Neuron, Vol. 35, 951–960, August 29, 2002, Copyright 2002 by Cell Press Extinction Antagonizes Olfactory Memory at the Subcellular Level and vertical MB lobes (Figure 1). The fine structure in the lobes defines three classes of Kenyon cells in distinct subsystems (/; /; and ; Crittenden et al., 1998; Martin Schwaerzel, Martin Heisenberg, and Troy Zars 1,2 Theodor Boveri Institut fu ¨ r Biowissenschaften Heisenberg, 1998; Zars, 2000). Efferent neurons from Lehrstuhl fu ¨ r Genetik und Neurobiologie the MBs have been identified which project to various Biozentrum anterior neuropil regions, contra- and ipsilateral MB Am Hubland, D-97074 lobes, and the lateral horn (Ito et al., 1998). From there, Wu ¨ rzburg large descending neurons relay signals through the cer- Germany vical connective to the ventral ganglion, as suggested from a report in larger flies (Strausfeld et al., 1984). The cAMP 2 nd -messenger cascade is important for Summary synaptic plasticity and memory formation (Byrne and Kandel, 1996; Lechner and Byrne, 1998). In general, Memory loss occurs by diverse mechanisms, as differ- cAMP is synthesized by a type 1 adenylyl cyclase (AC) ent time constants of performance decrement and in response to the coincidence of two signals, activated sensitivities to experimental manipulations suggest. heterotrimeric G protein mediating the unconditioned While the phenomena of memory decay, interference, stimulus (US) and Ca 2 /calmodulin (CAM) signaling ac- and extinction are well established behaviorally, little tivity in the pathway of the conditioned stimulus (CS). is known about them at the circuit or molecular level. This rise in cAMP is thought to activate protein kinase In Drosophila, odorant memories lasting up to 3 hr can A (PKA) and leads to a release of its regulatory subunits be localized to mushroom body Kenyon cells, a single (and eventually their degradation). The catalytic subunit neuronal level in the olfactory pathway. The plasticity of PKA can then phosphorylate various targets such as underlying this memory trace can be induced without ion channels and synaptic vesicle release machinery. Kenyon cell synaptic output. Experimental extinction, This protein phosphorylation is thought to mediate the i.e., presentation of the conditioned stimulus without short-term molecular changes underlying both synaptic the reinforcer, reduces memory performance and plasticity and memory formation. does so at the same circuit level as memory formation. In Drosophila, the cAMP signaling cascade has been Thus, unreinforced presentation of learned odorants shown to be important for synaptic plasticity and mem- antagonizes intracellularly the signaling cascade un- ory formation using mutations or transgenic manipula- derlying memory formation. tion all along the cAMP pathway: the G protein subunit, the type 1 AC, a cAMP-phosphodiesterase, PKA, and a Introduction potassium channel (as a putative substrate for PKA) (Connolly et al., 1996; Cowan and Siegel, 1986; Drain et Most of our memories are doomed, leaving us eventually al., 1991; Han et al., 1992; Levin et al., 1992; Nighorn et al., with vague, altered, or no recollections of what has hap- 1991; Qiu et al., 1991; Skoulakis et al., 1993; Zars et al., pened. Different processes may underlie such memory 2000a). There are several other molecules known to be loss. Besides decay, memory traces may be masked by involved in olfactory learning and memory in Drosophila interference with the consolidation process, or super- (e.g., Cheng et al., 2001; Grotewiel et al., 1998), indicat- ceded by an additional memory trace (re-learning; re- ing that our knowledge of the molecular machinery un- viewed in Bouton et al., 1999). While memory loss inher- derlying synaptic plasticity is incomplete. ent to the system (as above) is intensely studied For olfactory short-term memory formation, the cAMP behaviorally, little is known of the underlying molecular cascade is needed in a specific part of the olfactory or cellular mechanisms. We investigate here extinction pathway. Earlier experiments already indicated that the of a memory trace in a differential olfactory conditioning MBs are necessary for olfactory memory formation paradigm of the fly Drosophila melanogaster. (Connolly et al., 1996; de Belle and Heisenberg, 1994; Olfactory processing in insects follows a well-charac- Heisenberg et al., 1985). In addition, providing the wild- terized anatomical pathway. Odorant stimuli are re- type Rut-AC exclusively in the Kenyon cells of mutant ceived on the antennae and maxillary palps by receptor rutabaga (rut) flies restored to normal their short-term neurons projecting to the antennal lobes (Stocker, 1994; olfactory memory defect (Zars et al., 2000a). More evi- Strausfeld, 1976, 1998). There they meet two broad cate- dence that olfactory memory is stored in Kenyon cells gories of interneurons: antennal lobe intrinsic neurons came from the amnesiac (amn) gene, which is required and projection neurons. The latter extend to the lateral for middle-term memory (in the range of hours). Rescue horn and the calyces, the site of dendritic input to the of amn in the so-called DPM cells that intimately inner- mushroom bodies (MBs). The intrinsic neurons of the vate the MB lobes and anterior peduncle restores mid- MBs, the Kenyon cells, send their axons to the anterior dle-term memory (Waddell et al., 2000). Because one of part of the brain where some bifurcate to form the medial the amn products has some homology to the PACAP neuropeptide (Feany and Quinn, 1995) and this is thought to modulate AC function (Zhong, 1995), the amn product 1 Correspondence: [email protected] may prolong Rut-AC activation in the MBs to extend olfac- 2 Present address: Division of Biology, University of Missouri-Colum- bia, Columbia, Missouri 65211. tory memory. Finally, use of the temperature-dependent

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neuron, Vol. 35, 951–960, August 29, 2002, Copyright 2002 by Cell Press
Extinction Antagonizes OlfactoryMemory at the Subcellular Level
and vertical MB lobes (Figure 1). The fine structure inthe lobes defines three classes of Kenyon cells in distinctsubsystems (�/�; ��/��; and �; Crittenden et al., 1998;
Martin Schwaerzel, Martin Heisenberg,and Troy Zars1,2
Theodor Boveri Institut fur BiowissenschaftenHeisenberg, 1998; Zars, 2000). Efferent neurons fromLehrstuhl fur Genetik und Neurobiologiethe MBs have been identified which project to variousBiozentrumanterior neuropil regions, contra- and ipsilateral MBAm Hubland, D-97074lobes, and the lateral horn (Ito et al., 1998). From there,Wurzburglarge descending neurons relay signals through the cer-Germanyvical connective to the ventral ganglion, as suggestedfrom a report in larger flies (Strausfeld et al., 1984).
The cAMP 2nd-messenger cascade is important forSummarysynaptic plasticity and memory formation (Byrne andKandel, 1996; Lechner and Byrne, 1998). In general,Memory loss occurs by diverse mechanisms, as differ-cAMP is synthesized by a type 1 adenylyl cyclase (AC)ent time constants of performance decrement andin response to the coincidence of two signals, activatedsensitivities to experimental manipulations suggest.heterotrimeric G protein mediating the unconditionedWhile the phenomena of memory decay, interference,stimulus (US) and Ca2�/calmodulin (CAM) signaling ac-and extinction are well established behaviorally, littletivity in the pathway of the conditioned stimulus (CS).is known about them at the circuit or molecular level.This rise in cAMP is thought to activate protein kinaseIn Drosophila, odorant memories lasting up to 3 hr canA (PKA) and leads to a release of its regulatory subunitsbe localized to mushroom body Kenyon cells, a single(and eventually their degradation). The catalytic subunitneuronal level in the olfactory pathway. The plasticityof PKA can then phosphorylate various targets such asunderlying this memory trace can be induced withoution channels and synaptic vesicle release machinery.Kenyon cell synaptic output. Experimental extinction,This protein phosphorylation is thought to mediate thei.e., presentation of the conditioned stimulus withoutshort-term molecular changes underlying both synapticthe reinforcer, reduces memory performance andplasticity and memory formation.does so at the same circuit level as memory formation.
In Drosophila, the cAMP signaling cascade has beenThus, unreinforced presentation of learned odorantsshown to be important for synaptic plasticity and mem-antagonizes intracellularly the signaling cascade un-ory formation using mutations or transgenic manipula-derlying memory formation.tion all along the cAMP pathway: the G protein � subunit,the type 1 AC, a cAMP-phosphodiesterase, PKA, and aIntroductionpotassium channel (as a putative substrate for PKA)(Connolly et al., 1996; Cowan and Siegel, 1986; Drain et
Most of our memories are doomed, leaving us eventuallyal., 1991; Han et al., 1992; Levin et al., 1992; Nighorn et al.,
with vague, altered, or no recollections of what has hap-1991; Qiu et al., 1991; Skoulakis et al., 1993; Zars et al.,
pened. Different processes may underlie such memory 2000a). There are several other molecules known to beloss. Besides decay, memory traces may be masked by involved in olfactory learning and memory in Drosophilainterference with the consolidation process, or super- (e.g., Cheng et al., 2001; Grotewiel et al., 1998), indicat-ceded by an additional memory trace (re-learning; re- ing that our knowledge of the molecular machinery un-viewed in Bouton et al., 1999). While memory loss inher- derlying synaptic plasticity is incomplete.ent to the system (as above) is intensely studied For olfactory short-term memory formation, the cAMPbehaviorally, little is known of the underlying molecular cascade is needed in a specific part of the olfactoryor cellular mechanisms. We investigate here extinction pathway. Earlier experiments already indicated that theof a memory trace in a differential olfactory conditioning MBs are necessary for olfactory memory formationparadigm of the fly Drosophila melanogaster. (Connolly et al., 1996; de Belle and Heisenberg, 1994;
Olfactory processing in insects follows a well-charac- Heisenberg et al., 1985). In addition, providing the wild-terized anatomical pathway. Odorant stimuli are re- type Rut-AC exclusively in the Kenyon cells of mutantceived on the antennae and maxillary palps by receptor rutabaga (rut) flies restored to normal their short-termneurons projecting to the antennal lobes (Stocker, 1994; olfactory memory defect (Zars et al., 2000a). More evi-Strausfeld, 1976, 1998). There they meet two broad cate- dence that olfactory memory is stored in Kenyon cellsgories of interneurons: antennal lobe intrinsic neurons came from the amnesiac (amn) gene, which is requiredand projection neurons. The latter extend to the lateral for middle-term memory (in the range of hours). Rescuehorn and the calyces, the site of dendritic input to the of amn in the so-called DPM cells that intimately inner-mushroom bodies (MBs). The intrinsic neurons of the vate the MB lobes and anterior peduncle restores mid-MBs, the Kenyon cells, send their axons to the anterior dle-term memory (Waddell et al., 2000). Because one ofpart of the brain where some bifurcate to form the medial the amn products has some homology to the PACAP
neuropeptide (Feany and Quinn, 1995) and this is thoughtto modulate AC function (Zhong, 1995), the amn product1Correspondence: [email protected] prolong Rut-AC activation in the MBs to extend olfac-2 Present address: Division of Biology, University of Missouri-Colum-
bia, Columbia, Missouri 65211. tory memory. Finally, use of the temperature-dependent

Neuron952
Figure 1. The Mushroom Bodies of Drosophila
The mushroom bodies are a paired neuropil structure with a dendritic input (the calyx) and axonal projections that proceed anterior in thebrain where many of the intrinsic neurons (Kenyon cells) bifurcate to form the vertical and medial lobes. (A) The GAL4 driver 247 was usedin all behavioral experiments. Its expression pattern is shown here with an UAS-lacZ reporter, false colored in yellow. The brain neuropil wasvisualized with the a monoclonal antibody (nc82) recognizing synapses and is false colored with blue to green to give a sense of depth (topimage). The line 247 is largely specific to Kenyon cells (A, bottom image). Low levels of expression can be detected in some central brainglial cells (not evident here). (B and C) Quantification of Kenyon cell number using the UAS-nls-lacZ reporter in the line 247 revealedapproximately 700 cells labeled in males while in c772 males, approximately 850 Kenyon cells express GAL4. Scale bar represents 50 �m in(A) and 25 �m in (B) and (C). Abbreviations: ca � calyx, vl � vertical lobes, ml � medial lobes, p � peduncle.
blocker of synaptic transmission, shibirets1 (shi ts1, Kita- but, in addition, has expression in scattered neurons inthe central brain outside the MBs, including fibers in themoto, 2001) shows that Kenyon cell synapses are modu-
lated during memory formation even in the absence of antennal lobes, the ellipsoid body, and multiple unde-fined neuronal tracts (data not shown; Zars et al., 2000a).synaptic output from these cells (Dubnau et al., 2001;
McGuire et al., 2001). Quantification of the number of Kenyon cells in the twolines using the nuclear reporter UAS-nls-lacZ indicatesHere we show that extinction of olfactory memory is
an intracellular process rather than a re-learning phe- that GAL4 line 247 labels 711 � 17 cells/hemisphere inmales (mean � SD, n � 6) and 825 � 22 (n � 8) in femalesnomenon involving new memories in other neurons. This
“memory trace suppression” likely happens via a modifi- (Mader, 2001). The GAL4 line c772 similarly labels 842 �28 in males (n � 6) and 871 � 47 (n � 6) Kenyon cellscation of the molecular machinery of memory formation.
First, we demonstrate that the 30 min and 3 hr memory in females (Figure 1). Since a single mushroom body ismade up of approximately 2500 Kenyon cells (Technauis still contained in the mushroom body Kenyon cells.
Second, we localize extinction to the same set of Kenyon and Heisenberg, 1982), the following experiments ma-nipulate about 1/4 to 1/3 of these cells.cells implicated in memory formation. We propose that
in the Kenyon cell synapses, arrival of the CS withoutthe US stimulates a process antagonistic to the cAMP/ Memory FormationPKA signaling cascade. To determine if the olfactory memory stays in the Kenyon
cells, MB-rescued rut mutant flies were trained andtested at different time points following training. WeResultspreviously used several GAL4 lines with MB expressionto rescue the olfactory short-term memory defect of rutFor mapping a memory trace, a careful analysis of GAL4
expression in the lines used to target a respective ef- mutant flies (Zars et al., 2000a). We concentrate hereon the line 247 as it has the most specific mushroomfector gene is necessary (Rein et al., 2002). Examination
of the GAL4 line 247 with UAS-TAU or UAS-lacZ-KI2 as body expression. Wild-type flies show a small (�25%)memory decay over the 3 hr tested (Figure 2). MB-res-reporters shows that neurons of all three MB subsys-
tems (�/�; ��/��; and �) are labeled (Zars et al., 2000a cued rut mutant flies and wild-type flies with overexpres-sion of the rut transgene in the MBs show a performanceand Figure 1). Relatively more signal, however, is found
in the �/� and � subsystems than in ��/��. There is little similar to wild-type flies at all time points tested. Thisis in contrast to the rut mutant and rut mutant flies withexpression detected in the adult brain outside the MBs
with a weak signal found in a subset of the central brain’s the GAL4 driver or UAS-rut� effector alone, which showa drastically lower performance at all time points com-glial cells (data not shown). Another GAL4 line used in
this study is c772. It again labels all MB subsystems pared to MB-rescued rut mutant and wild-type flies.
Olfactory Memory Trace in MB Kenyon Cells953
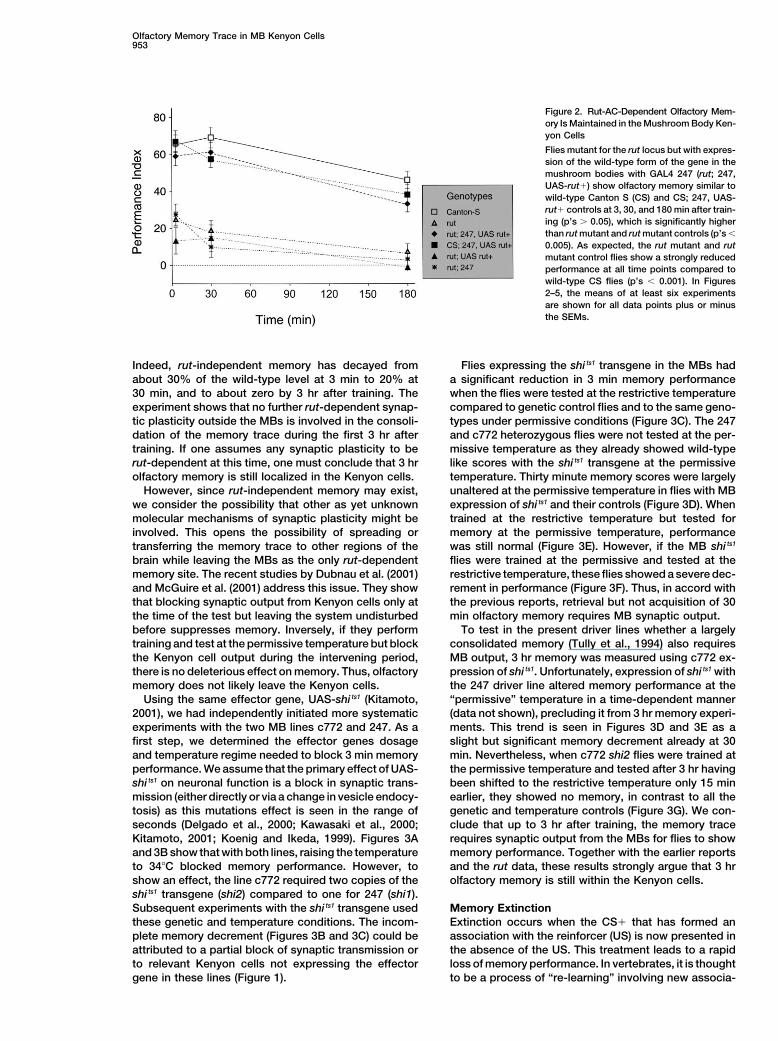
Figure 2. Rut-AC-Dependent Olfactory Mem-ory Is Maintained in the Mushroom Body Ken-yon Cells
Flies mutant for the rut locus but with expres-sion of the wild-type form of the gene in themushroom bodies with GAL4 247 (rut; 247,UAS-rut�) show olfactory memory similar towild-type Canton S (CS) and CS; 247, UAS-rut� controls at 3, 30, and 180 min after train-ing (p’s 0.05), which is significantly higherthan rut mutant and rut mutant controls (p’s
0.005). As expected, the rut mutant and rutmutant control flies show a strongly reducedperformance at all time points compared towild-type CS flies (p’s 0.001). In Figures2–5, the means of at least six experimentsare shown for all data points plus or minusthe SEMs.
Indeed, rut-independent memory has decayed from Flies expressing the shi ts1 transgene in the MBs hada significant reduction in 3 min memory performanceabout 30% of the wild-type level at 3 min to 20% at
30 min, and to about zero by 3 hr after training. The when the flies were tested at the restrictive temperaturecompared to genetic control flies and to the same geno-experiment shows that no further rut-dependent synap-
tic plasticity outside the MBs is involved in the consoli- types under permissive conditions (Figure 3C). The 247and c772 heterozygous flies were not tested at the per-dation of the memory trace during the first 3 hr after
training. If one assumes any synaptic plasticity to be missive temperature as they already showed wild-typelike scores with the shi ts1 transgene at the permissiverut-dependent at this time, one must conclude that 3 hr
olfactory memory is still localized in the Kenyon cells. temperature. Thirty minute memory scores were largelyunaltered at the permissive temperature in flies with MBHowever, since rut-independent memory may exist,
we consider the possibility that other as yet unknown expression of shi ts1 and their controls (Figure 3D). Whentrained at the restrictive temperature but tested formolecular mechanisms of synaptic plasticity might be
involved. This opens the possibility of spreading or memory at the permissive temperature, performancewas still normal (Figure 3E). However, if the MB shi ts1transferring the memory trace to other regions of the
brain while leaving the MBs as the only rut-dependent flies were trained at the permissive and tested at therestrictive temperature, these flies showed a severe dec-memory site. The recent studies by Dubnau et al. (2001)
and McGuire et al. (2001) address this issue. They show rement in performance (Figure 3F). Thus, in accord withthe previous reports, retrieval but not acquisition of 30that blocking synaptic output from Kenyon cells only at
the time of the test but leaving the system undisturbed min olfactory memory requires MB synaptic output.To test in the present driver lines whether a largelybefore suppresses memory. Inversely, if they perform
training and test at the permissive temperature but block consolidated memory (Tully et al., 1994) also requiresMB output, 3 hr memory was measured using c772 ex-the Kenyon cell output during the intervening period,
there is no deleterious effect on memory. Thus, olfactory pression of shi ts1. Unfortunately, expression of shi ts1 withthe 247 driver line altered memory performance at thememory does not likely leave the Kenyon cells.
Using the same effector gene, UAS-shi ts1 (Kitamoto, “permissive” temperature in a time-dependent manner(data not shown), precluding it from 3 hr memory experi-2001), we had independently initiated more systematic
experiments with the two MB lines c772 and 247. As a ments. This trend is seen in Figures 3D and 3E as aslight but significant memory decrement already at 30first step, we determined the effector genes dosage
and temperature regime needed to block 3 min memory min. Nevertheless, when c772 shi2 flies were trained atthe permissive temperature and tested after 3 hr havingperformance. We assume that the primary effect of UAS-
shi ts1 on neuronal function is a block in synaptic trans- been shifted to the restrictive temperature only 15 minearlier, they showed no memory, in contrast to all themission (either directly or via a change in vesicle endocy-
tosis) as this mutations effect is seen in the range of genetic and temperature controls (Figure 3G). We con-clude that up to 3 hr after training, the memory traceseconds (Delgado et al., 2000; Kawasaki et al., 2000;
Kitamoto, 2001; Koenig and Ikeda, 1999). Figures 3A requires synaptic output from the MBs for flies to showmemory performance. Together with the earlier reportsand 3B show that with both lines, raising the temperature
to 34�C blocked memory performance. However, to and the rut data, these results strongly argue that 3 hrolfactory memory is still within the Kenyon cells.show an effect, the line c772 required two copies of the
shi ts1 transgene (shi2) compared to one for 247 (shi1).Subsequent experiments with the shi ts1 transgene used Memory Extinction
Extinction occurs when the CS� that has formed anthese genetic and temperature conditions. The incom-plete memory decrement (Figures 3B and 3C) could be association with the reinforcer (US) is now presented in
the absence of the US. This treatment leads to a rapidattributed to a partial block of synaptic transmission orto relevant Kenyon cells not expressing the effector loss of memory performance. In vertebrates, it is thought
to be a process of “re-learning” involving new associa-gene in these lines (Figure 1).
Neuron954
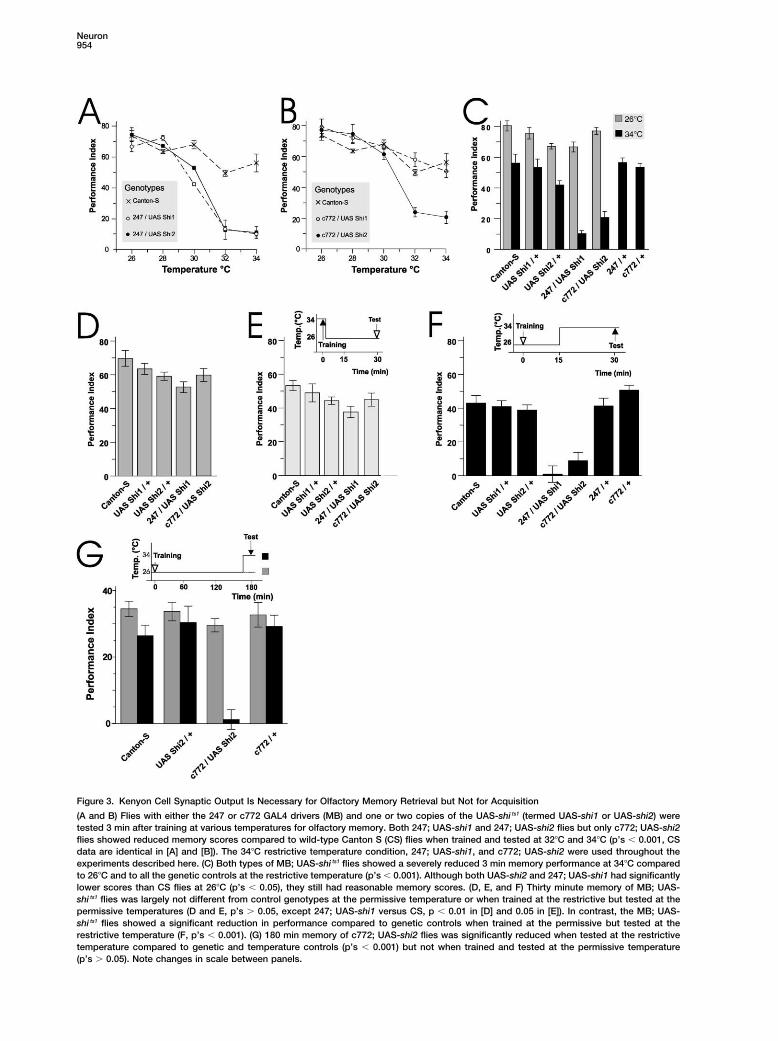
Figure 3. Kenyon Cell Synaptic Output Is Necessary for Olfactory Memory Retrieval but Not for Acquisition
(A and B) Flies with either the 247 or c772 GAL4 drivers (MB) and one or two copies of the UAS-shi ts1 (termed UAS-shi1 or UAS-shi2) weretested 3 min after training at various temperatures for olfactory memory. Both 247; UAS-shi1 and 247; UAS-shi2 flies but only c772; UAS-shi2flies showed reduced memory scores compared to wild-type Canton S (CS) flies when trained and tested at 32�C and 34�C (p’s 0.001, CSdata are identical in [A] and [B]). The 34�C restrictive temperature condition, 247; UAS-shi1, and c772; UAS-shi2 were used throughout theexperiments described here. (C) Both types of MB; UAS-shi ts1 flies showed a severely reduced 3 min memory performance at 34�C comparedto 26�C and to all the genetic controls at the restrictive temperature (p’s 0.001). Although both UAS-shi2 and 247; UAS-shi1 had significantlylower scores than CS flies at 26�C (p’s 0.05), they still had reasonable memory scores. (D, E, and F) Thirty minute memory of MB; UAS-shi ts1 flies was largely not different from control genotypes at the permissive temperature or when trained at the restrictive but tested at thepermissive temperatures (D and E, p’s 0.05, except 247; UAS-shi1 versus CS, p 0.01 in [D] and 0.05 in [E]). In contrast, the MB; UAS-shi ts1 flies showed a significant reduction in performance compared to genetic controls when trained at the permissive but tested at therestrictive temperature (F, p’s 0.001). (G) 180 min memory of c772; UAS-shi2 flies was significantly reduced when tested at the restrictivetemperature compared to genetic and temperature controls (p’s 0.001) but not when trained and tested at the permissive temperature(p’s 0.05). Note changes in scale between panels.
Olfactory Memory Trace in MB Kenyon Cells955
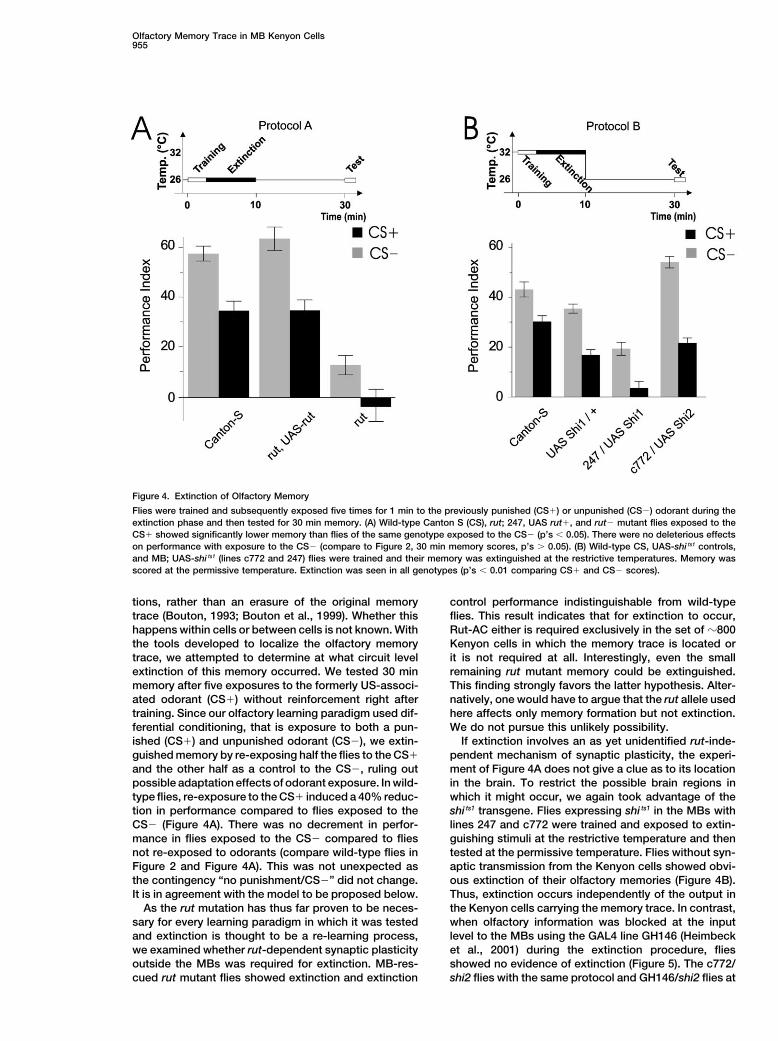
Figure 4. Extinction of Olfactory Memory
Flies were trained and subsequently exposed five times for 1 min to the previously punished (CS�) or unpunished (CS�) odorant during theextinction phase and then tested for 30 min memory. (A) Wild-type Canton S (CS), rut; 247, UAS rut�, and rut� mutant flies exposed to theCS� showed significantly lower memory than flies of the same genotype exposed to the CS� (p’s 0.05). There were no deleterious effectson performance with exposure to the CS� (compare to Figure 2, 30 min memory scores, p’s 0.05). (B) Wild-type CS, UAS-shi ts1 controls,and MB; UAS-shi ts1 (lines c772 and 247) flies were trained and their memory was extinguished at the restrictive temperatures. Memory wasscored at the permissive temperature. Extinction was seen in all genotypes (p’s 0.01 comparing CS� and CS� scores).
tions, rather than an erasure of the original memory control performance indistinguishable from wild-typeflies. This result indicates that for extinction to occur,trace (Bouton, 1993; Bouton et al., 1999). Whether this
happens within cells or between cells is not known. With Rut-AC either is required exclusively in the set of �800Kenyon cells in which the memory trace is located orthe tools developed to localize the olfactory memory
trace, we attempted to determine at what circuit level it is not required at all. Interestingly, even the smallremaining rut mutant memory could be extinguished.extinction of this memory occurred. We tested 30 min
memory after five exposures to the formerly US-associ- This finding strongly favors the latter hypothesis. Alter-natively, one would have to argue that the rut allele usedated odorant (CS�) without reinforcement right after
training. Since our olfactory learning paradigm used dif- here affects only memory formation but not extinction.We do not pursue this unlikely possibility.ferential conditioning, that is exposure to both a pun-
ished (CS�) and unpunished odorant (CS�), we extin- If extinction involves an as yet unidentified rut-inde-pendent mechanism of synaptic plasticity, the experi-guished memory by re-exposing half the flies to the CS�
and the other half as a control to the CS�, ruling out ment of Figure 4A does not give a clue as to its locationin the brain. To restrict the possible brain regions inpossible adaptation effects of odorant exposure. In wild-
type flies, re-exposure to the CS� induced a 40% reduc- which it might occur, we again took advantage of theshi ts1 transgene. Flies expressing shi ts1 in the MBs withtion in performance compared to flies exposed to the
CS� (Figure 4A). There was no decrement in perfor- lines 247 and c772 were trained and exposed to extin-guishing stimuli at the restrictive temperature and thenmance in flies exposed to the CS� compared to flies
not re-exposed to odorants (compare wild-type flies in tested at the permissive temperature. Flies without syn-aptic transmission from the Kenyon cells showed obvi-Figure 2 and Figure 4A). This was not unexpected as
the contingency “no punishment/CS�” did not change. ous extinction of their olfactory memories (Figure 4B).Thus, extinction occurs independently of the output inIt is in agreement with the model to be proposed below.
As the rut mutation has thus far proven to be neces- the Kenyon cells carrying the memory trace. In contrast,when olfactory information was blocked at the inputsary for every learning paradigm in which it was tested
and extinction is thought to be a re-learning process, level to the MBs using the GAL4 line GH146 (Heimbecket al., 2001) during the extinction procedure, flieswe examined whether rut-dependent synaptic plasticity
outside the MBs was required for extinction. MB-res- showed no evidence of extinction (Figure 5). The c772/shi2 flies with the same protocol and GH146/shi2 flies atcued rut mutant flies showed extinction and extinction
Neuron956
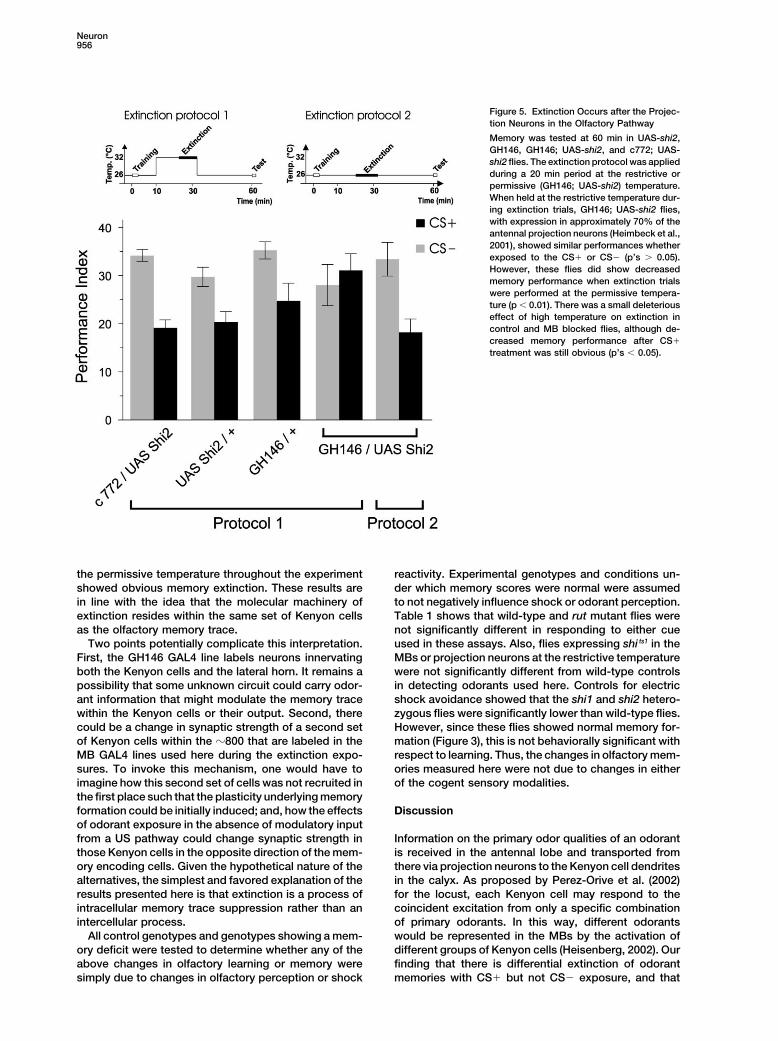
Figure 5. Extinction Occurs after the Projec-tion Neurons in the Olfactory Pathway
Memory was tested at 60 min in UAS-shi2,GH146, GH146; UAS-shi2, and c772; UAS-shi2 flies. The extinction protocol was appliedduring a 20 min period at the restrictive orpermissive (GH146; UAS-shi2) temperature.When held at the restrictive temperature dur-ing extinction trials, GH146; UAS-shi2 flies,with expression in approximately 70% of theantennal projection neurons (Heimbeck et al.,2001), showed similar performances whetherexposed to the CS� or CS� (p’s 0.05).However, these flies did show decreasedmemory performance when extinction trialswere performed at the permissive tempera-ture (p 0.01). There was a small deleteriouseffect of high temperature on extinction incontrol and MB blocked flies, although de-creased memory performance after CS�
treatment was still obvious (p’s 0.05).
the permissive temperature throughout the experiment reactivity. Experimental genotypes and conditions un-der which memory scores were normal were assumedshowed obvious memory extinction. These results are
in line with the idea that the molecular machinery of to not negatively influence shock or odorant perception.Table 1 shows that wild-type and rut mutant flies wereextinction resides within the same set of Kenyon cells
as the olfactory memory trace. not significantly different in responding to either cueused in these assays. Also, flies expressing shi ts1 in theTwo points potentially complicate this interpretation.
First, the GH146 GAL4 line labels neurons innervating MBs or projection neurons at the restrictive temperaturewere not significantly different from wild-type controlsboth the Kenyon cells and the lateral horn. It remains a
possibility that some unknown circuit could carry odor- in detecting odorants used here. Controls for electricshock avoidance showed that the shi1 and shi2 hetero-ant information that might modulate the memory trace
within the Kenyon cells or their output. Second, there zygous flies were significantly lower than wild-type flies.However, since these flies showed normal memory for-could be a change in synaptic strength of a second set
of Kenyon cells within the �800 that are labeled in the mation (Figure 3), this is not behaviorally significant withrespect to learning. Thus, the changes in olfactory mem-MB GAL4 lines used here during the extinction expo-
sures. To invoke this mechanism, one would have to ories measured here were not due to changes in eitherof the cogent sensory modalities.imagine how this second set of cells was not recruited in
the first place such that the plasticity underlying memoryformation could be initially induced; and, how the effects Discussionof odorant exposure in the absence of modulatory inputfrom a US pathway could change synaptic strength in Information on the primary odor qualities of an odorant
is received in the antennal lobe and transported fromthose Kenyon cells in the opposite direction of the mem-ory encoding cells. Given the hypothetical nature of the there via projection neurons to the Kenyon cell dendrites
in the calyx. As proposed by Perez-Orive et al. (2002)alternatives, the simplest and favored explanation of theresults presented here is that extinction is a process of for the locust, each Kenyon cell may respond to the
coincident excitation from only a specific combinationintracellular memory trace suppression rather than anintercellular process. of primary odorants. In this way, different odorants
would be represented in the MBs by the activation ofAll control genotypes and genotypes showing a mem-ory deficit were tested to determine whether any of the different groups of Kenyon cells (Heisenberg, 2002). Our
finding that there is differential extinction of odorantabove changes in olfactory learning or memory weresimply due to changes in olfactory perception or shock memories with CS� but not CS� exposure, and that
Olfactory Memory Trace in MB Kenyon Cells957
Table 1. Sensory Acuity Tests
Shock OdorantGenotype Avoidance Index Avoidance Index
Canton S 87.3 � 4.2 87.3 � 3.8rut2080 74.9 � 7.1 83.3 � 3.8rut2080; UAS-rut� 78.6 � 6.0 94.9 � 1.8rut2080; 247 71.5 � 6.1 89.7 � 3.1Canton S* 67.0 � 6.5 93.0 � 2.2UAS-shi1/�* 51.7 � 5.0 90.3 � 1.3UAS-shi2/�* 50.0 � 3.0 91.5 � 3.0247/�; UAS-shi1/�* 53.1 � 5.2 95.6 � 1.2c772/�; UAS-shi2/�* 58.1 � 5.2 91.6 � 2.2GH146/�; UAS-shi2/�* NA 89.3 � 1.3
Neither the rescue of the rutabaga olfactory learning phenotype northe blocking of Kenyon cell or projection neuron synaptic outputcauses behaviorally significant changes in electroshock or olfactorysensitivity. Wild-type Canton S (CS), rutabaga (rut) mutant, and flieswith blocked Kenyon cell or projection neuron synaptic transmission(at the restrictive temperature*) were tested for responses to boththe electroshock (except GH146) and the odorants used in the learn-ing experiments. There were no significant differences betweenwild-type CS flies and rut mutant flies in either assay (p’s 0.05).The flies with blocked Kenyon cell or projection neuron synaptictransmission were not different from wild-type CS flies at the ele-vated temperature in odorant avoidance. There was a significantdifference between CS flies and UAS-shi1 and UAS-shi2 flies inelectric shock avoidance (p’s 0.05) but comparing these flies,which learn normally (Fig 2), to the rest of the memory-impairedgenotypes’ electric shock avoidance scores at the elevated temper-ature show no significant differences. Thus, these changes are notsignificant for the olfactory learning assay. Means of 6–8 experi-ments per genotype are shown plus or minus SEMs.
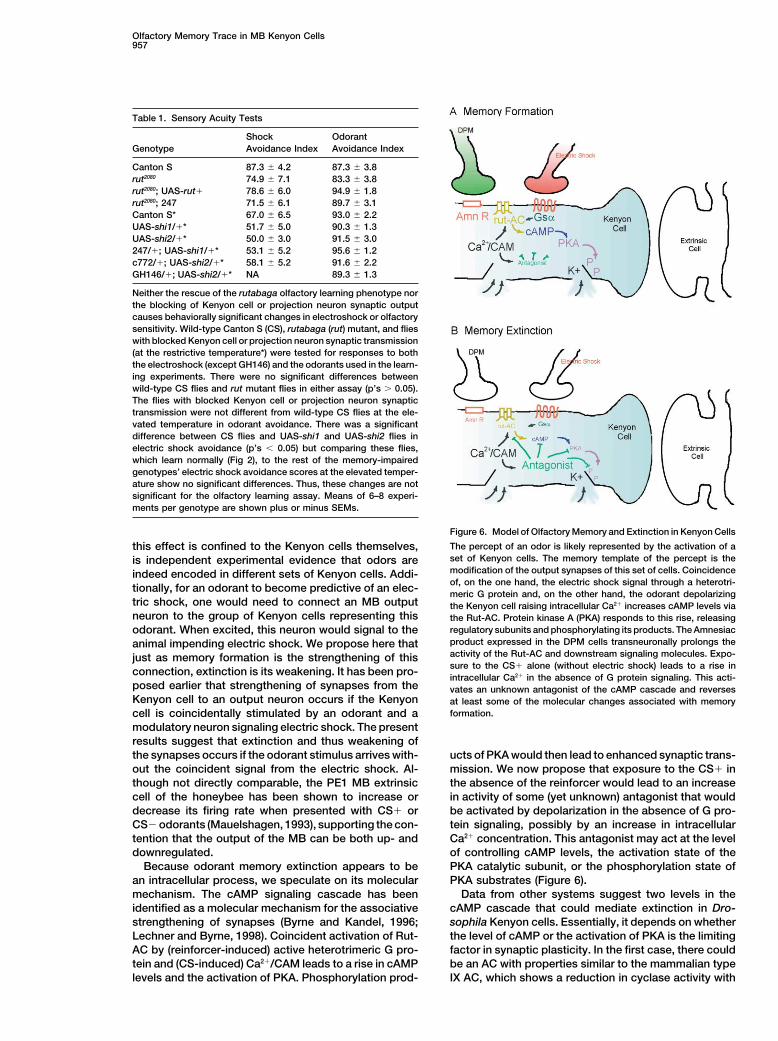
Figure 6. Model of Olfactory Memory and Extinction in Kenyon Cells
The percept of an odor is likely represented by the activation of athis effect is confined to the Kenyon cells themselves,set of Kenyon cells. The memory template of the percept is theis independent experimental evidence that odors aremodification of the output synapses of this set of cells. Coincidenceindeed encoded in different sets of Kenyon cells. Addi-of, on the one hand, the electric shock signal through a heterotri-tionally, for an odorant to become predictive of an elec- meric G protein and, on the other hand, the odorant depolarizing
tric shock, one would need to connect an MB output the Kenyon cell raising intracellular Ca2� increases cAMP levels vianeuron to the group of Kenyon cells representing this the Rut-AC. Protein kinase A (PKA) responds to this rise, releasing
regulatory subunits and phosphorylating its products. The Amnesiacodorant. When excited, this neuron would signal to theproduct expressed in the DPM cells transneuronally prolongs theanimal impending electric shock. We propose here thatactivity of the Rut-AC and downstream signaling molecules. Expo-just as memory formation is the strengthening of thissure to the CS� alone (without electric shock) leads to a rise inconnection, extinction is its weakening. It has been pro- intracellular Ca2� in the absence of G protein signaling. This acti-
posed earlier that strengthening of synapses from the vates an unknown antagonist of the cAMP cascade and reversesKenyon cell to an output neuron occurs if the Kenyon at least some of the molecular changes associated with memory
formation.cell is coincidentally stimulated by an odorant and amodulatory neuron signaling electric shock. The presentresults suggest that extinction and thus weakening ofthe synapses occurs if the odorant stimulus arrives with- ucts of PKA would then lead to enhanced synaptic trans-
mission. We now propose that exposure to the CS� inout the coincident signal from the electric shock. Al-though not directly comparable, the PE1 MB extrinsic the absence of the reinforcer would lead to an increase
in activity of some (yet unknown) antagonist that wouldcell of the honeybee has been shown to increase ordecrease its firing rate when presented with CS� or be activated by depolarization in the absence of G pro-
tein signaling, possibly by an increase in intracellularCS� odorants (Mauelshagen, 1993), supporting the con-tention that the output of the MB can be both up- and Ca2� concentration. This antagonist may act at the level
of controlling cAMP levels, the activation state of thedownregulated.Because odorant memory extinction appears to be PKA catalytic subunit, or the phosphorylation state of
PKA substrates (Figure 6).an intracellular process, we speculate on its molecularmechanism. The cAMP signaling cascade has been Data from other systems suggest two levels in the
cAMP cascade that could mediate extinction in Dro-identified as a molecular mechanism for the associativestrengthening of synapses (Byrne and Kandel, 1996; sophila Kenyon cells. Essentially, it depends on whether
the level of cAMP or the activation of PKA is the limitingLechner and Byrne, 1998). Coincident activation of Rut-AC by (reinforcer-induced) active heterotrimeric G pro- factor in synaptic plasticity. In the first case, there could
be an AC with properties similar to the mammalian typetein and (CS-induced) Ca2�/CAM leads to a rise in cAMPlevels and the activation of PKA. Phosphorylation prod- IX AC, which shows a reduction in cyclase activity with

Neuron958
Ca2�/calcineurin signaling and is important for learning tinction trials, implying that modifications in other neu-and memory in mice (Antoni et al., 1998; Paterson et al., rons compensated for this lack of reversal (Repa et al.,2000). If this were the case in the fly, then repeated 2001). Second, pharmacological alteration of the inferiordepolarization of the Kenyon cells would decrease the olive during extinction training of the eyelid response inlevels of cAMP in the cells. There is, indeed, a gene with the rabbit has revealed the importance of proper regula-high homology to a type IX AC in Drosophila; its role in tion of the US pathway for behavioral extinction to occur.learning and memory, however, has not yet been tested Decreasing or increasing activity of the climbing fibers(Iourgenko et al., 1997). Downstream in the cascade, a either blocked or induced extinction, respectively (Me-cAMP phosphodiesterase, responsive to Ca2� signaling, dina et al., 2002). Whether there is normally an inhibitorycould be activated by Kenyon cell depolarization. An signal connecting the tone (CS) pathway to the US path-alternative level of regulation could be on PKA itself way during tone alone presentation (mimicking a blockor its phosphorylated products. A model of synaptic of the excitatory input) is open. If this were true, thenplasticity in the Aplysia long-term sensitization system this would be an example of an intercellular mechanismincludes an activation of PKA by release and eventual of extinction. It will certainly be interesting to examinedegradation of its regulatory subunits that could lead to what the differences are that determine inter- versusa kinase that is independent of cAMP signaling (Bergold intracellular mechanisms of extinction.et al., 1992; Bernier et al., 1982; Chain et al., 1999; Green-berg et al., 1987). If this model is applicable to the Dro- Conclusionssophila Kenyon cells for olfactory memory, long-term We have shown here that olfactory memory can beactivation of PKA and continued phosphorylation of its formed and stored for up to 3 hr in about 1/3 of theproducts could be a mechanism of memory storage. Drosophila Kenyon cells (or a subset of these). ThisThe odorant-induced extinction would then antagonize memory can be acquired in the absence of Kenyon cellthis process by inducing the re-binding of the PKA- synaptic output and can still be abolished by blockingregulatory subunits to the catalytic subunits, or an enzyme the Kenyon cell output 3 hr later. This implies that thelike Calcineurin, a Ca2�/CAM-dependent phosphatase molecular mechanisms underlying 3 hr olfactory mem-could reverse the effects of PKA activity (Malleret et ory are located within Kenyon cells. Extinction mostal., 2001; Zeng et al., 2001). The molecular process of likely occurs by an intracellular suppression of the mem-extinction will probably involve a number of steps that ory trace, not by an intercellular mechanism. At the leveldecrease the forward signaling through cAMP and in- of the Kenyon cells, extinction is triggered by the CS�crease the reverse reactions. If the molecular memory if no signal arrives at the same time via the reinforcementinduction step is different from the extinction antagonist, pathway. At the molecular level, a depolarization-depen-then different time constants of enzymatic activity could dent increase of intracellular Ca2� may activate an an-account for spontaneous recovery of extinguished tagonist of cAMP or PKA signaling.memory performance, a commonly observed phenome-non in memory extinction. Precise determination of the Experimental Proceduresmolecules mediating the extinction process within the
Fly CareKenyon cells awaits further experimentation.Flies were raised at 25�C and 60% humidity with a 12:12 hr light:darkEarlier studies of extinction indicate that a partial shar-cycle on our standard cornmeal-based food (Guo et al., 1996). Theing of signal transduction pathways in memory acquisi-UAS-rut� #1 transgene (Zars et al., 2000b) was recombined onto
tion and extinction is rather common. That extinction the same chromosome as the 247 GAL4 enhancer (Zars et al., 2000a)of a memory uses a subset of the signaling pathways and put in wild-type Canton S and rut 2080 mutant background byneeded for its formation has been seen in fear condition- standard crosses. The rut 2080; UAS rut� and rut 2080; 247 lines were
similarly handled. Flies containing the UAS-shi ts1 transgene (Kita-ing in the mouse and rat, and conditioned taste avoid-moto, 2001) and a GAL4 driver (c772, GH146, and 247) (Tettamantiance in the rat. Lattal and Abel (2001) showed that inet al., 1997; Zars et al., 2000a) were F1 progeny of homozygousthe mouse, protein synthesis is needed for acquisitionparental lines.of spatial learning and fear conditioning, but protein
synthesis is not needed for extinction of that process. ImmunohistologyOn the other hand, for fear conditioning in the rat, the We examined 5-day-old adult flies containing both the GAL4 elementNMDA receptor in the amygdala is involved in both ac- and a reporter (UAS:lacZ-K12 for whole-cell expression or UAS:nls-
lacZ for nuclear expression). Whole-mount brains were dissectedquisition and extinction (Lee and Kim, 1998). A similarand fixed as described in Rein et al. (2002). Brains were double-partial use of signaling pathways in acquisition and ex-stained with nc82 to label the synaptic neuropil and anti-�-gal. Alltinction is seen in conditioned taste avoidance memoryantibody stainings were labeled using secondary antibodies cou-in rats (Berman and Dudai, 2001). In contrast to fearpled to Cy3.18 and Alexa488[0]. Further handling and confocal scan-
conditioning, however, protein synthesis was required ning methods are as in Rein et al. (2002) except that specimensin the insular cortex for both acquisition and extinction. with nuclear expression were scanned from posterior at an axialWhile these studies have addressed the molecular pro- resolution of about 1.25 �m. Nuclear expression patterns were addi-
tionally analyzed using a special AMIRA-module for manual Kenyoncess of extinction, they did not address the underlyingcell counting (Mader, 2001).circuit changes accompanying these phenomena.
Two studies have explored the intra/intercellular prob-Behavioral Experimentslem. First is an intriguing observation from electrophysi-Flies were trained to avoid Benzaldehyde or 3-Octanol with 5 mm
ological recording in the amygdala during fear condition- or 16 mm diameter cups, respectively, essentially as described (Tullying (Repa et al., 2001). The authors found some neurons and Quinn, 1985). Modifications to the teaching machine were car-that showed evidence of plasticity during acquisition, ried out to allow for four simultaneous experiments, shock grids
were also slightly altered (plans available on request). All rutabagaand these plastic changes were maintained through ex-

Olfactory Memory Trace in MB Kenyon Cells959
rescue and shi ts1 permissive experiments were performed at 26�C R.L. (1998). Tripartite mushroom body architecture revealed by anti-genic markers. Learn. Mem. 5, 38–51.and 85% relative humidity. The shi ts1 restrictive experiments were
carried out at 34�C and 85% relative humidity. Extinction consisted de Belle, J.S., and Heisenberg, M. (1994). Associative odor learningof five, 1 min exposures to the odorant previously associated with in Drosophila abolished by chemical ablation of MBs. Science 263,punishment (CS�), with a 1 min interstimulus interval. The “extinc- 692–695.tion control” experiment was the same except that the odorant
Delgado, R., Maureira, C., Oliva, C., Kidokoro, Y., and Labarca, P.not previously associated with punishment (CS�) was presented.(2000). Size of vesicle pools, rates of mobilization, and recycling atMemory was scored 30 or 60 min after the initial training sessionthe neuromuscular synapses of a Drosophila mutant, shibire. Neuronwas finished. Shock reactivity and olfactory acuity experiments used28, 941–953.stimuli exactly as in the training protocols. Statistically significantDrain, P., Folkers, E., and Quinn, W.G. (1991). cAMP-dependentdifferences were determined using ANOVAs and Duncan post hocprotein kinase and the disruption of learning in transgenic flies.tests when appropriate.Neuron 6, 71–82.
Acknowledgments Dubnau, J., Grady, L., Kitamoto, T., and Tully, T. (2001). Disruptionof neurotransmission in Drosophila mushroom body blocks retrieval
We thank B. Gerber for thoughtful insights on different aspects of but not acquisition of memory. Nature 411, 476–480.the project, M. Fischer for initial memory measurements, and S.
Feany, M.B., and Quinn, W.G. (1995). A neuropeptide gene definedKirschner, M. Stratmann, S. Nanu, C. Grubel, and B. Muhlbauer for
by the Drosophila memory mutant amnesiac. Science 268, 869–873.excellent technical assistance. We are grateful to H. Kadeschabek
Greenberg, S.M., Castellucci, V.F., Bayley, H., and Schwartz, J.H.and K. Oechsner for building the modified olfactory teaching ma-(1987). A molecular mechanism for long-term sensitization inchine. T. Kitamoto kindly provided the UAS-shi ts1 fly lines. This workAplysia. Nature 329, 62–65.was supported by SFB 554 from the DFG, Fonds der Chemischen
Industrie, BMBF (0311555), and Human Frontiers Science Program Grotewiel, M.S., Beck, C.D., Wu, K.H., Zhu, X.R., and Davis, R.L.(RG0143) to M.H., as well as a graduate student fellowship from the (1998). Integrin-mediated short-term memory in Drosophila. NatureGraduiertenkolleg “Arthropodenverhalten” to M.S. 391, 455–460.
Guo, A., Liu, L., Xia, S.-Z., Feng, C.-H., Wolf, R., and Heisenberg,Received: April 15, 2002 M. (1996). Conditioned visual flight orientation in Drosophila: depen-Revised: June 14, 2002 dence on age, practice, and diet. Learn. Mem. 3, 49–59.
Han, P.L., Levin, L.R., Reed, R.R., and Davis, R.L. (1992). PreferentialReferencesexpression of the Drosophila rutabaga gene in mushroom bodies,neural centers for learning in insects. Neuron 9, 619–627.Antoni, F.A., Palkovits, M., Simpson, J., Smith, S.M., Leitch, A.L.,Heimbeck, G., Bugnon, V., Gendre, N., Keller, A., and Stocker, R.F.Rosie, R., Fink, G., and Paterson, J.M. (1998). Ca2�/calcineurin-inhib-(2001). A central neural circuit for experience-independent olfactoryited adenylyl cyclase, highly abundant in forebrain regions, is impor-and courtship behavior in Drosophila melanogaster. Proc. Natl.tant for learning and memory. J. Neurosci. 18, 9650–9661.Acad. Sci. USA 98, 15336–15341.Bergold, P.J., Beushausen, S.A., Sacktor, T.C., Cheley, S., Bayley,Heisenberg, M. (1998). What do the mushroom bodies do for theH., and Schwartz, J.H. (1992). A regulatory subunit of the cAMP-insect brain? An introduction. Learn. Mem. 5, 1–10.dependent protein kinase down-regulated in Aplysia sensory neu-
rons during long-term sensitization. Neuron 8, 387–397. Heisenberg, M. (2002). Die Pilzkorper der Insekten—das TrojanischeBerman, D.E., and Dudai, Y. (2001). Memory extinction, learning Pferd fur die Hirnforschung? Neuroforum 8, 179–186.anew, and learning the new: dissociations in the molecular machin- Heisenberg, M., Borst, A., Wagner, S., and Byers, D. (1985). Dro-ery of learning in cortex. Science 291, 2417–2419. sophila mushroom body mutants are deficient in olfactory learning.Bernier, L., Castellucci, V.F., Kandel, E.R., and Schwartz, J.H. (1982). J. Neurogenet. 2, 1–30.Facilitatory transmitter causes a selective and prolonged increase Iourgenko, V., Kliot, B., Cann, M.J., and Levin, L.R. (1997). Cloningin adenosine 3�:5�-monophosphate in sensory neurons mediating and characterization of a Drosophila adenylyl cyclase homologousthe gill and siphon withdrawal reflex in Aplysia. J. Neurosci. 2, 1682– to mammalian type IX. FEBS Lett. 413, 104–108.1691.
Ito, K., Suzuki, K., Estes, P., Ramaswami, M., Yamamoto, D., andBouton, M.E. (1993). Context, time, and memory retrieval in the
Strausfeld, N.J. (1998). The organization of extrinsic neurons andinterference paradigms of Pavlovian learning. Psychol. Bull. 114,
their implications in the functional roles of the mushroom bodies in80–99.
Drosophila melanogaster Meigen. Learn. Mem. 5, 52–77.Bouton, M.E., Nelson, J.B., and Rosas, J.M. (1999). Stimulus gener-
Kawasaki, F., Hazen, M., and Ordway, R.W. (2000). Fast synapticalization, context change, and forgetting. Psychol. Bull. 125,
fatigue in shibire mutants reveals a rapid requirement for dynamin171–186.
in synaptic vesicle membrane trafficking. Nat. Neurosci. 3, 859–860.Byrne, J.H., and Kandel, E.R. (1996). Presynaptic facilitation revis-
Kitamoto, T. (2001). Conditional modification of behavior in Dro-ited: state and time dependence. J. Neurosci. 16, 425–435.sophila by targeted expression of a temperature-sensitive shibire
Chain, D.G., Casadio, A., Schacher, S., Hegde, A.N., Valbrun, M., allele in defined neurons. J. Neurobiol. 47, 81–92.Yamamoto, N., Goldberg, A.L., Bartsch, D., Kandel, E.R., and
Koenig, J.H., and Ikeda, K. (1999). Contribution of active zone sub-Schwartz, J.H. (1999). Mechanisms for generating the autonomouspopulation of vesicles to evoked and spontaneous release. J. Neuro-cAMP-dependent protein kinase required for long-term facilitationphysiol. 81, 1495–1505.in Aplysia. Neuron 22, 147–156.Lattal, K.M., and Abel, T. (2001). Different requirements for proteinCheng, Y., Endo, K., Wu, K., Rodan, A.R., Heberlein, U., and Davis,synthesis in acquisition and extinction of spatial preferences andR.L. (2001). Drosophila fasciclinII is required for the formation of odorcontext-evoked fear. J. Neurosci. 21, 5773–5780.memories and for normal sensitivity to alcohol. Cell 105, 757–768.Lechner, H.A., and Byrne, J.H. (1998). New perspectives on classicalConnolly, J.B., Roberts, I.J., Armstrong, J.D., Kaiser, K., Forte, M.,conditioning: a synthesis of Hebbian and non-Hebbian mechanisms.Tully, T., and O’Kane, C.J. (1996). Associative learning disrupted byNeuron 20, 355–358.impaired Gs signaling in Drosophila mushroom bodies. Science 274,
2104–2107. Lee, H., and Kim, J.J. (1998). Amygdalar NMDA receptors are criticalfor new fear learning in previously fear-conditioned rats. J. Neurosci.Cowan, T.M., and Siegel, R.W. (1986). Drosophila mutations that18, 8444–8454.alter ionic conduction disrupt acquisition and retention of a condi-
tioned odor avoidance response. J. Neurogenet. 3, 187–201. Levin, L.R., Han, P.L., Hwang, P.M., Feinstein, P.G., Davis, R.L.,and Reed, R.R. (1992). The Drosophila learning and memory geneCrittenden, J.R., Skoulakis, E.M., Han, K.A., Kalderon, D., and Davis,

Neuron960
rutabaga encodes a Ca2�/Calmodulin-responsive adenylyl cyclase. Zars, T. (2000). Behavioral functions of the insect mushroom bodies.Curr. Opin. Neurobiol. 10, 790–795.Cell 68, 479–489.
Zars, T., Fischer, M., Schulz, R., and Heisenberg, M. (2000a). Local-Mader, M. (2001). Analysis of expression patterns in the Drosophilaization of a short-term memory in Drosophila. Science 288, 672–675.mushroom bodies. Masters thesis, Julius-Maximilians-Universitaet
Wuerzburg, Wuerzburg, Germany. Zars, T., Wolf, R., Davis, R., and Heisenberg, M. (2000b). Tissue-specific expression of a type I adenylyl cyclase rescues the rutabagaMalleret, G., Haditsch, U., Genoux, D., Jones, M.W., Bliss, T.V.,mutant memory defect: in search of the engram. Learn. Mem. 7,Vanhoose, A.M., Weitlauf, C., Kandel, E.R., Winder, D.G., and Man-18–31.suy, I.M. (2001). Inducible and reversible enhancement of learning,
memory, and long-term potentiation by genetic inhibition of cal- Zeng, H., Chattarji, S., Barbarosie, M., Rondi-Reig, L., Philpot, B.D.,cineurin. Cell 104, 675–686. Miyakawa, T., Bear, M.F., and Tonegawa, S. (2001). Forebrain-spe-
cific calcineurin knockout selectively impairs bidirectional synapticMauelshagen, J. (1993). Neural correlates of olfactory learning para-plasticity and working/episodic-like memory. Cell 107, 617–629.digms in an identified neuron in the honeybee brain. J. Neurophysiol.Zhong, Y. (1995). Mediation of PACAP-like neuropeptide transmis-69, 609–625.sion by co-activation of Ras/Raf and cAMP signal transduction path-McGuire, S.E., Le, P.T., and Davis, R.L. (2001). The role of Drosophilaways in Drosophila. Nature 375, 588–592.mushroom body signaling in olfactory memory. Science 293, 1330–
1333.
Medina, J.F., Nores, W.L., and Mauk, M.D. (2002). Inhibition of climb-ing fibres is a signal for the extinction of conditioned eyelid re-sponses. Nature 416, 330–333.
Nighorn, A., Healy, M.J., and Davis, R.L. (1991). The cyclic AMPphosphodiesterase encoded by the Drosophila dunce gene is con-centrated in the mushroom body neuropil. Neuron 6, 455–467.
Paterson, J.M., Smith, S.M., Simpson, J., Grace, O.C., Sosunov,A.A., Bell, J.E., and Antoni, F.A. (2000). Characterisation of humanadenylyl cyclase IX reveals inhibition by Ca(2�)/calcineurin and dif-ferential mRNA polyadenylation. J. Neurochem. 75, 1358–1367.
Perez-Orive, J., Mazor, O., Turner, G., Cassenaer, S., Wilson, R.I.,and Laurent, G. (2002). Oscillations and sparsening of odor repre-sentation in the mushroom body. Science 297, 359–365.
Qiu, Y.H., Chen, C.N., Malone, T., Richter, L., Beckendorf, S.K., andDavis, R.L. (1991). Characterization of the memory gene dunce ofDrosophila melanogaster. J. Mol. Biol. 222, 553–565.
Rein, K., Zockler, M., Mader, M.T., Grubel, C., and Heisenberg, M.(2002). The Drosophila standard brain. Curr. Biol. 12, 227–231.
Repa, J.C., Muller, J., Apergis, J., Desrochers, T.M., Zhou, Y., andLeDoux, J.E. (2001). Two different lateral amygdala cell populationscontribute to the initiation and storage of memory. Nat. Neurosci.4, 724–731.
Skoulakis, E.M., Kalderon, D., and Davis, R.L. (1993). Preferentialexpression in mushroom bodies of the catalytic subunit of proteinkinase A and its role in learning and memory. Neuron 11, 197–208.
Stocker, R.F. (1994). The organization of the chemosensory systemin Drosophila melanogaster: A review. Cell Tissue Res. 275, 3–26.
Strausfeld, N.J. (1976). Atlas of an Insect Brain (Berlin: Springer-Verlag).
Strausfeld, N.J., Bassemir, U., Singh, R.N., and Bacon, J.P. (1984).Organizational principles of outputs from dipteran brains. J. InsectPhysiol. 30, 73–93.
Strausfeld, N.J., Hansen, L., Li, Y., Gomez, R.S., and Ito, K. (1998).Evolution, discovery, and interpretations of arthropod mushroombodies. Learn. Mem. 5, 11–37.
Technau, G., and Heisenberg, M. (1982). Neural reorganization dur-ing metamorphosis of the corpora pedunculata in Drosophila mela-nogaster. Nature 295, 405–407.
Tettamanti, M., Armstrong, J.D., Endo, K., Yang, M.Y., Furukubo-Tokunaga, K., Kaiser, K., and Reichert, H. (1997). Early developmentof the Drosophila mushroom bodies, brain centers for associativelearning and memory. Dev. Genes Evol. 207, 242–252.
Tully, T., and Quinn, W.G. (1985). Classical conditioning and reten-tion in normal and mutant Drosophila melanogaster. J. Comp. Phys-iol. [A] 157, 263–277.
Tully, T., Preat, T., Boynton, S.C., and Del Vecchio, M. (1994). Geneticdissection of consolidated memory in Drosophila. Cell 79, 35–47.
Waddell, S.R.M., Armstrong, J.D., Kitamoto, T., Kaiser, K., andQuinn, W.G. (2000). The amnesiac gene product is expressed in twoneurons in the Drosophila brain that are critical for memory. Cell103, 805–813.
Related Documents











