Hydrobiologia 354: 17–40, 1997. 17 c 1997 Kluwer Academic Publishers. Printed in Belgium. Phycoflora of the tropical high-mountain lake El Sol, Central Mexico, and some biogeographical relationships Alfonso G. Banderas-Tarabay Mexican Institute of Water Technology, Hydraulics Coordination, Environmental Hydraulics Area, Paseo Cuauhn´ ahuac 8532, Jiutepec, Morelos 62550, Mexico. FAX 91 73 208725 Received 3 October 1996; in revised form 8 July 1997; accepted 8 July 1997 Key words: phycoflora, freshwater, tropical, high-mountain lake, biogeography Abstract Algae collected from an oligotrophic crater lake were identified and compared with similar regional information. A total of 50 genera and 105 infrageneric taxa were identified, mostly Chlorophyta (66%), Cyanophyta (20%) and Bacillariophyta (7 genera: one central and six pennates). Relatively few Chlorococcales were found in this oligotrophic lake. The families most frequently found were Oedogoniaceae (18 species), Desmidiaceae (17), Zygne- mataceae (13), Oscillatoriaceae (11), and Chaetophoraceae (9). Two genera of Cyanophyta, seven of Chlorophyta and 59 species are first records for Mexico. Introduction The appearance of new species during the exploration of Mexico and tropical areas of the world is still common because of a scarcity of biodiversity studies (Ramamoorthy et al., 1993). Ortega (1985) stated that the continential phycoflora in Mexico has been large- ly ignored due to shortage of phycologists, few large water bodies, deficiencies in the selection of study sites and sampling methods, and the limited availability of specialized literature. She lists 837 infrageneric taxa prior to 1974, but more phycological studies will raise this number, as is shown by this paper on the phyto- benthos of an oligotrophic high-mountain crater lake. Description of site studied El Sol lake is located at 19 08 N and 99 45 W at an altitude of 4,170 m in the so called alpine, cold high-mountain region (Banderas et al., 1991) of the Nevado de Toluca Volcano belonging to the Mexican Neovolcanic Belt. It is a small oligotrophic crater lake with a maximum depth of 15 m and surface area of 0.23 km 2 . Its water is clear and continuously circu- lates. It was formed at the end of the last glacier age (Pleistocene), when the high, broken terrain where the volcano lies was formed by intense volcanic activi- ty mainly during the Miocene and Pliocene (Waitz, in Deevey, 1957). The basin is small and closed. The crater has a large central dome (El Ombligo) that covers the volcano’s main chimney and forms the eastern edge of the lake. Given the height of the dome (about 4320 masl), it seems that the floor of the crater descend- ed with the sinking of the underlying structures into the empty chamber of the volcano. This process could have occurred as described by Hutchinson (1975) for the origin of Lizard Island in Crater Lake, Oregon. The limnology of El Sol has been described by Banderas et al. (1991). This type of lakes is unique in Mexico and rare in the tropics (cf. Larson, 1989). The only aquatic vertebrate present is the rainbow trout (Salmo gairdneri), which was introduced at the middle of this century. Materials and methods Twenty-three sampling sites with different depths, sub- strates, slopes and orientations in the lake, were select- ed. For each depth (stratum), a number of sites were selected (0.1 m, 5 sites; 1 m, 5 sites; 2 m, 5 sites;

Ficoflora Tarabay
Nov 08, 2014
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hydrobiologia 354: 17–40, 1997. 17c 1997 Kluwer Academic Publishers. Printed in Belgium.
Phycoflora of the tropical high-mountain lake El Sol, Central Mexico,and some biogeographical relationships
Alfonso G. Banderas-TarabayMexican Institute of Water Technology, Hydraulics Coordination, Environmental Hydraulics Area,Paseo Cuauhnahuac 8532, Jiutepec, Morelos 62550, Mexico. FAX 91 73 208725
Received 3 October 1996; in revised form 8 July 1997; accepted 8 July 1997
Key words: phycoflora, freshwater, tropical, high-mountain lake, biogeography
Abstract
Algae collected from an oligotrophic crater lake were identified and compared with similar regional information.A total of 50 genera and 105 infrageneric taxa were identified, mostly Chlorophyta (66%), Cyanophyta (20%)and Bacillariophyta (7 genera: one central and six pennates). Relatively few Chlorococcales were found in thisoligotrophic lake. The families most frequently found were Oedogoniaceae (18 species), Desmidiaceae (17), Zygne-mataceae (13), Oscillatoriaceae (11), and Chaetophoraceae (9). Two genera of Cyanophyta, seven of Chlorophytaand 59 species are first records for Mexico.
Introduction
The appearance of new species during the explorationof Mexico and tropical areas of the world is stillcommon because of a scarcity of biodiversity studies(Ramamoorthy et al., 1993). Ortega (1985) stated thatthe continential phycoflora in Mexico has been large-ly ignored due to shortage of phycologists, few largewater bodies, deficiencies in the selection of study sitesand sampling methods, and the limited availability ofspecialized literature. She lists 837 infrageneric taxaprior to 1974, but more phycological studies will raisethis number, as is shown by this paper on the phyto-benthos of an oligotrophic high-mountain crater lake.
Description of site studied
El Sol lake is located at 19�080N and 99�450W atan altitude of 4,170 m in the so called alpine, coldhigh-mountain region (Banderas et al., 1991) of theNevado de Toluca Volcano belonging to the MexicanNeovolcanic Belt. It is a small oligotrophic crater lakewith a maximum depth of 15 m and surface area of0.23 km2. Its water is clear and continuously circu-lates. It was formed at the end of the last glacier age
(Pleistocene), when the high, broken terrain where thevolcano lies was formed by intense volcanic activi-ty mainly during the Miocene and Pliocene (Waitz,in Deevey, 1957). The basin is small and closed. Thecrater has a large central dome (El Ombligo) that coversthe volcano’s main chimney and forms the eastern edgeof the lake. Given the height of the dome (about 4320masl), it seems that the floor of the crater descend-ed with the sinking of the underlying structures intothe empty chamber of the volcano. This process couldhave occurred as described by Hutchinson (1975) forthe origin of Lizard Island in Crater Lake, Oregon.
The limnology of El Sol has been described byBanderas et al. (1991). This type of lakes is uniquein Mexico and rare in the tropics (cf. Larson, 1989).The only aquatic vertebrate present is the rainbow trout(Salmo gairdneri), which was introduced at the middleof this century.
Materials and methods
Twenty-three sampling sites with different depths, sub-strates, slopes and orientations in the lake, were select-ed. For each depth (stratum), a number of sites wereselected (0.1 m, 5 sites; 1 m, 5 sites; 2 m, 5 sites;
GSB: PIPS Nr.: 147214 BIO2KAPhy3739.tex; 11/11/1997; 13:42; v.5; p.1

18
3 m, 3 sites, 4 m, 2 sites; and 5, 10 and 15 m, 1site each one), in proportion to the lake’s area, whichwas considered to have a bucket shape with the areadecreasing with depth. This design reduces the num-ber of samples needed without sacrificing the repre-sentation of the algal community. Monthly visits weremade from April 1982 to April 1983, except in Januaryand February due to bad weather. The sampling siteswere reached from a boat using a lead weight (isobath-ic region) thus it is impossible that two samples mayhave been taken from the same site.
A weighed multihook anchor was dragged for twoor three meters and the entangled vegetation collect-ed. Samples were fixed using 4% formaline. Algaewere analyzed using an optical microscope with a clearchamber. Species were identified following publisheddescriptions and drawings were made and measure-ments taken of each species. Cellular diameter dataof filamentous species was presented in stem-and-leafdiagrams (Tukey, 1977). Material collected for thisstudy was placed in the MEXU Herbarium (Banderas,1986).
Species were classified (Appendix 1) according toSilva (1962). The classification of the angiospermswas based on Cronquist (1968). Following Thomasson(1956), the phycoflora of El Sol was compared withthat described by Ortega (1985), Mendoza-Gonzalez(1985) and Iltis (1991) to examine its biogeographicalrelationships.
Results
Flora list
Species found in this study and by Kusel-Fetzmann(1973) and Villalpando (1968) are listed in Appendix1. The Ceratium sp. and Triploceras sp. genera wereidentified from the remains of two epithecas and a cellwall, respectively, that appeared in the samples. Themorphometric separation of the Zygnema and Spirogy-ra (Chlorophyta, Zygnemataceae) was accomplishedconsidering the abundance and variety of both forms(Figures 1 and 2). These were dominant in samplesfrom sandy, shallow areas and in those without rhi-zobenthos.
(i) Zygnema and/or Zygnemopsis. This genus wascommon and abundant. The individual filamentswere found closely grouped around diameters of10, 20 and 30 �. These units are morphometrically
similar to those described by Iltis (1984) for theAndean lakes at altitudes similar to that of El Sol.
(ii) Spirogyra. This genus was scarce, but widely dis-tributed throughout the lake. Diameter and shapeof the cross wall indicates the presence of four sim-ilar groups, plus two additional groups of similarsize but with a different number of chloroplasts(Figures 79 y 80 in Appendix 1). Modified mirror-image stem-and-leaf diagram further differentiatessuch groups.
Floristics
A total of 105 infrageneric taxa were identified,two of which were angiosperms, with filamentousspecies being dominant. Chlorophyta contributed 69species (66%), with 3 species of Zygnema (Figure 1),and 2 species and 6 forms of Spirogyra (Figure 2).Cyanophyta were represented by 20 species (19%)two of which had heterocysts. Pyrrophyta and Chryso-phyta were represented by two species each and Xan-thophyta, Euglenophyta and Charophyta by one each.Bacillariophyta was represented by 7 genera (14% ofgenera), one centric and six pennates, that were notidentified to the species level.
The best represented families were Oedogoniaceae(18 species), Desmidiaceae (17), Zygnemataceae (13),Oscillatoriaceae (11), and Chaetophoraceae (9). Thisresult underscores the diversity of mostly seconddegree epiphytic Chaetophoraceae, growing on Oedo-gonium spp., which was mostly first degree epiphyticon rhizobenthic macrophytes (Ellatine brachysperma,Elocharis sp. and Nitella gracilis v. intermedia).
Subcommunities
Phytoplankton was dominated by Peridinium lom-nickii, P. willei, Dinobryon cilindricum v. alpinum,small diatoms and smaller species of Closteriumand Oscillatoria. Usually benthic species of Zygne-ma, Spirogyra, Desmidium swartzii and Gonatozy-gon aculeatum, were also found in the plankton(tychopelagic).
All the phytobenthic subcommunities described byRound (1981) were found. Sandy shallow areas with aslight slope housed a smaller diversity of filamentousspecies dominated by epipelic species. This was nottrue for a small bay located on the southwestern shoreof the lake, nor for the rocky areas with interstitial sandand mud where the winding course reduced the flowof water and favoured the deposition of the diaspores.
hy3739.tex; 11/11/1997; 13:42; v.5; p.2

19
Figure 1. Stem-and-leaf diagram of the morphometric separation of genus Zygnema. Numbers represent: in the column, tenths; beside thecolumn, cumulative frequencies; in parenthesis, the mode ubication; right hand, units in micrometers (�).
In these areas, colonies of Ellatine brachysperma wereabundant, together with Eleocharis sp.; Nitella gra-cilis v. intermedia dominated from a depth of 5 metersto the deep muddy regions. These three macrophytesbore an abundant subcommunity of epiphytes, main-ly Oedogoniales, and Gomphonema sp. and Lygnbyaperelegans. A second degree subcommunity of epi-phytes represented by Chaetophorales, and Tolypothrixnodosa, Nostoc paludosum and Chaetosphaeridiumpringsheimii, was found attached to oedogoniales.
In the epilythic subcommunity, Bulbochaete repan-da was observed in the upper, and Oedogonium grandev. majus in the lower strata. This subcommunity alsoincluded Stigeoclonium aestivalis, S. pusillum andDraparnaldia glomerata. The epipelic subcommuni-ty included the Zygnematales at mid-depth areas witha sandy bed. Species of Spirogyra were often foundat a low abundance, while Zygnema spp., Desmidi-um swartzii, Gonatozygon aculeatum, G. monotaeni-um and Pleurotaenium ehrenbergii were often found inabundance. Both filamentous and unicellular epipelicspecies formed part of the metaphytic subcommuni-ty in the shallow sandy areas and in the deep muddy
regions, accompanied by rhizobenthos and their epi-phytes.
Closterium lunula and Euglena acus v. acus werefound most often where the bed was muddy, the formerthroughout the depth profile, and the latter at depthsgreater than 10 m. Populations of hydra (Pelmato-hydra) and oligochaete annelids (Lumbricidae) werefound in samples from the deepest strata, where thelatter were specially abundant.
Biogeography
Of the 50 (100%) genera found in El Sol, 9 (18%)are first records for Mexico. These genera includ-ed 10 (9.9%) species: Aulosira sp., Pseudoanabaenaconstricta, Aphanochaete repens, Chaetonema orna-tum, Thammiochaete huberi, Pseudoendoclonium sp.,Gonatozygon aculeatum, G. monotaenium, Tetmemo-rus granulatus and Triploceras sp. At the species level,of the 103 algal taxa found in the lake, 59 (includingone angiosperm) were registered for the first time inMexico. The similarity of the phycoflora from El Soland that for the whole of Mexico (Ortega, 1984), is82% (genera) and 40.4% (species).
hy3739.tex; 11/11/1997; 13:42; v.5; p.3

20
Figure 2. Mirror-image stem-and-leaf diagram of the morphometric separation of the genus Spirogyra. On the left are the forms with replicatedcross wall; on the right are those with plane cross wall. Roman numbers indicate the distinct morphospecies. Bars represent the limits of thefilament diameter of Spirogyra jugalis (Sj) and S. setiformis (Ss). Units as in Figure 1.
Santiago Tilapa Lake, located at 2800 masl on theSW slope of the Ajusco mountain range (Mendoza-Gonzalez, 1985) lies also within the Neovolcanic Axis.It is found 35 km east-northeast of El Sol lake. Thesimilarity of this community, with 76 genera and 156species, with that of El Sol, was 64% for genera and12% for species. Santiago Tilapa Lake has a greatervariety of cyanophytes.
The algal community of the puna-type TiticacaLake (3810 masl) in the Bolivian Andes, includes 158species (Iltis, 1984 and 1991), 27 (63%) of the 43 gen-era (excluding the diatoms and angiosperms) in El Solare also found in the Titicaca. Of the 14 remaining,
Pseudoanabaena, Aphanochaete and Thamnniochaetehad not been found either in the Andes or in Mexico.
The Cyanophyta and Chlorococcales showed agreater diversity of genera and species in Santiago Tila-pa and Titicaca lakes than in El Sol, where the formswith heterocysts seems to be absent. At the species lev-el, 13 (14% of the flora in El Sol, excluding diatoms)are also present in the Titicaca but with a smaller vari-ety in desmids. The similarity between the floras inSantiago Tilapa and in Titicaca is greater than theirsimilarity with El Sol. The two former share 38 (50%)genera and 26 (17%) species, particularly the desmidsand chlorococcales.
hy3739.tex; 11/11/1997; 13:42; v.5; p.4

21
Discussion
The presence of several planktonic species identifiedin previous studies was confirmed here. Individualssmaller than the distance between the hooks of theanchor used, and those with rigid parts firmly adheredto the substrate, were probably under-represented inour samples. This may partially explain the low diver-sity of chlorococcales in El Sol. However, becauseorganisms from this group were not found in samplestaken using a 20� planckton net, it is likely that they areabsent from El Sol, perhaps because it is oligotrophic.It is precisely the presence of desmids and chlorococ-cales which accounts for the similarity between San-tiago Tilapa (meso-eutrophic) and Titicaca lakes. Thefrequent presence of unicellular species in the samplesmay indicate that they are abundant in the benthos andnot in the plankton. This is true for most of the diatomsand desmids that together constitute 26% of all speciesat El Sol.
The qualitative differences in vegetation betweensites were associated to differences in substrate. Asidefrom the obvious species-substrate relationships, quan-titative methods are required to detect less clearlydefined associations.
The fact that more than half of the species found inEl Sol had not been registered in Mexico may be dueto two factors:(1) The physiographic features of the basin may deter-
mine an unusual climate in a region otherwise trop-ical and semiarid, or
(2) The abundance of epiphytic species of the Oedogo-niaceae and Chaetophoracea families in the com-munity may just reflect that few studies on benthicalgae have been carried out or published in Mexico.These two factors suggest that, by increasing the
number of phycofloristic studies in the northern andhigher regions of Mexico, the number of recordedfreshwater algae species would increase. Most of theOscillatoriaceae were also newly reported, but thismay be due El Sol being oligotrophic. Additionalstudies may confirm that Aphanochaete, Tamniochaeteand Ellatine brachysperma can be considered atypicalin tropical regions and better adapted to oligotrophicwaters in temperate regions.
The relative isolation of El Sol with respect to otherwater bodies can be demonstrated by the presence ofNitella gracilis that seems to be widespread in Mexi-co (Ortega, 1985). According to Proctor (1980), thisspecies has a pantropical distribution and in isolatedregions may show an unusual combination of charac-
teristics. This observation was confirmed in the sam-ples from El Sol, where submonoic individuals with avariable number of dactilar cells and peculiar ramifi-cations were found. In spite of the isolation, the florain El Sol was similar at the generic (82%) and specific(40.4%) levels, to that for the whole of Mexico, to thatof the neighbouring Santiago Tilapa Lake (64% and12%), a representative of the lower regions, and to thatof the Andes (63% and 14%), a representative of theneotropical high-altitude region. This similarity wasclose in spite of distance. No regionalization is possi-ble based on this comparison. Otherwise, it reflects atrivial algal flora.
A comparison of El Sol with Santiago Tilapa andTiticaca lakes emphasized two features. The algaediversity (excluding diatoms) was greater in the Mexi-can lakes than in Titicaca, which is bigger. This pover-ty of flora in Titicaca has been pointed out by Iltis(1991). El Sol shares the same fraction of genera andspecies with the other two. However, the absence in theTiticaca of Aphanochaete, Tamniochaete and Elatinebrachysperma may only reflect a lack of systematicstudies of the periphyton in the Andean lakes (Iltis,1984). This is suggested by the occurrence of the firsttwo organisms as second degree epiphytes in El Sol.
The greater similarity (genera and species) of thephycoflora in El Sol with the known Mexican tropi-cal flora implies a common origin for Northamericanalgal floras. Loffler (1972), based on a study by Kusel-Fetzmann (1973), stated that there is a great similaritybetween the phytoplankton in the high lakes of CentralAmerica and the northern portion of the Andes dueto the presence of Peridinium (P. willei and P. volzii),Botryoccus braunii and Dinobryon spp., implying anample dispersion of species. The dominance of flagel-late species in the phytoplankton is characteristic ofAlpine lakes (Tilzer, 1973). However, Loffler (1972)also studied the zooplankton and found a clear region-alization in the harpacticoid fauna between the highaltitude regions of North, Central and South Amer-ica, attributable to the geographic isolation resultingfrom the submersion of a portion of Central Amer-ica during the Eocene and Miocene. He concludedthat passive dispersion processes may be less effectivefor zooplankton than previously considered. Recent-ly, Sarma et al. (1996) found 31 species of rotifers(Rotifera, Monogononta) of which 10 were registeredfor the first time in Mexico (Lecane inopinata, L. fur-cata, Lepadella rhomboides, Cephalodella tenuiseta,Trichocerca bidens, T. collaris, Testudinella emargin-ula, Taphrocampa annulosa, Aspelta lestes and Dicra-
hy3739.tex; 11/11/1997; 13:42; v.5; p.5

22
nophorus forcipatus). The authors defined this set ofspecies as transition fauna between the Neartic andNeotropical biota. Their physical-chemical data coin-cided with those of Banderas et al. (1991) and Loffler(1972), with a difference of 10 and 20 years each, indemonstrating that the limnological conditions werestable over this period.
This study was carried out with data from onlyone year (both physical-chemical and biological dates).The question may be posed as to whether this ecosys-tem is stable enough to use data from one year to char-acterize the dynamics over a longer period? It seemssafe to say that at least one more year of sampling isrequired to update information and to provide infor-mation on the lake community stability. However, thisstudy may be representative of middle term communityvariability because:(1) Except for the annual variations observed in this
study, the lake did not show changes dramaticenough to produce a substantial change in its olig-otrophic condition or in the quality of the oth-er components of the aquatic environment. Thus,the environment is quite predictable. According toSnoeijs and Prentice (1989), the composition ofspecies in the phycobenthic community is a con-servative property unless there is a catastrophicevent that radically changes the characteristics ofthe water or of the surrounding land that may, inturn, result in pollution of the water or induce achange in the trophic conditions.
(2) The aquatic environment within a volcano suchas El Sol, restricts the successful colonization bytropical species from lower regions.
Conclusions
The information concerning biodiversity in Mexico isinsufficient. Of the algal flora, only 837 infragener-ic algal taxa have been described, whereas in thisstudy 104 were identified, of which 59 were regis-tered for the first time in Mexico. This would indicatethat by increasing the number of studies, the measureddiversity would rise. The algal flora found in El Solwas similar to both the neartic and the Andean algalfloras, except for a lower diversity of chlorococcalesand desmids, which may be related to its oligotrophiccondition. Aphanochaete, Tamniochaete and Elatinebrachysperma can be regarded atypical for tropicalregions and more typical of oligotrophic waters fromtemperate climates. However, a study of the Andean
periphyton is required to better define the biogeograph-ic relationships between these regions. This study isrepresentative in the middle term because the systemexperienced no dramatic limnological changes duringa twenty-year period and the high altitude restricts thesuccessful colonization of species from lower regions.
Acknowledgements
This project was begun at the Phycology Laboratoryat the Biology Institute of the U.N.A.M., under DrMartha M. Ortega, and concluded at the PhycologyLaboratory of the Faculty of Sciences, of the same Uni-versity, under Dr Jorge Gonzalez-Gonzalez, to whomI express my gratitude. I also thank Dianne Haywardfor the English translation of the manuscript, and DrAndrea Raz-Guzman Macbeth,Dr Constantino Macıasand Alejandro Ruiz-Lopez for their review and helpfulcomments.
References
Banderas, A., 1986 (Ined). Coleccion de algas bentonicas del LagoEl Sol, en el crater del Volcan Nevado de Toluca, Mexico: Guıade Consulta. Instituto de Biologıa, Herbario MEXU. U.N.A.M.11 p.
Banderas, A., R. Gonzalez-Villela & G. de la Lanza, 1991. Limno-logical aspects of a high mountain lake in Mexico. Hydrobiologia224: 1–10.
Bourrelly, P., 1970. Les Algues d’Eau Douce. Initiation a laSystematique. Tome III: Les algues bleues et rouges. EditionsN. Boubee et Cie. Paris.
Bourrelly, P., 1972. Les Algues d’Eau Douce. Initiation a laSystematique. Tome I: Les algues vertes. Editions N. Boubeeet Cie. Paris.
Correll, D.S. & H.B. Correll, 1975. Aquatic and Wetland plants ofSouthwestern United States. Stanford Univ. Press. V. II. 1145 pp.
Cronquist, A., 1968. The Evolution and Classification of FloweringPlants. William C. Steere (Ed.), Thomas Nelson (Printers) Ltd.Great Britain.
Deevey, E.S., 1957. Limnologic studies in Middle America. Trans.Conn. Acad. Art and Sciences 39: 213–328.
Desikachary, T.V., 1959. Cyanophyta. Indian Council of AgriculturalResearch, New Delhi. 686 pp.
Hutchinson, G.E., 1975. A Treatise on Limnology. V. 1, P. 1. J. Wiley& Sons, Inc. 540 pp.
Iltis, A., 1984. Algues du Lac Titicaca et des lacs de la valle d’IchuKhota (Bolivia). Criptogamie Algol. 5: 85–108.
Iltis, A., 1991. Algues du Lac Titicaca Bolivien. Criptogamie, Algol.12: 213–230.
Islam, A.K.M.M., 1963. A Revision of the Genus Stigeoclonium.Beihefte Zur Nova Hedwigia. Heft 10. Verlag Von. J. Cramer(Germany) Weinheim.
Kusel-Fetzmann, E., 1973. Beitrage zur Kenntnis der Algenflorader Hochgebirge Zentralamerikas und der Nordlichen Anden.Hochgebirgsforschung, Heft 3, 5: 29–68.
hy3739.tex; 11/11/1997; 13:42; v.5; p.6

23
Loffler, H., 1972. Contribution to the limnology of high-mountainlakes in Central America. Int. Revue. ges. Hydrobiol. 57: 397–408.
Larson, D.W., 1989. Caldera lakes. Hydrobiologia, 171: 23–32.Mendoza-Gonzalez, A.C., 1985. Estudio florıstico ficologico esta-
cional de la Laguna de Victoria o de Santiago Tilapa, Mexico.Phytologia 58: 479–487.
Ortega, M.M., 1985. Algas Continentales Recientes de Mexico.U.N.A.M. 559 p.
Palmer, C.M., 1962. Algas en Abastecimientos de Agua. Ed. Inter-americana, S.A. Mexico. 91 p., 6 Lam.
Printz, H., 1964. Die Chaetophoralen der Binnengewasser, 367.Verlag Dr. W. Junk-Dan Haag. 376 pp.
Proctor, V.W., 1980. Historical biogeography of Chara (Charophy-ta): an appraisal of the Braun-Wood classification plus a falsiliablealternative for future consideration. J. Phycol. 16: 218–233.
Ralfs, J., 1848. The British Desmidieae. Reprint 1972. Verlag Von J.Cramer. Wheldom and Wesley Ltd. New York, N.Y. 226 pp. 35lam.
Rammamorthy, T.P., R. Bye, H.A. Lot & J. Fa (eds): BiologicalDiversity of Mexico: Origins and Distribution. Oxford UniversityPress, New York, 579 pp.
Randawa, M.S., 1959. Zygnemaceae. Indian Council of AgriculturalResearch, New Delhi, 478 pp.
Round, E., 1981. The Ecology of Algae. Cambridge, Univ. press.Great Britain: 653 pp.
Sarma, S.S., Elıas-Gutierrez & C. Serranıa-Soto, 1996. Rotifersfrom the high altitude crater lakes located at volcano Nevado deToluca, Mexico. Hydrobiologia 6: 33–38.
Silva, P.C., 1962. Classification of Algae. In Lewin (ed.), Physiologyand Biochemistry of Algae. Academic Press Inc., N. York: 827–837.
Snoeijs, P.J.M. & I.C. Prentice, 1989. Effects of cooling water dis-charge on the structure and dynamics of epilithic algal commu-nities in the northern Baltic. Hydriobiologia 184: 99–123.
Starmach, K., 1966. Flora Slodkowodna Polski. V. II: Cyanophyta-Sinice; Glaucophyta-Glaukofity. Polska akademia Nank, InstytutBotaniki. Warszawa: 807 pp.
Taylor, W., 1928. The alpine algal vegetation of mountains of BritishColumbia. Proc. Acad. Nat. Sci. Philad. 80: 45–114.
Thomasson, K., 1956. Reflections on artic and alpine lakes. Oikos7: 119–143.
Tiffany, L.H., 1930. The Oedogoniaceae. A Monograph. Publ. bythe Autor. Columbus, Ohio, 253 pp.
Tilzer, M.M., 1973. Diurnal periodicity in the phytoplankton assem-blage of a high-mountain lake. Limnol. Oceanogr. 18: 15–30.
Tukey, J.W., 1977. Exploratory Data Analysis. Addison-WesleyPubl. Co. Reding, Massachussets, 688 pp.
Villalpando, O., 1968. Algunos Aspectos Ecologicos del VolcanNevado de Toluca. Tesis Licenciatura. Fac. de Ciencias U.N.A.M.
West, W. & G.S. West, 1904. British Desmidiaceae. A. Monograph.V. 1. The Ray Society. Reprint for Johnson Reprint Corp, 1971.
Wetzel, R.G., 1975. Limnology. 2nd edn. 1983. Saunders CollegePubl., Mexico City, 767 pp.
Whitford, L.A. & G.J. Schumacher, 1969. A Manual of the Fresh-water Algae in North Carolina. Tech. Bul. 188. Published by TheNorth Carolina Agricultural Experiment Station.
Wood, R.D. & K. Imahori, 1964. A Revision of the Characeae.Weinghaim. Verlag Von J. Cramer. With 394 icones.
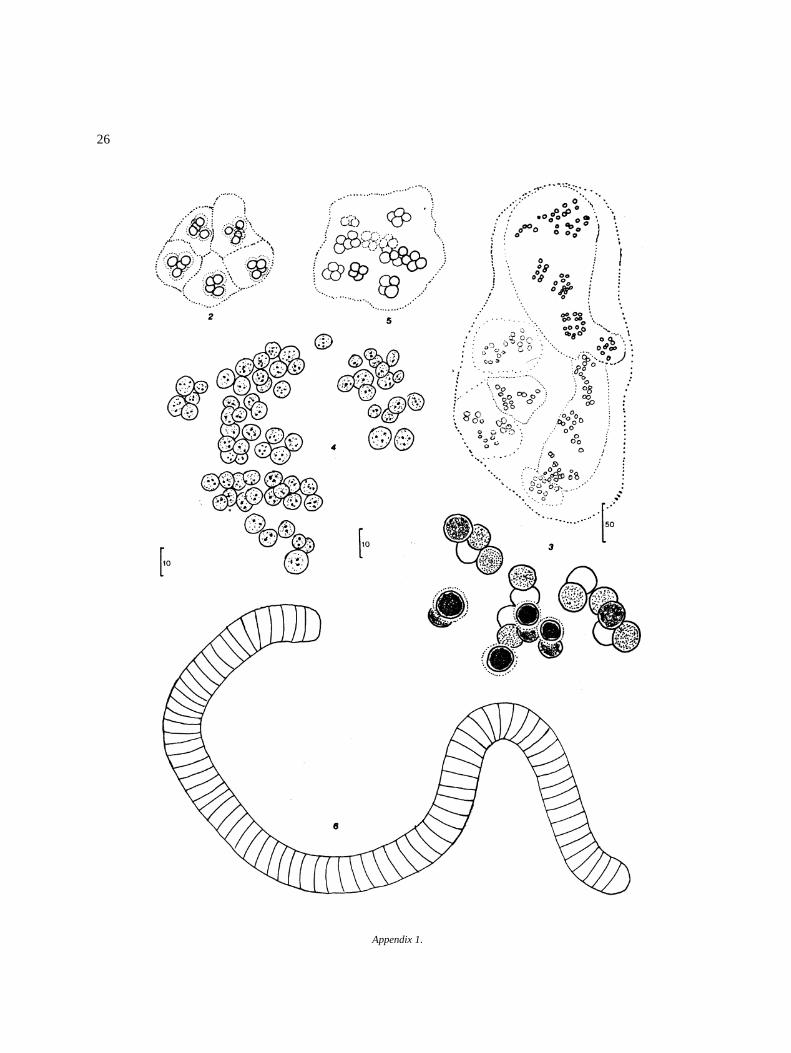
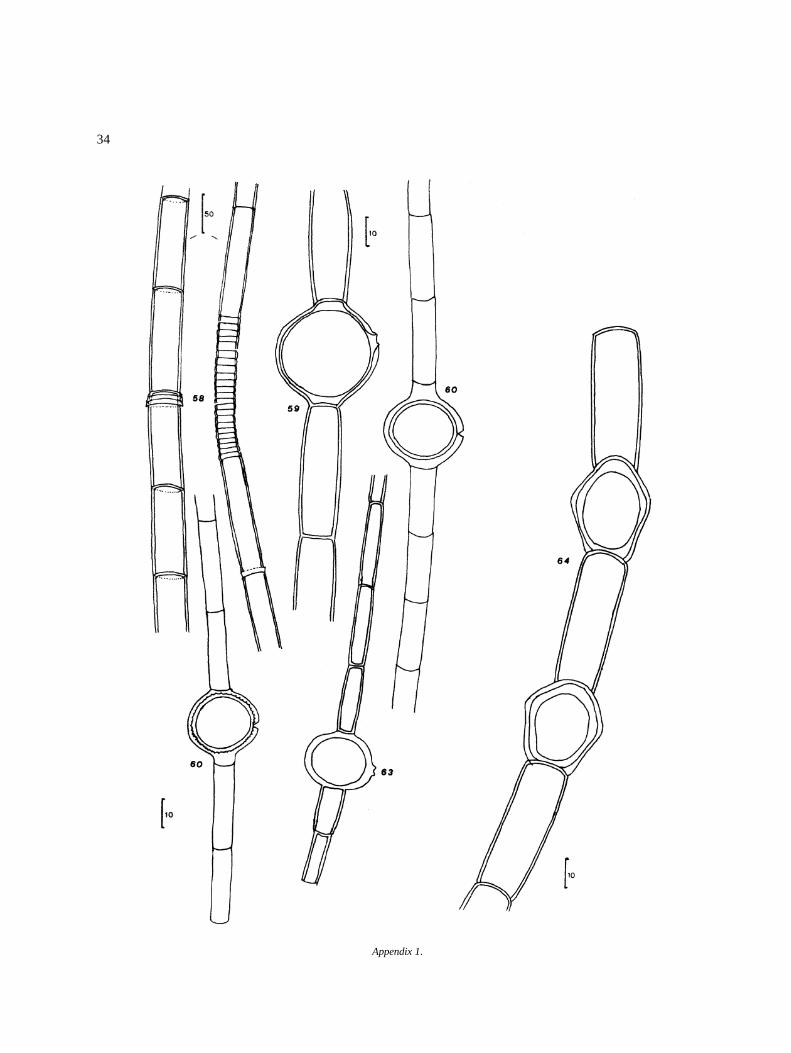
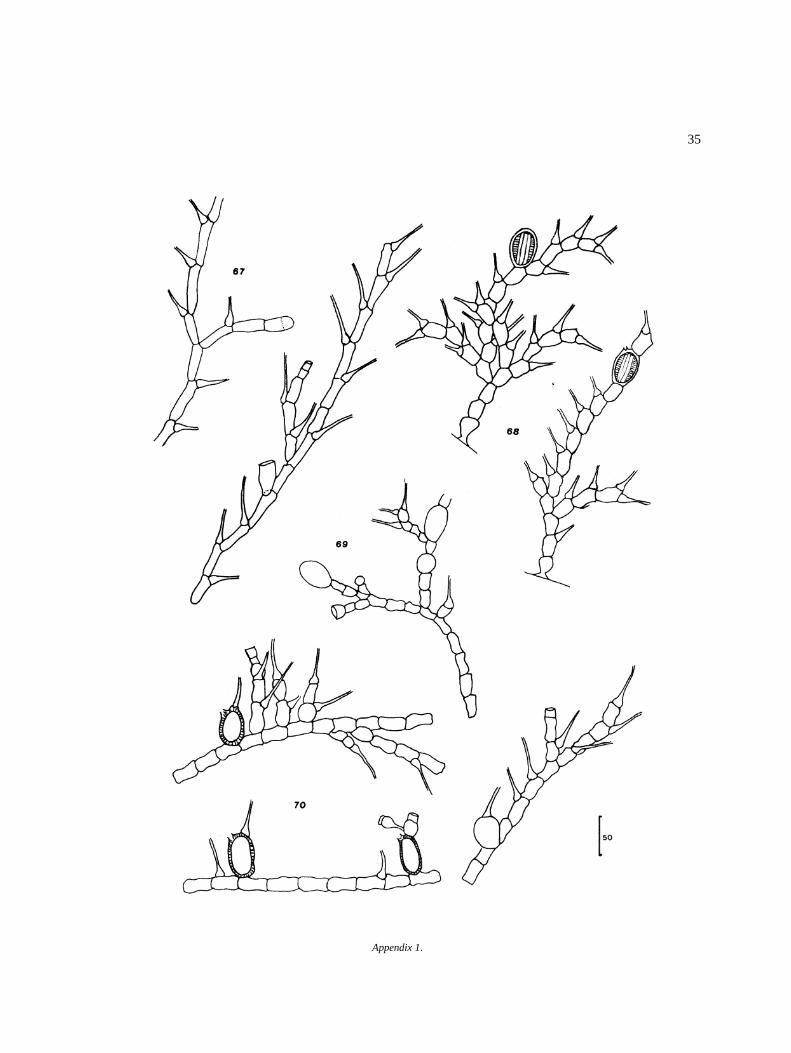
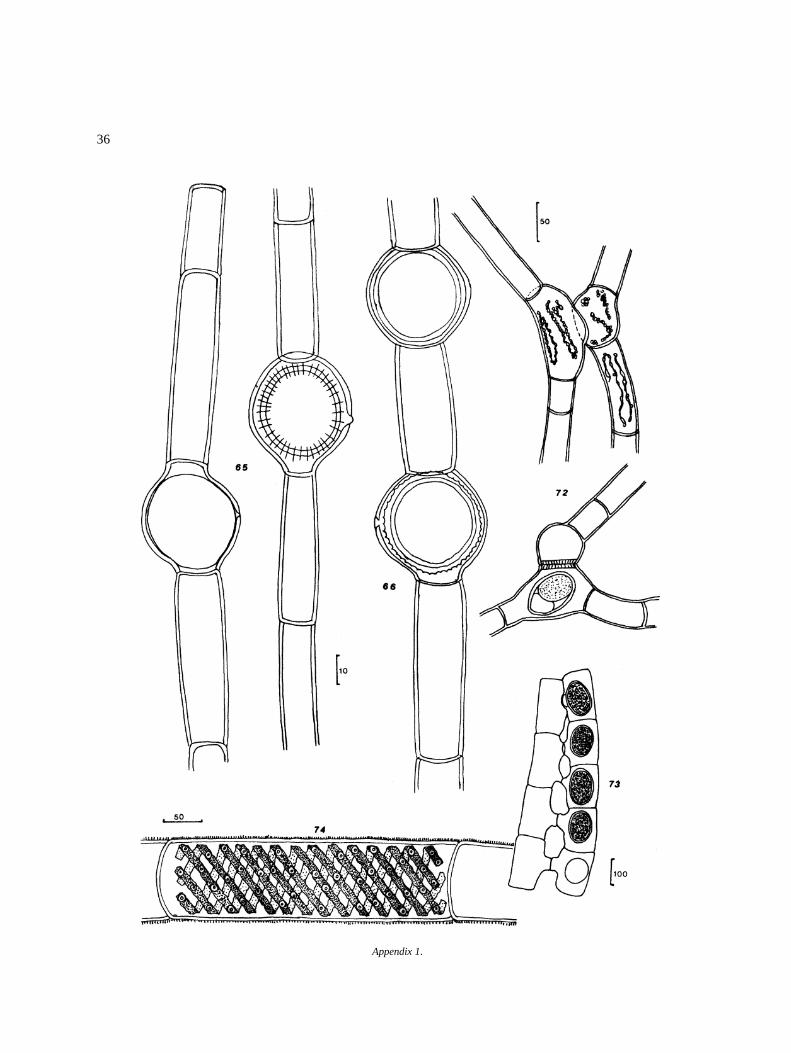
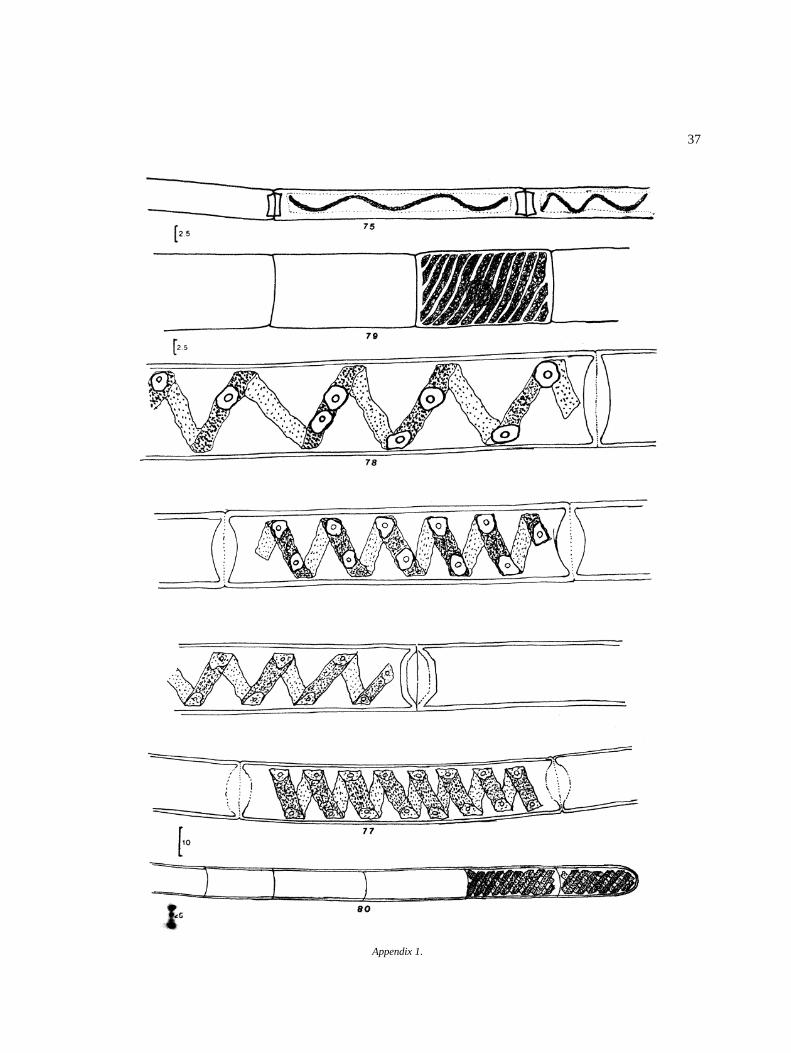
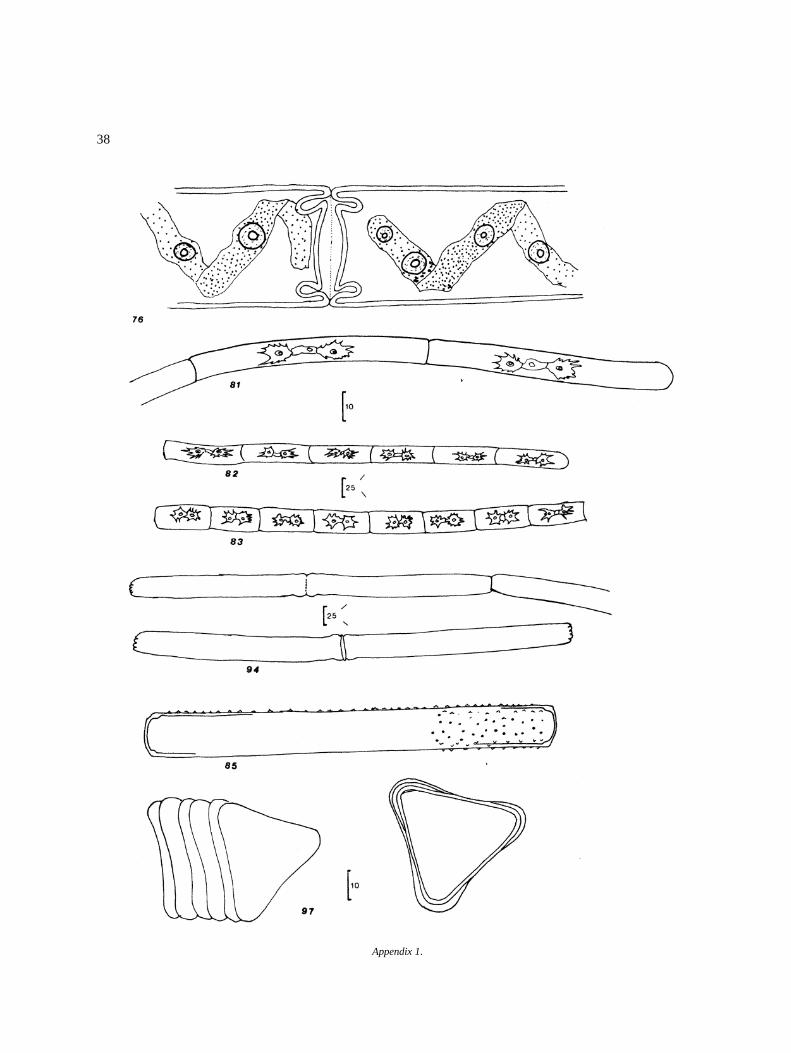
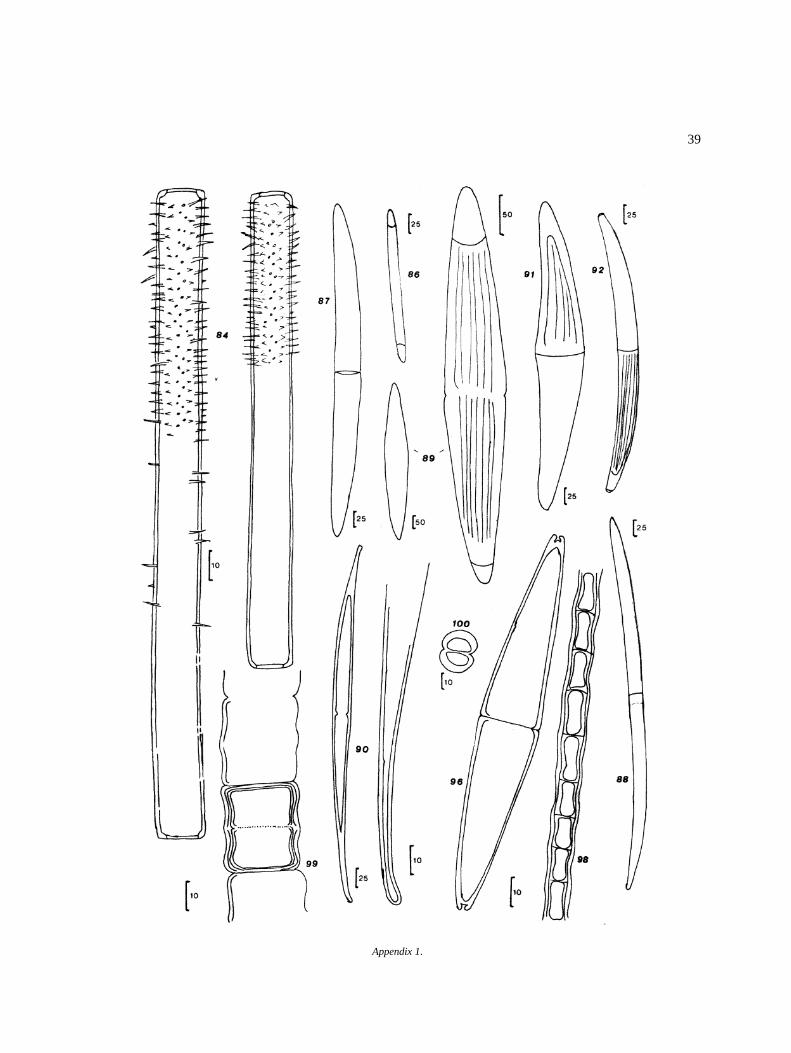
Appendix I
Algal species found in El Sol Lake, Nevado de Toluca,Mexico. Most species come from samples taken dur-ing April 1982–April 1983. Species reported by otherauthors in the lake (�) are included. Progressive num-bers in the right column correspond to that of figures inthe iconography. The bars in the icons are in microm-eters (�m).
DivisionClassOrderFamilyGenus and species Figure
CyanophytaChroococcalesChroococcaceae� Anacystis sp. (= Gloeothece, Aphanothece)Gloeocapsa aeruginosa (Carm.) Kutzing 2
Microcystis aeruginosa Kutzing 3M. robusta (= wesenbergii) (Clark) Nygaard 4Aphanocapsa rivularis (Carm.) Rabenhorst 5
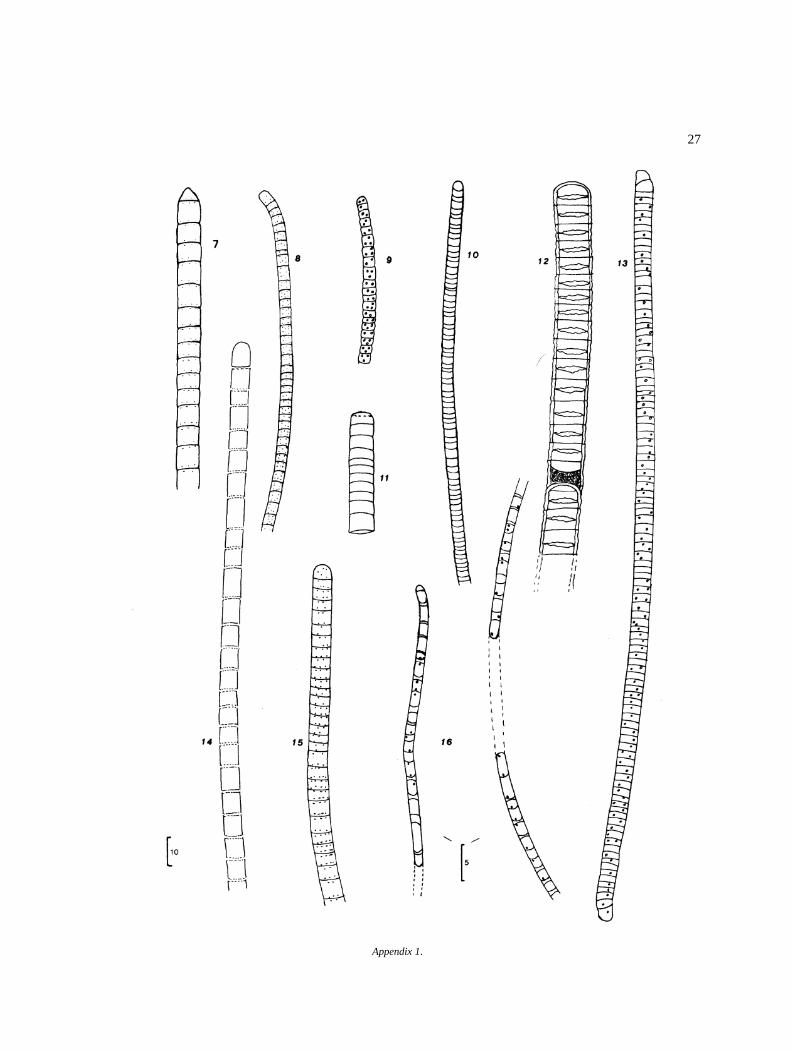
NostocalesOscillatoriaceaeOscillatoria curviceps Agardh 6O. formosa Bory 7O. jasorvensis Vouk 8O. mougeotii Kutzing 9O. obscura Bruhl & Biswas 10O. ornata Kutzing ex Gomont 11O. perornata Skuja 12O. proteus Skuja 13O. raoi De Toni, J. 14O. subbrevis Schmidle 15Lyngbya perelegans Lemmermann 16
NostocaceaeAulosira sp. 17Nostoc paludosum Kutzing 18Pseudoanabaenaconstricta (Szafer) Lauterborn 19ScytonemataceaeTolypothrix nodosa Bharadw 20
PyrrophytaDinophyceaePeridinialesPeridiniaceae� Peridinium lomnickii Woloszynska 21� P. willei Huitfeldt-Kaas 22CeratiaceaeCeratium sp.
Bacillariophyta
hy3739.tex; 11/11/1997; 13:42; v.5; p.7

24
CentrobacillariophyceaeEupodiscalesCoscinodiscaceae� Melosira sp.
PennatibacillariophyceaeFragilarialesFragilariaceae� Synedra sp.
NaviculalesNaviculaceae� Frustulia sp.
Cymbellaceae� Amphora sp.� Cymbella sp.Gomphonema sp.
SurirellalesSurirellaceae� Surirella sp.
ChrysophytaOchromonadalesDinobryaceaeDinobryon cylindricum v. alpinum(Imhof Bachmann) 30
Dinobryon sp. 31Xanthophyta
XanthophyceaeHeterococcalesCharaciopsidaceaeCharaciopsis longipes (Rab.) Borzi 32
EuglenophytaEuglenalesEuglenaceaeEuglena acus v. acus Ehrenberg 33
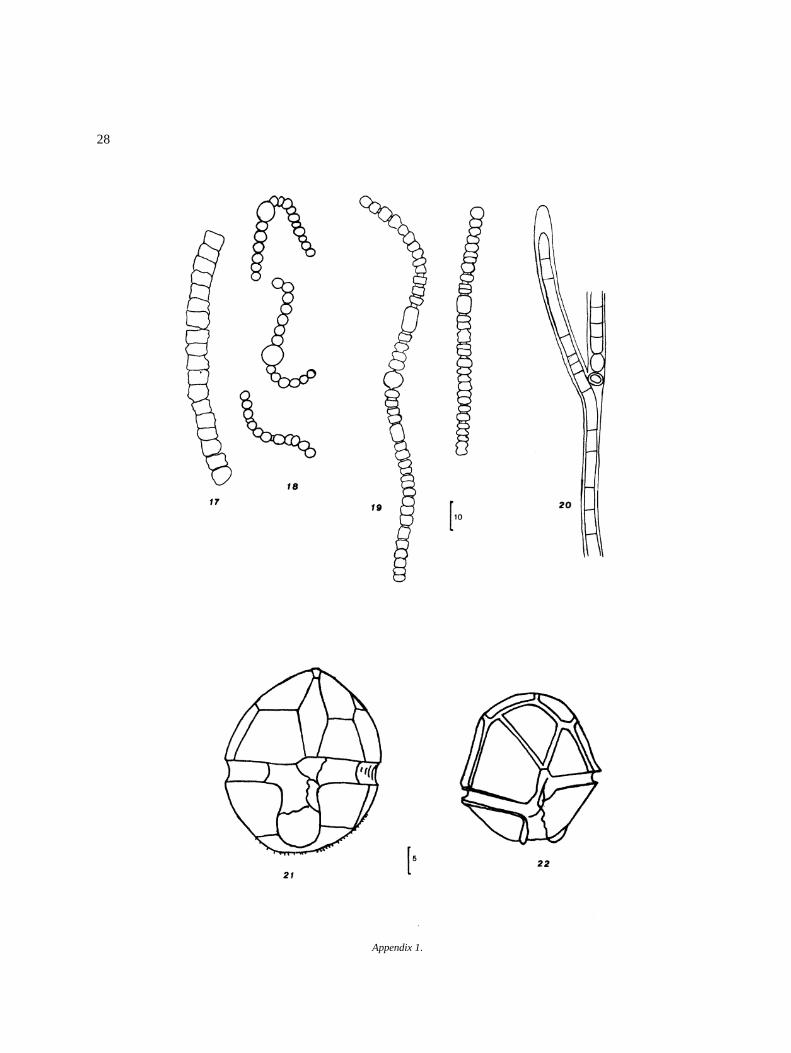
ChlorophytaVolvocalesTetrasporineaePalmellaceae� Sphaerocystis schroeteri Chodat
ChlorococcalesHydrodictyaceaeCharacium ornithocephalum A. Braun 35Pediastrum boryanum (Turpin) Meneghini 36P. duplex Meyen 37
ScenedesmaceaeScenedesmus armatus (Chodat) 38S. bijuga (ecornis ?) (Turpin) Lagerheim 39S. longispina v. longispina Chodat(= S. quadricauda v. longispina (Chodat)G.M. Smith) 40
S. quadricauda (Turpin) Brebisson 41S. quadricauda v. quadrispina (Chodat)
G.M. Smith 42UlotrichalesChaetophoraceaeAphanochaete repens Wolle 43Chaetonema ornatum Transeau 44Chaetophora incrassata Schrank 45Draparnaldia glomerata (Vaucher) Agardh 46D. simplex Meyer 47Stigeoclonium aestivale (Hazen) Collins 48S. pusillum (Lyngb.) Kutzing 49Thamniochaete huberi Gay 50Pseudoendoclonium sp. 51
ColeochaetaceaeChaetosphaeridium pringsheimii f. confertaKlebahn 52
OedogonialesOedogoniaceaeOedogonium acrosporum de Bary 53O. acrosporum v. bathmidosporum(Nordstedt) Hirn 54
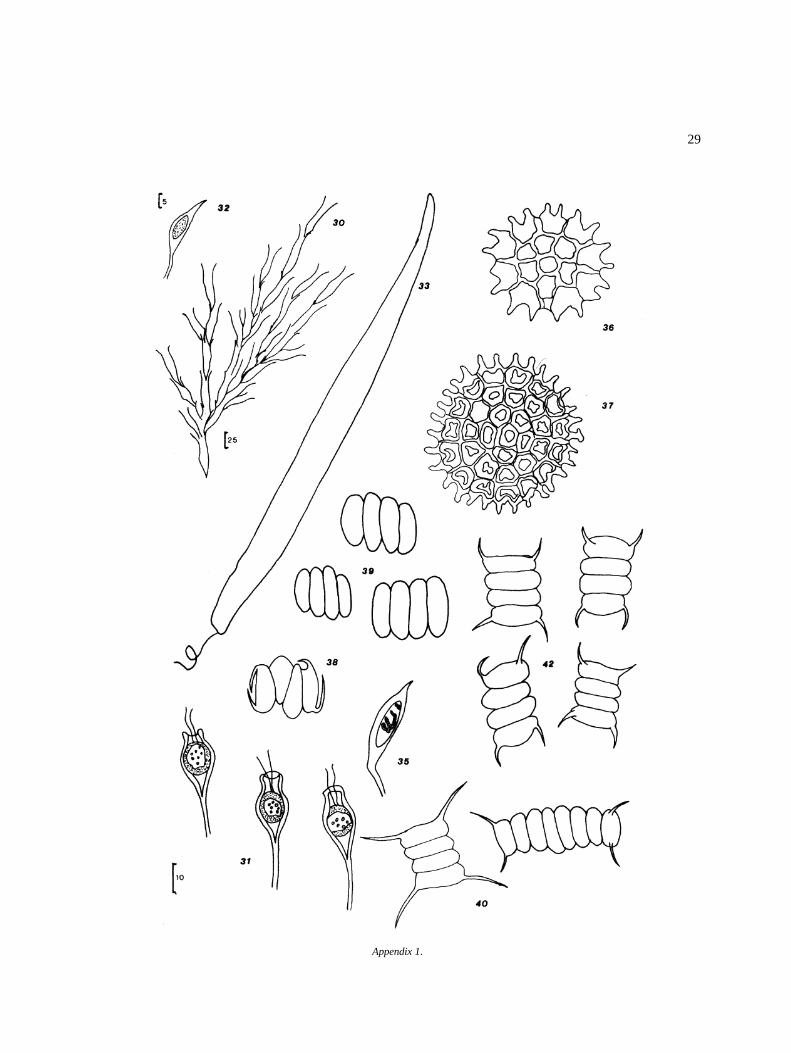
O. argentum HirnO. equinospermum A. Braun 56O. flavescens (Hans.) Wittrock 57O. grande v. majus Hansgirg 58O. inerme Hirn 59O. infimum Tiffany 60O. magnusii WittrockO. nanum WittrockO. rufescens v. exiguum Wittrock 63O. sexangulare v. majus Cleve 64O. sociale Wittrock 65O. suecicum f. australe Wittrock 66Bulbochaete congener Hirn 67B. debaryana Wittrock & Lund 68B. monile v. robusta Wittrock & Lund 69B. repanda Wittrock 70
ZygnematalesZynemataceaeMougeotia (?) sp.Sirogonium sticticum Kutzing 72Spirogyra jugalis Kutzing 73S. setiformis (Roth.) Kutzing 74Spirogyra sp. I 75Spirogyra sp. II 76Spirogyra sp. III 77Spirogyra sp. IV 78Spirogyra sp. a 79Spirogyra sp. b 80Zygnema o Zygnemopsis sp. 81Zygnema sp. 1 82
g cf. terrestre Randhawa
hy3739.tex; 11/11/1997; 13:42; v.5; p.8

25
Zygnema sp. 2 83MesotaeniaceaeGonatozygon aculeatum Hastings 84G. monotaenium de Bary 85
DesmidiaceaeClosterium abruptum West 86C. acerosum (Schrank) Ehrenberg 87C. acerosum v. elongatum Brebisson 88C. lunula Ehrenberg 89C. rostratum Ehrenberg 90C. ehrenberghii Meneghini 91C. striolatum Ehrenberg 92C. ulna FockePleurotaenium erhenbergii (Brebisson) de Bary 94� Euastrum oblongum (Grev.) RalfsTetmemorus granulatus f. minor Nordstedt 96Desmidium swartzii Agardh 97Hyalotheca undulata Nordstedt 98Cosmarium obliquum f. mayor (?) Nordstedt 99Cosmarium circulare 100� Micrasterias sp.Triploceras (?) sp.
CharophytaCharalesCharaceaeNitella gracilis v. intermedia (Nordst.) Wood 103
EspermatophytaAngiospermaEllatinaceaeEllatine brachysperma Gray
CyperaceaeEleocharis sp.
hy3739.tex; 11/11/1997; 13:42; v.5; p.9
26
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.10
27
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.11
28
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.12
29
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.13
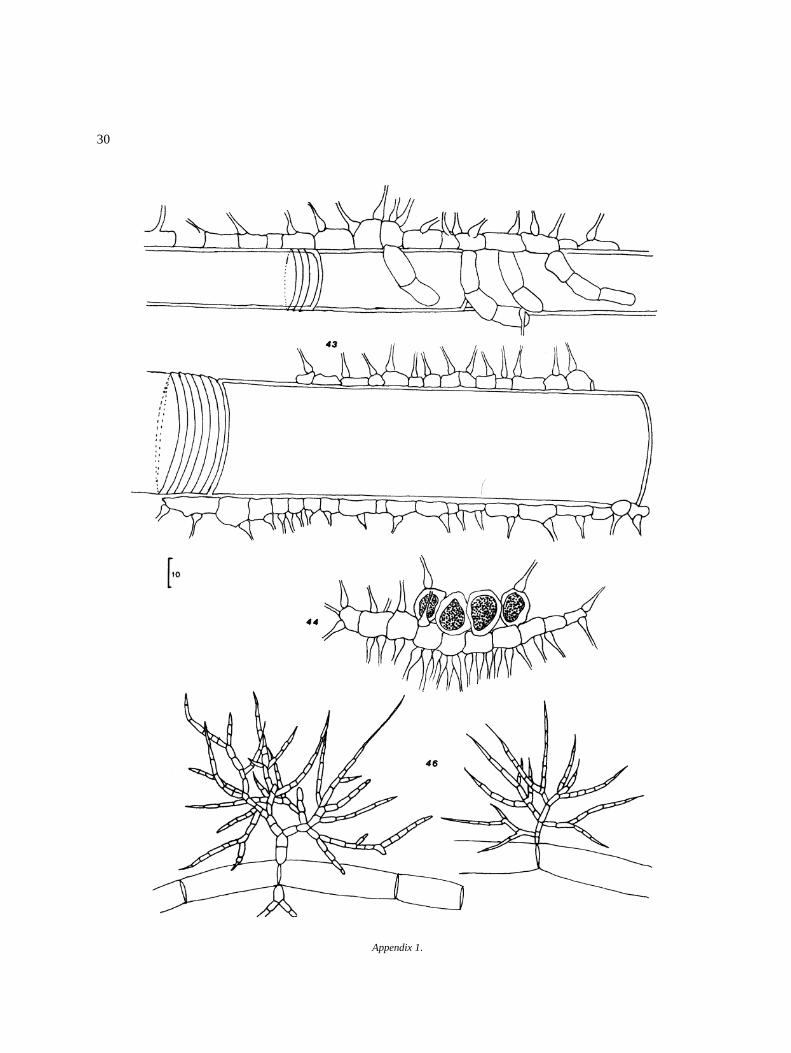
30
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.14
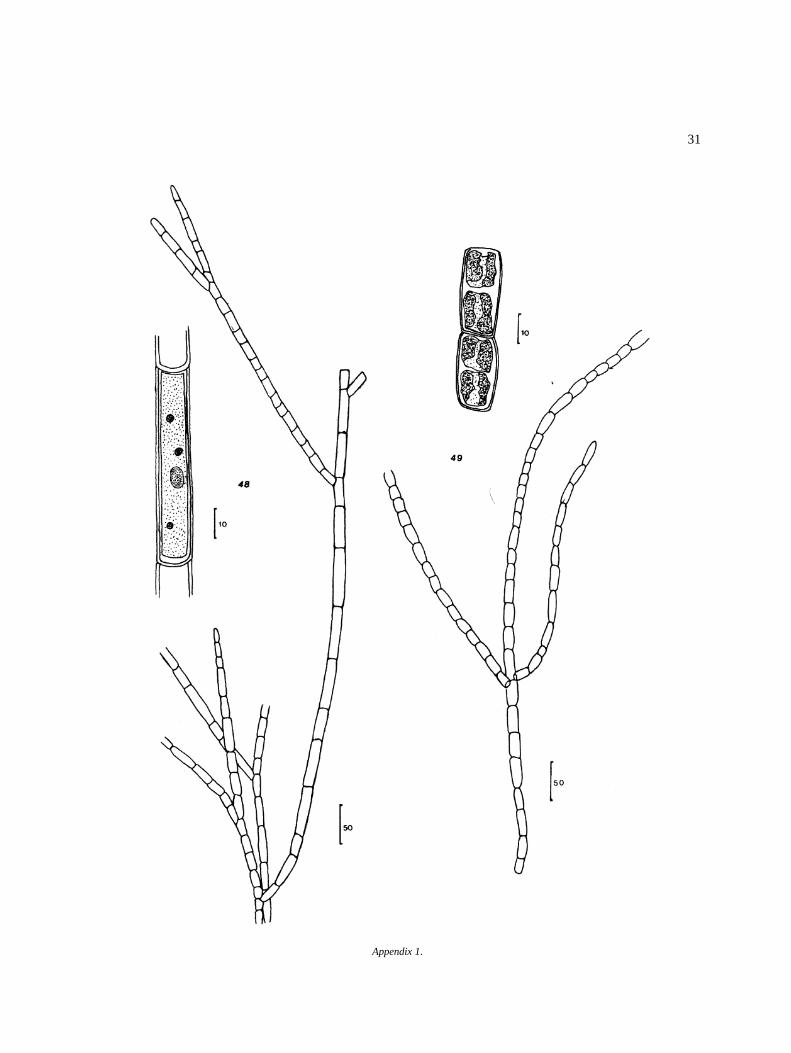
31
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.15
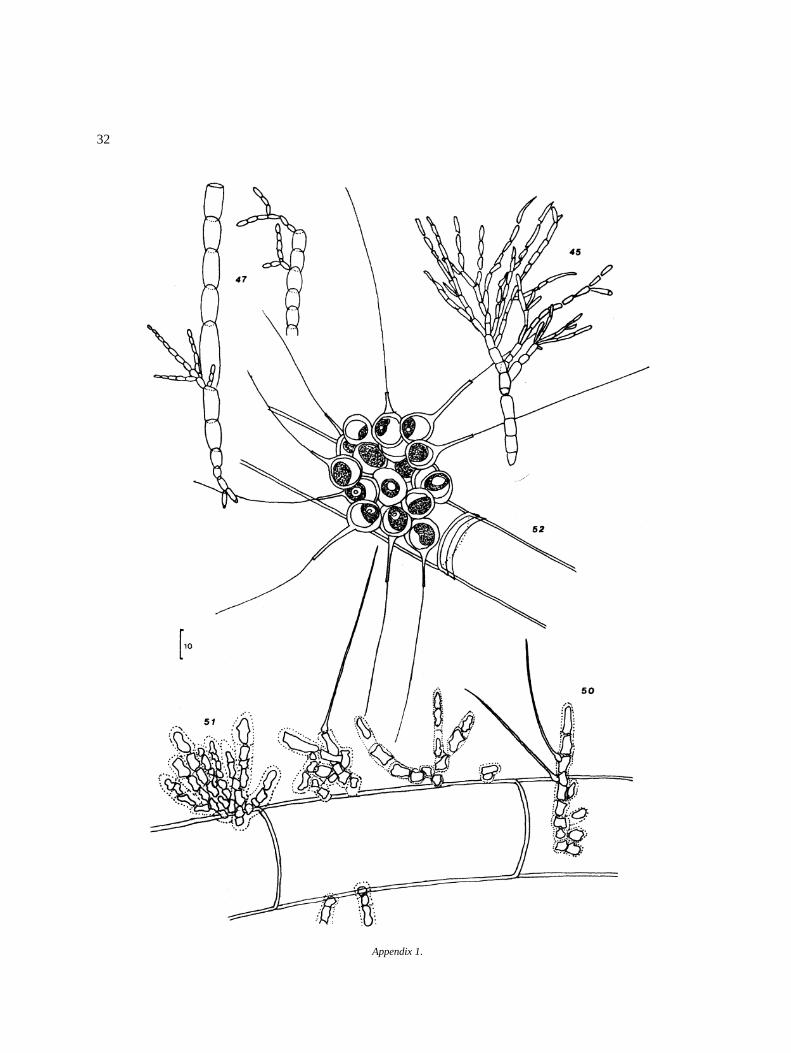
32
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.16
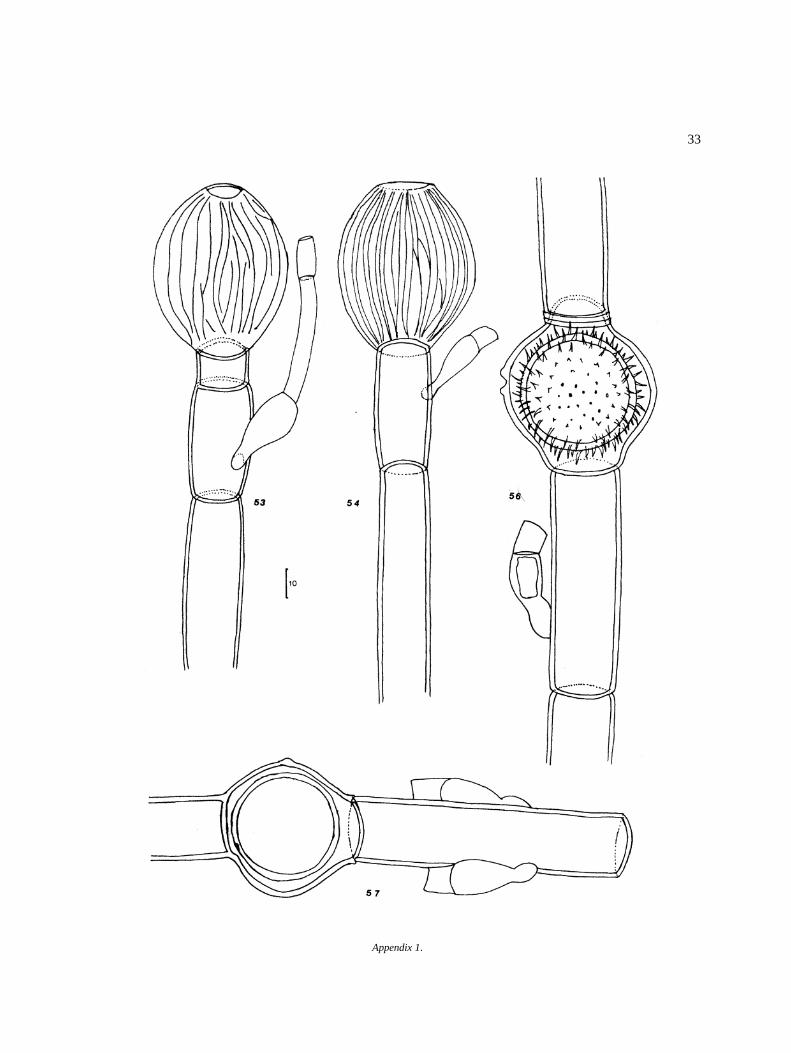
33
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.17
34
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.18
35
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.19
36
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.20
37
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.21
38
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.22
39
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.23

40
Appendix 1.
hy3739.tex; 11/11/1997; 13:42; v.5; p.24
Related Documents











