ACTA ZOOLOGICA BULGARICA Acta zool. bulg., 63 (1), 2011: 17-35 Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo (SW Bulgaria) Zlatozar Boev National Museum of Natural History, Bulgarian Academy of Sciences, 1, Tsar Osvoboditel Blvd., 1000 Sofia, Bulgaria; E-mail: [email protected]; [email protected] Abstract: A new species of small falcon of the group ‘ tinnunculus’ is described on the base of 16 bone finds of an unarticulated skeleton of an adult individual, dated late Miocene (Turolian - Maeotian, lower part of the zone MN 11-12). Diagnosis: A medium-sized fossil species in the genus Falco differing from the closest F. tinnunculus by: (1) coracoid - much shorter f. a. clavicularis in cranial view; (2) humerus - relatively longer diaphysis; (3) ulna - wider distal fourth of depressio m. brachialis; (4) tibiotarsus - longer base (more proximally positioned inception) of crista cnemialis lateralis, and less protruding area interarticu- laris; (5) tarsometatarsus - relatively shorter diaphysis. It is suggested that the splitting of the genus Falco into two major groups, ‘ tinnunculus’ and ‘cherrug’occurred at least in Late Pliocene, and probably it may be an indication of the super-genera/genera rank. The ‘ tinnunculus’ group is of more ancient origin and it was diversified in late Miocene. The ‘ cherrug ’ group is not recorded in Neogene and the all Tertiary. Its record appears even in Early Pleistocene, while the ‘tinnunculus’ group is well documented in Neogene of Europe since Late Pliocene. Key words: Falcons, Fossil birds, Falconiformes, Late Miocene, Bulgaria Introduction Genus Falco comprises 37 recent species, 15 of them from Africa, Madagascar and adjacent islands, 11 - from South-East Asia, Australia and New Zealand, and 7 - from North and South Americas. Ten species occur both in Europe and Asia (White et al, 1994). Olson (1985) suggests an Afro-South-Asian origin of the genus, in spite of the present-day concentration of the species of family Falconidae in South America. Few data are known on the fossil history of falcons Falco Linnaseus, 1758. A summary of the fossil record of the family has been given in other paper (Boev 1999). A total of 10 fossil species were described in the genus Falco until now, although re- cently only seven of them are considered as valid spe- cies (Olson 1985, Bochenski 1997, Mlikovsky 1996, 2002) - (1) F. antiquus Mourer-Chauvire (1975) (Middle Pleistocene of Noailles, France); (2) F. me- dius Umanskaya, 1981 (late Miocene (MN 11-13) from S Ukraine; the only known falcon of Miocene both of Eurasia and Africa; the oldest record of genus Falco at all); (3) Falco umanskajae, Sobolev 2003 (late Pliocene (MN 16) from the vicinities of Odessa, Ukraine); (4) F. bakalovi Boev, 1999 (late Pliocene of Varshets, Bulgaria). Mlikovsky (2002) considered F. antiquus as a synonym of F. cherrug Gray, 1844, and listed only two fossil species of genus Falco: F. medius and F. bakalovi. (5) Falco chowi Hou Lianhai 1982 is known from Pleistocene of China. Two fal- cons have been described from the New World locali- ties - (6) F. oregonus Brodkorb, 1946 (Early/Middle Pliocene of Oregon), and (7) F. kurochkini Sufirez & Olson , 2001 (Late Pleistocene/Holocene of Cuba). 17

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ACTA ZOOLOGICA BULGARICA Acta zool. bulg., 63 (1), 2011: 17-35
Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo (SW Bulgaria)
Z latozar B oev
National Museum of Natural History, Bulgarian Academy of Sciences, 1, Tsar Osvoboditel Blvd., 1000 Sofia, Bulgaria; E-mail: [email protected]; [email protected]
Abstract: A new species of small falcon of the group ‘tinnunculus’ is described on the base of 16 bone finds of anunarticulated skeleton of an adult individual, dated late Miocene (Turolian - Maeotian, lower part of the zone MN 11-12). Diagnosis: A medium-sized fossil species in the genus Falco differing from the closest F. tinnunculus by: (1) coracoid - much shorter f. a. clavicularis in cranial view; (2) humerus - relatively longer diaphysis; (3) ulna - wider distal fourth of depressio m. brachialis; (4) tibiotarsus - longer base (more proximally positioned inception) of crista cnemialis lateralis, and less protruding area interarticu- laris; (5) tarsometatarsus - relatively shorter diaphysis. It is suggested that the splitting of the genus Falco into two major groups, ‘tinnunculus’ and ‘cherrug’ occurred at least in Late Pliocene, and probably it may be an indication of the super-genera/genera rank. The ‘tinnunculus’ group is of more ancient origin and it was diversified in late Miocene. The ‘cherrug ’ group is not recorded in Neogene and the all Tertiary. Its record appears even in Early Pleistocene, while the ‘tinnunculus’ group is well documented in Neogene of Europe since Late Pliocene.
Key words: Falcons, Fossil birds, Falconiformes, Late Miocene, Bulgaria
IntroductionGenus Falco comprises 37 recent species, 15 o f them from Africa, Madagascar and adjacent islands, 11 - from South-East Asia, Australia and New Zealand, and 7 - from North and South Americas. Ten species occur both in Europe and Asia (White et a l, 1994). Olson (1985) suggests an Afro-South-Asian origin of the genus, in spite of the present-day concentration of the species of family Falconidae in South America.
Few data are known on the fossil history of falcons Falco Linnaseus, 1758. A summary of the fossil record of the family has been given in other paper (Boev 1999). A total o f 10 fossil species were described in the genus Falco until now, although recently only seven of them are considered as valid species (Olson 1985, Bochenski 1997, Mlikovsky 1996, 2002) - (1) F. antiquus M ourer-Chauvire (1975)
(Middle Pleistocene o f Noailles, France); (2) F. me- dius Umanskaya, 1981 (late Miocene (MN 11-13) from S Ukraine; the only known falcon o f Miocene both of Eurasia and Africa; the oldest record of genus Falco at all); (3) Falco umanskajae, Sobolev 2003 (late Pliocene (MN 16) from the vicinities o f Odessa, Ukraine); (4) F. bakalovi Boev, 1999 (late Pliocene of Varshets, Bulgaria). Mlikovsky (2002) considered F. antiquus as a synonym of F. cherrug G ray, 1844, and listed only two fossil species of genus Falco: F. medius and F. bakalovi. (5) Falco chowi Hou Lianhai 1982 is known from Pleistocene o f China. Two falcons have been described from the New World localities - (6) F. oregonus Brodkorb, 1946 (Early/Middle Pliocene o f Oregon), and (7) F. kurochkini Sufirez & Olson , 2001 (Late Pleistocene/Holocene o f Cuba).
17

Boev Z.
Abbreviations
Anatomical: dex. - dextra; dig. - digitus, digiti; dist.- distalis; f. a. - facies articularis; max. - maximum; proc. - processus, processi; prox. - proximalis; sin- sinistra; s. a. - sulcus articularis; tbt - tibiotarsus; tmt - tarsometatarsus; Institutional: NHM - Natural History Museum, formerly British Museum (Natural History), Tring; ISEAK - Institute of Systematics and Evolution o f Animals (Polish Academy of Sciences), Krakow; NMNHS - National Museum of Natural History (Bulgarian Academy o f Sciences), Sofia.
Material and MethodsThe finds originate from the vicinity o f the town of Hadzhidimovo near the town o f Gotse Delchev (Blagoevgrad District; SW Bulgaria), Hadzhidimovo-1 locality, 41.30 N, 23.52 E; UTM grid: GM 30; 500 m a. s. l. They come from gray- yellowish-coloured sands and clay sands o f oblique and complex inner stratification at a depth o f 1.001.50 m (Dimitar Kovachev, NMNNS - pers. comm.). All the 16 bone finds (NoNo NMNHS 12539-12545; 12559-12567; Fig. 1, 2) represent parts o f an unarticulated skeleton o f one adult individual. It was collected by D. Kovachev in 1980s. Material is kept in the NMNHS. The findings have been identified through reference to comparative bird collections of the ISEAK, NHM and NMNHS.
Description o f the manner o f bone measuring in fossil and recent Falco: coracoid: a - thickness of processus acrocoracoideus; b - width o f processus acrocoracoideus; c - thickness o f diaphysis in the cranial end o f impressio m. sternocoracoidei on the cranial side; d - length o f f. a. humeralis (Table 1); scapula dex. prox.: a - width o f column scapulae beyond f. a. humeralis; b - thickness o f column scapulae beyond f. a. humeralis (Table 2); humerus: a - maximum length o f bone; b - length o f crista pec- toralis from tuberculum dorsale; c - length o f fossa brachialis; d - thickness o f caput humeri; e - height of caput humeri on lateral side; f - thickness o f the diaphysis at the foramen nutrinium; g - width of the diaphysis at the foramen nutrinium (Table 3); ulna: a - width in the middle o f the diaphysis on the medial side; b - maximum width o f proximal epiphysis; c - length o f depressio m. brachialis; d - thickness
of proximal epiphysis; e - length (transversal diameter) o f cotyla dorsalis; f - longitudinal diameter of cotyla ventralis (Table 4); radius: a - width o f proximal epiphysis; b - thickness o f proximal epiphysis; c - minimum width o f proximal half o f diaphysis; d - thickness o f epiphysis in tuberculum bicipitale (Table 5); femur: a - thickness o f diaphysis in the end o f its 1st (proximal) forth of its length; b - width of diaphysis in the same point (Table 6); tibiotarsus: a - length o f crista fibularis; b - thickness of the diaphysis in the distal end o f crista fibularis; c - width of diaphysis in the distal end o f crista fibularis; d - maximum diagonal width o f proximal epiphysis; e - length from the proximal epiphysis to the distal end of the tibio-fibular symphysis (at the caudal side); f - length o f foramen interosseum distale (Table 7); tarsometatarsus: a - width o f diaphysis in the proximal end o f the fossa metatarsi I; b - thickness of dia- physis in the proximal end o f the fossa metatarsi I; c - minimum width o f diaphysis (Table 8); phalanx 1 dig. I pedis: a - minimum width o f the ‘diaphysis’; b - width o f f. a. prox.; c - height o f f. a. prox. (Table 9); phalanx 2 dig. II pedis dex.: a - minimum width of the ‘diaphysis’; b - width o f f. a. prox.; c - height of f. a. prox. (Table 10); phalanx 2 dig. III pedis dex.: a - minimum width o f the ‘diaphysis’; b - width o f f. a. prox.; c - height of f. a. prox.; d - diameter of trochlea articularis (Table 11); phalanx 3 dig. III pedis dex. a - minimum width of the ‘diaphysis’; b - width of f. a. prox.; c - height o f f. a. prox. (Table 12).
All measurements have been taken using calipers to 0.05 mm accuracy, but read to the 1st digit after decimal point. All generic names o f the binomi- nals are given abbreviated in the text and are in full in the Appendix I. ‘Smaller’, ‘much smaller’, ‘bigger’ or ‘much bigger’ in ‘Comparison and discussion’ section mean that the fossil specimen differs considerably in size from the specimens o f compared species, and thus their taxonomic identity is excluded.
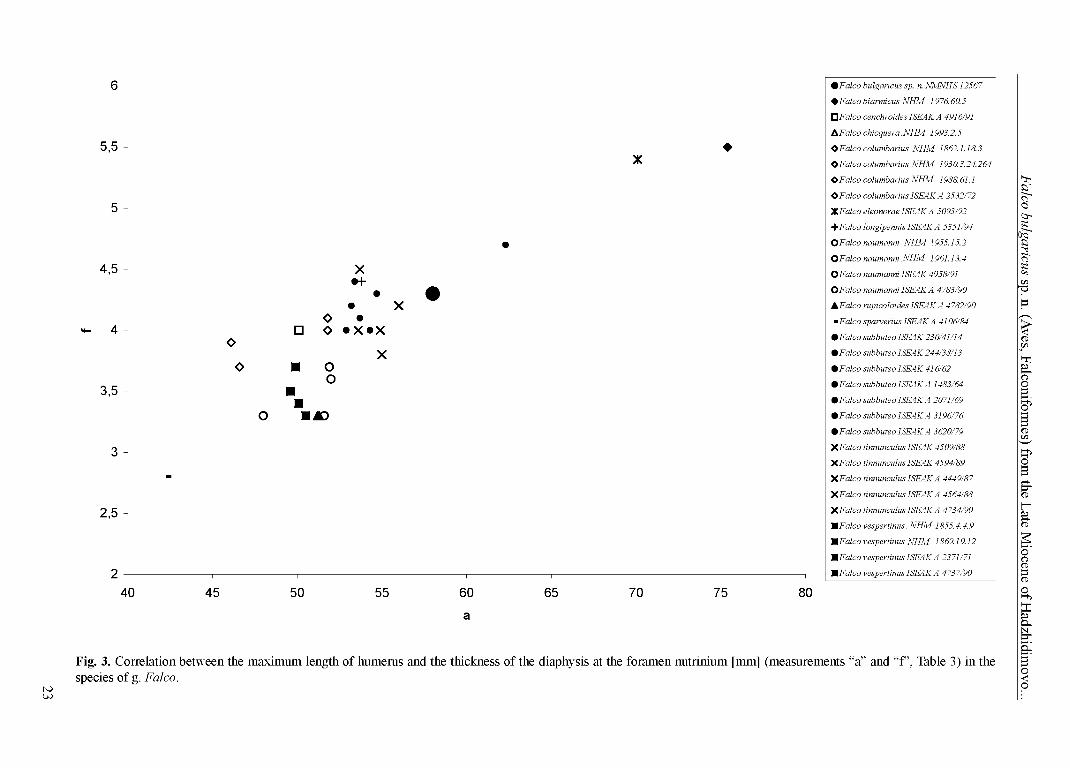
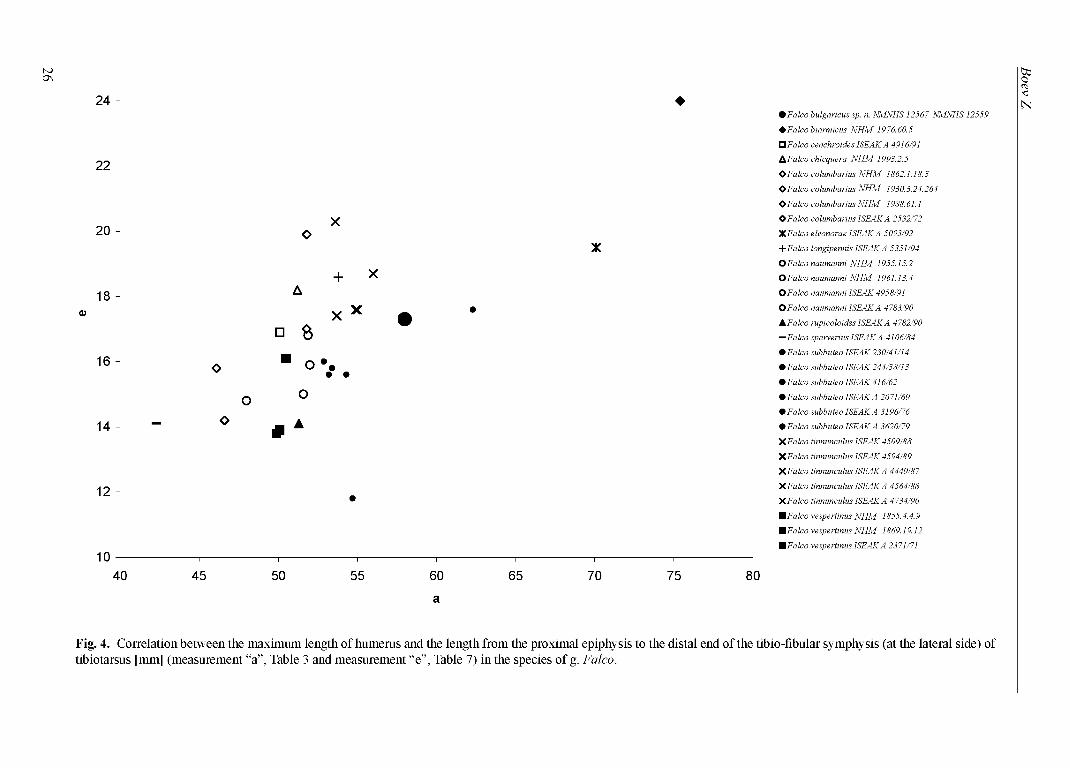
Two indices have been calculated with these measurements: (1) max. length o f humerus (Table 3 - a): thickness o f the diaphysis o f humerus at the foramen nutrinium (Table 3 - f) (Fig. 3); (2) max. length o f humerus (Table 3 - a): length from the proximal epiphysis to the distal end o f the tibiofibular symphysis (at the lateral side) o f tibiotarsus (Table 7 - e) (Fig. 4).
18
Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
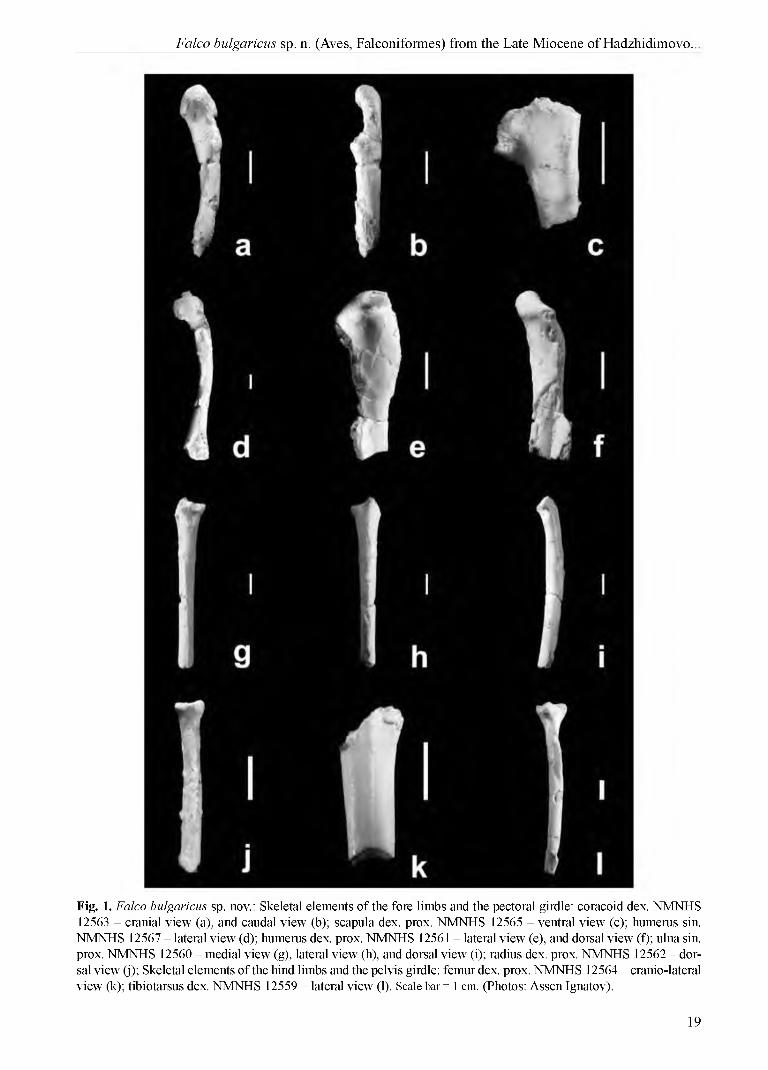
Fig. 1. Falco bulgaricus sp. nov.: Skeletal elements of the fore limbs and the pectoral girdle: coracoid dex. NMNHS 12563 - cranial view (a), and caudal view (b); scapula dex. prox. NMNHS 12565 - ventral view (c); humerus sin. NMNHS 12567 - lateral view (d); humerus dex. prox. NMNHS 12561 - lateral view (e), and dorsal view (f); ulna sin. prox. NMNHS 12560 - medial view (g), lateral view (h), and dorsal view (i); radius dex. prox. NMNHS 12562 - dorsal view (j); Skeletal elements of the hind limbs and the pelvis girdle: femur dex. prox. NMNHS 12564 - cranio-lateral view (k); tibiotarsus dex. NMNHS 12559 - lateral view (l). Scale bar = 1 cm. (Photos: Assen Ignatov).
19
Boev Z.
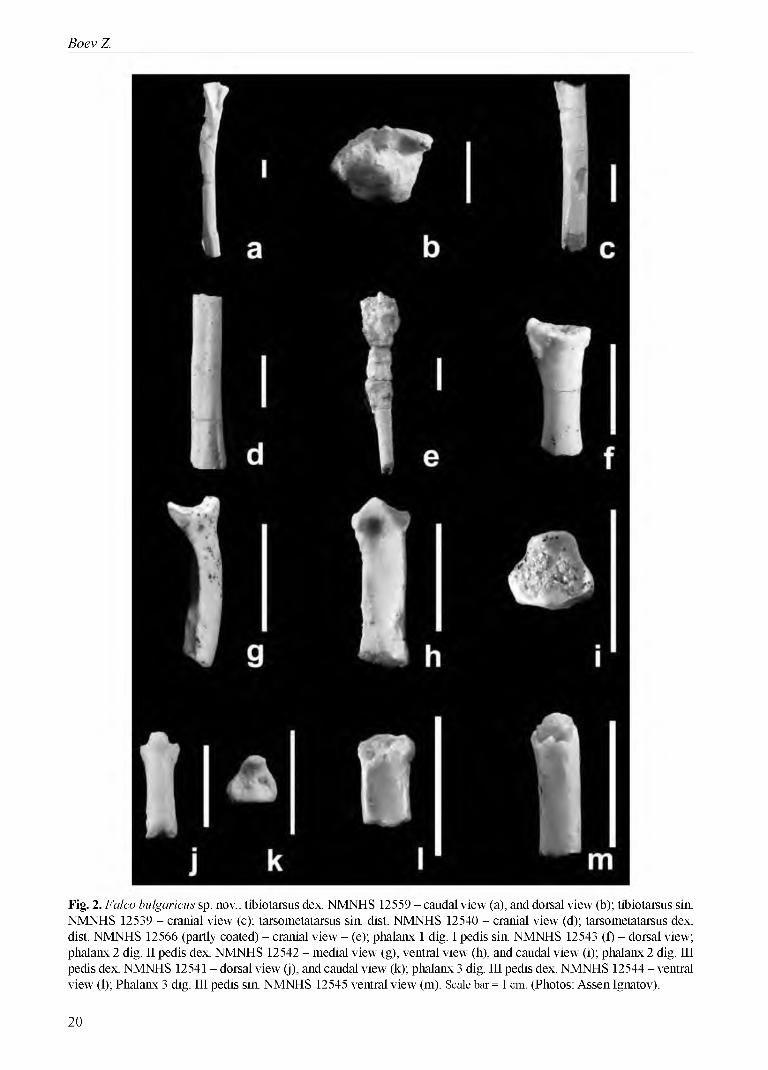
Fig. 2. Falco bulgaricus sp. nov.: tibiotarsus dex. NMNHS 12559 - caudal view (a), and dorsal view (b); tibiotarsus sin. NMNHS 12539 - cranial view (c); tarsometatarsus sin. dist. NMNHS 12540 - cranial view (d); tarsometatarsus dex. dist. NMNHS 12566 (partly coated) - cranial view - (e); phalanx 1 dig. I pedis sin. NMNHS 12543 (f) - dorsal view; phalanx 2 dig. II pedis dex. NMNHS 12542 - medial view (g), ventral view (h), and caudal view (i); phalanx 2 dig. III pedis dex. NMNHS 12541 - dorsal view (j), and caudal view (k); phalanx 3 dig. III pedis dex. NMNHS 12544 - ventral view (l); Phalanx 3 dig. III pedis sin. NMNHS 12545 ventral view (m). Scale bar = 1 cm. (Photos: Assen Ignatov).
20
Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
Table 1. Measurements of coracoid in fossil and recent Falco.
Species a b c dFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12563 3.4 5.7 3.8 ca. 5.1RecentFalco naumanni ISEAK A 4783/90 2.5 5.0 2.6 6.6Falco naumanni ISEAK A 4958/91 2.8 5.5 2.4 6.6Falco subbuteo ISEAK A 3196/76 2.8 6.0 2.9 7.2Falco subbuteo ISEAK A 2071/69 2.9 6.8 3.1 6.6Falco subbuteo ISEAK A 3620/79 2.8 6.4 3.0 6.8Falco subbuteo ISEAK 230/41/14 3.1 6.2 3.0 6.8Falco subbuteo ISEAK 244/38/13 3.0 5.8 2.8 7.3Falco subbuteo ISEAK 416/62 3.3 6.7 3.0 7.2Falco subbuteo ISEAK A 1483/64 2.8 5.8 2.8 6.5Falco tinnunculus ISEAK A 4449/87 2.7 6.0 2.6 5.8Falco tinnunculus ISEAK A 4734/90 2.7 6.2 2.9 6.5Falco tinnunculus ISEAK A 4564/88 3.0 6.3 3.9 6.8Falco tinnunculus ISEAK A 4594/89 2.9 6.7 2.9 6.1Falco tinnunculus ISEAK A 4509/88 2.8 6.5 3.1 6.1Falco vespertinus ISEAK A 5056/92 2.7 4.6 2.6 6.1Falco columbarius NMH 1988.61.1 2.8 6.9 4.0 6.3Falco columbarius ISEAK A 2532/72 2.6 5.9 2.7 6.9Falco eleonorae ISEAK A 5093/92 3.8 9.1 3.8 8.8Falco biarmicus NMH 1976.60.5 ca. 3.8 7.7 5.2 9.4Falco sparverius ISEAK A 4106/84 2.4 4.0 2.1 5.2Falco longipennis ISEAK A 5351/94 3.9 6.9 3.1 7.3Falco cenchroides ISEAK A 4916/91 2.7 4.7 2.7 5.6Falco rupicoloides ISEAK A 4782/90 2.6 5.8 2.7 6.4Falco chicquera NMH 1993.2.5 3.1 5.0 3.1 ca.5.3Falco columbarius NMH 1862.1.18.3 ca. 3.3 - 3.7 5.7Falco vespertinus NMH 1855.4.4.9 3.1 5.9 3.1 5.6Falco columbarius NMH 1930.3.24.264 2.7 5.7 3.1 5.9Falco naumanni NMH 1961.13.4 2.2 5.7 4.0 5.1Falco naumanni NMH 1955.15.2 2.6 6.3 3.9 6.0Falco vespertinus NMH 1869.19.12 2.7 4.6 2.5 4.9
The taxonomy follows W h i t e et al. (1994). The osteological terminology is after Ba u m e l & W i t m e r
(1993) and, in some respects, L i v e z e y & Zu s i , 2006. The chronostratigraphy follows Me i n (1990).
Short description of the siteAccording to the rich associated terrestrial megafauna this locality is dated back to the Late Miocene (Turolian - Maeotian, lower part o f the zone MN 11-12; ca. 7 million years ago). The associated land mammalian megafauna after Sp a s s o v (2002) includes 30 taxa: Mustelidae indet., Hyaenotheriun gr. wongii (Zd a n s k y , 1924), Hyaenictitheriini indet., cf. Miohyaenotherium bessarabicum Se m e n o v , 1989, Adcrocuta eximia (Ro t h a n d W a g n e r , 1854),
Table 2. Measurements of scapula prox. in fossil and recent Falco.
Species a bFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12565 4.0 2.5RecentFalco columbarius NMNHS 2/2002 3.8 2.0Falco columbarius NMNHS 1/1997 3.6 2.2Falco subbuteo NMNHS 1/1989 5.5 2.8Falco subbuteo NMNHS 3/1993 3.9 1.9Falco tinnunculus NMNHS 1/2002 2.8 1.8Falco tinnunculus NMNHS 7/1989 3.0 1.8Falco tinnunculus NMNHS 10/1992 4.0 2.4Falco vespertinus NMNHS 1/1991 2.8 2.0
21
Boev Z.
Table 3. Measurements of humerus in fossil and recent Falco.
Species a b c d e f gFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12561 - ca. 16.9 - 3.5 3.4 - -Falco bulgaricus sp. n. NMNHS 12567 ca. 58.0 ca. 17.0 6.7 - ca. 3.5 4.3 4.6RecentFalco biarmicus NMH 1976.60.5 75.4 23.9 10.9 5.6 5.6 5.5 6.0Falco cenchroides ISEAK A 4916/91 50.1 15.3 5.5 3.1 2.9 4.0 4.3Falco chicquera NMH 1993.2.5 51.2 16.0 5.1 3.4 3.7 3.3 4.2Falco columbarius NMH 1862.1.18.3 51.8 18.5 ca. 6.4 4.0 3.8 4.0 4.8Falco columbarius NMH 1930.3.24.264 46.1 15.0 6.6 3.5 3.4 3.9 4.4Falco columbarius NMH 1988.61.1 51.8 17.1 4.7 3.8 4.2 4.1 4.9Falco columbarius ISEAK A 2532/72 46.6 14.3 6.7 3.5 3.0 3.7 4.1Falco eleonorae ISEAK A 5093/92 70.1 19.9 7.9 5.3 5.0 5.4 6.2Falco longipennis ISEAK A 5351/94 53.8 17.7 6.6 4.1 3.6 4.4 5.0Falco naumanni NMH 1955.15.2 51.9 16.2 7.8 3.4 3.5 3.7 4.1Falco naumanni NMH 1961.13.4 48.0 8.9 7.0 2.9 3.3 3.3 3.5Falco naumanni ISEAK 4958/91 52.0 16.6 7.5 3.1 2.8 3.6 4.1Falco naumanni ISEAK A 4783/90 51.6 16.8 7.0 3.1 2.7 3.3 4.0Falco rupicoloides ISEAK A 4782/90 51.3 17.3 6.5 3.3 3.0 3.7 4.1Falco sparverius ISEAK A 4106/84 42.3 13.2 5.5 2.6 2.5 2.8 3.3Falco subbuteo ISEAK 230/41/14 53.4 16.5 6.4 3.9 3.7 4.4 4.8Falco subbuteo ISEAK 244/38/13 54.3 18.1 6.1 3.9 3.7 4.0 4.3Falco subbuteo ISEAK 416/62 62.3 20.0 6.8 4.3 3.8 4.7 5.2Falco subbuteo ISEAK A 1483/64 53.7 17.1 7.4 3.6 3.2 4.1 4.6Falco subbuteo ISEAK A 2071/69 53.2 16.9 6.6 4.0 3.5 4.2 4.8Falco subbuteo ISEAK A 3196/76 54.7 17.4 6.6 3.9 3.6 4.3 4.8Falco subbuteo ISEAK A 3620/79 52.9 17.6 7.9 4.3 3.5 4.0 4.6Falco tinnunculus ISEAK 4509/88 56.0 19.3 6.8 3.4 3.3 4.2 4.7Falco tinnunculus ISEAK 4594/89 53.6 17.2 7.3 3.3 3.1 4.0 4.1Falco tinnunculus ISEAK A 4449/87 55.0 19.4 7.6 3.3 2.9 3.8 4.2Falco tinnunculus ISEAK A 4564/88 53.7 18.4 7.7 3.1 3.0 4.5 4.1Falco tinnunculus ISEAK A 4734/90 54.9 18.9 7.0 3.4 3.2 4.0 4.5Falco vespertinus NMH 1855.4.4.9 50.5 18.4 7.0 3.3 3.8 3.3 3.8Falco vespertinus NMH 1869.19.12 50.1 16.5 6.6 2.9 3.1 3.4 3.8Falco vespertinus ISEAK A 2371/71 49.9 14.8 7.1 3.1 2.9 3.7 4.0Falco vespertinus ISEAK A 4737/90 49.6 16.7 6.7 3.1 2.8 3.5 3.8Falco vespertinus ISEAK A 5056/92 - 17.2 - 3.2 3.1 - -
Paramachairodus cf. orientalis gaudry, 1862,Machairodus giganteus (W agner, 1848),Metailurus cf. major Zdansky 1924, Gomphotheriidae indet., M ammut gr. borsoni, (Hays, 1834), Ancylotherium pentelicum (Gaudry & L a rte t , 1856),Chalicotheriinae indet., ? Ceratotherium neumayri (Osborn, 1900), Dicerorhinus pikermiensis G loger, 1841, Tapirus aff. jeanpiveteaui Boeuf, 1991, Hipparion gr. mediterraneum Roth & W agner,
1855, Hipparion cf. brachypus Hensel 1862, Hipparion ?platygenys Gromova, 1952,Microstonyx sp., Cervidae indet., Helladotherium duvernoyi (Gaudry & L arte t, 1856), Palaeotragus rouenii, Gazella (Procapra) sp., Palaeoryx sp., Palaeoreas lindermayeri W agner, 1848, Tragoportax sp. - l, Tragoportax sp. - 2, Hystrix primigenia, W agner, 1848) and Mesopithecus cf. delsoni Bonis & Bouvrain, G eraads & Koufos, 1990.
22
23
6 -I
5,5Ж
5
4,5
- 4
3,5
X
• Xо •о *х*х
оX
о ■ о о■
■О ■ JD
3
2,5 -
40 45 50 55 60
a
65 70 75 802
mFalco bulgaricus sp. n. NMNHS 12567
mFalco biarmicus N H M 1976.60.5
UFalco cenchroides ISEAK A 4916/91
OFalco chicqueraNHM 1993.2.5
OFalco columbarius N H M 1862.1.18.3
OFalco columbarius N H M 1930.3.24.264
OFalco columbarius N H M 1988.61.1
OFalco columbarius ISEAK A 2532/72
X Falco eleonorae ISEAK A 5093/92
+Falco longipennis ISEAK A 5351/94
OFalco naumanni N H M 1955.15.2
OFalco naumanni N H M 1961.13.4
OFalco naumanni ISEAK 4958/91
OFalco naumanni ISEAK A 4783/90
AFalco rupicoloides ISEAK A 4782/90
mFalco sparverius ISEAK A 4106/84
• Falco subbuteo ISEAK 230/41/14
• Falco subbuteo ISEAK 244/38/13
Ш Falco subbuteo ISEAK 416/62
• Falco subbuteo ISEAK A 1483/64
m Falco subbuteo ISEAK A 2071/69
m Falco subbuteo ISEAK A 3196/76
m Falco subbuteo ISEAK A 3620/79
X Falco tinnunculus ISEAK 4509/88
X Falco tinnunculus ISEAK 4594/89
OFalco tinnunculus ISEAK A 4449/87
X Falco tinnunculus ISEAK A 4564/88
XFalco tinnunculus ISEAK A 4734/90
MFalco vespertinus .N H M 1855.4.4.9
MFalco vespertinus .NHM 1869.19.12
M Falco vespertinus ISEAK A 2371/71
M Falco vespertinus ISEAK A 4737/90
Fig. 3. Correlation between the maximum length of humerus and the thickness of the diaphysis at the foramen nutrinium [mm] (measurements “a” and “f”, Table 3) in the species of g. Falco.
Falco bulgaricus
sp. n. (Aves, Falconiform
es) from the
Late M
iocene of H
adzhidimovo...
Boev Z.
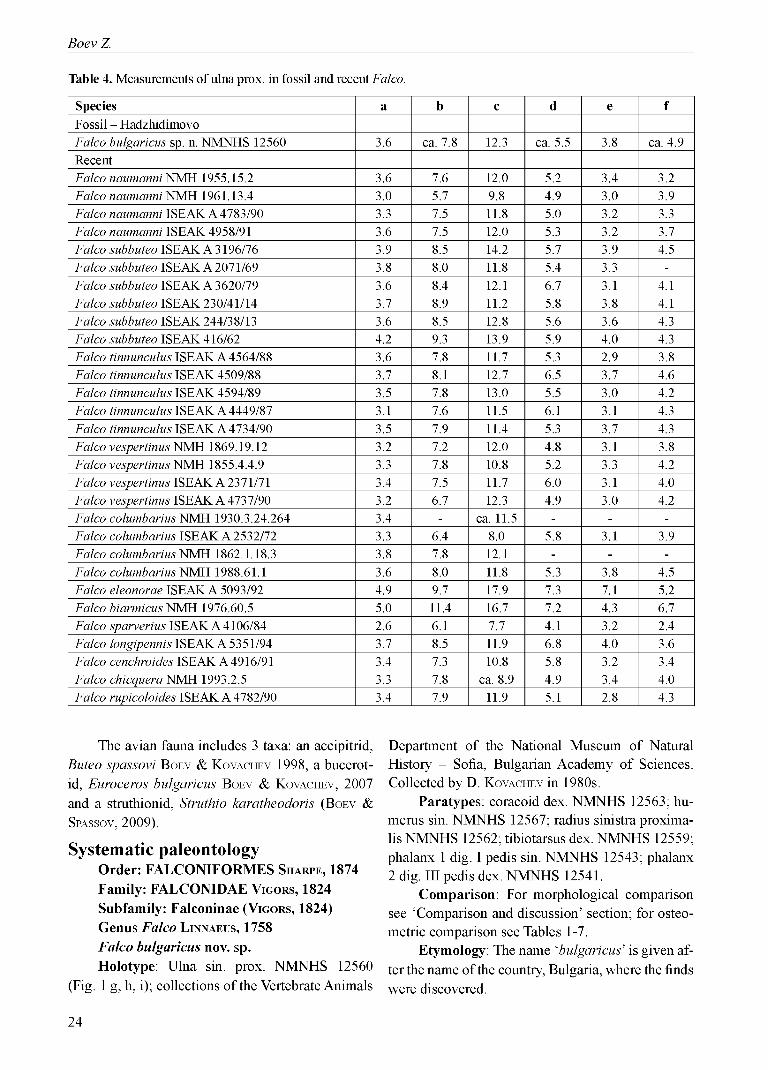
Table 4. Measurements of ulna prox. in fossil and recent Falco.
Species a b c d e fFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12560 3.6 ca. 7.8 12.3 ca. 5.5 3.8 ca. 4.9RecentFalco naumanni NMH 1955.15.2 3.6 7.6 12.0 5.2 3.4 3.2Falco naumanni NMH 1961.13.4 3.0 5.7 9.8 4.9 3.0 3.9Falco naumanni ISEAK A 4783/90 3.3 7.5 11.8 5.0 3.2 3.3Falco naumanni ISEAK 4958/91 3.6 7.5 12.0 5.3 3.2 3.7Falco subbuteo ISEAK A 3196/76 3.9 8.5 14.2 5.7 3.9 4.5Falco subbuteo ISEAK A 2071/69 3.8 8.0 11.8 5.4 3.3 -Falco subbuteo ISEAK A 3620/79 3.6 8.4 12.1 6.7 3.1 4.1Falco subbuteo ISEAK 230/41/14 3.7 8.9 11.2 5.8 3.8 4.1Falco subbuteo ISEAK 244/38/13 3.6 8.5 12.8 5.6 3.6 4.3Falco subbuteo ISEAK 416/62 4.2 9.3 13.9 5.9 4.0 4.3Falco tinnunculus ISEAK A 4564/88 3.6 7.8 11.7 5.3 2.9 3.8Falco tinnunculus ISEAK 4509/88 3.7 8.1 12.7 6.5 3.7 4.6Falco tinnunculus ISEAK 4594/89 3.5 7.8 13.0 5.5 3.0 4.2Falco tinnunculus ISEAK A 4449/87 3.1 7.6 11.5 6.1 3.1 4.3Falco tinnunculus ISEAK A 4734/90 3.5 7.9 11.4 5.3 3.7 4.3Falco vespertinus NMH 1869.19.12 3.2 7.2 12.0 4.8 3.1 3.8Falco vespertinus NMH 1855.4.4.9 3.3 7.8 10.8 5.2 3.3 4.2Falco vespertinus ISEAK A 2371/71 3.4 7.5 11.7 6.0 3.1 4.0Falco vespertinus ISEAK A 4737/90 3.2 6.7 12.3 4.9 3.0 4.2Falco columbarius NMH 1930.3.24.264 3.4 - ca. 11.5 - - -Falco columbarius ISEAK A 2532/72 3.3 6.4 8.0 5.8 3.1 3.9Falco columbarius NMH 1862.1.18.3 3.8 7.8 12.1 - - -Falco columbarius NMH 1988.61.1 3.6 8.0 11.8 5.3 3.8 4.5Falco eleonorae ISEAK A 5093/92 4.9 9.7 17.9 7.3 7.1 5.2Falco biarmicus NMH 1976.60.5 5.0 11.4 16.7 7.2 4.3 6.7Falco sparverius ISEAK A 4106/84 2.6 6.1 7.7 4.1 3.2 2.4Falco longipennis ISEAK A 5351/94 3.7 8.5 11.9 6.8 4.0 3.6Falco cenchroides ISEAK A 4916/91 3.4 7.3 10.8 5.8 3.2 3.4Falco chicquera NMH 1993.2.5 3.3 7.8 ca. 8.9 4.9 3.4 4.0Falco rupicoloides ISEAK A 4782/90 3.4 7.9 11.9 5.1 2.8 4.3
The avian fauna includes 3 taxa: an accipitrid,Buteo spassovi Bo e v & Ko v a c h e v 1998, a bucerot- id, Euroceros bulgaricus Bo e v & Ko v a c h e v , 2007 and a struthionid, Struthio karatheodoris (Bo e v & Sp a s s o v , 2009).
Systematic paleontologyOrder: FALCONIFORMES Sh a r p e , 1874 Family: FALCONIDAE Vi g o r s , 1824 Subfamily: Falconinae (Vi g o r s , 1824)Genus Falco Li n n a e u s , 1758 Falco bulgaricus nov. sp.Holotype: Ulna sin. prox. NMNHS 12560
(Fig. 1 g, h, i); collections o f the Vertebrate Animals
Department o f the National Museum of Natural History - Sofia, Bulgarian Academy o f Sciences. Collected by D. Ko v a c h e v in 1980s.
Paratypes: coracoid dex. NMNHS 12563; humerus sin. NMNHS 12567; radius sinistra proxima- lis NMNHS 12562; tibiotarsus dex. NMNHS 12559; phalanx 1 dig. I pedis sin. NMNHS 12543; phalanx 2 dig. III pedis dex. NMNHS 12541.
Comparison: For morphological comparison see ‘Comparison and discussion’ section; for osteo- metric comparison see Tables 1-7.
Etymology: The name ‘bulgaricus’ is given after the name o f the country, Bulgaria, where the finds were discovered.
24
Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
Table 5. Measurements of radius prox. in fossil and recent Falco.
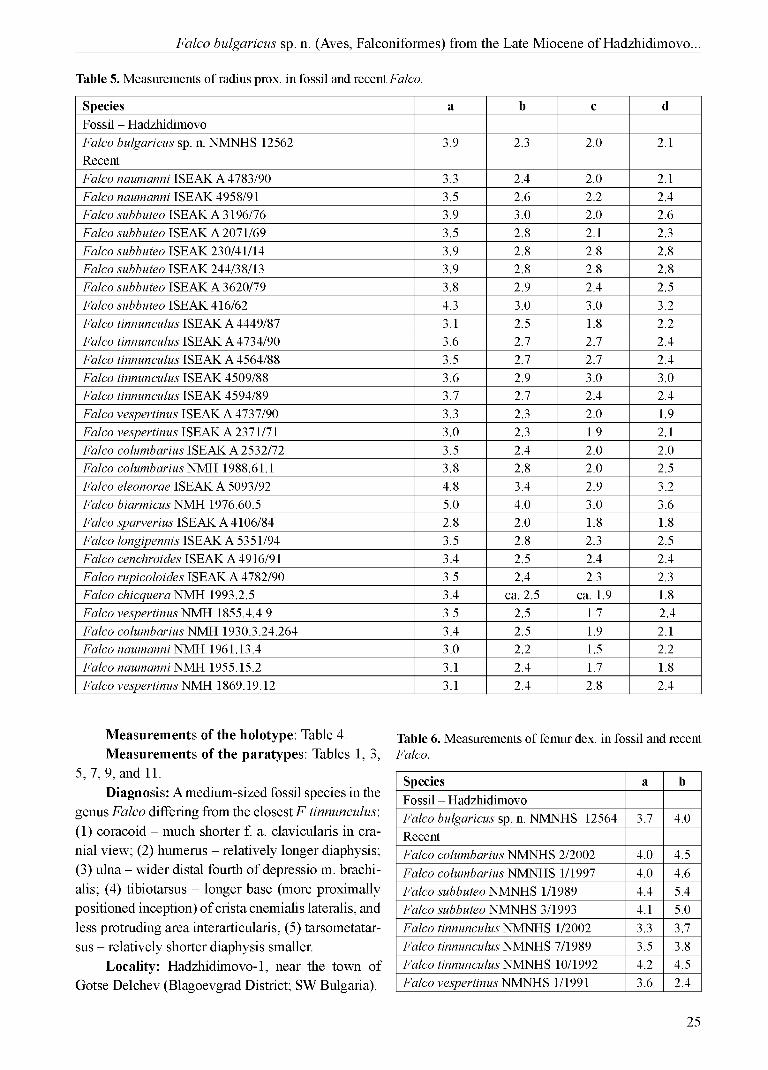
Species a b c dFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12562 3.9 2.3 2.0 2.1RecentFalco naumanni ISEAK A 4783/90 3.3 2.4 2.0 2.1Falco naumanni ISEAK 4958/91 3.5 2.6 2.2 2.4Falco subbuteo ISEAK A 3196/76 3.9 3.0 2.0 2.6Falco subbuteo ISEAK A 2071/69 3.5 2.8 2.1 2.3Falco subbuteo ISEAK 230/41/14 3.9 2.8 2.8 2.8Falco subbuteo ISEAK 244/38/13 3.9 2.8 2.8 2.8Falco subbuteo ISEAK A 3620/79 3.8 2.9 2.4 2.5Falco subbuteo ISEAK 416/62 4.3 3.0 3.0 3.2Falco tinnunculus ISEAK A 4449/87 3.1 2.5 1.8 2.2Falco tinnunculus ISEAK A 4734/90 3.6 2.7 2.7 2.4Falco tinnunculus ISEAK A 4564/88 3.5 2.7 2.7 2.4Falco tinnunculus ISEAK 4509/88 3.6 2.9 3.0 3.0Falco tinnunculus ISEAK 4594/89 3.7 2.7 2.4 2.4Falco vespertinus ISEAK A 4737/90 3.3 2.3 2.0 1.9Falco vespertinus ISEAK A 2371/71 3.0 2.3 1.9 2.1Falco columbarius ISEAK A 2532/72 3.5 2.4 2.0 2.0Falco columbarius NMH 1988.61.1 3.8 2.8 2.0 2.5Falco eleonorae ISEAK A 5093/92 4.8 3.4 2.9 3.2Falco biarmicus NMH 1976.60.5 5.0 4.0 3.0 3.6Falco sparverius ISEAK A 4106/84 2.8 2.0 1.8 1.8Falco longipennis ISEAK A 5351/94 3.5 2.8 2.3 2.5Falco cenchroides ISEAK A 4916/91 3.4 2.5 2.4 2.4Falco rupicoloides ISEAK A 4782/90 3.5 2.4 2.3 2.3Falco chicquera NMH 1993.2.5 3.4 ca. 2.5 ca. 1.9 1.8Falco vespertinus NMH 1855.4.4.9 3.5 2.5 1.7 2.4Falco columbarius NMH 1930.3.24.264 3.4 2.5 1.9 2.1Falco naumanni NMH 1961.13.4 3.0 2.2 1.5 2.2Falco naumanni NMH 1955.15.2 3.1 2.4 1.7 1.8Falco vespertinus NMH 1869.19.12 3.1 2.4 2.8 2.4
Measurements of the holotype: Table 4.Measurements of the paratypes: Tables 1, 3,
5, 7, 9, and 11.Diagnosis: A medium-sized fossil species in the
genus Falco differing from the closest F. tinnunculus: (1) coracoid - much shorter f. a. clavicularis in cranial view; (2) humerus - relatively longer diaphysis; (3) ulna - wider distal fourth o f depressio m. brachi- alis; (4) tibiotarsus - longer base (more proximally positioned inception) o f crista cnemialis lateralis, and less protruding area interarticularis; (5) tarsometatar- sus - relatively shorter diaphysis smaller.
Locality: Hadzhidimovo-1, near the town of Gotse Delchev (Blagoevgrad District; SW Bulgaria).
Table 6. Measurements of femur dex. in fossil and recentFalco.
Species a bFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12564 3.7 4.0RecentFalco columbarius NMNHS 2/2002 4.0 4.5Falco columbarius NMNHS 1/1997 4.0 4.6Falco subbuteo NMNHS 1/1989 4.4 5.4Falco subbuteo NMNHS 3/1993 4.1 5.0Falco tinnunculus NMNHS 1/2002 3.3 3.7Falco tinnunculus NMNHS 7/1989 3.5 3.8Falco tinnunculus NMNHS 10/1992 4.2 4.5Falco vespertinus NMNHS 1/1991 3.6 2.4
25
26
24 -
22 -
20 -
18 -
16 -
14
12 -
1040
X
ж+ X
X
□ еX
1 О » .
о
А
45 50 55 60
a
65 70 75
m Falco bulgaricus sp. n. NMNHS 12567 NMNHS 12559
OFalco biarmicus N H M 1976.60.5
UFalco cenchroides ISEAK A 4916/91
faFalco chicquera N H M 1993.2.5
OFalco columbarius N H M 1862.1.18.3
OFalco columbarius N H M 1930.3.24.264
OFalco columbariusNMM 1988.61.1
UFalco columbarius ISEAK A 2532/72
XFalco eleonorae ISEAK A 5093/92
+ Falco longipennis ISEAK A 5351/94
OFalco naumanni N H M 1955.15.2
OFalco naumanni N H M 1961.13.4
OFalco naumanni ISEAK 4958/91
OFalco naumanni ISEAK A 4783/90
OFalco rupicoloides ISEAK A 4782/90
—Falco sparverius ISEAK A 4106/84
Falco subbuteo ISEAK 230/41/14
Falco subbuteo ISEAK 244/38/13
Falco subbuteo ISEAK 416/62
Falco subbuteo ISEAK A 2071/69
Falco subbuteo ISEAK A 3196/76
Falco subbuteo ISEAK A 3620/79
XFalco tinnunculus ISEAK 4509/88
XFalco tinnunculus ISEAK 4594/89
XFalco tinnunculus ISEAK A 4449/87
OFalco tinnunculus ISEAK A 4564/88
Y,Falco tinnunculus ISEAK A 4734/90
MFalco vespertinus N H M ' 1855.4.4.9
MFalco vespertinus N H M 1869.19.12
M Falco vespertinus ISEAK A 2371/71
80
Fig. 4. Correlation between the maximum length of humerus and the length from the proximal epiphysis to the distal end of the tibio-fibular symphysis (at the lateral side) of tibiotarsus [mm] (measurement “a”, Table 3 and measurement “e”, Table 7) in the species of g. Falco.
Boev Z.
Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
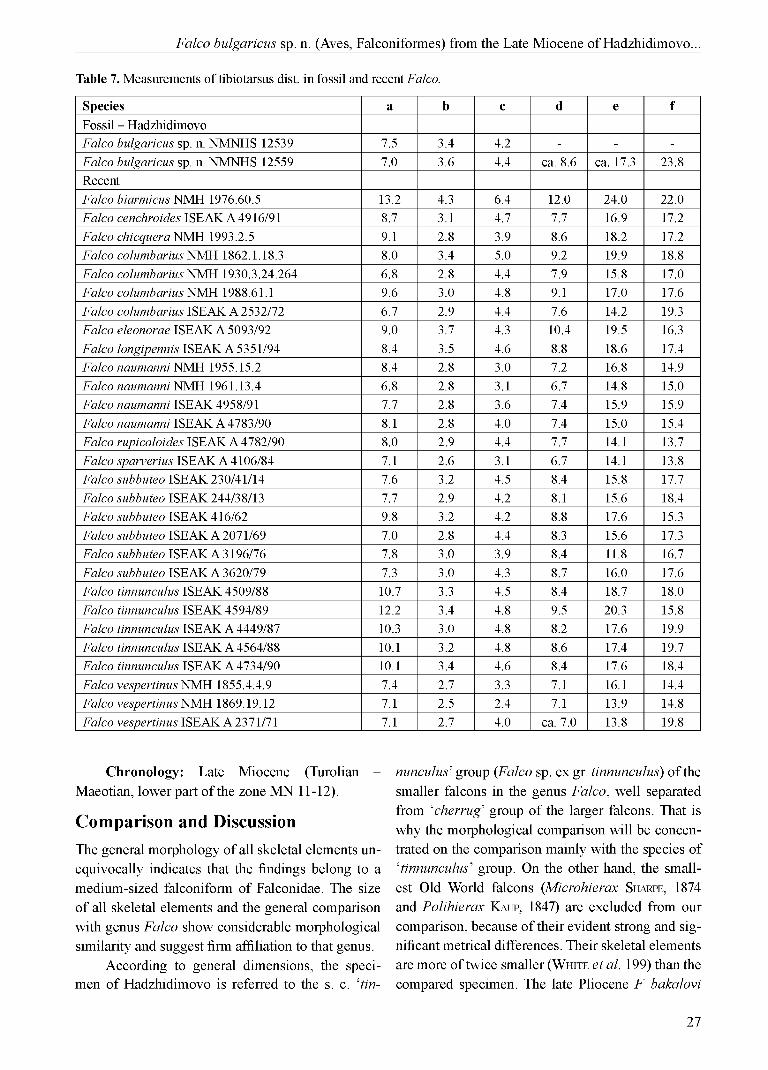
Table 7. Measurements of tibiotarsus dist. in fossil and recent Falco.
Species a b c d e fFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12539 7.5 3.4 4.2 - - -Falco bulgaricus sp. n. NMNHS 12559 7.0 3.6 4.4 ca. 8.6 ca. 17.3 23.8RecentFalco biarmicus NMH 1976.60.5 13.2 4.3 6.4 12.0 24.0 22.0Falco cenchroides ISEAK A 4916/91 8.7 3.1 4.7 7.7 16.9 17.2Falco chicquera NMH 1993.2.5 9.1 2.8 3.9 8.6 18.2 17.2Falco columbarius NMH 1862.1.18.3 8.0 3.4 5.0 9.2 19.9 18.8Falco columbarius NMH 1930.3.24.264 6.8 2.8 4.4 7.9 15.8 17.0Falco columbarius NMH 1988.61.1 9.6 3.0 4.8 9.1 17.0 17.6Falco columbarius ISEAK A 2532/72 6.7 2.9 4.4 7.6 14.2 19.3Falco eleonorae ISEAK A 5093/92 9.0 3.7 4.3 10.4 19.5 16.3Falco longipennis ISEAK A 5351/94 8.4 3.5 4.6 8.8 18.6 17.4Falco naumanni NMH 1955.15.2 8.4 2.8 3.0 7.2 16.8 14.9Falco naumanni NMH 1961.13.4 6.8 2.8 3.1 6.7 14.8 15.0Falco naumanni ISEAK 4958/91 7.7 2.8 3.6 7.4 15.9 15.9Falco naumanni ISEAK A 4783/90 8.1 2.8 4.0 7.4 15.0 15.4Falco rupicoloides ISEAK A 4782/90 8.0 2.9 4.4 7.7 14.1 13.7Falco sparverius ISEAK A 4106/84 7.1 2.6 3.1 6.7 14.1 13.8Falco subbuteo ISEAK 230/41/14 7.6 3.2 4.5 8.4 15.8 17.7Falco subbuteo ISEAK 244/38/13 7.7 2.9 4.2 8.1 15.6 18.4Falco subbuteo ISEAK 416/62 9.8 3.2 4.2 8.8 17.6 15.3Falco subbuteo ISEAK A 2071/69 7.0 2.8 4.4 8.3 15.6 17.3Falco subbuteo ISEAK A 3196/76 7.8 3.0 3.9 8.4 11.8 16.7Falco subbuteo ISEAK A 3620/79 7.3 3.0 4.3 8.7 16.0 17.6Falco tinnunculus ISEAK 4509/88 10.7 3.3 4.5 8.4 18.7 18.0Falco tinnunculus ISEAK 4594/89 12.2 3.4 4.8 9.5 20.3 15.8Falco tinnunculus ISEAK A 4449/87 10.3 3.0 4.8 8.2 17.6 19.9Falco tinnunculus ISEAK A 4564/88 10.1 3.2 4.8 8.6 17.4 19.7Falco tinnunculus ISEAK A 4734/90 10.1 3.4 4.6 8.4 17.6 18.4Falco vespertinus NMH 1855.4.4.9 7.4 2.7 3.3 7.1 16.1 14.4Falco vespertinus NMH 1869.19.12 7.1 2.5 2.4 7.1 13.9 14.8Falco vespertinus ISEAK A 2371/71 7.1 2.7 4.0 ca. 7.0 13.8 19.8
Chronology: Late Miocene (Turolian -Maeotian, lower part o f the zone MN 11-12).
Comparison and DiscussionThe general morphology of all skeletal elements unequivocally indicates that the findings belong to a medium-sized falconiform of Falconidae. The size o f all skeletal elements and the general comparison with genus Falco show considerable morphological similarity and suggest firm affiliation to that genus.
According to general dimensions, the specimen o f Hadzhidimovo is referred to the s. c. ‘tin-
nunculus’ group (Falco sp. ex gr. tinnunculus) o f the smaller falcons in the genus Falco, well separated from ‘cherrug’ group o f the larger falcons. That is why the morphological comparison will be concentrated on the comparison mainly with the species of ‘tinnunculus’ group. On the other hand, the smallest Old World falcons (Microhierax Sh a r p e , 1874 and Polihierax Ka u p , 1847) are excluded from our comparison, because o f their evident strong and significant metrical differences. Their skeletal elements are more o f twice smaller (Wh i t e et al. 199) than the compared specimen. The late Pliocene F. bakalovi
27
Boev Z.
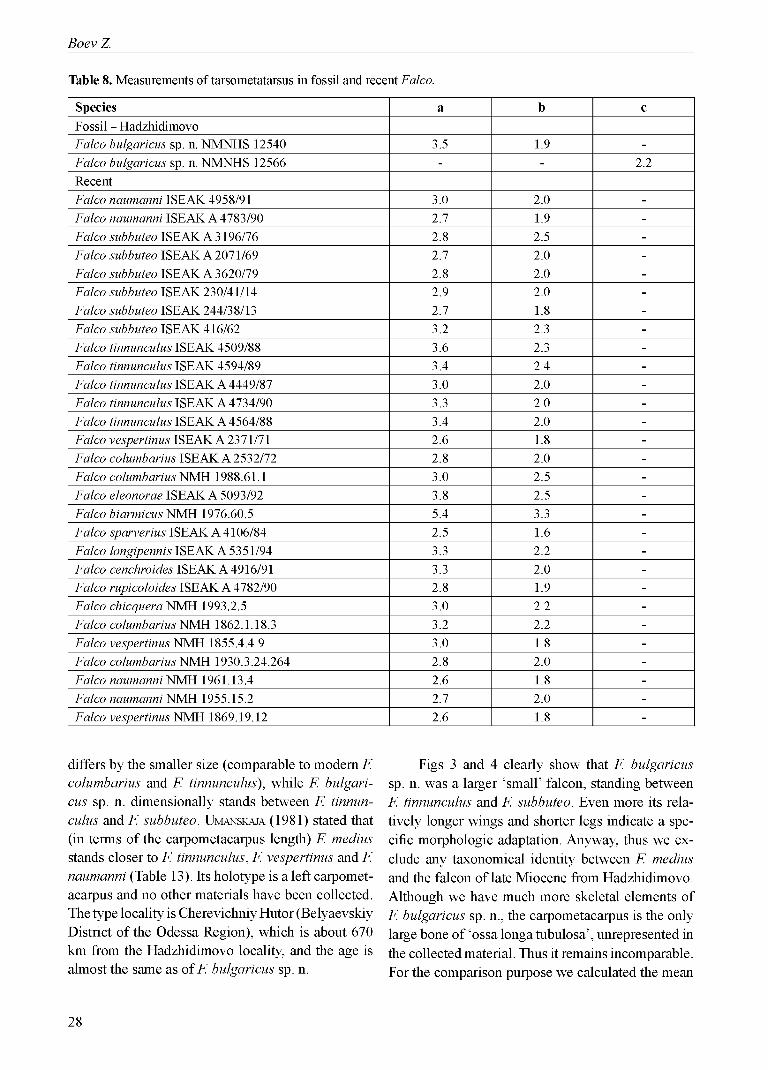
Table 8. Measurements of tarsometatarsus in fossil and recent Falco.
Species a b cFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12540 3.5 1.9 -Falco bulgaricus sp. n. NMNHS 12566 - - 2.2RecentFalco naumanni ISEAK 4958/91 3.0 2.0 -Falco naumanni ISEAK A 4783/90 2.7 1.9 -Falco subbuteo ISEAK A 3196/76 2.8 2.5 -Falco subbuteo ISEAK A 2071/69 2.7 2.0 -Falco subbuteo ISEAK A 3620/79 2.8 2.0 -Falco subbuteo ISEAK 230/41/14 2.9 2.0 -Falco subbuteo ISEAK 244/38/13 2.7 1.8 -Falco subbuteo ISEAK 416/62 3.2 2.3 -Falco tinnunculus ISEAK 4509/88 3.6 2.3 -Falco tinnunculus ISEAK 4594/89 3.4 2.4 -Falco tinnunculus ISEAK A 4449/87 3.0 2.0 -Falco tinnunculus ISEAK A 4734/90 3.3 2.0 -Falco tinnunculus ISEAK A 4564/88 3.4 2.0 -Falco vespertinus ISEAK A 2371/71 2.6 1.8 -Falco columbarius ISEAK A 2532/72 2.8 2.0 -Falco columbarius NMH 1988.61.1 3.0 2.5 -Falco eleonorae ISEAK A 5093/92 3.8 2.5 -Falco biarmicus NMH 1976.60.5 5.4 3.3 -Falco sparverius ISEAK A 4106/84 2.5 1.6 -Falco longipennis ISEAK A 5351/94 3.3 2.2 -Falco cenchroides ISEAK A 4916/91 3.3 2.0 -Falco rupicoloides ISEAK A 4782/90 2.8 1.9 -Falco chicquera NMH 1993.2.5 3.0 2.2 -Falco columbarius NMH 1862.1.18.3 3.2 2.2 -Falco vespertinus NMH 1855.4.4.9 3.0 1.8 -Falco columbarius NMH 1930.3.24.264 2.8 2.0 -Falco naumanni NMH 1961.13.4 2.6 1.8 -Falco naumanni NMH 1955.15.2 2.7 2.0 -Falco vespertinus NMH 1869.19.12 2.6 1.8 -
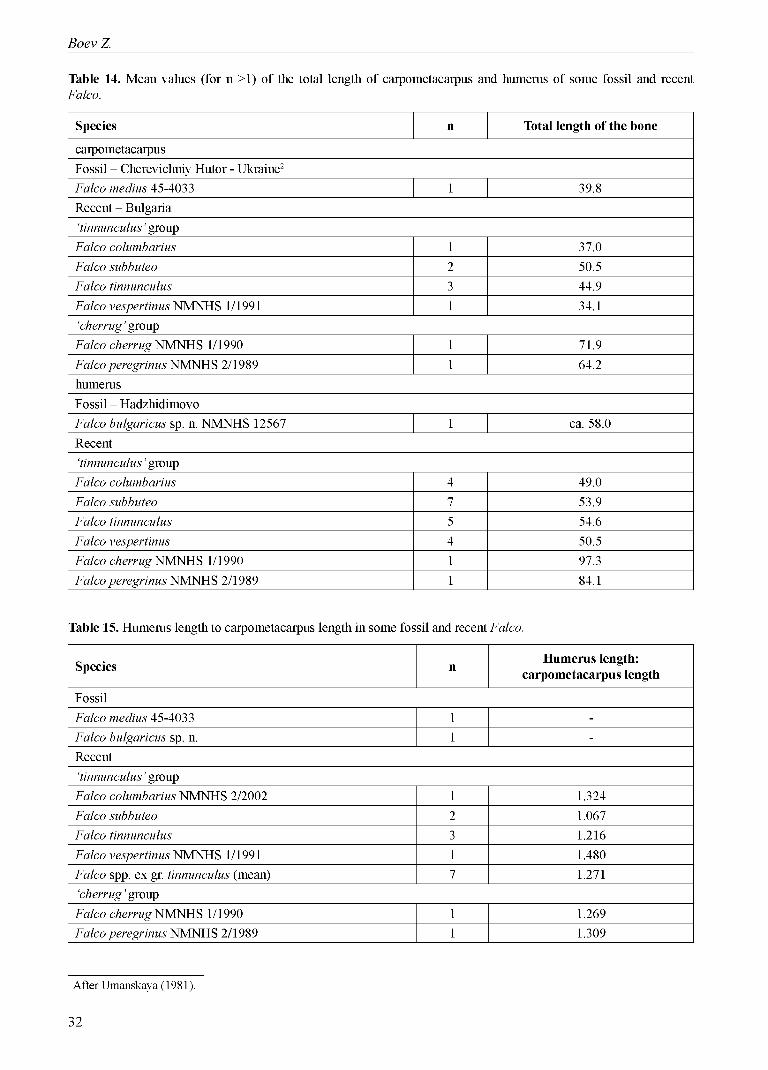
differs by the smaller size (comparable to modern F. columbarius and F. tinnunculus), while F. bulgaricus sp. n. dimensionally stands between F. tinnunculus and F. subbuteo. Um a n s k a j a (1981) stated that (in terms o f the carpometacarpus length) F. medius stands closer to F. tinnunculus, F. vespertinus and F. naumanni (Table 13). Its holotype is a left carpomet- acarpus and no other materials have been collected. The type locality is Cherevichniy Hutor (Belyaevskiy District o f the Odessa Region), which is about 670 km from the Hadzhidimovo locality, and the age is almost the same as o f F. bulgaricus sp. n.
Figs 3 and 4 clearly show that F. bulgaricus sp. n. was a larger ‘small’ falcon, standing between F. tinnunculus and F. subbuteo. Even more its relatively longer wings and shorter legs indicate a specific morphologic adaptation. Anyway, thus we exclude any taxonomical identity between F. medius and the falcon of late Miocene from Hadzhidimovo. Although we have much more skeletal elements of F. bulgaricus sp. n., the carpometacarpus is the only large bone o f ‘ossa longa tubulosa’, unrepresented in the collected material. Thus it remains incomparable. For the comparison purpose we calculated the mean
28

Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
values o f both carpometacarpus and humerus for the falcons (Table 14) and the ratio between the humerus length and the carpometacarpus length (Table 15). Thus the extraplolated humerus length o f Falco me- dius using the mean proportion (1.271) for ‘tinnun- culus’ group is ca. 50.6 mm, while the extrapolated carpometacarpus length o f Falco bulgaricus sp. n. using the same proportion is ca. 45.6 mm.
Skeletal elements of fore limbs and pectoral girdle
Coracoid dex. NMNHS 12563 (Fig. 1 - a, b; Table 1). F. biarmicus: much smaller size, and the relatively narrower humeral part of the bone; F. cenchroides: considerably larger size; F. rupi- coloides: larger size; F. chicquera: similar in size and general morphology; F. columbarius: thicker pr. acrocoracoideus (measurement ‘c’), thinner diaphysis (measurement ‘d ’); F. longipennis: thicker proc. acrocoracoideus (measurement ‘a ’); F. naumanni: larger, wider s. m. supracoracoidei, and less rounded proc. acrocoracoideus; F. subbuteo: thicker pr. acrocoracoideus (measurement ‘a ’) and almost twice wider impressio ligamenti acrocoracohumeralis; F. tinnunculus: similar both in general morphology and size, but sharper proc. acrocoracoideus, better developed longitudinal groove on the supracoracoidal surface, and much shorter f. a. clavicularis in cranial view; F. vespertinus: bigger, and the thicker proc. ac- rocoracoideus.
Scapula dex. prox. NMNHS 12565 (Fig. 1 - c; Table 2). F. columbarius: bigger and much thicker; F. subbuteo: thicker margo dorsalis at the area of collum scapulae; F. tinnununculus: more robust; F. vespertinus: bigger and more robust.
Humerus sin. NMNHS 12567 (Fig. 1 - d; Table 3) and humerus dex. prox. NMNHS 12561 (Fig. 1 - e, f; Table 3). F. biarmicus: resembles by the position o f the foramen nutritium, and by the presence o f two well developed transversal and parallel grooves on the lateral side o f the bone between the intumescentia, but differs by the much smaller size; F. chicquera: longer humerus, and the more proximally positioned foramen nutritium; F. colum- barius: considerably longer humeral bone and more proximally, instead distally, positioned foramen nu- tritium on the diaphysis. (The correlation between the measurements ‘a ’ and ‘b ’ in the fossil specimen
is 12.8, against 14.1 in F. columbarius); F. eleono- rae: much smaller; F. longipennis: longer and thinner diaphysis - the clearest difference; F. naumanni: larger, deeper foramen nutritium, deeper fossa m. brachialis, and longer humeral bone; F. sparverius: too smaller; F. tinnunculus: clearly differs in proportions as its diaphysis is relatively longer. Foramen nu- tritium is situated much more distally. Nevertheless most o f the details are similar, confirming its referring to ‘ tinnunculus’ group o f genus Falco; F. ves- pertinus: considerably larger size, the presence of two well developed transversal and parallel grooves (instead one hardly seen) on the lateral side o f the bone between the intumescentia and the neighboring narrower diaphysal parts (they are not represented in all recent small falcons /!/), and the longer and thinner diaphysis; F. subbuteo: smaller, more S-shape bent instead straight in dorsal view.
Ulna sin. prox. NMNHS 12560 (Fig. 1 - g, h, i; Table 4). F. biarmicus: smaller, and deeper depres- sio m. brachialis; F. cenchroides: larger, wider cotyla dorsalis; F. chicquera: larger, and deeper depressio m. brachialis; F. columbarius: more proximally positioned foramen nutritium is the only difference; F. eleonorae: much smaller; F. longipennis: smaller; F. sparverius: considerably larger; F. subbuteo: more proximally situated tuberculum bicipitale, and sharper linea intermuscularis; F tinnunculus: similar, but wider distal fourth o f depressio m. brachialis; F vespertinus: better marked longitudinal linea intermuscularis; Falco naumanni: larger, deeper fossa m. brachialis (esp. in its proximal end), and better developed tuberculum lig. collateralis ventralis.
Radius dex. prox. NMNHS 12562 (Fig. 1 - j; Table 5). F biarmicus: much smaller, better developed margo interosseus; F chicquera: slightly larger; F tinnunculus: shallower f. a. ulnaris; F ves- pertinus: larger and bigger tuberculum bicipitale; Falco columbarius: bigger tuberculum bicipitale; F subbuteo: slightly smaller; F tinnunculus: more protruding f. a. ulnaris.
Skeletal elements of the hind limbs and the pelvis
girdleFemur dex. prox. NMNHS 12564 (Fig. 1 - k;
Table 6). F columbarius: better developed linea intermuscularis cranialis; F subbuteo: slightly smaller and less developed impressiones iliotrochantericae;
29
Boev Z.
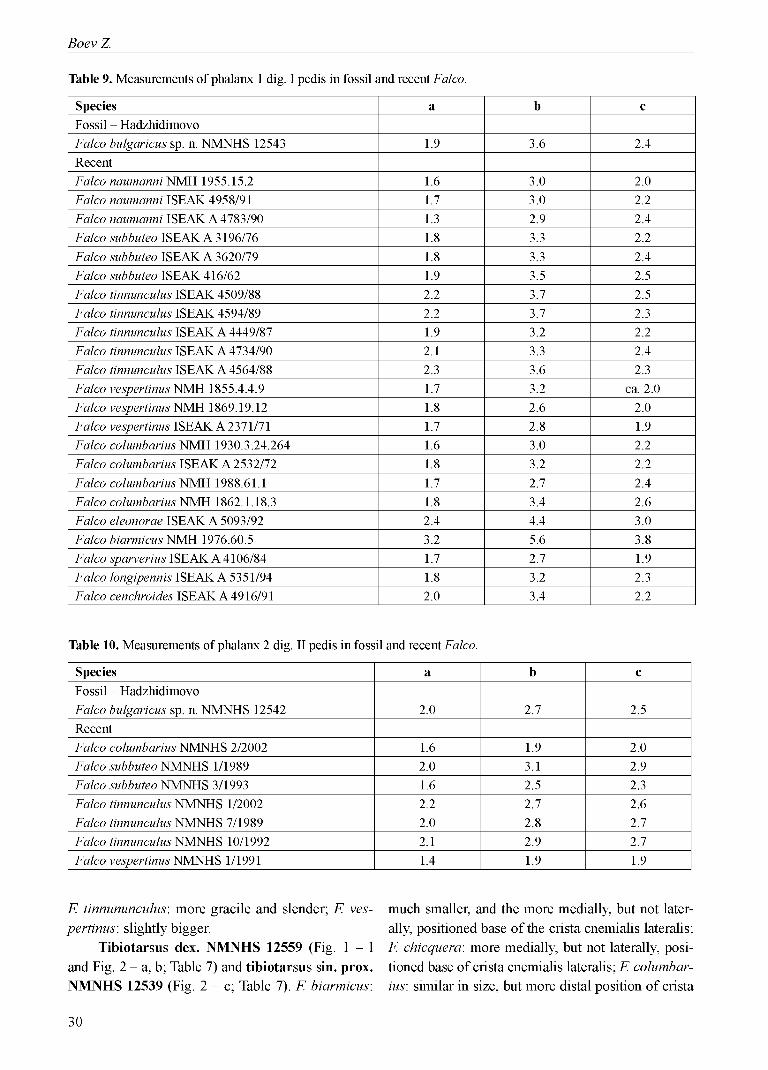
Table 9. Measurements of phalanx 1 dig. I pedis in fossil and recent Falco.
Species a b cFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12543 1.9 3.6 2.4RecentFalco naumanni NMH 1955.15.2 1.6 3.0 2.0Falco naumanni ISEAK 4958/91 1.7 3.0 2.2Falco naumanni ISEAK A 4783/90 1.3 2.9 2.4Falco subbuteo ISEAK A 3196/76 1.8 3.3 2.2Falco subbuteo ISEAK A 3620/79 1.8 3.3 2.4Falco subbuteo ISEAK 416/62 1.9 3.5 2.5Falco tinnunculus ISEAK 4509/88 2.2 3.7 2.5Falco tinnunculus ISEAK 4594/89 2.2 3.7 2.3Falco tinnunculus ISEAK A 4449/87 1.9 3.2 2.2Falco tinnunculus ISEAK A 4734/90 2.1 3.3 2.4Falco tinnunculus ISEAK A 4564/88 2.3 3.6 2.3Falco vespertinus NMH 1855.4.4.9 1.7 3.2 ca. 2.0Falco vespertinus NMH 1869.19.12 1.8 2.6 2.0Falco vespertinus ISEAK A 2371/71 1.7 2.8 1.9Falco columbarius NMH 1930.3.24.264 1.6 3.0 2.2Falco columbarius ISEAK A 2532/72 1.8 3.2 2.2Falco columbarius NMH 1988.61.1 1.7 2.7 2.4Falco columbarius NMH 1862.1.18.3 1.8 3.4 2.6Falco eleonorae ISEAK A 5093/92 2.4 4.4 3.0Falco biarmicus NMH 1976.60.5 3.2 5.6 3.8Falco sparverius ISEAK A 4106/84 1.7 2.7 1.9Falco longipennis ISEAK A 5351/94 1.8 3.2 2.3Falco cenchroides ISEAK A 4916/91 2.0 3.4 2.2
Table 10. Measurements of phalanx 2 dig. II pedis in fossil and recent Falco.
Species a b cFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12542 2.0 2.7 2.5RecentFalco columbarius NMNHS 2/2002 1.6 1.9 2.0Falco subbuteo NMNHS 1/1989 2.0 3.1 2.9Falco subbuteo NMNHS 3/1993 1.6 2.5 2.3Falco tinnunculus NMNHS 1/2002 2.2 2.7 2.6Falco tinnunculus NMNHS 7/1989 2.0 2.8 2.7Falco tinnunculus NMNHS 10/1992 2.1 2.9 2.7Falco vespertinus NMNHS 1/1991 1.4 1.9 1.9
F. tinnununculus: more gracile and slender; F. vespertinus: slightly bigger.
Tibiotarsus dex. NMNHS 12559 (Fig. 1 - l and Fig. 2 - a, b; Table 7) and tibiotarsus sin. prox. NMNHS 12539 (Fig. 2 - c; Table 7). F. biarmicus:
much smaller, and the more medially, but not laterally, positioned base o f the crista cnemialis lateralis; F. chicquera: more medially, but not laterally, positioned base o f crista cnemialis lateralis; F. columbarius: similar in size, but more distal position o f crista
30
Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
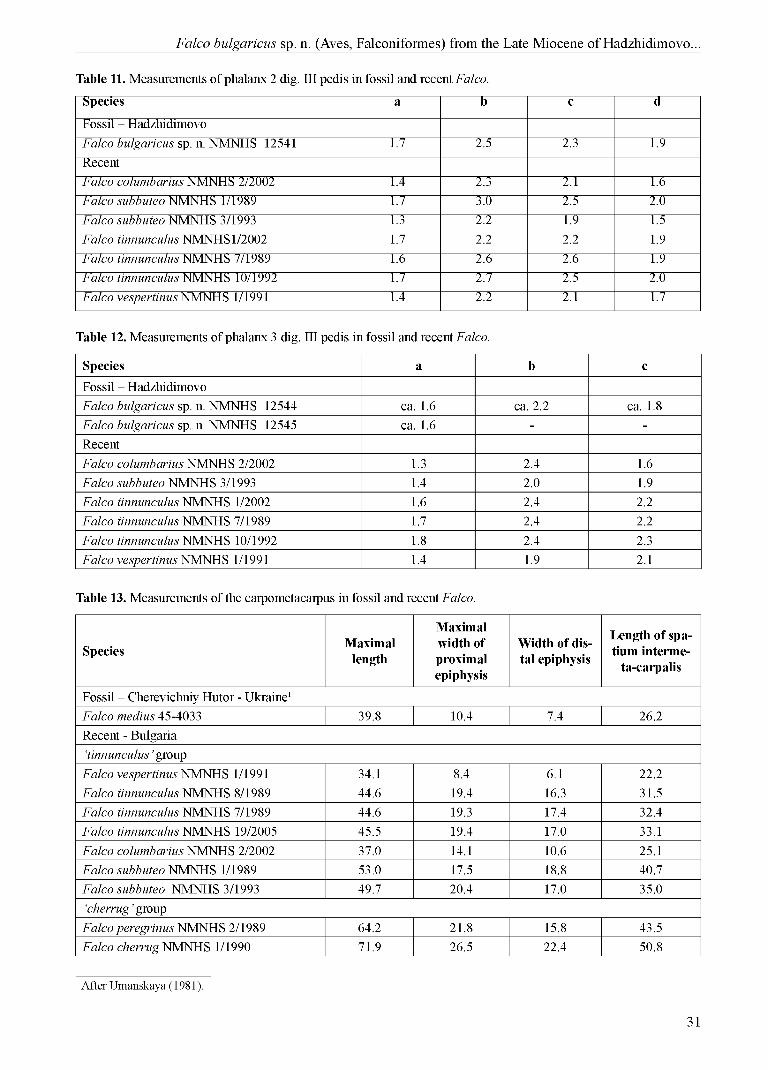
Table 11. Measurements of phalanx 2 dig. III pedis in fossil and recent Falco.
Species a b c d
Fossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12541 1.7 2.5 2.3 1.9RecentFalco columbarius NMNHS 2/2002 1.4 2.3 2.1 1.6Falco subbuteo NMNHS 1/1989 1.7 3.0 2.5 2.0Falco subbuteo NMNHS 3/1993 1.3 2.2 1.9 1.5Falco tinnunculus NMNHS1/2002 1.7 2.2 2.2 1.9Falco tinnunculus NMNHS 7/1989 1.6 2.6 2.6 1.9Falco tinnunculus NMNHS 10/1992 1.7 2.7 2.5 2.0Falco vespertinus NMNHS 1/1991 1.4 2.2 2.1 1.7
Table 12. Measurements of phalanx 3 dig. III pedis in fossil and recent Falco.
Species a b cFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12544 ca. 1.6 ca. 2.2 ca. 1.8Falco bulgaricus sp. n. NMNHS 12545 ca. 1.6 - -RecentFalco columbarius NMNHS 2/2002 1.3 2.4 1.6Falco subbuteo NMNHS 3/1993 1.4 2.0 1.9Falco tinnunculus NMNHS 1/2002 1.6 2.4 2.2Falco tinnunculus NMNHS 7/1989 1.7 2.4 2.2Falco tinnunculus NMNHS 10/1992 1.8 2.4 2.3Falco vespertinus NMNHS 1/1991 1.4 1.9 2.1
Table 13. Measurements of the carpometacarpus in fossil and recent Falco.
Species Maximallength
Maximal width of proximalepiphysis
Width of distal epiphysis
Length of spa- tium interme-
ta-carpalis
Fossil - Cherevichniy Hutor - Ukraine1Falco medius 45-4033 39.8 10.4 7.4 26.2Recent - Bulgaria‘tinnunculus ’ groupFalco vespertinus NMNHS 1/1991 34.1 8.4 6.1 22.2Falco tinnunculus NMNHS 8/1989 44.6 19.4 16.3 31.5Falco tinnunculus NMNHS 7/1989 44.6 19.3 17.4 32.4Falco tinnunculus NMNHS 19/2005 45.5 19.4 17.0 33.1Falco columbarius NMNHS 2/2002 37.0 14.1 10.6 25.1Falco subbuteo NMNHS 1/1989 53.0 17.5 18.8 40.7Falco subbuteo NMNHS 3/1993 49.7 20.4 17.0 35.0‘cherrug ’ groupFalco peregrinus NMNHS 2/1989 64.2 21.8 15.8 43.5Falco cherrug NMNHS 1/1990 71.9 26.5 22.4 50.8
After Umanskaya (1981).
31
Boev Z.
Table 14. Mean values (for n >1) of the total length of carpometacarpus and humerus of some fossil and recentFalco.
Species n Total length of the bone
carpometacarpusFossil - Cherevichniy Hutor - Ukraine2Falco medius 45-4033 1 39.8Recent - Bulgaria‘tinnunculus ’ groupFalco columbarius 1 37.0Falco subbuteo 2 50.5Falco tinnunculus 3 44.9Falco vespertinus NMNHS 1/1991 1 34.1‘cherrug ’ groupFalco cherrug NMNHS 1/1990 1 71.9Falco peregrinus NMNHS 2/1989 1 64.2humerusFossil - HadzhidimovoFalco bulgaricus sp. n. NMNHS 12567 1 ca. 58.0Recent‘tinnunculus ’ groupFalco columbarius 4 49.0Falco subbuteo 7 53.9Falco tinnunculus 5 54.6Falco vespertinus 4 50.5Falco cherrug NMNHS 1/1990 1 97.3Falco peregrinus NMNHS 2/1989 1 84.1
Table 15. Humerus length to carpometacarpus length in some fossil and recent Falco.
Species n Humerus length: carpometacarpus length
FossilFalco medius 45-4033 1 -Falco bulgaricus sp. n. 1 -Recent‘tinnunculus ’ groupFalco columbarius NMNHS 2/2002 1 1.324Falco subbuteo 2 1.067Falco tinnunculus 3 1.216Falco vespertinus NMNHS 1/1991 1 1.480Falco spp. ex gr. tinnunculus (mean) 7 1.271‘cherrug ’ groupFalco cherrug NMNHS 1/1990 1 1.269Falco peregrinus NMNHS 2/1989 1 1.309
After Umanskaya (1981).
32

Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
fibularis, and more medial, than lateral, inception of distal end of crista cnemialis cranialis; F. longipen- nis: dimensionally and morphologically similar, but sharper bend o f crista cnemialis cranialis; F. nauman- ni: bigger, relatively longer crista fibularis, uneven crista fibularis, and smaller general size o f bone; F. sparverius: much larger; F. subbuteo: more medially, but not laterally, positioned base o f the crista cnemia- lis lateralis, longer base of crista cnemialis lateralis, deeper fossa flexoria and longer foramen interosseum distale, nevertheless general dimensional similarity; F. tinnuncculus: longer base (more proximally positioned inception) o f crista cnemialis lateralis, and less protruding area interarticularis; F. vespertinus: considerably larger, and presence of a shallow concavity on the cranial surface of crista fibularis. F. subbuteo: slightly smaller, longer foramen interosseum distale.
Tarsometatarsus sin. dist. NMNHS 12540 (Fig. 2 - d; Table 8) and tarsometatarsus dex. dist. NMNHS 12566 (Fig. 2 - e; Table 8). F. biarmicus: much smaller size, and more cranio-caudally flat diaphysis; F. chicquera: lower crista plantaris me- dialis; F. columbarius: wider distal part o f diaphysis at foramen vasculare distale, considerably less developed crista plantaris mediana in plantar aspect, and less developed edge between the f. subcutanea lateralis and the f. dorsalis in dorsal aspect; F. nau- manni: larger size, shallower relief on distal part of dorsal surface o f tmt, and deeper groove o f foramen vasculare distale on the cranial surface; F. subbuteo: the same way as F. columbarius; F. tinnununculus: smaller dimensions o f tmt; F. vespertinus: bigger size, and lower crista plantaris medialis.
Phalanx 1 dig. I pedis sin. NMNHS 12543 (Fig. 2 - f; Table 9). F. columbarius: larger size and bigger asymmetry in proximal half o f the phalanx; F. naumanni: bigger size, deeper relief on ventral (plantar) surface, and straighter shaft; F. subbuteo: smaller and more concave in proximal end on facies plantaris; F. tinnununculus: slightly smaller, less concave f. plantaris in prox. end; F. vespertinus: larger.
Phalanx 2 dig. II pedis dex. NMNHS 12542 (Fig. 2 - g, h, i; Table 10). F. columbarius: bigger; F. subbuteo: slightly smaller, and more concave in proximal end of facies plantaris; F. tinnununculus: slightly smaller; F. vespertinus: bigger and more robust.
Phalanx 2 dig. III pedis dex. NMNHS 12541 (Fig. 2 - j, k; Table 11). F. columbarius: bigger,
plantar edge o f f. a. prox. less concave; F. subbuteo: more concave in proximal end o f facies plataris; F. tinnununculus: smaller, less concave profile o f trochlea articularis in dorsal view; F. vespertinus: thicker phalangeal body.
Phalanx 3 dig. III pedis dex. NMNHS 12544 (Fig. 2 - l; Table 12). and Phalanx 3 dig. III pedis sin. NMNHS 12545 (Fig. 2 - m; Table 12). F. columbarius: slightly bigger; F. subbuteo: smaller; F. tinnununculus: smaller; F. vespertinus: bigger and more robust.
General comparison. The comparison o f skeletal elements o f the fore-limbs/pectoral girdle and the hind-limbs/pelvic girdle clearly demonstrate more developed flight capability, i. e. F. bulgaricus sp. n. to a certain extent was more aerial than terrestrial, in comparison to its modern closer relatives. Its wings were relatively more developed, while the legs were less developed in comparison to the closest dimensionally and in many respects, morphologically, species. The compared specimen has relatively longer wings (longer proximal part, i. e. humeri) and shorter legs (shorter distal part, i.e. thinner, and possibly shorter, tarsometatarsi).
The two parallel transversal grooves (lineae intermusculares) on the lateral side o f the humeral bone bellow the intumescentia, marking the end of crista bicipitalis, are specific both for the species o f ‘ tinnunculus’ group, and for the specimen of Hadzhidimovo and they are lacking in all the compared species o f ‘cherrug’ group. Thus they may be a plesiomorphic ferature for the smaller falcons.
Conclusions
The presence o f parallel transversal grooves on humerus could be considered as one o f the slight osteological distinguishing features, suggesting an ancient, at least at Late Pliocene separation of these groups o f falcons - (1) ‘tinnunculus' , kestrels (including hobbies, g. Hypotriorchis Bo i e , 1826), and (2) ‘cherrug’, hierofalcons Hierofalco Cu v e r ,
1817 (including Peregrine Falcon Falco peregrinus Tu n s t a l l , 1771). On the other hand, the position of the foramen nutritium of humeral bone (NMNHS 12567) is more distal, approaching to F. vesperinus, instead to F. tinnunculus.
33

Boev Z.
Probably these major groups deserve to be ranked as separate super-genera/genera. According to the fossil record the large falcons (of ‘cher- rug’ group) appeared in the Early Pleistocene and they are not represented in the Tertiary record (Ml i k o v s k y , 2002). On the other side, the Neogene record o f small falcons (of ‘ tinnunculus’ group) in Europe is well documented (Bulgaria, Hungary and Ukraine, all o f Late Pliocene) (Ml i k o v s k y , 2002). The relatively younger origin o f ‘cherrug’’ complex
References
Ba u m e l J. J., L. M. Wi t m e r 1993. 4 Osteologia, in: Baumel, J., King, A., Breazile, J., Evans, H., Vanden Berge, J., (Eds.): - In: Handbook of Avian Anatomy, Nomina Anatomica Avium. Nutall Ornithological Club, 23:45-132.
Bo c h e n s k i Z., 1997. List of European fossil bird species. - Acta zoologica cracoviensia 40 (2): 293-33.
Bo e v , Z. 1999. Falco bakalovi sp. n. - a Late Pliocene falcon (Falconidae, Aves) from Varshets (W Bulgaria). - Geo- logicaBalcanica, 29 (1-2): 131-135.
Bo e v Z., D. Ko v a c h e v 1998. Buteo spassovi sp. n. - a Late Miocene Buzzard (Accipitridae, Aves) from SW Bulgaria. - Geologica Balcanica, 29 (1-2): 125-129.
Bo e v Z., D. Ko v a c h e v 2007. Euroceros bulgaricus gen. nov., sp. nov. from Hadzhidimovo (SW Bulgaria) (Late Miocene) - the first European record of Hornbills (Aves: Coraciiformes). - Geobios, 40: 39-49.
Bo e v Z., N. Sp a s s o v 2009. First record of ostriches (Aves, Stru- thioniformes, Struthionidae) from the late Miocene of Bulgaria with taxonomic and zoogeographic discussion. - Geodiversitas, 31 (3): 493-507.
Bo e v Z. 2011. New fossil record of the Late Pliocene falcon (Falco bakalovi Boev, 1999) from type locality in Bulgaria. - Geologica Balcanica. (In press).
Li v e z e y В. C., R. L. Zu s i . 2006. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy: I.- Methods and Characters. - Bulletin of Carnegie Museum of Natural History, Pittsburgh, 37: 502-544.
Me i n P. 1990. Updating of MN zones. In: Lindsay E. H., Fahl- busch V., Mein P. (Eds.): - In: European Neogene mammal chronology. New York. Plenum Press, 73-90.
in comparison to smaller falcons is also confirmed by N i t t i n g e r et al. (2005).
Acknowledgments: The Short-term Visits Program of the Royal Society (London) and the National Museum of Natural History (Sofia) have supported the study. The author is very grateful to Mr Dimitar Kovachev for handing the material for examination and Dr. Robert Prys-Jones and Dr. Joanne Cooper (NHM) and Dr. Tereza Tomek and Dr. Zbigniew Bochenski (ISEAK) for providing excellent working conditions, which facilitated much of this study. Special thanks to Dr. Nikolay Spassov (NMNHS) for the helpful review of an earlier version of the manuscript.
Ml i k o v s k y J. 1996 (Ed.): Tertiary avian localities ofEurope. Acta universitatis Carolinae Geologica. Univerzita Karlova. Praha, 39 (1995): 519-852.
Ml i k o v s k y J., 2002. Cenozoic Birds of the World. Part 1, Europe.Praha, Ninox Press. 1-406.
Ni t t i n g e r F., Ha r i n g , E., Pi n s k e r , W., Wi n k , Mi c h a e l & Ga m a u f , A. 2005. Out ofAfrica? Phylogenetic relationships between Falco biarmicus and other hierofalcons (Aves Falconi- dae). - Journal o f ZoologicalSystematics and Evolutionary Research 43 (4): 321-331.
Ol s o n S. L. 1985. The fossil record of birds. - In: King, J. R., D. C. Parker, Eds. Avian Biology, Vol. VIII, Academic Press, New York, 79-252.
So b o l e v D. V 2003. New species of Pliocene falcon (Falconi- formes, Falconidae). - Vestnik zoologii, 37 (6): 85-87.
Sp a s s o v N. 2002. The Turolian Megafauna of West Bulgaria and the character of the Late Miocene ‘Pikermian biome’. - Bollettino della Societa Paleontologica Italiana, 41 (1): 69-81.
Su a r e z W., S. Ol s o n . 2001. A remarkable new species of small falcon from the Quaternary of Cuba (Aves: Falconidae: Falco). - Proceed. Biological Soc. Washington, 114 (2): 34-41.
Um a n s k a y a A. S. 1981. The Miocene birds of the Western Black Sea Coasts of the Ukrainian SSR. - Vestnik Zoologii, 17 (3): 17-21. (In Russian, English summary).
Wh i t e C. M, P. F. Ol s e n , L. F. Ki f f 1994. Family Falconidae (Falcons and Caracaras). - in: del Hoyo, J., A. Elliot, J. Sargatal (Eds.) Handbook of the Birds of the World. Vol. 2. New World Vultures to Guineafowl. Lynx Edicions, Barcelona. 216-275.
Received: 29.10.2010 Accepted: 13.01.2011
34

Falco bulgaricus sp. n. (Aves, Falconiformes) from the Late Miocene of Hadzhidimovo.
APPENDIX 1
Examined specimens belonging to recent species in the Falconidae
Lanner Falcon Falco biarmicus NHM 1976.60.5; Nankeen Kestrel Falco cenchroides ISEAK A 4916/91; Red-nacked Falcon Falco chicquera Da u d i n , 1800: NHM 1993.2.5; Merlin Falco columbarius Li n n a e u s , 1758: NHM 1862.1.18.3, NHM 1930.3.24.264, NHM 1988.61.1, ISEAK A 2532/72; Eleonora’s Falcon Falco eleonorae ISEAK A 5093/92; Australian Hobby Falco longipennis ISEAK A 5351/94; Lesser Kestrel Falco naumanni Fl e i s c h e r , 1818: NHM 1955.15.2, NHM 1961.13.4, ISEAK 4958/91, ISEAK A 4783/90, ISEAK A 4958/91; Greater Kestrel Falco rupicoloides ISEAK A 4782/90; American Kestrel Falco sparverius L i n n a e u s , 1758: ISEAK A 4106/84; Eurasian Hobby
Falco subboteo L i n n a e u s , 1758: ISEAK 230/41/14, ISEAK 244/38/13, ISEAK 416/62, ISEAK A 1483/64, ISEAK A 2071/69, ISEAK A 3196/76, ISEAK A 3620/79; Common Kestrel Falco tinnunculus L i n n a e u s , 1758: ISEAK 4509/88, ISEAK 4594/89, ISEAK A 4449/87, ISEAK A 4509/88, ISEAK A 4594/89, ISEAK A 4734/90; Red-footed Falcon Falco vespertinus Li n n a e u s , 1766: NHM 1855.4.4.9, NHM 1869.19.12, ISEAK A 2371/71, ISEAK A 4737/90, ISEAK A 5056/92 Collared Falconet Microhierax caerulescens (Li n n a e u s , 1758): NHM 2002.41.2; African Pygmy- falcon Polihierax semitorqutus (Sm i t h , 1846): NHM 2002.41.1.
35

Related Documents











