Fermentative hydrogen production by new marine Clostridium amygdalinum strain C9 isolated from offshore crude oil pipeline H.S. Jayasinghearachchi a , Sneha Singh a , Priyangshu M. Sarma a , Anil Aginihotri b , Banwari Lal a, * a Environmental and Industrial Biotechnology Division, The Energy and Resource Institute, Darbari Seth Block, Habitat Place, Lodhi Road, New Delhi, 110 003, India b Corporate HSF, Oil and Natural Gas Corporation, New Delhi, India article info Article history: Received 22 January 2010 Received in revised form 6 April 2010 Accepted 8 April 2010 Available online 21 May 2010 Keywords: Biohydrogen Clostridium amygdalinum Pentoses Starch abstract The present study investigated hydrogen production potential of novel marine Clostridium amygdalinum strain C9 isolated from oil water mixtures. Batch fermentations were carried out to determine the optimal conditions for the maximum hydrogen production on xylan, xylose, arabinose and starch. Maximum hydrogen production was pH and substrate dependant. The strain C9 favored optimum pH 7.5 (40 mmol H 2 /g xylan) from xylan, pH 7.5e8.5 from xylose (2.2e2.5 mol H 2 /mol xylose), pH 8.5 from arabinose (1.78 mol H 2 /mol arabinose) and pH 7.5 from starch (390 ml H 2 /g starch). But the strain C9 exhibited mixed type fermentation was exhibited during xylose fermentation. NaCl is required for the growth and hydrogen production. Distribution of volatile fatty acids was initial pH dependant and substrate dependant. Optimum NaCl requirement for maximum hydrogen production is substrate dependant (10 g NaCl/L for xylose and arabinose, and 7.5 g NaCl/L for xylan and starch). ª 2010 Professor T. Nejat Veziroglu. Published by Elsevier Ltd. All rights reserved. 1. Introduction Renewable energy resources have received considerable attention due to the depletion of fossil fuel and environmental pollution [1]. Among which, hydrogen is considered to be an ideal energy carrier with a high energy content of 122 KJ/g as it produces only water when it is combusted as a fuel or con- verted to electricity [1]. Hydrogen can be obtained via non-biological and biological processes. However, biological hydrogen production processes are friendlier to environment and less energy intensive than chemical and electrochemical processes. Further, dark fermentation is a more promising method considering its high evolution rate in the absence of any light source and versatility of the substrates used than photosyn- thetic hydrogen production [2]. At present, the dominant cost element in fermentative hydrogen production is the substrate [3]. Therefore, it is necessary to find low cost feedstock for commercial purposes. Renewable energy sources, such as cellulose, lignocelluloses or starch containing biomass constitute an abundant, inex- pensive and reliable raw material for biohydrogen production and offer considerable advantages [4,5]. Further, xylan, the major portion of hemicellulose of plant cell walls, and is the second most abundant renewable hemicellulosic poly- saccharide after cellulose [6]. Therefore, fermentative H 2 * Corresponding author. Tel.: þ91 11 24682100; fax: þ91 11 24682144. E-mail address: [email protected] (B. Lal). Available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/he international journal of hydrogen energy 35 (2010) 6665 e6673 0360-3199/$ e see front matter ª 2010 Professor T. Nejat Veziroglu. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.ijhydene.2010.04.034

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 3
Avai lab le a t www.sc iencedi rec t .com
journa l homepage : www.e lsev ier . com/ loca te /he
Fermentative hydrogen production by new marine Clostridiumamygdalinum strain C9 isolated from offshore crudeoil pipeline
H.S. Jayasinghearachchi a, Sneha Singh a, Priyangshu M. Sarma a, Anil Aginihotri b,Banwari Lal a,*aEnvironmental and Industrial Biotechnology Division, The Energy and Resource Institute, Darbari Seth Block, Habitat Place,
Lodhi Road, New Delhi, 110 003, IndiabCorporate HSF, Oil and Natural Gas Corporation, New Delhi, India
a r t i c l e i n f o
Article history:
Received 22 January 2010
Received in revised form
6 April 2010
Accepted 8 April 2010
Available online 21 May 2010
Keywords:
Biohydrogen
Clostridium amygdalinum
Pentoses
Starch
* Corresponding author. Tel.: þ91 11 2468210E-mail address: [email protected] (B. L
0360-3199/$ e see front matter ª 2010 Profedoi:10.1016/j.ijhydene.2010.04.034
a b s t r a c t
The present study investigated hydrogen production potential of novel marine Clostridium
amygdalinum strain C9 isolated from oil water mixtures. Batch fermentations were carried
out to determine the optimal conditions for the maximum hydrogen production on xylan,
xylose, arabinose and starch. Maximum hydrogen production was pH and substrate
dependant. The strain C9 favored optimum pH 7.5 (40 mmol H2/g xylan) from xylan, pH
7.5e8.5 from xylose (2.2e2.5 mol H2/mol xylose), pH 8.5 from arabinose (1.78 mol H2/mol
arabinose) and pH 7.5 from starch (390 ml H2/g starch). But the strain C9 exhibited mixed
type fermentation was exhibited during xylose fermentation. NaCl is required for the
growth and hydrogen production. Distribution of volatile fatty acids was initial pH
dependant and substrate dependant. Optimum NaCl requirement for maximum hydrogen
production is substrate dependant (10 g NaCl/L for xylose and arabinose, and 7.5 g NaCl/L
for xylan and starch).
ª 2010 Professor T. Nejat Veziroglu. Published by Elsevier Ltd. All rights reserved.
1. Introduction considering its high evolution rate in the absence of any light
Renewable energy resources have received considerable
attention due to the depletion of fossil fuel and environmental
pollution [1]. Among which, hydrogen is considered to be an
ideal energy carrier with a high energy content of 122 KJ/g as it
produces only water when it is combusted as a fuel or con-
verted to electricity [1].
Hydrogen can be obtained via non-biological and biological
processes. However, biological hydrogen production
processes are friendlier to environment and less energy
intensive than chemical and electrochemical processes.
Further, dark fermentation is a more promising method
0; fax: þ91 11 24682144.al).ssor T. Nejat Veziroglu. P
source and versatility of the substrates used than photosyn-
thetic hydrogen production [2].
At present, the dominant cost element in fermentative
hydrogen production is the substrate [3]. Therefore, it is
necessary to find low cost feedstock for commercial purposes.
Renewable energy sources, such as cellulose, lignocelluloses
or starch containing biomass constitute an abundant, inex-
pensive and reliable raw material for biohydrogen production
and offer considerable advantages [4,5]. Further, xylan, the
major portion of hemicellulose of plant cell walls, and is the
second most abundant renewable hemicellulosic poly-
saccharide after cellulose [6]. Therefore, fermentative H2
ublished by Elsevier Ltd. All rights reserved.

i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 36666
production from cellulosic feedstock or from lignocellulosic
wastes could be competitive with fossil fuel-derived H2,
providing a plausible approach to practical biohydrogen
production in the future [7].
Further, it is interestingly noted that any studies have been
conducted to screen hydrogen producing bacteria from fresh
water fields. But few studies have been identified salt-tolerant
strains from fresh water fields. Hydrogen producing bacterial
strains can be directly screened from marine environments,
but few such bacteria can be cultivated in the laboratory.
Therefore, aim of the present study was to report moderately
halophilic novel marine strain of Clostridium amygdalinum
strain C9 which can effectively utilize xylan, xylose and arab-
inose to produce hydrogen under mesophilic condition, iso-
lated fromoil watermixtures. Hence, the presentworkmainly
focuses on the optimization of the efficiency of fermentative
hydrogen production under different pH, temperature, NaCl
and different substrate concentrations.
2. Methods
2.1. Oil water samples collection
Oil water mixture samples (97% oil, 3% water) were collected
from offshore sea buried crude oil pipelines in Bombay High,
India into 50 ml of anaerobic sterilized serum bottles con-
taining 1 ml of 2% Na2S. Samples were collected during
December, 2007eJanuary, 2008.
2.2. Medium preparation
PeptoneYeast (PY) liquidmediumcomposedof (g/L);Peptone20,
yeast extract 5,NaCl 5,Na2S2O3 0.03, ferric citrate 0.03,was used
for the isolation of the strain C9 Cysteine-hydro chloride was
added to a final concentration of 300 mg/L. The pH of the
medium was adjusted to 7.5 with 1N KOH at an ambient
temperature before autoclaving. The PYB medium was boiled
under a stream of O2-free N2 gas and aliquots of 6 ml were
anaerobically dispensed into 10 ml serum bottles. The bottles
were then sealed and autoclaved for 20 min at 120 �C. Prior toinoculation, 0.1ml of 2%filter sterilizedNa2S.9H2O and 0.1ml of
10% filter sterilized Na2CO3were injected separately by syringe.
2.3. Isolation of the strain C9
Samples (0.1 ml) were inoculated in liquid PY medium. All the
culture bottleswere incubated at 40 �C for 3e5 days depending
on the growth. Sub culturing was done in the same medium
after 3 days of inoculation. All the sub cultures were incubated
at 40 �C under atmospheric pressure.
The sub cultures were serially diluted and 10 mL of serially
diluted cultures (10�7 and 10�8 dilutions) were plated on
anaerobically prepared PY solid medium that was solidified
with 12% agar. Culture plates were incubated at 40 �C in the
anaerobic chamber (N2:H2:CO2; 5:5:90) for 2e4 days till colo-
nies appeared [8]. Culture plateswere observed daily andwell-
isolated colonies were transferred to the liquid PYP medium
with the help of sterile loop. Liquid cultures were incubated at
40 �C as described above. Gram staining was routinely
performed to check the purity of the cultures. Cells were
observed under a light microscope (Olympus, Japan) and pure
strain C9 was routinely cultivated in anaerobically prepared
Trypticase Soy Broth (TSB).
2.4. Strain identification
GenomicDNAfromthepure isolateC9wasextractedaccording
to the previously described procedures [9]. The 16S rRNA gene
of the isolatewas amplifiedby PCRusingMicroSec full GeneKit
(Applied Biosystems, UK) as per the manufactures instruc-
tions. Amplified product was sequenced using the DyeDeoxy
Terminator Cycle sequencing Kit (Applied Biosystems, UK) as
directed in the manufacturer’s protocol with an automatic
DNA sequencer (Model 300; Applied Biosystems, USA).
The sequence of the strain C9 was compared with other
related sequences available in GenBank using BLAST pro-
gramme [10]. Further, the sequence of strain C9 was aligned
with closely related sequences found in GenBank, using
CLUSTAL W, and pair wise evolutionary distances were
computed using Jukes-Canter Model [11]. Phylogenetic anal-
ysis was performed using Phylip version 3.67 [12] and
a phylogenetic tree was constructed using the neighbour-
joining algorithm [13]. Confidence in the tree topologies was
evaluated by re-sampling 100 bootstrap trees [14].
2.5. Study of the growth properties of the strain C9
The strain was cultured in liquid basal anaerobic medium
composed of (g/L): NH4Cl2 0.5; Yeast 5; K2HPO4 0.25; KCl 0.002;
MgCl2 .6H2O 0.125; NH4HCO3 0.4; Peptone 1; NH4H2PO4 0.4;
NaH2PO4 0.5 and 1ml of trace element solution. Trace element
solution was composed of (mg/L): CaCl2 50; MgCl2.6H2O 100;
FeCl2 25; NaCl 10; CoCl2.6H2O 5; MnCl2.4H2O 5; AlCl3 2.5;
(NH4)6MO7O25 15; H3PO4 5; NiCl2 5; CuCl2.5H2O 5; ZnCl2 5.
Temperature profile (35, 40, 45 and 55 �C) and NaCl require-
ment (0.0e7%) (w/v) for the growth of the strain C9 was
studied in anaerobically prepared liquid basal medium sup-
plementedwith 5 g/L glucose as carbon source. Further, ability
of utilization of different carbon sources for the growth of the
strain C9 was tested in liquid basal medium described above.
Following carbon sources were used at a final concentration of
5 g/L: sucrose, fructose, xylose, xylan, ribose, raffinose, ram-
nose, arabinose, xylan, rhamnose, galactose, cellobiose,
starch and cellulose. All the growth experiments were per-
formed in triplicate, using 60 ml serum bottles containing
10 ml of anaerobic basal medium.
2.5.1. Optimization of hydrogen production on differentcarbon sources in batch fermentationBatch dark fermentation experiments were carried out in 1 L
anaerobic bioreactor with 50 ml of liquid anaerobic basal
medium. Reducing agent such as Na2S or L-cysteine hydro-
chloride was not used during this study. Pre-grown inoculum
acquired during exponential growth phase was added at 5%
(v/v) using air tight sterilized syringe.
Influence of different initial pH levels for the hydrogen
production on different substrates (xylan, xylose, starch and
arabinose) was studied. The initial pH levels ranging from 5.5
to 9.5 with 0.5 increments were selected for this study. The pH

Fig. 1 e Neighbour-joining tree showing the phylogenetic
position of isolated strain, Clostridium spp. C9, based on the
16S rRNA sequences of the genus Clostridium. The numbers
at the nodes indicate the levels of bootstrap percentages
based on the neighbour-joining of 100 replicates. Bar
indicates the nucleotide substitution per site. GenBank
accession numbers are given in parenthesis.
i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 3 6667
of the mediumwas adjusted with 1M HCl or 1M NaOH prior to
dispensing the medium into the 1 L anaerobic bioreactors.
Influence of different NaCl concentration on growth and
hydrogen production was also studied. Different NaCl
concentrations testedwere 2.5, 7.5, 10, 20 and 25, 35 and 45 g/L.
Further, effect of initial substrate concentration on
hydrogenproductionwas investigated inbatchexperimentsas
described above. Optimal pH levels exhibited by the strain C9
for themaximumhydrogenproduction ondifferent substrates
wereusedas initial culturepH. Initial substrate concentrations
testedwere5, 10, 15, 20and25g/L for each individual substrate.
Distribution of secondary metabolites during fermentation
on different substrates at different pH levels studied. Further,
influenceof initial substrateconcentrationonthedistributionof
various secondary metabolites was also analysed in this study.
2.6. Invivo hydrogen evolution assay undermicroaerophilic conditions
In vivo hydrogen evolution assay under microaerophilic
condition was measured by using a gas chromatograph as
described below. Methylviolegen reduced by sodium dithion-
ite was used as an electron donor. Ten millilitres of serum
vials with butyl rubber stoppers were used for the assay. The
assay mixture contains: 1080e1380 mL of 100 mM potassium
phosphate (K2HPO4/KH2PO4) pH 6.8e7; 200 mL of 1 M sodium
dithionite; 200 mL of 10% Triton X-100; 200e500 mL of anaero-
bically grown culture (OD600nm 1.2). The reaction vessel was
afterwards closed up with the rubber stopper and crimpled
with aluminium caps. Then the serum bottles were removed
from the anaerobic chamber. The sealed bottles were vortex
for 10 s. Different amounts of air (1e4 ml) were added to the
reaction vessel with a syringe and incubated reaction vessels
in a water bath at 37 �C for 40 min. After incubation, 500 mL of
headspace gas samples were analysed. The assay was con-
ducted in triplicate.
2.7. Analytical methods
Bacterial cell mass in each individual bioreactor was deter-
mined by monitoring optical absorbance at OD600 nm in the
Hitachi U-2000 spectrophotometer (Tokyo, Japan) [20].
The headspace gas composition in each bioreactor was
directly analysed by the gas chromatography (GC 6890N, Agi-
lent, USA) equipped with thermal conductivity detector and
HP PLOTQ column (15M� 530 mm� 40 mm) as described in our
previous study [15].
Secondary metabolic products (Acetate, butyrate, iso-
butrate, valerate, isovalerate and propionate) in liquid phase
were measured with GC 6890N (Agilent, USA) equipped with
flame ionization detector and DB-WAXetr column
(30 m � 530 mm � 1 mm). High Performance Liquid Chroma-
tography (HPLC, Agilent 1100 series, USA) equipped with
Sugar-PAK.1 column (Water Research, USA) was used for the
detection of remaining sugars and ethanol production [15].
Further HPLC (Agilent 1100 series, USA) equipped with Ani-
nexR HPX-87H, (300mm� 7.8mm) column (BIO RAD, CA, USA)
was used to quantify succinate, formate and oleic acid
production. Sulphuric acid (0.0008 N) was used as the mobile
phase at a flow rate of 0.6 ml/min.
3. Results and discussion
3.1. General properties of the strain C9
The strain C9 was a facultative anaerobe and able to grow
under microaerophilic condition. The strain C9 grew from 30
to 42 �C with 37 �C for optimum growth. Weaker growth was
observed at 45 �C and no growth was observed beyond 45 �C.The strain C9 was moderately halophilic and able to grow up
to 6% NaCl (w/v) with optimum of 2e2.5% NaCl (w/v) in the
presence of glucose as a sole carbon source. The strain C9
could utilize glucose, sucrose, fructose, arabinose, xylan,
xylose, starch and ribose, raffinose, rhamose, cellobiose and
cellulose. The stain C9 could not utilized pectin.
3.2. Identification of the strain C9 based on the 16SrRNA gene sequence analysis
The nucleotide sequence of 16S rRNA gene of the strain C9was
deposited in the GenBank under accession of EU862317.
Consequently, the16S rRNAgenesequenceof thestrainC9was
compared with other sequences available in GenBank using
BLAST algorithm. BLAST search against the GenBank database
revealed that the strain C9 was belonging to the genus Clos-
tridium with all three algorithms used (neighbor-joining,
maximum parsimony and maximum likelihood) (Fig. 1).
Further the strain C9 was closely related to the C.
i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 36668
saccharolyticum (DSM2544) andC. amygdalinum (ATCCBAA-501)
with 98% similarity. Moreover, the strain C9 is closer to the
Clostridium xylanolyticum (ATCC 4963), Clostridium aerotoleras
(DSM 5434) and Clostridium celerecrescenswith 96% similarity.
3.3. Optimization of pH on hydrogen production ondifferent substrates under mesophilic condition
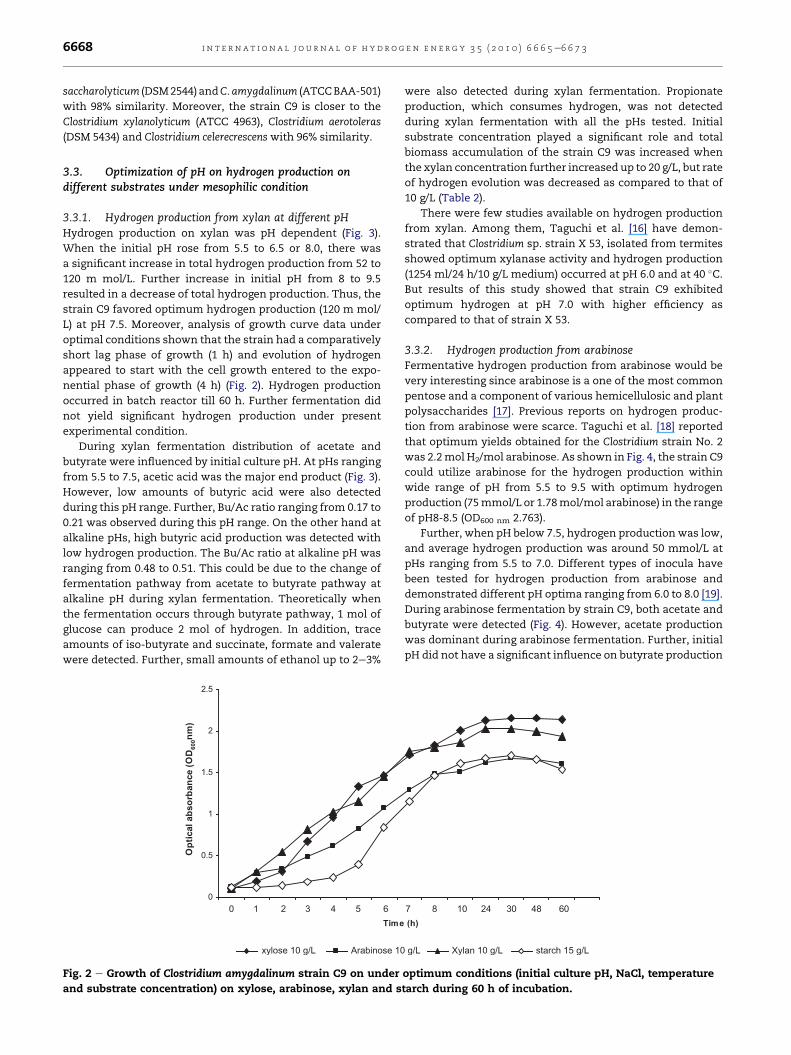
3.3.1. Hydrogen production from xylan at different pHHydrogen production on xylan was pH dependent (Fig. 3).
When the initial pH rose from 5.5 to 6.5 or 8.0, there was
a significant increase in total hydrogen production from 52 to
120 m mol/L. Further increase in initial pH from 8 to 9.5
resulted in a decrease of total hydrogen production. Thus, the
strain C9 favored optimum hydrogen production (120 m mol/
L) at pH 7.5. Moreover, analysis of growth curve data under
optimal conditions shown that the strain had a comparatively
short lag phase of growth (1 h) and evolution of hydrogen
appeared to start with the cell growth entered to the expo-
nential phase of growth (4 h) (Fig. 2). Hydrogen production
occurred in batch reactor till 60 h. Further fermentation did
not yield significant hydrogen production under present
experimental condition.
During xylan fermentation distribution of acetate and
butyrate were influenced by initial culture pH. At pHs ranging
from 5.5 to 7.5, acetic acid was the major end product (Fig. 3).
However, low amounts of butyric acid were also detected
during this pH range. Further, Bu/Ac ratio ranging from 0.17 to
0.21 was observed during this pH range. On the other hand at
alkaline pHs, high butyric acid production was detected with
low hydrogen production. The Bu/Ac ratio at alkaline pH was
ranging from 0.48 to 0.51. This could be due to the change of
fermentation pathway from acetate to butyrate pathway at
alkaline pH during xylan fermentation. Theoretically when
the fermentation occurs through butyrate pathway, 1 mol of
glucose can produce 2 mol of hydrogen. In addition, trace
amounts of iso-butyrate and succinate, formate and valerate
were detected. Further, small amounts of ethanol up to 2e3%
Fig. 2 e Growth of Clostridium amygdalinum strain C9 on under
and substrate concentration) on xylose, arabinose, xylan and s
were also detected during xylan fermentation. Propionate
production, which consumes hydrogen, was not detected
during xylan fermentation with all the pHs tested. Initial
substrate concentration played a significant role and total
biomass accumulation of the strain C9 was increased when
the xylan concentration further increased up to 20 g/L, but rate
of hydrogen evolution was decreased as compared to that of
10 g/L (Table 2).
There were few studies available on hydrogen production
from xylan. Among them, Taguchi et al. [16] have demon-
strated that Clostridium sp. strain X 53, isolated from termites
showed optimum xylanase activity and hydrogen production
(1254 ml/24 h/10 g/L medium) occurred at pH 6.0 and at 40 �C.But results of this study showed that strain C9 exhibited
optimum hydrogen at pH 7.0 with higher efficiency as
compared to that of strain X 53.
3.3.2. Hydrogen production from arabinoseFermentative hydrogen production from arabinose would be
very interesting since arabinose is a one of the most common
pentose and a component of various hemicellulosic and plant
polysaccharides [17]. Previous reports on hydrogen produc-
tion from arabinose were scarce. Taguchi et al. [18] reported
that optimum yields obtained for the Clostridium strain No. 2
was 2.2mol H2/mol arabinose. As shown in Fig. 4, the strain C9
could utilize arabinose for the hydrogen production within
wide range of pH from 5.5 to 9.5 with optimum hydrogen
production (75mmol/L or 1.78mol/mol arabinose) in the range
of pH8-8.5 (OD600 nm 2.763).
Further, when pH below 7.5, hydrogen production was low,
and average hydrogen production was around 50 mmol/L at
pHs ranging from 5.5 to 7.0. Different types of inocula have
been tested for hydrogen production from arabinose and
demonstrated different pH optima ranging from 6.0 to 8.0 [19].
During arabinose fermentation by strain C9, both acetate and
butyrate were detected (Fig. 4). However, acetate production
was dominant during arabinose fermentation. Further, initial
pH did not have a significant influence on butyrate production
optimum conditions (initial culture pH, NaCl, temperature
tarch during 60 h of incubation.
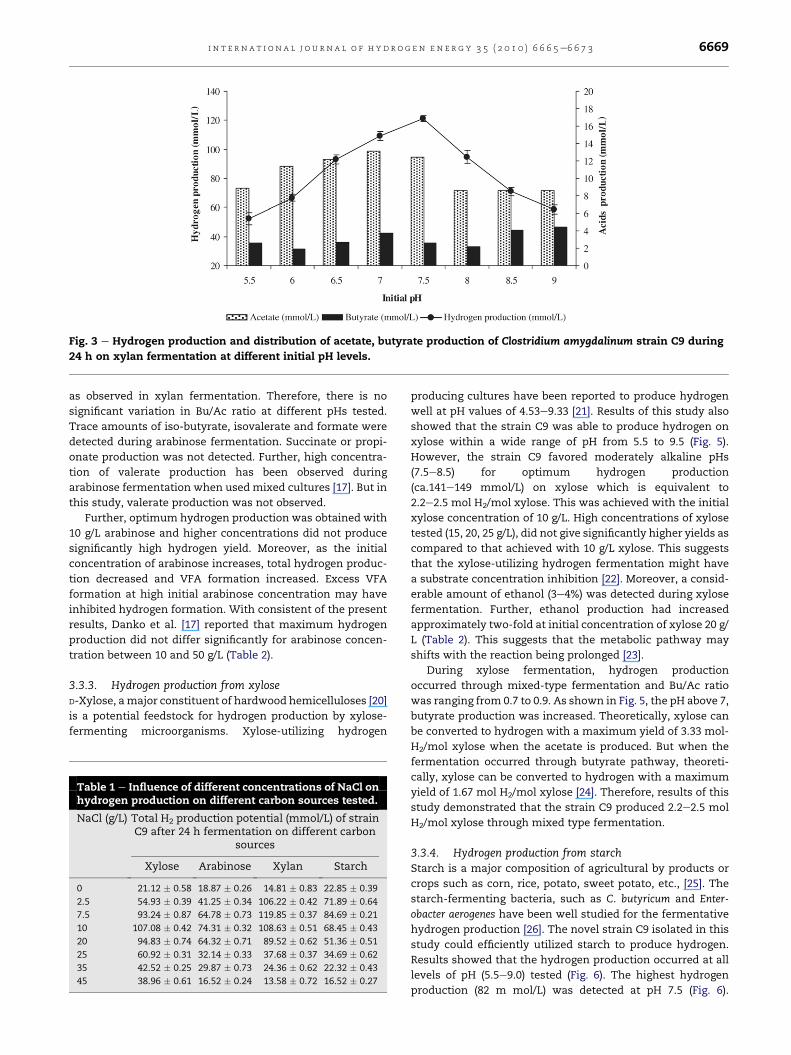
Fig. 3 e Hydrogen production and distribution of acetate, butyrate production of Clostridium amygdalinum strain C9 during
24 h on xylan fermentation at different initial pH levels.
i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 3 6669
as observed in xylan fermentation. Therefore, there is no
significant variation in Bu/Ac ratio at different pHs tested.
Trace amounts of iso-butyrate, isovalerate and formate were
detected during arabinose fermentation. Succinate or propi-
onate production was not detected. Further, high concentra-
tion of valerate production has been observed during
arabinose fermentation when usedmixed cultures [17]. But in
this study, valerate production was not observed.
Further, optimum hydrogen production was obtained with
10 g/L arabinose and higher concentrations did not produce
significantly high hydrogen yield. Moreover, as the initial
concentration of arabinose increases, total hydrogen produc-
tion decreased and VFA formation increased. Excess VFA
formation at high initial arabinose concentration may have
inhibited hydrogen formation. With consistent of the present
results, Danko et al. [17] reported that maximum hydrogen
production did not differ significantly for arabinose concen-
tration between 10 and 50 g/L (Table 2).
3.3.3. Hydrogen production from xyloseD-Xylose, amajor constituent of hardwood hemicelluloses [20]
is a potential feedstock for hydrogen production by xylose-
fermenting microorganisms. Xylose-utilizing hydrogen
Table 1 e Influence of different concentrations of NaCl onhydrogen production on different carbon sources tested.
NaCl (g/L) Total H2 production potential (mmol/L) of strainC9 after 24 h fermentation on different carbon
sources
Xylose Arabinose Xylan Starch
0 21.12 � 0.58 18.87 � 0.26 14.81 � 0.83 22.85 � 0.39
2.5 54.93 � 0.39 41.25 � 0.34 106.22 � 0.42 71.89 � 0.64
7.5 93.24 � 0.87 64.78 � 0.73 119.85 � 0.37 84.69 � 0.21
10 107.08 � 0.42 74.31 � 0.32 108.63 � 0.51 68.45 � 0.43
20 94.83 � 0.74 64.32 � 0.71 89.52 � 0.62 51.36 � 0.51
25 60.92 � 0.31 32.14 � 0.33 37.68 � 0.37 34.69 � 0.62
35 42.52 � 0.25 29.87 � 0.73 24.36 � 0.62 22.32 � 0.43
45 38.96 � 0.61 16.52 � 0.24 13.58 � 0.72 16.52 � 0.27
producing cultures have been reported to produce hydrogen
well at pH values of 4.53e9.33 [21]. Results of this study also
showed that the strain C9 was able to produce hydrogen on
xylose within a wide range of pH from 5.5 to 9.5 (Fig. 5).
However, the strain C9 favored moderately alkaline pHs
(7.5e8.5) for optimum hydrogen production
(ca.141e149 mmol/L) on xylose which is equivalent to
2.2e2.5 mol H2/mol xylose. This was achieved with the initial
xylose concentration of 10 g/L. High concentrations of xylose
tested (15, 20, 25 g/L), did not give significantly higher yields as
compared to that achieved with 10 g/L xylose. This suggests
that the xylose-utilizing hydrogen fermentation might have
a substrate concentration inhibition [22]. Moreover, a consid-
erable amount of ethanol (3e4%) was detected during xylose
fermentation. Further, ethanol production had increased
approximately two-fold at initial concentration of xylose 20 g/
L (Table 2). This suggests that the metabolic pathway may
shifts with the reaction being prolonged [23].
During xylose fermentation, hydrogen production
occurred through mixed-type fermentation and Bu/Ac ratio
was ranging from 0.7 to 0.9. As shown in Fig. 5, the pH above 7,
butyrate production was increased. Theoretically, xylose can
be converted to hydrogen with a maximum yield of 3.33 mol-
H2/mol xylose when the acetate is produced. But when the
fermentation occurred through butyrate pathway, theoreti-
cally, xylose can be converted to hydrogen with a maximum
yield of 1.67 mol H2/mol xylose [24]. Therefore, results of this
study demonstrated that the strain C9 produced 2.2e2.5 mol
H2/mol xylose through mixed type fermentation.
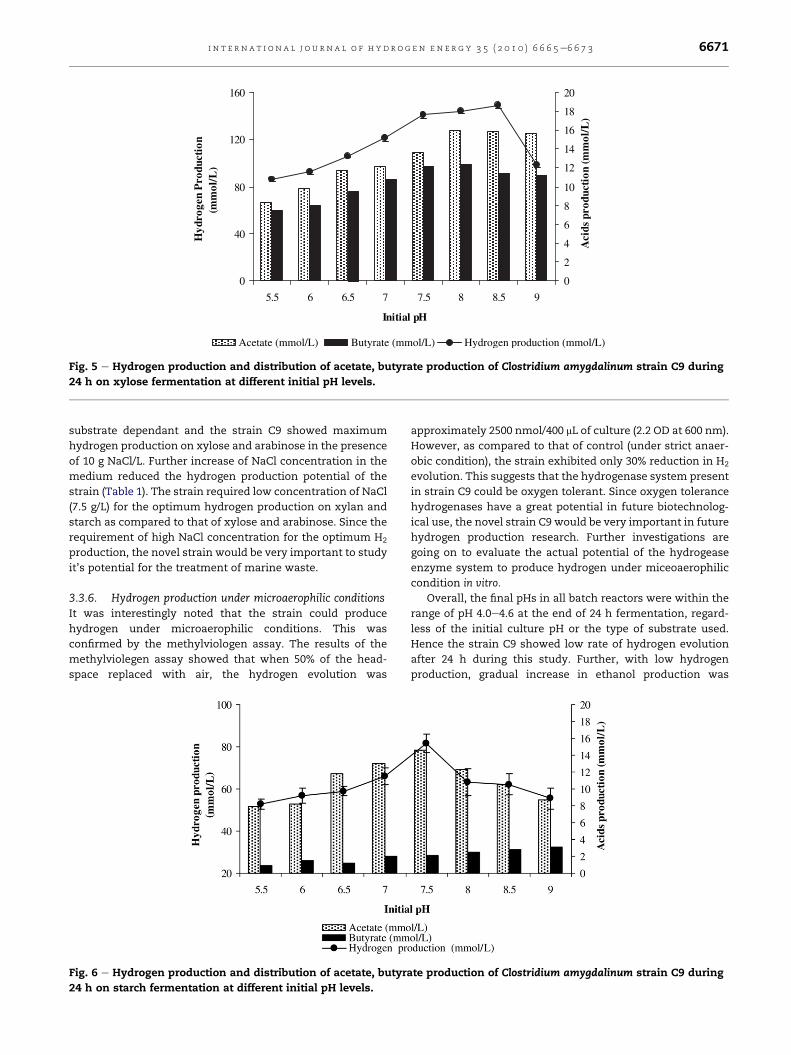
3.3.4. Hydrogen production from starchStarch is a major composition of agricultural by products or
crops such as corn, rice, potato, sweet potato, etc., [25]. The
starch-fermenting bacteria, such as C. butyricum and Enter-
obacter aerogenes have been well studied for the fermentative
hydrogen production [26]. The novel strain C9 isolated in this
study could efficiently utilized starch to produce hydrogen.
Results showed that the hydrogen production occurred at all
levels of pH (5.5e9.0) tested (Fig. 6). The highest hydrogen
production (82 m mol/L) was detected at pH 7.5 (Fig. 6).
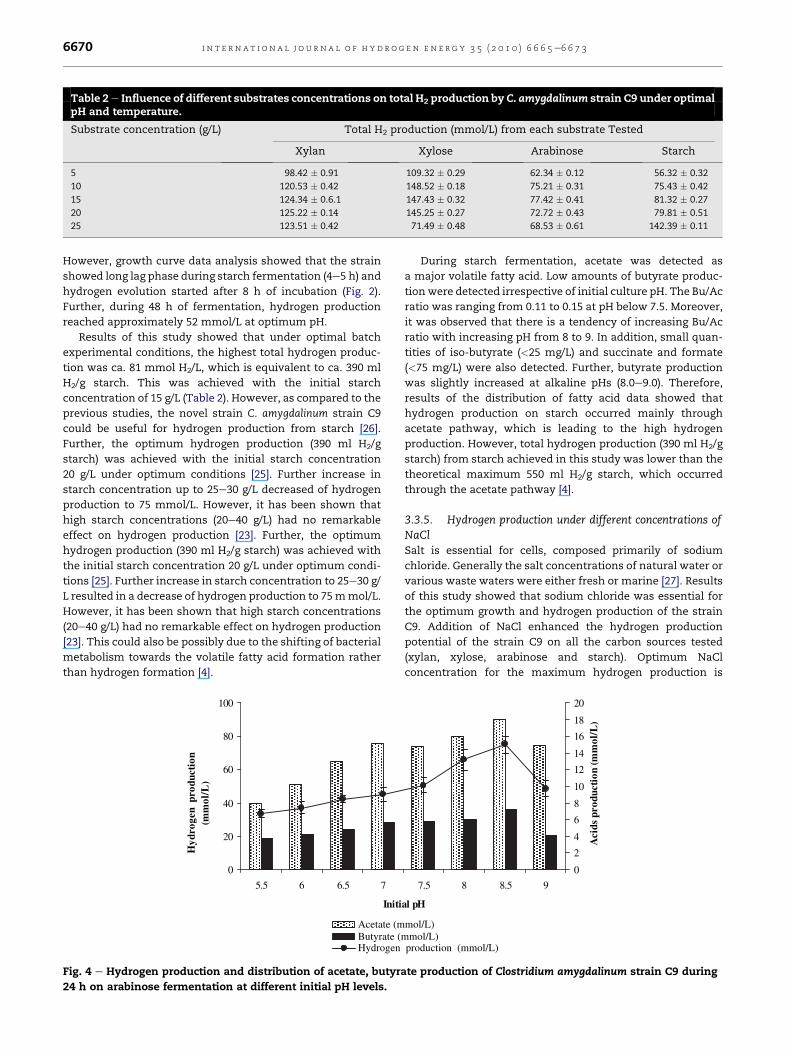
Table 2e Influence of different substrates concentrations on total H2 production by C. amygdalinum strain C9 under optimalpH and temperature.
Substrate concentration (g/L) Total H2 production (mmol/L) from each substrate Tested
Xylan Xylose Arabinose Starch
5 98.42 � 0.91 109.32 � 0.29 62.34 � 0.12 56.32 � 0.32
10 120.53 � 0.42 148.52 � 0.18 75.21 � 0.31 75.43 � 0.42
15 124.34 � 0.6.1 147.43 � 0.32 77.42 � 0.41 81.32 � 0.27
20 125.22 � 0.14 145.25 � 0.27 72.72 � 0.43 79.81 � 0.51
25 123.51 � 0.42 71.49 � 0.48 68.53 � 0.61 142.39 � 0.11
i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 36670
However, growth curve data analysis showed that the strain
showed long lag phase during starch fermentation (4e5 h) and
hydrogen evolution started after 8 h of incubation (Fig. 2).
Further, during 48 h of fermentation, hydrogen production
reached approximately 52 mmol/L at optimum pH.
Results of this study showed that under optimal batch
experimental conditions, the highest total hydrogen produc-
tion was ca. 81 mmol H2/L, which is equivalent to ca. 390 ml
H2/g starch. This was achieved with the initial starch
concentration of 15 g/L (Table 2). However, as compared to the
previous studies, the novel strain C. amygdalinum strain C9
could be useful for hydrogen production from starch [26].
Further, the optimum hydrogen production (390 ml H2/g
starch) was achieved with the initial starch concentration
20 g/L under optimum conditions [25]. Further increase in
starch concentration up to 25e30 g/L decreased of hydrogen
production to 75 mmol/L. However, it has been shown that
high starch concentrations (20e40 g/L) had no remarkable
effect on hydrogen production [23]. Further, the optimum
hydrogen production (390 ml H2/g starch) was achieved with
the initial starch concentration 20 g/L under optimum condi-
tions [25]. Further increase in starch concentration to 25e30 g/
L resulted in a decrease of hydrogen production to 75mmol/L.
However, it has been shown that high starch concentrations
(20e40 g/L) had no remarkable effect on hydrogen production
[23]. This could also be possibly due to the shifting of bacterial
metabolism towards the volatile fatty acid formation rather
than hydrogen formation [4].
0
20
40
60
80
100
5.5 6 6.5 7
Initi
oitcudorpnegordy
Hn
)L/lo
mm(
Acetate (mButyrate (Hydrogen
Fig. 4 e Hydrogen production and distribution of acetate, butyr
24 h on arabinose fermentation at different initial pH levels.
During starch fermentation, acetate was detected as
a major volatile fatty acid. Low amounts of butyrate produc-
tion were detected irrespective of initial culture pH. The Bu/Ac
ratio was ranging from 0.11 to 0.15 at pH below 7.5. Moreover,
it was observed that there is a tendency of increasing Bu/Ac
ratio with increasing pH from 8 to 9. In addition, small quan-
tities of iso-butyrate (<25 mg/L) and succinate and formate
(<75 mg/L) were also detected. Further, butyrate production
was slightly increased at alkaline pHs (8.0e9.0). Therefore,
results of the distribution of fatty acid data showed that
hydrogen production on starch occurred mainly through
acetate pathway, which is leading to the high hydrogen
production. However, total hydrogen production (390 ml H2/g
starch) from starch achieved in this study was lower than the
theoretical maximum 550 ml H2/g starch, which occurred
through the acetate pathway [4].
3.3.5. Hydrogen production under different concentrations ofNaClSalt is essential for cells, composed primarily of sodium
chloride. Generally the salt concentrations of natural water or
various waste waters were either fresh or marine [27]. Results
of this study showed that sodium chloride was essential for
the optimum growth and hydrogen production of the strain
C9. Addition of NaCl enhanced the hydrogen production
potential of the strain C9 on all the carbon sources tested
(xylan, xylose, arabinose and starch). Optimum NaCl
concentration for the maximum hydrogen production is
7.5 8 8.5 9
al pH
0
2
4
6
8
10
12
14
16
18
20
/lom
m(noitcudorp
sdicA
L)
mol/L) mmol/L) production (mmol/L)
ate production of Clostridium amygdalinum strain C9 during
0
40
80
120
160
5.5 6 6.5 7 7.5 8 8.5 9
Initial pH
oitcudorP
negordyH
n)
L/lom
m(
0
2
4
6
8
10
12
14
16
18
20
/lom
m(noitcudorp
sdicA
L)
Acetate (mmol/L) Butyrate (mmol/L) Hydrogen production (mmol/L)
Fig. 5 e Hydrogen production and distribution of acetate, butyrate production of Clostridium amygdalinum strain C9 during
24 h on xylose fermentation at different initial pH levels.
i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 3 6671
substrate dependant and the strain C9 showed maximum
hydrogen production on xylose and arabinose in the presence
of 10 g NaCl/L. Further increase of NaCl concentration in the
medium reduced the hydrogen production potential of the
strain (Table 1). The strain required low concentration of NaCl
(7.5 g/L) for the optimum hydrogen production on xylan and
starch as compared to that of xylose and arabinose. Since the
requirement of high NaCl concentration for the optimum H2
production, the novel strain would be very important to study
it’s potential for the treatment of marine waste.
3.3.6. Hydrogen production under microaerophilic conditionsIt was interestingly noted that the strain could produce
hydrogen under microaerophilic conditions. This was
confirmed by the methylviologen assay. The results of the
methylviolegen assay showed that when 50% of the head-
space replaced with air, the hydrogen evolution was
Fig. 6 e Hydrogen production and distribution of acetate, butyra
24 h on starch fermentation at different initial pH levels.
approximately 2500 nmol/400 mL of culture (2.2 OD at 600 nm).
However, as compared to that of control (under strict anaer-
obic condition), the strain exhibited only 30% reduction in H2
evolution. This suggests that the hydrogenase system present
in strain C9 could be oxygen tolerant. Since oxygen tolerance
hydrogenases have a great potential in future biotechnolog-
ical use, the novel strain C9 would be very important in future
hydrogen production research. Further investigations are
going on to evaluate the actual potential of the hydrogease
enzyme system to produce hydrogen under miceoaerophilic
condition in vitro.
Overall, the final pHs in all batch reactors were within the
range of pH 4.0e4.6 at the end of 24 h fermentation, regard-
less of the initial culture pH or the type of substrate used.
Hence the strain C9 showed low rate of hydrogen evolution
after 24 h during this study. Further, with low hydrogen
production, gradual increase in ethanol production was
te production of Clostridium amygdalinum strain C9 during

i n t e rn a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 36672
detected with xylose and xylan fermentation. This suggests
that microbial shift from favorable hydrogen/acid production
phase to energy-consuming ethanol production phase. This
shift may occur when the pH was further down to 4.5 or
below [24].
4. Conclusion
This study reported the hydrogen production potential of
moderately halophilic novel marine strain of C. amygdalinum
C9 isolated from sea buried crude oil pipelines. The strain was
identified based on 16S rRNA gene sequence. Hydrogen
production by strain C 9 was pH and substrate dependant.
Further, the strain favored alkaline pHs for optimum
hydrogen production of xylose (pH 7.5e8.5), xylan (pH 7.5),
arabinose (pH 8.0) and starch (pH 7.5). NaCl is essential for
optimum growth and hydrogen production. The strain C9
preferred high NaCl concentrations for optimum hydrogen
production on xylose and arabinose (10 g NaCl/L), and Xylan
and starch (7.5 g NaCl/L). Finally, it is very interestingly to note
that the strain had a great potential to produce hydrogen
under microaerophilic conditions. Therefore, the results dis-
cussed in this study could be used to assess the feasibility of
using pure bacterial strain in converting renewable energy
resources such as pentose or starch to clean H2. Hence this
study provides useful information for the design and opera-
tion of a fermentation processes using highly abundant
renewable energy resources like xylan, xylose, arabinose and
starch.
Acknowledgements
We are indebted to the R&D centre of Hindustan Petroleum
Cooperation Ltd., Mumbai and the Corporate Health Safety
Environment division of Oil and Natural Gas Corporation for
financial support of this study and also Department of
Biotechnology, Govt. of India for partial support. The
authors are thankful to Dr R. K. Pachauri, DG, TERI, New
Delhi, for providing infrastructure facility to carry out the
present study. We thank Abu Swealsh, Laboratory techni-
cian, Microbial Biotechnology Division, TERI for collection of
samples.
r e f e r e n c e s
[1] Kim D-H, Kim S-H, Shin S-H. Sodium inhibition offermentative hydrogen production. Int J Hydrogen Energy2009;34:3295e304.
[2] Yuan Z, Yang H, Zhi X, Shen J. Enhancement effect of L-cysteine on dark fermentative hydrogen production. Int JHydrogen Energy 2008;33:6535e40.
[3] Ren N-Q, Cao G-L, Guo W-Q, Wang A-J, Ahu Y-H, Liu B-F,Xu J-F. Biological hydrogen production from corn stove bymoderately thermophile Thermoanaerobacteriumthermosaccharolyticum W16. Int J Hydrogen Energy 2010;35:2708e12.
[4] Argun H, Kargi F, Kadapdan IK, Oztekin R. Batch darkfermentation of wheat starch to hydrogen gas: effects of theinitial substrate and biomass concentration. Int J HydrogenEnergy 2008;33:6109e15.
[5] KotaySM,DasD. Biohydrogenas a renewableenergy resources-prospectsandpotentials. Int JHydrogenEnergy2008;33:258e63.
[6] Mohana S, Shah A, Divecha J, Madamwar D. Xylanaseproduction by Burkholderia sp. DMAX strain under solid statefermentation using distillery spent wash. BioresourceTechnol 2008;99:7553e64.
[7] Saratale GD, Chen S-D, Lo YC, Saratale RG, Chang S J-.Outlook of biohydrogen production from lignocellulosicfeedstock using dark fermentation- a review. J Sci Ind Res2008;67:962e79.
[8] Miller TL, Wollin MJ. A serum bottle modification of theHungate technique for the cultivating obligate anaerobes.Appl Microbiol 1974;27:985e7.
[9] Sambrook J, Fritsch EF, Maniatis T. Molecular cloninig:a laboratory manual. 2nd ed. Cold Spring Harbor, NY:ColdSpring Harbor Laboratory; 1989.
[10] Altschul SF, Madden TL, Schaffer AA, Zhang Z, Miller W,LipmanDJ. GappedBLASTandPSI-BLAST: a newgeneration ofprotein database search programme. Nucl Acids Res 1997;25:3389e402.
[11] Jukes TH, Cantor CR. Evolution of protein molecules. In:Munro HN, editor. Mammalian protein metabolism. NewYork: Academic Press; 1969. p. 211e32.
[12] Felsenstein J. PHYLIP (Phylogeny Inference Package) version3.67. Seattle: Department of Genome Sciences, University ofWashington; 2007.
[13] Saitou N, Nei M. The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Mol Biol Evol1987;4:405e25.
[14] Felsenstein J. Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 1995;30:783e91.
[15] Jayasinghearachchi HS, Sarma PM, Singh S, Aginihotri A,Mandal AK, Lal B. Fermentative hydrogen production by twonovel strains of Enterobacter aerogenes HGN-2 and HT 34isolated from sea buried crude oil pipelines. Int J HydrogenEnergy 2009;34:7197e207.
[16] Taguchi F, Hasegawa K, Saito-Taki T, Hara K. Simultaneousproduction of xylanase and hydrogen using xylan in batchculture of Clostridium sp. strain X53. J Ferment Bioeng 1996;81:178e80.
[17] Danko AS, Abreu AA, Alves MM. Effect of arabinoseconcentration of dark fermentation hydrogen using differentmixed cultures. Int J Hydrogen Energy 2008;33:4527e33.
[18] Taguchi F, Mizukami N, Hasegawa K, Saito-Taki T. Microbialconversion of arabinose and xylose to hydrogen by a newlyisolated Clostridium sp. no. 2. Can J Microbiol 1994;40:228e33.
[19] Abreu AA, Danko AS, Costa JC, Ferreira EC, Alves MM.Inoculum type response to different pHs on biohydrogenproduction from L-arabinose, a component of hemicellulosicbiopolymers. Int J Hydrogen Energy 2009;34:1744e51.
[20] Lo YC, Chen WM, Hung CH, Chen SD, Chang JS. Darkfermentation from sucrose and xylose using H2-producingindigenous bacteria: feasibility and kinetic studies.Water Res 2008;42:827e49.
[21] Taguchi F, Mizukami N, Saito-Taki T, Hasegawa K. Hydrogenproduction from continuous fermentation of xylose duringgrowth of Clostridium sp. strain no. 2. Can J Microbiol 1990;41:536e40.
[22] Lin CY, Cheng CH. Fermentative hydrogen production fromxylose using anaerobic mixed microflora. Int J HydrogenEnergy 2006;31:832e40.
[23] Yang H, Shen J. Effect of ferrous iron concentration onanaerobic bio-hydrogen production from soluble starch. Int JHydrogen Energy 2006;31:2137e46.

i n t e r n a t i o n a l j o u r n a l o f h y d r o g e n en e r g y 3 5 ( 2 0 1 0 ) 6 6 6 5e6 6 7 3 6673
[24] Ren N, Li J, Li B, Wabg Y, Liu S. Biohydrogen production frommolasses by anaerobic fermentation with a pilot-scalebioreactor system. Int J Hydrogen Energy 2006;31:2147e57.
[25] Lin C-Y, Chang C-C, Hung C-H. Fermentative hydrogenproduction from starch using natural mixed cultures. Int JHydrogen Energy 2008;33:2445e53.
[26] Yokoi H, Tokushige T, Hirose J, Hayashi S, Takasaki Y. H2
production from starch by amixed culture of ClostridiumbutyricumandEnterobacteraerogenes. BiotechnolLett1998;10:43e7.
[27] Zhu D, Wang G, Qiao H, Cai J. Fermentative hydrogenproduction by new marine Pantoea agglomerans isolated fromthe mangrove sludge. Int J Hydrogen Energy 2008;33:6116e23.
Related Documents











