Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity towards the opportunistic pathogen, Hafnia alvei. Immunobiology (2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012 ARTICLE IN PRESS G Model IMBIO-51190; No. of Pages 7 Immunobiology xxx (2014) xxx–xxx Contents lists available at ScienceDirect Immunobiology jo ur nal ho me page: www.elsevier.com/locate/imbio Ficolin-3 activity towards the opportunistic pathogen, Hafnia alvei Mateusz Michalski a,b , Anna St. Swierzko a,∗ , Jolanta Lukasiewicz c , Aleksandra Man-Kupisinska c , Iwona Karwaciak c , Patrycja Przygodzka a , Maciej Cedzynski a a Institute of Medical Biology, Polish Academy of Sciences, Lodowa 106, 93-232 Lodz, Poland b Institute of Microbiology, Biotechnology and Immunology, University of Lodz, Banacha 12/16, 90-237 Lodz, Poland c Ludwik Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Weigla 12, 53-114 Wroclaw, Poland a r t i c l e i n f o Article history: Received 5 June 2014 Received in revised form 11 August 2014 Accepted 11 August 2014 Available online xxx Keywords: Complement Ficolin-3 H-ficolin Hafnia Innate immunity Lipopolysaccharide (LPS) Phagocytosis a b s t r a c t Ficolin-3 (also called H-ficolin or Hakata antigen) is a complement-activating pattern recognition molecule, possessing a fibrinogen-like domain involved in carbohydrate binding. Amongst human ficol- ins, Ficolin-3 has the highest concentration in serum and is the most potent lectin pathway activator in vitro. Evidence for its physiological function is sparse, although its deficiency has been suggested to increase susceptibility to infections. The specificity of Ficolin-3 is poorly characterized and currently few ligands are known. Here we report agglutination of Hafnia alvei, a Gram-negative enteric commensal bacterium and opportunist pathogen, in the presence of recombinant Ficolin-3 and calcium. Ficolin-3 also augmented phagocytosis of H. alvei by macrophages and displayed bactericidal activity. Addition- ally, Ficolin-3 inhibited host cells’ response to TLR4/MD-2/CD14-LPS dependent NF-B activation. This is the first demonstration of protective activity of Ficolin-3 against a human bacterial pathogen. Although human Ficolin-3 does not recognise and bind to common pathogenic bacteria, it could be an important component of innate immunity providing protection, for example, from commensal flora that can cause extraintestinal, opportunistic infections. © 2014 Elsevier GmbH. All rights reserved. Introduction Ficolin-3 (H-ficolin, Hakata antigen, thermolabile -2 macroglobulin) was first detected as an autoantigen precipi- tated by antibodies in sera of some systemic lupus erythematosus (SLE) patients (Yae et al. 1991). It is an oligomer (tetra- to octamer) of basic subunits, each consisting of three identical 34 kDa polypep- tide chains (Sugimoto et al. 1998; Hummelshoj et al. 2008). Like other ficolins, Ficolin-3 possesses an N-terminal cysteine-rich collagen-like domain and a C-terminal fibrinogen-like domain (Sugimoto et al. 1998). The collagen-like region forms complexes with Mannose-binding lectin (MBL)-associated serine proteases (MASP), which, after interaction of the fibrinogen-like domain with target structures, become activated and initiate the lectin pathway of complement activation (Matsushita et al. 2002). Although the related human Ficolin-1 and Ficolin-2 are >80% mutually identical Abbreviations: MASP, Mannose-binding lectin-associated serine protease; MBL, mannose-binding lectin. ∗ Corresponding author. Tel.: +48 42 2723607; fax: +48 42 2723630. E-mail address: [email protected] (A. St. Swierzko). in sequence, Ficolin-3 has only 48% sequence identity to the others (Sugimoto et al. 1998). In contrast to Ficolins-1 and -2 (and the collectins), Ficolin-3 is resistant to bacterial collagenases, possibly supporting its potential importance at sites of infection (Hummelshoj et al. 2008). Ficolin-3 was shown to be synthesized by hepatocytes, bile duct epithelial cells, ciliated bronchial- and type II alveolar epithe- lial cells. The level of its synthesis in lung exceeds that in the liver (Akaiwa et al. 1999; Hummelshoj et al. 2008). A frame-shift mutation in exon 5 (+1637delC) of the FCN3 gene causes Ficolin-3 deficiency and was suggested to increase susceptibility to severe recurrent infections and neonatal invasive enterocolitis (Munthe- Fog et al. 2009; Schlapbach et al. 2011). Although Ficolin-3 has been implicated in innate immunity (Matsushita et al. 2002), it does not seem to recognize common bacterial pathogens (Krarup et al. 2005). It binds avidly to an extra- cellular polysaccharide (EPS) of one strain of Aerococcus viridans (Tsujimura et al. 2001), isolated from contaminated human plasma. It also binds certain lipopolysaccharides (LPS) including those from Salmonella typhimurium, S. minnesota and Escherichia coli O111. Beside complement activation upon binding to isolated A. viridans EPS, bactericidal activity of Ficolin-3 was demonstrated http://dx.doi.org/10.1016/j.imbio.2014.08.012 0171-2985/© 2014 Elsevier GmbH. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

I
F
MAPa
b
c
a
ARRAA
KCFHHILP
I
mt(otoc(w(tor
m
h0
ARTICLE IN PRESSG ModelMBIO-51190; No. of Pages 7
Immunobiology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Immunobiology
jo ur nal ho me page: www.elsev ier .com/ locate / imbio
icolin-3 activity towards the opportunistic pathogen, Hafnia alvei
ateusz Michalskia,b, Anna St. Swierzkoa,∗, Jolanta Lukasiewiczc,leksandra Man-Kupisinskac, Iwona Karwaciakc,atrycja Przygodzkaa, Maciej Cedzynskia
Institute of Medical Biology, Polish Academy of Sciences, Lodowa 106, 93-232 Lodz, PolandInstitute of Microbiology, Biotechnology and Immunology, University of Lodz, Banacha 12/16, 90-237 Lodz, PolandLudwik Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Weigla 12, 53-114 Wroclaw, Poland
r t i c l e i n f o
rticle history:eceived 5 June 2014eceived in revised form 11 August 2014ccepted 11 August 2014vailable online xxx
eywords:omplementicolin-3
a b s t r a c t
Ficolin-3 (also called H-ficolin or Hakata antigen) is a complement-activating pattern recognitionmolecule, possessing a fibrinogen-like domain involved in carbohydrate binding. Amongst human ficol-ins, Ficolin-3 has the highest concentration in serum and is the most potent lectin pathway activatorin vitro. Evidence for its physiological function is sparse, although its deficiency has been suggested toincrease susceptibility to infections. The specificity of Ficolin-3 is poorly characterized and currently fewligands are known. Here we report agglutination of Hafnia alvei, a Gram-negative enteric commensalbacterium and opportunist pathogen, in the presence of recombinant Ficolin-3 and calcium. Ficolin-3also augmented phagocytosis of H. alvei by macrophages and displayed bactericidal activity. Addition-
-ficolinafnia
nnate immunityipopolysaccharide (LPS)hagocytosis
ally, Ficolin-3 inhibited host cells’ response to TLR4/MD-2/CD14-LPS dependent NF-�B activation. This isthe first demonstration of protective activity of Ficolin-3 against a human bacterial pathogen. Althoughhuman Ficolin-3 does not recognise and bind to common pathogenic bacteria, it could be an importantcomponent of innate immunity providing protection, for example, from commensal flora that can causeextraintestinal, opportunistic infections.
© 2014 Elsevier GmbH. All rights reserved.
ntroduction
Ficolin-3 (H-ficolin, Hakata antigen, thermolabile �-2acroglobulin) was first detected as an autoantigen precipi-
ated by antibodies in sera of some systemic lupus erythematosusSLE) patients (Yae et al. 1991). It is an oligomer (tetra- to octamer)f basic subunits, each consisting of three identical 34 kDa polypep-ide chains (Sugimoto et al. 1998; Hummelshoj et al. 2008). Likether ficolins, Ficolin-3 possesses an N-terminal cysteine-richollagen-like domain and a C-terminal fibrinogen-like domainSugimoto et al. 1998). The collagen-like region forms complexesith Mannose-binding lectin (MBL)-associated serine proteases
MASP), which, after interaction of the fibrinogen-like domain with
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
arget structures, become activated and initiate the lectin pathwayf complement activation (Matsushita et al. 2002). Although theelated human Ficolin-1 and Ficolin-2 are >80% mutually identical
Abbreviations: MASP, Mannose-binding lectin-associated serine protease; MBL,annose-binding lectin.∗ Corresponding author. Tel.: +48 42 2723607; fax: +48 42 2723630.
E-mail address: [email protected] (A. St. Swierzko).
ttp://dx.doi.org/10.1016/j.imbio.2014.08.012171-2985/© 2014 Elsevier GmbH. All rights reserved.
in sequence, Ficolin-3 has only 48% sequence identity to theothers (Sugimoto et al. 1998). In contrast to Ficolins-1 and -2(and the collectins), Ficolin-3 is resistant to bacterial collagenases,possibly supporting its potential importance at sites of infection(Hummelshoj et al. 2008).
Ficolin-3 was shown to be synthesized by hepatocytes, bileduct epithelial cells, ciliated bronchial- and type II alveolar epithe-lial cells. The level of its synthesis in lung exceeds that in theliver (Akaiwa et al. 1999; Hummelshoj et al. 2008). A frame-shiftmutation in exon 5 (+1637delC) of the FCN3 gene causes Ficolin-3deficiency and was suggested to increase susceptibility to severerecurrent infections and neonatal invasive enterocolitis (Munthe-Fog et al. 2009; Schlapbach et al. 2011).
Although Ficolin-3 has been implicated in innate immunity(Matsushita et al. 2002), it does not seem to recognize commonbacterial pathogens (Krarup et al. 2005). It binds avidly to an extra-cellular polysaccharide (EPS) of one strain of Aerococcus viridans(Tsujimura et al. 2001), isolated from contaminated human plasma.
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
It also binds certain lipopolysaccharides (LPS) including thosefrom Salmonella typhimurium, S. minnesota and Escherichia coliO111. Beside complement activation upon binding to isolated A.viridans EPS, bactericidal activity of Ficolin-3 was demonstrated

ING ModelI
2 unobio
(F(notFiiL
M
N
Fnt0
B
PoBcp
P
mo
G
PSkb
aHttk
C
lLUlCdtm
ARTICLEMBIO-51190; No. of Pages 7
M. Michalski et al. / Imm
Tsujimura et al. 2002). Previously, we reported the interaction oficolin-3 with the O-specific polysaccharides of Hafnia alvei LPSstrains 23, PCM 1200, 1203, 1205) (Swierzko et al. 2012), a Gram-egative commensal of the human gut and a rare but significantpportunistic pathogen (Gunthard and Pennekamp 1996). We usedhis model to obtain insight into the possible biological role(s) oficolin-3 in innate immunity. Specifically, we investigated the abil-ty of Ficolin-3 to agglutinate bacteria, its bactericidal effect, andts contribution to phagocytosis as well as its modifying effect onPS-dependent cell activation.
aterials and methods
ormal human serum
Normal human serum (NHS) was used as a source of humanicolin-3. Serum was obtained from a healthy volunteer withormal Ficolin-3 and -2 levels (25 �g/ml and 3.9 �g/ml, respec-ively) and low Ficolin-1 and MBL concentrations (0.6 �g/ml and.4 �g/ml, respectively).
acterial strains and culture conditions
H. alvei strains PCM 1200 and PCM 1222 came from theolish Collection of Microorganisms (Ludwik Hirszfeld Institutef Immunology and Experimental Therapy, Wroclaw, Poland).acteria were cultured in LB medium. Some experiments (flowytometry) were performed after inactivation of bacteria with 0.5%henol for 30 min at 37 ◦C.
reparation of LPS
LPS was extracted from bacterial cells by the hot phenol/waterethod (Westphal and Jann 1965) and purified as described previ-
usly (Petersson et al. 1997).
FP-expressing bacteria
To obtain the green fluorescent protein (GFP)-tagged H. alveiCM 1200 and PCM 1222 bacteria, the pGLO plasmid was used.ince H. alvei strains are naturally resistant to ampicillin, theanamycin resistance gene was cloned into pGLO. It was amplifiedy PCR, using the following primers:
Forward: 5′-GCCAAGCTTTCAGAAGAACT-3′
Reverse: 5′-ATGAATTCGATGATTGAACAAGA-3′.The product of amplification was cloned into the pGLO plasmid
nd then subjected to a double restriction digestion with EcoRI andindIII enzymes (Thermo Scientific, USA). The ligation was used
o transform the competent H. alvei cells (CaCl2 method). Posi-ive clones were screened on solid LB medium containing 50 �g/mlanamycin and 0.05% arabinose.
ell lines and culture conditions
The human macrophage-like cell line, U-937, came from the col-ection of the Laboratory of Immunobiology of Infections (IMB PAS,odz, Poland). The cells were routinely grown in DMEM (Gibco,SA) supplemented with 10% fetal bovine serum (FBS) (Gibco),
-glutamine/penicillin/streptomycin (Gibco). HEK293/TLR4-MD2-
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
D14 and HEK293/null cells (Invivogen, USA) were grown asescribed above, in DMEM additionally supplemented with blas-icidin, hygromycin B and normocin (Invivogen), according to the
anufacturer’s protocol.
PRESSlogy xxx (2014) xxx–xxx
Agglutination assay
The agglutination assay was, in general, performed as describedby Ariki et al. (2011) GFP-expressing bacteria were incubatedwith 100 ng of recombinant Ficolin-3 (R&D System, USA) in 5 mMTris–HCl, pH 7.4, containing 0.15 M NaCl, 2 mM CaCl2, and 2%(w/v) BSA at 37 ◦C for 4 h and observed in a TE-200U fluores-cent microscope (Nikon, Japan). In some experiments (to estimatewhether agglutination of bacteria is calcium-dependent), 2 mMEDTA (Sigma–Aldrich, USA) replaced CaCl2 in the assay buffer (Arikiet al. 2011).
Agglutination rate was estimated spectrophotometrically bymeasurement of bacterial sedimentation, as described by Ofek et al.(2001), modified. Briefly, bacteria were suspended to a density of1.3 OD600 in Tris-buffered saline (TBS), supplemented with 10 mMCaCl2, 5 mM MgCl2 and 0.1% BSA (pH 7.4). In some experiments,20 mM EDTA replaced CaCl2 and MgCl2. Next, 50 �l of bacterialsuspension was transferred to polystyrene cuvettes (Eppendorf,Germany). After addition of 50 �l (2 �g) of recombinant Ficolin-3 (R&D System) or buffer alone, mixtures were gently rotated at37 ◦C for 45 min. The aggregation of bacteria was then monitored(at RT) by recording the optical density at 600 nm for 2 h, using theEppendorf BioPhotometer. Starting OD was considered as 100%.
Bactericidal assay
H. alvei PCM 1200 bacteria were grown in LB medium overnightand then diluted in fresh LB (1:25) and further cultured for20 min. Thereafter, 5 �l 100-fold dilutions of bacteria were addedto serially diluted sera in a total volume of 105 �l in microplates(Nunc, Denmark). After 30 min of incubation at 37 ◦C, Alamar Bluereagent (Life Technologies, USA) was added. Survival of bacte-ria was measured according to the manufacturer’s protocol. Toinhibit the activity of classical and alternative pathways of comple-ment, serum was pre-treated with sodium polyanethole sulfonate(SPS, Sigma–Aldrich) (0.4 mg/ml) for 20 min on ice (Palarasah et al.2010). Ficolin-3- (and H. alvei PCM 1200-reactive antibodies)-depleted serum was prepared by thrice incubation of pre-diluted1:10 serum with 5 mg of dry mass of H. alvei PCM 1200 for 30 min at4 ◦C, followed by centrifugation (10,000 × g, 2 min). Alternatively,Ficolin-3 was depleted by incubation of serum in an anti-Ficolin-3mAb (clone 4H5, Hycult, The Netherlands) coated-plate. Antibody-depleted serum was performed similarly, using protein A/G agarose(Sigma–Aldrich). Serum was diluted in a buffer containing 25 mMHepes, 5 mM CaCl2, 5 mM MgCl2 and 155 mM NaCl (pH 7.4) unlessotherwise indicated.
C4 deposition assay
Ficolin-3-dependent lectin pathway activation by bacterial cellswas estimated in a modified C4 deposition assay (Petersen et al.2001; Swierzko et al. 2012). Briefly, the microtitre plates (Nunc)were coated with dry mass (10 �g/ml) of paraformaldehyde-inactivated bacteria (according to Neth et al. 2000) and incubatedovernight at 4 ◦C. The plates were washed with TBS supplementedwith 5 mM Ca2+ (TBS-Ca2+) and 0.05% Tween-20 and then blockedwith 0.1% BSA in TBS-Ca2+. Normal or Ficolin-3- (and H. alveiPCM 1200-reactive antibodies)-depleted human serum (10-folddiluted in 0.1% BSA/MBL-binding buffer) was added and incubatedovernight (4 ◦C). The MBL-binding buffer, containing 20 mM Tris,1 M NaCl, 10 mM CaCl2, pH 7.4 (high ionic strength) was used toprevent the activation of complement via the classical pathway
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
(Petersen et al. 2001).Next, low Ficolin-3 serum (Swierzko et al. 2012), pre-diluted
1:3000 in 0.1% BSA/TBS-Ca2+, was added as a source of C4.After incubation for 2 h at 37 ◦C, deposited C4c was detected
ING ModelI
nobio
wIeauU
P
(F(AcGaiSacCmww(
P
micoSfiawTMriwC
N
swtwBotFawUMaBtnq
(SD) from at least three experiments. The statistical significanceof differences was assessed by using Tukey’s multiple comparisontest after analysis of variance; p values of <0.05 were consideredsignificant.
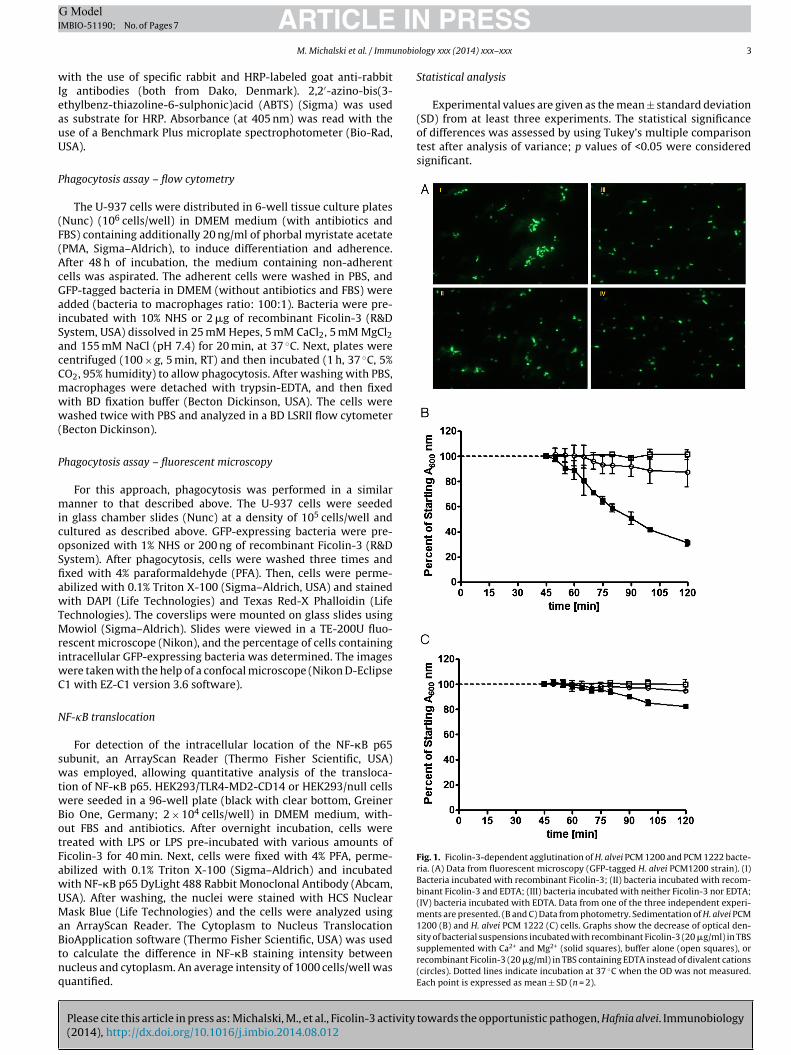
Fig. 1. Ficolin-3-dependent agglutination of H. alvei PCM 1200 and PCM 1222 bacte-ria. (A) Data from fluorescent microscopy (GFP-tagged H. alvei PCM1200 strain). (I)Bacteria incubated with recombinant Ficolin-3; (II) bacteria incubated with recom-binant Ficolin-3 and EDTA; (III) bacteria incubated with neither Ficolin-3 nor EDTA;(IV) bacteria incubated with EDTA. Data from one of the three independent experi-ments are presented. (B and C) Data from photometry. Sedimentation of H. alvei PCM
ARTICLEMBIO-51190; No. of Pages 7
M. Michalski et al. / Immu
ith the use of specific rabbit and HRP-labeled goat anti-rabbitg antibodies (both from Dako, Denmark). 2,2′-azino-bis(3-thylbenz-thiazoline-6-sulphonic)acid (ABTS) (Sigma) was useds substrate for HRP. Absorbance (at 405 nm) was read with these of a Benchmark Plus microplate spectrophotometer (Bio-Rad,SA).
hagocytosis assay – flow cytometry
The U-937 cells were distributed in 6-well tissue culture platesNunc) (106 cells/well) in DMEM medium (with antibiotics andBS) containing additionally 20 ng/ml of phorbal myristate acetatePMA, Sigma–Aldrich), to induce differentiation and adherence.fter 48 h of incubation, the medium containing non-adherentells was aspirated. The adherent cells were washed in PBS, andFP-tagged bacteria in DMEM (without antibiotics and FBS) weredded (bacteria to macrophages ratio: 100:1). Bacteria were pre-ncubated with 10% NHS or 2 �g of recombinant Ficolin-3 (R&Dystem, USA) dissolved in 25 mM Hepes, 5 mM CaCl2, 5 mM MgCl2nd 155 mM NaCl (pH 7.4) for 20 min, at 37 ◦C. Next, plates wereentrifuged (100 × g, 5 min, RT) and then incubated (1 h, 37 ◦C, 5%O2, 95% humidity) to allow phagocytosis. After washing with PBS,acrophages were detached with trypsin-EDTA, and then fixedith BD fixation buffer (Becton Dickinson, USA). The cells wereashed twice with PBS and analyzed in a BD LSRII flow cytometer
Becton Dickinson).
hagocytosis assay – fluorescent microscopy
For this approach, phagocytosis was performed in a similaranner to that described above. The U-937 cells were seeded
n glass chamber slides (Nunc) at a density of 105 cells/well andultured as described above. GFP-expressing bacteria were pre-psonized with 1% NHS or 200 ng of recombinant Ficolin-3 (R&Dystem). After phagocytosis, cells were washed three times andxed with 4% paraformaldehyde (PFA). Then, cells were perme-bilized with 0.1% Triton X-100 (Sigma–Aldrich, USA) and stainedith DAPI (Life Technologies) and Texas Red-X Phalloidin (Life
echnologies). The coverslips were mounted on glass slides usingowiol (Sigma–Aldrich). Slides were viewed in a TE-200U fluo-
escent microscope (Nikon), and the percentage of cells containingntracellular GFP-expressing bacteria was determined. The images
ere taken with the help of a confocal microscope (Nikon D-Eclipse1 with EZ-C1 version 3.6 software).
F-�B translocation
For detection of the intracellular location of the NF-�B p65ubunit, an ArrayScan Reader (Thermo Fisher Scientific, USA)as employed, allowing quantitative analysis of the transloca-
ion of NF-�B p65. HEK293/TLR4-MD2-CD14 or HEK293/null cellsere seeded in a 96-well plate (black with clear bottom, Greinerio One, Germany; 2 × 104 cells/well) in DMEM medium, with-ut FBS and antibiotics. After overnight incubation, cells werereated with LPS or LPS pre-incubated with various amounts oficolin-3 for 40 min. Next, cells were fixed with 4% PFA, perme-bilized with 0.1% Triton X-100 (Sigma–Aldrich) and incubatedith NF-�B p65 DyLight 488 Rabbit Monoclonal Antibody (Abcam,SA). After washing, the nuclei were stained with HCS Nuclearask Blue (Life Technologies) and the cells were analyzed using
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
n ArrayScan Reader. The Cytoplasm to Nucleus TranslocationioApplication software (Thermo Fisher Scientific, USA) was usedo calculate the difference in NF-�B staining intensity betweenucleus and cytoplasm. An average intensity of 1000 cells/well wasuantified.
PRESSlogy xxx (2014) xxx–xxx 3
Statistical analysis
Experimental values are given as the mean ± standard deviation
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
1200 (B) and H. alvei PCM 1222 (C) cells. Graphs show the decrease of optical den-sity of bacterial suspensions incubated with recombinant Ficolin-3 (20 �g/ml) in TBSsupplemented with Ca2+ and Mg2+ (solid squares), buffer alone (open squares), orrecombinant Ficolin-3 (20 �g/ml) in TBS containing EDTA instead of divalent cations(circles). Dotted lines indicate incubation at 37 ◦C when the OD was not measured.Each point is expressed as mean ± SD (n = 2).
ING ModelI
4 unobio
R
Fa
e(I(PtFwo
Ps1uoccrbm
F1pvsda
ARTICLEMBIO-51190; No. of Pages 7
M. Michalski et al. / Imm
esults
icolin-3 agglutinates bacteria and contributes to the bactericidalctivity of serum
Recombinant Ficolin-3 was demonstrated to agglutinate GFP-xpressing H. alvei PCM 1200 bacteria, in the presence of CaCl2Fig. 1A-I). Addition of EDTA markedly reduced that ability (Fig. 1A-I). No agglutination was observed in the absence of Ficolin-3Fig. 1A-III and A-IV) or after incubation of Ficolin-3 with H. alveiCM 1222 bacteria (data not shown). Furthermore, by measuringhe optical density of the bacterial suspension, it was shown thaticolin-3 agglutinated approximately 70% of H. alvei PCM 1200 cellsithin 2 h (Fig. 1B). A more modest effect (approx. 20%) was also
bserved with H. alvei PCM 1222 (Fig. 1C).To determine the anti-bacterial activity of Ficolin-3, H. alvei
CM 1200 bacteria were incubated with serially diluted NHS (i),erum depleted of Ficolin-3-MASP complexes and H. alvei PCM200-recognizing antibodies (ii), serum depleted of immunoglob-lins (iii) or serum pre-treated with SPS (iv). The effectivenessf absorption (Ficolin-3 concentration, normal immunoglobulinsoncentration and ability of absorbed sera to activate C3) was
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
hecked in a specific ELISA (data not shown). The survival of bacte-ia was estimated spectrophotometrically with the use of Alamarlue reagent. Pre-absorption of serum with H. alvei PCM 1200 dryass significantly reduced its bactericidal activity in comparison
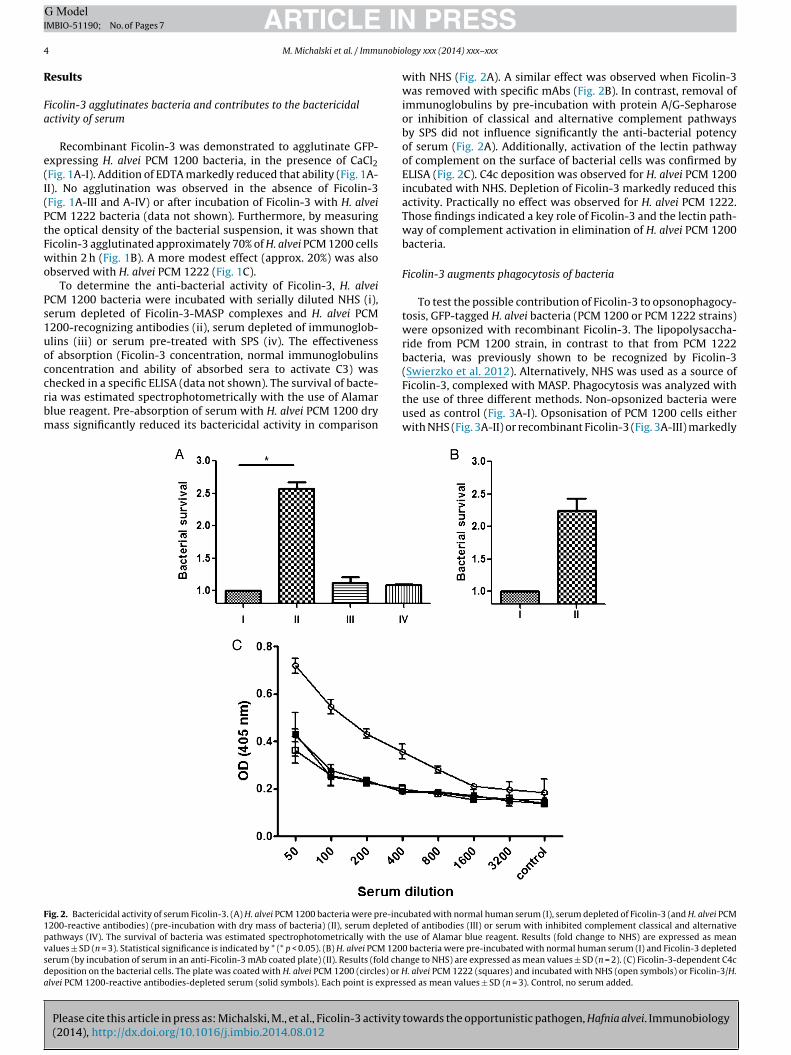
ig. 2. Bactericidal activity of serum Ficolin-3. (A) H. alvei PCM 1200 bacteria were pre-inc200-reactive antibodies) (pre-incubation with dry mass of bacteria) (II), serum depleteathways (IV). The survival of bacteria was estimated spectrophotometrically with the
alues ± SD (n = 3). Statistical significance is indicated by * (* p < 0.05). (B) H. alvei PCM 120erum (by incubation of serum in an anti-Ficolin-3 mAb coated plate) (II). Results (fold chaeposition on the bacterial cells. The plate was coated with H. alvei PCM 1200 (circles) or Hlvei PCM 1200-reactive antibodies-depleted serum (solid symbols). Each point is expres
PRESSlogy xxx (2014) xxx–xxx
with NHS (Fig. 2A). A similar effect was observed when Ficolin-3was removed with specific mAbs (Fig. 2B). In contrast, removal ofimmunoglobulins by pre-incubation with protein A/G-Sepharoseor inhibition of classical and alternative complement pathwaysby SPS did not influence significantly the anti-bacterial potencyof serum (Fig. 2A). Additionally, activation of the lectin pathwayof complement on the surface of bacterial cells was confirmed byELISA (Fig. 2C). C4c deposition was observed for H. alvei PCM 1200incubated with NHS. Depletion of Ficolin-3 markedly reduced thisactivity. Practically no effect was observed for H. alvei PCM 1222.Those findings indicated a key role of Ficolin-3 and the lectin path-way of complement activation in elimination of H. alvei PCM 1200bacteria.
Ficolin-3 augments phagocytosis of bacteria
To test the possible contribution of Ficolin-3 to opsonophagocy-tosis, GFP-tagged H. alvei bacteria (PCM 1200 or PCM 1222 strains)were opsonized with recombinant Ficolin-3. The lipopolysaccha-ride from PCM 1200 strain, in contrast to that from PCM 1222bacteria, was previously shown to be recognized by Ficolin-3(Swierzko et al. 2012). Alternatively, NHS was used as a source of
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
Ficolin-3, complexed with MASP. Phagocytosis was analyzed withthe use of three different methods. Non-opsonized bacteria wereused as control (Fig. 3A-I). Opsonisation of PCM 1200 cells eitherwith NHS (Fig. 3A-II) or recombinant Ficolin-3 (Fig. 3A-III) markedly
ubated with normal human serum (I), serum depleted of Ficolin-3 (and H. alvei PCMd of antibodies (III) or serum with inhibited complement classical and alternativeuse of Alamar blue reagent. Results (fold change to NHS) are expressed as mean0 bacteria were pre-incubated with normal human serum (I) and Ficolin-3 depletednge to NHS) are expressed as mean values ± SD (n = 2). (C) Ficolin-3-dependent C4c. alvei PCM 1222 (squares) and incubated with NHS (open symbols) or Ficolin-3/H.
sed as mean values ± SD (n = 3). Control, no serum added.
ARTICLE IN PRESSG ModelIMBIO-51190; No. of Pages 7
M. Michalski et al. / Immunobiology xxx (2014) xxx–xxx 5
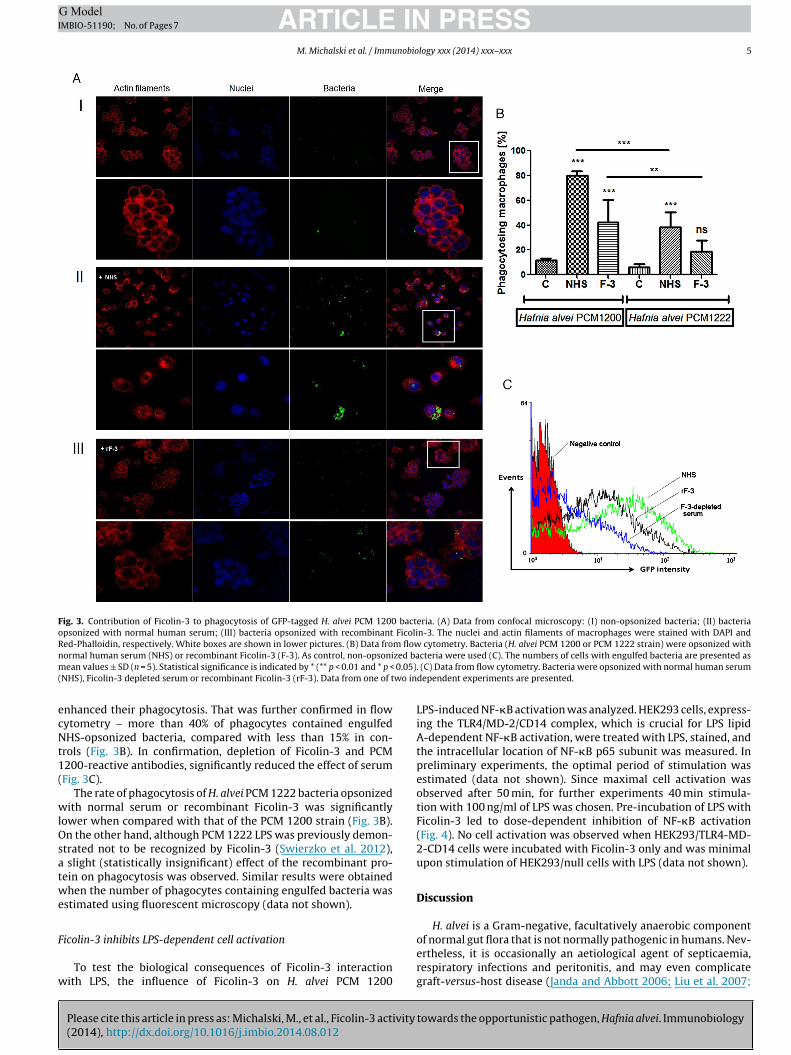
Fig. 3. Contribution of Ficolin-3 to phagocytosis of GFP-tagged H. alvei PCM 1200 bacteria. (A) Data from confocal microscopy: (I) non-opsonized bacteria; (II) bacteriaopsonized with normal human serum; (III) bacteria opsonized with recombinant Ficolin-3. The nuclei and actin filaments of macrophages were stained with DAPI andRed-Phalloidin, respectively. White boxes are shown in lower pictures. (B) Data from flow cytometry. Bacteria (H. alvei PCM 1200 or PCM 1222 strain) were opsonized withn zed bam 0.05)( wo in
ecNt1(
wlOsatwe
F
w
ormal human serum (NHS) or recombinant Ficolin-3 (F-3). As control, non-opsoniean values ± SD (n = 5). Statistical significance is indicated by * (** p < 0.01 and * p <
NHS), Ficolin-3 depleted serum or recombinant Ficolin-3 (rF-3). Data from one of t
nhanced their phagocytosis. That was further confirmed in flowytometry – more than 40% of phagocytes contained engulfedHS-opsonized bacteria, compared with less than 15% in con-
rols (Fig. 3B). In confirmation, depletion of Ficolin-3 and PCM200-reactive antibodies, significantly reduced the effect of serumFig. 3C).
The rate of phagocytosis of H. alvei PCM 1222 bacteria opsonizedith normal serum or recombinant Ficolin-3 was significantly
ower when compared with that of the PCM 1200 strain (Fig. 3B).n the other hand, although PCM 1222 LPS was previously demon-
trated not to be recognized by Ficolin-3 (Swierzko et al. 2012), slight (statistically insignificant) effect of the recombinant pro-ein on phagocytosis was observed. Similar results were obtainedhen the number of phagocytes containing engulfed bacteria was
stimated using fluorescent microscopy (data not shown).
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
icolin-3 inhibits LPS-dependent cell activation
To test the biological consequences of Ficolin-3 interactionith LPS, the influence of Ficolin-3 on H. alvei PCM 1200
cteria were used (C). The numbers of cells with engulfed bacteria are presented as. (C) Data from flow cytometry. Bacteria were opsonized with normal human serumdependent experiments are presented.
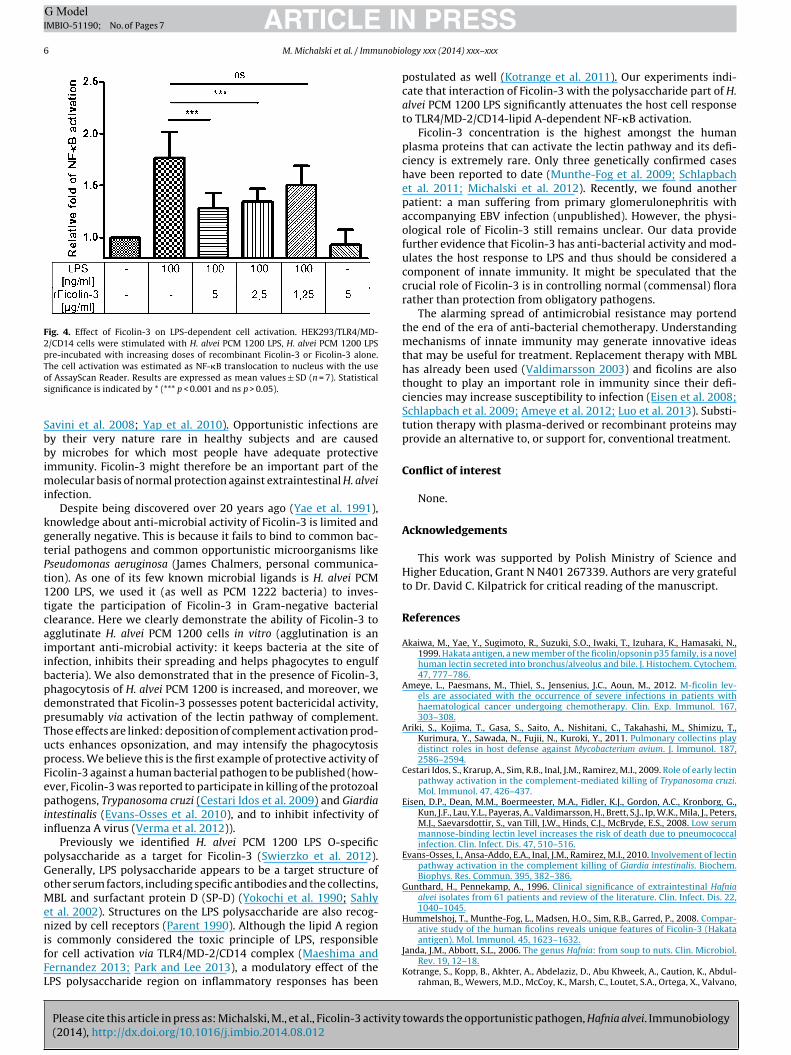
LPS-induced NF-�B activation was analyzed. HEK293 cells, express-ing the TLR4/MD-2/CD14 complex, which is crucial for LPS lipidA-dependent NF-�B activation, were treated with LPS, stained, andthe intracellular location of NF-�B p65 subunit was measured. Inpreliminary experiments, the optimal period of stimulation wasestimated (data not shown). Since maximal cell activation wasobserved after 50 min, for further experiments 40 min stimula-tion with 100 ng/ml of LPS was chosen. Pre-incubation of LPS withFicolin-3 led to dose-dependent inhibition of NF-�B activation(Fig. 4). No cell activation was observed when HEK293/TLR4-MD-2-CD14 cells were incubated with Ficolin-3 only and was minimalupon stimulation of HEK293/null cells with LPS (data not shown).
Discussion
H. alvei is a Gram-negative, facultatively anaerobic component
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
of normal gut flora that is not normally pathogenic in humans. Nev-ertheless, it is occasionally an aetiological agent of septicaemia,respiratory infections and peritonitis, and may even complicategraft-versus-host disease (Janda and Abbott 2006; Liu et al. 2007;
ARTICLE ING ModelIMBIO-51190; No. of Pages 7
6 M. Michalski et al. / Immunobio
Fig. 4. Effect of Ficolin-3 on LPS-dependent cell activation. HEK293/TLR4/MD-2/CD14 cells were stimulated with H. alvei PCM 1200 LPS, H. alvei PCM 1200 LPSpre-incubated with increasing doses of recombinant Ficolin-3 or Ficolin-3 alone.Tos
Sbbimi
kgtPt1tcaiibpdpTupFepii
pGoMenifFL
he cell activation was estimated as NF-�B translocation to nucleus with the usef AssayScan Reader. Results are expressed as mean values ± SD (n = 7). Statisticalignificance is indicated by * (*** p < 0.001 and ns p > 0.05).
avini et al. 2008; Yap et al. 2010). Opportunistic infections arey their very nature rare in healthy subjects and are causedy microbes for which most people have adequate protective
mmunity. Ficolin-3 might therefore be an important part of theolecular basis of normal protection against extraintestinal H. alvei
nfection.Despite being discovered over 20 years ago (Yae et al. 1991),
nowledge about anti-microbial activity of Ficolin-3 is limited andenerally negative. This is because it fails to bind to common bac-erial pathogens and common opportunistic microorganisms likeseudomonas aeruginosa (James Chalmers, personal communica-ion). As one of its few known microbial ligands is H. alvei PCM200 LPS, we used it (as well as PCM 1222 bacteria) to inves-igate the participation of Ficolin-3 in Gram-negative bacteriallearance. Here we clearly demonstrate the ability of Ficolin-3 togglutinate H. alvei PCM 1200 cells in vitro (agglutination is anmportant anti-microbial activity: it keeps bacteria at the site ofnfection, inhibits their spreading and helps phagocytes to engulfacteria). We also demonstrated that in the presence of Ficolin-3,hagocytosis of H. alvei PCM 1200 is increased, and moreover, weemonstrated that Ficolin-3 possesses potent bactericidal activity,resumably via activation of the lectin pathway of complement.hose effects are linked: deposition of complement activation prod-cts enhances opsonization, and may intensify the phagocytosisrocess. We believe this is the first example of protective activity oficolin-3 against a human bacterial pathogen to be published (how-ver, Ficolin-3 was reported to participate in killing of the protozoalathogens, Trypanosoma cruzi (Cestari Idos et al. 2009) and Giardia
ntestinalis (Evans-Osses et al. 2010), and to inhibit infectivity ofnfluenza A virus (Verma et al. 2012)).
Previously we identified H. alvei PCM 1200 LPS O-specificolysaccharide as a target for Ficolin-3 (Swierzko et al. 2012).enerally, LPS polysaccharide appears to be a target structure ofther serum factors, including specific antibodies and the collectins,BL and surfactant protein D (SP-D) (Yokochi et al. 1990; Sahly
t al. 2002). Structures on the LPS polysaccharide are also recog-ized by cell receptors (Parent 1990). Although the lipid A region
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
s commonly considered the toxic principle of LPS, responsibleor cell activation via TLR4/MD-2/CD14 complex (Maeshima andernandez 2013; Park and Lee 2013), a modulatory effect of thePS polysaccharide region on inflammatory responses has been
PRESSlogy xxx (2014) xxx–xxx
postulated as well (Kotrange et al. 2011). Our experiments indi-cate that interaction of Ficolin-3 with the polysaccharide part of H.alvei PCM 1200 LPS significantly attenuates the host cell responseto TLR4/MD-2/CD14-lipid A-dependent NF-�B activation.
Ficolin-3 concentration is the highest amongst the humanplasma proteins that can activate the lectin pathway and its defi-ciency is extremely rare. Only three genetically confirmed caseshave been reported to date (Munthe-Fog et al. 2009; Schlapbachet al. 2011; Michalski et al. 2012). Recently, we found anotherpatient: a man suffering from primary glomerulonephritis withaccompanying EBV infection (unpublished). However, the physi-ological role of Ficolin-3 still remains unclear. Our data providefurther evidence that Ficolin-3 has anti-bacterial activity and mod-ulates the host response to LPS and thus should be considered acomponent of innate immunity. It might be speculated that thecrucial role of Ficolin-3 is in controlling normal (commensal) florarather than protection from obligatory pathogens.
The alarming spread of antimicrobial resistance may portendthe end of the era of anti-bacterial chemotherapy. Understandingmechanisms of innate immunity may generate innovative ideasthat may be useful for treatment. Replacement therapy with MBLhas already been used (Valdimarsson 2003) and ficolins are alsothought to play an important role in immunity since their defi-ciencies may increase susceptibility to infection (Eisen et al. 2008;Schlapbach et al. 2009; Ameye et al. 2012; Luo et al. 2013). Substi-tution therapy with plasma-derived or recombinant proteins mayprovide an alternative to, or support for, conventional treatment.
Conflict of interest
None.
Acknowledgements
This work was supported by Polish Ministry of Science andHigher Education, Grant N N401 267339. Authors are very gratefulto Dr. David C. Kilpatrick for critical reading of the manuscript.
References
Akaiwa, M., Yae, Y., Sugimoto, R., Suzuki, S.O., Iwaki, T., Izuhara, K., Hamasaki, N.,1999. Hakata antigen, a new member of the ficolin/opsonin p35 family, is a novelhuman lectin secreted into bronchus/alveolus and bile. J. Histochem. Cytochem.47, 777–786.
Ameye, L., Paesmans, M., Thiel, S., Jensenius, J.C., Aoun, M., 2012. M-ficolin lev-els are associated with the occurrence of severe infections in patients withhaematological cancer undergoing chemotherapy. Clin. Exp. Immunol. 167,303–308.
Ariki, S., Kojima, T., Gasa, S., Saito, A., Nishitani, C., Takahashi, M., Shimizu, T.,Kurimura, Y., Sawada, N., Fujii, N., Kuroki, Y., 2011. Pulmonary collectins playdistinct roles in host defense against Mycobacterium avium. J. Immunol. 187,2586–2594.
Cestari Idos, S., Krarup, A., Sim, R.B., Inal, J.M., Ramirez, M.I., 2009. Role of early lectinpathway activation in the complement-mediated killing of Trypanosoma cruzi.Mol. Immunol. 47, 426–437.
Eisen, D.P., Dean, M.M., Boermeester, M.A., Fidler, K.J., Gordon, A.C., Kronborg, G.,Kun, J.F., Lau, Y.L., Payeras, A., Valdimarsson, H., Brett, S.J., Ip, W.K., Mila, J., Peters,M.J., Saevarsdottir, S., van Till, J.W., Hinds, C.J., McBryde, E.S., 2008. Low serummannose-binding lectin level increases the risk of death due to pneumococcalinfection. Clin. Infect. Dis. 47, 510–516.
Evans-Osses, I., Ansa-Addo, E.A., Inal, J.M., Ramirez, M.I., 2010. Involvement of lectinpathway activation in the complement killing of Giardia intestinalis. Biochem.Biophys. Res. Commun. 395, 382–386.
Gunthard, H., Pennekamp, A., 1996. Clinical significance of extraintestinal Hafniaalvei isolates from 61 patients and review of the literature. Clin. Infect. Dis. 22,1040–1045.
Hummelshoj, T., Munthe-Fog, L., Madsen, H.O., Sim, R.B., Garred, P., 2008. Compar-ative study of the human ficolins reveals unique features of Ficolin-3 (Hakata
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
antigen). Mol. Immunol. 45, 1623–1632.Janda, J.M., Abbott, S.L., 2006. The genus Hafnia: from soup to nuts. Clin. Microbiol.
Rev. 19, 12–18.Kotrange, S., Kopp, B., Akhter, A., Abdelaziz, D., Abu Khweek, A., Caution, K., Abdul-
rahman, B., Wewers, M.D., McCoy, K., Marsh, C., Loutet, S.A., Ortega, X., Valvano,

ING ModelI
nobio
K
L
L
M
M
M
M
N
O
P
P
P
P
P
ARTICLEMBIO-51190; No. of Pages 7
M. Michalski et al. / Immu
M.A., Amer, A.O., 2011. Burkholderia cenocepacia O polysaccharide chain con-tributes to caspase-1-dependent IL-1beta production in macrophages. J. Leukoc.Biol. 89, 481–488.
rarup, A., Sorensen, U.B., Matsushita, M., Jensenius, J.C., Thiel, S., 2005. Effect ofcapsulation of opportunistic pathogenic bacteria on binding of the pattern recog-nition molecules mannan-binding lectin, L-ficolin, and H-ficolin. Infect. Immun.73, 1052–1060.
iu, C.H., Lin, W.J., Wang, C.C., Lee, K.L., Tsai, M.C., 2007. Young-infant sepsis com-bined with urinary tract infection due to Hafnia alvei. J. Formos. Med. Assoc. 106,S39–S43.
uo, F., Sun, X., Wang, Y., Wang, Q., Wu, Y., Pan, Q., Fang, C., Zhang, X.L., 2013. Ficolin-2 defends against virulent Mycobacteria tuberculosis infection in vivo, and itsinsufficiency is associated with infection in humans. PLOS ONE 8, e73859.
aeshima, N., Fernandez, R.C., 2013. Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front. Cell. Infect. Microbiol. 3, 3.
atsushita, M., Kuraya, M., Hamasaki, N., Tsujimura, M., Shiraki, H., Fujita, T., 2002.Activation of the lectin complement pathway by H-ficolin (Hakata antigen). J.Immunol. 168, 3502–3506.
ichalski, M., Szala, A., Swierzko St, A., Lukasiewicz, J., Maciejewska, A., Kilpatrick,D.C., Matsushita, M., Domzalska-Popadiuk, I., Borkowska-Klos, M., Sokolowska,A., Szczapa, J., Lugowski, C., Cedzynski, M., 2012. H-ficolin (ficolin-3) con-centrations and FCN3 gene polymorphism in neonates. Immunobiology 217,730–737.
unthe-Fog, L., Hummelshoj, T., Honore, C., Madsen, H.O., Permin, H., Garred, P.,2009. Immunodeficiency associated with FCN3 mutation and ficolin-3 defi-ciency. N. Engl. J. Med. 360, 2637–2644.
eth, O., Jack, D.L., Dodds, A.W., Holzel, H., Klein, N.J., Turner, M.W., 2000. Mannose-binding lectin binds to a range of clinically relevant microorganisms andpromotes complement deposition. Infect. Immun. 68, 688–693.
fek, A., Mesika, M., Kalina, Y., Keisari, R., Podschun, H., Sahly, D., Chang, D., McGre-gor, E., Crouch, 2001. Surfactant protein D enhances phagocytosis and killingof unncapsulated phase variants of Klebsiella pneumoniae. Infect. Immun. 69,24–33.
alarasah, Y., Skjoedt, M.O., Vitved, L., Andersen, T.E., Skjoedt, K., Koch, C., 2010.Sodium polyanethole sulfonate as an inhibitor of activation of complementfunction in blood culture systems. J. Clin. Microbiol. 48, 908–914.
arent, J.B., 1990. Membrane receptors on rat hepatocytes for the inner core regionof bacterial lipopolysaccharides. J. Biol. Chem. 265, 3455–3461.
ark, B.S., Lee, J.O., 2013. Recognition of lipopolysaccharide pattern by TLR4 com-plexes. Exp. Mol. Med. 45, e66.
etersen, S.V., Thiel, S., Jensen, L., Steffensen, R., Jensenius, J.C., 2001. An assay for themannan-binding lectin pathway of complement activation. J. Immunol. Methods
Please cite this article in press as: Michalski, M., et al., Ficolin-3 activity
(2014), http://dx.doi.org/10.1016/j.imbio.2014.08.012
257, 107–116.etersson, C., Niedziela, T., Jachymek, W., Kenne, L., Zarzecki, P., Lugowski, C.,
1997. Structural studies of the O-specific polysaccharide of Hafnia alvei strainPCM 1206 lipopolysaccharide containing d-allothreonine. Eur. J. Biochem. 244,580–586.
PRESSlogy xxx (2014) xxx–xxx 7
Sahly, H., Ofek, I., Podschun, R., Brade, H., He, Y., Ullmann, U., Krouch, E., 2002. Sur-factant protein D binds selectively to Klebsiella pneumoniae lipopolysaccharidescontaining mannose-rich O-antigens. J. Immunol. 169, 3267–3274.
Savini, V., Di Bartolomeo, E., Catavitello, C., Talia, M., Manna, A., Febbo, F., Balbinot, A.,Di Bonaventura, G., Di Bartolomeo, P., Piccolomini, R., D’Antonio, D., 2008. Graftversus host disease-related Hafnia alvei colonization and probable infection. J.Med. Microbiol. 57, 1167–1169.
Schlapbach, L.J., Aebi, C., Hansen, A.G., Hirt, A., Jensenius, J.C., Ammann, R.A., 2009.H-ficolin serum concentration and susceptibility to fever and neutropenia inpaediatric cancer patients. Clin. Exp. Immunol. 157, 83–89.
Schlapbach, L.J., Thiel, S., Kessler, U., Ammann, R.A., Aebi, C., Jensenius, J.C., 2011.Congenital H-ficolin deficiency in premature infants with severe necrotisingenterocolitis. Gut 60, 1438–1439.
Sugimoto, R., Yae, Y., Akaiwa, M., Kitajima, S., Shibata, Y., Sato, H., Hirata, J., Okochi,K., Izuhara, K., Hamasaki, N., 1998. Cloning and characterization of the Hakataantigen, a member of the ficolin/opsonin p35 lectin family. J. Biol. Chem. 273,20721–20727.
Swierzko, A., Lukasiewicz, J., Cedzynski, M., Maciejewska, A., Jachymek, W.,Niedziela, T., Matsushita, M., Lugowski, C., 2012. New functional ligands forficolin-3 among lipopolysaccharides of Hafnia alvei. Glycobiology 22, 267–280.
Tsujimura, M., Ishida, C., Sagara, Y., Miyazaki, T., Murakami, K., Shiraki, H., Okochi,K., Maeda, Y., 2001. Detection of serum thermolabile beta-2 macroglycoprotein(Hakata antigen) by enzyme-linked immunosorbent assay using polysaccharideproduced by Aerococcus viridans. Clin. Diagn. Lab. Immunol. 8, 454–459.
Tsujimura, M., Miyazaki, T., Kojima, E., Sagara, Y., Shiraki, H., Okochi, K., Maeda, Y.,2002. Serum concentration of Hakata antigen, a member of the ficolins, is linkedwith inhibition of Aerococcus viridans growth. Clin. Chim. Acta 325, 139–146.
Valdimarsson, H., 2003. Infusion of plasma-derived mannan-binding lectin (MBL)into MBL-deficient humans. Biochem. Soc. Trans. 31, 768–769.
Verma, A., White, M., Vathipadiekal, V., Tripathi, S., Mbianda, J., Ieong, M., Qi, L.,Taubenberger, J.K., Takahashi, K., Jensenius, J.C., Thiel, S., Hartshorn, K.L., 2012.Human H-ficolin inhibits replication of seasonal and pandemic influenza Aviruses. J. Immunol. 189, 2478–2487.
Westphal, O., Jann, K., 1965. Bacterial lipopolysaccharide extraction with phenol-water and further application of the procedure. Methods Carbohydr. Chem. 5,83–89.
Yae, Y., Inaba, S., Sato, H., Okochi, K., Tokunaga, F., Iwanaga, S., 1991. Isolation andcharacterization of a thermolabile beta-2 macroglycoprotein (’thermolabile sub-stance’ or ’Hakata antigen’) detected by precipitating (auto) antibody in seraof patients with systemic lupus erythematosus. Biochim. Biophys. Acta 1078,369–376.
Yap, D.Y., Lau, S.K., Lamb, S., Choy, B.Y., Chan, T.M., Lai, K.N., Tang, S.C., 2010. An
towards the opportunistic pathogen, Hafnia alvei. Immunobiology
unusual organism for PD-related peritonitis: Hafnia alvei. Perit. Dial. Int. 30,254–255.
Yokochi, T., Inoue, Y., Kimura, Y., Kato, N., 1990. Strong interaction of lipopolysac-charides possessing the mannose homopolysaccharides with complement andits relation to adjuvant action. J. Immunol. 144, 3106–3110.
Related Documents