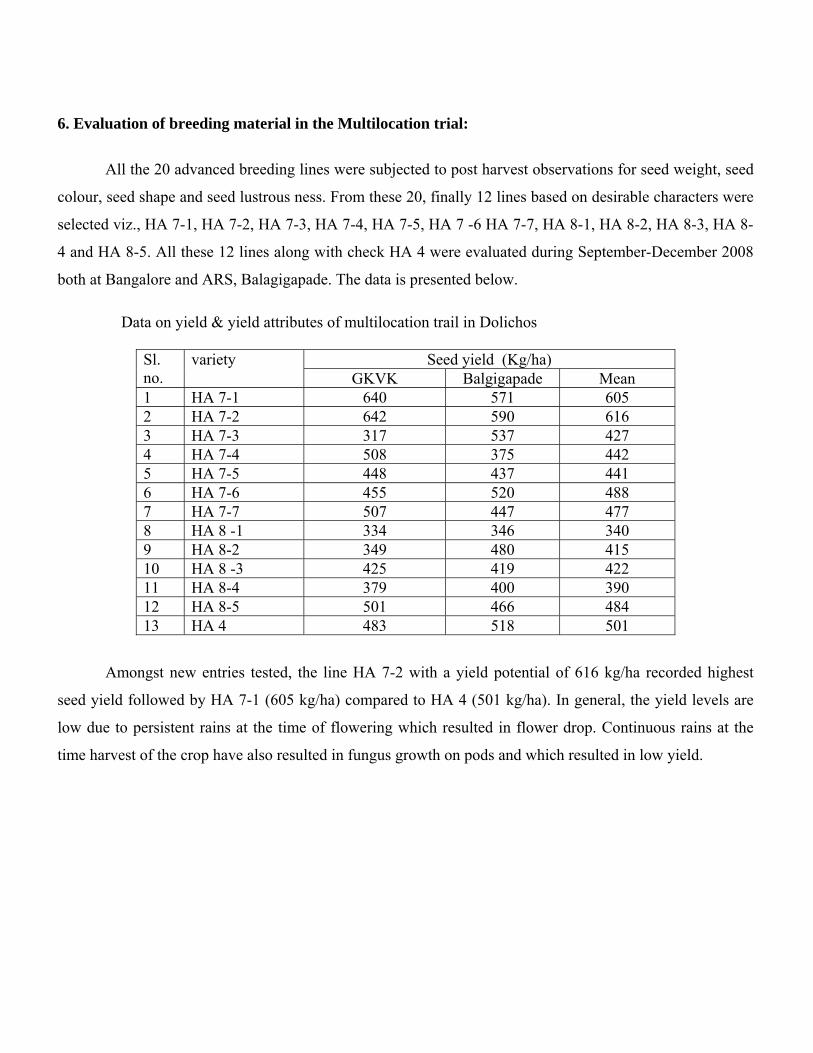
both at Bangalore and ARS, Balagigapade. The data is presented below. Data on yield & yield attributes of multilocation trail in Dolichos
Seed yield (Kg/ha) Sl. no.
variety GKVK Balgigapade Mean
1 HA 7-1 640 571 605 2 HA 7-2 642 590 616 3 HA 7-3 317 537 427 4 HA 7-4 508 375 442 5 HA 7-5 448 437 441 6 HA 7-6 455 520 488 7 HA 7-7 507 447 477 8 HA 8 -1 334 346 340 9 HA 8-2 349 480 415 10 HA 8 -3 425 419 422 11 HA 8-4 379 400 390 12 HA 8-5 501 466 484 13 HA 4 483 518 501
Amongst new entries tested, the line HA 7-2 with a yield potential of 616 kg/ha recorded highest
seed yield followed by HA 7-1 (605 kg/ha) compared to HA 4 (501 kg/ha). In general, the yield levels are
low due to persistent rains at the time of flowering which resulted in flower drop. Continuous rains at the
time harvest of the crop have also resulted in fungus growth on pods and which resulted in low yield.

6. Development of mapping population based on the molecular markers
To develop the mapping populations, the following crosses were carried out. In some crosses, very few
seeds were obtained.
sl Male line Female line 1 GL 148 CPI 24973 2 GL 148 CPI 31113 3 GL 148 CPI 60216 4 GL 153 CPI 24973 5 GL 153 CPI 31113 6 GL 153 CPI 60216 7 GL 147 CPI 24973 8 GL 147 CPI 31113 9 GL 147 CPI 60216 10 GL 48 CPI 24973 11 GL 48 CPI 31113 12 GL 48 CPI 60216 13 HA 4 CPI 24973 14 HA 4 CPI 31113 15 HA 4 CPI 60216
F1 generations were raised only in the following crosses where sufficient seeds were obtained. Plants
were raised under nylon nets for selfing. Though sufficient seed material were obtained from the above
crosses, we have selected only two crosses namely HA 4 x CPI 31113 and HA 4 x CPI 60216 to develop
mapping populations and Recombinant Inbred Lines. At maturity, F1 seeds were harvested, dried and stored
with proper care.
1 GL 148 CPI 60216 2 GL 153 CPI 31113 3 GL 153 CPI 60216 4 GL 147 CPI 60216 5 HA 4 CPI 24973 6 HA 4 CPI 31113 7 HA 4 CPI 60216

F2 material
To raise F2 populations, we have selected only two cross combinations namely, HA 4 x CPI 31113
and HA 4 x CPI 60216. These crosses were selected based on diversity on both phenotypic and molecular
data. Leaf samples were collected from all the individual plants from both the crosses for DNA analysis for
the Venkatesha’s s molecular project.

Variability for pod characters
Variability for seed characters

7. Development of Recombinant Inbred Lines
Two crosses involving local and with wild parents (HA 4 x CPI 31113, HA 4 x CPI 60125) which
were the part of Venkatesha’s s molecular project were selected for development of RIL populations. At the
time of flower bud initiation, all the individual F2 plants were selfed by using nylon net bags to avoid cross
pollination through insects. Only selfed seeds were harvested for forwarding to next generations.
8. Inheritance studies in the crosses HA 4 x CPI 31113 and HA 4 x CPI 60126
The genetic inheritance pattern of introgressed characters in the cross HA 4 X CPI 31113 revealed
that growth habit, flower bud colour, pod curvature and seed colour were controlled by a single dominant
gene and genes segregating in F2 as 3:1 ratio, Where as branch orientation and pod constriction were
controlled by two dominant threshold genes (11:5) indicating controlled by two genes. While pod fragrance
and stem pigmentation were controlled by two genes but complementary gene action governing this
character.
The genetic inheritance pattern in the other cross HA 4 X CPI 60126 revealed that growth habit
governed by two dominant threshold genes and segregating ratio of 11:5. Whereas branch orientation,
Flower bud colour and pod constriction were governed by single dominant gene and F2 segregation ratio of
3:1, while pod curvature, pod fragrance and stem pigmentation controlled by two genes with complementary
gene action (9:7) governing this character.

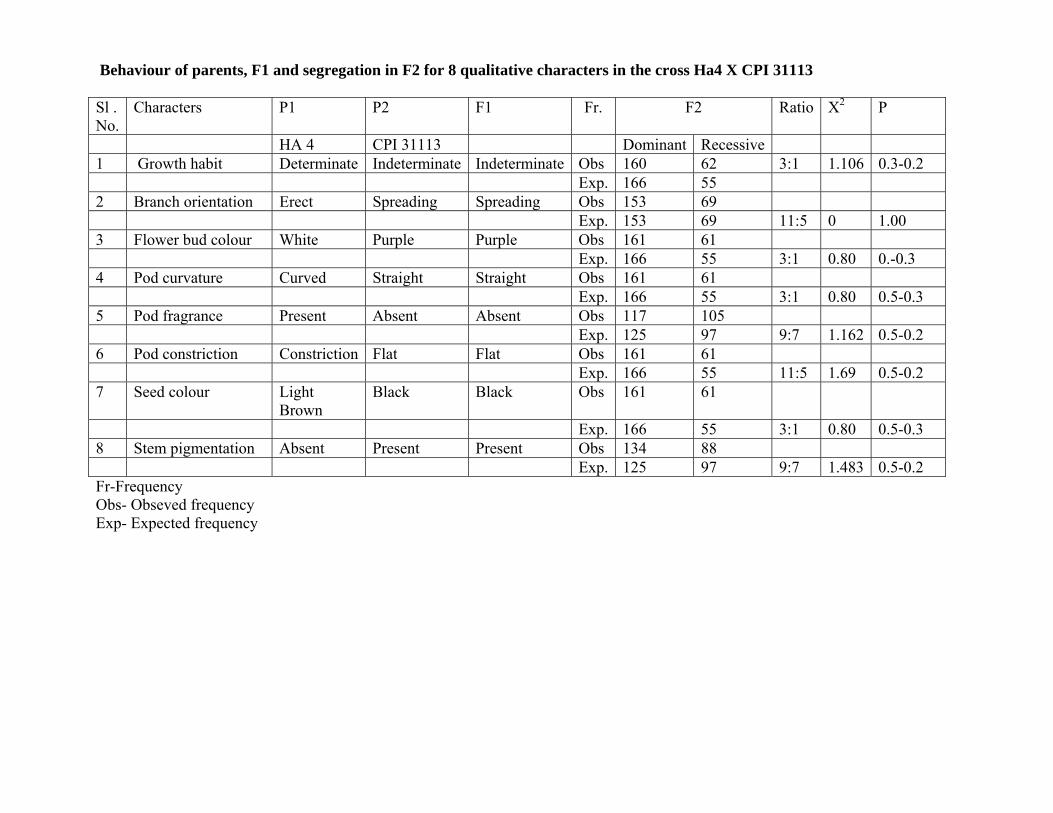
Behaviour of parents, F1 and segregation in F2 for 8 qualitative characters in the cross Ha4 X CPI 31113
Sl . No.
Characters P1 P2 F1 Fr. F2 Ratio X2 P
HA 4 CPI 31113 Dominant Recessive 1 Growth habit Determinate Indeterminate Indeterminate Obs 160 62 3:1 1.106 0.3-0.2 Exp. 166 55 2 Branch orientation Erect Spreading Spreading Obs 153 69 Exp. 153 69 11:5 0 1.00 3 Flower bud colour White Purple Purple Obs 161 61 Exp. 166 55 3:1 0.80 0.-0.3 4 Pod curvature Curved Straight Straight Obs 161 61 Exp. 166 55 3:1 0.80 0.5-0.3 5 Pod fragrance Present Absent Absent Obs 117 105 Exp. 125 97 9:7 1.162 0.5-0.2 6 Pod constriction Constriction Flat Flat Obs 161 61 Exp. 166 55 11:5 1.69 0.5-0.2 7 Seed colour Light
Brown Black Black Obs 161 61
Exp. 166 55 3:1 0.80 0.5-0.3 8 Stem pigmentation Absent Present Present Obs 134 88 Exp. 125 97 9:7 1.483 0.5-0.2 Fr-Frequency Obs- Obseved frequency Exp- Expected frequency
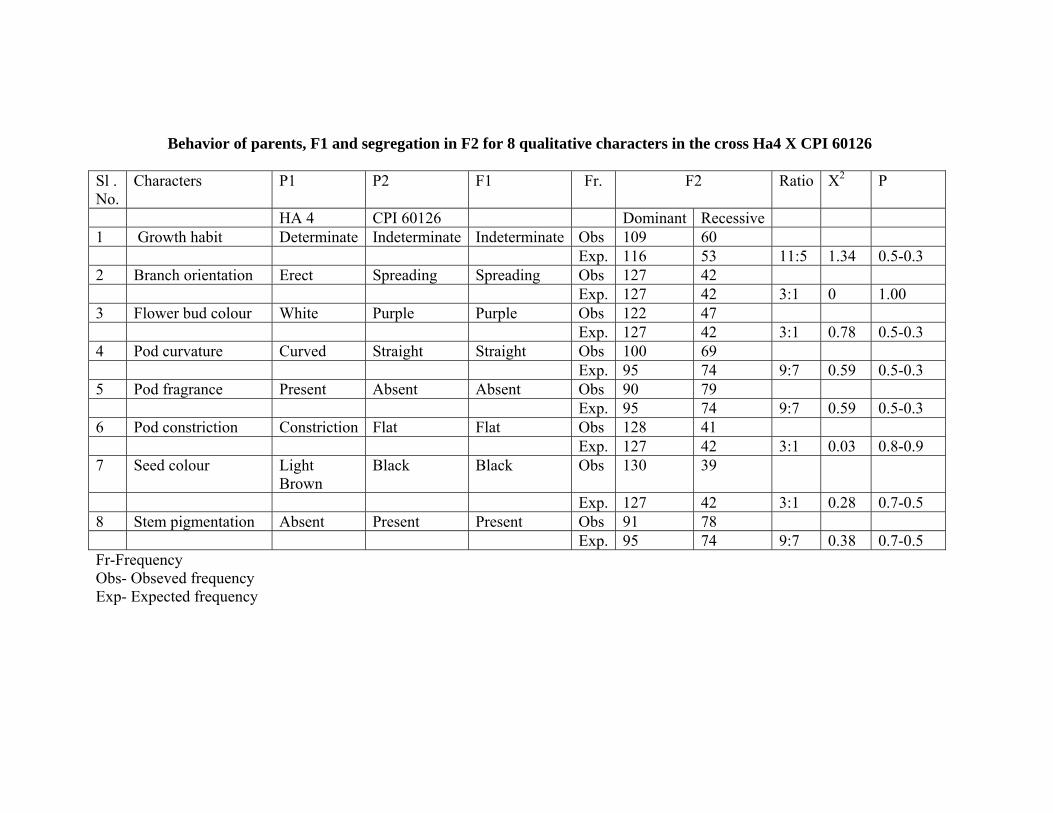
Behavior of parents, F1 and segregation in F2 for 8 qualitative characters in the cross Ha4 X CPI 60126 Sl . No.
Characters P1 P2 F1 Fr. F2 Ratio X2 P
HA 4 CPI 60126 Dominant Recessive 1 Growth habit Determinate Indeterminate Indeterminate Obs 109 60 Exp. 116 53 11:5 1.34 0.5-0.3 2 Branch orientation Erect Spreading Spreading Obs 127 42 Exp. 127 42 3:1 0 1.00 3 Flower bud colour White Purple Purple Obs 122 47 Exp. 127 42 3:1 0.78 0.5-0.3 4 Pod curvature Curved Straight Straight Obs 100 69 Exp. 95 74 9:7 0.59 0.5-0.3 5 Pod fragrance Present Absent Absent Obs 90 79 Exp. 95 74 9:7 0.59 0.5-0.3 6 Pod constriction Constriction Flat Flat Obs 128 41 Exp. 127 42 3:1 0.03 0.8-0.9 7 Seed colour Light
Brown Black Black Obs 130 39
Exp. 127 42 3:1 0.28 0.7-0.5 8 Stem pigmentation Absent Present Present Obs 91 78 Exp. 95 74 9:7 0.38 0.7-0.5 Fr-Frequency Obs- Obseved frequency Exp- Expected frequency
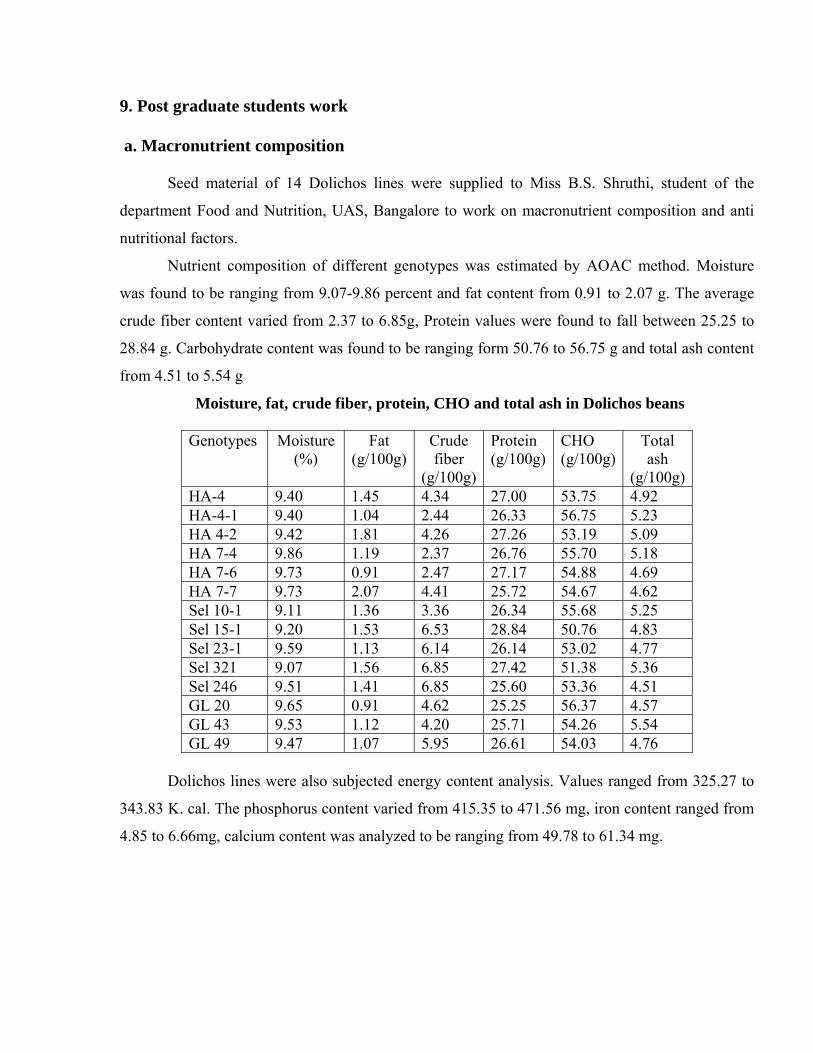
9. Post graduate students work a. Macronutrient composition
Seed material of 14 Dolichos lines were supplied to Miss B.S. Shruthi, student of the
department Food and Nutrition, UAS, Bangalore to work on macronutrient composition and anti
nutritional factors.
Nutrient composition of different genotypes was estimated by AOAC method. Moisture
was found to be ranging from 9.07-9.86 percent and fat content from 0.91 to 2.07 g. The average
crude fiber content varied from 2.37 to 6.85g, Protein values were found to fall between 25.25 to
28.84 g. Carbohydrate content was found to be ranging form 50.76 to 56.75 g and total ash content
from 4.51 to 5.54 g
Moisture, fat, crude fiber, protein, CHO and total ash in Dolichos beans
Genotypes Moisture (%)
Fat (g/100g)
Crude fiber
(g/100g)
Protein (g/100g)
CHO (g/100g)
Total ash
(g/100g)HA-4 9.40 1.45 4.34 27.00 53.75 4.92 HA-4-1 9.40 1.04 2.44 26.33 56.75 5.23 HA 4-2 9.42 1.81 4.26 27.26 53.19 5.09 HA 7-4 9.86 1.19 2.37 26.76 55.70 5.18 HA 7-6 9.73 0.91 2.47 27.17 54.88 4.69 HA 7-7 9.73 2.07 4.41 25.72 54.67 4.62 Sel 10-1 9.11 1.36 3.36 26.34 55.68 5.25 Sel 15-1 9.20 1.53 6.53 28.84 50.76 4.83 Sel 23-1 9.59 1.13 6.14 26.14 53.02 4.77 Sel 321 9.07 1.56 6.85 27.42 51.38 5.36 Sel 246 9.51 1.41 6.85 25.60 53.36 4.51 GL 20 9.65 0.91 4.62 25.25 56.37 4.57 GL 43 9.53 1.12 4.20 25.71 54.26 5.54 GL 49 9.47 1.07 5.95 26.61 54.03 4.76
Dolichos lines were also subjected energy content analysis. Values ranged from 325.27 to
343.83 K. cal. The phosphorus content varied from 415.35 to 471.56 mg, iron content ranged from
4.85 to 6.66mg, calcium content was analyzed to be ranging from 49.78 to 61.34 mg.
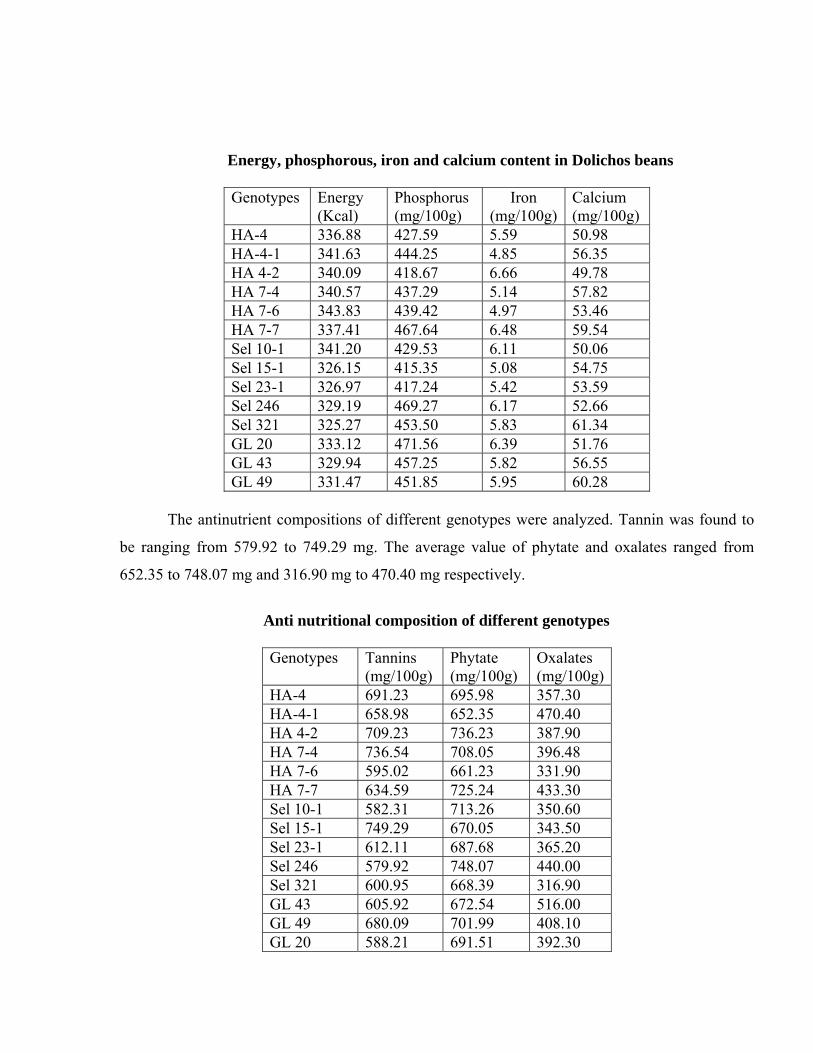
Energy, phosphorous, iron and calcium content in Dolichos beans
Genotypes Energy
(Kcal) Phosphorus (mg/100g)
Iron (mg/100g)
Calcium (mg/100g)
HA-4 336.88 427.59 5.59 50.98 HA-4-1 341.63 444.25 4.85 56.35 HA 4-2 340.09 418.67 6.66 49.78 HA 7-4 340.57 437.29 5.14 57.82 HA 7-6 343.83 439.42 4.97 53.46 HA 7-7 337.41 467.64 6.48 59.54 Sel 10-1 341.20 429.53 6.11 50.06 Sel 15-1 326.15 415.35 5.08 54.75 Sel 23-1 326.97 417.24 5.42 53.59 Sel 246 329.19 469.27 6.17 52.66 Sel 321 325.27 453.50 5.83 61.34 GL 20 333.12 471.56 6.39 51.76 GL 43 329.94 457.25 5.82 56.55 GL 49 331.47 451.85 5.95 60.28
The antinutrient compositions of different genotypes were analyzed. Tannin was found to
be ranging from 579.92 to 749.29 mg. The average value of phytate and oxalates ranged from
652.35 to 748.07 mg and 316.90 mg to 470.40 mg respectively.
Anti nutritional composition of different genotypes
Genotypes Tannins (mg/100g)
Phytate (mg/100g)
Oxalates (mg/100g)
HA-4 691.23 695.98 357.30 HA-4-1 658.98 652.35 470.40 HA 4-2 709.23 736.23 387.90 HA 7-4 736.54 708.05 396.48 HA 7-6 595.02 661.23 331.90 HA 7-7 634.59 725.24 433.30 Sel 10-1 582.31 713.26 350.60 Sel 15-1 749.29 670.05 343.50 Sel 23-1 612.11 687.68 365.20 Sel 246 579.92 748.07 440.00 Sel 321 600.95 668.39 316.90 GL 43 605.92 672.54 516.00 GL 49 680.09 701.99 408.10 GL 20 588.21 691.51 392.30

The genotypes Sel 15-1, HA 4-2, GL 49 and HA7-7 had higher nutrient content with
regard to protein, iron, calcium and phosphorus respectively. The genotypes Sel 246, HA 4-1 and
Sel 321 had lower antinutrient composition for tannin, phytate and oxalate respectively.
b. Development of food products from Dolichos
Green seeds of selected two lines (GL 43 and GL 49) were given to Miss B.S. Shruthi, student of
the department Food and Nutrition, UAS, Bangalore for development of food products. Six
products were developed from these two genotypes viz., Upma, Usli, Sambar, Palya, Khichidi and
fried Khara. The products were evaluated by 5 point hedonic scale for appearance, aroma, texture,
taste and overall acceptability. All the products were accepted. From the mean scores of
organoleptic characteristics, the data that varies to a maximum of 7.5 per cent with a character
shows marginal differences through the data was found significant. Therefore, it could be either of
the genotype GL 43 or GL 49 is suitable for preparation of these products.
1.

I0. Molecular work
a. Diversity studies in Lablab purpureus:
One hundred & ninety two primer pairs (table I) received from Dr.D.J.Kim, were used to
screen the selected 10 genotypes of Lablab purpureus. (HA3, Mac-1, SRL, Rongai, Highworth,
CPI36903, CPI52544, CPI52552, CPI52535 and ILRI6536) to assess the genetic diversity within
Lablab and to gain some understanding of the transferability of gene specific markers to Lablab
purpureus. These 10 genotypes comprises both Indian material and exotic material (please refer
table 2). In this study I have used the SSR markers, intron directed primers and few primers
designed based on Lablab sequence. And also I did AFLP studies for the selected 42 Indian
genotypes of Lablab purpureus with six primer.
Table I: Primers used for screening 10 genotypes of Lablab purpureus
DNA Name Sequence forward Sequence reverse Species CA906441-1 CTTCCCCACTGATTCATTTCTGTT CACTCCCCTTTGTTCTTGATTATG P. coccineus
CA909226-1 GATCTCTTTCACGCTGGACAT CACATAGCGACATGCTAGAACACT P. coccineus
CA910489-1 AGAAAAAGGAGGGAAATGCTGCTA TTGCCTCCGCTTTCTTTATCTTTC P. coccineus
CA910598-1 CCTCCAATTCGGTTCCAGCACTTC GCCCTTGATCTTCCTCTTTTTACG P. coccineus
CA911600-1 AACTGGTTCATCGCATTACTCTTC TGGCATCACCTTGTTTGGCATTAG P. coccineus
CA911990-1 TTTCATCCCCAGCATCAGCAGTTT TGGGGGTGGGGCAGAATCA P. coccineus
CA912170-1 TGGAGAAAGCATGGCAGGATACT CCCCGGAGGCAAGATTTCAA P. coccineus
CA914165-1 GATAATTTTGACGAGGGAGCAGA GGTGGAGAACAGGAGCAAAGTATT P. coccineus
CA912542-1 TTATTGGAGTTGCCCTGTTCTTCA GCCCCGTCTGCATTCCTTTGT P. coccineus
CA906286-1 CGGGCGGGGGAGATTGT CCCAGCCCACGTAAGGTAAGAAC P. coccineus
CA910928-1 CGGAATCTGGTCGGCCTACATCTA TGATACCCCTCCGGCAACAAG P. coccineus
CA907856-1 ACACTTGAGGGAAAGCCATCA TTCCAAGCCAAAGCAATAAC P. coccineus
M80799-1 ACGGCGGGCAATGTCAA AGTTCACGTGCTCCGCTGTTAT V. unguiculata CA906298-1 TAGTTGACCGGTTATCTGTTGTTT GCGCCTACGAGCATCACTTA P. coccineus Pc_CL2167-1 CATCACGCGATAATACACGACT TCTCTGCCGAAGGTTATGGA No items found CA912634-1 GACGGAATGAAGTTGAAGGTGGAT AGATGGGCCAAGATATTTTAGACT P. coccineus
CA913150-1 CGGAGATGGTGGCGTGTTATT CTTGTGCAGCCTTGTTTTCAGC P. coccineus
CA911895-1 CATCTGCTTTTGCCCATTACG CACCAAGCCGCTCTAGCAAGTTAT P. coccineus
CA911460-1 GAGCTGCAACAAGGAATGGATG ATGAGGGCTAACAGGAAAAGAAGG P. coccineus
CA902380-1 GGCCCCCTCAGACAGAT AGATAAATTTGGGCTTGAA P. coccineus
CA907467-1 AGCCTGTCAATTCCAAACTCCACT ATCCAACTTCAGCACCCAGACAAA P. coccineus
CA907742-1 TCGGTTGAAAAAGAAAAGCAGAAG ATAGGCACTCAGGAAGGGAATGTC P. coccineus
BQ481755-1 CATGGCGAAGTAACAAAAGTGGTC ATATGCAGGATGAGGAAGATGGTT P. vulgaris Pc_CL65-1 CTCGTCTCGCCACCCAGTTCT AGTCACCGCTCTCCACCCTTCT No items found CA900698-1 TTGTTCTCAGCGCAATGTGG CTGAGCACCAAAGCCCCTGACA P. coccineus
CA900229-1 TGCCGAGTCATCCCGAACAT CTCCAAAGCTGAAGCGAACCA P. coccineus
CA914163-1 ATGGTGAATGCCTGAAACAAATAC CATGACAAATGCCTGAATAAGACT P. coccineus
CA912177-1 CTTGGTCCGCCATACTCTGAT TTTTCTTTCCTCCTCTGGTATGG P. coccineus
AJ225088-3 GGTATGATGGCCGTGAGTGGT TAAAGCTGTACAACGGCGATGAA V. unguiculata
AJ225088-5 GAACACCACCGCCACACG GCACTAGCATTAACGCAAGAAACT V. unguiculata
AF287258-3 TAGCGCCATGACATTTACCTCTTA GCTCCGTTTTCTGCTTTATCTACA V. unguiculata
AF287258-5 CATTGGGCGAAAAGAAAGGAA CGCAAAAAGAAGGAGAGCAATAGA V. unguiculata
AF287258-1 AGGGGGTGAGCTTGTTTTTG TATTGCGAAGCCTTTAGTAGACCT V. unguiculata
AF287258-1C GTTTGATGGCATACTGGGACTTG CTCTTCCTCTGTTTTGCGGTTTA V. unguiculata
X91836-5 ATCGCGAAAAATGAATTATCTGGT TTATCGTCGGTGAAAACTGCT V. unguiculata
X91836-5C CCGGAAACATGGCATTATTATTAG CCATTGCCTCGTTCCCATCTT V. unguiculata
X91836-3 AGTCTCCACCTCCCCCTTCT TTTAGCATTTTGAGGCACTGATTC V. unguiculata
AB027154-3 ATAAAATACCACAACAAAGGAGAT CTTATTGGTTGATGTAGTGGAT V. unguiculata
AB028025-3 CGTTGCGCCGCCTTTTAGTTTAC TCAATTTGGCAGGATGAACC V. unguiculata
AB030293-3 ATACGCCCCCGGAATCTAC TAGCGGAACAAAACAAGAAGAATA V. unguiculata
AB030294-3 AAGGTGGTGAGGGCGAGAAG ACCAACCAAACCACAGAAATGAC V. unguiculata
AB030295-5 TGGGCGAGCTTACAACATACACA CGGCCCCACTTCCACCTT V. unguiculata
AB030295-3 TCGCATTCGCCCATCTGAG GCTTAGCATTGCTTACTCCATTCA V. unguiculata
AB030295-1 GTGCCTGCTGACATTGAAACTG ACGGAATCCCCAAGCAAAGTC V. unguiculata
AB038691-3 GACGAAGGCTTGGCTCTCC TTACCCACAAAAGTAAATCAAAT V. unguiculata
AB038692-3 AAACATTTGGCGTTTGAAGT CAGCATATGCATGAACCAAGTAGC V. unguiculata
AB056448-3 GCTTCAATTTGCTCCTTATGG TCATTTGTTTTATTTGAAGTGTCG V. unguiculata
AB056448-1 TGTTTTGGGCCATCAGAGG CGTCCAATTTATTCGCTACAAGTT V. unguiculata
AY189137-5 TGTTATCTGGGTTTTCTCGTCTTG TGATGGCGCTAGTGATGATTTT V. unguiculata
AY189137-3 GACACCGGGCGTATCCTT CTTGCTTATTATATGTTGCCTTAG V. unguiculata
AY189137-1 TTCGTGTCTGGGGGAGGAT TACGCCCGGTGTCATAGTGTT V. unguiculata
AY189138-3 GGCTGCAAGGGTTCTCAATG CAACAATATGCCTCCTTCTGC V. unguiculata
AY193835-3 AAATGGTTGCTTTCTCTGACA GCAACATTTGTATGGGGAACCT V. unguiculata
AY193836-3 GGGCAACCAAACCGTGTG ATGGAAGCAGAAATTTGAGTAAC V. unguiculata
AY193837-3 ATATCGGCGCCTCTTCCCTACAGT GACATAAAACTCCCACGAAATCAG V. unguiculata
AY193837-1 CCATTTGTACCACCCAGGAG ATCGGCAATGACAGGAACA V. unguiculata
AY257179-3 AACTCAGGCAGATCGCTTTGT TACATGTAATTTTTCTTCAACTCT V. unguiculata
AY257179-1 AGCCAAGCCTCTGCCATTC GACAAAGCGATCTGCCTGAGTT V. unguiculata
D83970-3 TCAACCAGTATAATCGCAAGACAT CCAGCGACATCATCACAACAATAA V. unguiculata
D83971-3 GCTTGGGGCTTGAATTTACTCCT AATGCAAACTCTTACAAACCACAC V. unguiculata
D83971-1 CCAGCTTTGAAGGGGACTCT TGGGCAATTGCAACATCTCT V. unguiculata
D83972-3 AGAAAGGGATAGTGGACAAGATTA TTACATATCCATTGGCAGAACATC V. unguiculata
D88121-3 CCTAACGATGTGGCAGAAGC ATGGCTAGATTTGAGTGAGGATTG V. unguiculata
D88122-3 CGCCGGACGAGGAGTAT AAAAGAAATTGGGTTAAAAAGTAT V. unguiculata
U30875-5 TTGGGATTAAGCTTCTGATTTTGA ATTGATGGCGCTAGTGATGATT V. unguiculata
U30875-3 TTTGGAGGCAAGGGCTACTA TTGCTTATTATATGTTGCCTTAGA V. unguiculata
U30875-1 CAATTCACGAGGCCTCTAAAAAG TACGCCCGGTGTCATAGTGTT V. unguiculata
U30895-5 TGCCACCGCTTCTGCTTATCTCAC CGCCCCCGAAGTCACAATG V. unguiculata
U30895-3 GGAGGCTGGGAAGATAGAAGACT ATGCGAGATCAAGATAAGCCTAAA V. unguiculata
U30895-1 ATTGTTGCCGGTGACGTTATTATT ACTCGGGGTATGTTATCTGTGAAA V. unguiculata
U30896-3 GTCGGGATATTGGCTGGAGAG GAGATGGGGGTGGATGAATAGTT V. unguiculata
U30896-1 TCGGCAGAAGGTTCCAAGTTT GATACCCCATTCAATTCTCCTCTG V. unguiculata
U33205-3 CTTTTTGGATTGGTGCGTGAT AACAATTTGATATGGGGAACCTC V. unguiculata
U61379-3 GGGCCCATGGACCTCAAAC TAAGAACGCAAAGAACGAAACATC V. unguiculata
U61379-5 GTCCGGTGGTTCTGCTCGTGTC CTGCCCGGGTGGAAAGGAAC V. unguiculata
U61396-3 AGAACCAGCTCAGCCGAAATCAGA GGAGCGTGCTATGGGCTTATG V. unguiculata
U61396-1 TTAAACCGCAAAACAGAGGAAGAG CATCGGGCTTGGCATTTTATC V. unguiculata
U61396-5 TTCCCTCATTTCTCTTCTTTTCTC GCCGGACCTGTACTTAGCAT V. unguiculata
U85250-3 AAATAGCATTGCCAGTGAAGA GTAAGCGACTAATGATAAAGATAA V. unguiculata
U92656-5 GTCCGCGAACACGAACA CCATTTGGGATTAGTAGTTTCATT V. unguiculata
U92656-3 GGGGTTGCCAGTGAAGGAGAG TGCTAACCAACACCACCAGATAAC V. unguiculata
X74821-3 ACCAGAGATGTTTTGAGTGATTAT GTACAAGCCAAGTACTCTAACAAG V. unguiculata
X74821-5 CCATCAATAGAATCGGAGTGTT GGCTGCCCCCATTCCTTTAT V. unguiculata
X79604-3 AATTGCGGCGTTACCTACC ATAGATATGATGATTCCCTGGTGA V. unguiculata
X86030-3 CTACCCCTACCTCTATAACTCACC CAATACAAACGCATGGGCTACAC V. unguiculata
X88800-3 TGGAGTGCGGCAGAGGACA TTATTGAAAAGTGATGGAAGTTAT V. unguiculata
X88800-5 GAGCGTAGCAGTGGTGGTGTGG TCCCCCGGTTGTTTCGTGAGAC V. unguiculata
X88801-3 CCCGGCCGATGTGTTGACCTCTA CAACATAACATGCGCCTACAAT V. unguiculata
X88802-1 CTCCGCCGACGACGCAACACA AGAAGCCGGCACTCCCAAAAACAG V. unguiculata
X88803-3 CAGCTGCTTTTGCCCATTTC TCCGCGAAGAGCATTACAA V. unguiculata
X89400-3 TTTGCGCAGAATTTATGGGTTTTG TAATTTTATCAATCAACGCATCTT V. unguiculata
X89400-1C CATCGAGACTTGAAACCACAGAA CAACTGGGGTAGAATAATGACGAG V. unguiculata
X90487-3 CTGGGCTCCTTTGTGGTTTTG CACACCCTTTTGCCCTTCTC V. unguiculata
X98608-3 CCCCACCCAACGAAGATGAA AAAGATACAAAGATACCCAGAAGC V. unguiculata
Y08624-1 AATCAAGGCTTCGTTTTCATCATC GCTTGCGCTGCCTTCTCTT V. unguiculata
Z21954-3 GTCGCAGGAAACCAGAACAG TGCGAATTTAGGGCATCAA V. unguiculata
CA906441-1 CTTCCCCACTGATTCATTTCTGTT CACTCCCCTTTGTTCTTGATTATG P. coccineus CA909226-1 GATCTCTTTCACGCTGGACAT CACATAGCGACATGCTAGAACACT P. coccineus CA910489-1 AGAAAAAGGAGGGAAATGCTGCTA TTGCCTCCGCTTTCTTTATCTTTC P. coccineus CA910598-1 CCTCCAATTCGGTTCCAGCACTTC GCCCTTGATCTTCCTCTTTTTACG P. coccineus CA911600-1 AACTGGTTCATCGCATTACTCTTC TGGCATCACCTTGTTTGGCATTAG P. coccineus CA911990-1 TTTCATCCCCAGCATCAGCAGTTT TGGGGGTGGGGCAGAATCA P. coccineus CA912170-1 TGGAGAAAGCATGGCAGGATACT CCCCGGAGGCAAGATTTCAA P. coccineus CA914165-1 GATAATTTTGACGAGGGAGCAGA GGTGGAGAACAGGAGCAAAGTATT P. coccineus CA912542-1 TTATTGGAGTTGCCCTGTTCTTCA GCCCCGTCTGCATTCCTTTGT P. coccineus CA906286-1 CGGGCGGGGGAGATTGT CCCAGCCCACGTAAGGTAAGAAC P. coccineus CA910928-1 CGGAATCTGGTCGGCCTACATCTA TGATACCCCTCCGGCAACAAG P. coccineus CA907856-1 ACACTTGAGGGAAAGCCATCA TTCCAAGCCAAAGCAATAAC P. coccineus M80799-1 ACGGCGGGCAATGTCAA AGTTCACGTGCTCCGCTGTTAT V. unguiculata CA906298-1 TAGTTGACCGGTTATCTGTTGTTT GCGCCTACGAGCATCACTTA P. coccineus Pc_CL2167-1 CATCACGCGATAATACACGACT TCTCTGCCGAAGGTTATGGA CA912634-1 GACGGAATGAAGTTGAAGGTGGAT AGATGGGCCAAGATATTTTAGACT P. coccineus CA913150-1 CGGAGATGGTGGCGTGTTATT CTTGTGCAGCCTTGTTTTCAGC P. coccineus CA911895-1 CATCTGCTTTTGCCCATTACG CACCAAGCCGCTCTAGCAAGTTAT P. coccineus CA911460-1 GAGCTGCAACAAGGAATGGATG ATGAGGGCTAACAGGAAAAGAAGG P. coccineus CA902380-1 GGCCCCCTCAGACAGAT AGATAAATTTGGGCTTGAA P. coccineus CA907467-1 AGCCTGTCAATTCCAAACTCCACT ATCCAACTTCAGCACCCAGACAAA P. coccineus CA907742-1 TCGGTTGAAAAAGAAAAGCAGAAG ATAGGCACTCAGGAAGGGAATGTC P. coccineus BQ481755-1 CATGGCGAAGTAACAAAAGTGGTC ATATGCAGGATGAGGAAGATGGTT P. vulgaris Pc_CL65-1 CTCGTCTCGCCACCCAGTTCT AGTCACCGCTCTCCACCCTTCT CA900698-1 TTGTTCTCAGCGCAATGTGG CTGAGCACCAAAGCCCCTGACA P. coccineus CA900229-1 TGCCGAGTCATCCCGAACAT CTCCAAAGCTGAAGCGAACCA P. coccineus CA914163-1 ATGGTGAATGCCTGAAACAAATAC CATGACAAATGCCTGAATAAGACT P. coccineus CA912177-1 CTTGGTCCGCCATACTCTGAT TTTTCTTTCCTCCTCTGGTATGG P. coccineus AJ225088-3 GGTATGATGGCCGTGAGTGGT TAAAGCTGTACAACGGCGATGAA V. unguiculata AJ225088-5 GAACACCACCGCCACACG GCACTAGCATTAACGCAAGAAACT V. unguiculata AF287258-3 TAGCGCCATGACATTTACCTCTTA GCTCCGTTTTCTGCTTTATCTACA V. unguiculata AF287258-5 CATTGGGCGAAAAGAAAGGAA CGCAAAAAGAAGGAGAGCAATAGA V. unguiculata AF287258-1 AGGGGGTGAGCTTGTTTTTG TATTGCGAAGCCTTTAGTAGACCT V. unguiculata

AF287258-1C GTTTGATGGCATACTGGGACTTG CTCTTCCTCTGTTTTGCGGTTTA V. unguiculata X91836-5 ATCGCGAAAAATGAATTATCTGGT TTATCGTCGGTGAAAACTGCT V. unguiculata X91836-5C CCGGAAACATGGCATTATTATTAG CCATTGCCTCGTTCCCATCTT V. unguiculata X91836-3 AGTCTCCACCTCCCCCTTCT TTTAGCATTTTGAGGCACTGATTC V. unguiculata AB027154-3 ATAAAATACCACAACAAAGGAGAT CTTATTGGTTGATGTAGTGGAT V. unguiculata AB028025-3 CGTTGCGCCGCCTTTTAGTTTAC TCAATTTGGCAGGATGAACC V. unguiculata AB030293-3 ATACGCCCCCGGAATCTAC TAGCGGAACAAAACAAGAAGAATA V. unguiculata AB030294-3 AAGGTGGTGAGGGCGAGAAG ACCAACCAAACCACAGAAATGAC V. unguiculata AB030295-5 TGGGCGAGCTTACAACATACACA CGGCCCCACTTCCACCTT V. unguiculata AB030295-3 TCGCATTCGCCCATCTGAG GCTTAGCATTGCTTACTCCATTCA V. unguiculata AB030295-1 GTGCCTGCTGACATTGAAACTG ACGGAATCCCCAAGCAAAGTC V. unguiculata AB038691-3 GACGAAGGCTTGGCTCTCC TTACCCACAAAAGTAAATCAAAT V. unguiculata AB038692-3 AAACATTTGGCGTTTGAAGT CAGCATATGCATGAACCAAGTAGC V. unguiculata AB056448-3 GCTTCAATTTGCTCCTTATGG TCATTTGTTTTATTTGAAGTGTCG V. unguiculata AB056448-1 TGTTTTGGGCCATCAGAGG CGTCCAATTTATTCGCTACAAGTT V. unguiculata AF077224 AGCTGAAGCCGCCACCATA AGCAGCAGCCTTAAACTCATCAA V. unguiculata X89400 ATTGTTAGGTTGCAGGATGTAGTG TGCCACAGAGAATTTGATAGAGGA V. unguiculata AF279252 AGCTTCGCCAACTCCTCCATCAT TCATCAGGGTCAAAGCTCATCAAT V. radiata AF151961 TTCCGCCATCACCTCTTCATTCC CCGGCGACGTCAGGGTTCTTA V. radiata AF402602 CCAGGGTTCAGGTTCCATCC CCCTGTTGCATACTTACGGTCTCT P. vulgaris D13557 GAACGGATCCCAGAACGAG GGAACACCCAAATCATCAAATAA V. radiata AY189907 TGCAGTGGGAGTATCAAGTT CTGCACCATTCCAGTCACCCT P. vulgaris U85250 ACTGGGCACAATTCCTATCTGACA GGTGGCCATCTTTGCAACTTTAGC V. unguiculata BQ481569 TTTGTGTTCGTCTGAGGCGTCTTC CCCGGGTCAGTTCATGTTGCTAC P. vulgaris AB056453 ATGGGCAGGGAGCGACAGATTTAT CAGTTGCCCCGGGTGCTTCC V. unguiculata AB062360 CCCTCGGCTATAGCATTGAAGAC ACGCATAAACAAAGAGGCTGGACT P. lunatus CA901109 CCAGCCAGCCATACCCATCT CTCGACCCACATAATCTTTCAGGA P. coccineus CA901635 TAGGAGAAATGGGGTTGGCTTGAA TAAACTTGCTCGGCGTGTAATGAT P. coccineus
CA901660 AACGGGGTCCCAGATGTTGT TCTCTAGGCCTTGTTGCTCA P. coccineus
CA910054 CCGCCGGCAAGAAAGTTATCAT GGCAGCACCATCAGCAAGGAA P. coccineus
CA912710 GGTGGCTGTTGGATGTGGATG CTGTGCCGCCAAGCATTCC P. coccineus
CA909621 GAGCTTCCGTTTCATACCATTAG CTTTCCCTCGCCTTCAAATA P. coccineus
D10266 TACGAGGCATTTGGTTTGACAGTG AGCCGGTTCCTCCATTTCTT V. radiata CK151423 TTTTCATGCGACTAGTTTATCCTT AACTGGCACTCTTTCTGGTCTTA V. unguiculata CB968060 ATTTCCGAGCTTGACACATT CAAAACCACCAAGCCACATTC V. unguiculata Pc_CL338 AGCTCCAGATACTTGTTGTTGTTC ATATGGGGGTGAGGGTGACG P. coccineus
CA906101 AACACGCGGTACTACGAAATCCTC CTCCGCGTCTCTGTCTCCTACCTC P. coccineus
Pc_CL593 ATCCTGTTTCGGTGCTGCTTCTTT GGCTTCCTCCGGCGATTCC P. coccineus
AF165998 TTTTGCCCAAGGAGGTAG TTTGATTTATAAGTGGGTAGAAGA V. unguiculata U08140 GACTTTGGCCTCTCCGTTTTCTTC ATCTATATGTCCCTTCAATACTGC V. radiata U10419 TCCCACGATCTGTTTGAGC CTGCATTAGTTTTTGTTTAGATTG P. vulgaris
U28645 GGGAAGCTTGGGAGAATAGTTTTG GATTTCCCTGCTGGTTTGGTCTCT P. vulgaris
U54703 TCGGTAAGAAAAAGGAAGAAGAGA ATGAGGTGGCAGCAGGTGGTG P. vulgaris
U70531 GGCTGAATTACTACCCTCCGTGTC AGCAGCTCTTGTATCTCCCATTTG P. vulgaris
U92656 CGATGAAGATCCGCGAAACTAT TCTGGCCTCTGGAATGTGTC V. unguiculata X53603 TTGGCCCCATTAAAATAGTGACA CCTGGTTGGGTGGGGTGCTC P. vulgaris U20809 TCCGCAACAAGAGTGAACAG GACAGCAGCAATATCTACCAACC V. radiata BQ481672 ATTTTTGGTGTGCTTTCGTTTAT TCCGTGGCTTGCTGATTAG P. vulgaris CA900138 CACCGTCACCATATTCTCTGC TCTCATTAGGCCGTGCTGAC P. coccineus
CA898279 CCGTAGGATTGCTGATGAGG TTCCGGTTATGTAAAAGATGTCC P. coccineus

CA898807 TGCTGGTTTTGCGGAGGTT CCCTGTAGGCTTCCATCATCTCTT P. coccineus
CA898809 GTACCCAGTCACCAATCATAAAG AATCCCAACAACCAAGTCATCAGA P. coccineus
CA899061 GTTCTCCCCACCCTTCACATC CACGTTGCAGGCTAATAAAATAAA P. coccineus
CA899306 GAAGGGCTTTGGATGGTC TGGGAGTTACGCAAAATAGGT P. coccineus
CA899524 CTCTTCTTCAGGCTCACTCACACT TCGCGATCGGAATAACAAG P. coccineus
AY193836 TGCTACGGCCATCCAACACC AGCCAACCAACCTCCACCAAC V. unguiculata CA901208 GGGGAGCTGTCACAAGAAGAAAT TGCACATCCCCAGTCAATA P. coccineus
CA902017 ACTTCAAGGAGATGGGAGGTTATT TGTGTTATTGGTTGAGGCAGGAG P. coccineus
CA902145 TTGATGACTTCCACCGGTTCTTA ATTTCGCTATGGAGGCTTCAA P. coccineus
CA902154 ACAAGGCTGATTTAGGTTCTCC TGTATCAGCGCGTCTTTCAT P. coccineus
CA906247 ATCTGGGCAAGCAAAGAGGT AGAAGTTCCCATTCGTAAGTTGTT P. coccineus
CA906300 TATTAGAGGGATTGAGTGGGATGC CAAGCTTCAAATATACTTCTTCAGG P. coccineus
BQ481774 GACGTCGTTGTGCTCTCG GTTGGGTACCCAAAAACTCG P. vulgaris Table 2: Lablab purpureus accessions used for screening the gene specific markers
Sl.No. Lablab accessions Country of origin 1 HA3 India 2 Mac-1 India 3 SRL India 4 Rongai Kenya 5 Highworth Ex.India 6 CPI36903 Ex. Ukrane 7 CPI52544 India 8 CPI52552 India 9 CPI52535 India 10 ILRI6536 Ex. Ethiopia
II. Phenotypic and molecular characterization of germplasm
The 224 accessions collected and maintained in the KT funded Dolichos were used for
screening with the selected primers. For screening of these 224 genotypes we have selected 13
primer pairs (Table 3) to see the genetic diversity within Lablab purpureus These 13 primers were
selected based on the research article we published in the journal of ‘Plant Genetic Resources:
Characterization & Utilization’. The criteria used for the selection of primer was PCR
amplification pattern like presence / absence of bands, banding patterns and polymorphism on
1.5% agarose gel among the 10 genotypes of Lablab purpureus (HA3, Mac-1, SRL, Rongai,
Highworth, CPI36903, CPI52544, CPI52552, CPI52535 and ILRI6536).

Table 3: Primers used to screen the 224 germplasm
Sl no. Sequence ID Species of origin Sequence information Sequence class 1 AF 067417 Lablab purpureus fril cDNA EST derived* 2 AY 583516 Lablab purpureus 5S rDNA Nuclear DNA: rDNA repeat3 AB 176567 Lablab purpureus Met 2 cDNA EST derived* 4 AY 049047 Lablab purpureus pDLT cDNA EST derived* 5 AY 189907-1 Phaseolus vulgaris Genic Genomic DNA** 6 BQ 481672 Phaseolus vulgaris Genic cDNA EST derived* 7 AY 193836-1 Vigna unguiculata Genic cDNA EST derived* 8 AF 327406-1 Vigna unguiculata Genic cDNA EST derived* 9 AJ 225088-3 Vigna unguiculata Genic cDNA EST derived* 10 AW 256794 Medicago truncatula TC(17) cDNA EST derived*** 11 AW 684360 Medicago truncatula CGA(8) cDNA EST derived*** 12 AW 127626 Medicago truncatula GTTT(7) cDNA EST derived*** 13 AW 186493 Glycine max CTT(13) cDNA EST derived*** *: aiming for intron spanning from genomic DNA template, **: gene where the exon & intron positions are known, primers amplify across an intron, ***: containing an SSR portion that is spanned by the primer
Molecular marker development
We have sequenced gene fragments from nine Lablab genotypes GL48, GL147, GL153,
HA-4, CPI24973, CPI31113, Highworth & Rongai (for the origin of these Lablab accession please
refer table VII) along with a sample from cowpea genotype C-152 corresponding to fril (mannose
lectin mRNA), pDLL (double headed Bowman-Birk inhibitor gene), pDLT (double headed
Bowman-Birk inhibitor gene), 5S rDNA (Lablab purpureus 5S ribosomal RNA gene), and two
metallothionine genes Met1, Met2 (these primers are designed based on Lablab sequence),
AF287258 (aspartic proteinase gene from Vigna unguiculata), AF151961 (1-aminocyclopropane-
1-carboxylic acid synthase gene) & D13557 (mRNA for catalase) (from Vigna radiata).
The sequencing was done using Big dye V 3.1 terminator mix which contains Taq
polymerase, dNTPs, ddNTPs labeled with fluorescent dye, PCR buffer, Sequence buffer and single
primer (i.e. either forward or reverse primer). First I amplified the genomic template DNA of the
parental genotypes with the specific primers and the PCR conditions were as follows: 0.1µmol/l of
each forward and reverse primer, 200 µmol/l each dNTP, 1U of Taq DNA polymerase (Invitrogen,
UK) and the appropriate reaction buffers, 20–50 ng of DNA, in a 10 µl volume. For all species and
all primer combinations the following touch-down cycling regime was used: 940 C/3 min; (940
C/30 s; 580 C/30 s; 720 C/60 s) repeat for 10 cycles reducing the annealing temperature by 0.50

C/cycle; (940 C/30 s; 530 C/30 s; 720 C/60 s) repeat 34 cycles; 720 C/10 min; 120 C/30 min. PCR
products (5 µl of each) were visualized on 2% agarose for amplification. Then 1µl of PCR
amplified product is used as template DNA, 1µl of Big dye, 1 µl of sequence buffer, 1µl of 3.2 µM
primer and the PCR conditions were as follows: 960 C/1 min; (960 C/10 s: 560 C/5 s; 600 C/4 min)
repeat for 24 cycles: 100 C 10 minutes to amplify for the sequencing experiment. The PCR
amplified DNA was sequenced using DNA analyzer from ABI. I have sequenced gene fragments
5S rDNA of amplicon size 300 base pair, fril of amplicon size 300 base pair, pDLL of amplicon
size 200 base pair, pDLT 200 base pair of amplicon size, AF287258 of amplicon size 400 base
pair, AF151961 of amplicon size 400 base pair, D13557 of amplicon size 800 base pair,
metallothionine genes Met1 of amplicon size 1000 base pair & Met2 of amplicon size 800 base
pair from the above nine Lablab genotypes plus one cowpea genotype
These DNA sequencing results showed single nucleotide polymorphism among the exotic
material in particular with CPI24973 being noticeably distinct; the Indian materials are identical
except in one case i.e. HA4 (table 6).
Table 6: DNA Sequence polymorphism among the Lablab accessions Met 1 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 CPI24973 C C T C C C G A A T T A C G T G G C A CPI31113 C T T C C T A A A C T A C A - - G T T Rongai T C - - C C G A T T A G T A - - A C A Highworth T C - - G C G G A T A G C A - - G C A HA-4 T C - - C C G A T T A G T A - - A C A GL48 T C - - C C G A T T A G T A - - A C A GL147 T C - - C C G A T T A G T A - - A C A GL153 T C - - C C G A T T A G T A - - A C A Met 2
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 CPI24973 T C T TTG G TT A T C G G - G A TCTG C A G T G CPI31113 T C T TTG G TT A T T A G - T G ---- C T A T T CPI60126 C G C --- - -- T A T A T G T A TCTG G T A T T Rongai T C T TTG G TT A T T A G - G A TCTG G A G A T Highworth T C T TTG G TT A T T A G - G A TCTG G A G A T HA-4 T C T TTG G TT A T T A G - G A TCTG G A G A T GL48 T C T TTG G TT A T T A G - G A TCTG G A G A T GL147 T C T TTG G TT A T T A G - G A TCTG G A G A T GL153 T C T TTG G TT A T T A G - G A TCTG G A G A T

D13557 AF287258 1 2 3 4 CPI24973 T T ------------- A CPI31113 C A AGCTCAACGTATA GRongai C A AGCTCAACGTATA A HA-4 C A AGCTCAACGTATA A GL48 C A AGCTCAACGTATA A GL147 C A AGCTCAACGTATA A GL153 C A AGCTCAACGTATA A AF151961 5S rRNA 1 2 CPI24973 C A CPI31113 T A CPI60126 C A Rongai C A Highworth C A HA-4 C G GL48 C A GL147 C A GL153 C A
The molecular diversity found among the Indian materials and the African materials can be
seen in Table IX. Based on the Met2 genic sequencing data at the end of my trip in JIC 2007 I
designed a pair of allele specific primers (Soleimani et al, 2003) to distinguish between CPI 24973
& the Indian genotypes by making use of single nucleotide polymorphism. When we amplify with
these allele specific primers the alleles were not distinguished as expected. Here we have used two
general primers and an allele specific primer in a PCR reaction (Soleimani et al, 2003). It was
expected a common band in both CPI 24973 and HA 4 and a lower band in one genotype
depending on the allele specific primer. But when it was amplified with HA4 allele specific primer
I got a common band and a lower band in both the genotypes, where as in case of CPI 24973 allele
specific primer I got a lower band in CPI 24973 as expected but I did not get the common band in
both the genotypes.
1 2 3 CPI24973 A T A CPI31113 T C G CPI60126 T C G Rongai T T A Highworth T T A HA-4 T T A GL48 T T A GL147 T T A GL153 T T A
1 2 3 4 CPI24973 - T G A Rongai C T G T Highworth C G A T HA-4 C T G T GL147 C T G T GL153 C T G T

Table 7: Molecular diversity within the selected Lablab accessions CPI24973 CPI31113 Rongai HA-4 GL48 GL147 GL153
CPI24973 0.00 0.68 0.42 0.46 0.42 0.42 0.42
CPI31113 0.68 0.00 0.56 0.59 0.56 0.56 0.56
Rongai 0.42 0.56 0.00 0.02 0.00 0.00 0.00
HA-4 0.46 0.59 0.02 0.00 0.02 0.02 0.02
GL48 0.42 0.56 0.00 0.02 0.00 0.00 0.00
GL147 0.42 0.56 0.00 0.02 0.00 0.00 0.00
GL153 0.42 0.56 0.00 0.02 0.00 0.00 0.00
III. Generation of mapping populations for constructing genetic map
a. Selection of distant genotypes for wide crosses based on molecular and phenotypic data
To construct a genetic map we need distant/ wide parents for making crosses to develop the
F2 mapping populations. If there is variability among the parents we are going to get variation
within the segregating populations (i.e. in F2 mapping population), if there is no variability among
the parents we can not expect variation within the segregating populations. Therefore selection of
distant genotypes for crosses is very much important in generating the mapping populations for
genetic map construction.
Based on the phenotypic data and genotypic data we have selected three genotypes GL 153,
GL 48& GL 147 in addition to HA-4 which is a released variety that is photo insensitive,
determinate, has early maturity and high fragrance. Morphological traits considered for selection
are Growth habit, Photo-sensitivity, crop duration, Pod fragrance and the 100 seed dry weight
(Table 4). These four parents were used as female parents and crossed with the CPI 24973, CPI
31113 and (this CPI material was brought from JIC during our 2006 visit). Therefore we made 12
crosses using these 7 parents (Table 5).

Table 4: Important phenotypic traits of parents used for crossing
Name of the genotypes Phenotypic characters GL 48 GL 147 GL 153 HA 4 CPI 24973 CPI 31113 CPI
60216 Growth habit Indeterminate Determinate Indeterminate Photo sensitivity Sensitive Insensitive Sensitive Crop duration Long (Annual) Short Long (Perennial) Pod fragrance Medium High Nil 100 seed weight (dry)
15.2g 14.5g 18.5g 19.8g 6.85g 6.93g 6.9g
Table 5: Details of the crosses attempted
♀/♂ CPI24973 CPI31113
GL48 X X
GL147 X X
GL153 X X
HA-4 X X
b. Development of F2 populations for genetic map construction
The F1 seeds along with their parents were germinated in water cups (4cm in diameter &
10cm in height) in the greenhouse. 30 days after sowing the seedlings were planted in the field. I
have selected the true F1 crosses by observing the phenotypes in the field, those which resemble
the female parents (due to selfing) were rejected and the other types were retained. There were
only 3 true F1 crosses and the successful F1 crosses are GL 153 X CPI 31113, HA-4 X CPI 31113
& HA-4 X as highlighted in Table VI. (The failure in crossing may be due to the inappropriate
selection of the female flower, that the female flower may be selected after the anther dehices since
the F1 plants were resembling the female parent). The phenotypic observations were recorded till
harvest. At the onset of flower initiation the whole plot was covered with net to avoid the possible
cross pollination. i.e. the F1 plants were allowed to self. DNA was extracted from the F1 plants and
the F2 seeds were collected and preserved.

Molecular marker development (Work done in JIC during Aug-Sept 2007) (contd.)
Work done in JIC during June-July 2008
We have designed 32 primer pairs based on the information available in cowpea genomic
database (http://cowpeagenomics.med.virginia.edu/CGKB) and received 26 primer pairs from
Dr.Mike Timko. We used all 58 primer pairs (details are in table X) to amplify products from nine
Lablab purpureus genotypes GL48, GL147, GL153, HA4, Rongai, High worth, CPI24973,
CPI31113, and a Cowpea genotype C-152. When these PCR amplicon was separated on 2%
agarose gel I have identified monomorphic band in 33 out of 58 primer combinations. In 13 out of
58 primer combinations there was no amplification in Lablab purpureus. But in 2 out of 58 primer
combinations there was no amplification even in cowpea (this may be due to failed PCR). In 17
out of 58 primer combinations, few Lablab purpureus were amplified. To see the length
polymorphism, when we separated the PCR amplicon on 5% horizontal non denaturing Poly
Acrylamide Gel four primer pairs showed polymorphism, in turn which were monomorphic on 2%
agarose gel (Table XI). Simultaneously we sequenced the genic fragments from nine Lablab
purpureus genotypes GL48, GL147, GL153, HA4, Rongai, Highworth, CPI24973, CPI31113, and
a Cowpea genotype C-152. These DNA sequencing results showed single nucleotide
polymorphism and indels among the exotic material and Indian materials.
By using the sequence data of CP5 gene fragment I have designed allele specific primers
(Ye et al, 2001) to distinguish between the CPI genotypes and the Indian genotypes and also by
using the earlier sequencing data of Met2 I have designed two more allele specific primers (Ye et
al, 2001) to distinguish between CPI24973 and the Indian genotypes. This time I have used four
primer combinations for allele specific primer, that means two allele specific primers and two
general primers (Ye et al, 2001), but in my earlier allele specific PCR I used only three primers
(one allele specific primers and two common primer).
Cowpea SSR primers used to screen Lablab purpureus genotypes Sl. no
Primer name
Sequence forward ( 5’ to 3’) Sequence reverse ( 5’ to 3’)
1 CP1 TTGGAGAGGGACAAAATTGC ATTGTGCCACGTGGTAGTGA 2 CP2 TCCTTTCTCTGCCTCAGGTG GCTCTTTGCCTCCATTCTCC 3 CP3 ACCGGTGAAAGTAGGACCAC GGCTGATTGATCTGGCCATC 4 CP4 GGAGTTAGCCCACTCTCGTTC CAGAGCCTCTCCACTACTACAGC 5 CP5 AGCTCCTCATCAGTGGGATG CATTGCCACCTCTTCTAGGG 6 CP6 GGGGGAGAGAGAGAGAGAGAGA TTCTCCCCCTATGTGGACCT 7 CP7 GAGGAGGAGGAGGATCTGACA CTTCTGCAGGCTTGTGGTTC 8 CP8 ACCATAGCAGCTCCCCATTC GGTCTAGGACAAGGTCCTGGTAG 9 CP9 AGGAGGAGGAGGAGGAGGAG GTCTGGGAGACCTGGATTAGC 10 CP10 AGGAGGAGGAGGAGGAGGAG GTCTGGGAGACCTGGATTAGC 11 CP11 CCACAGTGGAAAACCACACC GGTGGTGGTGGAGTGAAGAG 12 CP12 CACTCATCTGCCTCTCAACG GCAGTCTACTCCCAGTACCACAC 13 CP13 CACCACCACTCCAAAGAAGC CTAACGCTGCAGCCTCATCA 14 CP14 GGTTTATGGGTGGGAGGTGT GTATCTGGCTGTCGTGACCA 15 CP15 ACGAGGCTCAAGATCAGACG GGAGCTCTTCCTTGAGAGTGG 16 CP16 CACCCTTCACCACCGTTTAC GTACACGAGGTGCTCCATTCC 17 CP17 ACCACCACCACTGTTCTTCTTC TTACCTTCTGGACTGGGAGAGA 18 CP18 GTGTAAGAAGCATGCAGAGGTG AGTACTAGAATGGGGCCATGTG 19 CP19 CTTCCCTGTGTTGCTGTTACTG CTCGCACGAGTAGAAGAGAAGA 20 CP20 AAGTCCTTACACCACCACTCCA GAGTCCGAAGAACGACTTGTG 21 CP21 CTCTCTCCATTCCCAAGTGTCT GAAGAAGAACAGTGGTGGTGGT 22 CP22 AGATGTCGCTCCCTCTGTTG GAGGAGGAGGAGGAGGAGAAG 23 CP23 CGAGGGAGTACGAGTTCAGC GTCTCGACCTTGACGTCGTT 24 CP24 AGCGTCAGAGACTGGGAAGA GAAGAGAGAGTGGTGGTGGTG 25 CP25 GGTGGCAATGAGCCTTACAC AGATGTGGAGGGCATGGTAG 26 CP26 CGACCACCATAACTCGGAAG GGGAGAAACAGAGGGAGAGAG 27 CP27 CGCAACTCACCTTTCCTCAG CGTTGAGATGCAGGGAGAAC 28 CP28 CGCAACTCACCTTTCCTCAG CGTTGAGATGCAGGGAGAAC 29 CP29 TCTCGAGAACGATGGTGGAG AAGGACTGAAGCGAGAGACG 30 CP30 GCGTGTCTTGCCAACTTCTC ATATCCTCCTCCTCCACCAC 31 CP31 GCTACATAGTCCCCACCAAGC TGTCAGATCCTCCTCCTCCTC 32 CP32 CGACTCCACCTTGTTCTTGC GGTGGTGGTGACAGAGTGAAG 33 CP115 GGGAGTGCTCCGGAAAGT TTCCCTATGAACTGGGAGATCTAT 34 CP117 GTGGAAGGAATGGGTCCAG AGGAAATTTGCATTCCCTTGT 35 CP163 CACTTTCTCCTAAGCACTTTTGC AAGTGAAGCATCATGTTAGCC 36 CP171 GTAGGGAGTTGGCCACGATA CAACCGATGAAAAAGTGGACA 37 CP181 GGGTGCTTTGCTCACATCTT TCCATGTGTTTATGACGCAAA 38 CP197 TGAATGGAGCAACTTCTTGGA GTTGCACTTGGTTGCCCTAT 39 CP201 GGTTTCCTAGTTGGGAAGGAA ATTATGCCATGGAGGGTTCA 40 CP215 CAGAAGCGGTGAAAATTGAAC GCATGTTGCTTTGACAATGG 41 CP239 CACCCCCGTACACACACAC CACTTAAATTTTCACCAGGCATT 42 CP333 CAAAGGGTCATCAGGATTGG TTTAAGCAGCCAAGCAGTTGT 43 CP359 TGAAAACAACGATATGCAGAAG TCAGTCTTAGAATTGAGTTTTCTTCG 44 CP391 TGCCTATGCTTATGCCTGTG GATGCCTGTTACTTGCCTTCT

45 CP395 GTTGTGAGCTTCCCCAGATG AATTTTGAACCCACCACCAG 46 CP397 TCATGGGTTAAATTTGCTTCAA AAACCATGTGGTTGTTGCAC 47 CP403 TGCAATATGGACCAGAAGAAA ATGCCCCAACAACAACATTT 48 CP431 CCTCAACACCTTTTGGAAGGA CAAATGCACCTCCTGTGCTA 49 CP433 CAACTTCACAGCCCTCAA TTGAAGGTATGGCCTTTTGTTT 50 CP435 TGCTCATCGTGCTTTGTCTT CACTTCAGACTTAGAGCGAAGAA 51 CP443 GCTCGGATATGGTCCTGAAA TCAGTGTCAGCACCATCCC 52 CP573 CAGAATCCTTGTGAACCTG TTTCGCAATATGCCCTTTTC 53 CP605 AAAGAGATACACATGCCTAACA GACCAACAGCGACTTTGAGC 54 Y1 GATATAGAATAGCATATTTAACATATTAG GTTGAAAGTTTGATAGTAAAGTGG 55 Y21 GAGAACTTCACGCACAATAG CGCGGTAGCATGATTGAATTTTG 56 Y26 CTAAATTATAATATTCGTCGGTC GGTTAAGGAAAAGAGGGTAGG 57 Y31 CTATTGGAATCTTGCCGTTG CTTTACCTTTATGCAAACCAATTC 58 Y45 CGATTATCCTGGCTAACGATG GGATCTGAGATAGTGTGAC Amplification pattern of Cowpea SSR primers in Lablab purpureus genotypes
Band size (base pair)
Genotypes** Sl.no
Primer name
CP Lablab 1 2 3 4 5 6 7 8 9 10
Remarks
1 CP1 200 D (200, 400) - - - - ++ ++ + ++ - + U* 2 CP2 200 S (220) + + + + + + + +
(indel) + + Poly on PAGE,
indels 3 CP3 200 D (200, 800) + + + - ++ + ++ + + + U* 4 CP4 250 M(200-
1000) - - - - + + + + + + U*
5 CP5 300 S (300) + + + + + + + + + + SNPs 6 CP6 800 S (250) + + + + + + + + + + Mono on PAGE 7 CP7 200 M(200-
1000) + + + + + + + + + + U*
8 CP8 200 S (200) + + + + + + + + (indel)
+ + Poly on PAGE, indels
9 CP9 200 S (200) + + + + + + + + + + Mono on PAGE 10 CP10 200 S (200) + + + + + + + + + + Mono on PAGE 11 CP11 200 M (450) + - + - + + + + + + Mono on PAGE 12 CP12 300 S (700) - - - - + + + - - + Mono on PAGE 13 CP13 100 D (100-300) + + + + ++ ++ + + + + U* 14 CP14 600 S(800) + + + + + + + + + + Mono on PAGE 15 CP15 200 S (200) + + + + + + + + + + Mono on PAGE 16 CP16 200 S (250) + + + + + + + + + + Mono on PAGE 17 CP17 200 S (200) + + + - + + + + + + Mono on PAGE 18 CP18 220 S (200) + - + - + + + + - - Mono on PAGE 19 CP19 200 - - - - - - - - - - + No amp in Lablab 20 CP20 200 - - - - - - - - - - + No amp in Lablab 21 CP21 250 - - - - - - - - - - + No amp in Lablab 22 CP22 200 M(200-800) + + + - + + + + + + Mono on PAGE 23 CP23 200 S (200) + + + + + + + + + + Mono on PAGE
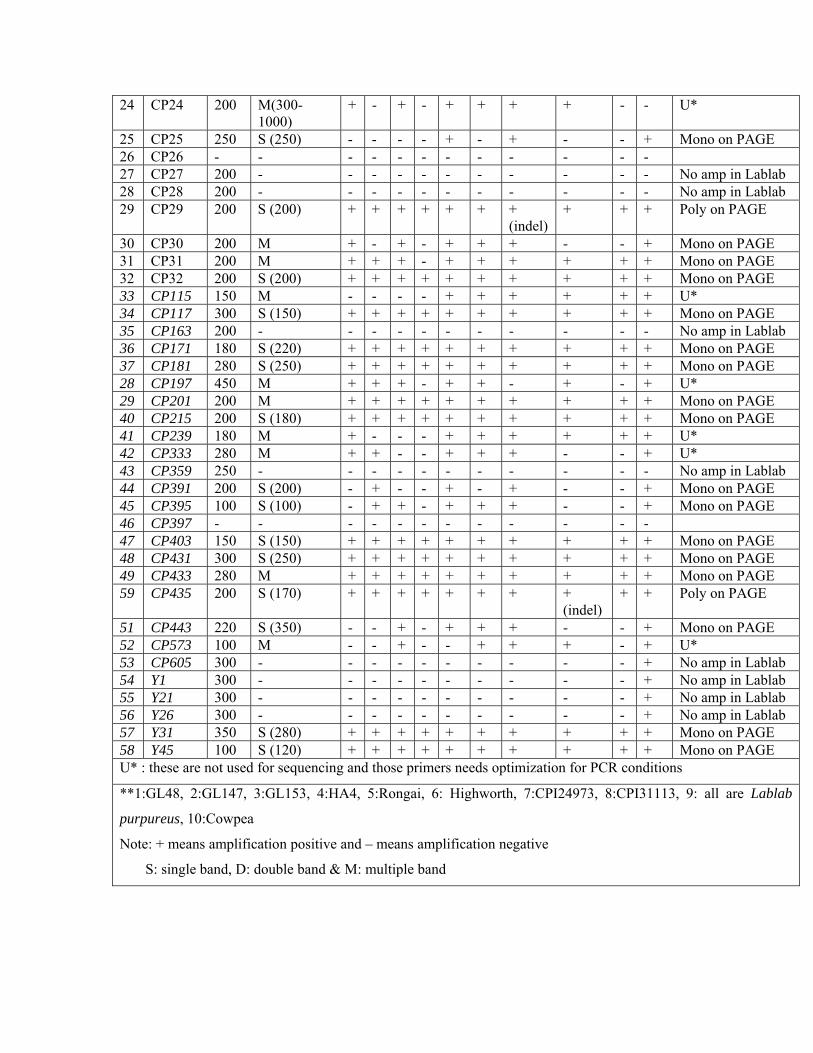
24 CP24 200 M(300-1000)
+ - + - + + + + - - U*
25 CP25 250 S (250) - - - - + - + - - + Mono on PAGE 26 CP26 - - - - - - - - - - - - 27 CP27 200 - - - - - - - - - - - No amp in Lablab 28 CP28 200 - - - - - - - - - - - No amp in Lablab 29 CP29 200 S (200) + + + + + + +
(indel) + + + Poly on PAGE
30 CP30 200 M + - + - + + + - - + Mono on PAGE 31 CP31 200 M + + + - + + + + + + Mono on PAGE 32 CP32 200 S (200) + + + + + + + + + + Mono on PAGE 33 CP115 150 M - - - - + + + + + + U* 34 CP117 300 S (150) + + + + + + + + + + Mono on PAGE 35 CP163 200 - - - - - - - - - - - No amp in Lablab 36 CP171 180 S (220) + + + + + + + + + + Mono on PAGE 37 CP181 280 S (250) + + + + + + + + + + Mono on PAGE 28 CP197 450 M + + + - + + - + - + U* 29 CP201 200 M + + + + + + + + + + Mono on PAGE 40 CP215 200 S (180) + + + + + + + + + + Mono on PAGE 41 CP239 180 M + - - - + + + + + + U* 42 CP333 280 M + + - - + + + - - + U* 43 CP359 250 - - - - - - - - - - - No amp in Lablab 44 CP391 200 S (200) - + - - + - + - - + Mono on PAGE 45 CP395 100 S (100) - + + - + + + - - + Mono on PAGE 46 CP397 - - - - - - - - - - - - 47 CP403 150 S (150) + + + + + + + + + + Mono on PAGE 48 CP431 300 S (250) + + + + + + + + + + Mono on PAGE 49 CP433 280 M + + + + + + + + + + Mono on PAGE 59 CP435 200 S (170) + + + + + + + +
(indel) + + Poly on PAGE
51 CP443 220 S (350) - - + - + + + - - + Mono on PAGE 52 CP573 100 M - - + - - + + + - + U* 53 CP605 300 - - - - - - - - - - + No amp in Lablab 54 Y1 300 - - - - - - - - - - + No amp in Lablab 55 Y21 300 - - - - - - - - - - + No amp in Lablab 56 Y26 300 - - - - - - - - - - + No amp in Lablab 57 Y31 350 S (280) + + + + + + + + + + Mono on PAGE 58 Y45 100 S (120) + + + + + + + + + + Mono on PAGE U* : these are not used for sequencing and those primers needs optimization for PCR conditions
**1:GL48, 2:GL147, 3:GL153, 4:HA4, 5:Rongai, 6: Highworth, 7:CPI24973, 8:CPI31113, 9: all are Lablab
purpureus, 10:Cowpea
Note: + means amplification positive and – means amplification negative
S: single band, D: double band & M: multiple band
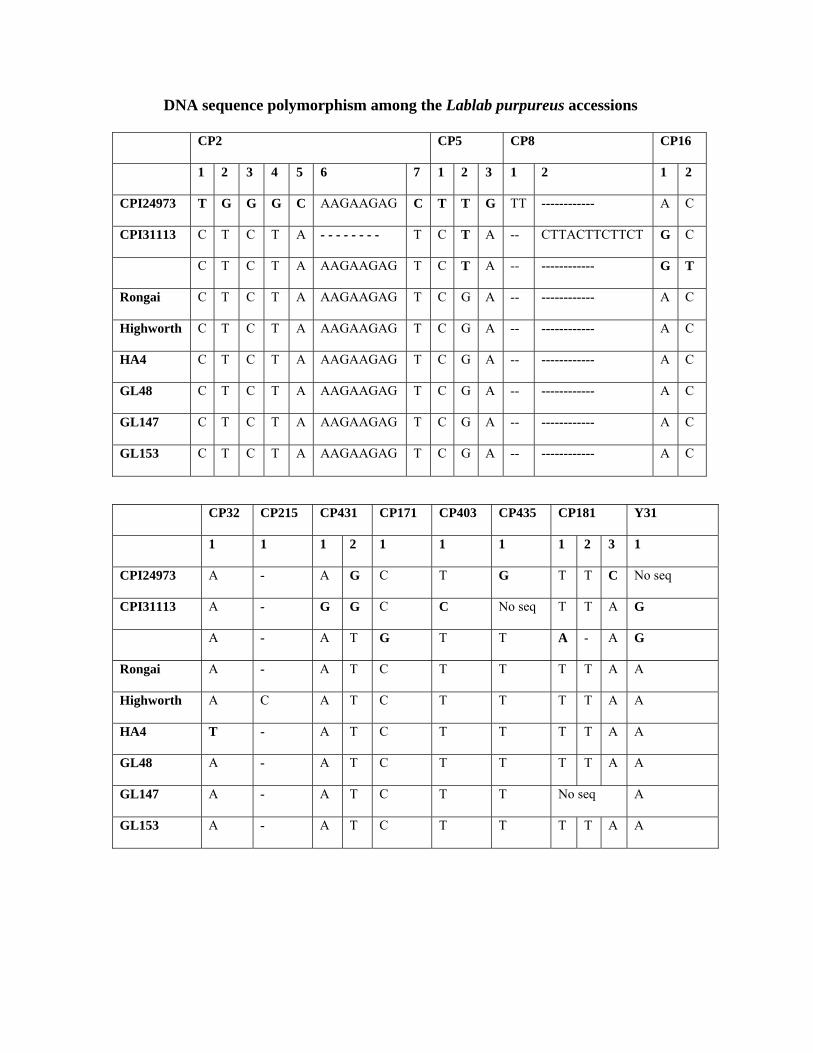
DNA sequence polymorphism among the Lablab purpureus accessions CP2 CP5 CP8 CP16
1 2 3 4 5 6 7 1 2 3 1 2 1 2
CPI24973 T G G G C AAGAAGAG C T T G TT ------------ A C
CPI31113 C T C T A - - - - - - - - T C T A -- CTTACTTCTTCT G C
C T C T A AAGAAGAG T C T A -- ------------ G T
Rongai C T C T A AAGAAGAG T C G A -- ------------ A C
Highworth C T C T A AAGAAGAG T C G A -- ------------ A C
HA4 C T C T A AAGAAGAG T C G A -- ------------ A C
GL48 C T C T A AAGAAGAG T C G A -- ------------ A C
GL147 C T C T A AAGAAGAG T C G A -- ------------ A C
GL153 C T C T A AAGAAGAG T C G A -- ------------ A C
CP32 CP215 CP431 CP171 CP403 CP435 CP181 Y31
1 1 1 2 1 1 1 1 2 3 1
CPI24973 A - A G C T G T T C No seq
CPI31113 A - G G C C No seq T T A G
A - A T G T T A - A G
Rongai A - A T C T T T T A A
Highworth A C A T C T T T T A A
HA4 T - A T C T T T T A A
GL48 A - A T C T T T T A A
GL147 A - A T C T T No seq A
GL153 A - A T C T T T T A A

: Molecular diversity among the nine genotypes based on Nei’s genetic distance
[Pair wise genetic distance matrix is a measurement of pair wise relatedness between individuals).
Summary of Amplification Success
Among the 58 primer pairs which I have used to screen the parental genotypes, four primer
pairs showed polymorphism on 5% horizontal non-denaturing Poly Acrylamide Gel, that means
hardly 7% length variant polymorphisms (The details are in Table XIV). In order to develop an F2
map from one or more of these populations, I need to generate more markers. The chromosome
number of Lablab purpureus 2n= 22, so we are going to get 12-15 linkage groups and if we
estimate 10 markers in each linkage group we need for about 150 polymorphic markers.
Pair wise genetic distance matrix
1 2 3 4 5 6 7 8 9
24973 31113 60216 GL48 GL147 GL153 HA4 HW Rongai
1. CPI 24973 0 0.6964 0.3008 0.6769 0.6836 0.4284 0.5038 0.5274 0.6017
2 .CPI 31113 0.6964 0 0.7045 0.7099 0.7052 0.7068 0.6908 0.6870 0.6958
3. 0.3008 0.7045 0 0.6676 0.6667 0.4296 0.4585 0.5078 0.5532
4. GL48 0.6769 0.7099 0.6676 0 0.3513 0.4533 0.4801 0.6543 0.3845
5. GL147 0.6836 0.7052 0.6667 0.3513 0 0.3950 0.5617 0.7165 0.4629
6. GL153 0.4284 0.7068 0.4296 0.4533 0.3950 0 0.2748 0.6886 0.2930
7. HA 0.5038 0.6908 0.4585 0.4801 0.5617 0.2748 0 0.6358 0.2030
8. HW 0.5174 0.6870 0.5078 0.6543 0.7165 0.6886 0.6358 0 0.6864
9. Rongai 0.6017 0.6958 0.5532 0.3845 0.4629 0.2930 0.2030 0.6864 0
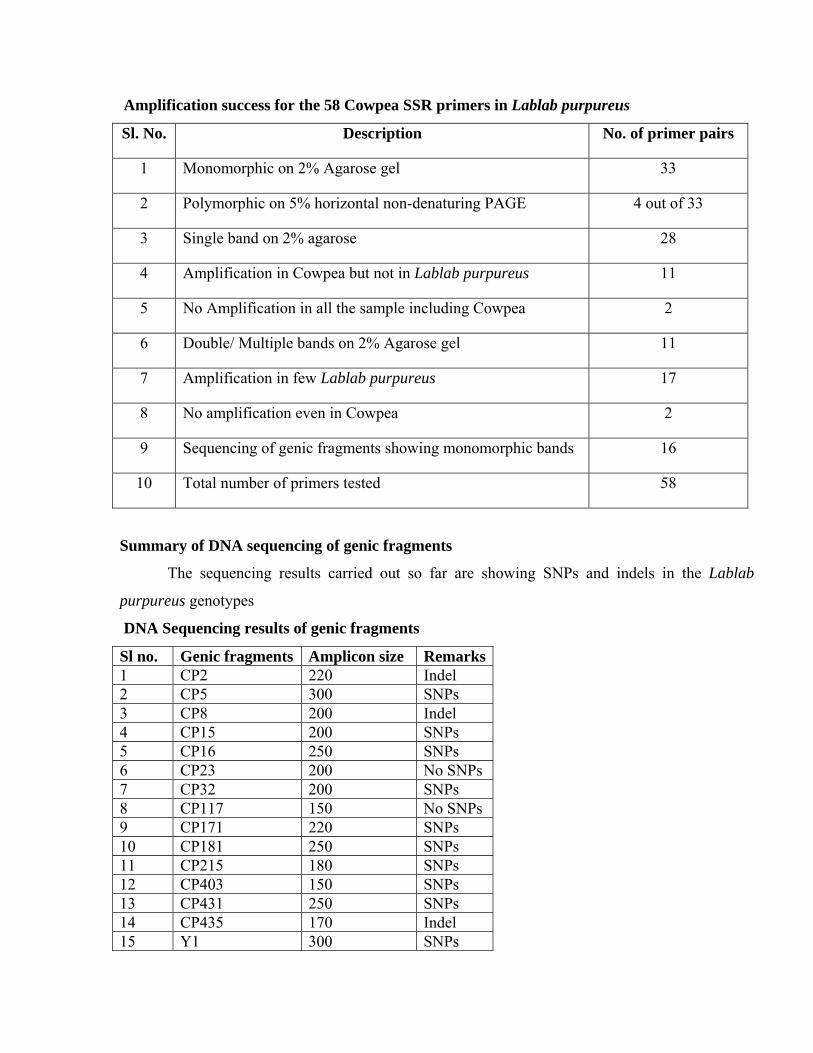
Amplification success for the 58 Cowpea SSR primers in Lablab purpureus
Sl. No. Description No. of primer pairs
1 Monomorphic on 2% Agarose gel 33
2 Polymorphic on 5% horizontal non-denaturing PAGE 4 out of 33
3 Single band on 2% agarose 28
4 Amplification in Cowpea but not in Lablab purpureus 11
5 No Amplification in all the sample including Cowpea 2
6 Double/ Multiple bands on 2% Agarose gel 11
7 Amplification in few Lablab purpureus 17
8 No amplification even in Cowpea 2
9 Sequencing of genic fragments showing monomorphic bands 16
10 Total number of primers tested 58
Summary of DNA sequencing of genic fragments
The sequencing results carried out so far are showing SNPs and indels in the Lablab
purpureus genotypes
DNA Sequencing results of genic fragments
Sl no. Genic fragments Amplicon size Remarks1 CP2 220 Indel 2 CP5 300 SNPs 3 CP8 200 Indel 4 CP15 200 SNPs 5 CP16 250 SNPs 6 CP23 200 No SNPs 7 CP32 200 SNPs 8 CP117 150 No SNPs 9 CP171 220 SNPs 10 CP181 250 SNPs 11 CP215 180 SNPs 12 CP403 150 SNPs 13 CP431 250 SNPs 14 CP435 170 Indel 15 Y1 300 SNPs

16 pDLL 200 SNPs 17 5S rRNA 300 SNPs 18 fril 300 SNPs 19 Met1 1000 SNPs 20 Met2 800 SNPs 21 AF 151961 400 SNPs 22 AF 287258 400 SNPs 23 D 13557 800 SNPs
Phenotypic characterization of F2 mapping populations:
We have taken 200 F2 seeds from two mapping populations HA 4 X CPI 31113 and HA 4
X CPI 60126 along with 10 seeds each from parents HA 4, CPI 31113 and CPI 60126. Seeds were
soaked on 03/08/2008 in water for 20 hours and then sowed in the polythene bags (12 X 8 cm)
containing soil mixture on 04/08/2008.
Three weeks old seedlings 140 each from two mapping populations were transplanted in
the field on 21/08/2008 at the spacing of 1.5 X 1.25 meter.DNA was extracted from all the F2
individuals and their parents during September- October 2008.
The phenotypic observations were recorded on the traits like
Stem pigmentation
Petiole pigmentation
Mid rib colour
Growth habit
Flower bud colour
First flower initiation
Pod curvature
Pod constriction
Pod fragrance
Number of seeds per pod
100 seed weight (Dry)
Seed colour
Length of the pod
Width of the pod
Branch orientation

Variability in F2 mapping population
Variability in F2mapping population
Separation of G-DNA on 0.8% agarose


Related Documents











