Microanatomical Variation of the Nasal Capsular Cartilage in Newborn Primates TIMOTHY D. SMITH, 1,2 * JAMES B. ROSSIE, 3 GREGORY M. COOPER, 4,5 EMILY L. DURHAM, 6 ROBIN M. SCHMEIG, 2 BETH A. DOCHERTY, 6 CHRISTOPHER J. BONAR, 7 AND ANNE M. BURROWS 2,6 1 School of Physical Therapy, Slippery Rock University, Slippery Rock, Pennsylvania 2 Department of Anthropology, University of Pittsburgh, Pittsburgh, Pennsylvania 3 Department of Anthropology, Stony Brook University, Stony Brook, New York 4 Department of Surgery, University of Pittsburgh, Pittsburgh, Pennsylvania 5 Department of Oral Biology, University of Pittsburgh, Pittsburgh, Pennsylvania 6 Department of Physical Therapy, Duquesne University, Pittsburgh, Pennsylvania 7 Dallas World Aquarium, Dallas, Texas ABSTRACT The breakdown of nasal capsule cartilage precedes secondary pneu- matic expansion of the paranasal sinuses. Recent work indicates the nasal capsule of monkeys undergoes different ontogenetic transformations regionally (i.e., ossification, persistence as cartilage, or resorption). This study assesses nasal capsule morphology at the perinatal age in a taxo- nomically broad sample of non-human primates. Using traditional histo- chemical methods, osteopontin immunohistochemistry and tartrate- resistant acid phosphatase procedure, the cartilage of the lateral nasal wall (LNC) was studied. At birth, matrix properties differ between portions of the LNC that ultimately form elements of the ethmoid bone and regions of the LNC that have no postnatal (descendant) structure. The extent of carti- lage that remains in the paranasal parts of the LNC varies among species. It is fragmented in species with the greatest extent of maxillary and/or frontal pneumatic expansion. Conversely, greater continuity of the LNC is noted in newborns of species that lack maxillary and/or frontal sinuses as adults. Chondroclasts occur adjacent to elements of the ethmoid bone, along the margin of the nasal tectum, and/or along islands of cartilage that bear no signs of ossification. Chondroclasts are prevalent along remnants of the paranasal LNC in tamarin species (Leontopithecus, Saguinus), which have extensive frontal and maxillary bone pneumatization. Taken together, the morphological observations indicate that the localized loss of cartilage might be considered a critical event at the onset of secondary pneumatization, facilitated by rapid recruitment of chondro-/osteoclasts, possibly occurring simultaneously in cartilage and bone. Anat Rec, 295:950–960, 2012. V C 2012 Wiley Periodicals, Inc. Key words: craniofacial; development; growth; paranasal sinus; pars anterior; pars intermedia INTRODUCTION A firm understanding of the ontogeny of the lateral wall of the nasal capsule (LNC, also termed the paries nasi) elucidates two different phases of paranasal sinus development. Primary pneumatization is the process by which paranasal spaces extend outward from the nasal fossa as recesses (Sperber, 2000). Recesses (the primor- dial sinuses) develop into ‘‘true’’ sinuses by undergoing Grant sponsor: National Science Foundation; Grant number: BCS-0820751. *Correspondence to: Timothy D. Smith, Ph.D., School of Physical Therapy, Slippery Rock University, Slippery Rock, PA 16057. Fax: 724-738-2113. E-mail: [email protected] Received 12 December 2011; Accepted 28 February 2012. DOI 10.1002/ar.22448 Published online 27 March 2012 in Wiley Online Library (wileyonlinelibrary.com). THE ANATOMICAL RECORD 295:950–960 (2012) V V C 2012 WILEY PERIODICALS, INC.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Microanatomical Variation of the NasalCapsular Cartilage in Newborn Primates
TIMOTHY D. SMITH,1,2* JAMES B. ROSSIE,3 GREGORY M. COOPER,4,5
EMILY L. DURHAM,6 ROBIN M. SCHMEIG,2 BETH A. DOCHERTY,6
CHRISTOPHER J. BONAR,7 AND ANNE M. BURROWS2,6
1School of Physical Therapy, Slippery Rock University, Slippery Rock, Pennsylvania2Department of Anthropology, University of Pittsburgh, Pittsburgh, Pennsylvania3Department of Anthropology, Stony Brook University, Stony Brook, New York4Department of Surgery, University of Pittsburgh, Pittsburgh, Pennsylvania
5Department of Oral Biology, University of Pittsburgh, Pittsburgh, Pennsylvania6Department of Physical Therapy, Duquesne University, Pittsburgh, Pennsylvania
7Dallas World Aquarium, Dallas, Texas
ABSTRACTThe breakdown of nasal capsule cartilage precedes secondary pneu-
matic expansion of the paranasal sinuses. Recent work indicates the nasalcapsule of monkeys undergoes different ontogenetic transformationsregionally (i.e., ossification, persistence as cartilage, or resorption). Thisstudy assesses nasal capsule morphology at the perinatal age in a taxo-nomically broad sample of non-human primates. Using traditional histo-chemical methods, osteopontin immunohistochemistry and tartrate-resistant acid phosphatase procedure, the cartilage of the lateral nasal wall(LNC) was studied. At birth, matrix properties differ between portions ofthe LNC that ultimately form elements of the ethmoid bone and regions ofthe LNC that have no postnatal (descendant) structure. The extent of carti-lage that remains in the paranasal parts of the LNC varies among species.It is fragmented in species with the greatest extent of maxillary and/orfrontal pneumatic expansion. Conversely, greater continuity of the LNC isnoted in newborns of species that lack maxillary and/or frontal sinuses asadults. Chondroclasts occur adjacent to elements of the ethmoid bone, alongthe margin of the nasal tectum, and/or along islands of cartilage that bearno signs of ossification. Chondroclasts are prevalent along remnants of theparanasal LNC in tamarin species (Leontopithecus, Saguinus), which haveextensive frontal and maxillary bone pneumatization. Taken together,the morphological observations indicate that the localized loss ofcartilage might be considered a critical event at the onset of secondarypneumatization, facilitated by rapid recruitment of chondro-/osteoclasts,possibly occurring simultaneously in cartilage and bone. Anat Rec,295:950–960, 2012. VC 2012 Wiley Periodicals, Inc.
Key words: craniofacial; development; growth; paranasalsinus; pars anterior; pars intermedia
INTRODUCTION
A firm understanding of the ontogeny of the lateralwall of the nasal capsule (LNC, also termed the pariesnasi) elucidates two different phases of paranasal sinusdevelopment. Primary pneumatization is the process bywhich paranasal spaces extend outward from the nasalfossa as recesses (Sperber, 2000). Recesses (the primor-dial sinuses) develop into ‘‘true’’ sinuses by undergoing
Grant sponsor: National Science Foundation; Grant number:BCS-0820751.
*Correspondence to: Timothy D. Smith, Ph.D., School ofPhysical Therapy, Slippery Rock University, Slippery Rock, PA16057. Fax: 724-738-2113. E-mail: [email protected]
Received 12 December 2011; Accepted 28 February 2012.
DOI 10.1002/ar.22448Published online 27 March 2012 in Wiley Online Library(wileyonlinelibrary.com).
THE ANATOMICAL RECORD 295:950–960 (2012)
VVC 2012 WILEY PERIODICALS, INC.

secondary pneumatization, or expansion beyond the bor-ders of the nasal capsule, into facial bones (Weiglein,1999; Maier, 2000; Rossie, 2006; Smith et al., 2011).Using osteological evidence alone, secondary pneumati-zation can be inferred using cross-age series ofmammals (e.g., Rossie, 2006). Paranasal sinuses form byactively excavating bone along paranasal recesses(Witmer, 1999; Smith et al., 2005, 2011). More direct evi-dence is seen histologically. For example, breakdown ofnasal capsule cartilage that delimits a paranasal recessis considered a necessary prelude to secondary pneuma-tization (Wang et al., 1994; Maier, 2000; Rossie, 2006;Smith et al., 2008). In a previous report, Smith et al.(2008) documented the fate of the nasal capsule cartilagein perinatal tamarins (Saguinus geoffroyi). In newbornS. geoffroyi, the nasal capsule is partially fragmented(anteriorly) while the remainder is in various stages ofendochondral replacement by bone (posteriorly). Exami-nation of an age series of tamarins indicates that thebreakdown or replacement of nasal capsular cartilage isa relatively rapid process, extending from late fetal toearly infant stages (Smith et al., 2008).
To better understand pneumatization as a growthmechanism, longitudinal radiographic analyses (e.g.,Shah et al., 2003) and mathematical modeling (e.g., Zol-likofer and Weissman, 2008) have been used. Usingmicroanatomical methods, some authors have sought tounderstand cellular attributes of sinus mucosa (Satoet al., 1998; Wojtowicz et al., 2002) or the wall of sinusesand recesses (Smith et al., 2005, 2008, 2010, 2011). Inthis study, early (perinatal) spatial relationships andmorphology of capsular cartilage is investigated across abroad array of primate species. Variations among speciesin our sample create natural experiments for elucidatinggrowth processes. For example, our previous reportshave shown that the perinatal time period is a dynamicphase of paranasal sinus formation in at least somemonkeys, with some species undergoing secondary pneu-matic expansion of the maxillary sinus (Smith et al.,2005, 2008, 2010, 2011). Moreover, different sinusesdevelop on different ontogenetic schedules in monkeys(Rossie, 2006; Smith et al., 2008). Specifically, it is possi-ble to study one sinus during secondary pneumatization(the maxillary sinus) and another prior to secondarypneumatization (the frontal recess) in the same species(Smith et al., 2008). Finally, numerous species of prima-tes fail to form one or both of these sinuses in adults(Rossie, 2006; Tuckmantel et al., 2009; Smith et al.,2010). This remarkable diversity is used as a basis forthis study, where the morphology, matrix characteristics,and the role of chondro-/osteoclastic activity in the onsetof secondary pneumatization are studied.
Based on our earlier observation that different regionsof the nasal capsule of S. geoffroyi have different devel-opmental fates (ossification, persistence as cartilage, orapparent resorption) (Smith et al., 2008), here we assessnasal capsule morphology at the perinatal age in a taxo-nomically broad sample of non-human primates. Inaddition to traditional histochemical methods, we useosteopontin (OPN) immunohistochemistry and the tar-trate-resistant acid phosphatase (TRAP) procedure toprovide additional details about the nasal capsule. OPNhas been implicated in bone remodeling as well asresorption of mineralized matrix in the epiphyseal plate(Reinholt et al., 1990; Sugiyama et al., 2001; Weizmann
et al., 2005). OPN is also expressed in nasal capsule car-tilage, particularly in elements of the ethmoid bone(Smith et al., 2008). The exact function of TRAP isunknown, but it may be vital to chondro-/osteoclast func-tion (Hayman et al., 1996; Halleen et al., 1999). The TRAPprocedure identifies chondro-/osteoclasts and theirprecursors (Colnot and Helms, 2001; Yeh and Popowics,2011). The aims of this study are to: 1) characterize thecartilage matrix and chrondrocytes in the anterior nasalcapsule and paranasal regions and 2) establish the pres-ence or absence of cells that resorb bone and cartilage(osteoclasts or chondroclasts) adjacent to the nasalcapsule.
MATERIALS AND METHODS
Sample
Twenty-seven cadaveric primates were studied(Table 1). All specimens were available as a result of natu-ral deaths from captive populations in zoos or researchcenters. Most specimens used in this study were fixedin 10% buffered formalin after death. Some were frozenand subsequently fixed in formalin. The sample includeda diverse range, seven species of anthropoids and eightspecies of strepsirrhines (including lemurs and bushba-bies). Four of the species of monkeys were available aslarger samples (Table 1), collected specifically for the studyof early stages of pneumatization (Smith et al., 2010,2011). All specimens had been prepared for histologicalobservations in previous studies (e.g., Smith et al., 2003,2005, 2007). For this study, archived, unstained sections ofthese specimens were used. All specimens had a complete,undamaged nasal capsule on one or both sides.
Histological Preparation of Specimens
Most aspects of specimen preparation were previouslydescribed (Smith et al., 2005, 2007, 2011). Briefly, allspecimens were decalcified using a sodium citrate–for-mic acid solution and processed for paraffin embedding.Each paraffin block was serially sectioned at 10–12 lm.Selected sections were mounted on slides for stainingwith hematoxylin and eosin or other procedures for gen-eral micromorphology. Selected sections were mounted
TABLE 1. Sample and species
Species n
AnthropoidsCallithrix jacchus (common marmoset) 4Cebuella pygmaea (pygmy marmoset) 1Leontopithecus rosalia (lion tamarin) 4Saguinus geoffroyi (Geoffroy’s tamarin) 1Saguinus oedipus (cotton-top tamarin) 3Pithecia pithecia (saki monkey) 1Saimiri boliviensis (squirrel monkey) 4
StrepsirrhinesEulemur macaco (black lemur) 1Eulemur mongoz (mongoose lemur) 1Lemur catta (ringtail lemur) 2Propithecus verrauxi (Verraux’s sifaka) 1Microcebus murinus (gray mouse lemur) 1Mirza coquereli (Coquerel’s dwarf lemur) 1Galago moholi (lesser bushbaby) 1Otolemur crassicaudatus (greater bushbaby) 1
MICROANATOMICAL VARIATION OF THE NASAL CAPSULAR CARTILAGE 951
on slides or archived (the paraffin section partly pressedto paper and carefully boxed) as a means to save them,unstained, for alternate procedures. In this study,unstained sections were retrieved and used for threeprocedures. First, sections at selected anteroposteriorcross-section levels were stained using an alcian blueprocedure, which is used to the stain proteoglycans indeveloping cartilage, including matrix of the epiphysealplate (Roach, 1997; Hamrick, 1999).
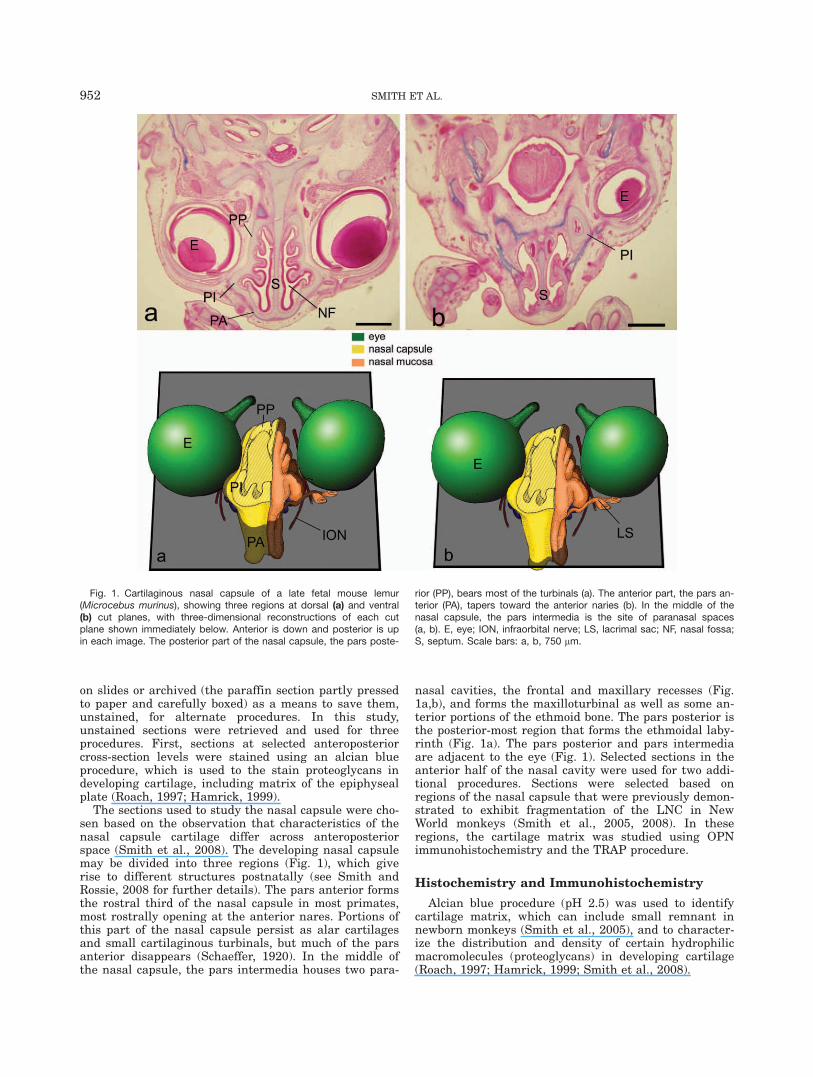
The sections used to study the nasal capsule were cho-sen based on the observation that characteristics of thenasal capsule cartilage differ across anteroposteriorspace (Smith et al., 2008). The developing nasal capsulemay be divided into three regions (Fig. 1), which giverise to different structures postnatally (see Smith andRossie, 2008 for further details). The pars anterior formsthe rostral third of the nasal capsule in most primates,most rostrally opening at the anterior nares. Portions ofthis part of the nasal capsule persist as alar cartilagesand small cartilaginous turbinals, but much of the parsanterior disappears (Schaeffer, 1920). In the middle ofthe nasal capsule, the pars intermedia houses two para-
nasal cavities, the frontal and maxillary recesses (Fig.1a,b), and forms the maxilloturbinal as well as some an-terior portions of the ethmoid bone. The pars posterior isthe posterior-most region that forms the ethmoidal laby-rinth (Fig. 1a). The pars posterior and pars intermediaare adjacent to the eye (Fig. 1). Selected sections in theanterior half of the nasal cavity were used for two addi-tional procedures. Sections were selected based onregions of the nasal capsule that were previously demon-strated to exhibit fragmentation of the LNC in NewWorld monkeys (Smith et al., 2005, 2008). In theseregions, the cartilage matrix was studied using OPNimmunohistochemistry and the TRAP procedure.
Histochemistry and Immunohistochemistry
Alcian blue procedure (pH 2.5) was used to identifycartilage matrix, which can include small remnant innewborn monkeys (Smith et al., 2005), and to character-ize the distribution and density of certain hydrophilicmacromolecules (proteoglycans) in developing cartilage(Roach, 1997; Hamrick, 1999; Smith et al., 2008).
Fig. 1. Cartilaginous nasal capsule of a late fetal mouse lemur(Microcebus murinus), showing three regions at dorsal (a) and ventral(b) cut planes, with three-dimensional reconstructions of each cutplane shown immediately below. Anterior is down and posterior is upin each image. The posterior part of the nasal capsule, the pars poste-
rior (PP), bears most of the turbinals (a). The anterior part, the pars an-terior (PA), tapers toward the anterior naries (b). In the middle of thenasal capsule, the pars intermedia is the site of paranasal spaces(a, b). E, eye; ION, infraorbital nerve; LS, lacrimal sac; NF, nasal fossa;S, septum. Scale bars: a, b, 750 lm.
952 SMITH ET AL.

For OPN immunohistochemistry, selected sectionsfrom the anterior and middle nasal capsule regions wereobtained from Callithrix jacchus, Leontopithecus rosalia,Saguinus oedipus, and Saimiri boliviensis. Two lemurs(Eulemur spp.) were studied for comparison. The assaywas conducted according to Vector protocols (and seeSmith et al., 2005). Briefly, labeling of OPN wasachieved by incubating slides with OPN antibody (SantaCruz Biotech, CA) at a 1:250 dilution in 2% donkeyserum for 30 min at RT, with a subsequent incubationwith donkey anti-goat biotinylated secondary antibody(1:250 in phosphate buffered saline) for 30 min at RT. AVector ABC kit (Vector Laboratories, CA) was used tostain immunoreactive structures, using diaminobenzi-dine (DAB) as chromogen. Following treatment withDAB, slides were counterstained with Harris’ hematoxy-lin (Surgipath, IL). OPN staining was consideredpositive if staining was more intense than that found incomparable tissue on negative control slides that wereincubated in the absence of primary antibody.
TRAP assays were conducted on selected sections ofthe four monkey species studied above, according toSigma protocol. Slides were deparaffinized and rehy-drated through serial alcohols for TRAP staining (acidphosphatase leukocyte TRAP assay, Sigma kit 387A).Slides were then refixed in 3.7% formaldehyde for 3 minat RT. Slides were rinsed for 2 min with dH2O thenstained with diazotized fast garnet GBC solution (45 mLdH2O, 0.5 mL Fast Garnet GBC base solution, 0.5 mLsodium nitrile solution, 0.5 mL naphthol AS-BI phos-phate solution, 2 mL acetate solution, and 1 mL tartratesolution for 1 hr at 37�C in the dark). Following TRAPstaining, slides were rinsed with dH2O and counter-stained with Gill’s hematoxylin followed by an 8-minwash with running tap water. TRAP staining was con-sidered positive if staining in cells with a reddish-browncolor and verified by comparison to similar cells in slideswhere hematoxylin was used but the TRAP procedurewas omitted. Adjacent sections were used when possible,allowing comparison of the same cells as prepared withand without the TRAP procedure.
RESULTS
General Observations
At birth, the nasal capsule is fragmentary laterally inall species (Figs. 2, 3). More centralized projections, theturbinals, remain at least partially cartilaginous(Fig. 2a,b). Laterally, however, there are gaps in the parsanterior and pars intermedia. Alcian blue staining inten-sity varies regionally, in many cases being more intensein the septal cartilage than the LNC (e.g., Figs. 2a,c, 3d,f).
The inferior scroll of the pars intermedia that housesthe maxillary recess (Figs. 1b, 2a, 3a,c,f) is mostly orcompletely absent, as a cartilaginous structure, in allprimates at birth. In strepsirrhines, the only cartilagi-nous remnant of this scroll is its medial projection, themaxilloturbinal; even this portion is mostly ossified inmost lemurs and all bushbabies. In some smaller strep-sirrhines (cheirogaleids), some cartilage of the inferiorpart of the pars intermedia persists at the base of themaxillary recess. In anthropoids, the pars intermedia isabsent in the region that borders the maxillary recesswith two exceptions. First, the root of the maxilloturbi-nal, which is ossified in most of the monkeys, but still
cartilaginous in tamarins, forms the lateral margin ofthe maxillary recess (see also Smith et al., 2008). Sec-ond, along the superior and lateral border of themaxillary recess, remnants of the pars intermedia areseen in some tamarins (Saguinus spp.) and S. boliviensis.Remnants are largest in S. boliviensis (Fig. 3a), where itsurrounds the inferolateral sides of the recess in somespecimens. The matrix of this remnant is lightly stainedwith alcian blue and is irregular in contour (Fig. 3a).
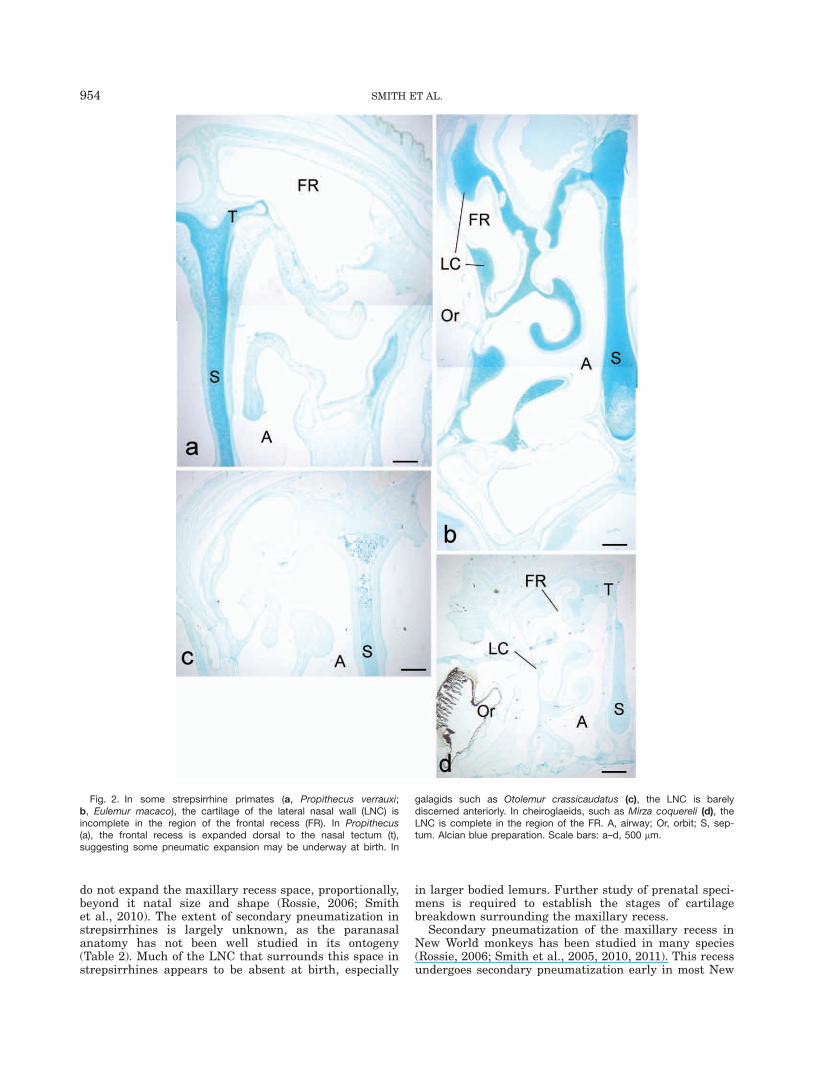
The region of the frontal recess is nearly or completelydevoid of cartilage in some strepsirrhines, such as Propi-thecus verrauxi (Fig. 2a) and all the galagids studied(e.g., Fig. 2c). In Eulemur spp. (Fig. 2b) and Lemurcatta, this recess is bordered medially by cartilage, whilethe capsular cartilage extends most laterally across therecess in cheirogaleids (Fig. 2d). The frontal recess issmall at birth in the newborn monkeys studied. In itsnascent region, near the middle meatus, the extent ofcartilage is variable. In tamarins, the tectum in limitedand the cartilage lateral to it is fragmented (Fig. 3b,c).In Pithecia pithecia (Fig. 3d), C. jacchus (Fig. 3e), andS. boliviensis (Fig. 3f), the nasal tectum is more exten-sive and continuous with cartilage along the LNC.
Immunohistochemistry and TRAP Procedure
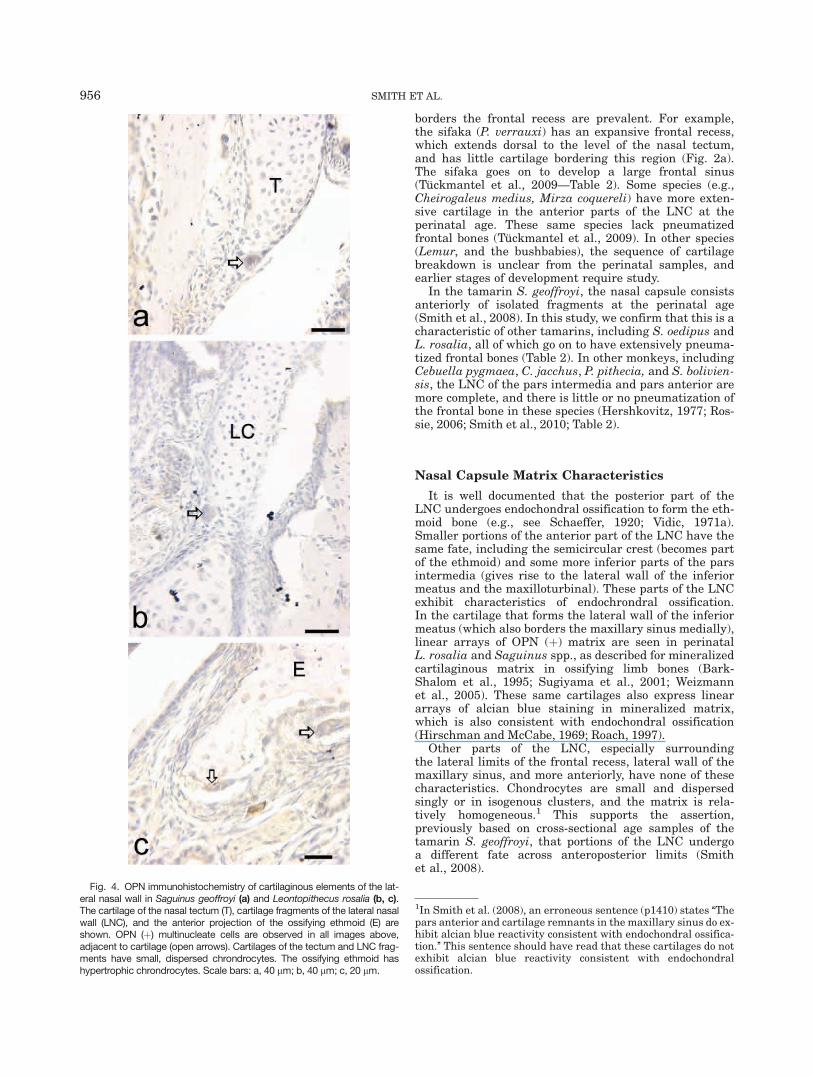
In the specimens studied immunohistochemically, thecartilage of the pars anterior and nasal tectum is OPN(�), with small chondrocytes (Fig. 4a–c). In tamarins,OPN (þ) multinucleate cells are observed near the nasaltectum and pars anterior. These cells are closely associ-ated with bone adjacent to the cartilage (Fig. 4a) but arealso observed adjacent to or between small islands ofcartilage (Fig. 4b,c) or at the margins of the nasal tec-tum (Figs. 4c, 5). Multinucleate cells at or near themargins of the nasal tectum are TRAP (þ) (Fig. 5).
In tamarins, the unossified tip of the maxilloturbinalis OPN (�) (Fig. 6a). The root of the maxilloturbinal, ad-jacent to the medial margin of the maxillary sinus, isossified in all primates studied except tamarins. In tam-arins, the maxilloturbinal root is cartilaginous. Thiscartilage is OPN (þ) (Fig. 6b) and has hypertrophicchondrocytes and TRAP (þ) cells (Fig. 7).
DISCUSSION
Several authors have suggested that local breakdown ofnasal capsule cartilage is an important event that allowsthe growth of sinuses to occur more rapidly by secondarypneumatization (Wang et al., 1994; Maier, 2000; Smithet al., 2008). The results of this study provide support forthis assertion, as those species that form maxillary orfrontal sinuses have extensive reductions or local gaps inthe LNC at sites that are documented to undergo pneuma-tization. In this study, we have documented dynamicaspects of nasal capsule morphology in primates that maybe significant to pneumatization patterns.
Nasal Capsule Morphology
Maxillary recess
Although a maxillary paranasal space is nearly ubiq-uitous in mammals, secondary pneumatization from theprimordial sinus space (i.e., the maxillary recess) is not(see Rossie, 2006; Smith et al., 2010, 2011). Some species
MICROANATOMICAL VARIATION OF THE NASAL CAPSULAR CARTILAGE 953
do not expand the maxillary recess space, proportionally,beyond it natal size and shape (Rossie, 2006; Smithet al., 2010). The extent of secondary pneumatization instrepsirrhines is largely unknown, as the paranasalanatomy has not been well studied in its ontogeny(Table 2). Much of the LNC that surrounds this space instrepsirrhines appears to be absent at birth, especially
in larger bodied lemurs. Further study of prenatal speci-mens is required to establish the stages of cartilagebreakdown surrounding the maxillary recess.
Secondary pneumatization of the maxillary recess inNew World monkeys has been studied in many species(Rossie, 2006; Smith et al., 2005, 2010, 2011). This recessundergoes secondary pneumatization early in most New
Fig. 2. In some strepsirrhine primates (a, Propithecus verrauxi;b, Eulemur macaco), the cartilage of the lateral nasal wall (LNC) isincomplete in the region of the frontal recess (FR). In Propithecus(a), the frontal recess is expanded dorsal to the nasal tectum (t),suggesting some pneumatic expansion may be underway at birth. In
galagids such as Otolemur crassicaudatus (c), the LNC is barelydiscerned anteriorly. In cheiroglaeids, such as Mirza coquereli (d), theLNC is complete in the region of the FR. A, airway; Or, orbit; S, sep-tum. Alcian blue preparation. Scale bars: a–d, 500 lm.
954 SMITH ET AL.
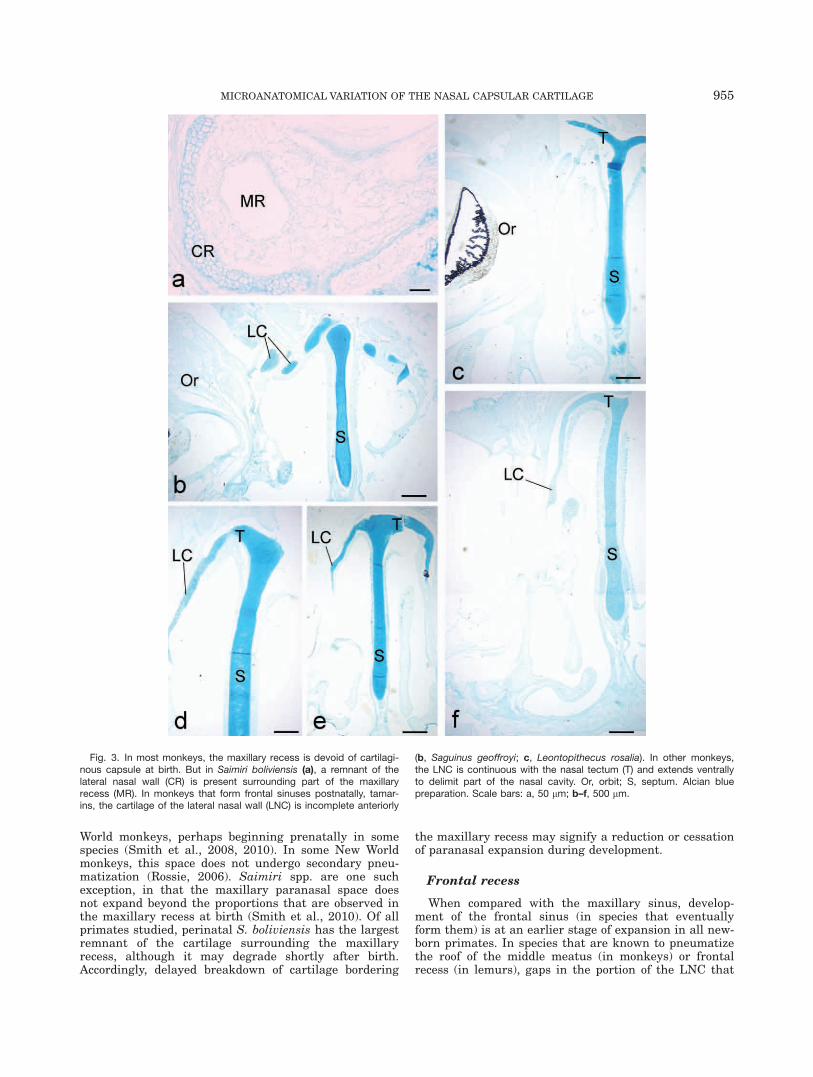
World monkeys, perhaps beginning prenatally in somespecies (Smith et al., 2008, 2010). In some New Worldmonkeys, this space does not undergo secondary pneu-matization (Rossie, 2006). Saimiri spp. are one suchexception, in that the maxillary paranasal space doesnot expand beyond the proportions that are observed inthe maxillary recess at birth (Smith et al., 2010). Of allprimates studied, perinatal S. boliviensis has the largestremnant of the cartilage surrounding the maxillaryrecess, although it may degrade shortly after birth.Accordingly, delayed breakdown of cartilage bordering
the maxillary recess may signify a reduction or cessationof paranasal expansion during development.
Frontal recess
When compared with the maxillary sinus, develop-ment of the frontal sinus (in species that eventuallyform them) is at an earlier stage of expansion in all new-born primates. In species that are known to pneumatizethe roof of the middle meatus (in monkeys) or frontalrecess (in lemurs), gaps in the portion of the LNC that
Fig. 3. In most monkeys, the maxillary recess is devoid of cartilagi-nous capsule at birth. But in Saimiri boliviensis (a), a remnant of thelateral nasal wall (CR) is present surrounding part of the maxillaryrecess (MR). In monkeys that form frontal sinuses postnatally, tamar-ins, the cartilage of the lateral nasal wall (LNC) is incomplete anteriorly
(b, Saguinus geoffroyi; c, Leontopithecus rosalia). In other monkeys,the LNC is continuous with the nasal tectum (T) and extends ventrallyto delimit part of the nasal cavity. Or, orbit; S, septum. Alcian bluepreparation. Scale bars: a, 50 lm; b–f, 500 lm.
MICROANATOMICAL VARIATION OF THE NASAL CAPSULAR CARTILAGE 955
borders the frontal recess are prevalent. For example,the sifaka (P. verrauxi) has an expansive frontal recess,which extends dorsal to the level of the nasal tectum,and has little cartilage bordering this region (Fig. 2a).The sifaka goes on to develop a large frontal sinus(Tuckmantel et al., 2009—Table 2). Some species (e.g.,Cheirogaleus medius, Mirza coquereli) have more exten-sive cartilage in the anterior parts of the LNC at theperinatal age. These same species lack pneumatizedfrontal bones (Tuckmantel et al., 2009). In other species(Lemur, and the bushbabies), the sequence of cartilagebreakdown is unclear from the perinatal samples, andearlier stages of development require study.
In the tamarin S. geoffroyi, the nasal capsule consistsanteriorly of isolated fragments at the perinatal age(Smith et al., 2008). In this study, we confirm that this is acharacteristic of other tamarins, including S. oedipus andL. rosalia, all of which go on to have extensively pneuma-tized frontal bones (Table 2). In other monkeys, includingCebuella pygmaea, C. jacchus, P. pithecia, and S. bolivien-sis, the LNC of the pars intermedia and pars anterior aremore complete, and there is little or no pneumatization ofthe frontal bone in these species (Hershkovitz, 1977; Ros-sie, 2006; Smith et al., 2010; Table 2).
Nasal Capsule Matrix Characteristics
It is well documented that the posterior part of theLNC undergoes endochondral ossification to form the eth-moid bone (e.g., see Schaeffer, 1920; Vidic, 1971a).Smaller portions of the anterior part of the LNC have thesame fate, including the semicircular crest (becomes partof the ethmoid) and some more inferior parts of the parsintermedia (gives rise to the lateral wall of the inferiormeatus and the maxilloturbinal). These parts of the LNCexhibit characteristics of endochrondral ossification.In the cartilage that forms the lateral wall of the inferiormeatus (which also borders the maxillary sinus medially),linear arrays of OPN (þ) matrix are seen in perinatalL. rosalia and Saguinus spp., as described for mineralizedcartilaginous matrix in ossifying limb bones (Bark-Shalom et al., 1995; Sugiyama et al., 2001; Weizmannet al., 2005). These same cartilages also express lineararrays of alcian blue staining in mineralized matrix,which is also consistent with endochondral ossification(Hirschman and McCabe, 1969; Roach, 1997).
Other parts of the LNC, especially surroundingthe lateral limits of the frontal recess, lateral wall of themaxillary sinus, and more anteriorly, have none of thesecharacteristics. Chondrocytes are small and dispersedsingly or in isogenous clusters, and the matrix is rela-tively homogeneous.1 This supports the assertion,previously based on cross-sectional age samples of thetamarin S. geoffroyi, that portions of the LNC undergoa different fate across anteroposterior limits (Smithet al., 2008).
Fig. 4. OPN immunohistochemistry of cartilaginous elements of the lat-eral nasal wall in Saguinus geoffroyi (a) and Leontopithecus rosalia (b, c).The cartilage of the nasal tectum (T), cartilage fragments of the lateral nasalwall (LNC), and the anterior projection of the ossifying ethmoid (E) areshown. OPN (þ) multinucleate cells are observed in all images above,adjacent to cartilage (open arrows). Cartilages of the tectum and LNC frag-ments have small, dispersed chrondrocytes. The ossifying ethmoid hashypertrophic chrondrocytes. Scale bars: a, 40 lm; b, 40 lm; c, 20 lm.
1In Smith et al. (2008), an erroneous sentence (p1410) states ‘‘Thepars anterior and cartilage remnants in the maxillary sinus do ex-hibit alcian blue reactivity consistent with endochondral ossifica-tion.’’ This sentence should have read that these cartilages do notexhibit alcian blue reactivity consistent with endochondralossification.
956 SMITH ET AL.
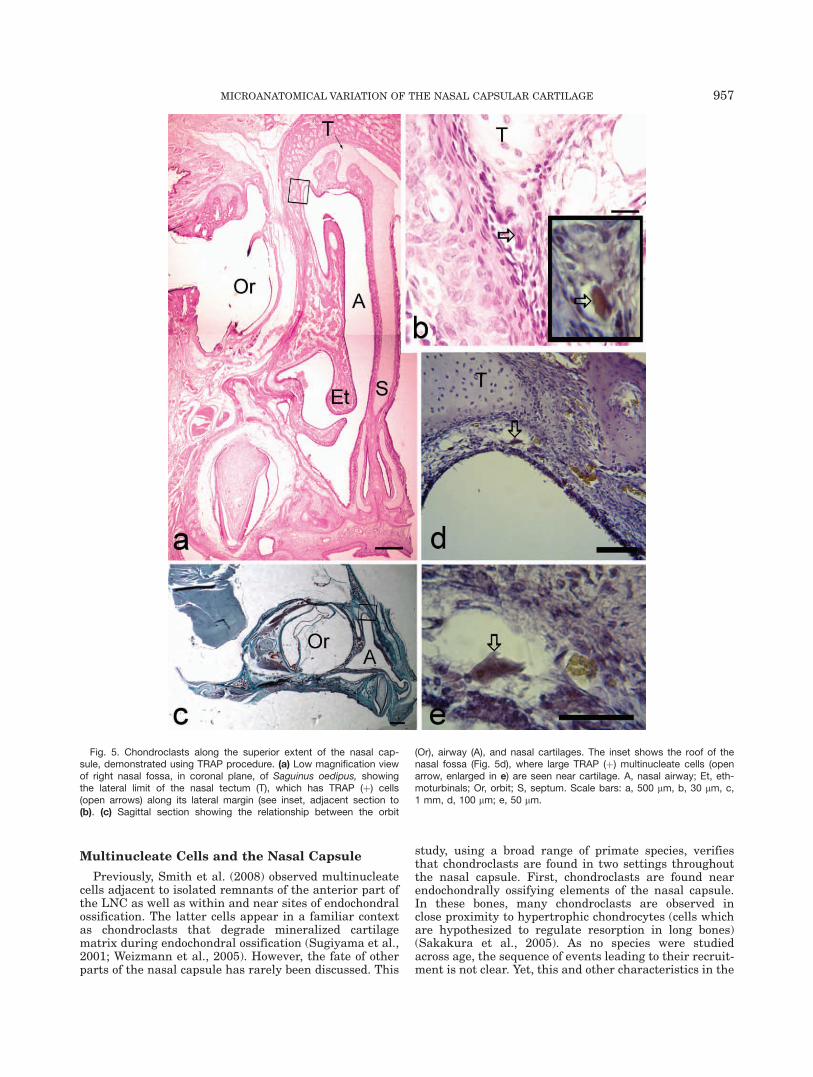
Multinucleate Cells and the Nasal Capsule
Previously, Smith et al. (2008) observed multinucleatecells adjacent to isolated remnants of the anterior part ofthe LNC as well as within and near sites of endochondralossification. The latter cells appear in a familiar contextas chondroclasts that degrade mineralized cartilagematrix during endochondral ossification (Sugiyama et al.,2001; Weizmann et al., 2005). However, the fate of otherparts of the nasal capsule has rarely been discussed. This
study, using a broad range of primate species, verifiesthat chondroclasts are found in two settings throughoutthe nasal capsule. First, chondroclasts are found nearendochondrally ossifying elements of the nasal capsule.In these bones, many chondroclasts are observed inclose proximity to hypertrophic chondrocytes (cells whichare hypothesized to regulate resorption in long bones)(Sakakura et al., 2005). As no species were studiedacross age, the sequence of events leading to their recruit-ment is not clear. Yet, this and other characteristics in the
Fig. 5. Chondroclasts along the superior extent of the nasal cap-sule, demonstrated using TRAP procedure. (a) Low magnification viewof right nasal fossa, in coronal plane, of Saguinus oedipus, showingthe lateral limit of the nasal tectum (T), which has TRAP (þ) cells(open arrows) along its lateral margin (see inset, adjacent section to(b). (c) Sagittal section showing the relationship between the orbit
(Or), airway (A), and nasal cartilages. The inset shows the roof of thenasal fossa (Fig. 5d), where large TRAP (þ) multinucleate cells (openarrow, enlarged in e) are seen near cartilage. A, nasal airway; Et, eth-moturbinals; Or, orbit; S, septum. Scale bars: a, 500 lm, b, 30 lm, c,1 mm, d, 100 lm; e, 50 lm.
MICROANATOMICAL VARIATION OF THE NASAL CAPSULAR CARTILAGE 957
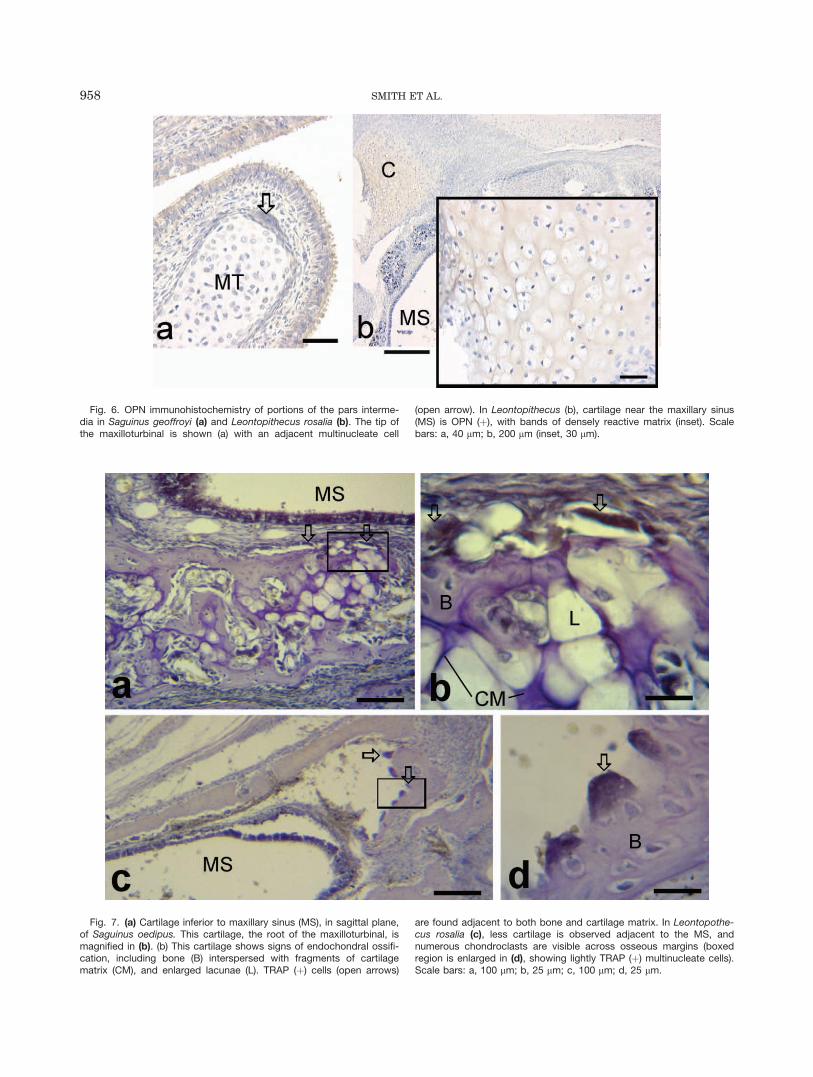
Fig. 6. OPN immunohistochemistry of portions of the pars interme-dia in Saguinus geoffroyi (a) and Leontopithecus rosalia (b). The tip ofthe maxilloturbinal is shown (a) with an adjacent multinucleate cell
(open arrow). In Leontopithecus (b), cartilage near the maxillary sinus(MS) is OPN (þ), with bands of densely reactive matrix (inset). Scalebars: a, 40 lm; b, 200 lm (inset, 30 lm).
Fig. 7. (a) Cartilage inferior to maxillary sinus (MS), in sagittal plane,of Saguinus oedipus. This cartilage, the root of the maxilloturbinal, ismagnified in (b). (b) This cartilage shows signs of endochondral ossifi-cation, including bone (B) interspersed with fragments of cartilagematrix (CM), and enlarged lacunae (L). TRAP (þ) cells (open arrows)
are found adjacent to both bone and cartilage matrix. In Leontopothe-cus rosalia (c), less cartilage is observed adjacent to the MS, andnumerous chondroclasts are visible across osseous margins (boxedregion is enlarged in (d), showing lightly TRAP (þ) multinucleate cells).Scale bars: a, 100 lm; b, 25 lm; c, 100 lm; d, 25 lm.
958 SMITH ET AL.

nasal capsule of tamarins suggest some similarities toendochondrally ossifying long bones. For example, someremnants of the posterior part of the LNC have matrixcharacteristics of endochondrally ossifying bone (seeabove; and see Smith et al., 2008).
In other locations, chondroclasts are observed in anentirely distinct context. In the anterior part of the LNCof tamarins, cartilage remnants occur in isolated islands.Here, chondroclasts are adjacent to the margins of carti-lage that show no features consistent with endochondralossification. Indeed, many of these islands of cartilageare anterior to the ethmoid bone. The sample studiedhere revealed some remnants that were partially brokendown, and these too have OPN (�) matrix. These dis-tinctions from the locations where endochondralossification occurs strongly suggest there are twodistinct mechanisms of chondro-/osteoclast recruitmentin the nasal capsule. Distinct fates of viscerocranial car-tilages have been described elsewhere in the facialskeleton. For example, chondroclasts appear to resorbMeckel’s cartilage without the involvement of the signal-ing system described for long bone ossification, such asRANKL (Sakakura et al., 2005).
Findings on chondroclast distribution also bear on theprocess of pneumatization. Chondroclasts are observedalong both cartilaginous and bony surfaces in newborn pri-mates. In some of the species studied, pneumaticexpansion of the sinus cavity is most certainly underway atbirth, based on mapping of bone cells (Smith et al., 2005,2011) and cross-age comparisons of sinus size (Smith et al.,2008, 2010). This offers a specific context for cartilagebreakdown. Recruitment of chondro-/osteoclasts may berapid, possibly occurring simultaneously in cartilage andbone in regions of secondary pneumatization.
SUMMARY AND CONCLUSIONS
At birth, matrix properties differ between portions of thenasal capsule that ultimately form elements of the ethmoid
bone and regions of the nasal capsule that have no post-natal (descendant) structure. This distinction, regardingexpression of OPN and distribution of glycosaminoglycans,applies broadly to primates. However, the extent of carti-lage that remains in the pars anterior and pars intermediavaries. To some degree, the extent of cartilage present atbirth reflects incipient sites of sinus expansion. Primatespecies known to form large frontal sinuses have an incom-plete cartilaginous LNC at birth. Several species in whichthe frontal bone does not pneumatize have more completelateral cartilaginous walls. One interpretation could bethat this large remnant delays pneumatization. On theother hand, such remnants may simply reflect the pace ofpneumatic expansion (none, in the case of S. boliviensis,Rossie, 2006; Smith et al., 2010).
It should be noted that bushbabies (Otolemur crassi-cadatus and Galago moholi) have little remaining LNCat birth but appear to have no pneumatization of thefrontal bone (Tuckmantel et al., 2009). Thus, the timingof breakdown may coincide with the advent of pneumaticexpansion into bone is some species, but the LNC isreplaced or resorbed regardless of whether secondarypneumatization occurs. We suggest certain findingsherein, such as the great number of observations ofchondroclasts near anterior nasal capsule fragments intamarins, may be a manifestation of a specific stage ofpneumatization and may occur earlier or later acrossspecies. A complete picture of the temporal pattern ofpneumatization in primates is lacking, primarily becausea broad fetal to postnatal age series is elusive except forvery few species (e.g., see Smith et al., 2008).
Finding a non-primate animal model to understandpneumatization, preferably a less rare, faster reproduc-ing taxon would seem a desirable route to further ourunderstanding of pneumatization. Yet unique character-istics of primates make this challenging. One aspect oflife history that distinguishes the order Primates frommost other mammals is the relatively prolonged gesta-tion length (Martin, 1983). It may be precisely this
TABLE 2. Species characteristics in the paranasal regions
Species
Secondary pneumatization
Refs.Of maxilla Of frontal
AnthropoidsCallithrix jacchus Y ?a Rossie (2006); Hershkovitz (1977)Cebuella pygmaea Y N Hershkovitz (1977)Leontopithecus rosalia Y Y Smith et al. (2005); Rossie (2006)Saguinus geoffroyi Y Y Smith et al. (2005), (2008)Saguinus oedipus Y Y Rossie (2006)Pithecia pithecia Y N Hershkovitz (1977)Saimiri boliviensis N N Rossie (2006)
Strepsirrhinesb
Eulemur macaco ? Y Tuckmantel et al. (2009)Eulemur mongoz ? Y Tuckmantel et al. (2009)Lemur catta ? N Rossie, unpublished dataPropithecus verrauxi ? Y Tuckmantel et al. (2009)Microcebus murinus ? N Tuckmantel et al. (2009)Mirza coquereli ? ? –Galago moholi ? N Rossie, unpublishedOtolemur crassicaudatus ? N Tuckmantel et al. (2009)
N, no; Y, yes.aHershkovitz remarked on variation in frontal pneumatization in the genus Callithrix, with some species having small‘‘cells’’ communicating with the middle meatus.bSecondary pneumatization of the maxilla has not been investigated in strepsirrhines.
MICROANATOMICAL VARIATION OF THE NASAL CAPSULAR CARTILAGE 959

characteristic that makes the perinatal age a dynamicperiod in nasal capsule development in primates. Other,more altricial, species have much less ossified andless fragmented nasal capsule in the perinatal period(e.g., rats—Vidic, 1971b, Tupaia glis—Smith, unpub-lished data) compared to primates. A broader difficultyexists that our understanding of nasal capsule ontogenyand fate in a comparative perspective is poor. This studyillustrates that such knowledge may inform our under-standing of secondary pneumatization. Focal capsularcartilage breakdown should be regarded as part ofthe process of secondary pneumatization, with chondro-clasts breaking down cartilage as one of the earliestevents. Ontogenetic studies of diverse vertebrate taxa(specifically certain mammals and birds) are needed to fur-ther our understanding of this unique growth mechanism.
ACKNOWLEDGEMENT
We thank T. Popowics for helpful discussions on theTRAP procedure.
LITERATURE CITED
Bark-Shalom T, Schickler M, Knopov V, Shapira R, Hurwitz S,Pines M. 1995. Synthesis and phosphorylation of osteopontin byavian epiphyseal growth-plate chondrocytes as affected by differ-entiation. Comp Biochem Physiol 111C:49–59.
Colnot CI, Helms JA. 2001. A molecular analysis of matrix remodelingand angiogenesis during long bone development. Mech Dev 100:245–250.
Halleen JM, R€ais€anen S, Salo JJ, Reddy SV, Roodman GD,Hentunen TA, Lehenkari PP, Kaija H, Vihko P, V€a€an€anen HK.1999. Intracellular fragmentation of bone resorption productsby reactive oxygen species generated by tartrate-resistant acidphosphatase. J Biol Chem 274:22907–22910.
Hamrick MW. 1999. Development of epiphyseal structure and func-tion in Didelphis virginiana (Marsupiala, Didelphidae). J Morphol239:283–296.
Hayman AR, Jones SJ, Boyde A, Foster D, Colledge WH, CarltonMB, Evans MJ, Cox TM. 1996. Mice lacking tartrate-resistantacid phosphatase (Acp 5) have disrupted endochondral ossificationand mild osteopetrosis. Development 122:3151–3162.
Hershkovitz P. 1977. Living new world monkeys (Platyrrhini). Vol.1.Chicago: University of Chicago Press.
Hirschmann A, McCabe DM. 1969. Staining of intracellular granulesin fresh epiphyseal cartilage by cationic dyes. Calcif Tissue Int 4:260–268.
Maier W. 2000. Ontogeny of the nasal capsule in cercopithecoids: acontribution to the comparative and evolutionary morphology ofcatarrhines. In: Whitehead PF, Jolly CJ, editors. Old World mon-keys. Cambridge: Cambridge University Press. p 99–132.
Martin RD. 1983. Human brain evolution in an ecological context(James Arthur lecture on the evolution of the human brain, no.52, 1982). American Museum of Natural History, New York, NY.
Reinholt F, Hultenby K, Oldenberg A, Heinegard D. 1990. Osteo-pontin—a possible anchor of osteoclasts to bone. Proc Natl AcadSci USA 87:4473–4475.
Roach HI. 1997. New aspects of endochondral ossification in thechick: chondrocyte apoptosis, bone formation by former chondro-cytes, and acid phosphotase activity in the endochondral bone ma-trix. J Bone Min Res 12:795–805.
Rossie J. 2006. Ontogeny and homology of the paranasal sinuses inPlatyrrhini (Mammalia: Primates). J Morphol 267:1–40.
Sakakura Y, Tsuruga E, Irie K, Hosokawa Y, Nakamura H, YajimaT. 2005. Immunolocalization of receptor activator of nuclear fac-tor-jB ligand (RANKL) and osteoprotegerin (OPG) in Meckel’scartilage compared with developing endochondral bones in mice.J Anat 207:325–337.
Sato I, Sunohara M, Mikami A, Watanabe K, Sato T. 1998.Immunocytochemical study of the maxilla and maxillary sinusduring human fetal development. Okajimas Folia Anat Jpn 75:205–216.
Schaeffer JP. 1920. The nose, paranasal sinuses, nasolacrimalpassageways, and olfactory organ in man. Philadelphia: P. Blakis-ton’s Son & Co.
Shah RK, Dhingra JK, Carter BL. 2003. Paranasal sinus develop-ment: a radiographic study. Laryngoscope 113:205–299.
Smith TD, Rossie JB. 2008. Nasal Fossa of Mouse and DwarfLemurs (primates, cheirogaleidae). Anat Rec 291:895–915.
Smith TD, Bhatnagar KP, Bonar CJ, Shimp KL, Mooney MP, SiegelMI. 2003. Ontogenetic characteristics of the vomeronasal organ inSaguinus geoffroyi and Leontopithecus rosalia with comparisonsto other primates. Am J Phys Anthropol 121:342–353.
Smith TD, Bhatnagar KP, Rossie JB, Docherty BA, Burrows AM,Mooney MP, Siegel MI. 2007. Scaling of the first ethmoturbinal innocturnal strepsirrhines: olfactory and respiratory surfaces. AnatRec 290:215–237.
Smith TD, Rossie JB, Cooper GM, Mooney MP, Siegel MI. 2005.Secondary pneumatization in the maxillary sinus of callitrichidprimates: insights from immunohistochemistry and bone cell dis-tribution. Anat Rec 285:677–689.
Smith TD, Rossie JB, Cooper GM, Carmody KA, Schmieg RM,Bonar CJ, Mooney MP, Siegel MI. 2010. The maxillary sinusin three genera of new world monkeys: factors that constrain sec-ondary pneumatization. Anat Rec, 293:91–107.
Smith TD, Rossie JB, Cooper GM, Schmieg RM, Bonar CJ,Mooney MP, Siegel MI. 2011. Comparative micro CT and histo-logical study of maxillary pneumatization in four species of NewWorld monkeys: the perinatal period. Am J Phys Anthropol 144:392–410.
Smith TD, Rossie JB, Docherty BA, Cooper GM, Bonar CJ, SilverioAL, Burrows AM. 2008. Fate of the nasal capsular cartilages inprenatal and perinatal tamarins (Saguinus geoffroyi) and extentof secondary pneumatization of maxillary and frontal sinuses.Anat Rec 291:1397–1413.
Sperber G. 2000. Craniofacial development. Hamilton: B.C. Decker.Sugiyama H, Imada M, Sasaki A, Ishino Y, Kawata T, Tanne K.2001. The expression of osteopontin with condylar remodeling ingrowing rats. Clin Orthod Res 4:194–199.
Tuckmantel S, R€ollin A, Muller AE, Soligo C. 2009. Facial correlatesof frontal bone pneumatisation in strepsirrhine primates. MammBiol 74:25–35.
Vidic B. 1971a. The morphogenesis of the lateral nasal wall in theearly prenatal life of man. Am J Anat 130:121–139.
Vidic B. 1971b. The prenatal morphogenesis of the lateral nasalwall in the rat (Mus rattus). J Morphol 133:303–317.
Wang R-G, Jiang S-C, Gu R. 1994. The cartilaginous nasal capsuleand embryonic development of human paranasal sinuses. J Oto-laryngol 23:239–243.
Weiglein AH. 1999. Development of the paranasal sinuses inhumans. In: Koppe T, Nagai H, Alt K, editors. The paranasalsinuses of higher primates. Berlin: Quintessence. p 35–50.
Weizmann S, Tong A, Reich A, Genina O, Yayon A, Monsanego-Ornan E. 2005. FGF upregulates osteopontin in epiphysealgrowth plate chondrocytes: implications for endochondral ossifica-tion. Matrix Biol 24:520–529.
Witmer LM. 1999. The phylogenetic history of the paranasalair sinuses. In: Koppe T, Nagai H, Alt KW, editors. The paranasalsinuses of higher primates. Berlin: Quintessence. p 21–34.
Wojtowicz A, Montella A, Bandiera P, Wlodarski K, Wojtowicz K,Kochanowska I, Ostrowski K. 2002. Human mucosal epitheliuminvolvement in prenatal growth of maxillary sinuses. Ital J AnatEmbryol 107:257–265.
Yeh K-D, Popowics T. 2011. Molecular and structural assess-ment of alveolar bone during tooth eruption and function inthe miniature pig, Sus Scrofa. Anat Histol Embryol 40:283–291.
Zollikofer CPE, Weissman JD. 2008. A morphogenetic model of cra-nial pneumatization based on the invasive tissue hypothesis.Anat Rec 291:1446–1454.
960 SMITH ET AL.
Related Documents











