Printed in Sweden Copyright @ 1980 by Academic Press, Inc. All rights of reproduction in any form reserved 0014.4827/80/010191-09$02.W/O Experimental Cell Research 12.5 (1980) 191-199 FATE OF PROTEINS SYNTHESIZED IN MITOCHONDRIA OF CULTURED MAMMALIAN CELLS REVEALED BY ELECTRON MICROSCOPE RADIOAUTOGRAPHY E. KNECHT, J. HERNANDEZ-YAGO, A. MARTINEZ-RAMGN and S. GRISOLiA lnstituto de Investigaciones Citoldgicas de la Caja de Ahorros, Valencia - 10, Spain SUMMARY Synthesis of mitochondrial proteins was studied in a monkey kidney epithelium cell line by electron microscope radioautography after pulse label for 1 h with [3H]leucine in the presence of cycloheximide followed by chase in the absence of the antibiotic. At all chase times tested the highest specific activities were found over mitochondria and lysosomes (dense bodies). Based on regression analysis of the decay curve of mitochondrial labeling, there appeared to be two main mitochondrial populations with apparent half-lives of l-2 h and 6 days. In parallel experiments, cells were labelled with [3H]leucine in the presence of cycloheximide plus chloramphenicol. Chloramphenicol markedly inhibited labelling over mitochondria and dense bodies, whereas it had no significant effect on labelling of other cell compartments (nucleus, endoplasmic reticulum, remainder cytoplasm). The significance of the cycloheximide-insensitive and chloramphenicol- sensitive lysosomal labelling is discussed. The data suggest that in cultured cells lysosomes are involved in the degrading of mitochondrial proteins. The mechanism by which cells degrade their proteins is not well understood. Al- though there is evidence for alternate sys- tems of intracellular protein breakdown, lysosomes have been frequently proposed as the main proteolytic site [l]. However, direct lysosomal participation in this pro- cess remains to be demonstrated. In order to limit the problem we have concentrated our studies on mitochondrial proteins. Since mitochondrial enzymes such as carbamyl phosphate synthetase, malic dehydrogenase and glutamate de- hydrogenase have half-lives of 7.7, 2.6 and ca 1 day, respectively [2], bulk or unregu- lated autophagy seems unlikely. Although non-lysosomal proteases specific for pyri- doxal-requiring enzymes have been re- ported in mitochondrial preparations [3], 13-791816 evidence has been presented pointing to a main role for lysosomes in mitochondrial proteolysis [4, 51. If lysosomes participate extensively in the degradation of mito- chondrial proteins, they could contain these proteins or their hydrolytic products which could then be detected with the aid of radio- active or other markers. Electron microscope radioautography, which allows fine localization, could pro- vide a new approach for studying protein degradation. In the present paper, we report a successful demonstration, by fine struc- tural radioautographical techniques, of the fate of intracellular labelled proteins under conditions where mitochondrial protein was the primary source of label. We have ac- complished this by taking advantage of the capability of mitochondria to synthesize Exp Cd Res 12.5 (1980)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Printed in Sweden Copyright @ 1980 by Academic Press, Inc.
All rights of reproduction in any form reserved 0014.4827/80/010191-09$02.W/O
Experimental Cell Research 12.5 (1980) 191-199
FATE OF PROTEINS SYNTHESIZED IN MITOCHONDRIA
OF CULTURED MAMMALIAN CELLS REVEALED BY
ELECTRON MICROSCOPE RADIOAUTOGRAPHY
E. KNECHT, J. HERNANDEZ-YAGO, A. MARTINEZ-RAMGN and S. GRISOLiA
lnstituto de Investigaciones Citoldgicas de la Caja de Ahorros,
Valencia - 10, Spain
SUMMARY Synthesis of mitochondrial proteins was studied in a monkey kidney epithelium cell line by electron microscope radioautography after pulse label for 1 h with [3H]leucine in the presence of cycloheximide followed by chase in the absence of the antibiotic. At all chase times tested the highest specific activities were found over mitochondria and lysosomes (dense bodies). Based on regression analysis of the decay curve of mitochondrial labeling, there appeared to be two main mitochondrial populations with apparent half-lives of l-2 h and 6 days. In parallel experiments, cells were labelled with [3H]leucine in the presence of cycloheximide plus chloramphenicol. Chloramphenicol markedly inhibited labelling over mitochondria and dense bodies, whereas it had no significant effect on labelling of other cell compartments (nucleus, endoplasmic reticulum, remainder cytoplasm). The significance of the cycloheximide-insensitive and chloramphenicol- sensitive lysosomal labelling is discussed. The data suggest that in cultured cells lysosomes are involved in the degrading of mitochondrial proteins.
The mechanism by which cells degrade their proteins is not well understood. Al- though there is evidence for alternate sys- tems of intracellular protein breakdown, lysosomes have been frequently proposed as the main proteolytic site [l]. However, direct lysosomal participation in this pro- cess remains to be demonstrated.
In order to limit the problem we have concentrated our studies on mitochondrial proteins. Since mitochondrial enzymes such as carbamyl phosphate synthetase, malic dehydrogenase and glutamate de- hydrogenase have half-lives of 7.7, 2.6 and ca 1 day, respectively [2], bulk or unregu- lated autophagy seems unlikely. Although non-lysosomal proteases specific for pyri- doxal-requiring enzymes have been re- ported in mitochondrial preparations [3],
13-791816
evidence has been presented pointing to a main role for lysosomes in mitochondrial proteolysis [4, 51. If lysosomes participate extensively in the degradation of mito- chondrial proteins, they could contain these proteins or their hydrolytic products which could then be detected with the aid of radio- active or other markers.
Electron microscope radioautography, which allows fine localization, could pro- vide a new approach for studying protein degradation. In the present paper, we report a successful demonstration, by fine struc- tural radioautographical techniques, of the fate of intracellular labelled proteins under conditions where mitochondrial protein was the primary source of label. We have ac- complished this by taking advantage of the capability of mitochondria to synthesize
Exp Cd Res 12.5 (1980)

192 E. Knecht et al.
10
L@
5
L T
2-4 da Fig. I. Abscissa: time (hours); ordinate: cell density (cells/ml x 10m5). O-O, Cycloheximide-treated cells; m-m, control cells (treated with buffer instead of cycloheximide).
Growth curves of MS cells growing exponentially (-4x 105 cells/ml) after exposure to cycloheximide (200 pg/ml) for 1 h and washing three times with drug- free medium.
some of their proteins and of the different sensitivities of mitoribosomes and cyto- plasmic ribosomes to inhibitors [6].
MATERIALS AND METHODS
Cell culture and analytical procedures MS cells, an established monkey kidney proximal epithelium cell line, were cultured as described previ- ously [7]. The number of cells and the effect of cyclo- heximide (CX) and CX plus chloramphenicol on cell growth was determined with a Coulter counter. The ef- fects of inhibitors of protein synthesis on the incor- ooration of 13Hlleucine into cultured cells was deter- mined by &bating exponentially growing cells (-5x IO5 cells/ml) in culture medium without and with inhibitors. After ‘10 min, 5 pCi/ml [3H]leucine (57 Ci/ mmol) was added. Cells were incubated for 1 h at 37°C washed three times with fresh medium contain- ing 2 mM cold leucine, in the presence of inhibitors and again three times in their absence. Then the cells were immediately detached from the flasks and washed bv centrifueation. first with 10 ml ice-cold basal medium (lackiig serum and antibiotics) and then twice with 10 ml phosphate-buffered saline. The washed cells were precipitated with 5 % trichloroacetic acid (TCA). After dissolving the precipitated protein in 0.2 N NaOH, the radioactivity was determined by liquid scintillation counting. Protein was assayed by the method of Lowry et al. [S].
Autoradiography Cells were labelled and washed as described above, except that [3H]leucine was added to give 300 &i/ml. The cells were labelled in the presence of CX (200 pg/
Table 1. Effects of cycloheximide and/or chloramphenicol on [3H]leucine incorpora- tion into proteins by MS cells
Inhibitor” cpm/mg protein*
% In- hibi- tionc
None Cycloheximide Chloramphenicol Cycloheximide+
chloramphenicol
102 041 f4 581 3 113+ 282 96.9
101 83lf4 360 -
2 295f 229 97.8
0 Cells were incubated with r3H]leucine in the pres- ence or absence of cycloheximide (200 pg/ml) and/or chloramphenicol (100 pg/ml) as described in Materials and Methods. * Data are given as mean of 4 experiments + S.E.M. c Data are expressed as % of the control incorporation inhibited by antibiotics.
ml), CX plus chloramphenicol (100 pg/ml) and/or CX plus erythromycin (100 &ml). Washed cells were im- mediately fixed or further-incubated for different chase periods in fresh medium containing 8 mM cold leucine in the absence of antibiotics. Culture medium was re- placed by fresh medium plus non-labelled leucine at 3. 6 and 24 h chase. Cells were fixed for 1 h with 0.1 M Na-cacodylate buffered (pH 7.3) glutaraldehyde (2 %), washed overnight, post-fixed 45 min in 1% osmium tetroxide dissolved in 0.05 M acetate Verorml buffer, pH 7.3, dehydrated in acetone and embedded in Vestopal W by standard methods. Silver-to-gold sec- tions were mounted on celloidin-coated slides, and stained for 1 h with 2% uranyl acetate. A carbon film about 10 nm thick was evaporated over the sections which were then coated with an Ilford L4 emulsion using the ‘dipping’ technique. The slides were stored in light-tight boxes for l&l2 weeks, and the radioauto- graphs were developed, after 30 set gold latensifica- tion. in Phenidon-ascorbic acid. Sections were re- moved from the slides, mounted on copper grids, im- mersed in isoamyl acetate for 20 min and treated with alkaline lead citrate for 1 min.
For each time point, 40 randomly selected areas of about 200 pm2 from two different experiments were counted; they gave essentially the same results. The distribution of silver grains over different cell compart- ments in the radioautographs was determined with the ‘resolution boundary circle’ with five points [9, 101,
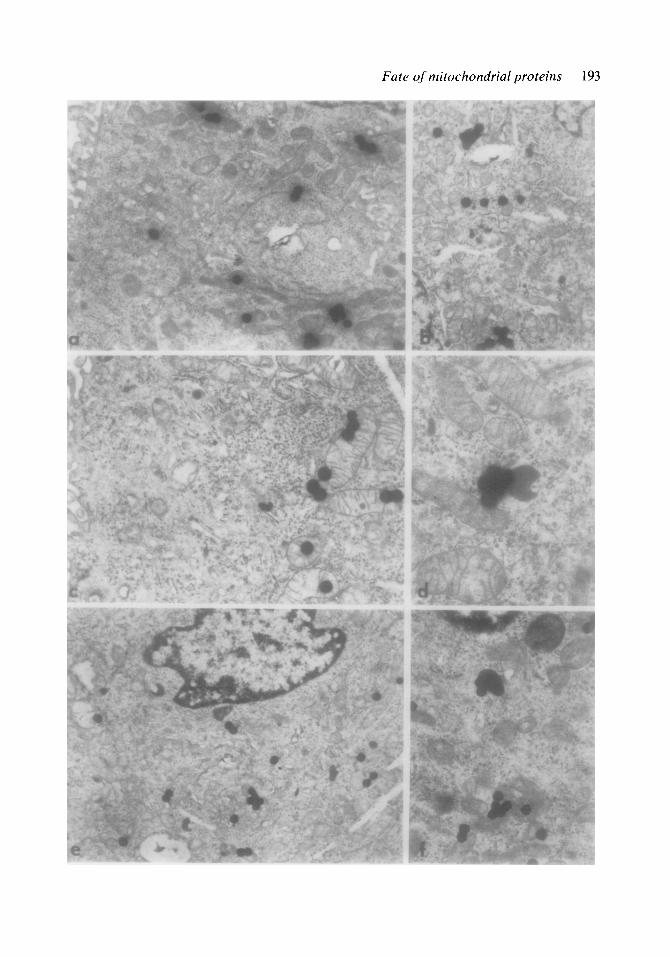
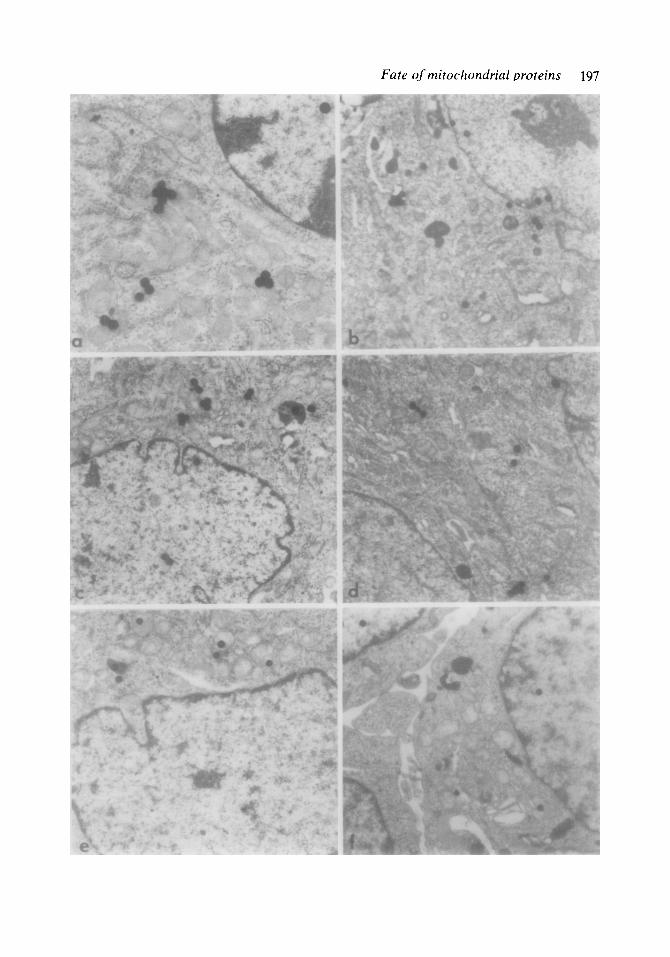
Fig. 2. Electron microscopic radioautographs of MS cells labelled with [3H]leucine in the presence of cyclo- heximide. The cells were washed and (a) immediatelv fixed or incubated again with cold let&e for (b) 3, (c, d) 6 or (e, j) 12 h. The silver grains are mainly lo- cated over mitochondria and lysosomes. (a) X 12 000; If;’ xg7g; (c) x16000; (d) x25000; (e) x10000;
x .
El-p Cc//Rc.r 125 (1980)
Fate of mitochondrial proteins 193
194 E. Knecht et al.
Table 2. Specific activities of different cell compartments at various periods of chase after labelling with [3H]leucine in the presence of cycloheximide
Chase time
Cell compartment
Mitochondria Spec. act. Surface counted
Dense bodies Spec. act. Surface counted
Reticulum Spec. act. Surface counted
0 h 3h 6h 12 h 24 h 48 h
135.8f7.7 94.7k4.8 75.9f4.1 67.4k4.0 52.5f4.5 30.5k2.2 747.7 737.9 711.3 700.4 701.2 696.2
93.5kll.l 74.5k8.7 81.5k10.9 71.4k8.0 51.3k6.4 25.Ok4.0 83.0 86.9 77.2 71.6 85.8 76.1
44.5f5.3 33.8k2.9 29.153.7 25.3t3.0 19.1t2.1 11.7L1.3 234.9 244.8 225.4 257.9 253.5 242.6
Remainder cytoplasm SDec. act. 38.5k1.8 33.4k2.2 29.5k2.3 26.151.7 20.6k 1.8 12.9kO.8 Surface counted 4 700.4 4 552.9 4 198.4 4 121.2 4 290.9 4 106.1
Nucleus Spec. act. 35.6k 1.9 26.9k1.9 22.9f2.1 21.9f2.3 18.1f2.1 12.1+1.3 Surface counted 2 534.2 2 634.0 2 551.1 2 594.9 2 663.3 2 543.5
Total cell Spec. act. 47.1f1.6 37.2fl.7 32.1k2.0 28.851.7 22.gk 1.6 14.3f0.8
Specific activities were calculated as described in Materials and Methods. The values represent the mean kS.E.M. Surface data are given in pm*.
and the relative volume occupied by these compart- ments by the ‘point hit method’ [lo] using a double lattice test system with 2200 points (for nuclei, mito- chondria and remainder cytoplasm) and 3600 (for endoplasmic reticulum and dense bodies). Specific ac- tivity is defined as the number of grains per unit area of each cell compartment and referring to 100 prn2. Relative grain concentration was calculated by divid- ing the percentage of all grains associated with any given compartment by the percentage of the total area occupied by that compartment. The mean value of these counts t standard error of the mean (S.E.M.) represented the final count.
RESULTS AND DISCUSSION
To label mitochondrial proteins selectively, CX was used as an inhibitor. The effects of pre-treatment with CX, at various doses and for various times, on growth of MS cells and on protein synthesis were first tested. From these data, subsequent experiments were done at 200 pglml CX and with 1 h ex- posure to r3H]leucine.
The electron microscopic appearance of
MS cells has been described previously [7]. At the concentrations used, CX gave rise to small, line structural changes, i.e. Golgi areas were reduced to some extent and there was a nearly complete absence of autophagic vacuoles. Other components of the lysosomal system and other organelles appeared unaltered.
As shown in fig. 1, the effects of CX on cell growth were largely reversible (possibly there is a short lag). The growth curve of CX plus chloramphenicol was virtually in- distinguishable from that of CX-treated cells. Table 1 shows that the incorporation of [3H]leucine was inhibited 96-97% when CX was added to the incubation medium. Chloramphenicol (100 pglml) had little ef- fect on total protein synthesis. When cells were labelled in the presence of CX plus chloramphenicol, there was an additional inhibition over the CX-sensitive [“Hlleu-
Fate of mitochondrial proteins 195
Table 3. Effect of erythromycin on cyclo- heximide insensitive labelling of the dif- ferent cell compartmentsa
Relative specific activity
Cell compartment Cyclo- heximide
Cyclo- heximide+ erythro- mycin
Mitochondria 2.88f0.17 2.77kO.10 Dense bodies 1.98k0.25 1.81f0.22 Reticulum 0.95f0.12 1.00f0.09 Remainder cytoplasm 0.82kO.05 0.84kO.03 Nucleus 0.76kO.05 0.79kO.05
a Cells were incubated for 1 h with [3H]leucine in the presence of cycloheximide (200 pg/ml) and cyclo- heximide+erythromycin (100 pg/ml), immediately fixed and prepared for electron microscope radioauto- graphy. Relative specific activities were calculated as described in Materials and Methods.
tine incorporation. These results indicate that a significant portion of the radioactivity found in CX-treated MS cells after 1 h [3H]- leucine incorporation is due to mito- chondrial protein synthesis.
In all intervals after administration of [3H]leucine in the presence of CX, radio- autographs revealed a great number of sil- ver grains overlying the mitochondria, and a smaller number of grains associated with other cell compartments (fig. 2). The spe- cific activities associated with the different compartments considered at each interval (table 2) indicated that only mitochondria and lysosomes (dense bodies) had a con- centration of label significantly higher than the mean value for the cell. The possibility that some CX-insensitive radioactivity found in the cells could be due to myco- plasma contamination was rendered un- likely, since labelling of cells in the pres- ence of CX plus erythromycin (100 pglml) gave similar results (table 3), as estimated by the Student’s t-test (p<O.OS).
To investigate in more detail the nature of
mitochondrial proteins labelled in the pres- ence of CX, we assumed that the decay of mitochondrial radioactivity was due, first, to loss of labelled mitochondrial proteins and second, to cell and mitochondrial divi- sion. This last contribution can be easily corrected for on the assumption that the total mitochondrial area per cytoplasmic area remains constant through the cell cycle [ 111. When the corrected values for mito- chondrial radioactivity decay (obtained by multiplying the specific activity by the growth rate at each time, fig. 3) were plotted semilogarithmically, the best fitted regres- sion lines were obtained when two lines were considered indicating two major groups of mitochondrial proteins, i.e. with apparent short and long half-lives (t&) of 1.6 h and 6 days, respectively (fig. 4). Re- utilization of [3H]leucine may introduce an overestimation of these t+. However, since the large volume of culture medium, con- taining excess unlabelled leucine relative to cell volume, dilutes the isotope to low con- centrations and since the frequent change of medium eliminates radioactive precursor (especially that resulting from turnover of short-lived proteins, which make the major contribution to the labelled pool), re-utiliza- tion should be minimal. Although there must be many proteins with differing ti
:
P /
1 /I
/
/
A' /
* A,A’ ‘5612 24 46
Fig. 3. Abscissa: time (hours); ordinate: growth rate. Growth rate of MS cells under the conditions of the
radioautographic experiment. The growth rate was cal- culated from the ratio of the cell density on the time indicated and the initial cell density.
ExpCeURes 125 (1980)
196 E. Knecht et al.
:
Calculation of the specific activity of dense bodies showed them to be heavily labelled (although the % of all grains over- laying them was small, dense bodies make
. up only a small portion of the cell volume (- 1%)). The possibility that the presence of silver grains in dense bodies might be due to background or to radiation emitted by heavily labelled adjacent mitochondria was excluded, since serial sections of dense bodies also appeared labelled, even when they lay well separated from mitochondria. Moreover, endoplasmic reticulum, which also often appears closely associated with mitochondria, did not show a significant labelling. Therefore, since mitochondria represent the major site in the cell of CX- insensitive r3H]leucine labelling, the most probable cause for the lysosomal labelling is the presence in dense bodies of proteins of mitochondrial origin. However, an alterna- tive explanation for the lysosomal labelling is that most of the CX-insensitive proteins synthesized in the cytoplasm are trans- ported to lysosomes. The nature of these proteins is at present unknown. Jeffreys & Craig [IS] have indicated that in cultured cells little of this label can be detected auto- radiographically as discrete bands in poly- acrylamide gel electrophoresis. Therefore,
Fig. 4. Abscissa: time (hours); ordinnfe: no. of silver grains/100 pm* mitochondria.
Decay curve of radioactive proteins in MS mito- chondria. The experimental curve (W-U) was decom- nosed into two exnonential functions: slowly renewed proteins (A---Aj and rapidly renewed- proteins (O---O, obtained bv extrapolation of the first line to tero time and subtraction from the experimental points). The calculated half-lives are 1.6 h and 6.0 days.
values, even within the two classes of pro- teins that we have distinguished, such a gross classification has been shown to be valid in bacterial and mammalian cells [12, 131. It also becomes apparent from fig. 4
1 .\ lo \ \ b \
I \
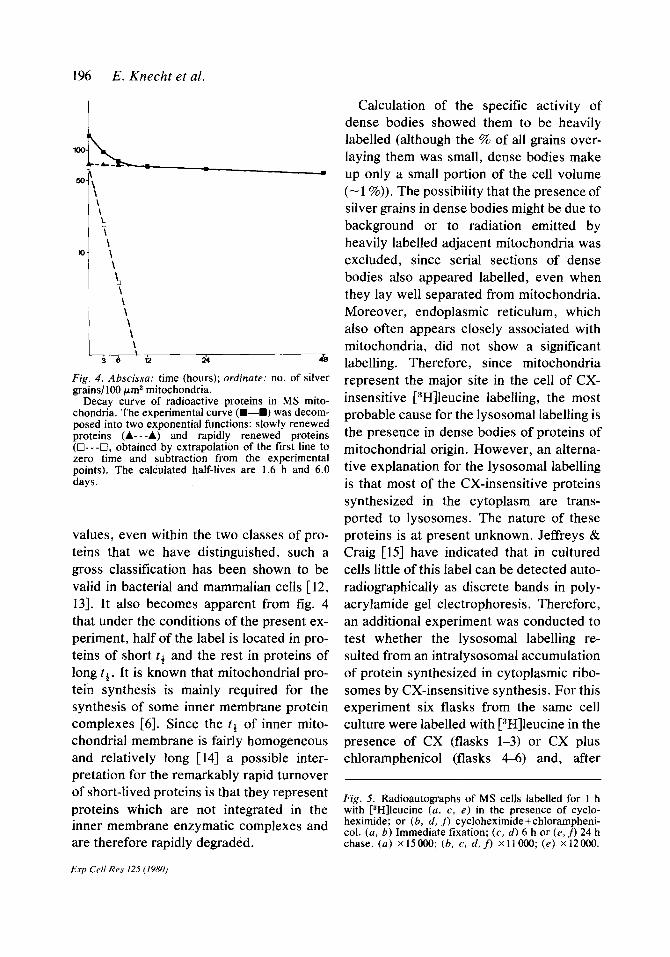
that under the conditions of the present ex- an additional experiment was conducted to periment, half of the label is located in pro- test whether the lysosomal labelling re- teins of short tt and the rest in proteins of sulted from an intralysosomal accumulation long tt . It is known that mitochondrial pro- of protein synthesized in cytoplasmic ribo- tein synthesis is mainly required for the somes by CX-insensitive synthesis. For this synthesis of some inner membrane protein experiment six flasks from the same cell complexes [6]. Since the t1 of inner mito- culture were labelled with [3H]leucine in the chondrial membrane is fairly homogeneous presence of CX (flasks l-3) or CX plus and relatively long [14] a possible inter- chloramphenicol (flasks 4-6) and, after pretation for the remarkably rapid turnover of short-lived proteins is that they represent proteins which are not integrated in the
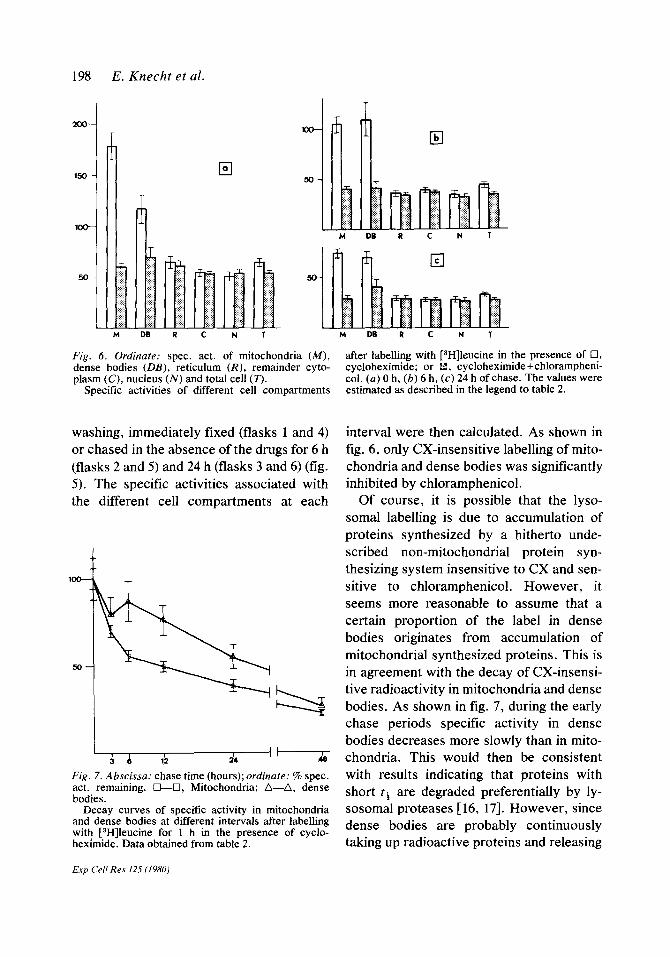
Fig. 5. Radioautographs of MS cells labelled for 1 h with r3H]leucine (a, c, e) in the presence of cyclo-
inner membrane enzymatic complexes and heximide; or (b, d, J) cycloheximide+chlorampheni-
are therefore rapidly degraded. col. (a, 6) Immediate fixation; (c, d) 6 h or (e. f) 24 h chase. (a) x15000; (b, c, d,J) x11000;(e) x12000.
Exp CeNRes 125(1980)
Fate of mitochondrial proteins 197
198 E. Knecht et al.
m- 150 100 50 El
DR R C N ..:.: . . . ::;: n : .:. :::: .:.... :.: : :: . . . . :: . ‘::; : 1 Fig. 6. Ordinate: spec. act. of mitochondria (M), after labelling with [3H]leucine in the presence of 0, dense bodies (DB), reticulum (I?), remainder cyto- cycloheximide; or q , cycloheximide+chlorampheni- plasm (C), nucleus (N) and total cell (T). col. (a) 0 h, (b) 6 h, (c) 24 h of chase. The values were
Specific activities of different cell compartments estimated as described in the legend to table 2.
washing, immediately fixed (flasks 1 and 4) or chased in the absence of the drugs for 6 h (flasks 2 and 5) and 24 h (flasks 3 and 6) (fig. 5). The specific activities associated with the different cell compartments at each
c
Fig. 7. Abscissa: chase time (hours); ordinate: % spec. act. remaining. 0-Q Mitochondria; A-A, dense bodies.
Decay curves of specific activity in mitochondria and dense bodies at different intervals after labelling with [3H]leucine for 1 h in the presence of cyclo- heximide. Data obtained from table 2.
DB R C N 1 M DB R C N T
interval were then calculated. As shown in fig. 6, only CX-insensitive labelling of mito- chondria and dense bodies was significantly inhibited by chloramphenicol.
Of course, it is possible that the lyso- somal labelling is due to accumulation of proteins synthesized by a hitherto unde- scribed non-mitochondrial protein syn- thesizing system insensitive to CX and sen- sitive to chloramphenicol. However, it seems more reasonable to assume that a certain proportion of the label in dense bodies originates from accumulation of mitochondrial synthesized proteins. This is in agreement with the decay of CX-insensi- tive radioactivity in mitochondria and dense bodies. As shown in fig. 7, during the early chase periods specific activity in dense bodies decreases more slowly than in mito- chondria. This would then be consistent with results indicating that proteins with short tt are degraded preferentially by ly- sosomal proteases [ 16, 171. However, since dense bodies are probably continuously taking up radioactive proteins and releasing
Exp Cell Res 125 (1980)

protein degradation products, the dis- crepancy in dense bodies labelling during early and late chase periods may simply reflect the different velocities of transloca- tion of mitochondrial proteins versus elimi- nation of labelled protein and products thereof from dense bodies.
Obviously it is difficult to quantitate how much of the radioactivity incorporated into mitochondria passes to dense bodies, though some obviously does. Also, the pos- sibility that the labelling in dense bodies corresponds to partially degraded proteins (e.g. large peptides) cannot be excluded. Moreover, although low molecular sub- stances are leached out from cells fixed and embedded for electron microscopy, in some cases small peptides may be retained by trapping inside a matrix of fixed proteins [18]. Therefore, whether there is initiation of degradation in mitochondria (and/or cytoplasm) and completion in lysosomes is not known, nor whether degradation of mitochondrial proteins also occurs in mito- chondria [19]. The mechanism of entry of mitochondrial proteins into lysosomes re- mains speculative. However, an autophagic segregation process of the whole mito- chondria seems unlikely, at least in the early chase periods, because typical auto- phagic vacuoles containing mitochondria were uncommon and because CX, which inhibits protein degradation [20], also in- hibits formation of autophagic vacuoles INI.
Nevertheless, it is clear from the work presented here that a fraction of proteins synthesized in mitochondria is indeed de- graded in lysosomes, and this tends to con- firm the involvement of lysosomes in the turnover of other mitochondrial proteins.
Fate of mitochondrial proteins 199
Further work using more specific labelling, e.g. immunocytochemical localization of other mitochondrial proteins, may clarify the problem.
We thank Dr F. Thompson and Dr V. Rubio for critical reading of the manuscript, and R. Wallace, A. Mon- taner, I. Salazar and F. Rip011 for technical assistance.
REFERENCES 1.
2.
3.
4. 5. 6.
7.
8.
9.
10.
11.
12. 13. 14.
15.
16.
17. 18.
19.
20.
21.
Goldberg, A L & St John, A C, Ann rev biochem 45 (1976) 747. Nicoletti, M, Guerri, C & Grisolia, S, Eur j bio- them 75 (1977) 583. Katunuma, N, Kominami, E, Kobayashi, K, Ban- no, Y, Suzuki, K, Chichibu, K, Hamaguchi, Y & Katsunuma, T, Eur j biochem 52 (1975) 37. Wieland, 0 H, FEBS lett 52 (1975) 44. Rubio, V & Grisolia, S, FEBS lett 75 (1977) 281. Schatz, G & Mason, T L, Ann rev biochem 43 (1974) 51. Knecht, E & Hernandez, J, Cell tissue res 193 (1978) 473. Lowry, 0 H, Rosebrough, N J, Fat-r, A L & Ran- dall, R J, J biol them 193 (1951) 265. Salpeter, M M & McHenry, F A, Advanced tech- niques in biological electron microscopy. Analysis of autoradiograms (ed J K Koehler) p. 113. Springer Verlag, Heidelberg (1973). Weibel, E R &Bolender, R-P; Principles and tech- niques of electron microscopy. Biological applica- tions (ed M A Hayat) vol. 3, p. 237. Van Nostrand Reinhold Company, New York (1973). Kolb-Bachofen, V & Vogel, W, Exp cell res 94 (1975) 95. Pine, M J, Ann rev microbial 26 (1972) 103. Poole, B & Wibo, M, J biol them 248 (1973) 6221. Waterlow, J C, Garlick, P J & Millward, D J, Protein turnover in mammalian tissues and in the whole body. North-Holland Publishing Company, Amsterdam (1978). J$freys, A J & Craig, I W, Eur j biochem 68 (1976)
Seg’al, H I, Winkler, J R & Miyagi, M P, J biol them 249 (1974) 6364. Tanaka, K & Ichihara, A, Exp cell res 99 (1976) 1. Grille, T A I, Ogunnaike, P 0 & Faoye, S, J endocrinol51 (1971) 645. Kal’nov, S L, Serebryakova, NV, Zubatov, A S & Luzikov, V N, Biokhimiya SSSR 43 (1978) 662. Grisolia, S, Rivas, J, Wallace, R & Mendelson, J, Biochem biophys res commun 77 (1977) 367. Pfeiffer, U, Verh dtsch ges path 60 (1976) 28.
Received May 23, 1978 Revised version received April 26, 1979 Accepted May 22, 1979
Exp Ce//Res 125 (1980)
Related Documents











