F-Actin Structure Destabilization and DNase I Binding Loop Fluctuations Mutational Cross-Linking and Electron Microscopy Analysis of Loop States and Effects on F-Actin Zeynep A. Oztug Durer 1 , Karthikeyan Diraviyam 2 , David Sept 2 , Dmitri S. Kudryashov 1 and Emil Reisler 1 ⁎ 1 Department of Chemistry and Biochemistry, and Molecular Biology Institute, University of California at Los Angeles, Los Angeles, CA 90095, USA 2 Biomedical Engineering and Center for Computational Medicine and Bioinformatics, University of Michigan, Ann Arbor, MI 48109, USA Received 13 August 2009; received in revised form 30 October 2009; accepted 2 November 2009 Available online 6 November 2009 The conformational dynamics of filamentous actin (F-actin) is essential for the regulation and functions of cellular actin networks. The main contribution to F-actin dynamics and its multiple conformational states arises from the mobility and flexibility of the DNase I binding loop (D-loop; residues 40–50) on subdomain 2. Therefore, we explored the structural constraints on D-loop plasticity at the F-actin interprotomer space by probing its dynamic interactions with the hydrophobic loop (H-loop), the C- terminus, and the W-loop via mutational disulfide cross-linking. To this end, residues of the D-loop were mutated to cysteines on yeast actin with a C374A background. These mutants showed no major changes in their polymerization and nucleotide exchange properties compared to wild-type actin. Copper-catalyzed disulfide cross-linking was investigated in equi- molar copolymers of cysteine mutants from the D-loop with either wild- type (C374) actin or mutant S265C/C374A (on the H-loop) or mutant F169C/C374A (on the W-loop). Remarkably, all tested residues of the D- loop could be cross-linked to residues 374, 265, and 169 by disulfide bonds, demonstrating the plasticity of the interprotomer region. However, each cross-link resulted in different effects on the filament structure, as detected by electron microscopy and light-scattering measurements. Disulfide cross- linking in the longitudinal orientation produced mostly no visible changes in filament morphology, whereas the cross-linking of D-loop residues N 45 to the H-loop, in the lateral direction, resulted in filament disruption and the presence of amorphous aggregates on electron microscopy images. A similar aggregation was also observed upon cross-linking the residues of the D-loop (N 41) to residue 169. The effects of disulfide cross-links on F-actin stability were only partially accounted for by the simulations of current F- actin models. Thus, our results present evidence for the high level of conformational plasticity in the interprotomer space and document the link between D-loop interactions and F-actin stability. © 2009 Elsevier Ltd. All rights reserved. Edited by R. Craig Keywords: actin; cross-linking; molecular dynamics simulations Introduction Actin is a major component of the cytoskeleton in eukaryotic cells that is capable of self-assembly into filaments and higher-order structures that participate in numerous cellular processes. Cellular functions of actin are intricately regulated by actin- binding proteins and filament dynamics. Thus, an in-depth understanding of actin-based cellular events requires comprehension of the structure *Corresponding author. E-mail address: [email protected]. Abbreviations used: F-actin, filamentous actin; D-loop, DNase I binding loop; H-loop, hydrophobic loop; G-actin, globular actin; TMR, tetramethylrhodamine-5-maleimide; EM, electron microscopy; WT, wild type; MTS-6, 1,6-hexanediyl bis (methanethiosulfonate); TCEP, tris (2-carboxyethyl) phosphine. doi:10.1016/j.jmb.2009.11.001 J. Mol. Biol. (2010) 395, 544–557 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2009 Elsevier Ltd. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1016/j.jmb.2009.11.001 J. Mol. Biol. (2010) 395, 544–557
Available online at www.sciencedirect.com
F-Actin Structure Destabilization and DNase I BindingLoop FluctuationsMutational Cross-Linking and Electron Microscopy Analysis ofLoop States and Effects on F-Actin
Zeynep A. Oztug Durer1, Karthikeyan Diraviyam2, David Sept2,Dmitri S. Kudryashov1 and Emil Reisler1⁎
1Department of Chemistry andBiochemistry, and MolecularBiology Institute, University ofCalifornia at Los Angeles,Los Angeles, CA 90095, USA2Biomedical Engineering andCenter for ComputationalMedicine and Bioinformatics,University of Michigan, AnnArbor, MI 48109, USAReceived 13 August 2009;received in revised form30 October 2009;accepted 2 November 2009Available online6 November 2009
*Corresponding author. E-mail [email protected] used: F-actin, filam
DNase I binding loop; H-loop, hydroglobular actin; TMR, tetramethylrhoEM, electron microscopy; WT, wild1,6-hexanediyl bis (methanethiosulf(2-carboxyethyl) phosphine.
0022-2836/$ - see front matter © 2009 E
The conformational dynamics of filamentous actin (F-actin) is essential forthe regulation and functions of cellular actin networks. The maincontribution to F-actin dynamics and its multiple conformational statesarises from the mobility and flexibility of the DNase I binding loop (D-loop;residues 40–50) on subdomain 2. Therefore, we explored the structuralconstraints on D-loop plasticity at the F-actin interprotomer space byprobing its dynamic interactions with the hydrophobic loop (H-loop), the C-terminus, and the W-loop via mutational disulfide cross-linking. To thisend, residues of the D-loop were mutated to cysteines on yeast actin with aC374A background. These mutants showed no major changes in theirpolymerization and nucleotide exchange properties compared to wild-typeactin. Copper-catalyzed disulfide cross-linking was investigated in equi-molar copolymers of cysteine mutants from the D-loop with either wild-type (C374) actin or mutant S265C/C374A (on the H-loop) or mutantF169C/C374A (on the W-loop). Remarkably, all tested residues of the D-loop could be cross-linked to residues 374, 265, and 169 by disulfide bonds,demonstrating the plasticity of the interprotomer region. However, eachcross-link resulted in different effects on the filament structure, as detectedby electron microscopy and light-scattering measurements. Disulfide cross-linking in the longitudinal orientation produced mostly no visible changesin filament morphology, whereas the cross-linking of D-loop residues N45to the H-loop, in the lateral direction, resulted in filament disruption and thepresence of amorphous aggregates on electron microscopy images. Asimilar aggregation was also observed upon cross-linking the residues ofthe D-loop (N41) to residue 169. The effects of disulfide cross-links on F-actinstability were only partially accounted for by the simulations of current F-actin models. Thus, our results present evidence for the high level ofconformational plasticity in the interprotomer space and document the linkbetween D-loop interactions and F-actin stability.
© 2009 Elsevier Ltd. All rights reserved.
Edited by R. Craig
Keywords: actin; cross-linking; molecular dynamics simulationsress:
entous actin; D-loop,phobic loop; G-actin,damine-5-maleimide;type; MTS-6,onate); TCEP, tris
lsevier Ltd. All rights reserve
Introduction
Actin is a major component of the cytoskeletonin eukaryotic cells that is capable of self-assemblyinto filaments and higher-order structures thatparticipate in numerous cellular processes. Cellularfunctions of actin are intricately regulated by actin-binding proteins and filament dynamics. Thus, anin-depth understanding of actin-based cellularevents requires comprehension of the structure
d.

545F-Actin Structure and DNase I Binding Loop
and conformational dynamics of globular actin (G-actin) and filamentous actin (F-actin).The atomic structure of G-actin suggests consid-
erable mobility and flexibility within the molecule,1
while a normal-mode analysis of G-actin ascribesthis mobility to the movement of subdomains andseveral dynamic elements, including the DNase Ibinding loop (D-loop; residues 39–50), the hydro-phobic loop (H-loop; residues 264–273), the C-terminus (including residues 374 and 375), and theW-loop (residues 165–172).2 Consistent with this,the D-loop and the C-terminus are disordered inmost crystal structures of G-actin.3–6 However,crystallographic studies demonstrated also that theD-loop can adopt specific conformational states.For example, the D-loop was seen as a β-strand inactin complexed to DNase I,1 while it formed ashort α-helix in the crystal of ADP tetramethylrho-damine-5-maleimide (TMR)-bound actin7 and wasunstructured in ATP TMR-bound actin,3,8 indi-cating the conformational plasticity inherent in thisstructural loop.In contrast to over 30 crystal structures reported
for G-actin alone and in complex with its bindingpartners, the information on F-actin structurederives from models built from fiber diffractiondata and electron microscopy (EM) reconstructions.The first and most frequently used actin model, the“Holmes model,” was built to 8 Å resolution byfitting the atomic structure of G-actin complexedwith DNase I to the X-ray fiber diffraction dataobtained from oriented F-actin gels in the presence ofphalloidin.1,9 The main filament contacts weresuggested to be between subdomain 3 of the upperprotomer and subdomain 4 of the lower protomer,supported by additional subdomain 1 and subdo-main 2 contacts involving the W-loop, the C-terminus, and D-loop residues 40–45. In addition,the two filament strands were hypothesized to bestabilized by the insertion of the H-loop from alateral protomer into a hydrophobic pocket at theinterface between two neighboring protomers in theopposite strand. Although the H-loop hypothesis isconsistent with results from several experimentalstudies (for a review, see Reisler and Egelman10), theloop was also shown to distribute among differentstates and orientations.11,12 This loop dynamicsappears essential for filament stability, since lockingof the H-loop to the protein backbone by cross-linking in a yeast actin mutant leads to filamentdestruction.11,13
The original refinements of the Holmes modelpreserved its main features while introducingchanges in the location and orientation of the D-loop and the C-terminus of actin.14,15 In 2003,Holmes et al. revised the actin filament model byusing an ADP TMR G-actin structure with a helicalD-loop.16 In this newer model, the helical D-loop ispositioned to interact with the W-loop in a protomerabove it in a manner resembling the binding ofWH2-domain-containing proteins to actin.17
The Holmes models and their subsequent refine-ments were built on the assumption that the mono-
mer structure is invariant upon its incorporation intothe filament. This assumption was partially sup-ported by the recent highest-resolutionOdamodel.18It has been revealed that G-actin-to-F-actin transitioninvolves a simple relative rotation of the two majordomains by about 20° to yield a flat actin molecule inthe filament. The second feature of this model is thatthe D-loop adopts an extended open loop confor-mation positioned along the hydrophobic cleftbetween subdomain 1 and subdomain 3.The differences noted among the various actin
models in the conformation and location of dynamicelements such as the D-loop can be viewed asreflecting the dynamic nature of the filament. In fact,a recent high-resolution cryo-EMmap of actin scruinbundle assigns numerous conformational states tothe filament.12 In that study, subdomain 2 displayedangular disorder, resulting in an average 10° rota-tion of subunits from their ideal positions, in agree-ment with prior EM reconstructions of F-actin.19,20
EM studies also revealed that variable tilts21 andtwists22 form spontaneously in pure filaments ofactin. These dynamic motions can be modulated byactin-binding proteins such as cofilin, which wasshown to shift the mean twist of the protomers by 5°by stabilizing one of the preexisting states.23
Actin models and experimental evidence suggestthat the dynamic loops of monomeric actin formimportant interprotomer contacts within the fila-ment24 and probably determine its conformationalstates. In this context, the D-loop is of particularinterest, since it represents the most flexible regionon actin, while its modifications or proteolyticcleavage directly affect actin polymerization.25–28Therefore, we investigated the plasticity of the D-loop (residues 40–50) by probing its dynamic inter-protomer interactions with the H-loop, the C-terminus, and the W-loop via disulfide cross-linkingwithin copolymers formed with cysteine mutantsfrom each element (Fig. 1). The characterization ofthese mutants revealed no major changes comparedto the wild-type (WT) actin. Strikingly, all testedresidues of the D-loop (mutated to cysteine) could becross-linked to the C-terminus (residue 374), the H-loop (residue 265), and the W-loop (residue 169) bydisulfide bonds, demonstrating the plasticity of theinterprotomer region. However, differences in cross-linking rates and extents reveal the link betweenfilament stability and specific D-loop states. Takentogether with our light-scattering and EM observa-tions, these results show the extensive conforma-tional plasticity of the interprotomer interface, whichallows also for the presence of intrinsically destabi-lized states of the filament.
Results
The goal of this study has been to demonstrate theplasticity of the D-loop and the inherent dynamics ofthe actin filament resulting from the interactionsbetween the D-loop, the C-terminus, the W-loop,and the H-loop by using copper-catalyzed disulfide

Fig. 1. A three-protomer Oda18
model of the actin filament, withthe cross-linked regions high-lighted. Residues of the D-loop areshown in red, the C-terminal resi-due C374 is shown in blue, the W-loop residue Y169 (mutated tocysteine in this study) is shown inblue, and the H-loop residue S265 isshown in green (mutated to cys-teine in this study). Individual pro-tomers are shown in pink, green,and blue ribbons.
546 F-Actin Structure and DNase I Binding Loop
cross-linking. To this end, we have generated single-cysteine mutants of the D-loop (residues 40–50) andresidue 169 by mutating the yeast actin (Saccharo-myces cerevisiae) gene along with the mutation ofnative cysteine 374 to an alanine. Therefore, eachmutant has a single reactive cysteine that isavailable for disulfide cross-linking. All actin muta-tions, except for M44C/C374A, were found toproduce viable yeast colonies, and the propertiesof the resulting actins were characterized. For asimplified notation, all actin mutants are denotedby a single-letter code (e.g., “C44” instead of M44C/C374A).
Functional characterization of mutants
To test the functional integrity of actinmutants, wecarried out nucleotide exchange and polymerizationexperiments. The exchange of fluorescent ɛ-ATPwith excess ATP can be used to detect mutationeffects on the nucleotide binding cleft. The C374Amutant exchanged ɛ-ATP at rates similar to those ofWT actin, indicating that the differences observed forvariousmutants can be attributed to the introductionof new cysteines (Fig. 2a). Mutant C48 (G48C/C374A) and mutant C49 (Q49C/C374A) exchangedɛ-ATP at rates similar to those of WT, whereasmutants C47 (M47C/C374A) – C45 (V45C/C374A)andmutants C43 (I43C/C374A) –C40 (H40/C374A)exhibited up to twofold slower rates compared to theWT protein (Fig. 2a). Since these mutations are notobserved in the cleft-forming residues, the relativelysmall changes observed are of allosteric nature.
Next, we monitored the polymerization of mutantactins via an increase in light scattering to detect ifthe mutations affect the nucleation and elongationbehaviors of actin. All actins, except for the C40 andC50 (K50C/C374A) mutants, polymerized at similarrates, indicating little, if any, change in nucleation orelongation steps (Fig. 2b). Modification of histidine40, which is suggested to participate in extensiveinterprotomer contacts,9 has been shown previouslyto inhibit actin polymerization.25 Hence, it is notsurprising that the cysteine replacement of thisresidue inhibits actin polymerization. The slownucleation phase observed for C50 might be due tothe loss of undefined ionic interactions with thereplacement of the native K50 (Fig. 2b). In bothcases, the negative effect of mutations can becompensated for by forming copolymers with WT,C265 (S265C/C374A), or C169 (F169C/C374A),which polymerizes at rates that are only slightlyslower than those of other copolymers (see Figs. 4aand 6b for C50).
Intermolecular cross-linking in F-actin and itsstructural consequences
Disulfide (zero length) and dibromobimane (4.4 Åon average) cross-linkings between residues 41–265and residues 41–374 had been demonstratedpreviously.24 This work extends the disulfidecross-linking to all residues of the D-loop betweenresidues 40 and 50 (except for residue 44, asmentioned above). These residues were cross-linkedto C-terminus residue 374, H-loop residue 265, and

Fig. 2. Functional characteriza-tion of mutant actins. (a) Ratios ofthe ɛ-ATP exchange rate of mutantactin to the ɛ-ATP exchange rate ofWT actin. The ɛ-ATP exchangeassays were performed as describedin Materials and Methods. Theratios of the calculated rates arepresented, with error bars indicat-ing the mean error. For WT actin,the rate of ɛ-ATP exchange was0.0157 ± 0.0018 s− 1 . (b) Actin(5.0 μM) polymerization by MgCl2(3.0 mM), as monitored by lightscattering at 325 nm. Plots werenormalized to reveal kinetic differ-ences in polymerization reactions.For clarity, only representative po-lymerization data are given: datafor WT (overlapping with C41 andC47) in black, data for C49 [over-lapping with C48, C45, and C42(G42C/C374A)] in blue, data forC46 in green, and data for C43 ingray. The mutant (C40) in cyancompleted its polymerization at6000 s under these conditions.
547F-Actin Structure and DNase I Binding Loop
W-loop residue 169 in equimolar polymers of D-loop mutants with either WT (C374) actin or mutantC265 or C169, thus allowing for a maximum pos-sible cross-linking yield of 50% (Materials andMethods). In control experiments, we exposedfilaments of WT actin or single reactive cysteine-carrying mutants to the same oxidizing conditionsas used for the copolymers, and we did not detectany cross-linked dimers (data not shown). Also, themonomers of WT and mutants did not form cross-linked dimers under reducing conditions. Thisagrees with previous results with WT, C41 (Q41C/C374S), and C265, and shows that the disulfidebonds are formed specifically between the reactivecysteines in the copolymer.24
Disulfide cross-linking of D-loop mutants withthe C-terminus
The 20 μM CuSO4-catalyzed disulfide cross-link-ing of D-loop mutants with C374 reached a plateauof 41.6±3.0% cross-linked dimers within 30 s or less
(representative copolymers are shown in Fig. 3a).Under these conditions, the F-actin-stabilizing toxinphalloidin29 did not appear to affect the rate andextent (41.4±2.3% dimers) of the cross-linkingreaction (Fig. 3a). However, with the increasedkinetic resolution of these reactions achieved atlow CuSO4 concentrations (5.0 μM), the rate ofcross-linking C41, C43, and C50 to C374 was about30–50% faster in the presence of phalloidin,consistent with previous results for C41 (Fig. 3b).30
Earlier reports have shown that the cross-linksintroduced either by disulfide bonds or by chemicalreagents can alter filament morphology and mayresult in filament destruction.11,13 Thus, we checkedthis possibility using both light-scattering and EMobservations. As shown in Fig. 4a, disulfide cross-linking to C374 either had no effect (with C41, C45,and C47) or caused only a slight decrease in lightscattering (with C43 and C50). As confirmed by EM,all control copolymers exhibited the regular mor-phology of yeast actin filaments, and the disulfidecross-linked filaments appeared normal for C41,

Fig. 3. Disulfide cross-links be-tween the D-loop and C265 andC374. (a) SDS-PAGE (nonreducing)of representative disulfide orMTS-6cross-linked yeast F-actin copoly-mers of D-loop mutants with C374and C265 actins. The cross-linkedpairs are indicated above the col-umns. The (−) and (+) signs indicatethe presence or the absence of20 μM phalloidin, CuSO4, or MTS-6 in the reaction mixtures. The timepoints at which the aliquots wereremoved are indicated in eachpanel. (b) Stabilization of filamentsby phalloidin increases the rates oflongitudinal cross-linking. The ob-served rate constants in the pres-ence (white circles) and in theabsence (black circles) of phalloidinfor the oxidation of copolymersC43–C374 (right) and copolymersC50–C374 (left) were calculated byfitting the percentage of monomersto a first-order exponential decayequation. The ratios of cross-linkingrates in the presence of phalloidinto cross-linking rates in the absenceof phalloidin were 1.44±0.12 and1.41±0.14 for C43–C374 and C50–C374 filaments, respectively. (c) Thepercentage of representative cross-linked dimers formed between D-loop and H-loop mutants in 30 s.The cross-linking products wereanalyzed on nonreducing SDS-PAGE after treatment with 2.0 mMN-ethyl maleimide. Error bars rep-resent the mean percentage error,and mean values are given insidethe bars.
548 F-Actin Structure and DNase I Binding Loop
C45, and C47 (data not shown). The oxidized fila-ments of C43 also had regular morphology, indicat-ing that the decrease in light scattering by cross-linking might be due to depolymerization from thefilament ends, with no detectable effect on their fila-ment structure (Fig. 4b). For C50, the oxidized fila-ments appeared shorter and kinked and, in manycases, were also severed or fragmented (Fig. 4b).Overall, these observations demonstrate that theresidues of the D-loop are highly dynamic, with eachone of them residing, at least transiently, in closeproximity to the C-terminus. These dynamicmotionsproduce multiple conformational states, all of whichare accommodated within the filament structure,
including those that—if locked—destabilize thefilament.
Disulfide cross-linking of D-loop mutants withH-loop residue C265
In contrast to similar disulfide cross-linking ratesof D-loop cysteine mutants to C374 (at 20 μMCu2+),the C-terminal region of the D-loop cross-links toC265 on the H-loop more slowly than its N-terminalresidues (Fig. 3a and c). These differences in cross-linking rates can be detected by monitoring theamount of dimers formed in 30 s. As shown in Fig.3c, residues 41, 43, and 45 cross-link equally well
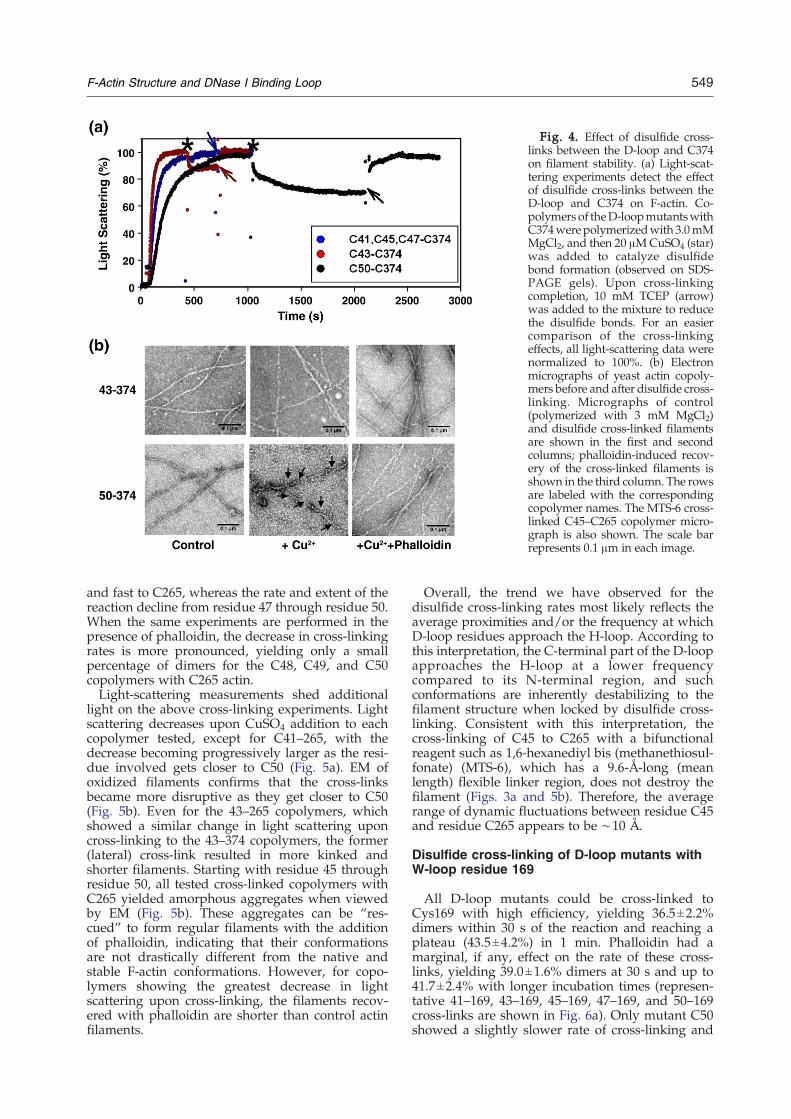
Fig. 4. Effect of disulfide cross-links between the D-loop and C374on filament stability. (a) Light-scat-tering experiments detect the effectof disulfide cross-links between theD-loop and C374 on F-actin. Co-polymers of theD-loopmutantswithC374werepolymerizedwith 3.0mMMgCl2, and then 20 μMCuSO4 (star)was added to catalyze disulfidebond formation (observed on SDS-PAGE gels). Upon cross-linkingcompletion, 10 mM TCEP (arrow)was added to the mixture to reducethe disulfide bonds. For an easiercomparison of the cross-linkingeffects, all light-scattering data werenormalized to 100%. (b) Electronmicrographs of yeast actin copoly-mers before and after disulfide cross-linking. Micrographs of control(polymerized with 3 mM MgCl2)and disulfide cross-linked filamentsare shown in the first and secondcolumns; phalloidin-induced recov-ery of the cross-linked filaments isshown in the third column. The rowsare labeled with the correspondingcopolymer names. The MTS-6 cross-linked C45–C265 copolymer micro-graph is also shown. The scale barrepresents 0.1 μm in each image.
549F-Actin Structure and DNase I Binding Loop
and fast to C265, whereas the rate and extent of thereaction decline from residue 47 through residue 50.When the same experiments are performed in thepresence of phalloidin, the decrease in cross-linkingrates is more pronounced, yielding only a smallpercentage of dimers for the C48, C49, and C50copolymers with C265 actin.Light-scattering measurements shed additional
light on the above cross-linking experiments. Lightscattering decreases upon CuSO4 addition to eachcopolymer tested, except for C41–265, with thedecrease becoming progressively larger as the resi-due involved gets closer to C50 (Fig. 5a). EM ofoxidized filaments confirms that the cross-linksbecame more disruptive as they get closer to C50(Fig. 5b). Even for the 43–265 copolymers, whichshowed a similar change in light scattering uponcross-linking to the 43–374 copolymers, the former(lateral) cross-link resulted in more kinked andshorter filaments. Starting with residue 45 throughresidue 50, all tested cross-linked copolymers withC265 yielded amorphous aggregates when viewedby EM (Fig. 5b). These aggregates can be “res-cued” to form regular filaments with the additionof phalloidin, indicating that their conformationsare not drastically different from the native andstable F-actin conformations. However, for copo-lymers showing the greatest decrease in lightscattering upon cross-linking, the filaments recov-ered with phalloidin are shorter than control actinfilaments.
Overall, the trend we have observed for thedisulfide cross-linking rates most likely reflects theaverage proximities and/or the frequency at whichD-loop residues approach the H-loop. According tothis interpretation, the C-terminal part of the D-loopapproaches the H-loop at a lower frequencycompared to its N-terminal region, and suchconformations are inherently destabilizing to thefilament structure when locked by disulfide cross-linking. Consistent with this interpretation, thecross-linking of C45 to C265 with a bifunctionalreagent such as 1,6-hexanediyl bis (methanethiosul-fonate) (MTS-6), which has a 9.6-Å-long (meanlength) flexible linker region, does not destroy thefilament (Figs. 3a and 5b). Therefore, the averagerange of dynamic fluctuations between residue C45and residue C265 appears to be ∼10 Å.
Disulfide cross-linking of D-loop mutants withW-loop residue 169
All D-loop mutants could be cross-linked toCys169 with high efficiency, yielding 36.5±2.2%dimers within 30 s of the reaction and reaching aplateau (43.5±4.2%) in 1 min. Phalloidin had amarginal, if any, effect on the rate of these cross-links, yielding 39.0±1.6% dimers at 30 s and up to41.7±2.4% with longer incubation times (represen-tative 41–169, 43–169, 45–169, 47–169, and 50–169cross-links are shown in Fig. 6a). Only mutant C50showed a slightly slower rate of cross-linking and

Fig. 5. Effect of disulfide cross-links between the D-loop and C265 on filament stability. (a) Light-scatteringexperiments detect the effect of disulfide cross-links between the D-loop and C265 on F-actin. All experiments wereperformed as described in the legend to Fig. 4a. The star and the arrows denote the addition of 20 μMCuSO4 and 10 mMTCEP, respectively. (b) Electron micrographs of yeast actin copolymers before and after disulfide cross-linking.Micrographs of control and disulfide cross-linked filaments are presented in the first and second columns; phalloidin-induced recovery of the cross-linked filaments is shown in the third column. The MTS-6 cross-linked C45–C265copolymer micrograph is also shown. The scale bar represents 0.1 μm in each image.
550 F-Actin Structure and DNase I Binding Loop
formed 28.2±1.7% and 31.1±1.9% dimers at 30 swith or without phalloidin, respectively. This islikely to result from a longer average distancebetween C50 and C169.According to the 2003 Holmes model and the Oda
model of F-actin, C169 is within 5–10 Å of D-loopresidues. In the more recent Oda model,18 C43, C44,C45, C46 (G46C/C374A), and C47 are predicted tobe close to C169. Therefore, we did not expect to see
filament destruction upon disulfide formation forthese residues. However, all copolymers showedsome (slight to major) decrease in light scatteringuponCuSO4 addition (Fig. 6b). As indicated by light-scattering decrease, the cross-linked copolymers ofC43–C169, C45–C169, and C50–C169 did not appearin EM images as filaments, but mainly as amorphousaggregates (Fig. 6d). Similar but larger aggregateswere observed for C47–C169 (Fig. 6f), consistent

Fig. 6. SDS-PAGE (nonreducing) and the effect of disulfide cross-links between the D-loop and C169 on actinfilaments. (a) SDS-PAGE of disulfide cross-linked for 30 s and 10 min F-actin (10 μM) copolymers of D-loop mutants withthe C169 mutant in the presence of 20 μM phalloidin. The copolymers are identified above the columns. (b) Light-scattering experiments detect the effect of disulfide cross-links between the D-loop and C169 on F-actin. All experimentswere performed as described in the legend to Fig. 4. The star and the arrows denote the addition of 20 μM CuSO4 and10 mM TCEP, respectively. The right column shows the EM images of oxidized F-actin copolymers C41–C169 (c), C45–C169 (d), and C47–C169 (f). The EM image of filaments regenerated with the addition of phalloidin to aggregates shownin (d) is given in (e).
551F-Actin Structure and DNase I Binding Loop
with the light-scattering results (Fig. 6b). All aggre-gates could be transformed into filaments by phal-loidin, as shown in Fig. 6e for the restored C45–C169filament. For the C41–C169 copolymer, which showsthe least decrease in light scattering upon cross-linking, the filaments appeared irregular and short-ened (Fig. 6c). These results show that D-loopflexibility or plasticity is needed for filament stabil-ity, as its cross-linking to theW-loop,which hasmorerestricted motions compared to the C-terminus,destroys the filament.
Molecular dynamics simulations of F-actinmodels
Using the coordinates from the Oda and Holmesactin filament models, we performed 30-ns molec-ular dynamics simulations on eight protomer F-actinstructures that we constructed for each model.9,18 Inthe Holmes model, we replaced the existing mono-mer structure with the ADP TMR-bound actinwhere the D-loop is in the form of a short α-helix.7
Specifically, we looked at the Cα–Cα distances
between the residues of the D-loop and residues169, 265, and 374 to test if the models explain theobserved cross-linking effects. In addition, we con-sidered also the distances between the same residuesin the 13-protomer actin scruin bundle model (Congmodel), in which all protomers were depicted tohave their unique orientation and conformation fordynamic regions such as the H-loop and the D-loop.12 Overall, the analysis of average Cα–Cα dis-tances between the D-loop and targeted residuesrevealed that no single model provides a compre-hensive description for the observed effects of disul-fide cross-links summarized in Table 1. In otherwords, they each offered a partial description for ourcross-linking results.Firstly, neither the Oda model nor the Holmes
model predicts the disulfide cross-links between C-terminal residue 374 and all D-loop residues, sinceaverage Cα–Cα distances are at least 9 Å (i.e., toofar for disulfide bond formation) (Fig. 7a). A fewcross-links are expected to occur to C374 (in Fig. 7afrom 43–45 in the Oda model and 46 and 47 in theHolmes model), but other cross-links could disrupt
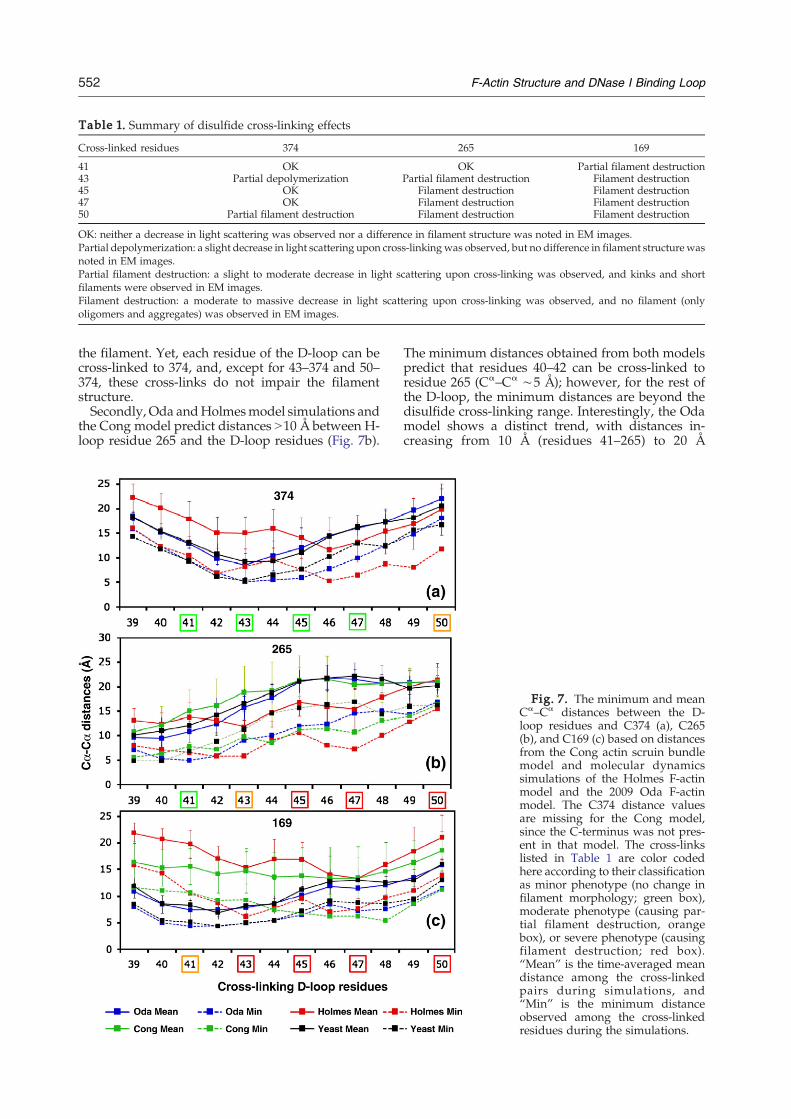
Table 1. Summary of disulfide cross-linking effects
Cross-linked residues 374 265 169
41 OK OK Partial filament destruction43 Partial depolymerization Partial filament destruction Filament destruction45 OK Filament destruction Filament destruction47 OK Filament destruction Filament destruction50 Partial filament destruction Filament destruction Filament destruction
OK: neither a decrease in light scattering was observed nor a difference in filament structure was noted in EM images.Partial depolymerization: a slight decrease in light scattering upon cross-linkingwas observed, but no difference in filament structure wasnoted in EM images.Partial filament destruction: a slight to moderate decrease in light scattering upon cross-linking was observed, and kinks and shortfilaments were observed in EM images.Filament destruction: a moderate to massive decrease in light scattering upon cross-linking was observed, and no filament (onlyoligomers and aggregates) was observed in EM images.
552 F-Actin Structure and DNase I Binding Loop
the filament. Yet, each residue of the D-loop can becross-linked to 374, and, except for 43–374 and 50–374, these cross-links do not impair the filamentstructure.Secondly, Oda andHolmesmodel simulations and
the Cong model predict distances N10 Å between H-loop residue 265 and the D-loop residues (Fig. 7b).
The minimum distances obtained from both modelspredict that residues 40–42 can be cross-linked toresidue 265 (Cα–Cα ∼5 Å); however, for the rest ofthe D-loop, the minimum distances are beyond thedisulfide cross-linking range. Interestingly, the Odamodel shows a distinct trend, with distances in-creasing from 10 Å (residues 41–265) to 20 Å
Fig. 7. The minimum and meanCα–Cα distances between the D-loop residues and C374 (a), C265(b), and C169 (c) based on distancesfrom the Cong actin scruin bundlemodel and molecular dynamicssimulations of the Holmes F-actinmodel and the 2009 Oda F-actinmodel. The C374 distance valuesare missing for the Cong model,since the C-terminus was not pres-ent in that model. The cross-linkslisted in Table 1 are color codedhere according to their classificationas minor phenotype (no change infilament morphology; green box),moderate phenotype (causing par-tial filament destruction, orangebox), or severe phenotype (causingfilament destruction; red box).“Mean” is the time-averaged meandistance among the cross-linkedpairs during simulations, and“Min” is the minimum distanceobserved among the cross-linkedresidues during the simulations.

553F-Actin Structure and DNase I Binding Loop
(between residues 45 and 265 and between residues50 and 265). This trend coincides with cross-links tothe H-loop disrupting the filament starting fromresidue 43. Taken together, our analysis indicatesthat the models would predict only part of thedisulfide cross-links and/or their effects, pointing tomore extensive dynamic rearrangements in theinterprotomer space than previously thought.Finally, the interresidue distances between W-loop
residue 169 and the D-loop residues obtained fromsimulations are the only set of data displaying dis-crete differences between the Oda model and theHolmes model of F-actin (Fig. 7c). Holmes modelsimulations give an average distance of 15 Å betweenresidue 169 and the D-loop, whereas Oda modelsimulations result in distances of less than 10 Å (witha minimum of ∼5 Å) from residue 169 to residue 40through residue 45. Thus, our D-loop–C169 disulfidecross-linking data appear more consistent with thevalues of the Oda model. However, such cross-linksdestroy the filament structure, as shown by light-scattering and EM experiments (Fig. 6b and c), andthese destabilizing effects can be rationalized betterby the much higher average distances observed inHolmesmodel simulations. For example, the averagedistance of C45–C169 is 10Å (while facing each other)and 17 Å (while facing away) in the Oda andHolmesmodels, respectively. Overall, C169–D-loop cross-linkings and simulation results suggest that, in thefilament, theD-loopmight be in dynamic equilibriumbetween an ordered state (2003Holmesmodel)16 anda disordered extended state (Oda model), and revealthat blocking this conformational exchange by cross-linking destroys filament integrity.
Discussion
The goal of ourworkwas to explore the constraintson D-loop plasticity at the interprotomer space, asdetermined by its interactions with other dynamicelements of actin, including the C-terminus, the W-loop, and the H-loop. The participation of thesestructural loops in interprotomer contacts in the actinfilament was predicted by the models of F-actin,9,14,15 and solution evidence confirmed suchinteractions.24 However, the solution results restedon the use of either a single-residue mutant in the D-loop, C41 in yeast actin,11,24,27,31 or proteolytic diges-tions as a tool for examining the environment of theN-terminal or C-terminal part of the D-loop.26,28
Thus, a comprehensive analysis of the motions andinteractions of the entire loop across the interproto-mer space has yet to be pursued. The combineddisulfide cross-linking, light-scattering, and EMstudies presented here offer such an analysis.
Structural states of F-actin include intrinsicallydestabilizing transient conformations
Actin filament was demonstrated to have multiplestructural states, depending on the bound nucleo-tide, divalent cations, and binding partners, by EM,
limited proteolysis, fluorescence spectroscopy, andradiolytic protein footprinting.28,32–38 Previous worksuggests that these states might arise from thedynamic interactions of subdomain 2 with its longi-tudinal and/or lateral contact sites in the filament.The disulfide cross-linking results of the presentwork add credence to this view and demonstratethat the interplay between the D-loop in subdomain2 and other dynamic elements of F-actin, includingthe C-terminus, the H-loop, and the W-loop, is moreextensive than predicted by the simulations of F-actin models.9,12,16,18 The fact that each testedresidue of the D-loop could be cross-linked toC374, C265, and C169 clearly reflects the plasticityand mobility of the interface between actin proto-mers. However, as we have shown by EM and light-scatteringmeasurements, some of themultiple statescreated by dynamic rearrangements of these struc-tural loops intrinsically destabilize the filament. Thisis evidenced by filament disruption upon trappingof such states via disulfide cross-linking.
Close proximity of the D-loop's C-terminal to theH-loop creates destabilizing modes in F-actin
A well-known example of actin's structural statesis the 5° (to 162°) twist induced by the actin-severingprotein cofilin.23 This variable twist was also foundto be inherent in pure actin filaments, suggestingthat cofilin acts by stabilizing this preexisting con-former of the filament.22 This 162° state might behighly relevant to the states corresponding to ourdisulfide cross-links between D-loop residues andH-loop residue 265. Our data show that these cross-links impair filament stability, starting from residue43, as shown by EM images of severed and kinkedfilaments. As the residue number in the D-loopapproaches 50, the cross-links cause completefilament disruption, yielding small aggregates thatcan be restored into short filaments by phalloidin.This rescue indicates that the conformations of thecross-linked actins can still be filament compatiblegiven additional stabilizing and/or bridging factors.Since the rate and extent of disulfide cross-linkingcan be viewed as a qualitative reflection of the realaverage proximities of the targeted thiol residues,our results suggest that the C-terminus of the D-loopapproaches the H-loop at a lower frequency than itsN-terminus, with the latter proximity appearingnecessary for filament stability.Intriguingly, the destabilized filament conformers
created by the motion/proximity of the C-terminalregion of the D-loop to the H-loop may be “used” bysevering proteins such as cofilin. Moreover, somepathogens may also capitalize on these states for cellinvasion. For example, the cross-linking of two actinmolecules between D-loop residue 50 and H-loopresidue 27039,40 induced by Vibrio choleraeMARTXVctoxin impairs actin polymerization by locking thesesubunits in a conformation incompatible to filamentelongation. Since residue 270 is close to residue 265used in this work, it is attractive to speculate that thetoxin traps actin in an unstable transient oligomeric

554 F-Actin Structure and DNase I Binding Loop
form present at the early stages of actin oligomer-ization, similar to the states we have identified inthis study.
Phalloidin stabilization of filaments has differenteffects on cross-linking rates
Filament stabilization by phalloidin, most likelythrough restriction of breathing motions in F-actin,29resulted in different effects on the rate of formation oflongitudinal and lateral disulfide cross-links. In thelongitudinal cross-links of the C-terminus and D-loops, phalloidin increased the cross-linking rates by30–50%, most likely by eliminating large-amplitudefluctuations of these elements and thus increasingthe probability of their cross-linking. In the lateral H-loop and D-loop cross-links, phalloidin inhibited theformation of cross-links from D-loop residue 47 andbeyond to residue 265. This inhibitory effect is alsocompatible with the reduced amplitude of loop fluc-tuations (i.e., the elimination of destabilizing statesthat bring residues 48–50 close to residue 265). In linewith this argument, incubation of C50–C265 copo-lymers with BeFx, an ADP-Pi analog known toreduce the amplitude of subdomain 2motions,33 alsoinhibited the formation of 50–265 cross-linkeddimers (data not shown). Notably, higher frequencystates detected by 43–265 and 45–265 cross-links,which are unaffected by phalloidin, demonstrate theexistence of these mildly destabilizing states also inthe presence of filament-stabilizing factors.
The conformational plasticity of the D-loop isrequired for F-actin stability
Because the W-loop has more restricted motionsand flexibility than the C-terminus and the H-loop,the disulfide cross-linking data provide, in this case,a stronger set of constraints on the conformation andposition of the D-loop residues. The D-loop can becross-linked to C169 from every position tested withsimilar efficiency, demonstrating that it mightassume a position in the hydrophobic cleft (betweensubdomain 1 and subdomain 3) as proposed byDominguez.17 It was further hypothesized in thatstudy that the D-loop might bind in this cleft in ahelical conformation similar to that observed for theWH2-domain-containing proteins. Our experimentsshow that locking C169 to any residue tested in theD-loop, except for C41, disrupts the filamentstructure and results in actin aggregation. Surpris-ingly, this is also true for the C45–C169 cross-link,which is modeled to face C169 within an averagedistance of 10 Å (and a minimum of 6.5 Å) in ourOda model simulations, allowing for a possibledisulfide bond formation between these residues.18
The same pair of residues was 17 Å on average and9.6 Å minimum in Holmes model simulations (withthe D-loop in a helical conformation), suggestingthat a disulfide cross-link between them wouldimpair filament stability.16 Thus, our data stronglysuggest that conformational fluctuations that bringD-loop residues close to C169 destabilize the
filament. This is evidenced by filament aggregationthrough trapping of these D-loop–C169 states bydisulfide cross-linking. Dynamic rearrangementsand/or conformational exchange of the D-loop isessential for the dissipation of these destabilizingfilament modes and filament stability. It is an intri-guing possibility, yet to be tested, that the Holmesand Oda models might represent ordered anddisordered conformers of the D-loop in the filament.An alternative possibility is that the cross-linking ofD-loop and C169 may preclude some conformation-al states of the H-loop within the interprotomerspace from the opposite strand.9,41
The interplay between the dynamic loops ofF-actin is functionally important
The ability of F-actin to undergo conformationalrearrangements is likely to play an important rolein the regulation of actin-mediated functions in thecell.10 In line with this view, the longitudinal cross-link between residue 41 on the D-loop and residue374 on the C-terminus inhibits actomyosin forcegeneration in the absence of morphologicalchanges in the cross-linked filament structures.42,43
Therefore, these dynamic fluctuations are requiredfor the function of actin, possibly for actin tointeract with its many binding partners in eukary-otic cells. Our work, along with previous studies,demonstrates that the highly conserved dynamicloops of actin are essential for these conformationalrearrangements.11,13,30,35,41,44,45 Interestingly, theunusual properties of the Leishmania actin46,47 wereattributed to sequence alterations in the H-loop orthe D-loop. Therefore, it is likely that differentorganisms tailor these structural elements to aproper dynamic functioning of their actin networks.
Conformational plasticity of actin filaments
It has been recognized that actin isoforms can havestructural and dynamic properties different fromeach other. For example, yeast actin filaments arereported to be slightly more flexible than skeletalactin filaments, as evidenced by fluorescence,48 limi-ted proteolysis,49 and three-dimensional EM recons-tructions.50 This raises the possibility that theinconsistency observed between our cross-linkingresults (based on yeast actin) and current actinmodels (based on skeletal actin) might be a result ofisoform specificity. To address this concern, we haveperformed molecular dynamics simulations of yeastactin filaments using the Oda model as startingpoint. Molecular dynamics simulations of skeletal F-actin and yeast F-actin are remarkably similar andshow only minor differences in the distancesbetween cross-linked residues (Fig. 7a–c, black andblue symbols). Therefore, the conformational flexi-bility demonstrated by our data is unlikely to beisoform specific and is probably a common feature ofactin filament dynamics.In conclusion, our study presents unambiguous
evidence for a high level of conformational plasticity

555F-Actin Structure and DNase I Binding Loop
in the interprotomer interface in F-actin formed withthe participation of dynamic loops, including the D-loop, the H-loop, the W-loop, and the C-terminus.The disulfide cross-linking of the D-loop to each ofthe given loops reveals the coexistence of destabiliz-ing and stabilizing states of the filament, which maylead tomultiple binding surfaces needed for filamentfunctions. The fact that the structural effects ofdisulfide cross-links can only be partially describedby the dynamic modeling simulations of currentactin filament models implies that these dynamicrearrangements in the subunit–subunit interface aremore extensive than predicted by these models.Therefore, our cross-linking results and the cross-linking effects on filament integrity provide struc-tural constraints that can be used to describe andmodel the dynamic modes of actin filaments.
Materials and Methods
Cloning and mutagenesis
The QuikChange Site-Directed Mutagenesis kit (Strata-gene) was used for the preparation of actin mutants. Yeastactin (S. cerevisiae) mutations were generated on pRS314containing the TRP1 marker and the C374A actin geneunder the control of the native actin promoter. Then, yeasthaploid cells with mutated actins were generated with theresulting plasmids, as described previously.51 The prepa-ration of mutants Q41C/C374S, S265C/C374A, andK50C/C374A is described elsewhere.24,40,51 Forwardprimers used for the cloning and site-directedmutagenesisof actin genes are given in Table 2. Reverse primers arecomplementary to the forward primers given.
Purification of actin mutants
Yeast actin was purified as described earlier.24 Theprotein was eluted from a DNase I affinity column withbuffer A [10 mM Tris (pH 7.8), 1 mM DTT, 0.2 mM ATP,and 0.2 mM CaCl2] containing 20–25% sucrose and 50%formamide, and was then applied directly to a DEAEcolumn. Actin was eluted from the DEAE column in thesame buffer containing 350 mM KCl. Purified G-actin wasdialyzed and stored in buffer B [10 mM Hepes (pH 7.6),0.2 mM CaCl2, 0.2 mM ATP, and 1.0 mM DTT] on ice.
Actin cross-linking and SDS-PAGE analysis
Immediately prior to cross-linking, DTT was removedfrom actin mutants (20 μM) by passing them through
Table 2. Forward primers used for site-directed mutagenesis
Mutation
H40C 5′ CG42C 5′ GI43C 5′ GM44C 5′ CCAV45C 5′ CAAGAG46C 5M47C 5′ CAAG48C 5′Q49C 5′ C
Sephadex G-50 spin columns equilibrated with buffer B.The heteromixture of actins was prepared bymixing equalmolar amounts of mutant actins to yield a combined finalconcentration of 10 μM. This hetero-G-actin mixture wasthen polymerized with 3.0 mMMgCl2 at 22 °C for 20 min.Copper or MTS reagents were added at a 1:2 actin:cross-linking catalyst/reagent mole ratio, and aliquots wereremoved at the desired time points. The reactions werestopped by the addition of 2.0 mM N-ethylmaleimide andSDS electrophoresis sample buffer without reducingreagents. The cross-linking products were run on nonre-ducing 10% SDS-PAGE and visualized by Coomassiestaining. The density values for protein bands werecalculated by Scion Image (Alpha 4.0.3.2), and plotswere generated by Sigma Plot 11 (Systat Software) orExcel 2007 (Microsoft Office).
Light-scattering assays
Actin polymerization, disulfide cross-linking, and di-sulfide reduction were monitored via light-scatteringmeasurements, with the PTI fluorometer set at 325 nmfor excitation and emission wavelengths. Actin polymer-ization was induced by adding 3.0 mM MgCl2 to theprotein solution at 5.0 μM for standard polymerizationassays and at 10 μM for experiments with copolymercross-linking experiments. CuSO4 (20 μM) was added tocatalyze disulfide bond formation in copolymers, andonce the plateau had reached 10 mM, DTT or tris (2-carboxyethyl) phosphine (TCEP) was added to reduce thedisulfide bonds.
Nucleotide exchange
ATP exchange in G-actin (1–2 μM) was followedthrough the fluorescence of ɛ-ATP. The release of ɛ-ATPfrom actin in buffer B (containing 5 μM ɛ-ATP) wasinitiated with the addition of 100 μMATP. The subsequentdecrease in the fluorescence of released ɛ-ATP wasmonitored with a PTI fluorometer, with the excitationand emission wavelengths set at 350 nm and 410 nm,respectively. The rates of exchange were obtained byfitting the fluorescence decay data to single-exponentialexpression.
Electron microscopy
The copolymers (10 μM) were prepared as describedabove and divided into two fractions. DTT (1.0 mM) wasadded to the first fraction, which was used as controlfilament. The second fraction was incubated with 20 μMCuSO4 for 45 min. Phalloidin (20 μM) was added to afraction removed from the oxidized sample and incubated
Forward primer
GTCGGTAGACCAAGATGTCAAGGTATCATGGTCGG 3′TAGACCAAGACACCAATGTATCATGGTCGGTATG 3′ACCAAGACACCAAGGTTGTATGGTCGGTATGGGTC 3′AGACACCAAGGTATCTGTGTCGGTATGGGTCAAAAAG 3′CACCAAGGTATCATGTGTGGTATGGGTCAAAAAGACTC 3′′ CCAAGGTATCATGGTCTGTATGGGTCAAAAAG 3′GGTATCATGGTCGGTTGTGGTCAAAAAGACTCCTAC 3′
GTATCATGGTCGGTATGTGTCAAAAAGACTCCTAC 3ATGGTCGGTATGGGTTGTAAAGACTCCTACGTTGG 3′

556 F-Actin Structure and DNase I Binding Loop
for 15 min. Eventually, all fractions were diluted to a finalconcentration of 2.5 μM with the corresponding polymer-ization buffers and applied to 400-mesh carbon-coatedcopper grids coated with formvar films (EM Sciences).After 60 s of adsorption, the grids were blotted dry andtreated with 1% uranyl acetate for 45 s. The grids wereexamined with a JEOL 1200EX-II electron microscopeoperated at 80 kV. The EM images were analyzed usingIMAGE J software.
Molecular dynamics simulations
We used three models of actin filament for simulationpurposes. The first was the Holmes model,9 where wereplaced the existing monomer structure with the newerADP actin crystal structure (1J6Z).7 The second filamentmodel was the Oda et al. structure (2ZWH).18 The finalstructure was a yeast filament structure based on the Odafilament model. Since yeast actin and muscle actin have88% identity with no insertions or deletions, we were ableto use Modeller52 to create the homology model. Althougha yeast G-actin structure does exist, using the Oda F-actinmodel as template was important, since the subdomains ofthe monomer are slightly rearranged in the filament. Weconstructed eight monomer filaments for each model,solvated the systems with TIP3P water, added neutraliz-ing ions, and set the ionic strength to 50 mM. Using themolecular dynamics package NAMD,53 we performed thesimulations of each filament using the CHARMM27 forcefield, an NPT ensemble at 1 atm pressure and 300 Ktemperature, a 10 Å cutoff for van der Waals interactionswith a switching distance of 8.5 Å, and particle meshEwald for long-range electrostatics. Covalent bonds withhydrogens were held rigid, allowing us to take 2-fs timesteps. Each system was heated in 50-K steps andequilibrated at 300 K for 5 ns, and we performedproduction runs of 30 ns, saving structures every 50 psfor analysis.
Acknowledgements
We thank Dr. Xiange Zheng for help with themolecular dynamics simulations. This work wassupported by US Public Health Service grant GM-077190 to E.R., National Science Foundation grantMCB 0316269 to E.R., and National Science Foun-dation grant GM067246 to D.S.
References
1. Kabsch, W., Mannherz, H. G., Suck, D., Pai, E. F. &Holmes, K. C. (1990). Atomic structure of the actin:DNase I complex. Nature, 347, 37–44.
2. Tirion, M. M. & ben-Avraham, D. (1993). Normalmode analysis of G-actin. J. Mol. Biol. 230, 186–195.
3. Graceffa, P. & Dominguez, R. (2003). Crystal structureof monomeric actin in the ATP state. Structural basisof nucleotide-dependent actin dynamics. J. Biol. Chem.278, 34172–34180.
4. McLaughlin, P. J., Gooch, J. T., Mannherz, H. G. &Weeds, A. G. (1993). Structure of gelsolin segment 1-actin complex and the mechanism of filamentsevering. Nature, 364, 685–692.
5. Otterbein, L. R., Cosio, C., Graceffa, P. & Dominguez,R. (2002). Crystal structures of the vitamin D-bindingprotein and its complex with actin: structural basis ofthe actin–scavenger system. Proc. Natl Acad. Sci. USA,99, 8003–8008.
6. Robinson, R. C.,Mejillano,M., Le, V. P., Burtnick, L.D.,Yin, H. L. & Choe, S. (1999). Domain movement ingelsolin: a calcium-activated switch. Science, 286,1939–1942.
7. Otterbein, L. R., Graceffa, P. & Dominguez, R. (2001).The crystal structure of uncomplexed actin in the ADPstate. Science, 293, 708–711.
8. Zheng, X., Diraviyam, K. & Sept, D. (2007). Nucleotideeffects on the structure and dynamics of actin. Biophys.J. 93, 1277–1283.
9. Holmes, K. C., Popp, D., Gebhard, W. & Kabsch, W.(1990). Atomic model of the actin filament. Nature,347, 44–49.
10. Reisler, E. & Egelman, E. H. (2007). Actin structureand function: what we still do not understand. J. Biol.Chem. 282, 36133–36137.
11. Scoville, D., Stamm, J. D., Toledo-Warshaviak, D.,Altenbach, C., Phillips, M., Shvetsov, A. et al. (2006).Hydrophobic loop dynamics and actin filamentstability. Biochemistry, 45, 13576–13584.
12. Cong, Y., Topf,M., Sali, A.,Matsudaira, P., Dougherty,M., Chiu, W. & Schmid, M. F. (2008). Crystallographicconformers of actin in a biologically active bundle offilaments. J. Mol. Biol. 375, 331–336.
13. Shvetsov, A., Stamm, J. D., Phillips, M., Warshaviak,D., Altenbach, C., Rubenstein, P. A. et al. (2006).Conformational dynamics of loop 262–274 in G- andF-actin. Biochemistry, 45, 6541–6549.
14. Lorenz, M., Popp, D. & Holmes, K. C. (1993).Refinement of the F-actin model against X-ray fiberdiffraction data by the use of a directed mutationalgorithm. J. Mol. Biol. 234, 826–836.
15. Tirion, M. M., ben-Avraham, D., Lorenz, M. &Holmes, K. C. (1995). Normal modes as refinementparameters for the F-actin model. Biophys. J. 68, 5–12.
16. Holmes, K. C., Angert, I., Kull, F. J., Jahn, W. &Schroder, R. R. (2003). Electron cryo-microscopyshows how strong binding of myosin to actin releasesnucleotide. Nature, 425, 423–427.
17. Dominguez, R. (2004). Actin-binding proteins—aunifying hypothesis. Trends Biochem. Sci. 29, 572–578.
18. Oda, T., Iwasa, M., Aihara, T., Maeda, Y. & Narita, A.(2009). The nature of the globular- to fibrous-actintransition. Nature, 457, 441–445.
19. Belmont, L. D., Orlova, A., Drubin, D. G. & Egelman,E. H. (1999). A change in actin conformation associ-ated with filament instability after Pi release. Proc.Natl Acad. Sci. USA, 96, 29–34.
20. Egelman, E. H. & DeRosier, D. J. (1991). Angulardisorder in actin: is it consistent with generalprinciples of protein structure? J. Mol. Biol. 217,405–408.
21. Galkin, V. E., VanLoock, M. S., Orlova, A. &Egelman, E. H. (2002). A new internal mode in F-actin helps explain the remarkable evolutionaryconservation of actin's sequence and structure.Curr. Biol. 12, 570–575.
22. Galkin, V. E., Orlova, A., Lukoyanova, N., Wriggers,W. & Egelman, E. H. (2001). Actin depolymerizingfactor stabilizes an existing state of F-actin and canchange the tilt of F-actin subunits. J. Cell Biol. 153,75–86.
23. Galkin, V. E., Orlova, A., VanLoock, M. S., Shvetsov,A., Reisler, E. & Egelman, E. H. (2003). ADF/cofilin

557F-Actin Structure and DNase I Binding Loop
use an intrinsic mode of F-actin instability to disruptactin filaments. J. Cell Biol. 163, 1057–1066.
24. Kim, E., Wriggers, W., Phillips, M., Kokabi, K.,Rubenstein, P. A. & Reisler, E. (2000). Cross-linkingconstraints on F-actin structure. J. Mol. Biol. 299,421–429.
25. Hegyi, G., Premecz, G., Sain, B. & Muhlrad, A. (1974).Selective carbethoxylation of the histidine residues ofactin by diethylpyrocarbonate. Eur. J. Biochem. 44,7–12.
26. Schwyter, D. H., Kron, S. J., Toyoshima, Y. Y.,Spudich, J. A. & Reisler, E. (1990). Subtilisin cleavageof actin inhibits in vitro sliding movement of actinfilaments over myosin. J. Cell Biol. 111, 465–470.
27. Kim, E., Motoki, M., Seguro, K., Muhlrad, A. &Reisler, E. (1995). Conformational changes in sub-domain 2 of G-actin: fluorescence probing by dansylethylenediamine attached to Gln-41. Biophys. J. 69,2024–2032.
28. Khaitlina, S. Y., Moraczewska, J. & Strzelecka-Golaszewska, H. (1993). The actin/actin interactionsinvolving the N-terminus of the DNase-I-bindingloop are crucial for stabilization of the actinfilament. Eur. J. Biochem. 218, 911–920.
29. Dancker, P., Low, I., Hasselbach, W. & Wieland, T.(1975). Interaction of actin with phalloidin: polymer-ization and stabilization of F-actin. Biochim. Biophys.Acta, 400, 407–414.
30. Scoville, D., Stamm, J. D., Altenbach, C., Shvetsov, A.,Kokabi, K., Rubenstein, P. A. et al. (2009). Effects ofbinding factors on structural elements in F-actin.Biochemistry, 48, 370–378.
31. Kim, E. & Reisler, E. (1996). Intermolecular couplingbetween loop 38–52 and the C-terminus in actinfilaments. Biophys. J. 71, 1914–1919.
32. Mossakowska, M., Moraczewska, J., Khaitlina, S. &Strzelecka-Golaszewska, H. (1993). Proteolytic remov-al of three C-terminal residues of actin alters themonomer–monomer interactions. Biochem. J. 289,897–902.
33. Muhlrad, A., Cheung, P., Phan, B. C., Miller, C. &Reisler, E. (1994). Dynamic properties of actin.Structural changes induced by beryllium fluoride. J.Biol. Chem. 269, 11852–11858.
34. Orlova, A. & Egelman, E. H. (1992). Structural basisfor the destabilization of F-actin by phosphate releasefollowing ATP hydrolysis. J. Mol. Biol. 227, 1043–1053.
35. Orlova, A. & Egelman, E. H. (1993). A conformationalchange in the actin subunit can change the flexibilityof the actin filament. J. Mol. Biol. 232, 334–341.
36. Moraczewska, J., Strzelecka-Golaszewska, H., Moens,P. D. & dos Remedios, C. G. (1996). Structural changesin subdomain 2 of G-actin observed by fluorescencespectroscopy. Biochem. J. 317, 605–611.
37. Strzelecka-Golaszewska,H.,Moraczewska, J., Khaitlina,S. Y. & Mossakowska, M. (1993). Localization of thetightly bound divalent-cation-dependent and nucleo-tide-dependent conformation changes inG-actin usinglimited proteolytic digestion. Eur. J. Biochem. 211,731–742.
38. Guan, J. Q., Takamoto, K., Almo, S. C., Reisler, E. &Chance, M. R. (2005). Structure and dynamics of theactin filament. Biochemistry, 44, 3166–3175.
39. Kudryashov, D. S., Cordero, C. L., Reisler, E. &Satchell, K. J. (2008). Characterization of the enzymat-ic activity of the actin cross-linking domain from theVibrio cholerae MARTX Vc toxin. J. Biol. Chem. 283,445–452.
40. Kudryashov, D. S., Durer, Z. A., Ytterberg, A. J.,Sawaya, M. R., Pashkov, I., Prochazkova, K. et al.(2008). Connecting actin monomers by iso-peptidebond is a toxicity mechanism of the Vibrio choleraeMARTX toxin. Proc. Natl Acad. Sci. USA, 105,18537–18542.
41. Shvetsov, A., Musib, R., Phillips, M., Rubenstein, P. A.& Reisler, E. (2002). Locking the hydrophobic loop262–274 to G-actin surface by a disulfide bridgeprevents filament formation. Biochemistry, 41,10787–10793.
42. Kim, E., Bobkova, E., Hegyi, G., Muhlrad, A. &Reisler, E. (2002). Actin cross-linking and inhibition ofthe actomyosin motor. Biochemistry, 41, 86–93.
43. Prochniewicz, E. & Thomas, D. D. (1997). Perturba-tions of functional interactions with myosin inducelong-range allosteric and cooperative structuralchanges in actin. Biochemistry, 36, 12845–12853.
44. Orlova, A. & Egelman, E. H. (1995). Structuraldynamics of F-actin: I. Changes in the C terminus. J.Mol. Biol. 245, 582–597.
45. Orlova, A., Prochniewicz, E. & Egelman, E. H. (1995).Structural dynamics of F-actin: II. Cooperativity instructural transitions. J. Mol. Biol. 245, 598–607.
46. Kapoor, P., Sahasrabuddhe, A. A., Kumar, A., Mitra,K., Siddiqi, M. I. & Gupta, C. M. (2008). Anunconventional form of actin in protozoan hemofla-gellate, Leishmania. J. Biol. Chem. 283, 22760–22773.
47. Sahasrabuddhe, A. A., Bajpai, V. K. & Gupta, C. M.(2004). A novel form of actin in Leishmania: molecularcharacterisation, subcellular localisation and associa-tion with subpellicular microtubules. Mol. Biochem.Parasitol. 134, 105–114.
48. Prochniewicz, E. & Thomas, D. D. (1999). Differencesin structural dynamics of muscle and yeast actinaccompany differences in functional interactions withmyosin. Biochemistry, 38, 14860–14867.
49. Kim, E., Miller, C. J. & Reisler, E. (1996). Polymer-ization and in vitro motility properties of yeast actin:a comparison with rabbit skeletal alpha-actin.Biochemistry, 35, 16566–16572.
50. Orlova, A., Chen, X., Rubenstein, P. A. & Egelman, E.H. (1997). Modulation of yeast F-actin structure by amutation in the nucleotide-binding cleft. J. Mol. Biol.271, 235–243.
51. Feng, L., Kim, E., Lee, W. L., Miller, C. J., Kuang, B.,Reisler, E. & Rubenstein, P. A. (1997). Fluorescenceprobing of yeast actin subdomain 3/4 hydrophobicloop 262–274. Actin–actin and actin–myosin interac-tions in actin filaments. J. Biol. Chem. 272, 16829–16837.
52. Eswar,N.,Webb,B.,Marti-Renom,M.A.,Madhusudhan,M. S., Eramian, D., Shen, M. Y. et al. (2006).Comparative protein structure modeling using Mod-eller. Curr. Protoc. Bioinf.; chapt. 5, unit 5.6.
53. Phillips, J. C., Braun, R., Wang, W., Gumbart, J.,Tajkhorshid, E., Villa, E. et al. (2005). Scalablemolecular dynamics with NAMD. J. Comput. Chem.26, 1781–1802.
Related Documents



![Review Actin-targeting natural products: structures ... · actin-binding proteins actively break or ‘sever’ actin filaments [e.g. actin-depolymerizing factor (ADF) and cofilin].](https://static.cupdf.com/doc/110x72/5f0f85bd7e708231d44494d0/review-actin-targeting-natural-products-structures-actin-binding-proteins-actively.jpg)

![CYTOSKELETON NEWS - fnkprddata.blob.core.windows.net · Dynamic remodeling of the actin cytoskeleton [i.e., rapid cycling between filamentous actin (F-actin) and monomer actin (G-actin)]](https://static.cupdf.com/doc/110x72/609edd2b88630103265d18ee/cytoskeleton-news-dynamic-remodeling-of-the-actin-cytoskeleton-ie-rapid-cycling.jpg)





