DEVELOPMENTAL DYNAMICS 206.343-353 (1996) Expression of the cCdx-B Homeobox Gene in Chick Embryo Suggests its Participation in Rostrocaudal Axial Patterning AIXA V. MORALES, ENRIQUE J. de la ROSA, AND FLORA de PABLO Department of Cell and Deuelopmental Biology, Centro de Investigaciones Biolbggicas, CSIC, E-28006 Madrid, Spain ABSTRACT cCdx-B (formerly cHox-cad 2) is a chick homeobox-containing gene related to the Drosophila caudal. Compared with other caudal homologues, its similarity is highest with the mu- rine Cdx-4. In the present study, we characterize the localization of cCdx-B transcripts to the cau- dal region of the embryo by using reverse tran- scription-polymerase chain reaction (RT-PCR) and, in detail, by using in situ hybridization. Chick embryos from gastrulation to early organogenesis were hybridized with digoxigenin-labeled ribo- probes, and the pattern of expression of cC&x-B mRNA was analyzed in wholemount embryos and in tissue sections. In the early gastrula, tran- scripts were localized in a gradient through the caudal half of the embryo, in the epiblast and the mesoderm cells, but not including Hensen’s node. During neurulation, cC&-B transcripts were found more rostrally, with high levels localized in Hensen’s node and the posterior neural plate. Ex- pression was also high in paraxial mesoderm, with a rostral limit in the most recently formed somite. There was no expression in definitive en- doderm. During late neurulation and tail bud for- mation, cCdx-B mRNA expression regressed pos- teriorly and was finally confined to the tail bud region. This pattern of expression of cCdx-B, reg ulated in time and space, is different from that of the other known chick caudal homologue, cCdx-A. Both genes may play a coordinated role in the posterior axial patterning of the chick embryo, whereas cCdx-B may specify further the identity of the tail region. o 1996 Wiley-Liss, Inc. Key words: Homeobox, Caudal, Neurulation, Hensen’s node INTRODUCTION The characterization of the molecular mechanisms underlying vertebrate axis formation during early de- velopment has progressed remarkably by applying con- cepts and molecular tools derived from Drosophila. Multiple genes that are structurally similar to the Dro- sophila genes involved in embryonic axis and pattern formation have been cloned and characterized in ver- tebrates as those of the Hox complexes, homologous to the homeotic complex or HOM-C in Drosophila (for re- @ 1996 WILEY-LISS, INC. views, see McGinnis and Krumlauf, 1992; Krumlauf, 1994). Other Drosophila genes are required to specify the identities of the most anterior and the most poste- rior regions in the anteroposterior (A-P) axis and are expressed in unique A-P domains in early embryos. Among them, orthodenticle (otd) and empty spiracles (ems) seem to play an important role in the head seg- mentation of Drosophila (Dalton et al., 1989; Cohen and Jurgens, 1990).Their counterparts in mouse (Otx I and 2 and Em 1 and 2) are expressed in overlapping domains in forebrain and midbrain (Simeone et al., 1992; Rubenstein et al., 1994) rostrally to the expres- sion domains of the Hox-C genes. Less is known about the genes involved in pattern- ing of the embryo posterior region, but caudal (cad appears to be one of them. The highest level of cad protein is found in Drosophila embryos before gastru- lation at the posterior end region (Macdonald and Struhl, 1986; Mlodzik and Gehring, 1987a) and seems to play a role in the organization of this region (Mac- donald and Struhl, 1986; Mlodzik et al., 1990; Schulz and Tautz, 1995). Vertebrate genes containing a ho- meodomain similar to that of cad have been found in mouse (Duprey et al., 1988; James and Kazenwadel, 1991; Gamer and Wright, 19931, rat (Freund et al., 19921, hamster (German et al., 1992), chick (Frumkin et al., 1991; Serrano et al., 1993), Xenopus (Blumberg et al., 1991; Northrop and Kimelman, 1994), and ze- brafish (Joly et al., 1992). The expression patterns of these vertebrate cad counterparts resemble the zygotic expression patterns of Drosophila in both early and late development. Interestingly, in some vertebrates, there is more than one member of the cad family, and it is an open question whether each gene has a different role. The chick embryo provides an excellent vertebrate model system for the study of early development, be- cause it is easily accessible during gastrulation and early neurulation. The first gene of the cad family re- ported in chick, cCdx-A, is activated early in the three layers of the gastrula. Its expression disappears early in neurulation, although it is up-regulated again in Received November 2,1995; accepted January 29,1996. Address reprint requests/correspondence to Dr. F. de Pablo, Centro de Investigaciones Biol6gicas, CSIC, Velbquez 144, E-28006 Madrid, Spain.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DEVELOPMENTAL DYNAMICS 206.343-353 (1996)
Expression of the cCdx-B Homeobox Gene in Chick Embryo Suggests its Participation in Rostrocaudal Axial Patterning AIXA V. MORALES, ENRIQUE J. de la ROSA, AND FLORA de PABLO Department of Cell and Deuelopmental Biology, Centro de Investigaciones Biolbggicas, CSIC, E-28006 Madrid, Spain
ABSTRACT cCdx-B (formerly cHox-cad 2) is a chick homeobox-containing gene related to the Drosophila caudal. Compared with other caudal homologues, its similarity is highest with the mu- rine Cdx-4. In the present study, we characterize the localization of cCdx-B transcripts to the cau- dal region of the embryo by using reverse tran- scription-polymerase chain reaction (RT-PCR) and, in detail, by using in situ hybridization. Chick embryos from gastrulation to early organogenesis were hybridized with digoxigenin-labeled ribo- probes, and the pattern of expression of cC&x-B mRNA was analyzed in wholemount embryos and in tissue sections. In the early gastrula, tran- scripts were localized in a gradient through the caudal half of the embryo, in the epiblast and the mesoderm cells, but not including Hensen’s node. During neurulation, cC&-B transcripts were found more rostrally, with high levels localized in Hensen’s node and the posterior neural plate. Ex- pression was also high in paraxial mesoderm, with a rostral limit in the most recently formed somite. There was no expression in definitive en- doderm. During late neurulation and tail bud for- mation, cCdx-B mRNA expression regressed pos- teriorly and was finally confined to the tail bud region. This pattern of expression of cCdx-B, reg ulated in time and space, is different from that of the other known chick caudal homologue, cCdx-A. Both genes may play a coordinated role in the posterior axial patterning of the chick embryo, whereas cCdx-B may specify further the identity of the tail region. o 1996 Wiley-Liss, Inc.
Key words: Homeobox, Caudal, Neurulation, Hensen’s node
INTRODUCTION The characterization of the molecular mechanisms
underlying vertebrate axis formation during early de- velopment has progressed remarkably by applying con- cepts and molecular tools derived from Drosophila. Multiple genes that are structurally similar to the Dro- sophila genes involved in embryonic axis and pattern formation have been cloned and characterized in ver- tebrates as those of the Hox complexes, homologous to the homeotic complex or HOM-C in Drosophila (for re-
@ 1996 WILEY-LISS, INC.
views, see McGinnis and Krumlauf, 1992; Krumlauf, 1994). Other Drosophila genes are required to specify the identities of the most anterior and the most poste- rior regions in the anteroposterior (A-P) axis and are expressed in unique A-P domains in early embryos. Among them, orthodenticle (otd) and empty spiracles (ems) seem to play an important role in the head seg- mentation of Drosophila (Dalton et al., 1989; Cohen and Jurgens, 1990). Their counterparts in mouse (Otx I and 2 and E m 1 and 2) are expressed in overlapping domains in forebrain and midbrain (Simeone et al., 1992; Rubenstein et al., 1994) rostrally to the expres- sion domains of the Hox-C genes.
Less is known about the genes involved in pattern- ing of the embryo posterior region, but caudal (cad appears to be one of them. The highest level of cad protein is found in Drosophila embryos before gastru- lation at the posterior end region (Macdonald and Struhl, 1986; Mlodzik and Gehring, 1987a) and seems to play a role in the organization of this region (Mac- donald and Struhl, 1986; Mlodzik et al., 1990; Schulz and Tautz, 1995). Vertebrate genes containing a ho- meodomain similar to that of cad have been found in mouse (Duprey et al., 1988; James and Kazenwadel, 1991; Gamer and Wright, 19931, rat (Freund et al., 19921, hamster (German et al., 1992), chick (Frumkin et al., 1991; Serrano et al., 1993), Xenopus (Blumberg et al., 1991; Northrop and Kimelman, 1994), and ze- brafish (Joly et al., 1992). The expression patterns of these vertebrate cad counterparts resemble the zygotic expression patterns of Drosophila in both early and late development. Interestingly, in some vertebrates, there is more than one member of the cad family, and it is an open question whether each gene has a different role.
The chick embryo provides an excellent vertebrate model system for the study of early development, be- cause it is easily accessible during gastrulation and early neurulation. The first gene of the cad family re- ported in chick, cCdx-A, is activated early in the three layers of the gastrula. Its expression disappears early in neurulation, although it is up-regulated again in
Received November 2, 1995; accepted January 29,1996. Address reprint requests/correspondence to Dr. F. de Pablo, Centro
de Investigaciones Biol6gicas, CSIC, Velbquez 144, E-28006 Madrid, Spain.

344 MORALES ET AL.
endodermal organs later in development (Doll and Niessing, 1993; Frumkin et al., 1993, 1994).
We found a second member of the chick cud family, cCdx-B, whose expression is highest during neurula- tion, as shown by using the reverse transcription-poly- merase chain reaction (RT-PCR; Serrano et al., 1993). In the present study, we were interested in analyzing in detail cCdx-B expression and comparing its localiza- tion to other vertebrate cad-related genes. In situ hy- bridization of whole embryos from gastrulation (em- bryonic day 0.5 (E0.51, corresponding to stages 4-6 of Hamburger and Hamilton (1951; HH4-6) to the begin- ning of organogenesis (E2.5; HH15-17) showed that cCdx-B mRNA expression peaks later and is main- tained longer in the caudal (posterior) region than cCdx-A, with partially overlapping cellular localiz,a- tion.
RESULTS cCdx-B Protein Homeodomain Comparison With Other Cud-Related Genes
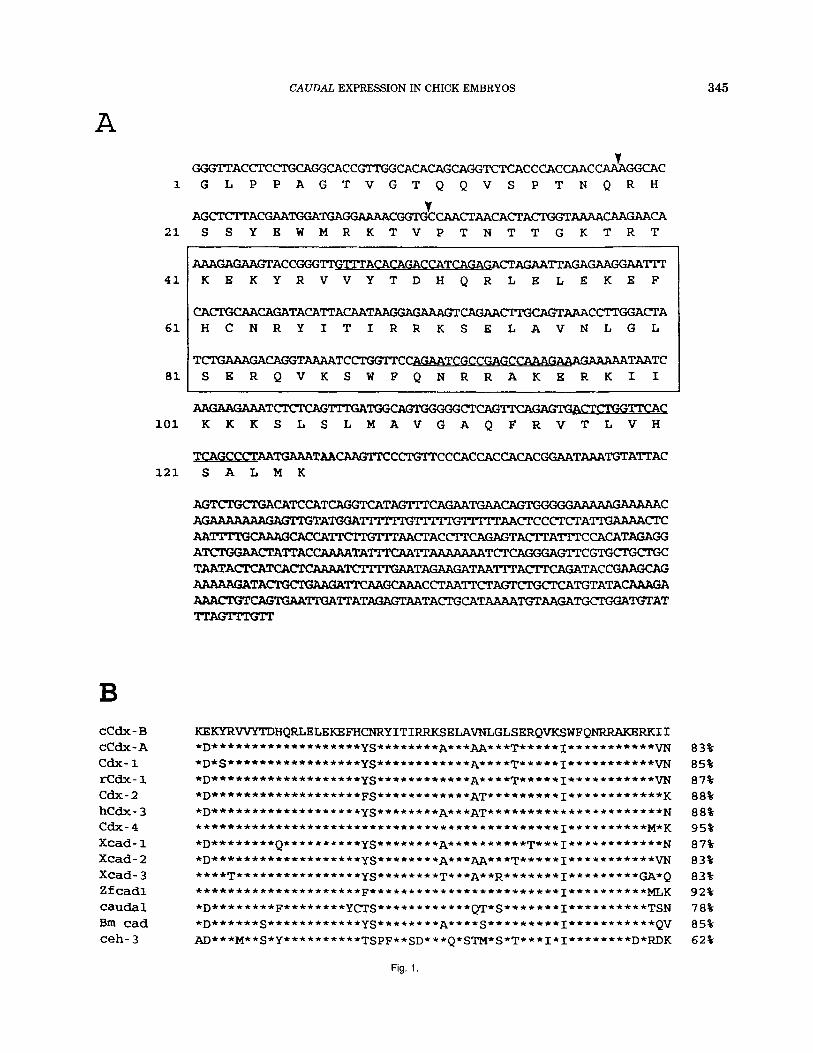
We previously reported the cloning and homeobox sequence of cHox-cud 2 (Serrano et al., 1993), which is now named cCdx-B, according to the report on cCdx-A by Frumkin et al. (1993). The available clone, repre- senting a partial cDNA of the 2.6 Kb cCdx-B tran- script, encodes putatively for 90 amino acids in open reading frame upstream of the homeobox and includes the translational stop codon, 35 amino acids down- stream of the homeobox (Fig. 1A).
At the amino acid level, the cC&-B homeobox was similar to the cud-related homeoboxes (Fig. lB), with a 78% identity with the Drosophilu protein and identities ranging from 62% with the C. eleguns ceh-3, to 95% with the mouse Cdx-4. It was only 83% identical to cCdx-A , perhaps implying slightly different roles.
Temporal Pattern of cC&-B Gene Expression in Early Chick Embryos
By using a sensitive RT-PCR method, we examined the expression of cCdx-B mRNA from unincubated blastoderm (EO) to E3 in the whole chick embryo and in the rostra1 and caudal regions at E1.5 (taking Hensen’s node as a limit; Fig. 2). cCdx-B transcripts were al- ready present at low levels at EO (corresponding to HH1-21, then they increased, reaching the highest ex- pression level a t the beginning of neurulation (El; HH7-8, 1-6 somites), and later decreased during orga- nogenesis (E2-3; HH15-18, 25-35 somites). The p-uc- tin cDNA amplified in parallel as a control showed a very different pattern, with the signal increasing pro- gressively from EO to E2 (results not shown; see PBrez- Villamil et al., 1994). At E1.5 (HH9-10, 7-12 somites), the signal was almost exclusively detected in the em- bryonic caudal region. These findings confirm and ex- tend our previous report on cCdx-B expression re- stricted on time (Serrano et al., 1993).
Regionalization of the cC&-B Gene Expression In situ hybridization analysis, performed on whole-
mount embryos at stages HH4-17 (E0.5-2.51, localized cCdx-B mRNA expression at the caudal half of the gas- trula, extending rostrally to Hensen’s node early in neurulation and becoming restricted to the tail bud region late in neurulation. During gastrulation, a t stage HH4, (Fig. 3B) cCdx-B mRNA was found in the posterior half of the embryo in an increasing rostrocau- dal gradient. In addition, the highest level was found in the border between the area pellucida and the area opaca, decreasing progressively in the proximity of the primitive streak. Hensen’s node was negative at this stage, whereas, at stage HH6, cCdx-B expression ex- tended to Hensen’s node (Fig. 3C).
During early neurulation, a t stage HH8 (Fig. 3D,G), cCdx-B mRNA was localized at high levels in the pos- terior neural plate and in some mesodermal compo- nents but, apparently, not in the notochord (see below). The cCdx-B transcripts were found in the lateral plate mesoderm, in the intermediate mesoderm, and in the paraxial mesoderm, which becomes subdivided into somites (Fig. 3G). Low levels of cCdx-B mRNA were expressed in the extraembryonic area pellucida (Fig. 3C-E). The localization of this mRNA in the area pel- lucida was restricted at HH8 to a narrow stripe that longitudinally flanked the embryo up to the head fold, regressing caudally later a t HH10. The anterior limit of cCdx-B expression showed a decreasing gradient in the paraxial mesoderm; thus, it was not sharply delim- ited. The border appears to lie in the presomitic region just posterior to the youngest somite (somite 6 in stage HH8+; Fig. 3D,G). In the adjacent neural tube, the anterior border extended rostrally, coinciding with the last one or two somites (Fig. 3D,E,G,H). This was con- firmed by the analysis of sectioned embryos that is de- scribed below.
In embryos at stage HHlO (Fig. 3E,H), the expres- sion of cCdx-B was similar to that described for HH8, with the signal regressing further caudally and a more restricted expression in the extraembryonic area pel-
Fig. 1. A: cDNA sequence of cCdx-6 partial clone and deduced amino acid sequence of the putative protein. The amino acid sequence is numbered. The homeodomain sequence is boxed. The C region corre- sponds to the sequence between arrowheads. The sequences corre- sponding to the upstream, nested, and downstream primers used in the reverse transcription-polymerase chain reaction (RT-PCR) are under- lined. The translational stop codon (TAA) is in boldface. B: Alignment of derived amino acid sequences of the caudal (cad)-related homeoboxes of several species. Residues identical to the cCdx-6 sequence are indi- cated by an asterisk. On the right, percentages of identity at the amino acid level between the cCdx-6 homeobox and the other homeoboxes are shown. Sequence sources: chick cCdx-A, Frumkin et al. (1991); mouse Cdx-7, Duprey et al. (1988); rat Cdx-7, Freund et al. (1992); mouse Cdx-2, James and Kazenwadel (1991); hamster hCdx-3, German et al. (1992); mouse Cdx-4 Gamer and Wright (1993); Xenopus Xcadl and Xcad2, Blumberg et al. (1991); Xenopus Xcad3, Northrop and Kimelman (1994); zebra fish Zf-cad 7 , Joly et al. (1992); Drosophila cad, Mlodzik et al. (1985); Bornbix mori 6m cad, Xu et al. (1994); and C. ekgans ceh-3, Waring and Kenyon (1991).
CAUDAL EXPRESSION IN CHICK EMBRYOS
A 345
CCdx- B C C d x - A cdx- 1 r C d x - 1 cdx- 2 h C d x - 3 cdx- 4 X c a d - 1 X c a d - 2 X c a d - 3 Zf cad1 caudal Bm cad ceh- 3
21
4 1
61
81
101
S S Y B W M R K T V P T N T T G K T R T
WGAGAAGTACCGGGTI'GT" ACACAGACCA TCAGAW CTAGAA'ITAGAGAAGGAATIT K E K Y R V V Y T D H Q R L E L E K E P
C ~ ~ T ~ C ~ C T A H C N R Y I T I R R K S E L A V N L G L
TCTGAAAGACAGGTAAAATCCTGGTTC --cCA?iAGAAA GAAAAATAATC S E R Q V K S W P Q N R R A K E R K I I
A A G A A G A A A T C T C T C A G " G A ~ C T C A G ~ G A G T G A C T ~ ~ C K K K S L S L M A V G A Q F R V T L V H
T C A G C C C T A A T G A A A T A A C A A G T T C C ~ T T C C C A C C A C ~ ~ C ~ T ~ ~ T A T T A C 121 S A L M K
83% 85% 87% 88% 88% 95% 87% 83% 83% 92% 78% 85% 62%
Fig. 1.

MORALES ET AL 346
A B
- 1.5
Fig. 2 Developmental changes of cCdx-6 gene expression during gastrulation, neurulation, and early organogenesis. A: A 230 bp cCdx-6- specific fragment was amplified by RT-PCR (30 cycles) using 1.25 Fg of total RNA from unincubated blastoderm (EO), E0.5, E l , E2, and E3 chick embryos, and the products were analyzed by Southern blot hybridization. The autoradiography was exposed for 72 hrs at -70°C. B: Similar ex- periment with RNA from rostral (R) and caudal (C) regions of E l .5 em- bryos. N, negative control without RNA template.
lucida. In embryos at stage 4417 (30 somites; Fig. 3F), cCdx-B transcripts were even more restricted to the posterior end of the embryo, being localized at the tail bud region, including the neural tube and the meso- derm-generating area. The control sense riboprobe was used with embryos of the same stages described above and produced no signal (Fig. 3A,I and data not shown).
Cellular Localization of cC&-B Transcripts During Embryogenesis
cCdx-B transcripts were found in the epiblast and mesoderm of the gastrula and, later during neurula- tion, in the neuroectoderm, Hensen’s node, and pre- somitic mesoderm, as shown in serial cross sections of the hybridized wholemounts. Embryos at gastrulation (stage HH5; Fig. 4A,B) showed the maximal levels of cCdx-B transcripts in the epiblast at the level of the primitive streak (Fig. 4, diagram), specially in the basal cytosol-rich portion of this cell layer (pseudo- stratified columnar epithelium). The signal was lower in the ridge of the primitive streak, where cells from the epiblast are recruited to become mesodermal cells. The signal became even lower as the mesodermal cells separated from the vicinity of the primitive streak. No signal could be observed in the definitive endoderm. This cell layer specificity in the pattern of expression remained similar in embryos of stage HH7 (Fig. 4C ,D).
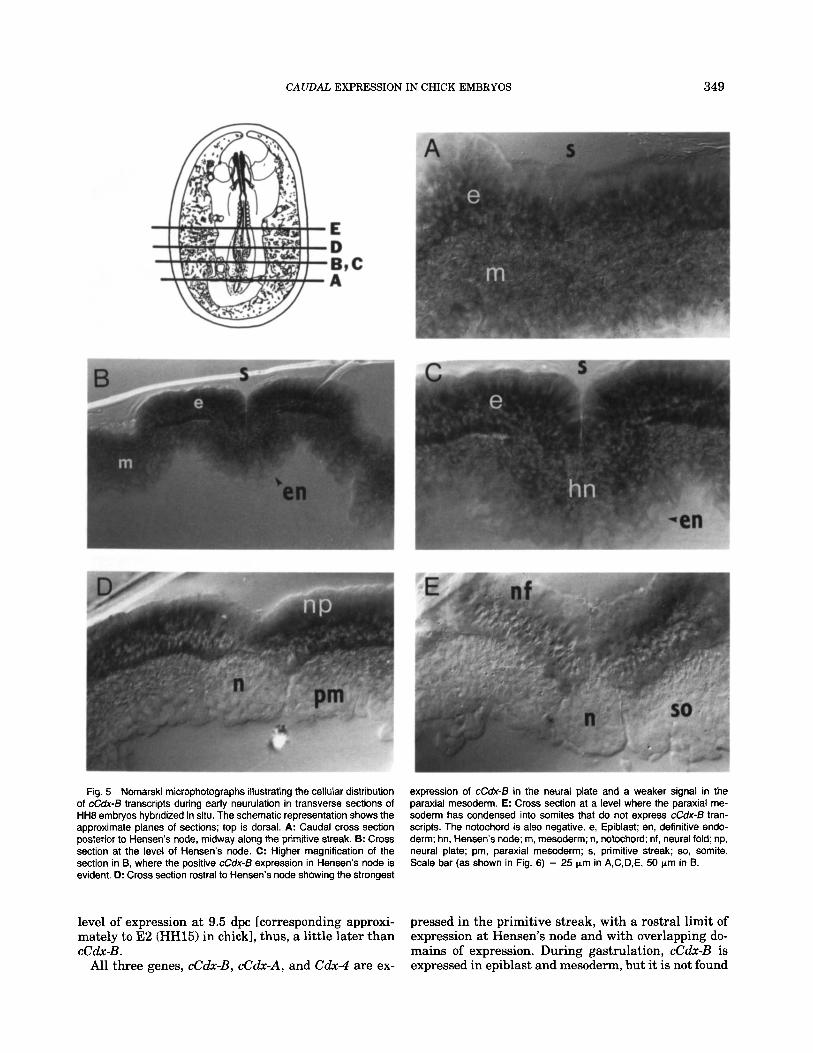
In embryos at neurulation (stages HH8-101, when the highest level of cCdx-B expression was found by RT-PCR, the strongest signal still appeared in the epi- blast a t the level of the primitive streak in regression (Fig. 5A). Lower expression was found in the meso- derm, and no signal was observable in the definitive endoderm (data not shown). At the level of Hensen’s node (Figs. 5B,C, 6A,B), a high level of cCdx-B mRNA
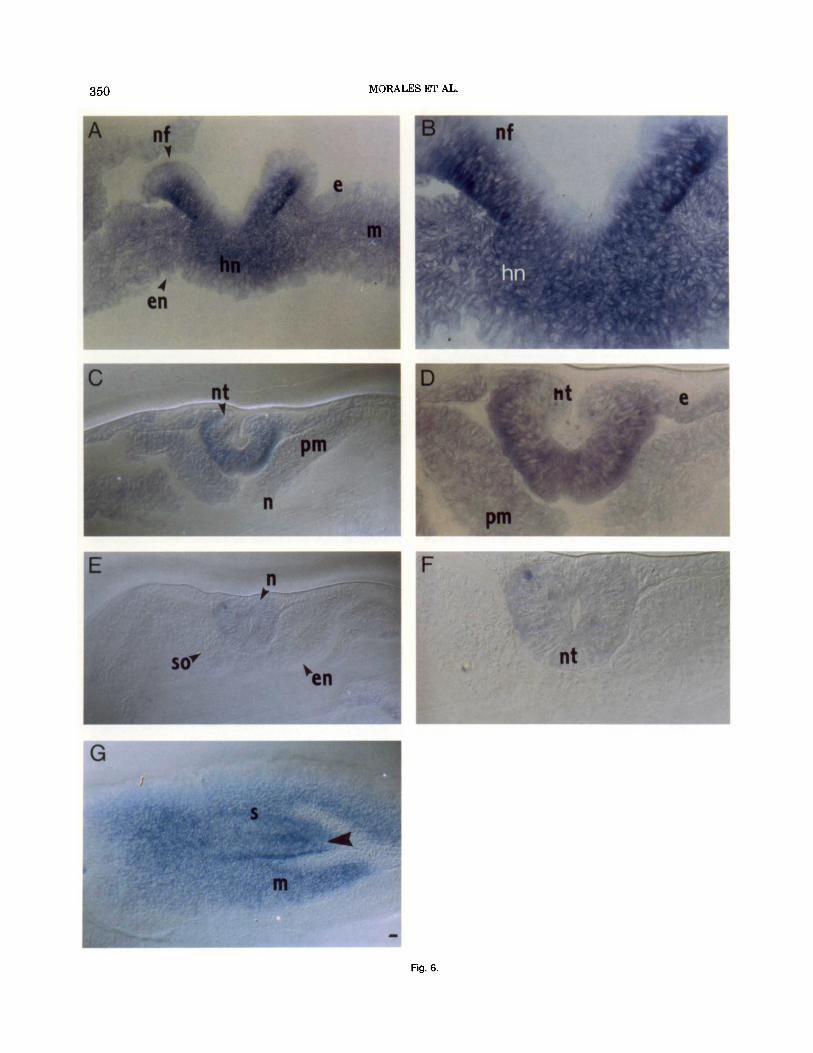
expression was found in this structure as well as in the ectoderm and the mesoderm. Rostra1 to Hensen’s node, a high signal was localized in the neuroectoderm (Figs. 5D,E, 6C,D). The largest amount of cCdx-B transcripts was found lateral to the primitive streak (Fig. 5B,C) and in the neural plate (Figs. 5D, 6D). In both struc- tures, the hybridization signal decorated a band close to the basal membrane and another intermediate, less marked band in more rostral sections (Figs. 5D, 6D). In the neural tube, the transcripts close to the basal mem- brane were more abundant in the ventrolateral zone (where the first motoneurons will arise) and were less abundant in the dorsal zone, showing a gradient pat- tern (Fig. 6D). Expression was lower in the epiblast lateral to the neural plate and in the paraxial meso- derm, and mRNA was not detected in the notochord (Figs. 5D, SC,D). When the axial mesoderm condensed into somites, the expression of cCdx-B disappeared from the mesodermal cells while remaining detectable in the neuroectoderm, although at lower levels (Figs. 5E, 6E,F). The anterior limit of cCdx-B expression in the neural tissue extended to coincide with the last one or two somites.
Embryos at stage HH17 (primary neurulation com- pleted) were sectioned horizontally to analyze cC&-B expression in the tail bud region. The cCdx-B mRNA was present in the mesoderm generating area, in the ridge of the primitive streak, and in a very distal axial line that may relate to secondary neurulation in that area (Holmdahl, 1925; O’Rahilly and Muller, 1994; Fig. 6G). There was no expression of cCdx-B in the groove of the streak or in any anterior structures, such as neural tube, somites, or other rostral structures, a t this time.
DISCUSSION Shaping an embryo requires specification of axes and
regionalization into increasingly smaller morphoge- netic fields, which are the Hox gene major players in this latter process (for reviews, see McGinnis and Krumlauf, 1992; Krumlauf, 1994). The expression pat- terns of the vertebrate cad-related homeobox genes, like cC&-B, presented in this work, Zf-cad 1 (Joly et al., 19921, Cdx-1 (Meyer and Gruss, 19931, Cdx-4 (Gamer and Wright, 1993) and cCdx-A (Frumkin et al., 1991, 1993), strongly suggest the involvement of this family of genes in rostrocaudal axis patterning at crit- ical periods of embryogenesis, namely, gastrulation
Fig. 3 A-I: Developmental pattern of cCdx-6 gene expression by in situ hybridization of chick embryos. In toto in situ hybridization was per- formed on chick embryos at stages 4 (HH4; Hamburger and Hamilton, 1951; A,B), HH6 (C), HH8 (D,G), HHlO (E,H,I), and HH17 (F). A and I show control hybridizations with the sense riboprobe. The embryos are oriented with rostral at the top. G-l show a close-up views of the caudal region of different neurulating chick embryos. Arrowheads point to the rostral limits of expression of cCdx-B gene in B, the intermediate meso- derm in G, and the positive tail bud in F. hn, Hensen’s node; Im, lateral mesoderm; np, neural plate; pm, paraxial mesoderm; so, somite; ys, youngest somite. Scale bar = 40 pm in A,B,G-I, 50 Fm in C, 100 pm in D-F.
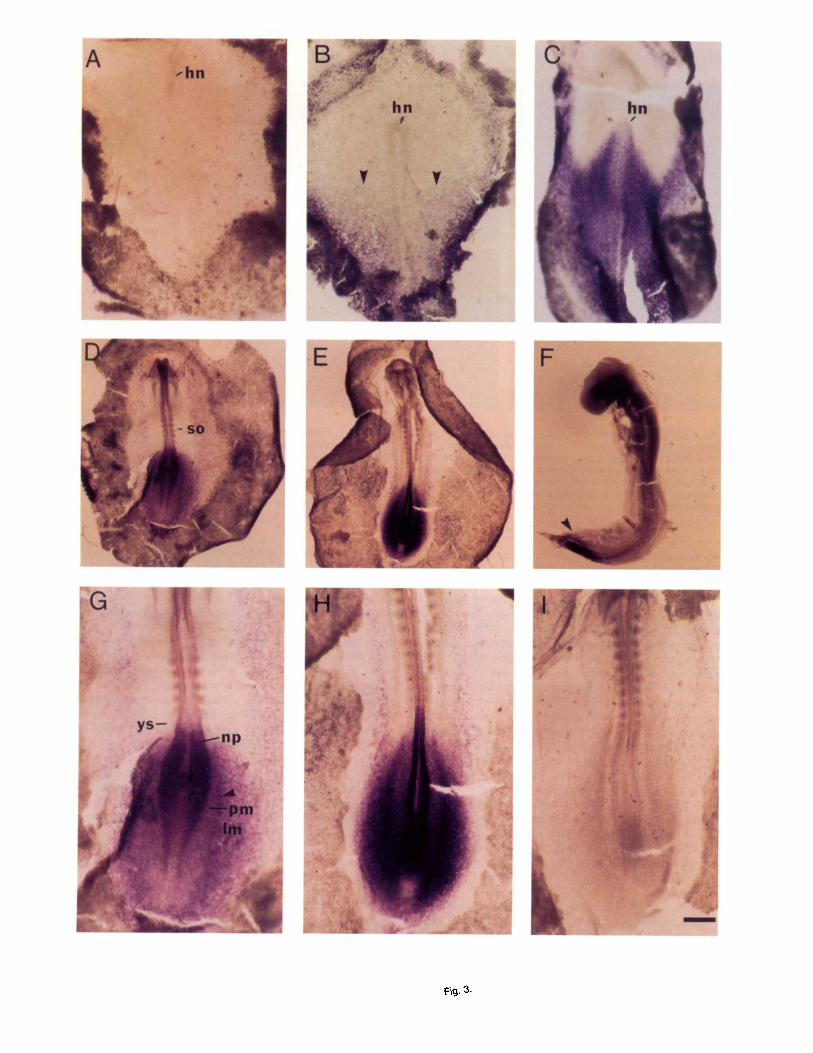
Fig. 3.

348 MORALES ET AL.
Fig. 4 Cellular localization of cCdx-13 in the primitive streak during gastrulation. Transverse sections of in situ hybridized embryos at HH5 (A,B) and HH7 (C,D). A and D are bright field, and Band C are Nomarski pictures. Inset: Schematic representation of an HH5 embryo showing the approximate plane of section corresponding to A and 8. A: The staining is localized mainly in the epiblast and is localized more weakly in the mesoderm. B: A close-up view of the primitive streak shows that highest
expression in the epiblast is localized in the basal layer, and less mRNA expression is found in the mesoderm. C: Control using the sense ribo- probe of cCdx-€3 with an HH7 embryo. D: cCdx-€3 expression in HH7 at the primitive streak level is similar to that described in HH5 (B), being the highest signal in the basal layer of the epiblast. e, Epiblast; en, definitive endoderm; m, mesoderm; s, primitive streak. Scale bar = 100 pm in A, 25 pm in B,C, 50 pm in D.
and neurulation. The transient, restricted expression of cCdx-B along the axis may contribute to modulating the expression of multiple Hox genes.
In Drosophila and other invertebrates as well as in zebrafish, only one cud gene has been found. Con- versely, in Xenopus (Blumberg et al., 1991; Northrop and Kimelman, 1994), chick (Frumkin et al., 1991; Se- rrano et al., 19931, and rodents (Duprey et al., 1988; James and Kazenwadel, 1991; German et al., 1992; Gamer and Wright, 19931, there are multiple members of this gene family with slightly different patterns of expression in each species (see below). This, in turn, suggests that they constitute a system of axial signals with perhaps some overlapping and some complemen- tary functions. Comparing the deduced amino acid se- quences of cCdx-B homeobox with other cad-related genes, the highest identities are observed with Cdx-4 (95%) and Zf-cad 1 (92%). Another conserved region, the so-called C region, which is located upstream of the homeobox (Gamer and Wright, 1993; Fig. lA), also ex- hibits high similarity between these three genes (84% with Cdx-4 and 67% with Zf-cad 1). Such conservation suggests that the corresponding genes might have sim- ilar functions.
Expression of cCdx-B mRNA is detected by RT-PCR in the unincubated embryo (in our conditions, blasto- derm at HH1-2) as early as cCdx-A, the other chick
cud-related gene (Frumkin et al., 1993). In contrast, the murine Cdx-1 and Cdx-4 are found at later stages of gastrulation (Gamer and Wright, 1993; Meyer and Gruss, 1993). However, this could be due to the fact that in situ hybridization is less sensitive than the RT- PCR procedure used for this very early stage in this work. Besides the coincident early onset of transcrip- tion, there are some important differences in the tem- poral expression of the two chick cad-related genes. First, cCdx-B mRNA expression is highest during early neurulation, whereas the maximal cCdx-A expression appears in gastrulation (Frumkin et al., 1991). Second, cCdx-B mRNA is continuously expressed in the rostro- caudal axis, excluding the endoderm, at least until E3, whereas cCdx-A disappears transiently from this axis a t HH10, to reappear again in the epithelial layer of the gut endoderm and, transiently, in the mesenchyme of the intestine (Frumkin et al., 1993, 1994). These facts suggest that cCdx-B may regulate the axial pat- terning longer during development than cCak-A, whereas the latter may have an additional role in the development of the gut, as suggested by Frumkin et al. (1993,1994). The murine Cdx-4, with the highest iden- tity to cCdx-B, is continuously expressed from 7.0 dpc (gastrulation) until 10.5 dpc (late neurulation), resem- bling the temporal pattern of expression of cCdx-B mRNA. However, Cdx-4 mRNA reaches the maximum
CAUDAL EXPRESSION IN CHICK EMBRYOS 349
Fig. 5 Nomarski microphotographs illustrating the cellular distribution of cCdx-B transcripts during early neurulation in transverse sections of HH8 embryos hybridized in situ. The schematic representation shows the approximate planes of sections; top is dorsal. A: Caudal cross section posterior to Hensen's node, midway along the primitive streak. 6: Cross section at the level of Hensen's node. C: Higher magnification of the section in B, where the positive cCdx-B expression in Hensen's node is evident. D: Cross section rostral to Hensen's node showing the strongest
expression of cCdx-B in the neural plate and a weaker signal in the paraxial mesoderm. E: Cross section at a level where the paraxial me- soderm has condensed into somites that do not express cCdx-6 tran- scripts. The notochord is also negative. e, Epiblast; en, definitive endo- derm; hn, Hensen's node; m, mesoderm; n, notochord; nf, neural fold; np, neural plate; pm, paraxial mesoderm; s, primitive streak; so, somite. Scale bar (as shown in Fig. 6) = 25 pm in A,C,D,E, 50 pm in B.
level of expression at 9.5 dpc [corresponding approxi- mately to E2 (HH15) in chick], thus, a little later than cCdx-B .
All three genes, cCdx-B, cCdx-A, and Cdx-4 are ex-
pressed in the primitive streak, with a rostral limit of expression at Hensen's node and with overlapping do- mains of expression. During gastrulation, cCdx-B is expressed in epiblast and mesoderm, but it is not found
350 MORALES ET AL.
Fig. 6.

CAUDAL EXPRESSION IN CHICK EMBRYOS 351
in definitive endoderm, in contrast with cCdx-A, which is expressed in the three germ layers during the elon- gation of the primitive streak (Frumkin et al., 1993). Nevertheless, we have not studied whether cCdx-B is expressed in the hypoblast at stage HH3-4, coinciding with the elongation of the primitive streak. This anal- ysis has not been done for Cdx-4 either. At the time when the head fold of the neuroectoderm appears, cCdx-B expression spreads rostrally to Hensen's node, whereas cCdx-A expression remains caudal to the node, decreasing jointly with the regression of the primitive streak. The anterior border of expression of cCdx-B appears to lie in the presomitic region, just pos- terior to the youngest somite. The same pattern of ex- pression has been described for the murine Cdx-4, with the exception that, during neurulation, Cdx-4 tran- scripts are present at high levels in all three germ lay- ers (Gamer and Wright, 1993), whereas cCdx-B is ex- pressed only in the mesoderm and ectoderm. In fact, we have not observed expression of cCdx-B in the defini- tive endoderm during the first 3 days of chick develop- ment. Zf-cad 1 (Joly et al., 1992), which is expressed in posterior neuroectoderm, is also expressed in hypoblast and endoderm.
The gradient of cCdx-B mRNA moves posteriorly be- tween HH5 and HH17, with the maximal levels always associated with the proliferative posterior region of the embryo. In this area, generating cells receive posi- tional assignments within their respective germ lay- ers. Because cCdx-B encodes a putative transcriptional factor, its function may be related to regulation of the expression of the Hox genes or of the Eux genes (Dush and Martin, 1992). Recently, the Cdx-1 gene knock-out mouse showed an anterior homeotic transformation of vertebrae, possibly related to defects in the establish- ment of the anterior expression boundaries of Hox genes in the somites, such as Hoxc-5, Hoxc-6, Hoxc-8, Hoxa-7, and Hoxd-3. In addition, Cdx-1 seems to regu-
Fig. 6 Nomarski microphotographs illustrating the spatial distribution of cCdx-B transcripts during late neurulation. Transverse sections after in situ hybridization of cCdx-B on embryos at stage HHlO (A-F) and hori- zontal section of an embryo at HH17 (G). From A to F, the sections are arranged from caudal to rostral in a distribution similar to that shown in Figure 5; top is dorsal. A: Section at the level of the rostral tip of Hensen's node, where the neural folds can be observed. B: A higher magnification of the section in A. Strong staining can be observed in the cytoplasm of the cells in Hensen's node and in the neural folds. C: Cross section showing the neural tube almost closed and the paraxial mesoderm un- condensed. D: A higher magnification of the section in C showing a strong expression of cCdx-B in the neural tube and a weaker signal in the epiblast and the paraxial mesoderm. E: Cross section showing that the axial mesoderm condenses to form the somites and that cCdx-B mRNA expression in the mesodermal cells disappears. F: A higher magnification of the section in E showing only a few cells in the closed neural tube weakly expressing cCdx-B G: Caudal horizontal section of a HHI 7 em- bryo. Rostra1 is to the right. The cCdx-B mRNA appears in the mesoderm- generating area and in the primitive streak. The arrowhead marks an axial line that may relate to secondary neurulation. e, Epiblast; en, de- finitive endoderm; hn, Hensen's node; m, mesoderm; n, notochord; nf, neural fold; nt, neural tube; pm, paraxial mesoderm; s, primitive streak; so, somite. Scale bar = 50 pm in A,C,E,G, 25 pm in B,D,F.
late directly the Hoxa-7 promoter, as shown by in vitro transactivation (Subramanian et al., 1995). Similarly, in Drosophila, cad is involved in the regulation of de- formed, a gene of the HOM-C Complex (Mlodzick et al., 19901, as well as the pair-rule gene fushi tarmu (Dearolf et al., 1989) and abdominal gap genes such as knirps and giant (Schulz and Tautz, 1995). Drosophila embryos lacking the characteristic maternal cad mRNA gradient have defective abdominal boundaries, whereas embryos lacking zygotic cad expression have abnormal posterior terminalia. These findings reveal at least two functional periods for cad in Drosophila development (Macdonald and Struhl, 1986; Mlodzik and Gehring, 1987a,b). Late functions have also been suggested for the vertebrate cad-related gene family. Zf-cad 1 becomes restricted mostly to the posterior neu- ral tissue and cCdx-A becomes restricted mostly to the developing gut (Frumkin et al., 1991, 1994; Joly et al., 1992). Cdx-1 expression late in embryogenesis becomes restricted to the epithelial lining of the intestine at 14 dpc (Duprey et al., 1988). Although we cannot exclude another late period of expression for cCdx-B, this mRNA seems to be present only during gastrulation and neurulation stages, similar to the mouse Cdx-4 (Gamer and Wright, 1993). At these stages, cCdx-B should be contributing to the combinatorial signalling code that determines rostrocaudal polarity and, in par- ticular, to the regional specification of the tail region, including the spinal cord. Additional experiments are needed to unravel the relative contribution of cCdx-A and -B to the patterning of the chick embryo.
EXPERIMENTAL PROCEDURES Chick Embryos
Fertilized White Leghorn eggs (Avicola Grau, Ma- drid, Spain) were incubated at 38.2"C with 60-90% rel- ative humidity. The embryos were staged according to Hamburger and Hamilton (1951), referring also to the embryonic age (E) and, occasionally, to the number of somites (to allow for comparison with mouse embryos).
cDNA, Oligonucleotides, and Riboprobes The cloning of the chick cCdx-B cDNA (previously
cHox-cad 2) has been reported previously (Serrano et al., 1993). Oligonucleotides of 20 bases were used as primers in the RT-PCR (Fig. 1A). cCdx-B upstream (5 GTTTACACAGACCATCAGAG3') and downstream (5'AGGGCTGAGTGAACCAGAGT3') primers ampli- fied a region of 230 bp, and an internal oligonucleotide (5"M!CTTTGGCTCGGCGATTCT3') was used as a probe for blot hybridization.
To generate RNA probes for in situ hybridization, the 1,082 bp insert of pcHox-cad2/cCdx-B (Serrano et al., 1993) was subcloned into the Eco RI site of pBlue- script KS (+/-; Promega). The sense and antisense riboprobes were synthesized by incorporation of digox- igenin-UTP (Boehringer Mannheim), according to Wil- kinson and Nieto (1993).

352 MORALES ET AL.
RT-PCR and DNA Blot Hybridization The embryos were separated from the extraembry-
onic membranes (completely only after E1.5; some membranes remained in younger embryos), sectioned when indicated, rinsed in cold phosphate-buffered sa- line (PBS), frozen in dry ice, and lyophilized and stored at -70°C. Total RNA was extracted from whole or parts of the embryos by using the RNAzol method (CinnalBiotecx), and 1.25 pg was used for cDNA syn- thesis. Reverse transcription to obtain a cDNA and PCR for 30 cycles were carried out as described previ- ously (Perez-Villamil et al., 1994). The PCR-amplified products were gel fractionated, transferred to a nylon membrane, hybridized with 5'-end [32P]-labeled spe- cific internal oligonucleotide, and exposed to x-ray films as described (Perez-Villamil et al., 1994).
In Situ Hybridization Embryos of the appropriate stage were dissected as
indicated above, fixed in 4% formaldehyde in PBS:, and immediately processed for hybridization. The whole- mount in situ hybridization protocol for embryos with digoxigenin-labeled probes was adapted from Wilkin- son and Nieto (1993). Briefly, prior to hybridization, embryos were permeated with 0.1% Tween 20 in PBS (PBT), dehydrated, and rehydrated. Then, they were bleached with 1% hydrogen peroxide, treated with 2.5 pg/ml proteinase K, and refixed. Prehybridization. was carried out in 1 ml of 50% formamide, 5 x SSC, pH 5, 1% SDS, 250 pg/ml yeast tRNA, and 50 pg/ml hepa- rin a t 70°C overnight. Hybridization was done in the same prehybridization mix, adding the digoxigenin-la- beled probe (1 kg/ml). Before the incubation with the antidigoxigenin alkaline phosphatase-conjugated anti- body (Boehringer Mannheim; 1:2,000 dilution), non- specific binding was blocked with 10% lamb serum, and the antibody was preabsorbed with chick embryo pow- der. After incubation with the antibody overnight, the embryos were developed with nitro-blue tetrazolium salt (NBT) and 5-bromo-4-chloro-3-indolyl-phosphate (BCIP). The embryos were cleared in 50% glycerol and photographed using a Wilt-Leitz M-400 photomacro- scope.
Some of the embryos were then refixed, dehydrated with ethanol, and embedded in Historesin (Jung, Her- aeus Kulzer GmbH). Sections (25 pm thick) were placed on slides, mounted with Depex (Serva), and pho- tographed using a Zeiss Axiophot microscope with ei- ther transmitted light or Nomarski optics.
ACKNOWLEDGMENTS We thank A. Nieto for very useful advice with the in
situ hybridization protocol, C. Risueiio for help with sectioning techniques, J. Serrano for sequencing, and A. Pena for helpful discussions. We are grateful to A. Fairen for the use of the Zeiss microscopes and to A. Fainsod, L. Puelles, and S. Martinez for comments on the paper.
REFERENCES Blumberg, B., Wright, C.V., De Robertis, E.M., and Cho, K.W. (1991)
Organizer-specific homeobox genes in Xenopus laeuis embryos. Sci- ence 253:194-196.
Cohen, S.M., and Jurgens, G. (1990) Mediation of Drosophila head development by gap-like segmentation genes. Nature 346:482-485.
Dalton, D., Chadwick, R., and McGinnis, W. (1989) Expression and embryonic function of empty spiracles: A Drosophila homeobox gene with two patternings functions on the anterior-posterior axis of the embryo. Genes Dev. 3:1940-1956.
Dearolf, C.R., Topol, J., and Parker, C.S. (1989) The caudal gene prod- uct is a direct activator of fushi tarazu transcription during Dro- sophila embryogenesis. Nature 341340-343.
Doll, U., and Niessing, J. (1993) Continued expression of the chick caudal homologue in endodermally derived organs. Dev. Biol. 156: 155-163.
Duprey, P., Chowdhury, K., Dressler, G.R., Balling, R., Simon, D., Guenet, J.L., and Gruss, P. (1988) A mouse gene homologous to the Drosophila gene caudal is expressed in epithelial cells from the embryonic intestine. Genes Dev. 2:1647-1654.
Dush, M.K., and Martin, G.R. (1992) Analysis of mouse Evx genes: Evx-1 displays graded expression in the primitive streak. Dev. Biol. 151:273-287.
Freund, J.N., Boukamel, R., and Benazzouz, A. (1992) Gradient ex- pression of Cdx along the rat intestine throughout postnatal devel- opment. FEBS Lett. 314:163-166.
Frumkin, A,, Rangini, Z., Ben Yehuda, A,, Gruenbaum, Y., and Fain- sod, A. (1991) A chick caudal homologue, CHox-cad, is expressed in the epiblast with posterior localization and in the early endodermal lineage. Development 11 2:207-2 19.
Frumkin, A,, Haffner, R., Shapira, E., Tarcic, N., Gruenbaum, Y., and Fainsod, A. (1993) The chick CdxA homeobox gene and axial posi- tioning during gastrulation. Development 118553-562.
Frumkin, A., Pillemer, G., Haffner, R., Tarcic, N., Gruenbaum, Y., and Fainsod, A. (1994) A role for CdxA in gut closure and intestinal epithelia differentiation. Development 120:253-263.
Gamer, L.W., and Wright, C.V. (1993) Murine Cdx-4 bears striking similarities to the Drosophila caudal gene in its homeodomain se- quence and early expression pattern. Mech. Dev. 43:71-81.
German, M.S., Wang, J., Chadwick, R.B., and Rutter, W.J. (1992) Synergistic activation of the insulin gene by a LIM-homeo domain protein and a basic helix-loop-helix protein: Building a functional insulin minienhancer complex. Genes Dev. 6:2165-2176.
Hamburger, V., and Hamilton, H.L. (1951) A series of normal stages in the development of the chick embryo. J. Morphol. 88:49-92.
Holmdahl, D.E. (1925) Experimentelle Untersuchungen uber die Lage der Grenze zwischen primarer und sekundarer Korper- entwicklung beim Huhn. Anat. Am. 59:393-396.
James, R., and Kazenwadel, J . (1991) Homeobox gene expression in the intestinal epithelium of adult mice. J . Biol. Chem. 266:3246- 3251.
Joly, J.S., Maury, M., Joly, C., Duprey, P., Boulekbache, H., and Con- damine, H. (1992) Expression of a zebrafish caudal homeobox gene correlates with the establishment of posterior cell lineages at gas- trulation. Differentiation 50:75-87.
Krumlauf, R. (1994) Hox genes in vertebrate development. Cell 78: 191-201.
Macdonald, P.M., and Struhl, G. (1986) A molecular gradient in early Drosophila embryos and its role in specifying the body pattern. Nature 324537-545.
McGinnis, W., and Krumlauf, R. (1992) Homeobox genes and axial patterning. Cell 68:283-302.
Meyer, B.I., and Gruss, P. (1993) Mouse cdx-1 expression during gas- trulation. Development 117:191-203.
Mlodzic, M., and Gehring, W.J. (1987a) Expression of the caudal gene in the germ line of Drosophila: Formation of an RNA and protein gradient during early embryogenesis. Cell 48:465-478.
Mlodzic, M., and Gehring, W.J. (1987b) Hierarchy of the genetic in- teractions that specify the anteroposterior segmentation pattern of the Drosophila embryo as monitored by caudal protein expression. Development 101:421-435.

CAUDAL EXPRESSION IN CHICK EMBRYOS 353
Mlodzic, M., Fjose, A., and Gehring, W.J. (1985) Isolation of caudal, a Drosophila homeo box-containing gene with maternal expressin, whose transcripta form a concentration gradient at the pre-blasto- derm stage. EMBO J. 42961-2969.
Mlodzik, M., Gibson, G., and Gehring, W.J. (1990) Effects of ectopic expression of caudal during Drosophila development. Development
Northrop, J.L., and Kimelman, D. (1994) Dorsal-ventral differences in Xcad-3 expression in response to FGF-mediated induction in Xeno- pus. Dev. Biol. 161:490-503.
O’Rahilly, R., and Muller, F. (1994) Neurulation in the normal human embryo. Ciba Found. Symp. 181:70-82.
P6rez-Villami1, B., De la Rosa, E.J., Morales, A.V., and De Pablo, F. (1994) Developmentally regulated expression of the preproinsulin gene in the chick embryo during gastrulation and neurulation. En- docrinology 1352342-2350.
Rubenstein, J.L., Martinez, S., Shimamura, K., and Puelles, L. (1994) The embryonic vertebrate forebrain: The prosomeric model. Science
Schulz, C., and Tautz, D. (1995) Zygotic caudal regulation by hunch- back and its role in abdominal segment formation of the Drosophila embryo. Development 121:1023-1028.
109271-277.
266~578-580.
Serrano, J., Seavo, L., Roth, J., De la Rosa, E.J., and De Pablo, F. (1993) A novel chick homeobox-containing gene expressed in neu- rulating embryos. Biochem. Biophys. Res. Commun. 190:270-276.
Simeone, A., Acampora, D., Gulisano, M., Stornaiuolo, A., and Bon- cinelli, E. (1992) Nested expression domains of four homeobox genes in developing rostra1 brain. Nature 358:687-690.
Subramanian, V., Meyer, B. I., and Gruss, P. (1995) Disruption of the murine homeobox gene Cdz-1 affects axial skeletal identities by altering the mesodermal expression domains of Hox genes. Cell
Waring, D.A., and Kenyon, C. (1991) Regulation of cellular respon- siveness to inductive signals in the developing C. eZegans nervous system. Nature 350:712-715.
Wilkinson, D.G., and Nieto, M.A. (1993) Detection of messenger RNA by in situ hybridization to tissue sections and whole mounts. Meth- ods Enzymol. 225361-373.
Xu, X., Xu, P.X., and Suzuki, Y. (1994) A maternal homeobox gene, Bombyx caudal, forms both mRNA and protein concentration gra- dients spanning anteroposterior axis during gastrulation. Develop- ment 120:277-285.
83~641-653.
Related Documents











