Journal of Plant Physiology 162 (2005) 985—1002 Expression of transgenic stilbene synthases in wheat causes the accumulation of unknown stilbene derivatives with antifungal activity $ Liliya Serazetdinova ,1 , Klaus H. Oldach 2 ,HorstLo¨rz Institute of General Botany and Botanical Garden, AMP II, University of Hamburg, Ohnhorststrasse 18, D-22609 Hamburg, Germany Received 14 September 2004; accepted 18 November 2004 Summary The expression of foreign phytoalexins in a new host is thought to increase fungal resistance, since host-specific pathogens have not experienced selection for detoxifying or metabolising the novel antifungal compounds. Two resveratrol synthase genes vst1 and vst2 from grapevine (Vitis vinifera L.) and the pinosylvin synthase gene pss from pine (Pinus sylvestris L.) were stably transformed into bread wheat. The expression of the target genes is regulated by stress-inducible grapevine promoters. The vst1 and vst2 promoters were functional in wheat and retained their expression profiles described for grapevine. All vst and pss transgenic lines accumulated stilbene derivatives upon induction by UV light. The detected stilbenes showed a remarkable similarity to resveratrol and pinosylvin, however were found to be more hydrophilic than resveratrol and pinosylvin. Upon inoculation with the biotrophic pathogen Puccinia recondita f.sp. tritici several vst expressing wheat lines showed a significant reduction of disease symptoms (1979% to 2778%) compared to wild-type plants. The reduction of disease symptoms was even more obvious after inoculation with the facultative biotrophic pathogen Septoria nodorum Berk. and ranged from 42713% to 7174%. None of the four tested pss expressing lines showed a reduction in disease incidence. & 2005 Elsevier GmbH. All rights reserved. ARTICLE IN PRESS www.elsevier.de/jplph KEYWORDS Phytoalexins; Stilbenes; Transgenic wheat 0176-1617/$ - see front matter & 2005 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2004.11.005 $ This work was supported by the German Academic Exchange Service (post-graduate fellowship A/97/13719 to L.S.) and Norddeutsche Pflanzenzucht K&G (Hohenlieth, Germany). Corresponding author. E-mail address: [email protected] (L. Serazetdinova). 1 Present address: The Sainsbury Laboratory, John Innes Centre, Norwich Research Park, Norwich, NR4 7UH, UK. 2 Present address: Australian Centre for Plant Functional Genomics, University of Adelaide, Glen Osmond, Waite Campus, SA 5064, Australia.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
Journal of Plant Physiology 162 (2005) 985—1002
KEYWORDPhytoalexiStilbenes;Transgenic
0176-1617/$ - sdoi:10.1016/j.
$This workNorddeutsche P�CorrespondE-mail addr
1Present add2Present add
Australia.
www.elsevier.de/jplph
Expression of transgenic stilbene synthases inwheat causes the accumulation of unknownstilbene derivatives with antifungal activity$
Liliya Serazetdinova�,1, Klaus H. Oldach2, Horst Lorz
Institute of General Botany and Botanical Garden, AMP II, University of Hamburg, Ohnhorststrasse 18, D-22609Hamburg, Germany
Received 14 September 2004; accepted 18 November 2004
Sns;
wheat
ee front matter & 200jplph.2004.11.005
was supported by tflanzenzucht K&G (Hing author.ess: liliya.serazetdinoress: The Sainsbury Laress: Australian Centr
SummaryThe expression of foreign phytoalexins in a new host is thought to increase fungalresistance, since host-specific pathogens have not experienced selection fordetoxifying or metabolising the novel antifungal compounds. Two resveratrol synthasegenes vst1 and vst2 from grapevine (Vitis vinifera L.) and the pinosylvin synthase genepss from pine (Pinus sylvestris L.) were stably transformed into bread wheat. Theexpression of the target genes is regulated by stress-inducible grapevine promoters.The vst1 and vst2 promoters were functional in wheat and retained their expressionprofiles described for grapevine. All vst and pss transgenic lines accumulated stilbenederivatives upon induction by UV light. The detected stilbenes showed a remarkablesimilarity to resveratrol and pinosylvin, however were found to be more hydrophilicthan resveratrol and pinosylvin. Upon inoculation with the biotrophic pathogenPuccinia recondita f.sp. tritici several vst expressing wheat lines showed a significantreduction of disease symptoms (1979% to 2778%) compared to wild-type plants. Thereduction of disease symptoms was even more obvious after inoculation with thefacultative biotrophic pathogen Septoria nodorum Berk. and ranged from 42713% to7174%. None of the four tested pss expressing lines showed a reduction in diseaseincidence.& 2005 Elsevier GmbH. All rights reserved.
5 Elsevier GmbH. All rights reserved.
he German Academic Exchange Service (post-graduate fellowship A/97/13719 to L.S.) andohenlieth, Germany).
[email protected] (L. Serazetdinova).boratory, John Innes Centre, Norwich Research Park, Norwich, NR4 7UH, UK.e for Plant Functional Genomics, University of Adelaide, Glen Osmond, Waite Campus, SA 5064,

ARTICLE IN PRESS
L. Serazetdinova et al.986
Introduction
The synthesis of phytoalexins is one of the activedefence mechanisms in plants. Stilbenes representone of the many classes of phytoalexins. Thesephytoalexins are synthesised in a wide range ofunrelated plant species, including Vitaceae, Pina-ceae, Leguminosae and Polygonaceae (Gorham,1989) but are not present in important crops suchas wheat, barley or maize. The skeleton ofstilbenes is based on the trans-resveratrol structure(3,5,40-trihydroxystilbene) and the most commonstilbene is resveratrol (Fig. 1). Other monomericstilbenes found in Vitaceae are piceid (3-O-b-D-glucoside of resveratrol) and pterostilbene (3,5-dimethoxy-40-hydrostilbene) (Fig. 1). In Pinaceaestilbenes are represented by pinosylvin (30,50-dihydroxy-trans-stilbene, Fig. 1) and its mono-methylether.
Stilbenes have been the subject of numerousstudies due to their antifungal activity (Adrian etal., 1997). Several studies in grapevine haverevealed a positive correlation between the accu-mulation of stilbenes and disease resistance (Lang-cake and McCarthy, 1979; Dercks and Creasy, 1989).Resveratrol shows antifungal activity at concentra-tions ranging from 2.6 to 7� 10�4 M similar to theactivity range of other phytoalexins. Treatment ofconidia with sub-lethal or lethal concentrations ofstilbenes causes strong modification of fungalmembranes, resulting in structural abnormalitiesof fungal cells and cessation of respiration (Pontand Pezet, 1990; Adrian et al., 1997). Having a verysimilar chemical structure, resveratrol and pino-sylvin differ in the 40-hydroxyl group, which ispresent in resveratrol but absent in pinosylvin (Fig.1). It is known that a higher lipophily and higherelectrical charge of a phytoalexin correlates withits toxicity to fungi (Pont and Pezet, 1990). Beingmore lipophilic than resveratrol, pinosylvin mighthave higher fungistatical and fungitoxical activitythan resveratrol.
Both resveratrol and pinosylvin stilbenes areproducts of the phenylpropanoid pathway andsynthesised by stilbene synthases (STS). Based on
R3
R2
A
B
R1
trans-stilbene backbone
trans-stilbenes
Resveratrol-glucos
Resveratrol
Pinosylvin
Pterostilben
Figure 1. Chemical struct
substrate specificity, stilbene synthases aregrouped into two categories. Resveratrol synthases(RS, EC 2.3.1.95) in grapevine and peanut recruitthree molecules of p-coumaroyl-CoA and onemolecule of malonyl-CoA to synthesise resveratrol,while the pinosylvin synthase (PSS, EC 2.3.1.146) inpine uses cinnamoyl-CoA instead of p-coumaroyl-CoA for the synthesis of pinosylvin. All precursorsfor the biosynthesis of resveratrol and pinosylvinare common plant metabolites.
In grapevine, peanut and pine the stilbenesynthases are found as a cluster of genes that havea high homology in the coding region but differsignificantly in their promoter sequences. Theindividual members of the stilbene synthase genefamily show different expression profiles uponinduction and can be divided into two classes.Genes of the first class (vst1) are expressed earlyand strongly but with rapid degradation of themRNAs produced, whereas the genes of the secondclass including vst2 and pst3 are expressed laterand slowly activated, providing more stable mRNA.
Providing pine and grapevine stilbene synthasesrequire common plant metabolites (p-coumaroyl-CoA, cinnamoyl-CoA and malonyl-CoA) as theirprecursors, it was assumed that stilbene biosynth-esis depends only on the presence of a stilbenesynthase enzyme (Hain et al., 1990). A one-stepreaction in the biosynthesis of resveratrol andpinosylvin made them to attractive targets forgenetic engineering. The expression of foreignphytoalexins in a new host could result in increasedfungal resistance, since host-specific pathogens areunlikely to be able to detoxify or metabolise novelantifungal compounds. Indeed, expression of re-sveratrol-forming STS in dicotyledonous plantsincluding tobacco, tomato, and alfalfa resulted inaccumulation of either resveratrol or its glucosideand improved disease resistance (Hain et al., 1993;Thomzik et al., 1997; Hipskind and Paiva, 2000).Resveratrol synthase was also expressed in mono-cotyledonous plants including barley, rice, andwheat (Stark-Lorenzen et al., 1997; Leckband andLorz, 1998; Fettig and Hess, 1999). Transgenicbarley and rice have been shown to have improved
-R1 -R2 -R3
ide -OH - OC6H6O6 -OH
-OH - OH -OH
- OH -OH
-OH -OCH3 -OCH3
-H
ure of trans-stilbenes.

ARTICLE IN PRESS
Transgenic stilbene synthases in wheat 987
fungal resistance. So far heterologous expression ofpinosylvin-forming STS and its effects on diseaseresistance have not been reported. None of theprevious studies compare the expression of twodifferent classes of resveratrol synthase genes (vst1and vst2) in a heterologous system and their impacton the accumulation of stilbenes.
Here we report on stable transformation ofwheat with stilbene synthase genes originatingfrom grapevine (vst1, vst2) and pine (pss), theirexpression in transgenic plants, accumulation ofnovel stilbenes and improved disease resistanceagainst two wheat-specific pathogens, Pucciniarecondita f. sp. tritici and Septoria nodorum Berk.
Materials and methods
Plasmids and vector construction
Plasmids pVst1EPG, pVst1PG (Fig. 3A), and pGBI(Fig. 4) were described previously (Leckband andLorz, 1998). The vector pAct1DGUS (CAMBIA,Australia) contains the uidA reporter gene under
A
EcoRI
vst3
0
2.0 k
0.8
SspI
elicitor
H-box SA
ozone
ethyle
woun
-1100-1400-1550
B
-1700 - 1100-1400-2000
C
Figure 2. (A) Restriction map of the genomic clone pVst12t3coding regions of the stilbene synthase genes. Restriction sitepromoters are marked. (B, C) Schematic illustration of the poin 5 prime non-coding regions of the vst1 (B) and vst2 (C) ge
control of the Actin1 promoter from rice (McElroyet al., 1990) and the nos terminator from Agro-bacterium tumefaciens. The construct pAct1Dneo(Muller, 1994) contains the nptII-gene under controlof the Actin1 promoter from rice and the nosterminator. Vector pVst2 (not shown) containingthe genomic vst2 sequence was cloned by ligationof a 2.3 kb SspI fragment from plasmid pVst12t3 (R.Hain, Bayers, Fig. 2A) into the SmaI site of thevector pBlueskript sIIKS+. For the construction ofthe vector pVst2PG (Fig. 3A) the 0.8 kb 5 primeflanking region of the vst2 gene was amplified frompVst2 by PCR with M13 reverse primer and the vst2specific primer 50-GTAGGATCCCTGCTGAAATCG-30
with the introduced BamHI site), digested with SalIand BamHI and ligated with the SalI/BamHI frag-ment of pVst1EPG containing the uidA gene and nosterminator. Correct amplification and cloning wereverified by sequencing. The vector pVst2EPG (Fig.3A) was created from pVst2PG by the insertion ofthe HindIII fragment from pVst1EPG containing the4-fold enhancer element of the CaMV 35S promoter(Odel et al., 1985) in front of the vst2 promotersequence. The plasmid pEVst2 (Fig. 4) contains the2.3 kb long SspI fragment of the vst2 genomic
5000 10000
vst2 vst1
b 1.5 kb
EcoRI
kb
SspI PstI
ne
ding
ABA
light
MYB binding sites
- 800 + 100-200-500
-800 +100-200-500
(Hain et al., 1993; Wiese et al., 1994). Arrows indicates relevant to the cloning and functional analysis of the vstsition and distribution of the putative cis-acting elementsnes.
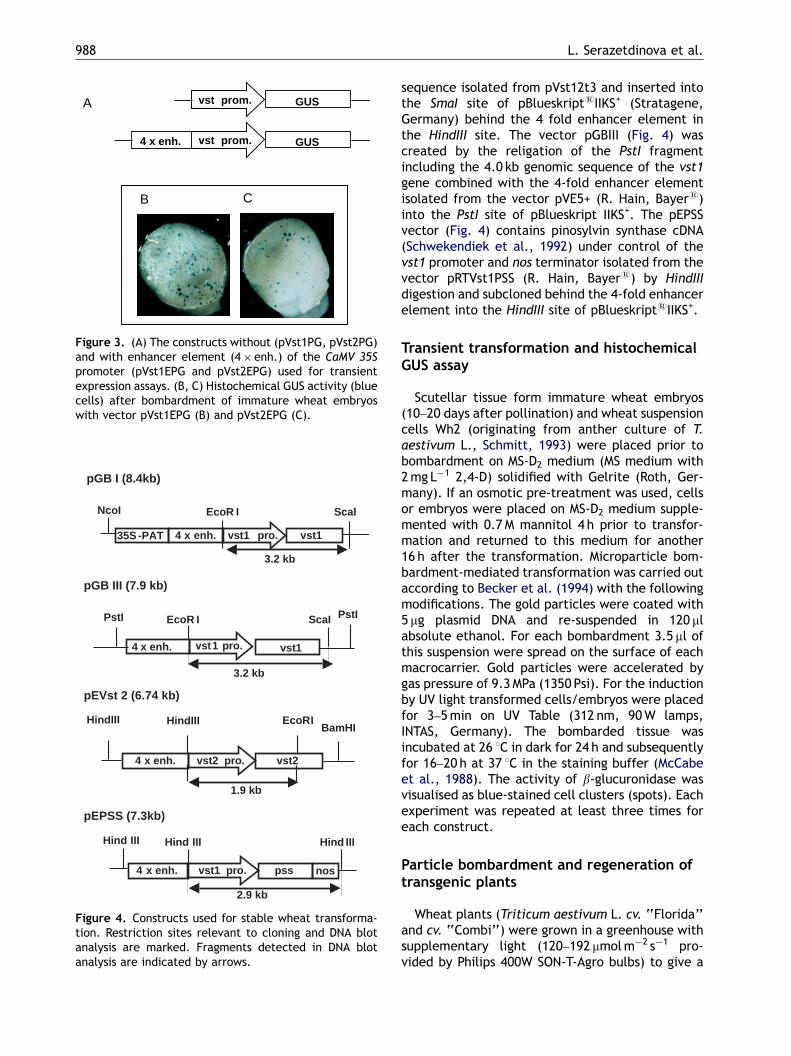
ARTICLE IN PRESS
A vst prom. GUS
vst prom. GUS4 x enh.
CB
Figure 3. (A) The constructs without (pVst1PG, pVst2PG)and with enhancer element (4� enh.) of the CaMV 35Spromoter (pVst1EPG and pVst2EPG) used for transientexpression assays. (B, C) Histochemical GUS activity (bluecells) after bombardment of immature wheat embryoswith vector pVst1EPG (B) and pVst2EPG (C).
pGB I (8.4kb)
3.2 kb
4 x enh. vst1 pro. vst1
NcoI EcoR I ScaI
35S -PAT
pGB III (7.9 kb)
4 x enh. vst1 pro. vst1
PstI EcoR I ScaI
3.2 kb
PstI
4 x enh. vst2 pro. vst2
HindIII EcoRI
pEVst 2 (6.74 kb)
1.9 kb
BamHIHindIII
pEPSS (7.3kb)
4 x enh. vst1 pro. pss
Hind III Hind III
2.9 kb
nos
Hind III
Figure 4. Constructs used for stable wheat transforma-tion. Restriction sites relevant to cloning and DNA blotanalysis are marked. Fragments detected in DNA blotanalysis are indicated by arrows.
L. Serazetdinova et al.988
sequence isolated from pVst12t3 and inserted intothe SmaI site of pBlueskriptsIIKS+ (Stratagene,Germany) behind the 4 fold enhancer element inthe HindIII site. The vector pGBIII (Fig. 4) wascreated by the religation of the PstI fragmentincluding the 4.0 kb genomic sequence of the vst1gene combined with the 4-fold enhancer elementisolated from the vector pVE5+ (R. Hain, Bayers)into the PstI site of pBlueskript IIKS+. The pEPSSvector (Fig. 4) contains pinosylvin synthase cDNA(Schwekendiek et al., 1992) under control of thevst1 promoter and nos terminator isolated from thevector pRTVst1PSS (R. Hain, Bayers) by HindIIIdigestion and subcloned behind the 4-fold enhancerelement into the HindIII site of pBlueskriptsIIKS+.
Transient transformation and histochemicalGUS assay
Scutellar tissue form immature wheat embryos(10–20 days after pollination) and wheat suspensioncells Wh2 (originating from anther culture of T.aestivum L., Schmitt, 1993) were placed prior tobombardment on MS-D2 medium (MS medium with2mg L�1 2,4-D) solidified with Gelrite (Roth, Ger-many). If an osmotic pre-treatment was used, cellsor embryos were placed on MS-D2 medium supple-mented with 0.7M mannitol 4 h prior to transfor-mation and returned to this medium for another16 h after the transformation. Microparticle bom-bardment-mediated transformation was carried outaccording to Becker et al. (1994) with the followingmodifications. The gold particles were coated with5 mg plasmid DNA and re-suspended in 120 mlabsolute ethanol. For each bombardment 3.5 ml ofthis suspension were spread on the surface of eachmacrocarrier. Gold particles were accelerated bygas pressure of 9.3MPa (1350 Psi). For the inductionby UV light transformed cells/embryos were placedfor 3–5min on UV Table (312 nm, 90W lamps,INTAS, Germany). The bombarded tissue wasincubated at 26 1C in dark for 24 h and subsequentlyfor 16–20 h at 37 1C in the staining buffer (McCabeet al., 1988). The activity of b-glucuronidase wasvisualised as blue-stained cell clusters (spots). Eachexperiment was repeated at least three times foreach construct.
Particle bombardment and regeneration oftransgenic plants
Wheat plants (Triticum aestivum L. cv. ‘‘Florida’’and cv. ‘‘Combi’’) were grown in a greenhouse withsupplementary light (120–192 mmolm�2 s�1 pro-vided by Philips 400W SON-T-Agro bulbs) to give a

ARTICLE IN PRESS
Transgenic stilbene synthases in wheat 989
16 h light period, and a temperature regime of 18/14 1C (light/dark). Scutellar tissue of immatureembryos was used for stable transformation. Isola-tion and transformation of scutellar tissue was doneaccording to Becker et al. (1994). The plasmidencoding the target gene and pAct1Dneo weremixed in a ratio 1.5:1.0. Cultivation of wheatembryos, callus induction and regeneration wasperformed as described by Kluth et al. (2002).
Induction of vst promoters in transgenicplants
For UV induction leaves were exposed for 10minto UV light (254 nm, 1200 J cm�2) in a crosslinker(Stratagene, Germany) followed by incubation ingrowth chamber with a 16 h light period, at a lightintensity of 120–144 mmolm�2 s�1, a temperatureregime of 18/14 1C (light/dark), and 70% relativehumidity. For wounding detached leaf segments orintact leaves were gently rubbed with wet seasand. After induction leaves were incubated inflasks with the cut surface submerged into water. Ifdetached leaf segments were induced, they werekept on damp filter paper in sealed Petri dishes. Forinduction with S. nodorum leaves were sprayedwith conidia suspension of S. nodorum (1� 106/mlwater with 0.01% (vol/vol) Tween 20 solution).
DNA blot analysis
Kanamycin-resistant plants were analysed byDNA blot analysis. Total genomic DNA was isolatedas described by Pallotta et al. (2000). Gel separa-tion, DNA blot analysis, hybridisation and detectionof transgenic DNA were performed as described byOldach et al. (2001) with minor modifications. 32P-dCTP labelled probes were synthesised with thePrime-It II Random Primer Labelling Kit (Strata-gene, Germany). Hybridisation and stringencywashed were carried out at 65 1C. Hybridised filterswere exposed to Hyperfilm (Kodak, USA) at �80 1Cfor 1–21 d with a 8-fold intensifier screen.
RNA blot analysis
Total RNA was isolated from leaves by usingTRIzol reagent (Gibco BRL, Germany) accordingto the manufacturer’s instructions. Total RNA(10–25 mg) was separated, blotted, and hybridisedwith 32P-dCTP labelled DNA probes (1.5–2.0�106 cpmml�1) at 67–68 1C as described by Sauter(1997). Blots were washed successively at 68 1C asdescribed by Oldach et al. (2001). Hybridised filterswere exposed to Hyperfilm (Kodak, USA) at �80 1C
with a 8-fold intensifier screen. Alternativelysignals were detected using Phosphorimager BioImaging Analyzer BAS-1000 and PCBAS 2.09 g soft-ware (Fujifilm, Raytest, Germany). To quantify theamount of specific transcripts in total RNA, filterswere successively hybridised to a specific probe andto the GAPDH or 18S rRNA probe. Prior to successivehybridisation hybridised probes were stripped offby washing the membrane twice in boiling 0.1%(w/v) SDS solution. The specific transcript amountwas calculated by normalising the signal intensityfrom the specific probe to the signal intensity ofGAPDH or 18S rRNA probe and expressed as arelative transcript amount for each sample.
Probes used for DNA and RNA hybridisation
PCR fragments or plasmid DNA fragments wereused for the synthesis of radioactively labelledprobes. The primers used for amplification of thevst1 promoter probe were 50-TATGATTCCGCGTT-CATGACT-30 and 50-GGAGGCTCATAGGTCGTTCAA-30
(Tan 56 1C). For the amplification of the vst2promoter probe the following primers were used50-GTCGAGCTATGGGTCAATTGA-30 and 50-GTAG-GATCCCTGCTGAAATCG-30 (Tan 59 1C). The vst1 cDNAprobe was amplified with primers 50-GAGGAAATTA-GAAACGCTCAACATGCC-30 and 50-GACAGTTCCAC-CTGCATAG-30 (Tan 59 1C). The vst2 cDNA probe wasamplified with primers 50-GACCGAGCTCAAGAA-GAAGT-30 and 50-ACGGAATGTAACAACGGTGAT-30
(Tan 59 1C). For the amplification of a pss cDNAprobe the following primers were used 50-GTGTGCGCGTTCGTGGAG-30 and 50-ACCGCGCCTT-TGAGTTGG-30 (Tan 56 1C). Wheat 18S rRNA fragment(0.4 kb) was amplified with primers: 50-CTGCCAG-TAGTCATATGCTTGTCT-30 and 50-CCCCGTGTCAG-GATTGG-30. Wheat GAPDH probe (0.6 kb) wasamplified with primers 50-AGGGTGGTGCAAA-GAAGGTCA-30 and 50-GATCCCCACTCGTTGTCGTA-30.
RT-PCR
Leaves induced by wounding and UV-irradiationwere incubated for 8 h (plants with the vst1promoter) and 24 h (plants with the vst2 promoter)prior to extraction of RNA. All S. nodorum-inoculated plants were incubated for 24 h at a highhumidity of 90% in the dark. Total RNA wasextracted according to Dresselhaus et al. (1996).DNAse I treatment and the first strand synthesiswere performed according to the manufacturer’sprotocol (MBI Fermentas, Germany). Specific in-tron-spanning primers were 50-GAGGAAATTAGAAACGCTCAACATGCC-30 and 50-GACAGTTCCACCTGCA-

ARTICLE IN PRESS
L. Serazetdinova et al.990
TAG-30 for the vst1 gene and 50-GACCGAGCTCAA-GAAGAAGT-30 and 50-ACGGAATGTAACAACGGTGAT-30
for the vst2 gene. As a reference GAPDH wasamplified with primers 50-AGGGTGGTGCAAA-GAAGGTCA-30 and 50-GATCCCCACTCGTTGTCGTA-30.
HPLC analysis of transgenic plants
Chemically pure standards, trans-resveratrol(30,40,50-trihydroxy-trans-stilbene, Sigma-Aldrich,Germany), pinosylvin (30,50-dihydroxy-trans-stil-bene) and trans-piceid (trans-resveratrol-3-b-D-glucoside, both from Apin Chemicals, UK) weredissolved in methanol. Leaves were induced by UVlight. Samples were taken 24 h after induction forthe vst1 and pss expressing plants (under thecontrol of the vst1 promoter) and 46 h afterinduction for the vst2 expressing plants taking intoaccount different timing of vst1 and vst2 transcriptaccumulation. Leave tissue (1–2 g) was extractedwith methanol/water mixture (8:2, v/v) as de-scribed by Jeandet et al. (1997). An aliquot of theextract (2ml) was purified on a MF C18 cartridge(Isolute, UK) and eluted with 1 volume of metha-nol/water solution. The eluate was evaporated todryness under argon at 36 1C and redissolved in400 ml methanol. For HPLC analysis 10 ml of eachsample was injected.
HPLC analysis was performed on a TSK gel ODS-80TM reversed-phase column (15 cm� 4.6mm,5 mm, Toyo Soda, Japan) preceded by a guardcolumn of LiChroCart 4-4 (100RP 18 endcapped,5 mm, Merck, Germany) using an Hewlett Packardsystem controller and sample injector series 1050,a photodiode array detector 1050 and 1046Afluorometer (Hewlett Packard, Germany). Phenoliccompounds were separated with a 15min lineargradient of 10mM ammonium acetate pH 3.5(solvent A) and methanol (solvent B) (0min 90%A–
10%B, 10min 20%A–80%B, 12min 90%A–10%B) withthe flow rate 1mlmin�1. Fluorometric detectionwas carried out at 315 nm excitation wavelengthand 380 nm emission wavelength. UV spectra weremeasured at 215–400 nm. Chromatograms wereanalysed with the software HPLC 3D Chem StationRev.SA.03.02 (Hewlett Packard, Germany). Toevaluate the efficiency of the extraction 100 mgof trans-resveratrol were added to 1 g of leavesand treated as described above. The recoveryof trans-resveratrol was 40%. The amount ofstilbenes in wheat extracts was calculated inresveratrol equivalents. Enzymatic hydrolysis ofputative resveratrol glucosides was performedwith b-D-glucosidase (Catalog No. 49290, Fluka,
Switzerland) as described by Hipskind and Paiva(2000).
Inoculation with Puccinia recondita andSeptoria nodorum
Transgenic plants of T1 and T2 progeny and wild-type control plants cv. ‘‘Combi’’ were grown ingrowth chamber with a 16 h light period, at a lightintensity of 120–144 mmolm�2 s�1, a temperatureregime of 18/14 1C (light:dark), and 70% relativehumidity. Plants from segregating lines were onlyevaluated if the presence of the target gene wasverified by PCR. For each individual line 10–15progeny plants were tested. Detached leaf assayswith P. recondita f. sp. tritici were carried out asdescribed (Oldach et al., 2001). Freshly harvestedconidia (7–10mg) were inoculated in a settlingtower giving the inoculum density of80–200 spores cm�2. The number of formed colo-nies was counted for ach leaf segment nine totwelve days after inoculation.
For the inoculation with S. nodorum Berkcommercial conidia suspension prepared from themixture of three field isolates of S. nodorum wasprovided by Prophyta GmbH (Germany). Detachedleaf assays with S. nodorum were carried out asdescribed by Arriano et al. (2001) with thefollowing modifications. Seedlings were evenlysprayed until run-off with S. nodorum conidiasuspension (1� 106/ml) in 0.01% Tween 20 solution.Leaves were left to dry for 1–2 h before 3.5 cm leafsegments were cut from primary fully expandedleaves. One leaf segment was tested per plant.Leaf segments were laid with adaxial side up onagar (0.8% (w/v), 50mg L�1 benzimidazole) across agap so that cut ends rested on agar. Strips of agarwere then laid over the cut edges to prevent dryingoff and senescence. Plastic boxes with leaf seg-ments were closed and incubated in dark at 25 1Cand 100% relative humidity for 20–24 h and thentransferred into a climate chamber at 20 1C,16 h day�1 light (150720 mEm�2 s�1) and 80% rela-tive humidity. Final evaluation was done 8 d afterinoculation when the percentage of diseased leafarea was scored for each leaf segment. For eachline mean value and standard deviation werecalculated and expressed in percents. Diseaseincidence of the wild-type control cv. ‘‘Combi’’was used as a reference (100%). Data werestatistically analysed using the Student t-test witha significance level of 5%. Wheat breeding lineshighly resistant (‘‘SN+’’) and susceptible (‘‘SN�’’)to Septoria leaf blotch served as controls and were

ARTICLE IN PRESS
Transgenic stilbene synthases in wheat 991
obtained from V. Aguilar, Institute of Plant ScienceETH Zurich (Switzerland).
Table 1. Histochemical GUS activity of the constructspVst1PG, pVst1EPG, pVst2PG, pVst2EPG and pAct1DGUSin wheat suspension cells. The constructs were biolisti-cally transformed in suspension cells (0.2–0.3 g per Petridish). An average number of blue spots (signals) wasdetermined for each construct from three repetitions.
Constructs Histochemical GUS activity
pAct1DGUS 66.0pVst1PG 1.7pVst1EPG 4.3pVst2PG 0.3pVst2EPG 0.7
Table 2. Influence of osmotic pre-treatment (osmot.)and exposure to UV light (UV) on histochemical GUSactivity of the constructs pVst1EPG, pVst2EPG andpAct1DGUS. The constructs were biolistically trans-formed in scutellum tissue of wheat immature embryosand the GUS activity was determined as an averagenumber of blue spots (signals) per embryo. Threerepetitions were carried out for each construct.
Treatments Experiment no.
11 33
No Osmot. UV No UV
pAct1DGUS 43.0 500.0 17.0 — —pVst1EPG 9.7 183.0 51.0 10.0 45.0pVst2EPG 1.4 43.0 5.5 3.0 20.0
Results
Functional analysis of vst1 and vst2promoters in cereal cells
In order to find a structural basis for differentialexpression of vst1 and vst2 genes we analysedgenomic sequences upstream from the vst1 andvst2 start codons (Fig. 2A). Sequence alignment hasshown a low overall homology (33%) between them.Beside nucleotide stretches unique for vst2, se-quence fragments highly similar and even identicalare present in the 5 prime regions of both the vst1and vst2 coding sequences. Significant differenceswere found in the composition and distribution ofputative cis-acting elements between the vst1 andvst2 5 prime flanking regions (Figs. 2B and C). Mostof the identified cis-acting elements in the vst1 5prime region are localised as a cluster between�960 and �110 bp relative to the start codon,whereas elements in the vst2 5 prime region areunevenly spread over the whole 2.0 kb fragment.The vst1 5 prime region contained two ethylene-responsive elements, had more salicylic acidresponsive elements and binding sites for MYBtranscription factors compared to the 5 primeregion of the vst2. By contrast, more elicitor-specific elements, wound-inducible elements, andlight-inducible elements were found in the vst2 5prime region. Apart from pathogen- and stress-inducible elements, both promoters contain ele-ments responsible for circadian regulation (notshown).
To investigate whether the grapevine promotersare active in wheat, 5 prime regions upstream ofthe vst1 and vst2 (cds) were analysed in transientexpression experiments. The 1.5 kb flanking regionof the vst1 cds and 0.8 kb flanking region of the vst2cds (Fig. 2A) were fused to the uidA gene encodingb-glucuronidase (GUS). The 0.8 kb vst2 fragmentwas chosen because it contained the majority ofpathogen- and stress-inducible cis-acting elementscompared to the whole 2.0 kb genomic sequence(Fig. 2C). The 4-fold enhancer element of the CaMV35S promoter (Odel et al., 1985) was insertedupstream of the vst promoters to increase thetranscription of the stilbene synthase genes inwheat.
The vst-GUS-constructs (Fig. 3A) without(pVst1PG, pVst2PG) and with enhancer element(pVst1EPG and pVst2EPG) were biolistically trans-
formed into scutellar tissue of immature wheatembryos and wheat suspension cells. HistochemicalGUS assays proved the activity of both tested vstpromoter sequences in wheat scutellum tissues(Figs. 3B and C) and suspension cells (Table 1). The4-fold enhancer intensified the transient expressionof both vst promoter sequences in wheat. Theactivity of the vst1 sequence was higher than thatof the vst2 sequence irrespective of the insertion ofthe 4-fold enhancer element (Figs. 3B and C, Table1). The transient expression of both vst promotersequences was higher in tissues that were inducedby abiotic stress factors, such as increased osmoticpressure and exposure to UV light (Table 2).
Our results indicate that both tested vst se-quences (0.8 kb vst2 and 1.5 kb vst1 flankingregions) are functional in cereals but differsignificantly in their activity. The 0.8 kb flankingregion of the vst2 cds was sufficient to driveinducible expression in wheat. Furthermore, theinsertion of the 4-fold enhancer element of theCaMV 35S promoter increases the expression level

ARTICLE IN PRESS
L. Serazetdinova et al.992
of both vst promoters in cereals but does notchange their relative activity. These results allowedus to use the inducible vst promoters in combina-tion with the 4-fold enhancer element to drive theexpression of STS genes in transgenic wheat.
Figure 5. DNA blot analysis of five pss T0 plants.Genomic DNA isolated from leaves of transgenic plantsand wild-type control (C) was hybridised with radio-actively labelled pss cDNA specific probe. For each T0
plant 25 mg of genomic DNA (lane u—uncut, lane a—digested with HindIII, lane b—digested with NcoI) waselectrophoretically separated and blotted. 4 out of 5analysed lines (59/1, 64/1, 64/3 and 64/6) show adifferent integration pattern. Two T0 plants (64/1 and64/2) possess an identical integration pattern of the pssgene and represent clonal lines. The 2.9 kb fragment(indicated by arrow) including promoter, pss cDNA andnos terminator is integrated in the genome of 4 out of 5analysed T0 lines (59/1, 64/1, 64/2 and 64/3). The T0
line 64/6 not showing the 2.9 kb fragment has only anincomplete integration of the pss expression cassette.
Stable transformation and inheritance ofvst1, vst2 and pss genes in transgenic wheat
To study the expression of stilbene synthasegenes in wheat we performed stable transformationexperiments. Immature embryos of the commercialbread wheat cultivars ‘‘Florida’’ and ‘‘Combi’’(Triticum aestivum L.) were used for stable biolistictransformation. Vectors with the vst genes underthe control of their endogenous inducible promo-ters and the pss gene under the control of the vst1promoter and nos terminator (Fig. 4) were co-transformed with vector pAct1Dneo containing theselection marker gene nptII mediating resistance tothe antibiotic kanamycin. Twenty eight indepen-dent transformation experiments resulted in 101kanamycin-resistant plants (Table 3). Genomic DNAblot analyses showed that 101 regenerants hadintegrated the vst1, vst2 or pss gene, as exempli-fied by Fig. 5 for pss transgenic plants. From 45transgenic lines, five showed a redundant integra-tion pattern and were characterised as clonal lines,whereas 42 transgenic lines originated from in-dependent integration events. The complete inte-gration of the target gene was proven in 34 out of42 transgenic lines (Table 3).
To investigate the inheritance of the transformedgenes, progenies of T0 plants were analysed bykanamycin spray test and subsequently by DNA blotprobed with the target gene. In 29 out of 30 testedlines, the integrated genes were inherited into thesubsequent generation. The co-segregation of theselection marker and the target gene was con-firmed in 24 out of 29 examined transgenic lines.The inheritance of target genes was analysed up tothe T2 generation by kanamycin spray test. Alltested 14 transgenic lines maintained the activityof the nptII gene.
Table 3. Overview of transgenic wheat plants
Targetgenes
Kanamycinresistantplants
DNA blotpositiveplants
IndependentT0 lines
T0 pcompinteg
vst1 15 13 13 13vst2 36 11 10 7pss 50 21 19 14Sum 101 45 42 34
Inducible expression of vst1, vst2 and pssgenes in wheat
Transgenic wheat plants were analysed for theinduction of vst promoters and accumulation ofstilbene synthase transcripts. In grapevine andtransgenic tobacco the vst1 promoter is rapidlyactivated and the vst1 transcripts accumulatebetween 2 and 8 h (h) after induction (Wiese etal., 1994; Fischer, 1994). To determine the timepoint of the maximal accumulation of the vst2transcripts we analysed the steady state level ofthe vst2 transcripts in leaf tissue of T0 line 30/7induced by wounding. RNA Blot hybridisation hasshown that the basal level of the vst2 transcriptsdid not change during the first 4.5 h after induction
lants withlete copiesration
Expressing T0
linesAnalysed T1
linesExpressingT1 lines
13 10 95 5 38 3 3
26 18 15

ARTICLE IN PRESS
Transgenic stilbene synthases in wheat 993
but rose substantially between 6 and 24 h afterinduction (Fig. 6).
The transcription of the vst1, vst2 and pss geneswas analysed in all transgenic lines after inductioneither by wounding or by UV light. Samples weretaken 8 h after wounding for the vst1 and pssexpressing plants and 24 h post induction for thevst2 expressing plants. Transcripts of the expectedsize, 1.20 kb for the vst1 and vst2 and the slightlysmaller 1.17 kb for the pss, were detected intransgenic wheat plants as exemplified in Fig. 7for the vst1 transcripts. The induced expressionvaried substantially between different transgeniclines (Fig. 7). Expression of the vst1, vst2 and pssgene was detected in nearly all analysed plants
Figure 6. The steady-state level of the vst2 transcriptsin T0 line 30/7. Leaves of the transgenic plant wereinduced by wounding with sand. Total RNA was extractedfrom non-induced and induced leaves at different timepoints after induction. For each probe 20 mg of total RNAwas electrophoretically separated, blotted and hybri-dised to a 3 prime vst2 probe (panel A) and subsequentlywith a 18 rRNA probe (panel B). The size of the vst2transcripts is indicated.
Figure 7. RNA blot analysis of vst1 T0 plants. Total RNAwas extracted from leaves 8 and 24 h after wounding. Foreach sample 15 mg of total RNA was electrophoreticallyseparated, blotted and hybridised with a 3 prime vst1probe (A). Specific vst1 transcripts (1.2 kb) were de-tected in all analysed plants. Subsequently the mem-brane was stripped and re-probed with a GAPDH probe asreference (B).
with complete integration of the target gene andremained stable in the subsequent generation(Table 3).
The inducibility of the vst1 and vst2 promotersand the splicing of vst1 and vst2 transcripts in thetransgenic wheat plants was analysed by RT-PCR.Leaves of T2 progeny plants of vst1 line 58/1 andvst2 line 32/7 were induced by wounding, UV lightor S. nodorum infection. The expected 493 bp cDNAfragment was amplified in samples isolated fromthe vst1 plant induced by UV light, wounding andinoculation with the pathogen (Fig. 8) and is theresult of splicing the 357 bp intron from the 850 bpgenomic vst1 sequence. No amplification productwas detected in cDNA from non-induced vst1plants. Similarly for the vst2 plants, a 449 bp cDNAfragment resulting from the splicing of a 136 bpintron from the 585 bp genomic sequence wasamplified in samples isolated from leaves inducedby wounding, UV light or S. nodorum infection (Fig.8). The same amplification product was visible innon-induced leaves of this vst2 line, indicating abasal level of the vst2 transcripts in this sample. Wehave observed an additional smaller cDNA fragmentof the approximate size of 300 bp in the samplesfrom this vst2 plant, which might result from thealternative splicing or incomplete copies of thetransgene.
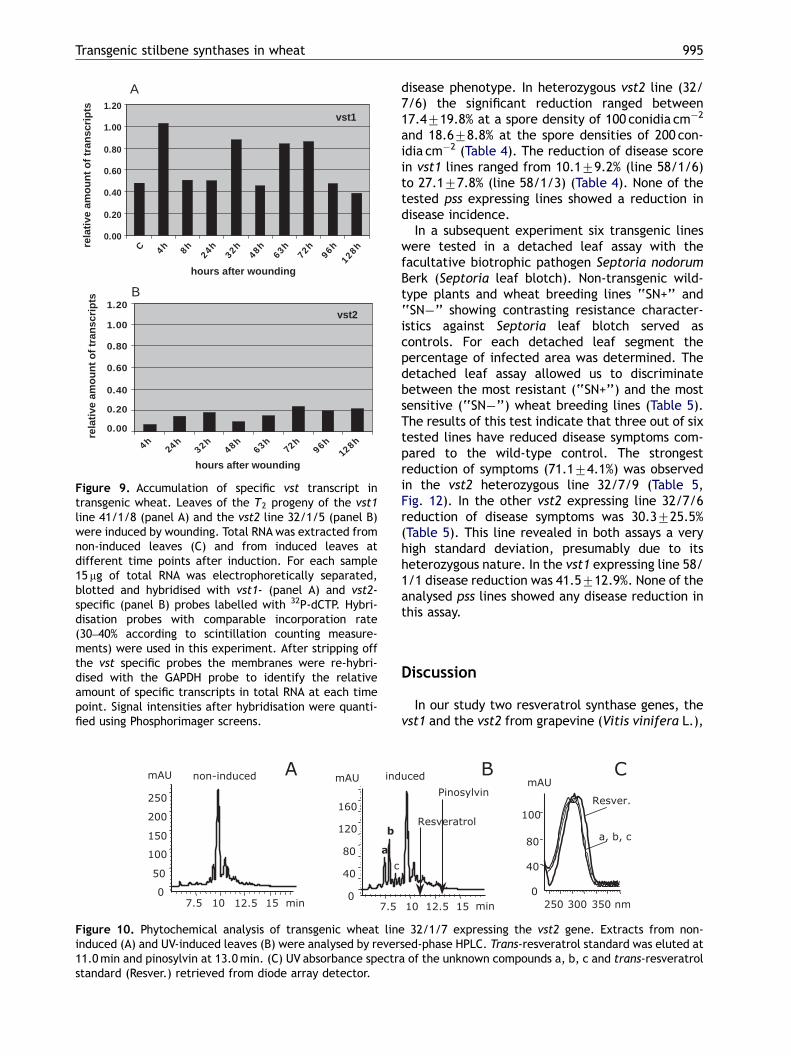
The transcription profiles of the vst1 and vst2genes in transgenic wheat were compared in T2
progeny plants by RNA blot analysis. Samples werecollected at different time points after wounding.The results have shown that the accumulation ofthe vst1 transcripts reached three maxima duringthe time course analysed: 4, 32 and 72 h afterinduction by wounding (Fig. 9A). By contrast, thevst2 transcripts reached less significant maximum32 and 72 h after induction and remained increasedbetween 96 and 128 h after induction (Fig. 9B). Forall lines examined, the vst1 and vst2 transcriptsaccumulated to different levels. Not only at theirfirst maximum (4 h for the vst1 transcripts and 32 hfor the vst2 transcripts) but also during the wholetime course after induction (0–128 h) the level ofthe vst1 transcripts remained four to six timeshigher than the level of the vst2 transcripts (Figs.9A and B). The same tendency was observed withother nine vst1 and four vst2 expressing lines (datanot presented).
Accumulation of stilbene derivatives intransgenic wheat lines
Individual STS expressing lines were analysed foraccumulation of stilbenes by reversed-phase HPLC

ARTICLE IN PRESS
Figure 8. RT-PCR of T2 progeny plants of the vst1 line 58/1/1 and the vst2 line 32/7/6. 1 mg total RNA isolated fromnon-induced leaves (control) and from leaves induced by wounding, UV light or inoculation with conidia of S. nodorumwas used in a reverse transcription reaction. RT-PCR products of the vst1 (493 bp) and the vst2 (449 bp) gene wereamplified with intron-flanking vst1/vst2 primers (panel A). Genomic fragments of the vst1 (850 bp) and the vst2 (585 bp)have been amplified from plasmid DNA (pGBI and pEVst2, respectively). In panel B the 620 bp product was amplifiedfrom the synthesized cDNA with intron-flanking GAPDH primers.
L. Serazetdinova et al.994
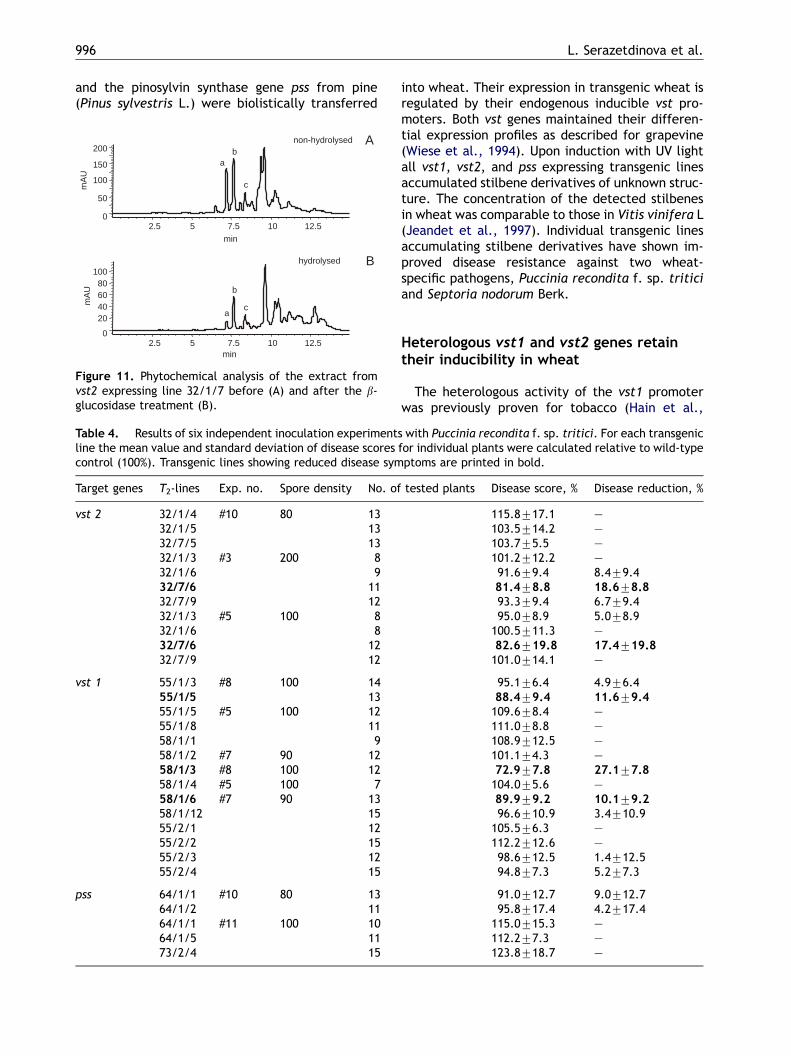
analysis. A HPLC profile of leaf extract from a vst2expressing line is shown in Fig. 10. The character-istic profile of phenolic compounds from non-induced leaves with one major peak and severalminor peaks (Fig. 10A) is identical to the profileobserved for non-transgenic plants (not shown).Fig. 10B shows the profile corresponding to theextract from leaves induced by UV light. In additionto the major peak, three unknown peaks with theretention times of 7.2, 7.6 and 8.4min appeared inthis extract. The same three peaks were detectedin all analysed vst1, vst2 and pss lines. Thepresence of the most and the least abundant peaks(retention time of 7.6 and 8.4min, respectively)and their relative abundance remained unchangedin all vst and pss expressing wheat lines. Thesepeaks were not detectable in non-transgenic con-trols and only as traces in non-induced transgenicplants. The UV spectra of the unknown peaks arealmost identical to those of trans-resveratrol withan absorbance maximum at 307 nm (Fig. 10C) andpinosylvin (not shown here). However, the reten-tion times of these three peaks (7.2, 7.6 and8.4min) were significantly shorter than those oftrans-resveratrol (11.0min) and pinosylvin(13.0min), indicating that these unknown com-pounds are more hydrophilic than the standards. Inorder to analyse whether their higher hydrophily iscaused by glycosylation of one or two hydroxylgroups of the stilbene backbone, we hydrolysedenzymatically the extracted stilbenes with b-D-glucosidase (EC 3. 2. 1. 21). A synthetic trans-resveratrol glucoside (trans-piceid) was used ascontrol. The glycosylated form of trans-resveratrol
peaked at a retention time of 9.5min and shifted to11.0min after hydrolysis. This new peak wascharacterised as trans-resveratrol based on theidentity of its retention time and UV spectrum tothose of trans-resveratrol (not shown). Althoughsome additional peaks appeared in the chromato-gram of hydrolysed extracts from transgenic plants,none of them represented either resveratrol orpinosylvin as exemplarily shown for the vst2 line inFig. 11. These results demonstrate that none of thethree unknown peaks represents trans-resveratrol-b-D-glucoside and so the nature of the identifiedpeaks remains unclear.
All twelve analysed T1 lines showed the accumu-lation of stilbene derivatives after induction by UVlight. The stilbene content varied drastically amonganalysed transgenic lines and ranged from 35 to190 mg of resveratrol equivalent per 1 g freshweight. No linear correlation was observed be-tween the concentration of the accumulatedstilbenes and the transcript levels of the integratedgenes (vst1, vst2 or pss).
Phytopathological analysis of transgenic lines
In the following experiments we analysed fungalresistance of the transgenic lines accumulating ahigh level of stilbene derivatives. Seven vst2, 13vst1 and four pss expressing lines were analysed ina detached leaf assay with the obligate biotrophicfungus Puccinia recondita f. sp. tritici (leaf rust ofwheat) (Table 4). Four transgenic lines (32/7/6, 58/1/3, 58/1/6, and 55/1/5) showed a reduced
ARTICLE IN PRESS
rela
tive
am
ou
nt
of
tran
scri
pts
rela
tive
am
ou
nt
of
tran
scri
pts
0.00
0.20
0.40
0.60
0.80
1.00
1.20
C 4h 8h
24h
32h
48h
63h
72h
96h128h
A
vst1
0.00
0.20
0.40
0.60
0.80
1.00
1.20
4h24h
32h48h
63h72h
96h128h
B
vst2
hours after wounding
hours after wounding
Figure 9. Accumulation of specific vst transcript intransgenic wheat. Leaves of the T2 progeny of the vst1line 41/1/8 (panel A) and the vst2 line 32/1/5 (panel B)were induced by wounding. Total RNA was extracted fromnon-induced leaves (C) and from induced leaves atdifferent time points after induction. For each sample15 mg of total RNA was electrophoretically separated,blotted and hybridised with vst1- (panel A) and vst2-specific (panel B) probes labelled with 32P-dCTP. Hybri-disation probes with comparable incorporation rate(30–40% according to scintillation counting measure-ments) were used in this experiment. After stripping offthe vst specific probes the membranes were re-hybri-dised with the GAPDH probe to identify the relativeamount of specific transcripts in total RNA at each timepoint. Signal intensities after hybridisation were quanti-fied using Phosphorimager screens.
mAU
0
50
100
150
200
250
min7.5 10 12.5 15
Anon-induced mAU
0
40
80
120
160
a
b
c
7.5
ind
Figure 10. Phytochemical analysis of transgenic wheat lininduced (A) and UV-induced leaves (B) were analysed by rever11.0min and pinosylvin at 13.0min. (C) UV absorbance spectrstandard (Resver.) retrieved from diode array detector.
Transgenic stilbene synthases in wheat 995
disease phenotype. In heterozygous vst2 line (32/7/6) the significant reduction ranged between17.4719.8% at a spore density of 100 conidia cm�2
and 18.678.8% at the spore densities of 200 con-idia cm�2 (Table 4). The reduction of disease scorein vst1 lines ranged from 10.179.2% (line 58/1/6)to 27.177.8% (line 58/1/3) (Table 4). None of thetested pss expressing lines showed a reduction indisease incidence.
In a subsequent experiment six transgenic lineswere tested in a detached leaf assay with thefacultative biotrophic pathogen Septoria nodorumBerk (Septoria leaf blotch). Non-transgenic wild-type plants and wheat breeding lines ‘‘SN+’’ and‘‘SN�’’ showing contrasting resistance character-istics against Septoria leaf blotch served ascontrols. For each detached leaf segment thepercentage of infected area was determined. Thedetached leaf assay allowed us to discriminatebetween the most resistant (‘‘SN+’’) and the mostsensitive (‘‘SN�’’) wheat breeding lines (Table 5).The results of this test indicate that three out of sixtested lines have reduced disease symptoms com-pared to the wild-type control. The strongestreduction of symptoms (71.174.1%) was observedin the vst2 heterozygous line 32/7/9 (Table 5,Fig. 12). In the other vst2 expressing line 32/7/6reduction of disease symptoms was 30.3725.5%(Table 5). This line revealed in both assays a veryhigh standard deviation, presumably due to itsheterozygous nature. In the vst1 expressing line 58/1/1 disease reduction was 41.5712.9%. None of theanalysed pss lines showed any disease reduction inthis assay.
Discussion
In our study two resveratrol synthase genes, thevst1 and the vst2 from grapevine (Vitis vinifera L.),
min10 12.5 15
B
Resveratrol
Pinosylvin
uced
nm250 300 350
mAU
0
40
80
100
a, b, c
C
Resver.
e 32/1/7 expressing the vst2 gene. Extracts from non-sed-phase HPLC. Trans-resveratrol standard was eluted ata of the unknown compounds a, b, c and trans-resveratrol
ARTICLE IN PRESS
L. Serazetdinova et al.996
and the pinosylvin synthase gene pss from pine(Pinus sylvestris L.) were biolistically transferred
min
min
2.5 5 7.5 10 12.5
mA
Um
AU
0
50
100
150
A
2.5 5 7.5 10 12.50
20406080
100B
a
a
c
c
b
b200non-hydrolysed
hydrolysed
Figure 11. Phytochemical analysis of the extract fromvst2 expressing line 32/1/7 before (A) and after the b-glucosidase treatment (B).
Table 4. Results of six independent inoculation experimentline the mean value and standard deviation of disease scorescontrol (100%). Transgenic lines showing reduced disease sym
Target genes T2-lines Exp. no. Spore density No. of
vst 2 32/1/4 #10 80 1332/1/5 1332/7/5 1332/1/3 #3 200 832/1/6 932/7/6 1132/7/9 1232/1/3 #5 100 832/1/6 832/7/6 1232/7/9 12
vst 1 55/1/3 #8 100 1455/1/5 1355/1/5 #5 100 1255/1/8 1158/1/1 958/1/2 #7 90 1258/1/3 #8 100 1258/1/4 #5 100 758/1/6 #7 90 1358/1/12 1555/2/1 1255/2/2 1555/2/3 1255/2/4 15
pss 64/1/1 #10 80 1364/1/2 1164/1/1 #11 100 1064/1/5 1173/2/4 15
into wheat. Their expression in transgenic wheat isregulated by their endogenous inducible vst pro-moters. Both vst genes maintained their differen-tial expression profiles as described for grapevine(Wiese et al., 1994). Upon induction with UV lightall vst1, vst2, and pss expressing transgenic linesaccumulated stilbene derivatives of unknown struc-ture. The concentration of the detected stilbenesin wheat was comparable to those in Vitis vinifera L(Jeandet et al., 1997). Individual transgenic linesaccumulating stilbene derivatives have shown im-proved disease resistance against two wheat-specific pathogens, Puccinia recondita f. sp. triticiand Septoria nodorum Berk.
Heterologous vst1 and vst2 genes retaintheir inducibility in wheat
The heterologous activity of the vst1 promoterwas previously proven for tobacco (Hain et al.,
s with Puccinia recondita f. sp. tritici. For each transgenicfor individual plants were calculated relative to wild-typeptoms are printed in bold.
tested plants Disease score, % Disease reduction, %
115.8717.1 —103.5714.2 —103.775.5 —101.2712.2 —91.679.4 8.479.481.478.8 18.678.893.379.4 6.779.495.078.9 5.078.9
100.5711.3 —82.6719.8 17.4719.8101.0714.1 —
95.176.4 4.976.488.479.4 11.679.4109.678.4 —111.078.8 —108.9712.5 —101.174.3 —72.977.8 27.177.8104.075.6 —89.979.2 10.179.296.6710.9 3.4710.9
105.576.3 —112.2712.6 —98.6712.5 1.4712.594.877.3 5.277.3
91.0712.7 9.0712.795.8717.4 4.2717.4
115.0715.3 —112.277.3 —123.8718.7 —

ARTICLE IN PRESS
Table 5. Results of the detached leaf assay with Septoria nodorum Berk. Non-transgenic wild-type genotype‘‘Combi’’ and breeding lines resistant (‘‘SN+’’) and susceptible (‘‘SN�’’) to S. nodorum served as controls. For each linethe mean value and standard deviation of the individual plants were calculated as percentage of diseased leaf arearelative to the wild-type control (100%).
Genotype (target gene) No. of plants analysed Diseased leaf area, % Disease score, % Disease reduction, %
‘‘SN�’’ 10 75.5725.9 118.9740.7 —‘‘SN+’’ 9 18.376.6 28.9710.4 71.1710.4‘‘Combi’’ 10 63.576.7 100.0710.5 032/7/6 (vst2) 7 44.3716.8 69.7725.5 30.3725.532/7/9 (vst2) 6 18.372.6 28.974.1 71.174.158/1/1 (vst1) 7 37.177.6 57.5712.9 41.5712.958/1/2 (vst1) 10 85.7715.0 135.0723.6 —64/1/1(pss) 7 71.7722.7 112.9735.7 —64/1/5 (pss) 10 80.8712.4 126.0719.6 —
Figure 12. Leaf segments after infection with Septoria nodorum Berk. Transgenic wheat lines 32/7/9 (vst2), 32/7/6(vst2) and 58/1/1 (vst1), wild-type genotype ‘‘Combi’’ as well as wheat breeding lines ‘‘SN+’’ (resistant) and ‘‘SN�’’(sensitive) were sprayed with conidia suspension of S. nodorum (1� 106/ml). Infection initiated from the cut edges ofthe leaf segments and spread towards the middle of the segment. Disease symptoms were assessed as percentage ofinfected leaf area 8 days after inoculation. Picture was taken 8 days after the inoculation when diseased leaf area ofeach individual segment was quantified.
Transgenic stilbene synthases in wheat 997
1993), tomato (Thomzik et al., 1997), rice (Stark-Lorenzen et al., 1997), barley and wheat (Leckbandand Lorz, 1998). Here we have shown that both thevst1 and vst2 promoter are functional in wheat. Sofar there had been no reports on the activity of thevst2 promoter in a heterologous system. As thetranscriptional activity of the vst promoters inwheat was very low, we combined the vst promo-ters with the enhancer element of the CaMV 35Spromoter (Odel et al., 1985) and increased theiractivity around three fold. The insertion of thisenhancer element in front of the vst promoters didnot inhibit their inducibility and did not alter theirspecific expression patterns as it was previouslyproven in tobacco and barley (Fischer, 1994;Leckband and Lorz, 1998).
In grapevine the vst promoters are induced bypathogens or their elicitors (Fischer, 1994) and byabiotic factors, including wounding (Leckband andLorz, 1998) and UV light (Jeandet et al., 1997). Wefound that in wheat both vst promoters wereinduced in response to wounding, UV light andpathogen infection. Expression of the foreign vst
genes in species that do not synthesise stilbenes,including tobacco, tomato, barley, rice, and wheat,indicates that transcription factors regulating ex-pression of the vst genes in grapevine are alsopresent in other species and must be ubiquitous.These transcription factors might be the same asthose involved in regulation of the phenylpropanoidmetabolism.
We found that the vst genes heterologouslyexpressed in wheat did not change their specificexpression patterns. The vst1 transcripts rapidlyaccumulated in wheat after induction and reachedseveral maxima as previously described for grape-vine and barley (Wiese et al., 1994; Leckband andLorz, 1998). Compared to the vst1 gene, thekinetics of the vst2 expression in heterologoussystem had not been studied before. We haveshown that the vst2 transcripts accumulatedsignificantly later and remained stable over alonger time period compared to the vst1 tran-scripts, resembling its expression in grapevine(Wiese et al., 1994). Apart from the temporaldifferences in expression, the expression level of

ARTICLE IN PRESS
L. Serazetdinova et al.998
the vst1 gene was four to six times higher than thatof the vst2 gene. This is in accordance with thefindings of Wiese et al. (1994) reporting on 10- to100-fold differences in the amount of accumulatedvst1 and vst2 mRNAs in grapevine.
We have observed several maxima in the inducedaccumulation of the vst transcripts consistent withthe presence of elements characteristic for circa-dian expression and numerous light-responsiveelements in the promoter regions of the vst genes.Circadian regulation of the expression was reportedfor the PR genes (Ward et al., 1991). It couldrepresent an evolutionary adaptation of the plantto pathogens with different timing of the plant-pathogen interaction and/or ensure a more con-tinuous supply for the fast degrading PR proteins.
Inducible accumulation of novel stilbenecompounds in wheat
HPLC analyses of methanolic extracts fromtransgenic wheat plants have shown that threeunknown compounds accumulate in transgenicplants after induction by UV light. Neither trans-resveratrol nor pinosylvin were detected in trans-genic wheat. In contrast to our results, freeresveratrol was detected in transgenic tobacco(Fischer, 1994) and tomato (Thomzik et al., 1997)expressing transgenic stilbene synthase by an ELISAanalysis. Fettig and Hess (1999) detected resvera-trol in transgenic and wild-type wheat plants usingacid hydrolysis of leaf extracts followed by HPLCanalysis. Latter stand in contrast to our findings, aswe did not detect any stilbene derivatives in wild-type wheat cv. ‘‘Combi’’. It is important to note,that esterified stilbene derivatives such as glycosy-lated forms, would release aglycones after acidhydrolysis. Stilbenes identified in our study weresignificantly more hydrophilic than trans-resvera-trol and pinosylvin. However, their UV spectra werecharacteristic for trans-stilbenes (Jeandet et al.,1997) and almost identical to those of trans-resveratrol and pinosylvin. In Vitis ssp. phenolicsubstances are often glucosylated in plants to makethem less toxic to the cell and more stable(Jeandet et al., 1997). The glycosylated form oftrans-resveratrol was also detected in transgenickiwi (Kobayashi et al., 2000), alfalfa (Hipskind andPaiva, 2000), and apple (Szankowski et al., 2003).However, b-glucosidase treatment did not releaseresveratrol or pinosylvin from our wheat extracts.Presumably stilbene derivatives accumulated inwheat contain polar moieties other than b-D-glucoside. Additional experiments are required to
clarify the nature of the stilbene derivativesaccumulated in wheat.
In our experiments the same three compoundswere detected in all vst and pss expressing lines.These compounds had different abundance, buttheir relative proportion remained unchanged in allvst and pss expressing wheat lines. These com-pounds may result from the use of alternativesubstrates and indicate a poor specificity of theresveratrol- and pinosylvin forming STSs in wheat. Asimilar suggestion was made in transgenic tobaccoexpressing the pss gene by Fischer (1994), whospeculated that instead of its original substratecinnamoyl-CoA (which might be limited in tobac-co), pinosylvin synthase accepts more abundant p-coumaroyl-CoA as a substrate and catalyses thesynthesis of resveratrol. The situation in transgenicwheat might be that both heterologously expressedSTSs are unable to recruit their specific substratesand use alternative substrates that may be avail-able in excess or at least be more abundant. Thepresence of three novel compounds in transgenicwheat suggests that either three different sub-strates are recruited by these enzymes, or thatthese different compounds represent subsequentsteps of stilbene modification in wheat. Theseassumptions are in agreement with the hypothesisof metabolic channelling, proposing that enzymesof the phenylpropanoid biosynthesis are organisedinto different membrane-associated multi-enzymecomplexes to control the flux of intermediatesbetween sequential reactions or between differentmetabolic branches. Common precursors are oftenunequally channelled through different pathways,and different isoforms of the enzymes may beorganised into different multienzyme complexes(Weisshaar and Jenkins, 1998; Winkel-Shirley,1999). Therefore, it is possible that the introducedstilbene synthases cannot enter the metabolicchannel or that the precursors are not freelyavailable in the cytoplasm.
The identified derivatives were present only intransgenic plants and their quantity rose signifi-cantly after induction with UV light concomitantwith the induction of the vst promoters. Apart fromthe homologous system of grapevine, inducibleaccumulation of resveratrol was demonstrated intobacco (Hain et al., 1990, 1993) and tomato(Thomzik et al., 1997). Inducible regulation ofstilbene biosynthesis is of significant advantage fora heterologous expression as it provides antimicro-bial compounds when they are required (e.g. afterpathogen attack or wounding) and does not inter-fere continuously with secondary biosyntheticpathways. A deleterious effect on the plantphenotype has been previously described when a

ARTICLE IN PRESS
Transgenic stilbene synthases in wheat 999
strong constitutive promoter had been chosen forthe expression of stilbene synthase. Male sterilityand altered pigment formation was observed intransgenic tobacco probably due to competitionbetween the introduced stilbene synthase and theendogenous chalcone synthase (EC 2.3.1.74) formetabolic intermediates (Fischer et al., 1997).
A semi-quantitative analysis of accumulatedstilbene derivatives in wheat has shown that theconcentration of stilbenes varied drastically indifferent transgenic lines, ranging from 35 to190 mg trans-resveratrol equivalents per g freshweight in high-level accumulating lines. Using thesame detection method, a comparable content oftrans-resveratrol (102 mg g�1 fresh weight) wasdetected in induced leaves of Vitis vinifera L(Jeandet et al., 1997). Leaves of transgenic kiwiaccumulated similar amounts of trans-piceid20–182 mg g�1 fresh weight (Kobayashi et al.,2000). A significantly lower trans-piceid contentof 0.5–20.0 mg g�1 fresh weight was detected intransgenic alfalfa (Hipskind and Paiva, 2000).
Surprisingly, in spite of the significant differencesin the activity of the vst1 and the vst2 promoters,no differences in accumulation of the stilbenederivatives were observed between the vst1, vst2and pss expressing lines. Furthermore, no correla-tion between the level of stilbene synthaseexpression and accumulation of stilbene derivativescould be found in our experiments. This resultindicates that accumulation of stilbenes mightcontinue until a certain threshold level is achieved.This upper limit might be reached when thesubstrates accepted by STSs are exhausted. Thiswould happen faster in a strongly expressing line,but more slowly in a line with a low transcript level.
Improved resistance to P. recondita f. sp.tritici and S. nodorum Berk in transgenicwheat
We investigated whether the accumulation ofunknown stilbene derivatives in wheat wouldimprove fungal resistance. Detached leaf assayswith the obligate biotrophic fungus Puccinia re-condita f. sp. tritici and facultative biotrophicpathogen Septoria nodorum Berk have shown thattransgenic lines expressing vst genes can haveimproved disease resistance.
Four out of 24 tested lines showed significantreduction of disease symptoms after leaf rustinfection ranging from 1979% to 2778%. An evenstronger effect was observed after Septoria leafblotch infection. Three out of six tested lines had areduction of disease symptoms ranging between
42713% and 7174%. None of the analysed pss linesshowed an improved resistance to S. nodorum or P.recondita f. sp. tritici, although comparable levelsof the accumulated stilbene derivatives weredetected in both vst and pss expressing lines afterinduction with UV light (data not shown).
Based on the fact that stilbenes are not synthe-sised in wild type wheat we refer the reduction ofsusceptibility in transgenic lines to the presence ofstilbene derivatives. Disease symptoms in trans-genic wheat were reduced but not completelyinhibited as described for a horizontal type ofresistance. This type of disease resistance wascharacteristic for other transgenic plant speciesexpressing stilbene synthase genes (Hain et al.,1993; Thomzik et al., 1997; Hipskind and Paiva,2000). Moreover, transgenic wheat plants showed adifferent level of resistance against two differentpathogens: a higher resistance against the faculta-tive pathogen S. nodorum (42713% and 7174%)compared to the obligate biotrophic pathogen P.recondita (1979%–2778%). A pathogen-dependentresistance response was also reported for trans-genic tomato. A significantly improved resistance(38–68%) was observed after inoculation with thehemibiotrophic pathogen Phytophtora infestans,whereas the necrotrophic pathogen Botrytis ciner-ea and the perthotrophic pathogen Alternariasolani were not inhibited (Thomzik et al., 1997).It is possible that S. nodorum is more sensitive tostilbene derivatives accumulated in transgenicwheat than P. recondita, or that P. recondita candegrade stilbenes accumulated in wheat. Numerousstudies identified the presence of a laccase-likestilbene oxidase in Botrytis cinerea that candetoxify stilbenes (Pezet et al., 1991; Adrian etal., 1998) and that the pathogenicity of Botrytiscinerea strains directly correlates with the degra-dation of stilbenes (Sbaghi et al., 1996).
In our experiments we have observed a bigvariation in disease resistance between differenttransgenic lines. Despite the similar steady-statelevels of the accumulated stilbene derivatives(measured at 24 and 46 h after induction in caseof the STS plants under the control of the vst1 andvst2 promoter, respectively) a few transgenic lineshave shown a higher reduction in susceptibility thanothers. The speed of stilbene accumulation mightplay a crucial role in the outcome of the plant-pathogen interaction. Thus, transgenic lines with ahigh basal level of STS transcripts (like some vst2lines) would have an increased level of the stilbenederivatives compared to other lines. Similarly, lineswith faster induction of the vst promoters (like thevst1 lines) would faster accumulate the effectiveconcentrations of the stilbenes after infection.

ARTICLE IN PRESS
L. Serazetdinova et al.1000
Additionally, the level of expression can be influ-enced by positional effects, which can positively ornegatively affect the transcription or post-tran-scriptional regulation of an integrated gene (Vau-chert and Fagard, 2001). The progenies of the sameparental lines, 58/1 and 32/7, have shown animproved disease resistance against both testedpathogens, indicating their transgenic genotypeseems to be more favourable for the expression ofthe STS genes and effective accumulation ofstilbene derivatives compared to other lines. It isunlikely that the insertion of the transgene wouldhave caused a mutation resulting in increasedresistance, the number of transgenic wheat lineswith improved resistance (six lines among 24 testedtransgenic wheat lines) is too high for such a rareevent.
Summarizing, our results demonstrate that het-erologous expression of stilbene synthases in wheatleads to inducible accumulation of unknown stil-bene derivatives. Their concentrations in wheat arecomparable to those of resveratrol in grapevine.Furthermore, transgenic wheat lines accumulatingstilbenes have shown improved disease resistanceagainst two pathogens. The accumulated stilbenederivatives either have a direct inhibitory effect onP. recondita and S. nodorum or may induce orintensify endogenous defence mechanisms. One ofthe most important questions to be answered in thefuture is the nature of the accumulated stilbenesand comprehensive analysis of their activity againstother economically important wheat pathogens.The antifungal effects of the stilbenes accumulatedin wheat could be further increased by theirmodification into more lipophilic substances. Forexample, ectopic expression of o-methyltransfer-ase could lead to the accumulation of methylatedforms of stilbenes that have higher fungitoxiceffects than non-methylated forms (Chiron et al.,2000).
Another promising application of the heterolo-gous expression of stilbene synthases in wheatcould result in beneficial effects on human health.Stilbenes were shown to have antioxidative, anti-mutagenic, immunoregulating, and anti-inflamma-tory effects that account for the health-promotingproperties of red wine (Pace-Asciak et al., 1995;Jang et al., 1997; Teguo et al., 1998). Apart fromresveratrol and its glucoside, other stilbenes suchas astringinin and 3,30,4-trihydroxystilbene havebeen shown to have activities described forresveratrol (Kimura et al., 1995; van Norren etal., 1997). It could therefore be assumed thatstilbene derivatives accumulated in transgenicwheat lines might also have health-promotingactivities.
Acknowledgments
The authors would like to thank Prof. R. Hain,Bayer CropScience AG (Monheim am Rhein, Ger-many) for providing the plasmids pVst12t3, pVE5+,and pRTVst1PSS. We are grateful to V. Aguilar,Institute of Plant Science ETH Zurich, (Zurich,Switzerland) for seeds of wheat breeding lines‘‘SN+’’ and ‘‘SN�’’. We express our gratitude to Dr.W. Pfau, Institute for Experimental and ClinicalPharmacology and Toxicology, University HospitalEppendorf (Hamburg, Germany) for the assistancewith HPLC analysis.
References
Adrian M, Jeandet P, Veneau J, Weston LA, Bessis R.Biological activity of resveratrol, a stilbene compoundfrom grapevines, against Botrytis cinerea, the causalagent for grey mould. J Chem Ecol 1997;23:1689–702.
Adrian M, Rajaei P, Jeandet P, Veneau J, Bessis R.Resveratrol oxidation in Botrytis cinerea conidia.Phytopathol 1998;88:472–6.
Arriano LS, Brading PA, Brown JKM. A detached seedlingleaf technique to study resistance to Mycosphaerellagraminicola (anamorph Septoria tritici) in wheat.Plant Pathol 2001;50:339–46.
Becker D, Brettschneider R, Lorz H. Fertile transgenicwheat from microprojectile bombardment of scutellartissue. Plant J 1994;5:299–307.
Chiron H, Drouet A, Claudot AC, Eckerskorn C, Trost M,Heller W, Ernst D, Sandermann H. Molecular cloningand functional expression of a stress-induced multi-functional O-methyltransferase with pinosylvinmethyltransferase activity from scots pine (Pinussylvestris L.). Plant Mol Biol 2000;44:733–45.
Dercks W, Creasy LL. The significance of stilbenephytoialexines in the Plasmopara viticola-grapewineinteraction. Physiol Mol Plant Pathol 1989;34:189–202.
Dresselhaus T, Hagel C, Lorz H, Kranz E. Isolation of a fulllength cDNA encoding calreticulin from PCR library ofin vitro zygotes of maize. Plant Mol Biol 1996;31:23–34.
Fettig S, Hess D. Expression of a chimeric stilbenesynthase gene in transgenic wheat lines. TransgenicRes 1999;8:179–89.
Fischer R. Optimierung der heterologen expression vonstilbensynthasegenen fur den Pflanzenschutz. Ph Ddissertation, University of Hohenheim, Faculty ofBiology, Hohenheim, Germany, 1994.
Fischer R, Budde I, Hain R. Stilbene synthase geneexpression causes changes in flower color and mailsterility in tobacco. Plant J 1997;11:489–98.
Gorham J. Stilbenes and phenantrens. In: Harborne JB,editor. Methods in plant biochemistry, plant phenolics.London, England: Academic Press; 1989. p. 159–96.
Hain R, Bieseler B, Kind H, Schroder G, Stocker R.Expression of stilbene synthase in Nicotiana tabacum

ARTICLE IN PRESS
Transgenic stilbene synthases in wheat 1001
results in synthesis of the phytoalexin resveratrol.Plant Mol Biol 1990;15:325–35.
Hain R, Reif HJ, Krause E, Langbartels R, Kindl H, VornamB, Wiese W, Schmelzer E, Schreier PH, Stocker RH,Stenzl K. Disease resistance results from foreignphytoalxine expression in a novel plant. Nature1993;361:153–6.
Hipskind JD, Paiva NL. Constitutive accumulation of aresveratrol-glucoside in transgenic alfalfa increasesresistance to Phoma medicagis. Mol Plant MicrobeInteract 2000;13:551–62.
Jang M, Chai L, Udeani GO, Slowing KV, Thomas CF,Beecher CWW, Fong HHS, Farnsworth NR, Kinghorn AD,Mehta RG, Moon RC, Pezzuto JM. Cancer chemopre-ventive activity of resveratrol, a natural productderived from grapes. Science 1997;275:218–20.
Jeandet P, Breuli AC, Adrian M, Westone LA, Debord S,Meunier P, Maume G, Bessis R. HPLC-analysis ofgrapewine phytoalxines coupling photodiode arraydetection and fluometry. Anal Chem 1997;69:5172–7.
Kimura Y, Okuda H, Kubo M. Effects of stilbenes isolatedfrom medical plants on arachidonate metabolism anddegranulation in human polymorphonuclear leuko-cytes. J Ethnopharmacol 1995;45:131–9.
Kluth A, Sprunck S, Becker D, Lorz H, Lutticke S. 50
deletion of a gbss1 promoter region from wheat leadsto changes in tissue and developmental specificities.Plant Mol Biol 2002;49:669–82.
Kobayashi S, Ding CK, Nakamura Y, Nakajima I, Matsu-moto R. Kiwifruits (Actiniidia deliciosa) transformedwith a Vitis stilbene synthase gene produce piceid(resveratrol-glucoside). Plant Cell Rep 2000;19:904–10.
Langcake P, McCarthy WV. The relationship of resveratrolproduction to infection of grapevine by Botrytiscinerea. Vitis 1979;18:244–53.
Leckband G, Lorz H. Transformation and expression of astilbene synthase gene of Vitis vinifera L. in barley andwheat for increased fungal resistance. Theor ApplGenet 1998;96:1004–12.
McCabe DE, Swain WF, Martinell BJ, Christou P. Stabletransformation of Soybean (Glycine max) by particlebombardment. Bio Technol 1988;6:923–6.
McElroy D, Thang W, Cao J, Wu R. Isolation of an efficientactin promoter for rice transformation. Plant Cell1990;2:163–71.
Muller E. Molekularbiologische Untersuchungen zur Va-riabilitat in transgenen Zellkulturen und in vitro-Regeneraten von Getreiden. Ph D dissertation, Uni-versity of Hamburg, Faculty of Biology, Hamburg,Germany, 1994.
Odel JT, Nagy F, Chua NH. Identification of DNA sequencesrequired for activity of the cauliflower mosaic virus35S promoter. Nature 1985;313:810–2.
Oldach KH, Becker D, Lorz H. Heterologous expression ofgenes mediating enhanced fungal resistance in trans-genic wheat. Mol Plant Microb Interact 2001;14:832–8.
Pace-Asciak CR, Hahn S, Diamandis EP, Soleas G, GoldbergDM. The red wine phenolics trans-resveratrol andquercetin block human platelet aggregation and
ecosanoid synthesis: implications for protectionagainst coronary heart disease. Clin Chem Acta1995;235:207–19.
Pallotta MA, Graham RD, Langridge P, Sparrow DHB,Barker SJ. RFLP mapping of manganese efficiency inbarley. Theor Appl Genet 2000;101:1100–8.
Pezet R, Pont V, Hoangvan K. Evidence for oxi-dative detoxification of pterostilbene and resveratrolby a laccase-like stilbene oxidase produced byBotrytis cinerea. Physiol Mol Plant Pathol 1991;39:441–50.
Pont V, Pezet R. Relation between the chemical structureand the biological activity of hydroxystilbenes againsBotrytis cinerea. J Phytoapathhol 1990;130:1–8.
Sauter M. Differential expression of a CAK (cdc2-activat-ing kinase)-like protein kinase, cyclins and cdc2 genesfrom rice during the cell cycle and in response togibberelin. Plant J 1997;11:181–90.
Sbaghi M, Jeandet P, Bessis R, Leroux P. Degradation ofstilbene-type phytoalexines in relation to the patho-genecity of Botrytis cinerea. Plant Pathol 1996;45:139–44.
Schmitt M. In vitro Kultur und Protoplastenregenerationvon Weizen (Triticum aestivum L.). Ph D dissertation,University of Hamburg, Faculty of Biology, Hamburg,Germany, 1993.
Schwekendiek A, Pfeffer G, Kindl H. Pine stilbenesynthase cDNA, a tool for probing environmentalstress. FEBS Lett 1992;301:41–4.
Stark-Lorenzen P, Nelke B, Hansler G, Muhlbach HP,Thomzik JE. Transfer of grapevine stilbene synthasegene to rice (Oryza sativa L.). Plant Cell Rep1997;16:668–73.
Szankowski I, Briviba K, Fleshhut J, Schonherr J,Jacobsen HJ, Kiesecker H. Transformation of apple(Malus domestica Borkh.) with the stilbene synthasegene from grapevine (Vitis vinifera L.) and a PGIPgene from kiwi (Actinidia deliciosa). Plant Cell Rep2003;22:141–9.
Teguo WP, Fauconneau B, Deffiex G, Huguet F, Vercau-teren J, Merillon JM. Isolation, identification, andantioxidant activity of three stilbene glucosides newlyextracted from Vitis vinifera cell cultures. J Nat Prod1998;61:655–7.
Thomzik JE, Stenzel K, Stocker R, Schreier PH, Hain R,Stahl DJ. Synthesis of a grapevine phytoalexin intransgenic tomatoes (Lycopersicon esculentum Mill.)conditions resistance against Phytophtora infestans.Physiol Mol Plant Pathol 1997;51:265–78.
van Norren K, Borggreven JMPM, Hovingh A, Willems HL,de Boo T, Elving LD, Berden JHM, DePont JJHHM.Antioxidant activity of the Stilbenes astringin, newliyextracted from Vitis vinifera cell cultures. Clin Chem1997;43:1092–3.
Vauchert H, Fagard M. Transcriptional gene silencing inplants: targets, inducers and regulators. Trends PlantSci 2001;17:27–35.
Ward ER, Uknes SJ, Williams SC, Dincher SS, WiederholdDL, Alexander DC, Ahl-Goy P, Metraux JH, Ryals JA.Coordinate gene activity in response to agents that

ARTICLE IN PRESS
L. Serazetdinova et al.1002
induce systemic acquired resistance. Plant Cell1991;3:1085–94.
Weisshaar B, Jenkins GI. Phenylpropanoid biosynthesisand its regulation. Curr Opin Plant Biol 1998;1:251–7.
Wiese W, Vornam B, Krause E, Kindl H. Structuralorganization and differential expression of three
stilbene synthase genes located on a 13 kbgrapevine DNA fragment. Plant Mol Biol 1994;26:667–77.
Winkel-Shirley B. Evidence for enzyme complexes in thephenylpropanoid and flavonoid pathways. PhysiolPlant 1999;107:142–9.
Related Documents











