Molecular & Biochemical Parasitology 193 (2014) 75–81 Contents lists available at ScienceDirect Molecular & Biochemical Parasitology Extracellular functions of glycolytic enzymes of parasites: Unpredicted use of ancient proteins Amaranta Gómez-Arreaza a , Hector Acosta a , Wilfredo Qui ˜ nones b , Juan Luis Concepción b , Paul A.M. Michels b,c , Luisana Avilán a,∗ a Laboratorio de Fisiología Animal, Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101, Venezuela b Laboratorio de Enzimología de Parásitos, Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101, Venezuela c Institute of Structural and Molecular Biology, School of Biological Sciences, University of Edinburgh, King’s Buildings, Edinburgh EH9 3JU, Scotland, UK a r t i c l e i n f o Article history: Received 16 January 2014 Received in revised form 10 February 2014 Accepted 24 February 2014 Available online 3 March 2014 Keywords: Exosomes Extracellular localization Glycolytic enzymes Host-parasite interactions Moonlighting functions Shedding vesicles a b s t r a c t In addition of their usual intracellular localization where they are involved in catalyzing reactions of car- bohydrate and energy metabolism by glycolysis, multiple studies have shown that glycolytic enzymes of many organisms, but notably pathogens, can also be present extracellularly. In the case of parasitic protists and helminths, they can be found either secreted or attached to the surface of the parasites. At these extracellular localizations, these enzymes have been shown to perform additional, very dif- ferent so-called “moonlighting” functions, such as acting as ligands for a variety of components of the host. Due to this recognition, different extracellular glycolytic enzymes participate in various important parasite–host interactions such as adherence and invasion of parasites, modulation of the host’s immune and haemostatic systems, promotion of angiogenesis, and acquisition of specific nutrients by the para- sites. Accordingly, extracellular glycolytic enzymes are important for the invasion of the parasites and their establishment in the host, and in determining their virulence. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Glycolysis, which is a metabolic pathway involved in ATP and NADH production, also plays a central role in metabolism by providing its intermediates to several other pathways. Glycoly- sis is found in almost all organisms and for this reason can be considered as an ancient process. Its most common form is the Embden–Meyerhof pathway, but other variants can be found in many organisms. It comprises 10 enzyme-catalyzed steps that con- vert glucose into pyruvate (Fig. 1). Although glycolysis has been extensively studied from kinetic, structural and functional points of view in both prokaryotic and eukaryotic organisms, this path- way and its enzymes continue revealing new features that help to deepen our understanding of the multiple functions of the process and its enzymes as exerted in different cells. Glycolysis occurs nor- mally in the cytosol and although considered as being freely soluble, Abbreviations: ALD, aldolase; ENO, enolase; GAPDH, glyceraldehyde-3- phosphate dehydrogenase; HK, hexokinase; PFK, phosphofructokinase; PGAM, phosphoglycerate mutase; PGI, glucose-6-phosphate isomerase; PYK, pyruvate kinase; TIM, triosephosphate isomerase. ∗ Corresponding author. Tel.: +58 274 2401307; fax: +58 274 2401286. E-mail address: [email protected] (L. Avilán). glycolytic enzymes often interact with each other in multi-enzyme complexes and with cellular structures such as the cytoskeleton or membranes, and such interactions can modulate their functional properties [1]. Moreover, in some organisms, as is the case for parasites belonging to the trypanosomatid group, glycolysis can also be organized within organelles related to peroxisomes named glycosomes [2]. Although the metabolism in parasites can vary as part of strate- gies to adapt their life style to the niches occupied in their hosts, the majority of parasitic protists and helminths appear to perform gly- colysis. Dependent on the nutrients found in their environment and the stage of their life cycle, many parasitic organisms rely preferen- tially on glucose catabolism to generate ATP. Moreover, in several parasites this catabolism involves solely glycolysis, without further metabolism of its end product pyruvate or lactate, as for example, in the bloodstream form of the sleeping sickness-causing kineto- plastid Trypanosoma brucei [3] and in the intra-erythrocytic stage of the malaria parasite Plasmodium falciparum [4]. 2. Glycolytic enzymes may have multiple functions Besides their role in glycolysis, glycolytic enzymes may func- tion in a variety of other cellular processes occurring at different http://dx.doi.org/10.1016/j.molbiopara.2014.02.005 0166-6851/© 2014 Elsevier B.V. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EU
APa
b
c
a
ARRAA
KEEGHMS
1
NpscEmveowdam
ppk
h0
Molecular & Biochemical Parasitology 193 (2014) 75–81
Contents lists available at ScienceDirect
Molecular & Biochemical Parasitology
xtracellular functions of glycolytic enzymes of parasites:npredicted use of ancient proteins
maranta Gómez-Arreazaa, Hector Acostaa, Wilfredo Quinonesb, Juan Luis Concepciónb,aul A.M. Michelsb,c, Luisana Avilána,∗
Laboratorio de Fisiología Animal, Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101, VenezuelaLaboratorio de Enzimología de Parásitos, Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101, VenezuelaInstitute of Structural and Molecular Biology, School of Biological Sciences, University of Edinburgh, King’s Buildings, Edinburgh EH9 3JU, Scotland, UK
r t i c l e i n f o
rticle history:eceived 16 January 2014eceived in revised form 10 February 2014ccepted 24 February 2014vailable online 3 March 2014
eywords:
a b s t r a c t
In addition of their usual intracellular localization where they are involved in catalyzing reactions of car-bohydrate and energy metabolism by glycolysis, multiple studies have shown that glycolytic enzymesof many organisms, but notably pathogens, can also be present extracellularly. In the case of parasiticprotists and helminths, they can be found either secreted or attached to the surface of the parasites.At these extracellular localizations, these enzymes have been shown to perform additional, very dif-ferent so-called “moonlighting” functions, such as acting as ligands for a variety of components of the
xosomesxtracellular localizationlycolytic enzymesost-parasite interactionsoonlighting functions
hedding vesicles
host. Due to this recognition, different extracellular glycolytic enzymes participate in various importantparasite–host interactions such as adherence and invasion of parasites, modulation of the host’s immuneand haemostatic systems, promotion of angiogenesis, and acquisition of specific nutrients by the para-sites. Accordingly, extracellular glycolytic enzymes are important for the invasion of the parasites andtheir establishment in the host, and in determining their virulence.
© 2014 Elsevier B.V. All rights reserved.
. Introduction
Glycolysis, which is a metabolic pathway involved in ATP andADH production, also plays a central role in metabolism byroviding its intermediates to several other pathways. Glycoly-is is found in almost all organisms and for this reason can beonsidered as an ancient process. Its most common form is thembden–Meyerhof pathway, but other variants can be found inany organisms. It comprises 10 enzyme-catalyzed steps that con-
ert glucose into pyruvate (Fig. 1). Although glycolysis has beenxtensively studied from kinetic, structural and functional pointsf view in both prokaryotic and eukaryotic organisms, this path-ay and its enzymes continue revealing new features that help to
eepen our understanding of the multiple functions of the processnd its enzymes as exerted in different cells. Glycolysis occurs nor-ally in the cytosol and although considered as being freely soluble,Abbreviations: ALD, aldolase; ENO, enolase; GAPDH, glyceraldehyde-3-hosphate dehydrogenase; HK, hexokinase; PFK, phosphofructokinase; PGAM,hosphoglycerate mutase; PGI, glucose-6-phosphate isomerase; PYK, pyruvateinase; TIM, triosephosphate isomerase.∗ Corresponding author. Tel.: +58 274 2401307; fax: +58 274 2401286.
E-mail address: [email protected] (L. Avilán).
ttp://dx.doi.org/10.1016/j.molbiopara.2014.02.005166-6851/© 2014 Elsevier B.V. All rights reserved.
glycolytic enzymes often interact with each other in multi-enzymecomplexes and with cellular structures such as the cytoskeleton ormembranes, and such interactions can modulate their functionalproperties [1]. Moreover, in some organisms, as is the case forparasites belonging to the trypanosomatid group, glycolysis canalso be organized within organelles related to peroxisomes namedglycosomes [2].
Although the metabolism in parasites can vary as part of strate-gies to adapt their life style to the niches occupied in their hosts, themajority of parasitic protists and helminths appear to perform gly-colysis. Dependent on the nutrients found in their environment andthe stage of their life cycle, many parasitic organisms rely preferen-tially on glucose catabolism to generate ATP. Moreover, in severalparasites this catabolism involves solely glycolysis, without furthermetabolism of its end product pyruvate or lactate, as for example,in the bloodstream form of the sleeping sickness-causing kineto-plastid Trypanosoma brucei [3] and in the intra-erythrocytic stageof the malaria parasite Plasmodium falciparum [4].
2. Glycolytic enzymes may have multiple functions
Besides their role in glycolysis, glycolytic enzymes may func-tion in a variety of other cellular processes occurring at different
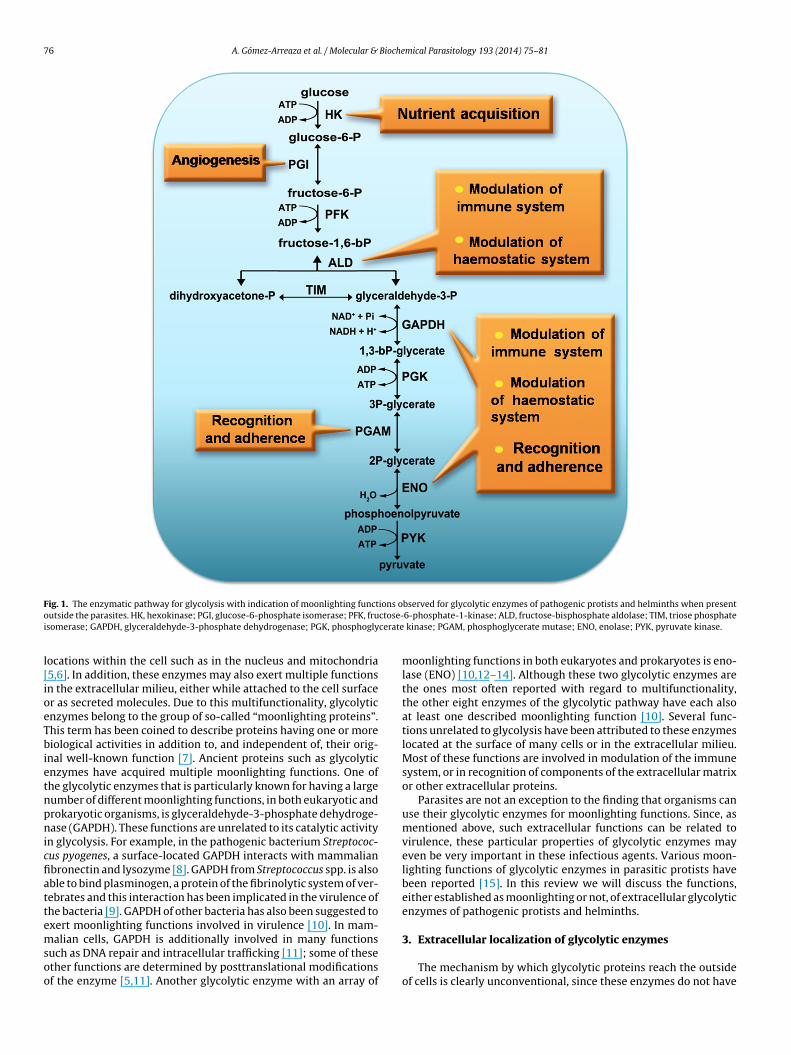
76 A. Gómez-Arreaza et al. / Molecular & Biochemical Parasitology 193 (2014) 75–81
F ions oo ctose-i cerate
l[ioeTbietnpnicfiattemsoo
ig. 1. The enzymatic pathway for glycolysis with indication of moonlighting functutside the parasites. HK, hexokinase; PGI, glucose-6-phosphate isomerase; PFK, frusomerase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; PGK, phosphogly
ocations within the cell such as in the nucleus and mitochondria5,6]. In addition, these enzymes may also exert multiple functionsn the extracellular milieu, either while attached to the cell surfacer as secreted molecules. Due to this multifunctionality, glycolyticnzymes belong to the group of so-called “moonlighting proteins”.his term has been coined to describe proteins having one or moreiological activities in addition to, and independent of, their orig-
nal well-known function [7]. Ancient proteins such as glycolyticnzymes have acquired multiple moonlighting functions. One ofhe glycolytic enzymes that is particularly known for having a largeumber of different moonlighting functions, in both eukaryotic androkaryotic organisms, is glyceraldehyde-3-phosphate dehydroge-ase (GAPDH). These functions are unrelated to its catalytic activity
n glycolysis. For example, in the pathogenic bacterium Streptococ-us pyogenes, a surface-located GAPDH interacts with mammalianbronectin and lysozyme [8]. GAPDH from Streptococcus spp. is alsoble to bind plasminogen, a protein of the fibrinolytic system of ver-ebrates and this interaction has been implicated in the virulence ofhe bacteria [9]. GAPDH of other bacteria has also been suggested toxert moonlighting functions involved in virulence [10]. In mam-
alian cells, GAPDH is additionally involved in many functionsuch as DNA repair and intracellular trafficking [11]; some of thesether functions are determined by posttranslational modificationsf the enzyme [5,11]. Another glycolytic enzyme with an array of
bserved for glycolytic enzymes of pathogenic protists and helminths when present6-phosphate-1-kinase; ALD, fructose-bisphosphate aldolase; TIM, triose phosphate
kinase; PGAM, phosphoglycerate mutase; ENO, enolase; PYK, pyruvate kinase.
moonlighting functions in both eukaryotes and prokaryotes is eno-lase (ENO) [10,12–14]. Although these two glycolytic enzymes arethe ones most often reported with regard to multifunctionality,the other eight enzymes of the glycolytic pathway have each alsoat least one described moonlighting function [10]. Several func-tions unrelated to glycolysis have been attributed to these enzymeslocated at the surface of many cells or in the extracellular milieu.Most of these functions are involved in modulation of the immunesystem, or in recognition of components of the extracellular matrixor other extracellular proteins.
Parasites are not an exception to the finding that organisms canuse their glycolytic enzymes for moonlighting functions. Since, asmentioned above, such extracellular functions can be related tovirulence, these particular properties of glycolytic enzymes mayeven be very important in these infectious agents. Various moon-lighting functions of glycolytic enzymes in parasitic protists havebeen reported [15]. In this review we will discuss the functions,either established as moonlighting or not, of extracellular glycolyticenzymes of pathogenic protists and helminths.
3. Extracellular localization of glycolytic enzymes
The mechanism by which glycolytic proteins reach the outsideof cells is clearly unconventional, since these enzymes do not have

A. Gómez-Arreaza et al. / Molecular & Biochemical Parasitology 193 (2014) 75–81 77
Table 1Determined moonlighting functions of extracellular glycolytic enzymes of parasites.
Glycolytic enzyme Moonlighting function Reference
Hexokinase (HK) Haemoglobin binding by Leishmania donovani [44]Glucose-6-phosphate isomerase (PGI) In vitro promotion of endothelial cell division by Echinococcus multilocularis [43]Aldolase (ALD) Plasminogen binding by various parasites
Activation of mouse macrophage protein tyrosine phosphatase-1 (SHP-1) by L.donovani
[33][38]
Glyceraldehyde-3-phosphatedehydrogenase (GAPDH)
Fibronectin binding by Trichomonas vaginalisPlasminogen binding by various parasitesComplement C3 binding and inactivation by Haemonchus contortus
[32][33][40]
Phosphoglycerate mutase (PGAM) Heparin binding motif in Leishmania spp. [30]Enolase (ENO) Gut epithelial interaction by Plasmodium spp.
y vari by St
[29]
ahsato[aErceTtwtcdffebiafpm[
otaefvwep
crgtedatipmi
ing this notion can be found in the reported surface localization ofglycolytic enzymes on exosome-like vesicles of prostate cells [27].Another aspect is that secretion of an enzyme can be triggered.For example, in Giardia lamblia trophozoites, ENO is localized in
Plasminogen binding bAntiphagocytic activity
recognizable export signal sequence. Several glycolytic enzymesave been found associated with secreted vesicles such as exo-omes, that are intracellularly derived from multivesicular bodiesnd released when they fuse with the plasma membrane. Alterna-ively, they have been found with shedding vesicles, another typef vesicles produced directly by budding of the plasma membrane16,17]. Such vesicles are thus formed by one of the different mech-nisms known for unconventional protein secretion by eukaryotes.xosomes and other types of secreted vesicles have also beeneported for protist parasites, such as trypanosomatids and api-omplexans, and for several helminths [18,19]. How glycolyticnzymes are sorted to such secreted vesicles is not yet known.he secretion of glycolytic enzymes by vesicles may be linkedo post-translational modifications that are sometimes associatedith moonlighting functions of proteins [11]. One may speculate
hat relocation of glycolytic enzymes to the plasma membranean help in targeting enzymes into vesicles to be formed by shed-ing. Such relocation of enzymes within the cytosol to the internalace of the plasma membrane has been previously documentedor mammalian cells [20]. Similarly, translocation of the glycolyticnzyme complexes from the cytosol to the pellicle (i.e. a tripleilayer structure characteristic of apicomplexan parasites) occurs
n extracellular forms of the parasite Toxoplasma gondii [21]. Inddition, the interaction of glycolytic enzymes with the internalace of the membrane may be achieved by acylation. For exam-le, palmitoylation of rabbit muscle phosphofructokinase (PFK)odulates the binding of this enzyme to the plasma membrane
22].Another possible mechanism that could play a role in sorting
f proteins into exosomes is through their interaction with pro-eins that are members of the tetraspanin family. These proteinsre part of membrane platforms in eukaryotes called tetraspanin-nriched microdomains. It has been suggested that such domainsound in exosomes can define the composition of the extracellularesicles [23]. Indeed, two tetraspanin proteins, CD81 and EWI-2,ere shown to recognize five glycolytic enzymes associated with
xosomes of human lymphoblasts: aldolase (ALD), GAPDH, phos-hoglycerate mutase (PGAM), ENO and pyruvate kinase (PYK) [23].
In summary, glycolytic enzymes could be driven to the extra-ellular milieu through secreted vesicles and, when released, alsoeassociate with the cell surface. This reassociation has been sug-ested for surface-bound ENO of Trichomonas vaginalis [24]. Despitehe evidence that glycolytic enzymes can be secreted throughxosomes, in the yeast Saccharomyces cerevisae it was clearlyemonstrated that ENO and glucose-6-phosphate isomerase (PGI)re secreted through a non-classical pathway driven by SNARE pro-eins, specifically the protein Tlg2p [25]. SNARE proteins participate
n the translocation of proteins between the Golgi apparatus andlasma membrane. Whether this secretion mechanism is comple-entary or related to that involving exosomes or shedding vesicless not yet known.
ous parasiteseinernema glaseri
[33][41]
4. Experimental evidence for the presence of extracellularglycolytic enzymes in parasitic protists and helminths
There is a growing amount of evidence for the presence ofglycolytic enzymes on the surface of several parasitic protistsand helminths. In addition, proteomic surveys of both groups ofparasites, involving determination of overall secretomes, secre-tion/excretion products, and purified secreted vesicles includingexosomes have also shown the presence of secreted glycolytic pro-teins. A detailed inventory of the relevant data as available in theliterature is given in the Supplementary Table S1. In almost all ofthese studies, ALD, GAPDH and ENO are the most commonly foundextracellular glycolytic proteins of parasites (Table 1 and Supple-mentary Table S1). In general, glycolytic enzymes can be associatedwith the parasite surface or present in the secretome, either withinsecreted vesicles – exosomes or shedding vesicles – or as freemolecules (Fig. 2). A parasite-derived glycolytic enzyme, GAPDH,has even been reported to be contained within secreted vesicles ofparasite-infected host cells [26]. Since secreted vesicles can mimicthe distribution of proteins in the cells from which they are derived,one may assume that glycolytic enzymes are also associated withthe external face of these vesicles (Fig. 2). Indeed, evidence concern-
Fig. 2. Schematic representation of export and localization of extracellular gly-colytic proteins in parasites. Exosomes or shedding vesicles may contain glycolyticproteins in their lumen or on their surface. Extracellular glycolytic proteins may alsobe present as free molecules or attached to the cell surface. These glycolytic proteinsare involved in various forms of parasite–host interactions.

7 Bioche
tletTb
5
crsm
5
tpemaceoPlrhitapttfibm
5
t[tiboaaawodgsiiptc
8 A. Gómez-Arreaza et al. / Molecular &
he cytoplasm. Interaction of the parasites with intestinal epithe-ial cells causes a considerable enhancement of the release of thisnzyme [28]. However, despite the large number of reports abouthe extracellular localization of glycolytic enzymes in parasites (seeable S1), there are only relatively few data about the roles exertedy these enzymes outside these organisms as discussed below.
. Functions of extracellular glycolytic enzymes of parasites
Parasite glycolytic enzymes may act as ligands for a variety ofomponents of the host. This binding would allow the parasites toecognize host cells for invasion, adhere to them, modulate haemo-tatic and immune systems, promote angiogenesis, or acquireolecules for nutrition.
.1. Adherence
Parasites employ a variety of strategies for recognition of hostissues and adherence to the cells they will invade. For exam-le, a peptide that binds to the surface of the mosquito midgutpithelium and inhibits the invasion of P. berghei was found to be aimetope of ENO detected on the surface of ookinetes of P. berghei
nd P. falciparum, suggesting that this extracellularly exposed gly-olytic enzyme has a function in recognizing and binding to anpithelial receptor [29]. Indeed anti-ENO antibodies inhibited theokinete invasion. Moreover, it was suggested that this function oflasmodium ENO might be shared with other extracellular moon-ighting functions, such as plasminogen recognition [29]. Similarly,elated to adherence and recognition of the extra cellular matrix, aeparin-binding motif is present in the 1,3-bisphosphoglycerate-
ndependent PGAM from Leishmania spp. [30]. It was suggestedhat this motif might be responsible for the interaction with hep-rin/heparin sulphate at the surface of host cells. This PGAM isresent in the secretome of these parasites [31] and could be athe cell surface. In another parasite, the sexually transmitted pro-ist T. vaginalis, the enzyme GAPDH, found at the surface, bindsbronectin [32], also a component of the extracellular matrix. Thisinding may be important to anchor the parasite to the extracellularatrix after disruption of the vaginal epithelium [32].
.2. Modulation of the haemostatic system
Parasites are able to interfere with the haemostatic system ofhe vertebrate host through interaction with fibrinolytic factors33]. This interaction involves the binding of plasminogen to pro-eins at the parasite surface or to secreted proteins. Plasminogens the zymogenic form of the serine protease plasmin, responsi-le for fibrin degradation in the blood clot after coagulation and inther physiological processes such as cell migration [34]. The inter-ction with plasminogen, recruited from the surrounding milieund then activated by its processing to plasmin, is known to playn important role in infection by pathogenic bacteria [9]. Like-ise, plasmin bound to surface-attached or secreted receptors
f parasites can help such protists and helminths to migrate byissolving blood clots, cross barriers and infect cells [33]. Plasmino-en binding, as demonstrated at least on the surface of vesiclesecreted from Leishmania mexicana [35], could also be part of anmmune evasion mechanism since plasmin can degrade antibod-
es. Three glycolytic enzymes, ENO, GAPDH and ALD, are among thelasminogen-binding proteins of parasites, a moonlighting func-ion they share with the corresponding enzymes from mammalianells, pathogenic fungi and bacteria.mical Parasitology 193 (2014) 75–81
5.3. Modulation of the immune system
The modulation and evasion of the immune response are sur-vival strategies used by both protists and helminths with a parasiticform of life. The mechanisms used to accomplish these strategiesare numerous [36,37]; they all involve effectors that are eitheron the surface of the parasites or molecules secreted by them.Glycolytic enzymes can be among the molecules that act as sucheffectors. For example, ALD can be part of the mechanisms toimpair macrophage function. It has been shown that this enzyme,secreted by L. donovani and imported into the cytoplasm of infectedmacrophages, interacts with the host cell’s protein tyrosine phos-phatase SHP-1 and so targets its signalling pathways [38]. It wasalso shown that ALD is released by L. donovani exosomes [31]. Theinteraction of SHP-1 with ALD could contribute to blocking thecapacity of the macrophages to mediate an immune response, thusfavouring the survival and replication of the parasites inside thephagolysosome. Indeed, ALD was demonstrated to be able to acti-vate SHP-1 in vitro. In addition, transfected macrophages express-ing the parasite’s ALD have an attenuated expression of nitric-oxidesynthase in response to interferon-� treatment [38]. Thus ALD isconsidered a secreted effector molecule of Leishmania delivered tothe host cell in order to modulate the immune response [39].
Another parasite glycolytic protein has been reported to beinvolved in a different kind of mechanism to modulate the immuneresponse. A GAPDH among the excretory/secretory products ofthe parasitic nematode Haemonchus contortus was identified asresponsible for a complement factor C3 binding activity [40]. C3 isrequired for the activation of each of the three complement path-ways – classical, alternative and lectin-dependent – of the innateimmune system in vertebrates. Binding of complement, acquired bythe parasite after a blood meal, could inhibit the formation of themembrane attack complex that would cause lysis of the parasite bydisruption of its cell membrane [40].
Finally, secreted ENO of the entomopathogenic helminth Stein-ernema glaseri has been suggested to suppress the immuneresponse in insects. It was shown that recombinant S. glaseri ENOaccelerated the infection by the pathogenic bacterium Xenorhab-dus poinarii and the fungus Metarhizium anisopliae in a bioassay forinfection of insect larvae [41].
5.4. Angiogenesis
In tumours, PGI that is known to act also as a cytokine, has a verywell-known function as autocrine motility factor (AMF); it stimu-lates the motility of cancer cells thus contributing to malignancy[42]. This PGI/cytokine promotes angiogenesis or formation of newblood vessels. Studies performed with recombinant PGI of the para-sitic helminth Echinococcus multilocularis showed that this proteinis able to stimulate division of endothelial cells in vitro [43]. PGIis found in the laminated layer at the surface of this parasite andequally in the vesicle fluid, that is an intraparasitic compartment ofthe larval form, the metacestode. It is hypothesized that PGI couldpromote angiogenesis around the metacestode in vivo, similarly towhat it does around tumour cells [43]. Moreover, recombinant E.multilocularis PGI enhances the proliferation of the parasite germi-nal layer cells in vitro suggesting also a role in the development ofthis worm.
5.5. Nutrient acquisition
Leishmania species acquire haem from the haemoglobin found
in the environment, since these parasites lack the synthesis path-way for this prosthetic group. In L. donovani, a haemoglobin-bindingprotein was found on the parasite’s surface in the flagellar pocket;it is possibly needed for endocytosis of the host protein [44]. This
Bioche
LaohThbs[eGt
5
pibcstWi
mecbelqtoigl
6l
ewgotllflrEppaeaaCbes
g
A. Gómez-Arreaza et al. / Molecular &
eishmania protein was identified as a hexokinase (HK); it is anuthentic HK, of which the sequence [45] aligns well with thatf other HKs such as those of Trypanosoma spp. and does notave an extra domain or strongly predicted transmembrane region.he N-terminal region of this Leishmania HK is responsible foraemoglobin binding. It is noteworthy that T. brucei is also capa-le of haemoglobin uptake from its host, but it employs a differenttrategy not involving a glycolytic protein as extracellular receptor46]. The utilization by a cell of a glycolytic protein to acquire nutri-nts from the environment is not without precedents. The enzymeAPDH of the bacterium Staphylococcus aureus acts as receptor in
he acquisition of transferrin-bound iron [47].
.6. Some hypothetical functions
Extracellular glycolytic enzymes have been linked to the apo-tosis process in some types of mammalian cells [48]. Moreover,
t has been proposed that apoptosis mimicry in pathogens coulde the effect of exposed glycolytic enzymes [48]. Apoptosis-likeell death has been recognized in unicellular parasites. It has beenuggested to mimic apoptotic features of multicellular organism toake advantage of the non-inflammatory response in the host [49].
hether apoptosis mimicry and externalized glycolytic enzymesn parasites are linked remains to be determined.
Because many glycolytic enzymes are found extracellularly, oneay wonder if such parasites have a functional complete or partial
xtracellular glycolytic route. The same question has been raisedoncerning the presence of extracellular glycolytic enzymes ofacteria [10]. For such speculative function to be carried out, sev-ral conditions must converge: the enzymes must be active andocated in a special environment within the host with the ade-uate concentration of substrates. To date, no data are availablehat support such a function for these enzymes outside pathogenicrganisms. Moreover, surface-bound ENO has been found inactiven Leishmania parasites [50,51]. However, a functional extracellularlycolytic pathway has been reported in prostasomes, exosome-ike microvesicles derived from the prostate [19].
. Evolutionary considerations about the extracellularocation of glycolytic enzymes
Four important questions arise about moonlighting glycolyticnzymes from an evolutionary point of view. First, one mayonder how moonlighting activities have arisen in proteins in
eneral and secondly, why and how this happened particularlyften in glycolytic enzymes. Third, and also pertinent for theopic of this review: how have intracellular enzymes during evo-ution succeeded in developing routes to reach an extracellularocalization where they exert their moonlighting functions? Andourth, why have similar glycolytic enzymes in different taxonomicineages, and notably in pathogenic organisms, independentlyecruited the same moonlighting function, such as, for example,NO that in many different organisms has become an extracellularlasminogen-binding protein? Indeed, there is no doubt that thearticipation in glycolysis/gluconeogenesis are the original and inlmost all extant organisms the predominant functions of glycolyticnzymes. The glycolytic/gluconeogenic functions of these proteinsre shared by organisms from all domains of life, and therefore mustlready have existed in the common ancestor of all these organisms.onsequently, moonlighting activities of glycolytic enzymes muste considered as “novelties from ancient material” that in differ-
nt organisms were independently ‘recruited’, often to execute theame physiological function.A protein with a new function is supposed to arise usually by aene duplication followed by modification of one of the duplicates
mical Parasitology 193 (2014) 75–81 79
as a result of accumulation of mutations and recombinations withother genes or gene fragments, while the other duplicate evolvesless drastically in order to conserve its original function. In addi-tion, sequences encoding new regulatory fragments may be addedto the modified gene to change the regulation of its expression orthe cellular localization of the new protein. If the modified proteinprovides an advantage to the organism, it is retained by naturalselection. Does this mechanism also play a role in the origin andevolution of moonlighting functions? In some cases this may be thecase. One copy among multiple genes may have changed, obtain-ing a new function without loosing much of its original identity sothat it is – correctly or not – still recognized as a glycolytic enzyme.Of course, if such a protein has mutated to such an extent that ithas lost its glycolytic function, the new, acquired activity cannot bedefined as a moonlighting one; it has become the only function ofthe protein; the identity of the protein has changed. In this case,evolution most likely will then rapidly erase the glycolysis-specificmotifs because the selective force to retain them has disappeared.A corollary of this scenario is that probably the presence of multipledifferent gene copies with only one of them having a moon-lighting function in addition to the original function representsa recent evolutionary process. However, one finds moonlightingfunctions in a glycolytic enzyme encoded by only a single geneor alternatively, as found in, for example, trypanosomatid para-sites, multiple, tandemly arranged genes coding for the same oralmost identical proteins [52]. This latter situation is the result ofregular gene conversions within the tandem repeats. When onlya single product with moonlighting functions is obtained fromone or multiple (nearly) identical genes, the non-glycolytic activ-ities may theoretically be attributed to either post-translationalmodification of part of the protein molecules, or to the presenceof features in the enzymes that provide new functions that arecompatible with the glycolytic functions. These features may beintrinsic to the glycolytic enzymes or have specifically evolved incertain cases. Examples of intrinsic features in glycolytic enzymesthat may have been exploited for moonlighting functions in otherprocesses are sites with affinities for ligands that participate in bothglycolysis and the other processes, such as nucleotides (e.g. cofac-tor NAD- and ATP-binding sites in dehydrogenases and kinases)and sugar- or sugar-phosphate-binding sites (in sugar kinases andsugar-phosphate isomerases). The cofactor-binding sites may alsobind other nucleotides or nucleic acids, while sugar(-phosphate)-binding sites may bind carbohydrates or glycoconjugates.
Alternatively, some features may have specifically evolved inglycolytic enzymes for use in moonlighting functions. This can beenvisaged for areas of the enzymes which do not have any specificfunction other than linking regions with defined function or sec-ondary structure, or for areas which are involved in protein–proteininteractions. In the latter case, it seems relevant that glycolyticenzymes are often associated together in specific configurationswithin multi-enzyme complexes and/or associated with intracel-lular macromolecular structures such as cytoskeleton proteins or(proteins in) membranes [1]. These protein-interaction regions ofglycolytic enzymes may have been exploited, during evolution, foralso making interactions with other proteins – the moonlightingfunctions – within the cell or extracellularly. Such regions have thenevolved and may even have been selected for dual or even multiplefunctions, such as glycolytic enzymes that have acquired multi-ple, extracellular receptor functions. The enzymes may also haveobtained additional regions or domains providing them the abilityto bind specific ligands extracellularly, without interfering with theglycolytic activity when the enzymes remain intracellularly.
As reported in other sections, glycolytic enzymes have beendetected within secreted vesicles or associated with the membraneof such organelles, at least in some cases exposed at their outer face.If glycolytic enzymes with potential moonlighting functions to be

8 Bioche
ecflb[ipovoaioto
dmfttteehratci–Itmtsoofsenaft
7v
bgdteteamtbirmt
0 A. Gómez-Arreaza et al. / Molecular &
xerted extracellularly are randomly, as part of the cytoplasm –ytosol and organelles – sequestered in the vesicles during theirormation, or specifically enriched therein remains to be estab-ished. There are several reasons why glycolytic enzymes may haveeen recruited readily for extracellular moonlighting functions33]. First, glycolytic enzymes are ubiquitous and usually presentn the cell at relatively high concentrations. Secondly, they areresent in the cytosol – or in the case of trypanosomes inside smallrganelles, the glycosomes – and are easily taken up in secretedesicles unlike proteins of larger organelles such as the nucleusr mitochondria. Nonetheless, it is also possible that the enzymesre enriched in the vesicles as a result of evolutionary acquisitionsnvolving a propensity for specific post-translational modificationsr specific sequence motifs that target the enzymes to vesicles. Thisargeting may be towards specific (but hypothetical) soluble matrixr membrane proteins of the vesicles to be formed.
It is noteworthy that corresponding enzymes are often, in highlyiverse (pathogenic or not) organisms, found to execute similaroonlighting functions. The most striking example is ENO, that is
ound as plasminogen receptor in many pathogenic bacteria, pro-ists and helminths and mammalian cells such as tumour cells,o facilitate migration, and crossing barriers, and thus to increasehe virulence. As argued above, this cannot be due to divergentvolution; the common function of ENO in organisms of all lin-ages is glycolysis/gluconeogenesis, and this activity must thusave been developed in the common ancestor. The plasminogen-eceptor function can only have been acquired late in evolution,fter the fibrinolytic system evolved in vertebrates. The acquisi-ion of plasminogen-binding in different organisms is thus due toonvergent evolution. It is most likely that features in the ENOmportant for the intracellular function could easily be recruited
probably by slight modification – to function also extracellularly.t can be imagined that a protein-interaction motif (for bindingo other glycolytic enzymes) could easily also be used for plas-
inogen binding. As a consequence, such a binding motif becamehe object of evolutionary optimization by selective forces actingimultaneously on the intracellular and extracellular functioningf ENO. The dual function was retained in all organisms where itriginated, but the evolutionary optimization of the binding sur-ace may have resulted in slight differences. Similar evolutionarycenarios could also be envisaged for other (glycolytic) enzymesxerting similar (extracellular) functions in highly different orga-isms. However, even related parasites may also have developedlternative strategies to exert similar functions, as illustrated aboveor haem acquisition by Leishmania and Trypanosoma [44,46]; onlyhe former species employs a glycolytic enzyme for it, HK.
. Potential implications for diagnosis and treatments –accination/chemotherapy
The extracellular glycolytic enzymes of parasites are known toe responsible for invasion, establishment in the host, and patho-enesis. Thus, these functions could serve as a basis for developingiagnostics as well as different types of therapeutic and prophylac-ic treatments. Indeed some studies have provided experimentalvidence that glycolytic enzymes can be used as antigens for pro-ective vaccination, although it is not clear which function of thenzymes is being targeted. For example, ENO has been proposed as
potential protective antigen since anti-ENO antibodies can protectice against the malarial parasite P. yoelii [53]. A similar situa-
ion has been described for ENO from Ascaris suum, where it haseen shown that ENO elicited a specific immune response in mice
mmunized with this enzyme, reducing the number of larvae thateached the liver and lungs after challenge infections in an A. suum-ouse model [54]. Since anti-ENO antibodies are also able to block
he invasion of Plasmodium spp. in the mosquito, ENO has been
mical Parasitology 193 (2014) 75–81
suggested as well as an antigen for a transmission blocking vac-cine [29]. Moreover, the ENO – plasminogen interaction peptide(EPIP), found to disrupt the interaction between the extracellu-lar enzyme and the fibrinolytic factor in Plasmodium spp. [55], hasbeen proposed as anti-plasmodium effector molecule to generaterefractory Anopheles mosquitoes incapable of sustaining parasitedevelopment [56].
Similarly, immunization of mice with E. multilocularis PGIresulted in a decrease of the metacestodes burden, after infection,by suppression of parasite development [35]. For leishmaniasis, thepotential of HK as a vaccine candidate was tested, based on thefinding of the enzyme’s function as a haemoglobin receptor in Ldonovani. Protection against L. donovani infection was obtained foran experimental period of 8 months after vaccination of mice andhamsters with HK DNA (full-length and N-terminal haemoglobin-binding domain) constructs [37].
Some glycolytic enzymes are well conserved between humanand their parasites and therefore one may wonder if they will besuitable for diagnostic purposes, in screening sera of (potentially)infected people; unique epitopes may have to be selected in suchcases. However other glycolytic proteins are poorly conserved; forinstance the PFK of trypanosomatids and humans show less than25% amino-acid sequence identity, while PGAM of these and sev-eral other parasites is not homologous to the enzyme catalyzing thecorresponding reaction in human. Such proteins may thus be goodcandidates for serological diagnosis. Indeed, glycolytic enzymeshave already been used as indicators of infection by parasites suchas P. falciparum [57]. In addition, glycolytic enzymes associated withsecreted vesicles of parasites could specifically serve for early diag-nosis and as biomarkers to determine virulence. The potential ofexosomes and other microvesicles as markers for parasitic diseasesis high since they can be captured from several body fluids [58].
8. Considerations for future research
Although an increasing number and diversity of moonlightingfunctions of glycolytic enzymes in eukaryotic and prokaryotic cellshas become apparent in the last few years, our present knowl-edge of the function of extracellular glycolytic proteins in parasitesresults so far from only a limited number of studies. In the yearsto come, combined proteomic surveys and inventories of secretedproteins of different parasitic organisms, and detailed functionalanalyses of each enzyme detected is required. One way for begin-ning to reveal additional functions, and the details of how thefunctions are exerted, is through a screening of binding propertiesagainst a variety of molecules in vitro. Additionally, other questionsremain to be answered: (i) How do the glycolytic enzymes reach thecell surface? (ii) How are they sorted into or onto exosomes or othertypes of vesicles? (iii) How are these enzymes arranged/locatedwithin the secreted vesicles? (iv) What is the activity of these extra-cellular glycolytic enzymes? (v) Which regions in the protein areresponsible for the moonlighting functions? (vi) How are extracel-lular enzymes attached to the surface of the parasites?
Acknowledgements
This study was supported by Fonacit, Venezuela, grant MC-2007000960. We apologize to those researchers whose work couldnot be cited here due to space limitation. However, an extensivereference list is provided with Table S1 in the Supplementary File.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at http://dx.doi.org/10.1016/j.molbiopara.2014.02.005.

Bioche
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[57] Mouatcho JC, Goldring JP. Malaria rapid diagnostic tests: challenges and
A. Gómez-Arreaza et al. / Molecular &
eferences
[1] Srere PA, Knull HR. Location-location-location. Trends Biochem Sci1998;23:319–20.
[2] Gualdrón-López M, Brennand A, Hannaert V, Quinones W, Cáceres AJ, BringaudF, et al. When, how and why glycolysis became compartmentalised in theKinetoplastea. A new look at an ancient organelle. Int J Parasitol 2012;42:1–20.
[3] Haanstra JR, van Tuijl A, van Dam J, van Winden W, Tielens AG, van Hellemond JJ,et al. Proliferating bloodstream-form Trypanosoma brucei use a negligible partof consumed glucose for anabolic processes. Int J Parasitol 2012;42:667–73.
[4] Roth Jr E. Plasmodium falciparum carbohydrate metabolism: a connectionbetween host cell and parasite. Blood Cells 1990;16:453–60.
[5] Sriram G, Martinez JA, McCabe ER, Liao JC, Dipple KM. Single-gene disorders:what role could moonlighting enzymes play. Am J Hum Genet 2005;76:911–24.
[6] Kim JW, Dang CV. Multifaceted roles of glycolytic enzymes. Trends Biochem Sci2005;30:142–50.
[7] Jeffery CJ. Moonlighting proteins. Trends Biochem Sci 1999;24:8–11.[8] Pancholi V, Fischetti VA. A major surface protein on group A Streptococci is a
glyceraldehyde-3-phosphate-dehydrogenase with multiple binding activity. JExp Med 1992;176:415–26.
[9] Sun H. The interaction between pathogens and the host coagulation system.Physiology (Bethesda) 2006;21:281–8.
10] Henderson B, Martin A. Bacterial virulence in the moonlight: multitasking bac-terial moonlighting proteins are virulence determinants in infectious disease.Infect Immun 2011;79:3476–91.
11] Tristan C, Shahani N, Sedlak TW, Sawa A. The diverse functions of GAPDH: viewsfrom different subcellular compartments. Cell Signal 2011;23:317–23.
12] Pancholi V, Multifunctional. �-Enolase: its role in diseases. Cell Mol Life Sci2001;58:902–20.
13] Avilán L, Gualdrón-López M, Quinones W, González-González L, Hannaert V,Michels PA, et al. Enolase: a key player in the metabolism and a probablevirulence factor of trypanosomatid parasites-perspectives for its use as a ther-apeutic target. Enzyme Res 2011;2011:932549.
14] Díaz-Ramos A, Roig-Borrellas A, García-Melero A, López-Alemany R. �-Enolase,a multifunctional protein: its role on pathophysiological situations. J BiomedBiotechnol 2012;2012:156795.
15] Collingridge PW, Brown RW, Ginger ML. Moonlighting enzymes in parasiticprotozoa. Parasitology 2010;137:1467–75.
16] Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immuneresponses. Nat Rev Immunol 2009;9:581–93.
17] Mathivanan S, Ji H, Simpson RJ. Exosomes: extracellular organelles importantin intercellular communication. J Proteomics 2010;73:1907–20.
18] Deatherage BL, Cookson BT. Membrane vesicle release in bacteria, eukaryotes,and archaea: a conserved yet underappreciated aspect of microbial life. InfectImmun 2012;80:1948–57.
19] Barteneva NS, Maltsev N, Vorobjev IA. Microvesicles and intercellular commu-nication in the context of parasitism. Front Cell Infect Microbiol 2013;3:49.
20] Raikar LS, Vallejo J, Lloyd PG, Hardin CD. Overexpression of caveolin-1 resultsin increased plasma membrane targeting of glycolytic enzymes: the struc-tural basis for a membrane associated metabolic compartment. J Cell Biochem2006;98:861–71.
21] Pomel S, Luk FC, Beckers CJ. Host cell egress and invasion induce marked relo-cations of glycolytic enzymes in Toxoplasma gondii tachyzoites. PLoS Pathog2008;4:e1000188.
22] Jenkins CM, Yang J, Sims HF, Gross RW. Reversible high affinity inhibition ofphosphofructokinase-1 by acyl-CoA: a mechanism integrating glycolytic fluxwith lipid metabolism. J Biol Chem 2011;286:11937–50.
23] Perez-Hernandez D, Gutiérrez-Vázquez C, Jorge I, López-Martín S, Ursa A,Sánchez-Madrid F, et al. The intracellular interactome of tetraspanin-enrichedmicrodomains reveals their function as sorting machineries toward exosomes.J Biol Chem 2013;288:11649–61.
24] Mundodi V, Kucknoor AS, Alderete JF. Immunogenic and plasminogen-binding surface-associated �-enolase of Trichomonas vaginalis. Infect Immun2008;76:523–31.
25] Miura N, Kirino A, Endo S, Morisaka H, Kuroda K, Takagi M, et al. Tracing putativetrafficking of the glycolytic enzyme enolase via SNARE-driven unconventionalsecretion. Eukaryot Cell 2012;11:1075–82.
26] Mantel PY, Hoang AN, Goldowitz I, Potashnikova D, Hamza B, Vorobjev I, et al.Malaria-infected erythrocyte-derived microvesicles mediate cellular commu-nication within the parasite population and with the host immune system. CellHost Microbe 2013;13:521–34.
27] Ronquist KG, Ek B, Stavreus-Evers A, Larsson A, Ronquist G. Human prostasomesexpress glycolytic enzymes with capacity for ATP production. Am J PhysiolEndocrinol Metab 2013;304:E576–82.
28] Ringqvist E, Palm JE, Skarin H, Hehl AB, Weiland M, Davids BJ, et al. Releaseof metabolic enzymes by Giardia in response to interaction with intestinalepithelial cells. Mol Biochem Parasitol 2008;159:85–91.
29] Ghosh AK, Coppens I, Gårdsvoll H, Ploug M, Jacobs-Lorena M. Plasmodium
ookinetes coopt mammalian plasminogen to invade the mosquito midgut. ProcNatl Acad Sci USA 2011;108:17153–8.30] Peysselon F, Launay G, Lisacek F, Duclos B, Ricard-Blum S. Comparative anal-ysis of Leishmania exoproteomes: implication for host-pathogen interactions.Biochim Biophys Acta 2013;1834:2653–62.
[
mical Parasitology 193 (2014) 75–81 81
31] Silverman JM, Clos J, de’Oliveira CC, Shirvani O, Fang Y, Wang C, et al. Anexosome-based secretion pathway is responsible for protein export fromLeishmania and communication with macrophages. J Cell Sci 2010;123:842–52.
32] Lama A, Kucknoor A, Mundodi V, Alderete JF. Glyceraldehyde-3-phosphatedehydrogenase is a surface-associated, fibronectin-binding protein of Tri-chomonas vaginalis. Infect Immun 2009;77:2703–11.
33] Figuera L, Gómez-Arreaza A, Avilán L. Parasitism in optima forma: exploitingthe host fibrinolytic system for invasion. Acta Trop 2013;128:116–23.
34] Castellino FJ, Ploplis VA. Structure and function of the plasminogen/plasminsystem. Thromb Haemost 2005;93:647–54.
35] Figuera L, Acosta H, Gómez-Arreaza A, Dávila-Vera D, Balza-Quintero A,Quinones W, et al. Plasminogen binding proteins in secreted membrane vesi-cles of Leishmania mexicana. Mol Biochem Parasitol 2013;187:14–20.
36] Zambrano-Villa S, Rosales-Borjas D, Carrero JC, Ortiz-Ortiz L. How pro-tozoan parasites evade the immune response. Trends Parasitol 2002;18:272–8.
37] Kamal SM, El Sayed Khalifa K. Immune modulation by helminthic infections:worms and viral infections. Parasite Immunol 2006;28:483–96.
38] Nandan D, Tran T, Trinh E, Silverman JM, Lopez M. Identification of leishmaniafructose-1,6-bisphosphate aldolase as a novel activator of host macrophage Srchomology 2 domain containing protein tyrosine phosphatase SHP-1. BiochemBiophys Res Commun 2007;364:601–7.
39] Lambertz U, Silverman JM, Nandan D, McMaster WR, Clos J, Foster LJ, et al.Secreted virulence factors and immune evasion in visceral leishmaniasis. JLeukoc Biol 2012;91:887–99.
40] Sahoo S, Murugavel S, Devi IK, Vedamurthy GV, Gupta SC, Singh BP,et al. Glyceraldehyde-3-phosphate dehydrogenase of the parasitic nematodeHaemonchus contortus binds to complement C3 and inhibits its activity. ParasiteImmunol 2013;35:457–67.
41] Liu H, Zeng H, Yao Q, Yuan J, Zhang Y, Qiu D, et al. Steinernema glaseri sur-face enolase: molecular cloning, biological characterization, and role in hostimmune suppression. Mol Biochem Parasitol 2012;185:89–98.
42] Furuta E, Okuda H, Kobayashi A, Watabe K. Metabolic genes in cancer: theirroles in tumor progression and clinical implications. Biochim Biophys Acta2010;1805:141–52.
43] Stadelmann B, Spiliotis M, Müller J, Scholl S, Müller N, Gottstein B, et al.Echinococcus multilocularis phosphoglucose isomerase (EmPGI): a glycolyticenzyme involved in metacestode growth and parasite–host cell interactions.Int J Parasitol 2010;40:1563–74.
44] Krishnamurthy G, Vikram R, Singh SB, Patel N, Agarwal S, Mukhopadhyay G,et al. Hemoglobin receptor in Leishmania is a hexokinase located in the flagellarpocket. J Biol Chem 2005;280:5884–91.
45] Guha R, Gupta D, Rastogi R, Vikram R, Krishnamurthy G, Bimal S, et al. Vacci-nation with leishmania hemoglobin receptor-encoding DNA protects againstvisceral leishmaniasis. Sci Transl Med 2013;5:202ra121.
46] Vanhollebeke B, De Muylder G, Nielsen MJ, Pays A, Tebabi P, Dieu M, et al. Ahaptoglobin–hemoglobin receptor conveys innate immunity to Trypanosomabrucei in humans. Science 2008;320:677–81.
47] Modun B, Morrissey J, Williams P. The staphylococcal transferrin receptor: aglycolytic enzyme with novel functions. Trends Microbiol 2000;8:231–7.
48] Ucker DS, Jain MR, Pattabiraman G, Palasiewicz K, Birge RB, Li H. Externalizedglycolytic enzymes are novel, conserved, and early biomarkers of apoptosis. JBiol Chem 2012;287:10325–43.
49] Lüder CG, Campos-Salinas J, Gonzalez-Rey E, van Zandbergen G. Impact ofprotozoan cell death on parasite–host interactions and pathogenesis. ParasitVectors 2010;3:116.
50] Quinones W, Pena P, Domingo-Sananes M, Cáceres A, Michels PA, Avilan L, et al.Leishmania mexicana: molecular cloning and characterization of enolase. ExpParasitol 2007;116:241–51.
51] Swenerton RK, Zhang S, Sajid M, Medzihradszky KF, Craik CS, Kelly BL, et al. Theoligopeptidase B of Leishmania regulates parasite enolase and immune evasion.J Biol Chem 2011;286:429–40.
52] Michels PAM, Hannaert V, Ernest I, Allert S. Organization, structure and evo-lutionary aspects of genes in trypanosomes. Biochem Soc Trans 1990;18:727–9.
53] Pal-Bhowmick I, Mehta M, Coppens I, Sharma S, Jarori GK. Protective proper-ties and surface localization of Plasmodium falciparum enolase. Infect Immun2007;75:5500–8.
54] Chen N, Yuan ZG, Xu MJ, Zhou DH, Zhang XX, Zhang YZ, et al. Ascarissuum enolase is a potential vaccine candidate against ascariasis. Vaccine2012;30:3478–82.
55] Wang S, Ghosh AK, Bongio N, Stebbings KA, Lampe DJ, Jacobs-Lorena M. Fightingmalaria with engineered symbiotic bacteria from vector mosquitoes. Proc NatlAcad Sci USA 2012;109:12734–9.
56] Wang S, Jacobs-Lorena M. Genetic approaches to interfere with malaria trans-mission by vector mosquitoes. Trends Biotechnol 2013;31:185–93.
prospects. J Med Microbiol 2013;62:1491–505.58] Vlassov AV, Magdaleno S, Setterquist R, Conrad R. Exosomes: current knowl-
edge of their composition, biological functions, and diagnostic and therapeuticpotentials. Biochim Biophys Acta 2012;1820:940–8.
Related Documents











