UNCORRECTED PROOF 2 Fatty acid control of nitric oxide production by macrophages 3 Thais Martins de Lima a, * , Larissa de Sa Lima b , Cristoforo Scavone b , Rui Curi a 4 a Department of Physiology and Biophysics, Institute of Biomedical Sciences, University of Sa ˜ o Paulo, Av. Prof Lineu Prestes, 1524, 5 05508-900, Sa ˜ o Paulo, Brazil 6 b Department of Pharmacology, Institute of Biomedical Sciences, University of Sa ˜ o Paulo, Av. Prof Lineu Prestes, 1524, 05508-900, Sa ˜ o Paulo, Brazil Received 17 April 2006; accepted 26 April 2006 Available online Edited by Sandro Sonnino 10 Abstract Modulation of macrophage functions by fatty acids 11 (FA) has been studied by several groups, but the effect of FA 12 on nitric oxide production by macrophages has been poorly 13 examined. In the present study the effect of palmitic, stearic, 14 oleic, linoleic, arachidonic, docosahexaenoic and eicosapentae- 15 noic acids on NF-jB activity and NO production in J774 cells 16 (a murine macrophage cell line) was investigated. All FA tested 17 stimulated NO production at low doses (1–10 lM) and inhibited 18 it at high doses (50–200 lM). An increase of iNOS expression 19 and activity in J774 cells treated with a low concentration of 20 FA (5 lM) was observed. The activity of NF-jB was time-depen- 21 dently enhanced by the FA treatment. The inhibitory effect of FA 22 on NO production may be due to their cytotoxicity, as observed 23 by loss of membrane integrity and/or increase of DNA fragmen- 24 tation in cells treated for 48 h with high concentrations. The re- 25 sults indicate that, at low concentrations FA increase NO 26 production by J774 cells, whereas at high concentrations they 27 cause cell death. 28 Ó 2006 Published by Elsevier B.V. on behalf of the Federation of 29 European Biochemical Societies. 30 Keywords: Fatty acid; Nitric oxide; NF-jB; iNOS expression; 31 iNOS activity; Cell death 32 33 1. Introduction 34 NO produced by activated macrophages has been shown to 35 regulate antimicrobial and antitumor activities. However, NO 36 production in excess causes tissue damage that is associated 37 with acute and chronic inflammation [1]. NO is synthesized 38 from L-arginine by NO synthase (NOS) using NADPH and 39 oxygen as cosubstrates [2]. Macrophages stimulated with lipo- 40 polysaccharide (LPS) and pro-inflammatory cytokines such as 41 interferon-c (IFN-c) and tumor necrosis factor a (TNF-a) [3,4] 42 produce large amounts of NO through inducible NOS (iNOS) 43 activity. The expression of iNOS is regulated by the nuclear 44 factor kappa B (NF-jB) in several cell types, including macro- 45 phages [5,6]. NF-jB plays an important role in controlling 46 inflammatory gene activation [7]. This transcription factor is 47 usually found in the cytosol as a heterodimer complex with 48 its inhibitory protein, IjB. When cells are stimulated with 49 LPS, phorbol esther or inflammatory cytokines, IjB is phos- 50 phorylated by IjB kinase and degraded. IjB phosphorylation 51 dissociates the dimmer and allows NF-jB to translocate to the 52 nucleus, where it activates target genes, including iNOS [8]. 53 Recent studies have shown that fatty acids (FA) can modu- 54 late NF-jB activation. In human monocytic THP-1 cells, lino- 55 leic (LA), a-linolenic (ALA) and docosahexaenoic (DHA) acids 56 decreased NF-jB DNA-binding activity [9]. Palmitic acid (PA) 57 increased NF-jB activity in 3T3-L1 adipocytes [10] and peri- 58 cytes [11]. On the other hand, palmitic (PA), oleic (OL) and lin- 59 oleic (LA) acids induced iKKb activation and decreased NF-jB 60 activity in endothelial cells [12]. Eicosapentaenoic (EPA) and 61 docosahexaenoic (DHA) acids decreased LPS induced NF-jB 62 activation in human kidney-2 (HK-2) cells [13]. 63 Several authors have shown modulation of macrophage 64 functions by FA [14–17]. The x 6 polyunsaturated FA usu- 65 ally stimulate the inflammatory response, whereas x 3 FA 66 have been considered as anti-inflammatory agents. The effect 67 of FA has been examined on cytokine, eicosanoids and reac- 68 tive oxygen species production [18–20], and adhesion molecule 69 expression [17,21]. However, the effect of FA on NO produc- 70 tion and iNOS expression in macrophages has been poorly 71 examined [22–25]. The studies have shown that x 3 FA, 72 especially DHA, markedly suppress NO production and iNOS 73 expression in murine macrophages. An increase in NO produc- 74 tion by macrophages from animals fed x 3 FA rich diets has 75 been also reported [26,27]. Up to now, however, there is no re- 76 port comparing the effect of the more abundant FA in plasma, 77 palmitic (saturated) and oleic acids (monounsaturated, x 9), 78 with x 3 (DHA and EPA) and x 6 (linoleic and arachi- 79 donic) FA. 80 As described above, NO production and iNOS expression are 81 regulated by NF-jB activation, and the activity of this tran- 82 scription factor can be modulated by FA. This information 83 led us to investigate the effect of various FA on NF-jB activity 84 and NO production in J774 cells (a murine macrophage cell 85 line). The following FA were studied: palmitic (PA), stearic 86 (SA), oleic (OA), linoleic (LA), arachidonic (AA), docosahexa- 87 enoic (DHA) and eicosapentaenoic (EPA) acids. 88 2. Materials and methods 89 2.1. Reagents 90 RPMI-1640 medium, HEPES, penicillin and streptomycin were 91 purchased from Invitrogen (Carlsbad, CA, USA). Fatty acids, LPS, 92 sulfanilamide, naphthylene diamine dihydrochloride, sodium pyro- 93 phosphate and sodium orthovanadate were obtained from Sigma (St. 94 Louis, MO, USA). Ethanol and phosphoric acid were purchased from 95 Merck (Frankfurter, Germany). Sodium bicarbonate and sodium ni- 96 trite were purchased from Labsynth products (Diadema, SP, Brazil). 97 DAF-DA was obtained from Molecular Probes (Eugene, OR, USA). * Corresponding author. Fax: +55 11 30917285. E-mail address: [email protected] (T.M. de Lima). 0014-5793/$32.00 Ó 2006 Published by Elsevier B.V. on behalf of the Federation of European Biochemical Societies. doi:10.1016/j.febslet.2006.04.091 FEBS Letters xxx (2006) xxx–xxx FEBS 30727 No. of Pages 9 6 May 2006; Disk Used ARTICLE IN PRESS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2
3
456
1011121314151617181920212223242526272829
3031
32
33
3435363738394041424344454647484950
FEBS Letters xxx (2006) xxx–xxx
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
Fatty acid control of nitric oxide production by macrophages
Thais Martins de Limaa,*, Larissa de Sa Limab, Cristoforo Scavoneb, Rui Curia
a Department of Physiology and Biophysics, Institute of Biomedical Sciences, University of Sao Paulo, Av. Prof Lineu Prestes, 1524,05508-900, Sao Paulo, Brazil
b Department of Pharmacology, Institute of Biomedical Sciences, University of Sao Paulo, Av. Prof Lineu Prestes, 1524, 05508-900, Sao Paulo, Brazil
Received 17 April 2006; accepted 26 April 2006
Available online
Edited by Sandro Sonnino
C
F
51525354555657585960616263646566676869707172
Abstract Modulation of macrophage functions by fatty acids(FA) has been studied by several groups, but the effect of FAon nitric oxide production by macrophages has been poorlyexamined. In the present study the effect of palmitic, stearic,oleic, linoleic, arachidonic, docosahexaenoic and eicosapentae-noic acids on NF-jB activity and NO production in J774 cells(a murine macrophage cell line) was investigated. All FA testedstimulated NO production at low doses (1–10 lM) and inhibitedit at high doses (50–200 lM). An increase of iNOS expressionand activity in J774 cells treated with a low concentration ofFA (5 lM) was observed. The activity of NF-jB was time-depen-dently enhanced by the FA treatment. The inhibitory effect of FAon NO production may be due to their cytotoxicity, as observedby loss of membrane integrity and/or increase of DNA fragmen-tation in cells treated for 48 h with high concentrations. The re-sults indicate that, at low concentrations FA increase NOproduction by J774 cells, whereas at high concentrations theycause cell death.� 2006 Published by Elsevier B.V. on behalf of the Federation ofEuropean Biochemical Societies.
Keywords: Fatty acid; Nitric oxide; NF-jB; iNOS expression;iNOS activity; Cell death
E 737475767778798081828384858687
88
89909192
UN
CO
RR1. Introduction
NO produced by activated macrophages has been shown to
regulate antimicrobial and antitumor activities. However, NO
production in excess causes tissue damage that is associated
with acute and chronic inflammation [1]. NO is synthesized
from LL-arginine by NO synthase (NOS) using NADPH and
oxygen as cosubstrates [2]. Macrophages stimulated with lipo-
polysaccharide (LPS) and pro-inflammatory cytokines such as
interferon-c (IFN-c) and tumor necrosis factor a (TNF-a) [3,4]
produce large amounts of NO through inducible NOS (iNOS)
activity. The expression of iNOS is regulated by the nuclear
factor kappa B (NF-jB) in several cell types, including macro-
phages [5,6]. NF-jB plays an important role in controlling
inflammatory gene activation [7]. This transcription factor is
usually found in the cytosol as a heterodimer complex with
its inhibitory protein, IjB. When cells are stimulated with
LPS, phorbol esther or inflammatory cytokines, IjB is phos-
phorylated by IjB kinase and degraded. IjB phosphorylation
*Corresponding author. Fax: +55 11 30917285.E-mail address: [email protected] (T.M. de Lima).
0014-5793/$32.00 � 2006 Published by Elsevier B.V. on behalf of the Feder
doi:10.1016/j.febslet.2006.04.091
TED
PR
OOdissociates the dimmer and allows NF-jB to translocate to the
nucleus, where it activates target genes, including iNOS [8].
Recent studies have shown that fatty acids (FA) can modu-
late NF-jB activation. In human monocytic THP-1 cells, lino-
leic (LA), a-linolenic (ALA) and docosahexaenoic (DHA) acids
decreased NF-jB DNA-binding activity [9]. Palmitic acid (PA)
increased NF-jB activity in 3T3-L1 adipocytes [10] and peri-
cytes [11]. On the other hand, palmitic (PA), oleic (OL) and lin-
oleic (LA) acids induced iKKb activation and decreased NF-jB
activity in endothelial cells [12]. Eicosapentaenoic (EPA) and
docosahexaenoic (DHA) acids decreased LPS induced NF-jB
activation in human kidney-2 (HK-2) cells [13].
Several authors have shown modulation of macrophage
functions by FA [14–17]. The x � 6 polyunsaturated FA usu-
ally stimulate the inflammatory response, whereas x � 3 FA
have been considered as anti-inflammatory agents. The effect
of FA has been examined on cytokine, eicosanoids and reac-
tive oxygen species production [18–20], and adhesion molecule
expression [17,21]. However, the effect of FA on NO produc-
tion and iNOS expression in macrophages has been poorly
examined [22–25]. The studies have shown that x � 3 FA,
especially DHA, markedly suppress NO production and iNOS
expression in murine macrophages. An increase in NO produc-
tion by macrophages from animals fed x � 3 FA rich diets has
been also reported [26,27]. Up to now, however, there is no re-
port comparing the effect of the more abundant FA in plasma,
palmitic (saturated) and oleic acids (monounsaturated, x � 9),
with x � 3 (DHA and EPA) and x � 6 (linoleic and arachi-
donic) FA.
As described above, NO production and iNOS expression are
regulated by NF-jB activation, and the activity of this tran-
scription factor can be modulated by FA. This information
led us to investigate the effect of various FA on NF-jB activity
and NO production in J774 cells (a murine macrophage cell
line). The following FA were studied: palmitic (PA), stearic
(SA), oleic (OA), linoleic (LA), arachidonic (AA), docosahexa-
enoic (DHA) and eicosapentaenoic (EPA) acids.
9394959697
2. Materials and methods
2.1. ReagentsRPMI-1640 medium, HEPES, penicillin and streptomycin were
purchased from Invitrogen (Carlsbad, CA, USA). Fatty acids, LPS,sulfanilamide, naphthylene diamine dihydrochloride, sodium pyro-phosphate and sodium orthovanadate were obtained from Sigma (St.Louis, MO, USA). Ethanol and phosphoric acid were purchased fromMerck (Frankfurter, Germany). Sodium bicarbonate and sodium ni-trite were purchased from Labsynth products (Diadema, SP, Brazil).DAF-DA was obtained from Molecular Probes (Eugene, OR, USA).
ation of European Biochemical Societies.

C
9899
100
101102103104105106107108109110111112113114
115116117118119120121122123124
125126127128129130131132133134135136137138139140141142143144145146147148149150151152
153154155156157158159160161162163164165166
167168169170171172173
174175176177178179180181182183184185186187188189190191192193194
195196197198199200201202203204205206207208209210211212213214215216217218219220221
222223224225226227228
229
230231232
2 T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
UN
CO
RR
E
Reagents for SDS–PAGE and immunoblotting were from Bio-RadLaboratories (Richmond, CA, USA). Antibodies were obtained fromSanta Cruz Biotechnology (Santa Cruz, CA, USA).
2.2. Culture conditions and fatty acid treatmentJ774 cells were grown in RPMI-1640 medium containing 10% fetal
calf serum (FCS). This medium was supplemented with glutamine(2 mM), HEPES (20 mM), streptomycin (10000 lg/mL), penicillin(10000 UI/mL) and sodium bicarbonate (24 mM). Cells were grownin 75 cm2 flasks containing 0.5–1 · 106 cells per mL. The cells werekept in a humidified atmosphere containing 5% CO2 at 37 �C.
Cells were treated with various concentrations (1–200 lM) of pal-mitic (PA), stearic (SA), oleic (OA), linoleic (LA), arachidonic (AA),eicosapentaenoic (EPA) and docosahexaenoic (DHA) acids for differ-ent periods (from 3 up to 48 h). The fatty acids were dissolved in eth-anol. The final concentration of ethanol in the culture medium did notexceed 0.5%. This concentration of ethanol is not toxic to the cells asreported by Siddiqui et al. [28].
2.3. Determination of nitric oxideThe content of nitrite was measured in the supernatant of cultured
cells based on the method described by Ding et al. [29]. Cells (5 · 105
per well) were seeded in 96 well plates and treated with 2.5 lg per mLLPS and different concentrations of FA for 48 h. At the end of the cul-ture period, 50 lL of the supernatant were removed and incubated withan equal volume of Griess reagent (1% sulfanilamide, 0.1% naphthylenediamine dihydrochloride, 2.5% H3PO4) at room temperature for10 min. The absorbance was determined at 550 nm. Nitrite concentra-tion was determined by using sodium nitrite as standard.
2.4. Determination of iNOS protein expressionJ774 cells were seeded in 25 cm flasks and treated with 2.5 lg per mL
LPS and 5 lM of the FA for 6, 12 and 24 h. At the end of the incuba-tion period, cells were immediately homogenized in 150 lL extractionbuffer (100 mM Trizma, pH 7.5; 10 mM EDTA; 10% sodium dodecylsulfate (SDS); 100 mM NaF; 10 mM sodium pyrophosphate; 10 mMsodium orthovanadate; at 100 �C) for 30 s. Samples were boiled for5 min and centrifuged at 12000 rpm, for 40 min, at 4 �C. Aliquots ofsupernatants were used for the measurement of total protein content,as described by Bradford [30]. Equal amount of protein of each sample(70 lg) was separated using 6% SDS–polyacrylamide gel. Westernblotting was carried out following the method described by Towbinet al. [31]. The proteins of the gel were transferred to a nitrocellulosemembrane at 120 V for 1 h. Non-specific bounds were blocked by incu-bating the membranes with 5% defatted milk in basal solution (10 mMTrizma, pH 7.5; 150 mM NaCl; 0.05% Tween 20) at room tempera-ture, for 2 h. Membranes were washed in basal solution three timesfor 10 min each and then incubated with anti-iNOS antibody in basalsolution containing 3% defatted milk, at room temperature, for 3 h.Membranes were washed three times for 10 min each and incubatedwith anti-IgG antibody linked to horseradish peroxidase in basal solu-tion containing 1% defatted milk, at room temperature, for 1 h. Fol-lowing another washing, membranes were incubated with substratefor peroxidase and chemiluminescence enhancer (Amersham Biosci-ences, Upsalla, SW) for 1 min and immediately exposed to X-ray filmfor 30 min. Films were then revealed in the conventional manner. Bandintensities were analysed using the ScionImage software (Scion Corpo-ration, MD, USA).
2.5. Measurement of iNOS activityiNOS activity was measured in J774 cells treated for 12, 24 and 48 h
with 2.5 lg per mL LPS and 5 lM FA. This assay is based on the bio-chemical conversion of LL-arginine to LL-citrulline by iNOS. Cells werehomogenized in ice-cold Tris–HCl buffer (20 mM Tris–HCl, 10 mMEDTA, and 10 mM EGTA, pH 7.4) using a Teflon homogenizer.The homogenates were centrifuged at 12000 · g for 5 min at 4 �C.Supernatants were removed and NOS assay was performed by incubat-ing (37 �C for 20 min) 150 lg (20 lL) of protein in a final volume of60 lL of assay mixture containing 50 mM Tris–HCl, 6 lM tetrahydro-biopterin, 2 lM FAD, 2 lM FMN, 10 mM NADPH, 100 mM LL-argi-nine/LL-[H3]-arginine (5 lCi/mL), 1 mM EDTA/EGTA. The reactionwas stopped with 1 mL of ice-cold stop buffer (50 mM HEPES and5 mM EDTA, pH 5.5) and 100 lL of cation-exchange resin (Dowex,
TED
PR
OO
F
Na+ form, equilibrated with 50 mM HEPES, pH 5.5) was added toeach reaction mixture to remove the excess of LL-[H3]-arginine. Aliquotswere collected into vials, scintillation liquid (6 mL) was added andradioactivity was quantified in a scintillation counter (Packard TRICARB 2100 TR Counters, Downers Grove, IL, USA). Protein concen-trations in samples were determined as described by Bradford [30] withbovine serum albumin as standard.
2.6. Electrophoretic mobility shift assayNFjB activation was evaluated after treatment of the cells for 3, 6,
12, 24 and 48 h with 5 lM fatty acids; concentrations in which a highproduction of NO was observed.
Nuclear extract from J774 cells was obtained as previously described[31]. Double-stranded oligonucleotides containing the NF-jB (5 0-AGTTGAGGGGACTTTCCCAGGC-3 0) consensus binding site [33]were end-labeled using T4 PNK and [c-32P]ATP (Amersham Biosci-ences). Binding reactions of the probes (30000 cpm) were performedwith 10 lg proteins from nuclear extract, at room temperature, for20 min, in 20 lL of the binding buffer consisting of 20 mM HEPES,pH 7.6, 50 mM KCl, 10% glycerol, 0.2 mM EDTA, 1 mM DTT and2 lg polydeoxyinosinic–deoxycytidylic acid (poly[dI–dC]). Competi-tive binding assays were conducted under the same conditions withthe addition of 2 pmol (100-fold molar excess) of unlabeled competitoroligonucleotides. The DNA–protein complexes were electrophoresedon 4% non-denaturing polyacrylamide gels, at 4 �C, in 45 mM Tris,45 mM borate and 1 mM EDTA buffer. The gels were dried and sub-jected to autoradiography. The blots were analysed by scanner densi-tometry (Image Master 1D�, Amersham Biosciences) and the resultsof the binding activity were expressed as arbitrary units.
2.7. Determination of fatty acid cytotoxicityFatty acid cytotoxicity was assessed by flow cytometry after treat-
ment of the cells for 48 h with high concentrations of fatty acids. Atthe end of the culture period, 0.5 mL of medium containing cells wereused to evaluate the membrane integrity. In this assay, 50 lL of a pro-pidium iodide (PI) solution (100 lg per mL in saline buffer) were addedto the cells. Propidium iodide is a highly water-soluble fluorescentcompound that cannot pass through intact membranes and is generallyexcluded from viable cells. It binds to DNA by intercalating betweenthe bases with little or no sequence preference. After 5 min incubationat room temperature, the cells were evaluated in a FACScalibur flowcytometry equipment (Becton Dickinson, CA, USA) by using the CellQuest software. Fluorescence was measured using the FL2 channel(Orange-red fluorescence – 585/42 nm). Ten thousand events were ana-lysed per experiment.
We also determined the percentage of cells with fragmented DNAafter the treatments using propidium iodide. In this assay, cells wereresuspended in a solution containing detergents that permeabilize thecells, which promptly incorporate the dye into DNA. Briefly, 0.5 mLof medium containing cells were centrifuged at 1000 · g, for 10 min,at 4 �C. The pellet was gently resuspended in 300 lL hypotonic solutioncontaining 50 lg/mL propidium iodide, 0.1% sodium citrate, and 0.1%Triton X-100. The cells were then incubated for 2 h at 4 �C. Fluores-cence was measured and analysed by flow cytometry as described above.
Both the lost of membrane integrity and/or DNA fragmentationwere considered signs of toxicity, regardless which one was first ob-served.
2.8. Statistical analysisResults are presented as means ± S.E.M. of 6–9 determinations from
2–3 experiments. Comparisons with control were performed by analy-sis of variance (2-way-ANOVA). Significant differences were analysedby the Bonferroni post-tests (Graph Pad Prism 4 – Graph Pad Soft-ware Inc., San Diego, CA, USA). The level of significance was set atP < 0.05.
3. Results
3.1. Effect of fatty acids on NO production
All FA tested stimulated NO production at low doses but
showed inhibitory effect at high concentrations (Fig. 1). The

233234235236237
238239240241242
T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx 3
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
stimulatory effect of the FA was more pronounced at concen-
trations between 2.5 and 5 lM (P < 0.001). At 5 lM, palmitic
acid was the less effective, inducing an increase of 32% in NO
production. Arachidonic acid, on the other hand, was the most
potent, increasing in 119% the production of NO. The crescent
UN
CO
RR
EC
EPA
0
2
4
6
8
10
12
14
16
0 1 2.5 5 10 25 50 100
μM
**
**
*
*
*
AA
02468
1012141618
0 1 2.5 5 10 25 50 100 150
μM
*
*
** *
**
OA
0
2
4
6
8
10
12
14
16
0 1 2.5 5 10 25 50 100 150 200
μM
*
* *
*
** *
*
PA
0
2
4
6
8
10
12
14
0 1 2.5 5 10 25 50
μM
**
* *
*
μM n
itrite
μM
nit
rite
μM
nit
rite
μM
nitr
ite
Fig. 1. Effects of fatty acids on NO production of J774 cells. The concentratreatment for 48 h with palmitic, stearic, oleic, linoleic, arachidonic, docosahLPS are shown. The supernatant was incubated with an equal volume of Grare presented as means ± S.E.M. of 16 determinations from four experiment
order of stimulatory effect at 5 lM was: PA (32%), SA (60%),
DHA (79%), LA (83%), EPA (89%), OA (94%), and AA
(117%). PA and EPA inhibited NO production at 50 lM. SA
suppressed NO production at 100 lM, AA and DHA at
150 lM, and OA and LA at 200 lM.
TED
PR
OO
FDHA
0
2
4
6
8
10
12
14
16
0 1 2.5 5 10 25 50 100 150
μM
*
***
*
*
LA
0
2
4
6
8
10
12
14
16
0 1 2.5 5 10 25 50 100 150 200
μM
*
*
*
*
***
*
SA
0
2
4
6
8
10
12
14
0 1 2.5 5 10 25 50 100
μM
*
*μM n
itrite
μM
nit
rite
μM
nit
rite
tion (lM) of nitrite measured in the supernatant of cultured cells afterexaenoic and eicosapentaenoic acids in the presence of 2.5 lg per mL
iess reagent and the absorbance was determined at 550 nm. The valuess. * P < 0.001 for comparison with control.

243244245246247248249250251252253254
2552
257258259260261
262263264265266267268269
270271272273274275276
277278279
4 T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
3.2. Effect of fatty acids on iNOS expression
In order to investigate the mechanisms involved in the regu-
lation of NO production by FA, we determined iNOS protein
expression in J774 cells treated for 6, 12 and 24 h with 5 lM
FA in the presence of LPS (2.5 lg/mL). Cells treated with
PA did not present changes in iNOS expression when com-
pared with those treated with LPS and ethanol at any period
of treatment. SA and OA treated cells presented higher iNOS
expression after 12 h (P < 0.001). Cells treated with LA, AA,
DHA and EPA showed increased iNOS expression after 6 h
of treatment (P < 0.001) and this effect remained up to 24 h
(Fig. 2).
3.3. Involvement of NFjB activation on modulation of iNOS
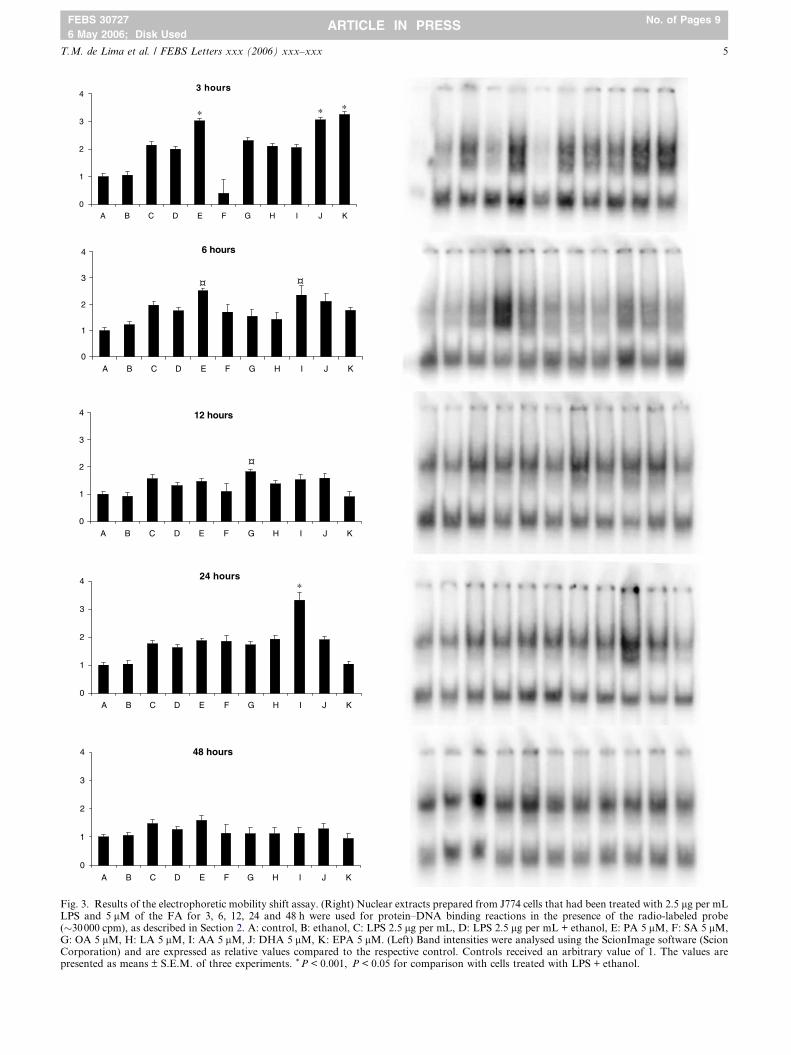
56 expression by FA
Treatment of J774 cells with 5 lM FA altered NFjB activa-
tion as observed in Fig. 3. However, the period of incubation
for the effect of FA to be observed varied considerably. LPS
caused NFjB activation and presented the more potent effect
after 3 h treatment (115%) but the addition of ethanol (vehicle)
UN
CO
RR
EC
Fig. 2. Effects of fatty acids on LPS induced iNOS expression. Cells were trWhole cell lysates were dissolved in a sample buffer and submitted to 8% Spolyclonal antibody. Band intensities were analysed using the ScionImagecompared to the respective control (LPS and ethanol). Controls received an aexperiments. * P < 0.001, # P < 0.01 for comparison with control.
OO
F
diminished NFjB activation by 16.6 ± 3.2% (means ± S.E.M.
of five experiments). PA, DHA and EPA increased NFjB acti-
vation after 3 h treatment, when compared to cells treated with
LPS and ethanol, by 51%, 52% and 62%, respectively. AA ex-
erted its stimulatory effect after 6 h of treatment (36%) and
reached its peak after 24 h (106%). OA increased NFjB activa-
tion after 12 h (39%). SA and LA did not affect NFjB activa-
tion at any period of treatment tested.
3.4. Effect of fatty acids on iNOS activity
Cells treated with AA for 12 h showed higher iNOS activity
when compared with the correspondent control (ethanol 12 h).
Exposure for 24 h to PA also led to an increased of iNOS
activity. Cells treated with OA, LA, DHA and EPA presented
higher iNOS activity after 48 h treatment. SA did not present
significant effect on iNOS activity (Fig. 4).
3.5. Effect of fatty acids on cell viability and DNA fragmentation
In order to evaluate if the inhibitory effect of the FA on NO
production at high concentrations was due to their cytotoxic-
TED
PR
eated for 6, 12 and 24 h with 2.5 lg per mL LPS and 5 lM of the FA.DS–PAGE. Western blotting was performed using mouse anti-iNOSsoftware (Scion Corporation) and are expressed as relative values
rbitrary value of 1. The values are presented as means ± S.E.M. of three
UN
CO
RR
EC
TED
PR
OO
F
* * *
¤ ¤
¤
*
3 hours
0
1
2
3
4
A B C D E F G H I J K
24 hours
0
1
2
3
4
A B C D E F G H I J K
48 hours
0
1
2
3
4
A B C D E F G H I J K
6 hours
0
1
2
3
4
A B C D E F G H I J K
12 hours
0
1
2
3
4
A B C D E F G H I J K
Fig. 3. Results of the electrophoretic mobility shift assay. (Right) Nuclear extracts prepared from J774 cells that had been treated with 2.5 lg per mLLPS and 5 lM of the FA for 3, 6, 12, 24 and 48 h were used for protein–DNA binding reactions in the presence of the radio-labeled probe(�30000 cpm), as described in Section 2. A: control, B: ethanol, C: LPS 2.5 lg per mL, D: LPS 2.5 lg per mL + ethanol, E: PA 5 lM, F: SA 5 lM,G: OA 5 lM, H: LA 5 lM, I: AA 5 lM, J: DHA 5 lM, K: EPA 5 lM. (Left) Band intensities were analysed using the ScionImage software (ScionCorporation) and are expressed as relative values compared to the respective control. Controls received an arbitrary value of 1. The values arepresented as means ± S.E.M. of three experiments. * P < 0.001, P < 0.05 for comparison with cells treated with LPS + ethanol.
T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx 5
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS

C
RO
OF
280281282283284285286287288289290291292293294295
296
297298299300301302303304305306307308309310311312313314
315316317318319320321322323324325326327328329330
pmol
.mg-1
.min
-1
0
0.05
0.1
0.15
0.2
0.25
0.3
OH+LPS PA SA OA LA AA DHA EPA
12 hours 24 hours 48 hours
¤
# #
*
*
¤
Fig. 4. Effects of fatty acids on iNOS activity. Cells were treated for 12, 24 and 48 h with 2.5 lg per mL LPS and 5 lM of the FA. iNOS activity wasdetermined after quantification of radioactivity of the LL-citrulline produced after conversion of LL-arginine to LL-citrulline by iNOS. The values arepresented as means ± S.E.M. of three experiments. * P < 0.001, # P < 0.01, P < 0.05 for comparison with the respective control (ethanol and LPS).
6 T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
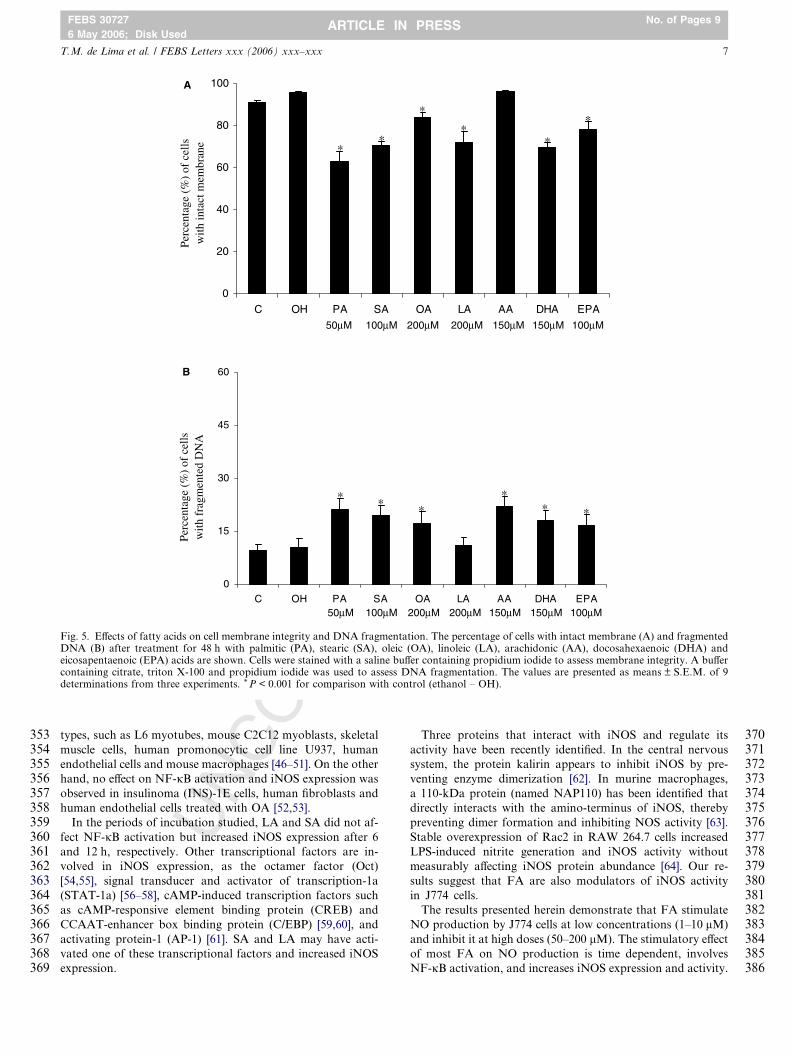
RE
ity, the percentage of cells with intact cell membrane and with
fragmented DNA after the treatment for 48 h was determined
(Fig. 5). All FA induced loss of membrane integrity, except
AA. Cells treated with PA (50 lM), SA (100 lM), OA
(200 lM), LA (200 lM), DHA (150 lM) and EPA (100 lM)
presented a significant decrease in cell membrane integrity by
34%, 26%, 12%, 24%, 27% and 18%, respectively. The treat-
ments also induced DNA fragmentation, except for LA. An in-
crease in the percentage of cells with fragmented DNA was
observed after treatment with PA (50 lM), SA (100 lM), OA
(200 lM), AA (150 lM), DHA (150 lM) and EPA (100 lM)
by 2-, 1.9-, 1.7-, 2.1-, 1.7- and 1.6-fold, respectively. Cells trea-
ted with ethanol, the vehicle used for FA preparation, did not
present loss of membrane integrity or induction of DNA frag-
mentation, indicating that the concentration of ethanol used
(0.5%) is not cytotoxic.
331332333334335336337338339340341342343344345346347348349350351352
UN
CO
R4. Discussion
The effect of different concentrations of various FA on nitric
oxide production by J774 cells and the possible mechanisms in-
volved were investigated in this study.
J774 cells cultivated for 48 h with FA and LPS showed high
production of NO as compared to control cells, especially at
low concentrations (1–10 lM). Cells treated with LA, AA,
DHA and EPA showed a stimulatory effect up to 25 lM. On
the other hand, higher concentration (50–200 lM) inhibited
NO production. These results seem controversial but they
may help to explain the discrepancy in the literature. Many
studies observed a decrease in NO production by mice macro-
phages and cell lineages after exposure to FA [22–25], whereas
others found an increase [27,34]. In addition to cell type and
period of stimulation, these controversial results may be also
due to the different concentrations of the FA used. The inhib-
itory effect of FA on NO production may be due to their cyto-
toxicity, as observed by loss of membrane integrity and/or
increase of DNA fragmentation in cells treated for 48 h with
TED
Phigh concentrations. FA toxicity has been reported in several
cell types, as a concentration and time-dependent effect of
these metabolites [35,36].
The stimulatory effect of FA was also observed by fluores-
cence microscopy using DAF-DA. Cells treated for 12 and
24 h presented high intracellular NO content, which dimin-
ished after 48 h of exposure to FA. These results corroborate
with the low level of NO in the medium of cells treated with
FA for 24 h (data not shown).
High production of NO is achieved when the expression of
inducible NOS is stimulated [3]. J774 cells treated with LPS
and FA (5 lM) presented higher expression of iNOS protein
than cells treated with LPS and ethanol (vehicle). The period
of incubations where they exerted their effects varied, and
may be related to the activity of the nuclear factor kappa B
(NF-jB). Activation of NF-jB can trigger inflammatory re-
sponses by transcriptional induction of several pro-inflamma-
tory proteins and enzymes that generate mediators of
inflammation (e.g. iNOS) [37,38]. J774 cells presented signifi-
cant NF-jB activity in basal conditions (absence of LPS and
FA) that was increased by two fold after LPS addition. The
FA tested, with exception of SA and LA, stimulated NF-jB
activation, and this effect occurred prior to or at the same incu-
bation period where an increase in iNOS protein level was ob-
served.
Several studies have shown a decrease in NF-jB activation
by DHA and EPA [39–44]. However, Maziere et al. [45] ob-
served an increase of NFjB activation in fibroblasts treated
with DHA and EPA and Camandola et al. [46] stated that
EPA (45 lM) exerts no effect on the nuclear translocation of
NF-jB in the human promonocytic cell line U937. In the pres-
ent study, DHA and EPA stimulated NO production in J774
cells at low concentrations (1–25 lM). Concomitantly, an in-
crease of NF-jB activation was observed at 5 lM, indicating
that this effect on NF-jB may raise iNOS protein level and,
consequently, NO production.
Stimulatory effect of PA, AA and OA on the nuclear trans-
location of NF-jB has also been observed in different cells
RR
EC
TED
PR
OO
F
353354355356357358359360361362363364365366367368369
370371372373374375376377378379380381382383384385386
0
20
40
60
80
100
C OH PA SA OA LA AA DHA EPA
Perc
enta
ge (
%)
of c
ells
w
ith in
tact
mem
bran
e
50μM 100μM 200μM 200μM 150μM
50μM μM100μM 200μM 100μM 200μM 150 150μM
Perc
enta
ge (
%)
of c
ells
w
ith f
ragm
ente
d D
NA
* *
*
** **
150μM 100μM
**
*
0
15
30
45
60
C OH PA SA OA LA AA DHA EPA
A
B
* *
Fig. 5. Effects of fatty acids on cell membrane integrity and DNA fragmentation. The percentage of cells with intact membrane (A) and fragmentedDNA (B) after treatment for 48 h with palmitic (PA), stearic (SA), oleic (OA), linoleic (LA), arachidonic (AA), docosahexaenoic (DHA) andeicosapentaenoic (EPA) acids are shown. Cells were stained with a saline buffer containing propidium iodide to assess membrane integrity. A buffercontaining citrate, triton X-100 and propidium iodide was used to assess DNA fragmentation. The values are presented as means ± S.E.M. of 9determinations from three experiments. * P < 0.001 for comparison with control (ethanol – OH).
T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx 7
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
UN
CO
types, such as L6 myotubes, mouse C2C12 myoblasts, skeletal
muscle cells, human promonocytic cell line U937, human
endothelial cells and mouse macrophages [46–51]. On the other
hand, no effect on NF-jB activation and iNOS expression was
observed in insulinoma (INS)-1E cells, human fibroblasts and
human endothelial cells treated with OA [52,53].
In the periods of incubation studied, LA and SA did not af-
fect NF-jB activation but increased iNOS expression after 6
and 12 h, respectively. Other transcriptional factors are in-
volved in iNOS expression, as the octamer factor (Oct)
[54,55], signal transducer and activator of transcription-1a
(STAT-1a) [56–58], cAMP-induced transcription factors such
as cAMP-responsive element binding protein (CREB) and
CCAAT-enhancer box binding protein (C/EBP) [59,60], and
activating protein-1 (AP-1) [61]. SA and LA may have acti-
vated one of these transcriptional factors and increased iNOS
expression.
Three proteins that interact with iNOS and regulate its
activity have been recently identified. In the central nervous
system, the protein kalirin appears to inhibit iNOS by pre-
venting enzyme dimerization [62]. In murine macrophages,
a 110-kDa protein (named NAP110) has been identified that
directly interacts with the amino-terminus of iNOS, thereby
preventing dimer formation and inhibiting NOS activity [63].
Stable overexpression of Rac2 in RAW 264.7 cells increased
LPS-induced nitrite generation and iNOS activity without
measurably affecting iNOS protein abundance [64]. Our re-
sults suggest that FA are also modulators of iNOS activity
in J774 cells.
The results presented herein demonstrate that FA stimulate
NO production by J774 cells at low concentrations (1–10 lM)
and inhibit it at high doses (50–200 lM). The stimulatory effect
of most FA on NO production is time dependent, involves
NF-jB activation, and increases iNOS expression and activity.

387388
389
390
391392393
394
395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430431432433434435436437438439440441442443444445446447448449450451452
453454
8 T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
The inhibitory effect of FA on NO production by J774 cells is
mainly due to their cytotoxicity.
4554564574584594604614624634644655. Uncited reference
[32].
Acknowledgements: The authors are grateful to the technical assistanceof G. de Souza, J.R. Mendonca and E.P. Portioli. This research is sup-ported by FAPESP, CNPq, and CAPES.
C
466467468469470471472473474475476477478479480481482483484485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527
UN
CO
RR
E
References
[1] MacMicking, J., Xie, Q.W. and Nathan, C. (1997) Nitric oxideand macrophage function. Annu. Rev. Immunol. 15, 323–350.
[2] Marletta, M.A., Hurshman, A.R. and Rusche, K.M. (1998)Catalysis by nitric oxide synthase. Curr. Opin. Chem. Biol. 2,656–663.
[3] Moncada, S., Palmer, R.M. and Higgs, E.A. (1991) Nitric oxide:physiology, pathophysiology, and pharmacology. Pharmacol.Rev. 43, 109–142.
[4] Nathan, C. and Xie, Q.W. (1994) Nitric oxide synthases: roles,tolls, and controls. Cell 78, 915–918.
[5] Lowenstein, C.J., Alley, E.W., Raval, P., Snowman, A.M.,Snyder, S.H., Russell, S.W. and Murphy, W.J. (1993) Macro-phage nitric oxide synthase gene: two upstream regions mediateinduction by interferon gamma and lipopolysaccharide. Proc.Natl. Acad. Sci. USA 90, 9730–9734.
[6] Xie, Q.W., Kashiwabara, Y. and Nathan, C. (1994) Role oftranscription factor NF-kappa B/Rel in induction of nitric oxidesynthase. J. Biol. Chem. 269, 4705–4708.
[7] Baldwin, A.S. (1996) The NF-kappa B and I kappa B proteins:new discoveries and insights. Annu. Rev. Immunol. 14, 649–683.
[8] Lenardo, M.J., Fan, C.M., Maniatis, T. and Baltimore, D. (1989)The involvement of NF-kappa B in beta-interferon gene regula-tion reveals its role as widely inducible mediator of signaltransduction. Cell 57, 287–294.
[9] Zhao, G., Etherton, T.D., Martin, K.R., Vanden Heuvel, J.P.,Gillies, P.J., West, S.G. and Kris-Etherton, P.M. (2005) Anti-inflammatory effects of polyunsaturated fatty acids in THP-1cells. Biochem. Biophys. Res. Commun. 336, 909–917.
[10] Ajuwon, K.M. and Spurlock, M.E. (2005) Palmitate activates theNF-kappa B transcription factor and induces IL-6 and TNFalphaexpression in 3T3-L1 adipocytes. J. Nutr. 135, 1841–1846.
[11] Cacicedo, J.M., Benjachareowong, S., Chou, E., Ruderman, N.B.and Ido, Y. (2005) Palmitate-induced apoptosis in cultured bovineretinal pericytes: roles of NAD(P)H oxidase, oxidant stress, andceramide. Diabetes 54, 1838–1845.
[12] Kim, F., Tysseling, K.A., Rice, J., Gallis, B., Haji, L., Giachelli,C.M., Raines, E.W., Corson, M.A. and Schwartz, M.W. (2005)Activation of IKKbeta by glucose is necessary and sufficient toimpair insulin signaling and nitric oxide production in endothelialcells. J. Mol. Cell. Cardiol. 39, 327–334.
[13] Li, H., Ruan, X.Z., Powis, S.H., Fernando, R., Mon, W.Y.,Wheeler, D.C., Moorhead, J.F. and Varghese, Z. (2005) EPA andDHA reduce LPS-induced inflammation responses in HK-2 cells:evidence for a PPAR-gamma-dependent mechanism. Kidney Int.67, 867–874.
[14] Kelly, D.G. (1999) Nutrition in inflammatory bowel disease.Curr. Gastroenterol. Rep. 1, 324–330.
[15] Hayashi, N., Tashiro, T., Yamamori, H., Takagi, K., Morishima,Y., Otsubo, Y., Sugiura, T., Furukawa, K., Nitta, H., Nakajima,N., Suzuki, N. and Ito, I. (1998) Effects of intravenous omega � 3and omega � 6 fat emulsion on cytokine production and delayedtype hypersensitivity in burned rats receiving total parenteralnutrition. J. Parenter. Enteral. Nutr. 22, 363–367.
[16] Tappia, P.S., Ladha, S., Clark, D.C. and Grimble, R.F. (1997)The influence of membrane fluidity, TNF receptor binding, cAMPproduction and GTPase activity on macrophage cytokine pro-duction in rats fed a variety of fat diets. Mol. Cell. Biochem. 166,135–143.
TED
PR
OO
F
[17] Yaqoob, P. (1998) Lipids and the immune response. Curr. Opin.Clin. Nutr. Metab. Care 1, 153–161.
[18] Yu, Y., Correl, P.H. and Vanden Heuvel, J.P. (2002) Conjugatedlinoleic acid decreases production of pro-inflammatory productsin macrophages: evidence for a PPARc-dependent mechanism.Biochim. Biophys. Acta 1581, 89–99.
[19] Ait-Said, F., Elalamy, I., Werts, C., Gomard, M.T., Jacquemin,C., Couetil, J.P. and Hatmi, M. (2003) Inhibition by eicosapen-taenoic acid of IL-1b-induced PGHS-2 expression in humanmicrovascular endothelial cells: involvement of lipoxygenase-derived metabolites and p38 MAPK pathway. Biochim. Biophys.Acta 1631, 77–84.
[20] Kadri-Hassani, N., Leger, C.L. and Descomps, B. (1995) Thefatty acid bimodal action on superoxide anion production byhuman adherent monocytes under phorbol 12-myristate 13-acetate or diacylglycerol activation can be explained by themodulation of protein kinase C and p47phox translocation. J.Biol. Chem. 270, 15111–15118.
[21] Cai, P., Kaphalia, B.S. and Ansari, G.A. (2005) Methyl palmitate:inhibitor of phagocytosis in primary rat Kupffer cells. Toxicology210, 197–204.
[22] Khair-el-Din, T.A., Sicher, S.C., Vazquez, M.A. and Lu, C.Y.(1996) Inhibition of macrophage nitric-oxide production and Ia-expression by docosahexaenoic acid, a constituent of fetal andneonatal serum. Am. J. Reprod. Immunol. 36, 1–10.
[23] Ohata, T., Fukuda, K., Takahashi, M., Sugimura, T. andWakabayashi, K. (1997) Suppression of nitric oxide productionin lipopolysaccharide-stimulated macrophage cells by omega 3polyunsaturated fatty acids. Jpn. J. Cancer Res. 88, 234–237.
[24] Lu, C.Y., Penfield, J.G., Khair-el-Din, T.A., Sicher, S.C., Kielar,M.L., Vazquez, M.A. and Che, L. (1998) Docosahexaenoic acid, aconstituent of fetal and neonatal serum, inhibits nitric oxideproduction by murine macrophages stimulated by IFN gammaplus LPS, or by IFN gamma plus Listeria monocytogenes. J.Reprod. Immunol. 38, 31–53.
[25] Komatsu, W., Ishihara, K., Murata, M., Saito, H. and Shinohara,K. (2003) Docosahexaenoic acid suppresses nitric oxide produc-tion and inducible nitric oxide synthase expression in interferon-gamma plus lipopolysaccharide-stimulated murine macrophagesby inhibiting the oxidative stress. Free Radic. Biol. Med. 34,1006–1016.
[26] Wallace, F.A., Neely, S.J., Miles, E.A. and Calder, P.C. (2000)Dietary fats affect macrophage-mediated cytotoxicity towardstumour cells. Immunol. Cell Biol. 78, 40–48.
[27] Bonatto, S.J., Folador, A., Aikawa, J., Yamazaki, R.K., Pizatto,N., Oliveira, H.H., Vecchi, R., Curi, R., Calder, P.C. andFernandes, L.C. (2004) Lifelong exposure to dietary fish oil altersmacrophage responses in Walker 256 tumor-bearing rats. Cell.Immunol. 231, 56–62.
[28] Siddiqui, R.A., Jenski, L.J., Neff, K., Harvey, K., Kovacs, R.J.and Stillwell, W. (2001) Docosahexaenoic acid induces apoptosisin Jurkat cells by a protein phosphatase-mediated process.Biochim. Biophys. Acta 1499, 265–275.
[29] Ding, Y., Ma, K. and Tsui, Z.C. (1998) Induction of nitricoxide production by ganglioside GM3 in murine peritonealmacrophages activated for tumor cytotoxicity. In Vivo 12,357–361.
[30] Bradford, M. (1976) A rapid and sensitive method for thequantification of microgram quantities of protein utilizingthe principle of protein-dye binding. Anal. Biochem. 72,248–254.
[31] Towbin, H., Staehelin, T. and Gordon, J. (1992) Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications 1979. Biotechnology 24,145–149.
[32] Andrews, N.C. and Faller, D. (1991) A rapid micropreparationtechnique for extraction of DNA-binding proteins from limitingnumbers of mammalian cells. Nucleic Acids Res. 19, 24991.
[33] Hang, C.H., Shi, J.X., Tian, J., Li, J.S., Wu, W. and Yin, H.X.(2004) Effect of systemic LPS injection on cortical NF-kappa Bactivity and inflammatory response following traumatic braininjury in rats. Brain Res. 1026, 23–32.
[34] Chaet, M.S., Garcia, V.F., Arya, G. and Ziegler, M.M. (1994)Dietary fish oil enhances macrophage production of nitric oxide.J. Surg. Res. 57, 65–68.

C
528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590
591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646647648649650651652653
T.M. de Lima et al. / FEBS Letters xxx (2006) xxx–xxx 9
FEBS 30727 No. of Pages 9
6 May 2006; Disk UsedARTICLE IN PRESS
NC
OR
RE
[35] Otton, R. and Curi, R. (2005) Toxicity of a mixture of fatty acidson human blood lymphocytes and leukaemia cell lines. Toxicol. InVitro 19, 749–755.
[36] Cury-Boaventura, M.F., Gorjao, R., de Lima, T.M., Newsholme,P. and Curi, R. (2006) Comparative toxicity of oleic and linoleicacid on human lymphocytes. Life Sci. 78, 1448–1456.
[37] Mercurio, F. and Manning, A.M. (1999) NF-kappa B as aprimary regulator of the stress response. Oncogene 18, 6163–6171.
[38] Yamamoto, Y. and Gaynor, R.B. (2001) Therapeutic potential ofinhibition of the NF-kappa B pathway in the treatment ofinflammation and cancer. J. Clin. Invest. 107, 135–142.
[39] Narayanan, B.A., Narayanan, N.K., Simi, B. and Reddy, B.S.(2003) Modulation of inducible nitric oxide synthase and relatedproinflammatory genes by the omega � 3 fatty acid docosahexa-enoic acid in human colon cancer cells. Cancer Res. 63, 972–979.
[40] Denys, A., Hichami, A. and Khan, N.A. (2005) n � 3 PUFAsmodulate T-cell activation via protein kinase C-alpha and -epsilonand the NF-kappa B signaling pathway. J. Lipid Res. 46, 752–758.
[41] Babcock, T.A., Helton, W.S., Hong, D. and Espat, N.J. (2002)Omega � 3 fatty acid lipid emulsion reduces LPS-stimulatedmacrophage TNF-alpha production. Surg. Infect. 3, 145–149.
[42] Babcock, T.A., Kurland, A., Helton, W.S., Rahman, A., Anwar,K.N. and Espat, N.J. (2003) Inhibition of activator protein-1transcription factor activation by omega � 3 fatty acid modula-tion of mitogen-activated protein kinase signaling kinases. J.Parenter. Enteral. Nutr. 27, 176–180.
[43] Lo, C.J., Chiu, K.C., Fu, R., Lo, R. and Helton, S. (1999) Fish oildecreases macrophage tumor necrosis factor gene transcription byaltering the NF-kappa B activity. J. Surg. Res. 82, 216–221.
[44] Theuer, J., Shagdarsuren, E., Muller, D.N., Kaergel, E., Honeck,H., Park, J.K., Fiebeler, A., Dechend, R., Haller, H., Luft, F.C.and Schunck, W.H. (2005) Inducible NOS inhibition, eicosapen-taenoic acid supplementation, and angiotensin II-induced renaldamage. Kidney Int. 67, 248–258.
[45] Maziere, C., Conte, M.A., Degonville, J., Ali, D. and Maziere,J.C. (1999) Cellular enrichment with polyunsaturated fatty acidsinduces an oxidative stress and activates the transcription factorsAP1 and NF-kappa B. Biochem. Biophys. Res. Commun. 265,116–122.
[46] Camandola, S., Leonarduzzi, G., Musso, T., Varesio, L., Carini,R., Scavazza, A., Chiarpotto, E., Baeuerle, P.A. and Poli, G.(1996) Nuclear factor jB is activated by arachidonic acid but notby eicosapentaenoic acid. Biochem. Biophys. Res. Commun. 229,643–647.
[47] Sinha, S., Perdomo, G., Brown, N.F. and O’Doherty, R.M.(2004) Fatty acid-induced insulin resistance in L6 myotubes isprevented by inhibition of activation and nuclear localization ofnuclear factor kappa B. J. Biol. Chem. 279, 41294–41301.
[48] Jove, M., Planavila, A., Sanchez, R.M., Merlos, M., Laguna, J.C.and Vazquez-Carrera, M. (2005) Palmitate induces tumor necrosisfactor-{alpha} expression in C2C12 skeletal muscle cells by amechanism involving protein kinase C and nuclear-factor {kap-pa}B activation. Endocrinology 13 (2), (Epub ahead of print).
[49] Weigert, C., Brodbeck, K., Staiger, H., Kausch, C., Machicao, F.,Haring, H.U. and Schleicher, E.D. (2004) Palmitate, but notunsaturated fatty acids, induces the expression of interleukin-6 inhuman myotubes through proteasome-dependent activation ofnuclear factor-kappa B. J. Biol. Chem. 279, 23942–23952.
[50] Park, J.Y., Kim, Y.M., Song, H.S., Park, K.Y., Kim, Y.M., Kim,M.S., Pak, Y.K., Lee, I.K., Lee, J.D., Park, S.J. and Lee, K.U.(2003) Oleic acid induces endothelin-1 expression through activa-tion of protein kinase C and NF-kappa B. Biochem. Biophys.Res. Commun. 303, 891–895.
U 654TED
PR
OO
F
[51] Choi, C.Y., You, H.J. and Jeong, H.G. (2001) Nitric oxide andtumor necrosis factor-alpha production by oleanolic acid vianuclear factor-kappa B activation in macrophages. Biochem.Biophys. Res. Commun. 288, 49–55.
[52] Kharroubi, I., Ladriere, L., Cardozo, A.K., Dogusan, Z., Cnop,M. and Eizirik, D.L. (2004) Free fatty acids and cytokines inducepancreatic beta-cell apoptosis by different mechanisms: role ofnuclear factor-kappa B and endoplasmic reticulum stress. Endo-crinology 145, 5087–5096.
[53] Hennig, B., Reiterer, G., Majkova, Z., Oesterling, E., Meerarani,P. and Toborek, M. (2005) Modification of environmentaltoxicity by nutrients: implications in atherosclerosis. Cardiovasc.Toxicol. 5, 153–160.
[54] Perrella, M.A., Pellacani, A., Wiesel, P., Chin, M.T., Foster, L.C.,Ibanez, M., Hsieh, C.M., Reeves, R., Yet, S.F. and Lee, M.E.(1999) High mobility group-I(Y) protein facilitates nuclear factor-kappa B binding and transactivation of the inducible nitric-oxidesynthase promoter/enhancer. J. Biol. Chem. 274, 9045–9052.
[55] Lee, B.S., Kim, Y.M., Kang, H.S., Kim, H.M., Pyun, K.H. andChoi, I. (2001) Octamer binding protein-1 is involved in inhibitionof inducible nitric oxide synthase expression by exogenous nitricoxide in murine liver cells. J. Biochem. 129, 77–86.
[56] Meraz, M.A., White, J.M., Sheehan, K.C., Bach, E.A., Rodig,S.J., Dighe, A.S., Kaplan, D.H., Riley, J.K., Greenlund, A.C.,Campbell, D., Carver-Moore, K., DuBois, R.N., Clark, R.,Aguet, M. and Schreiber, R.D. (1996) Targeted disruption of theStat1 gene in mice reveals unexpected physiologic specificity in theJAK-STAT signaling pathway. Cell 84, 431–442.
[57] Doi, M., Shichiri, M., Katsuyama, K., Ishimaru, S. and Hirata,Y. (2002) Cytokine-activated Jak-2 is involved in inducible nitricoxide synthase expression independent from NF-jB activation invascular smooth muscle cells. Atherosclerosis 160, 123–132.
[58] Ganster, R.W., Taylor, B.S., Shao, L. and Geller, D.A. (2001)Complex regulation of human inducible nitric oxide synthase genetranscription by Stat 1 and NF-kappa B. Proc. Natl. Acad. Sci.USA 98, 8638–8643.
[59] Eberhardt, W., Pluss, C., Hummel, R. and Pfeilschifter, J. (1998)Molecular mechanisms of inducible nitric oxide synthase geneexpression by IL-1beta and cAMP in rat mesangial cells. J.Immunol. 160, 4961–4969.
[60] Bhat, N.R., Feinstein, D.L., Shen, Q. and Bhat, A.N. (2002) p38MAPK-mediated transcriptional activation of inducible nitric-oxide synthase in glial cells. Roles of nuclear factors, nuclearfactor kappa B, cAMP response element-binding protein,CCAAT/enhancer-binding protein-beta, and activating transcrip-tion factor-2. J. Biol. Chem. 277, 29584–29592.
[61] Kizaki, T., Suzuki, K., Hitomi, Y., Iwabuchi, K., Onoe, K., Haga,S., Ishida, H., Ookawara, T., Suzuki, K. and Ohno, H. (2001)Negative regulation of LPS-stimulated expression of induciblenitric oxide synthase by AP-1 in macrophage cell line J774A.1.Biochem. Biophys. Res. Commun. 289, 1031–1038.
[62] Ratovitski, E.A., Alam, M.R., Quick, R.A., McMillan, A., Bao,C., Kozlovsky, C., Hand, T.A., Johnson, R.C., Mains, R.E.,Eipper, B.A. and Lowenstein, C.J. (1999) Kalirin inhibition ofinducible nitric-oxide synthase. J. Biol. Chem. 274, 993–999.
[63] Ratovitski, E.A., Bao, C., Quick, R.A., McMillan, A., Kozlovsky,C. and Lowenstein, C.J. (1999) An inducible nitric-oxide synthase(NOS)-associated protein inhibits NOS dimerization and activity.J. Biol. Chem. 274, 30250–30257.
[64] Kuncewicz, T., Balakrishnan, P., Snuggs, M.B. and Kone, B.C.(2001) Specific association of nitric oxide synthase-2 with Racisoforms in activated murine macrophages. Am. J. Physiol. RenalPhysiol. 281, F326–F336.
Related Documents











