ORIGINAL PAPER S. Gutie´rrez Æ J. Velasco Æ A. T. Marcos F. J. Ferna´ndez Æ F. Fierro Æ J. L. Barredo B. Dı´ez Æ J. F. Martı´n Expression of the cefG gene is limiting for cephalosporin biosynthesis in Acremonium chrysogenum Received: 16 May 1997 / Received revision: 3 July 1997 / Accepted: 12 July 1997 Abstract The conversion of deacetylcephalosporin C to cephalosporin C is inecient in most Acremonium chrysogenum strains. The cefG gene, which encodes deacetylcephalosporin C acetyltransferase, is expressed very poorly in A. chrysogenum as compared to other genes of the cephalosporin pathway. Introduction of additional copies of the cefG gene with its native pro- moter (in two dierent constructions with upstream regions of 1056 bp and 538 bp respectively) did not produce a significant increase of the steady-state level of the cefG transcript. Expression of the cefG gene from the promoters of (i) the glyceraldehyde-3-phosphate dehy- drogenase (gpd ) gene of Aspergillus nidulans, (ii) the glucoamylase (gla) gene of Aspergillus niger, (iii) the glutamate dehydrogenase (gdh) and (iv) the isopenicillin N synthase ( pcbC ) genes of Penicillium chrysogenum, led to very high steady-state levels of cefG transcript and to increased deacetylcephalosporin-C acetyltransferase protein concentration (as shown by immunoblotting) and enzyme activity in the transformants. Southern analysis showed that integration of the new construc- tions occurred at sites dierent from that of the endog- enous cefG gene. Cephalosporin production was increased two- to threefold in A. chrysogenum C10 transformed with constructions in which the cefG gene was expressed from the gdh or gpd promoters as a result of a more ecient acetylation of deacetylcephalosporin C. Introduction The last step of the biosynthesis of cephalosporin C (CPC) by Acremonium chrysogenum (syn. Cephalospor- ium acremonium) involves the acetylation of deacetyl- cephalosporin C (DAC) to CPC (Fig. 1) (Nu¨esch et al. 1987; Martı´n et al. 1994). It is well known that high levels of DAC are accumulated in many cephalosporin- producing strains (Fujisawa et al. 1973) although the origin of the accumulated DAC is an open question. The conversion of DAC into CPC is carried out by the en- zyme acetyl-CoA:DAC acetyltransferase (DAC-AT) that has been identified and partially purified from A. chrysogenum cell-free extracts (Fujisawa and Kanzaki 1975; Scheidegger et al. 1985). The gene (cefG) encoding this enzyme has been cloned in three dierent labora- tories (Gutie´ rrez et al. 1992; Matsuda et al. 1992; Mathison et al. 1993). The cefG gene is linked to cefEF but is transcribed in the opposite orientation as a 1.4-kb monocistronic mRNA (Gutie´rrez et al. 1992). Expres- sion of the CPC biosynthetic genes is growth-phase-de- pendent and reaches its maximal level when the culture has entered the production phase. Recent studies using three dierent strains of A. chrysogenum, ATCC 11550 Brotzu (wild type), CW19 and C10, showed that the cefG gene is transcribed poorly in the three strains early and late during the fermentation, as compared to three other genes ( pcbAB, pcbC, cefEF ) of the cephalosporin pathway (Velasco et al. 1994). Very little is known about the structure and sequence motifs of fungal promoters (Ballance 1986; MacKenzie et al. 1993; Dhawale and Lane 1993). Upstream acti- vating sequences occur in some highly expressed fungal genes (Punt et al. 1992). It is unknown whether the cefG promoter lacks specific enhancers or regulatory se- quences that may explain the low steady-state levels of cefG transcripts in the cells. Modification of the up- stream region or expression from strong fungal pro- moters may lead to overexpression of the gene and, therefore, to an increase in CPC production. It was, Appl Microbiol Biotechnol (1997) 48: 606–614 Ó Springer-Verlag 1997 S. Gutie´rrez Æ J. Velasco Æ A. T. Marcos F. J. Ferna´ndez Æ F. Fierro Æ J. F. Martı´n (&) Area of Microbiology, Department of Ecology Genetics and Microbiology, Faculty of Biology, University of Leo´n, 24071 Leo´n, and Institute of Biotechnology, INBIOTEC, Avda. del Real 1, E-24006 Leo´n, Spain Tel.: +34 87 291505 Fax: +34 87 291506 e-mail: [email protected] J. L. Barredo Æ B. Dı´ez Laboratorio de Ingenierı´a Gene´tica, Antibio´ticos S.A., 24080 Leo´n, Spain

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL PAPER
S. Gutie rrez á J. Velasco á A. T. MarcosF. J. Ferna ndez á F. Fierro á J. L. BarredoB. Dõ ez á J. F. Martõ n
Expression of the cefG gene is limiting for cephalosporin biosynthesisin Acremonium chrysogenum
Received: 16 May 1997 / Received revision: 3 July 1997 /Accepted: 12 July 1997
Abstract The conversion of deacetylcephalosporin C tocephalosporin C is ine�cient in most Acremoniumchrysogenum strains. The cefG gene, which encodesdeacetylcephalosporin C acetyltransferase, is expressedvery poorly in A. chrysogenum as compared to othergenes of the cephalosporin pathway. Introduction ofadditional copies of the cefG gene with its native pro-moter (in two di�erent constructions with upstreamregions of 1056 bp and 538 bp respectively) did notproduce a signi®cant increase of the steady-state level ofthe cefG transcript. Expression of the cefG gene from thepromoters of (i) the glyceraldehyde-3-phosphate dehy-drogenase (gpd ) gene of Aspergillus nidulans, (ii) theglucoamylase (gla) gene of Aspergillus niger, (iii) theglutamate dehydrogenase (gdh) and (iv) the isopenicillinN synthase ( pcbC ) genes of Penicillium chrysogenum,led to very high steady-state levels of cefG transcript andto increased deacetylcephalosporin-C acetyltransferaseprotein concentration (as shown by immunoblotting)and enzyme activity in the transformants. Southernanalysis showed that integration of the new construc-tions occurred at sites di�erent from that of the endog-enous cefG gene. Cephalosporin production wasincreased two- to threefold in A. chrysogenum C10transformed with constructions in which the cefG genewas expressed from the gdh or gpd promoters as a resultof a more e�cient acetylation of deacetylcephalosporinC.
Introduction
The last step of the biosynthesis of cephalosporin C(CPC) by Acremonium chrysogenum (syn. Cephalospor-ium acremonium) involves the acetylation of deacetyl-cephalosporin C (DAC) to CPC (Fig. 1) (NuÈ esch et al.1987; Martõ n et al. 1994). It is well known that highlevels of DAC are accumulated in many cephalosporin-producing strains (Fujisawa et al. 1973) although theorigin of the accumulated DAC is an open question. Theconversion of DAC into CPC is carried out by the en-zyme acetyl-CoA:DAC acetyltransferase (DAC-AT)that has been identi®ed and partially puri®ed from A.chrysogenum cell-free extracts (Fujisawa and Kanzaki1975; Scheidegger et al. 1985). The gene (cefG) encodingthis enzyme has been cloned in three di�erent labora-tories (Gutie rrez et al. 1992; Matsuda et al. 1992;Mathison et al. 1993). The cefG gene is linked to cefEFbut is transcribed in the opposite orientation as a 1.4-kbmonocistronic mRNA (Gutie rrez et al. 1992). Expres-sion of the CPC biosynthetic genes is growth-phase-de-pendent and reaches its maximal level when the culturehas entered the production phase. Recent studies usingthree di�erent strains of A. chrysogenum, ATCC 11550Brotzu (wild type), CW19 and C10, showed that thecefG gene is transcribed poorly in the three strains earlyand late during the fermentation, as compared to threeother genes ( pcbAB, pcbC, cefEF ) of the cephalosporinpathway (Velasco et al. 1994).
Very little is known about the structure and sequencemotifs of fungal promoters (Ballance 1986; MacKenzieet al. 1993; Dhawale and Lane 1993). Upstream acti-vating sequences occur in some highly expressed fungalgenes (Punt et al. 1992). It is unknown whether the cefGpromoter lacks speci®c enhancers or regulatory se-quences that may explain the low steady-state levels ofcefG transcripts in the cells. Modi®cation of the up-stream region or expression from strong fungal pro-moters may lead to overexpression of the gene and,therefore, to an increase in CPC production. It was,
Appl Microbiol Biotechnol (1997) 48: 606±614 Ó Springer-Verlag 1997
S. Gutie rrez á J. Velasco á A. T. MarcosF. J. Ferna ndez á F. Fierro á J. F. Martõ n (&)Area of Microbiology, Department of Ecology Geneticsand Microbiology, Faculty of Biology, University of Leo n,24071 Leo n, and Institute of Biotechnology, INBIOTEC,Avda. del Real 1, E-24006 Leo n, SpainTel.: +34 87 291505Fax: +34 87 291506e-mail: [email protected]
J. L. Barredo á B. Dõ ezLaboratorio de Ingenierõ a Gene tica, Antibio ticos S.A.,24080 Leo n, Spain

therefore, of interest to amplify the cefG gene with itsown promoter and to express it from other transcrip-tion-initiating sequences that are known to be e�cientlyused in ®lamentous fungi.
As reported in this article, expression of the nativecefG from other promoters with strong transcription-initiating ability leads to a large accumulation of thecefG transcript and also to high levels of DAC-ATprotein and increased CPC production.
Materials and methods
Microbial strains
A. chrysogenum (Cephalosporium acremonium) ATCC 11550 (wildtype, original isolate of Brotzu) and A. chrysogenum C10 (ATCC48272) an improved CPC producer released by PanLabs Labora-tories (Demain 1983) were used as host strains for expressionstudies. A. chrysogenum ATCC 20371 (a strain partially de®cient inDAC-AT that accumulates DAC) (Fujisawa et al. 1975) was usedfor complementation experiments. Escherichia coli ESS2231, a b-lactam-supersensitive strain, was used for routine bioassays ofCPC. E. coli DH5a and E. coli WK6 were used for plasmid puri-®cation (double-stranded or single-stranded DNA respectively).
Media and growth conditions
For transcription analysis, CPC production, DAC-AT activity andimmunoblotting studies all A. chrysogenum strains were grown inseed medium 1 (Shen et al. 1986) for 48 h. Samples of 5 ml seedcultures were used to inoculate 100 ml de®ned production (DP1)medium (Shen et al. 1986) in triple-ba�ed 500-ml conical ¯asks(Bellco). Samples were taken every 24 h and b-lactam antibioticswere determined by bioassay with E. coli ESS2231 as the test strainin plates with or without penicillinase (from Bacillus cereusUL1), asdescribed previously (Zanca andMartõ n 1983; Gutie rrez et al. 1992).
Quanti®cation of DAC and cephalosporin C by HPLC
Samples (100 ll) of the culture broths were taken and the proteinsprecipitated by addition of 100 ll methanol and centrifuged at10 000 g for 5 min. The supernatant was diluted tenfold and used(50-ll samples) for CPC analysis in a Beckman System Gold high-pressure liquid chromatograph (HPLC) equipped with a lBonda-pack C18 column (300 ´ 4 mm). DAC and CPC were eluted with amixture of solvents A (10 mM acetic acid/sodium acetate, pH 4.5)and B (100% acetonitrile) using a 0%±3% gradient of solvent B(0% at 0 min, 3% at 2 min and 3% between 2 min and 15 min)with a constant ¯ow of 1.3 ml/min. Under these conditions DACand CPC eluted with retention times of 3.1 min and 8.3 min res-pectively. Both compounds were identi®ed by coelution in theHPLC with authentic samples of CPC and DAC (provided by F.Salto, Antibio ticos, S.A., Leo n, Spain).
Construction of pULcefGL and pULcefGS
A 1472-bp fragment (cassette) containing the phleomycin-resis-tance gene of Streptoalloteichus hindustanus (ble), expressed fromthe Penicillium chrysogenum pcbC promoter and cyc1 terminator,was inserted (with appropiate end-®llings) into EcoRV-digestedpBC KS+, originating plasmid pULC43. This plasmid was di-gested with SalI and linked to a 2.7-kb SalI DNA fragment car-rying the A. chrysogenum cefG gene and the 5¢ upstream region.The resulting plasmid (7.5 kb) was named pULcefGL. pULcefGLwas partially digested with HindIII and religated to producepULcefGS (7.0 kb) (Fig. 1).
Coupling of the di�erent promoters to the cefG gene
pULpcbC
The promoter of the pcbC gene of P. chrysogenum was isolated as a761-bp NcoI-EcoRI fragment and coupled (after appropiate end-®lling) to the 1715-bp BglI-SalI fragment carrying the cefG gene ofA. chrysogenum with its own transcription terminator. The con-struction (as a 1814-bp PstI fragment) was inserted into the PstIsite of pULPAB43 (S. Gutie rrez, unpublished), which carries theble gene (expressed from the pcbAB promoter of P. chrysogenumand followed by the cyc1 terminator) as a selective marker. Theresulting plasmid, pULpcbC, has a size of 6.7 kb.
pULgla
The promoter of the gla gene of Aspergillus niger was isolated fromplasmid pAN52.7 (Punt et al. 1987) as a 650-bp BamHI-NcoIfragment and linked (after appropiate end-modi®cation) with a1716-bp fragment of pULGC1 (Gutie rrez S, unpublished), whichcarried the cefG gene, and coupled to a 1472-bp ``cassette'' carryingthe ble gene and cyc1 terminator (under the control of theP. chrysogenum pcbC promoter). The resulting plasmid (6.85 kb)was named pULgla.
pULgpdL
A 3100-bp NotI-BamHI fragment containing the promoter regionof the A. nidulans gpd gene from plasmids pAN52.1 (Punt et al.1987) was linked to a 1700-bp fragment of pULB7.2 (Gutie rrezet al. 1992) containing the cefG gene and to the 1472-bp ble ``cas-sette'', as indicated above. The resulting plasmid (9.8 kb) wasnamed pULgpdL (for the long upstream region).
pULgpdS
A shorter promoter region (700 bp SacI-NcoI) of the A. nidulansgpd gene was isolated from pAN52.1 and coupled as above to thecefG gene of A. chrysogenum.
pALgdh
A 0.8-kb PstI-BamHI fragment containing the P. chrysogenum gdhpromoter was fused in pBluescript II SK(+) to a 1.6-kb BamHI-XhoI fragment carrying the cefG gene from pALC8 to produceplasmid pALC9. The product of gdh promoter fusion to cefG wassubcloned as a 2.4-kb fragment from pALC9 and linked (after end-®lling) to pAL¯eo7 containing the ble gene (also under the gdhpromoter) and the CmR gene as markers, creating pALgdh. In thisconstruction the DAC-AT protein starts at Met-46 of the nativeprotein (44.4 kDa).
Isolation of RNA and Northern hybridization
Total RNA was extracted by the sodium dodecyl sulphate (SDS)/phenol method (Ausubel et al. 1987). RNA blotting and hybrid-izations were carried out as described previously (Gutie rrez et al.1991).
DAC acetyltransferase assays
DAC acetyltransferase activity was measured by monitoring thein vitro conversion of DAC and acetyl-coenzyme A into CPC byhigh-pressure liquid chromatography as described by Gutie rrezet al. (1992). Total protein in the cell extract was determined by theBradford method with ovalbumin as a standard.
607

Transformation of A. chrysogenum
Transformation of A. chrysogenum protoplasts was carried out asdescribed previously (Gutie rrez et al. 1991). Transformants wereselected in tryptic soy agar TSA (Difco) with sucrose (10.3%)supplemented with 10 lg/ml phleomycin.
Polyclonal antibodies against DAC-AT
DAC-AT protein, obtained by expression of the cefG gene in E. coliJM109(DE3) (Tabor and Richardson 1985) was puri®ed from in-clusion bodies by electroelution of the corresponding band from aSDS/polyacrylamide gel (SDS-PAGE; J. Velasco, S. Gutie rrez andJ. F. Martõ n, unpublished results). Puri®ed DAC-AT protein(100 lg in 500 ll phosphate-bu�ered saline), was emulsi®ed with500 ll complete Freund's adjuvant (Difco Laboratories Inc., De-troit, Mich., USA). This emulsion was used to immunize NewZealand rabbits by intradermal injection, using the protocol de-scribed by Dunbar and Schwoebel (1990). This immunizationprocess was repeated once a month for 3 months using incompleteFreund's adjuvant. After the immunization was completed, bloodserum was collected by centrifugation, and immunoglobulin G waspuri®ed by ammonium sulphate precipitation and fast protein liq-uid a�nity chromatography using a protein-A±Sepharose column(Pharmacia Biotech Inc.) as described by Harlow and Lane (1988).
Immunoblotting procedure
After SDS-PAGE (40 mg protein in each well), the proteins weretransferred to a polyvinylidene ¯uoride membrane (Immobilon-P,Millipore) using a Minitransblot electroblotting system (Bio Rad).The membranes were treated with the anti-DAC-AT antibodies(sera dilution 1/5000) in phosphate-bu�ered saline containing 1%bovine serum albumin and 0.05% Triton X-100 and incubated for90 min at room temperature. The membranes were then washedwith 1 M NaCl and treated with a commercial conjugate of mouseanti-IgG with alkaline phosphatase in the same bu�er for 30 min.The immunoreactive bands were then developed with a commercialstabilized substrate solution for alkaline phosphatase (Promega)for 10 min.
Results
E�ect of the cefG upstream region on gene expression
The initial construction pULB7.2 carried the entirecefEF and cefG genes in a 7.2-kb BamHI fragment.These two genes are transcribed in opposite orientations(Fig. 1) from a bidirectional promoter located in the930-bp intergenic region. Since an interaction of RNApolymerase molecules and transcription-initiation com-plexes of the two genes (including putative regulatoryproteins) might result in a poor expression of the cefGgene, two new constructions were made for comparativestudies: (i) pULcefGL in which the cefG gene was pre-ceded by a long upstream region (1056 bp) containingthe complete intergenic region, and (ii) pULcefGS con-taining a short upstream region (a 538-bp fragment) thatincludes the cefG promoter region but not the upstreamregion adjacent to the cefEF gene (Fig. 1). These twoconstructions were introduced into A. chrysogenumATCC 20371 and ten transformants (®ve with eachconstruction) were selected with high phleomycin-resis-tance levels. All the transformants showed an averagetwofold increase in the basal DAC-AT level of this strain
(from an average 17.0 units/mg protein in the untrans-formed strain to 32.7 units/mg protein and 38.9 units/mg protein in transformants with the short and longpromoter regions respectively) but the di�erences be-tween the DAC-AT activities of the short and longpromoter regions were small. Northern analysis oftransformants with the cefG gene under the long andshort promoter regions showed little di�erence in thesteady-state levels of the cefG transcript (not shown).
Expression of the cefG from otherfungal promoters results in high transcript levels
To establish whether recognition of the cefG promoterwas really limiting for cefG expression, the native tran-scription-initiation region of the cefG gene was replacedby the promoter of the glyceraldehyde-3-phosphate de-hydrogenase (gpd ) of A. nidulans in two di�erent con-structions pULgpdL and pULgpdS (the long and shortgpd upstream regions respectively) (Fig. 2). Similarconstructions were made using the promoters of theglucoamylase (gla) gene of Aspergillus niger (plasmidpULgla), the glutamate dehydrogenase gene (gdh) ofP. chrysogenum (plasmid pALgdh) and the isopenicillinN synthase gene (pcbC) of P. chrysogenum (plasmidpULpcbC). We avoided the use of homologousA. chrysogenum promoters to prevent disruption of theendogenous allele that are required for proper functioningof primary metabolism or for the early steps of CPCbiosynthesis (in the case of pcbC). In the constructionswith the gla and gpdS promoters the cefG gene was ob-tained from a cDNA library and has no introns. Allconstructions were introduced to two di�erent strains A.chrysogenum ATCC 11550 (wild type, Brotzu strain) andthe higher cephalosporin producer A. chrysogenum C10.Results were similar with both host strains.
An intense cefG transcript band was found in trans-formants with the gla promoter as compared to that ofthe untransformed ATCC11550 strain; a particularlyhigh cefG transcript level was observed with the pro-moters of the pcbC and gdh genes of P. chrysogenum.
To establish whether expression of cefG is still limitingin the high-CPC producer A. chrysogenum C10 (Demain1983; Ramos et al. 1986) two transformants of strain C10with each of the constructions were studied in detail.
Steady-state levels of the cefG transcript were deter-mined in samples taken at 48 h from cultures of alltransformants in de®ned DP1 medium. Constructionswith the gla promoter and A. chrysogenum C10 as con-trol were grown in DP1 medium supplemented withstarch to favor induction of the glucoamylase promoter.As shown in Fig. 3 (upper panel), steady-state transcriptlevels of the cefG gene in strain C10 (without transfor-mation) were very low as compared to those obtainedwith clones expressing the cefG gene from the pcbC, gpdand gdh promoters. No di�erences were observed in thetranscript of the actin gene, used as control, in the dif-ferent transformants (Fig. 3, lower panel).
608
Increase of DAC-AT protein, DAC-AT activity andcephalosporin production with di�erent promoters
The DAC-AT protein and DAC-AT activity levels werealso quanti®ed in the same samples (48 h in de®nedmedium) as those used for RNA extraction. Results(Table 1) showed some variability between individualclones. Higher levels of DAC-AT activity (100%±145%higher than in control untransformed A. chrysogenumC10) were found in clones transformed with gpdL, gpdS,pcbC and gdh constructions. From the industrial pointof view, analysis of a large number of individual cloneswould be preferable to select the best ones.
Similarly, immunoblotting studies (Fig. 4) showedthat strains transformed with the gpdS (lane 3) and gdh(lane 6) constructions synthesized higher amounts ofDAC-AT protein than did control untransformed cul-tures (lane 1). Construction with the gdh promoter
showed an additional band of 44.4 kDa indicating that asmaller protein, which is recognized by anti-DAC-ATantibodies, is formed in this construction.
Southern analysis revealed integration at sitesdi�erent from the endogenous cefG gene
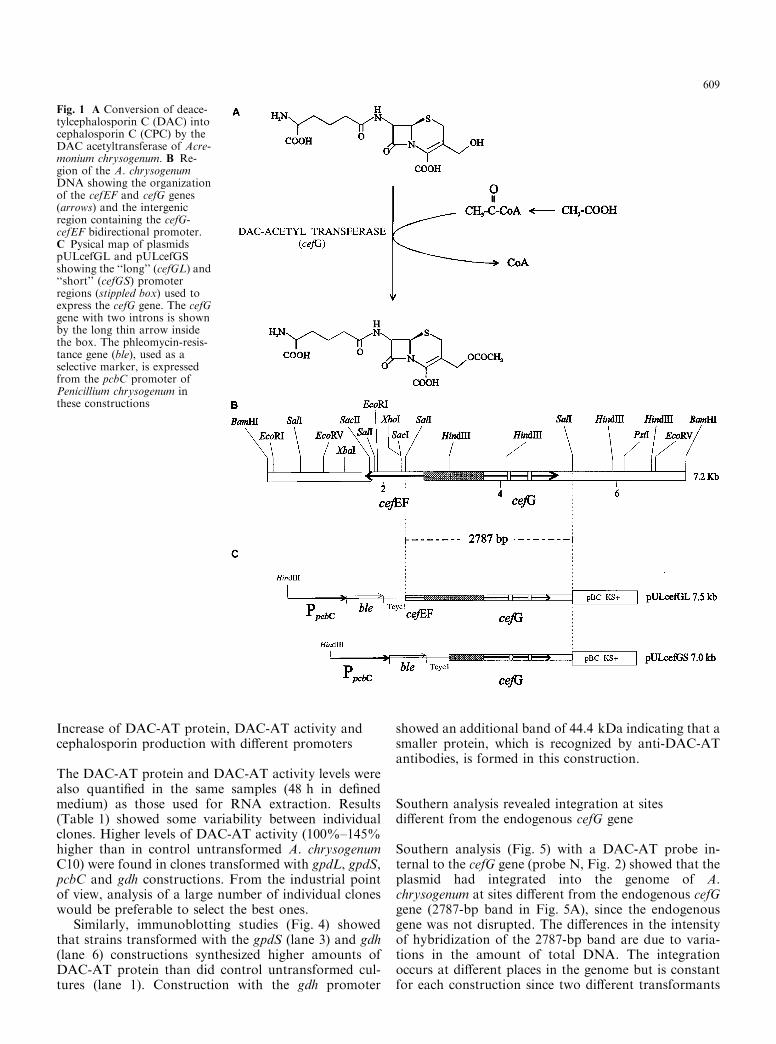
Southern analysis (Fig. 5) with a DAC-AT probe in-ternal to the cefG gene (probe N, Fig. 2) showed that theplasmid had integrated into the genome of A.chrysogenum at sites di�erent from the endogenous cefGgene (2787-bp band in Fig. 5A), since the endogenousgene was not disrupted. The di�erences in the intensityof hybridization of the 2787-bp band are due to varia-tions in the amount of total DNA. The integrationoccurs at di�erent places in the genome but is constantfor each construction since two di�erent transformants
Fig. 1 A Conversion of deace-tylcephalosporin C (DAC) intocephalosporin C (CPC) by theDAC acetyltransferase of Acre-monium chrysogenum. B Re-gion of the A. chrysogenumDNA showing the organizationof the cefEF and cefG genes(arrows) and the intergenicregion containing the cefG-cefEF bidirectional promoter.C Pysical map of plasmidspULcefGL and pULcefGSshowing the ``long'' (cefGL) and``short'' (cefGS) promoterregions (stippled box) used toexpress the cefG gene. The cefGgene with two introns is shownby the long thin arrow insidethe box. The phleomycin-resis-tance gene (ble), used as aselective marker, is expressedfrom the pcbC promoter ofPenicillium chrysogenum inthese constructions
609

isolated randomly showed the same hybridizationpattern (e.g. pULgpdS1 and pULgpdS2, Fig. 5A). Hy-bridizations with the probe P (internal to the phleomycin
resistance marker ble, Fig. 2), to determine whether thetransforming plasmid had integrated into the genome,showed positive hybridization in the transformants and
Fig. 2 Physical map of plas-mids pULgla, pULgpdS,pULgpdL, pULpcbC andpALgdh constructed to expressthe cefG gene from more e�-cient transcription-initiation re-gions (stippled box with arrow).The cefG gene with two intronsis shown by the thin arrowinside the box. Probes N and P,used in hybridization experi-ments, are indicated in theupper part of the ®gure. Thehybridizing bands expected bySouthern experiments, usingprobe N, are shown by a dashedline below each map
Fig. 3 Steady-state levels ofcefG transcript as shown byNorthern hybridization ofRNA extracted from 48-hcultures of A. chrysogenum C10untransformed (control) ortransformed with pULgla,pULgpdS, pULgpdL, pULp-cbC and pALgdh. Two trans-formants for each constructionare shown. The cefG transcript,1.4 kb, is indicated by arrows.Hybridizations were carried outwith probe N (upper panel )internal to the cefG gene (seeFig. 2) and with a probe inter-nal to the actin gene (0.83 kbNcoI-KpnI, Fidel et al. 1988)(lower panel )
610

no signal in the untransformed strain (Fig. 5B). Thehigh number of bands in some lanes indicated integra-tion in multiple loci in the host genome. Ampli®cationand/or recombination of the vector may have occurredin single transformants.
Overexpression of cefG results in increasedcephalosporin production
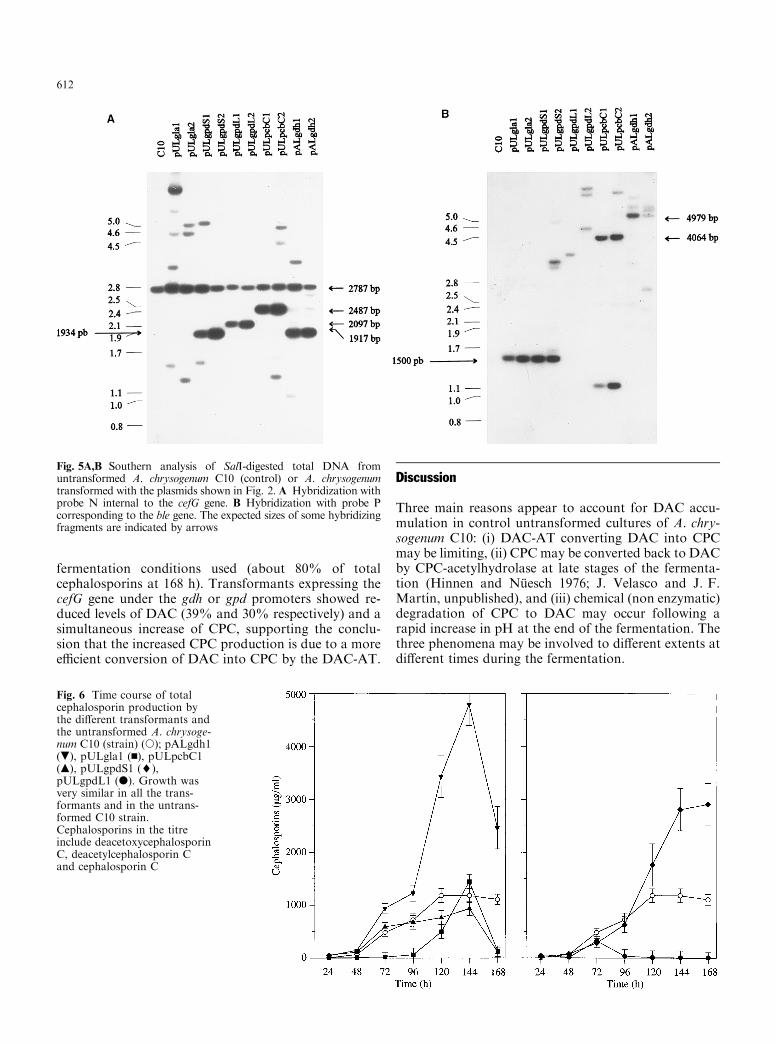
Production of CPC by the most representativeA. chrysogenum C10 transformants was tested in DP1medium. Two transformants of each construction,showing identical levels of phleomycin resistance, wererandomly selected. Transformant 1 of each constructionwas used in the liquid cultures. As shown in Fig. 6, thosetransformants in which the cefG is expressed from thegdh and gpdS promoters resulted in very high levels oftotal cephalosporins, as determined by bioassay (i.e. themixture of DAC and CPC). Growth of all transformantsfollowed the same pattern as that of the control (un-
transformed) C10 strain. Di�erences in dry weight werelower than 10% with respect to the control cultures,indicating that the observed di�erence in total cepha-losporin production is not due to an indirect e�ect of thetransforming DNA on growth of the strains. These re-sults agree well with the expected high expression of thegpd and gdh genes for primary metabolism. A goodcorrelation was obtained between the DAC-AT activi-ties of the di�erent transformants (Table 1) and theability to produce total cephalosporins (Fig. 6). Accu-mulation of total cephalosporins was delayed with res-pect to the cefG transcription time, which started after24 h (Velasco et al. 1994). Most of the promoters used inthis study are promoters of primary metabolism andthey are e�ciently transcribed at 48 h (not shown).
Expression of the cefG transcript from the gpdLpromoter, and cephalosporin production in transform-ants with this construction (pULgpdL), showed a strongburst of DAC-AT activity and cephalosporin produc-tion coinciding with the growth phase, but the enzymeactivity subsequently decreased and cephalosporin pro-duction was halted in repeated experiments. Immuno-blotting studies showed that transformants containingpULgpdL contained lower levels of DAC-AT proteinthan did transformants carrying pULgpdS (Fig. 4). Theunusual behaviour of transformants with the gpdLpromoter, when compared with expression from thegpdS promoters, suggests the implication of upstreamregulatory sequences on growth-phase-dependent ex-pression of this promoter. Indeed, an upstream activat-ing sequence has been described to occur in the 5¢ regionof the gpd promoter (Punt et al. 1992).
Increased conversion of DAC to CPC
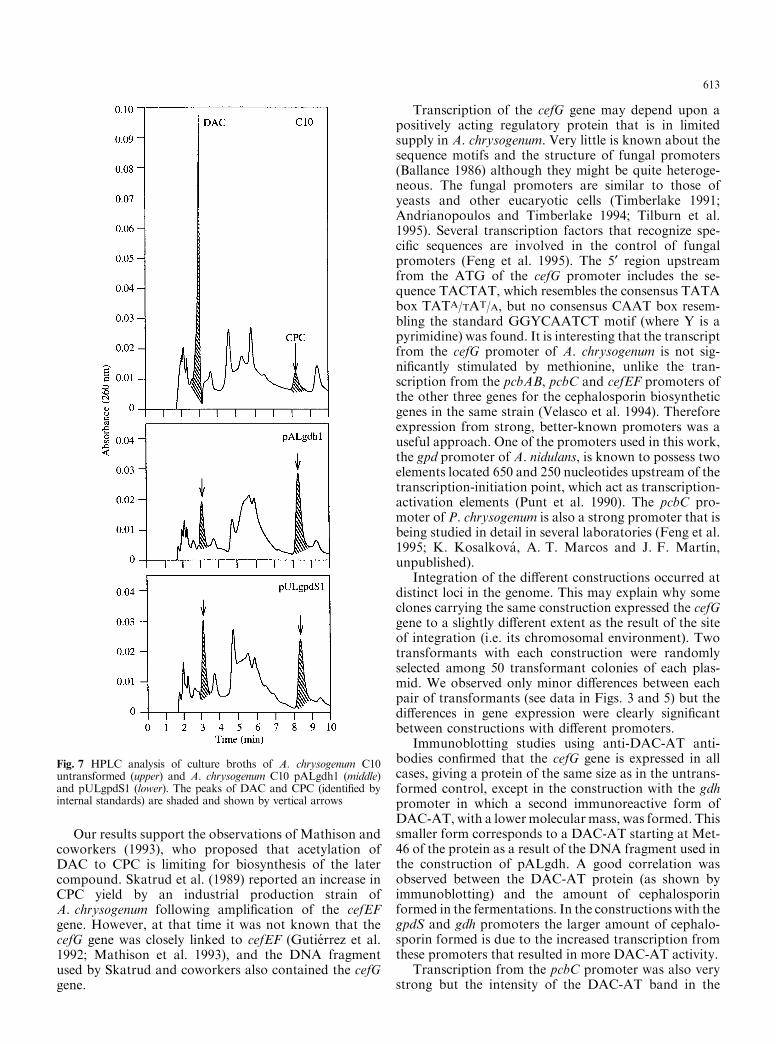
The high levels of cephalosporin obtained with thetransformants expressing cefG under the gpdS and gdhpromoters were due to an increased conversion of DACto CPC. As shown by HPLC analysis (Fig. 7), A. chry-sogenum C10 accumulated high levels of DAC under the
Table 1 DAC-acetyltransferaseactivity in transformants ofA. chrysogenum C10 containingthe cefG gene coupled todi�erent promoters
Transformants DAC-AT pkatals/mg of protein % of control
A. chrysogenum C10 (untransformed) 20.6 100A. chrysogenum C10-starch (untransformed)1 16.4 79.6pULgla1 14.7 71.3pULgla2 15.0 72.8pULgla1-starch1 15.8 96.31
pULgla2-starch1 20.2 123.21
pULgpdL1 50.6 245.6pULgpdL2 17.7 85.9pULgpdS1 39.2 190.3pULgpdS2 35.3 171.3pULpcbC1 45.0 218.4pULpcbC2 41.9 203.3pALgdh1 50.8 246.6pALgdh2 21.3 103.3
1A. chrysogenum C10-starch, gla1-starch and gla2-starch were grown in starch-supplemented DP1medium. Percentage of DAC-AT activity in the transformants with pULgla1 and pULgla2 are givenwith respect to the control A. chrysogenum C10 grown in starch-based medium
Fig. 4 Immunoblotting of acetyl-CoA:DAC acetyltransferase (DAC-AT ) protein in cultures of A. chrysogenum C10 (lane 1) andtransformants pULgla1 (lane 2), pULgpdS1 (lane 3), pULgpdL1(lane 4), pULpcbC1 (lane 5) and pALgdh1 (lane 6) using anti-DAC-AT antibodies. Sizes of standard proteins are indicated by arrows onthe left. The two proteins in lane 6 correspond to the endogenousDAC-AT (methionine 1) and the protein expressed from the gdhpromoter (methionine-46) (see text)
611
fermentation conditions used (about 80% of totalcephalosporins at 168 h). Transformants expressing thecefG gene under the gdh or gpd promoters showed re-duced levels of DAC (39% and 30% respectively) and asimultaneous increase of CPC, supporting the conclu-sion that the increased CPC production is due to a moree�cient conversion of DAC into CPC by the DAC-AT.
Discussion
Three main reasons appear to account for DAC accu-mulation in control untransformed cultures of A. chry-sogenum C10: (i) DAC-AT converting DAC into CPCmay be limiting, (ii) CPC may be converted back to DACby CPC-acetylhydrolase at late stages of the fermenta-tion (Hinnen and NuÈ esch 1976; J. Velasco and J. F.MartõÂ n, unpublished), and (iii) chemical (non enzymatic)degradation of CPC to DAC may occur following arapid increase in pH at the end of the fermentation. Thethree phenomena may be involved to di�erent extents atdi�erent times during the fermentation.
Fig. 5A,B Southern analysis of SalI-digested total DNA fromuntransformed A. chrysogenum C10 (control) or A. chrysogenumtransformed with the plasmids shown in Fig. 2. A Hybridization withprobe N internal to the cefG gene. B Hybridization with probe Pcorresponding to the ble gene. The expected sizes of some hybridizingfragments are indicated by arrows
Fig. 6 Time course of totalcephalosporin production bythe di�erent transformants andthe untransformed A. chrysoge-num C10 (strain) (s); pALgdh1(.), pULgla1 (n), pULpcbC1(m), pULgpdS1 (¨),pULgpdL1 (d). Growth wasvery similar in all the trans-formants and in the untrans-formed C10 strain.Cephalosporins in the titreinclude deacetoxycephalosporinC, deacetylcephalosporin Cand cephalosporin C
612
Our results support the observations of Mathison andcoworkers (1993), who proposed that acetylation ofDAC to CPC is limiting for biosynthesis of the latercompound. Skatrud et al. (1989) reported an increase inCPC yield by an industrial production strain ofA. chrysogenum following ampli®cation of the cefEFgene. However, at that time it was not known that thecefG gene was closely linked to cefEF (Gutie rrez et al.1992; Mathison et al. 1993), and the DNA fragmentused by Skatrud and coworkers also contained the cefGgene.
Transcription of the cefG gene may depend upon apositively acting regulatory protein that is in limitedsupply in A. chrysogenum. Very little is known about thesequence motifs and the structure of fungal promoters(Ballance 1986) although they might be quite heteroge-neous. The fungal promoters are similar to those ofyeasts and other eucaryotic cells (Timberlake 1991;Andrianopoulos and Timberlake 1994; Tilburn et al.1995). Several transcription factors that recognize spe-ci®c sequences are involved in the control of fungalpromoters (Feng et al. 1995). The 5¢ region upstreamfrom the ATG of the cefG promoter includes the se-quence TACTAT, which resembles the consensus TATAbox TATA/TAT/A, but no consensus CAAT box resem-bling the standard GGYCAATCT motif (where Y is apyrimidine) was found. It is interesting that the transcriptfrom the cefG promoter of A. chrysogenum is not sig-ni®cantly stimulated by methionine, unlike the tran-scription from the pcbAB, pcbC and cefEF promoters ofthe other three genes for the cephalosporin biosyntheticgenes in the same strain (Velasco et al. 1994). Thereforeexpression from strong, better-known promoters was auseful approach. One of the promoters used in this work,the gpd promoter of A. nidulans, is known to possess twoelements located 650 and 250 nucleotides upstream of thetranscription-initiation point, which act as transcription-activation elements (Punt et al. 1990). The pcbC pro-moter of P. chrysogenum is also a strong promoter that isbeing studied in detail in several laboratories (Feng et al.1995; K. Kosalkova , A. T. Marcos and J. F. Martõ n,unpublished).
Integration of the di�erent constructions occurred atdistinct loci in the genome. This may explain why someclones carrying the same construction expressed the cefGgene to a slightly di�erent extent as the result of the siteof integration (i.e. its chromosomal environment). Twotransformants with each construction were randomlyselected among 50 transformant colonies of each plas-mid. We observed only minor di�erences between eachpair of transformants (see data in Figs. 3 and 5) but thedi�erences in gene expression were clearly signi®cantbetween constructions with di�erent promoters.
Immunoblotting studies using anti-DAC-AT anti-bodies con®rmed that the cefG gene is expressed in allcases, giving a protein of the same size as in the untrans-formed control, except in the construction with the gdhpromoter in which a second immunoreactive form ofDAC-AT, with a lowermolecular mass, was formed. Thissmaller form corresponds to a DAC-AT starting at Met-46 of the protein as a result of the DNA fragment used inthe construction of pALgdh. A good correlation wasobserved between the DAC-AT protein (as shown byimmunoblotting) and the amount of cephalosporinformed in the fermentations. In the constructions with thegpdS and gdh promoters the larger amount of cephalo-sporin formed is due to the increased transcription fromthese promoters that resulted in more DAC-AT activity.
Transcription from the pcbC promoter was also verystrong but the intensity of the DAC-AT band in the
Fig. 7 HPLC analysis of culture broths of A. chrysogenum C10untransformed (upper) and A. chrysogenum C10 pALgdh1 (middle)and pULgpdS1 (lower). The peaks of DAC and CPC (identi®ed byinternal standards) are shaded and shown by vertical arrows
613

immunoblotting was not proportionally increased, sug-gesting that a translational control of the protein ex-pressed under the pcbC promoter may also occur.Translational control of gene expression is well knownin yeast (Hinnebusch 1993) and preliminary evidencesuggests that it is also involved in the control of pcbCand other genes involved in penicillin biosynthesis(MartõÂ n et al. 1997).
Acknowledgements This work was supported in part by a grant ofAntibio ticos, S.p.A. (Milan, Italy). J. Velasco and F. J. Ferna ndezreceived fellowships from the University of Leo n (Spain) andF. Fierro from the Junta de Castilla y Leo n (Valladolid, Spain). Wethank Antibio ticos, S.A. for providing plasmid pALgdh, DanielRamo n for providing a probe of the Aspergillus nidulans actin gene,Cees van den Hondel for providing plasmids pAN52.1 andpAN52.7, F. Salto and E. Bernasconi of Antibio ticos, S.p.A., andA. Vitaller and A. Collados of Antibio ticos, S.A. (Leo n, Spain) forvaluable discussions.
References
Andrianapoulos A, Timerlake WE (1994) The Aspergillus nidulansabaA gene encodes a transcriptional activator that acts as a ge-netic switch to control development.Mol Cell Biol 14: 2503±2525
Ausubel FM, Brent R, Kingston RE, Moore D, Smith JA, SeidmanJG, Struhl K (eds) (1987) Current protocols in molecular bi-ology. Wiley, New York
Ballance DJ (1986) Sequences important for gene expression in®lamentous fungi. Yeast 2: 229±236
Demain AL (1983) Strain exchange between industry and acade-mia. ASM News 49: 431
Dhawale SA, Lane AC (1993) Compilation of sequence-speci®cDNA-binding proteins implicated in transcriptional control infungi. Yeast 21: 5537±5546
Dunbar BS, Schwoebel ED (1990) Preparation of polyclonal anti-bodies. Methods Enzymol 182: 663±670
Feng B, Friedlin E, Marzluf GA (1995) Nuclear DNA-bindingproteins which recognize the intergenic control region of peni-cillin biosynthetic genes. Curr Genet 27: 351±358
Fidel S, Doonana JH, Morris NR (1988) Aspergillus nidulanscontains a single actin gene which has unique intron locationsand encodes a gamma-actin. Gene 70: 282±293
Fujisawa Y, Kanzaki T (1975) Role of acetyl-CoA:deacetylcepha-losporin C acetyltransferase in cephalosporin biosynthesis byCephalosporium acremonium. Agric Biol Chem 39: 2043±2048
Fujisawa Y, Shirafuji H, Kida M, Nara K, Yoneda M, Kanzaka T(1973) New ®ndings on CPC biosynthesis. Nature New Biol246: 154
Fujisawa Y, Shirafuji H, Kida M, Nara K, Yoneda M, Kanazaki T(1975) Accumulation of deacetylcephalosporin C by cephalo-sporin C negative mutants of Acremonium chrysogenum. AgricBiol Chem 39: 1295±1302
Gutie rrez S, Dõ ez B, Alvarez E, Barredo JL, Martõ n JF (1991)Expression of the penDE gene of Penicillium chrysogenum en-coding isopenicillin N acyltransferase in Cephalosporium acre-monium: production of benzylpenicillin by the transformants.Mol Gen Genet 225: 56±64
Gutie rrez S, Velasco J, Ferna ndez FJ, Martõ n JF (1992) The cefGgene of Cephalosporium acremonium is linked to the cefEF geneand encodes a deacetylcephalosporin C acetyltransferase closelyrelated to homoserine O-acetyltransferase. J Bacteriol 174:3056±3064
Harlow E, Lane D (1988) Antibodies: a laboratory manual. ColdSpring Harbor Laboratory, Cold Spring Harbor, New York
Hinnebusch AG (1993) Gene-speci®c translational control of theyeast GCN4 gene of phosphorylation of eurkaryotic initiationfactor 2. Mol Microbiol 10(2): 215±223
Hinnen A, NuÈ esch J (1976) Enzymatic hydrolysis of cephalosporinC by extracellular acetylhydrolase of Cephalosporium acre-monium. Antimicrob Agents Chemother 9: 824±830
MacKenzie DA, Jeenes DJ, Belshaw NJ, Archer DB (1993) Reg-ulation of secreted protein production by ®lamentous fungi:recent developments and perspectives. J Gen Microbiol 139:2295±2307
Martõ n JF, Gutie rrez S, Ferna ndez FJ, Velasco J, Fierro F, MarcosAT, Kosalkova K (1994) Expression of genes and processing ofenzymes for the biosynthesis of penicillins and cephalosporins.Antonie van Leeuwenhoek 65: 227±243
Martõ n JF, Gutie rrez S, Demain AL (1997) b-Lactams. In: Anke, T(ed) Fungal biotechnology. Chapman & Hall, Weinheim (inpress)
Mathison L, Soliday C, Stepan T, Aldrich T, Rambosek J (1993)Cloning, characterization, and use in strain improvement of theCephalosporium acremonium gene cefG encoding acetyl trans-ferase. Curr Genet 23: 33±41
Matsuda A, Sugiura H, Matsuyama K, Matsumoto H, Ichikawa S,Komatsu K-I (1992) Molecular cloning of acetyl coen-zymeA:deacetylcephalosporinCO-acetyltransferasecDNAfromAcremonium chrysogenum: sequence and expression of catalyticactivity in yeast. Biochem Biophys Res Commun 182: 995±1001
NuÈ esch J, Heim J, Treichler H-J (1987) The biosynthesis of sulfur-containing b-lactam antibiotics. Annu Rev Microbiol 41: 51±57
Punt PJ, Oliver RP, Dingemanse MA, Pouwels PH, HondelCAMJJ van den (1987) Transformation of Aspergillus based onthe hygromycin B resistance marker from Escherichia coli. Gene56: 117±124
Punt PJ, Dingemanse MA, Kuyvenhoven J, Soede RDM, PouwelsPH, Hondel CAMJJ van den (1990) Functional elements in thepromoter region of the Aspergillus nidulans gpdA gene, encodingglyceraldehyde-3-phosphate dehydrogenase. Gene 93: 101±109
Punt PJ, Kramer C, Kuyvenhoven A, Pouwels PH, Hondel CAMJJvan den (1992) An upstream activating sequence from theAspergillus nidulans gpdA gene. Gene 120: 67±73
Ramos FR, Lo pez-Nieto MJ, Martõ n JF (1986) Coordinate in-crease of isopenicillin N synthetase, isopenicillin N epimeraseand deacetoxycephalosporin C synthetase in a high cephalos-porin-producing mutant of A. chrysogenum and simultaneousloss of the three enzymes in a non-producing mutant. FEMSMicrobiol Lett 35: 123±127
Scheidegger A, Gutzwiller A, Kuenzi MT, Fiechter A, NuÈ esch J(1985) Investigation of acetyl-CoA:deacetylcephalosporinC O-acetyltransferase of Cephalosporium acremonium. J Bio-technol 3: 109±117
Shen Y-Q, Wolfe S, Demain AL (1986) Levels of isopenicillinN synthetase and deacetoxycephalosporin C synthetase inCephalosporium acremonium producing high and low levels ofcephalosporin C. Biotechnology 4: 61±64
Skatrud PL, Tietz AJ, Ingolia TD, Cantwell CA, Fisher DL,Chapman JL, Queener SW (1989) Use of recombinant DNA toimprove production of cephalosporin C by Cephalosporiumacremonium. Biotechnology 7: 476±485
Tabor S, Richardson CC (1985) A bacteriophage T7 RNA poly-merase/promoter system for controlled exclusive expression ofspeci®c genes. Proc Natl Acad Sci USA 82: 1074±1078
Tilburn J, Sarkar S, Widdick DA, Espeso EA, Orejas M, MungrooJ, PenÄ alva MA, Arst HN (1995) The Aspergillus PacC zinc®nger transcription factor mediates regulation of both acid-andalkaline-expressed genes by ambient pH. EMBO J 14: 779±790
Timberlake WE (1991) Cloning and analysis of fungal genes. In:Bennett JW, Lasure LL (eds) More gene manipulations infungi. Academic Press, San Diego, Calif, pp 51±85
Velasco J, Gutie rrez S, Ferna ndez FJ, Marcos AT, Areno s C,Martõ n JF (1994) Exogenous methionine increases levels ofmRNAs transcribed from pcbAB, pcbC, and cefEF gene, en-coding enzymes of the cephalosporin biosynthetic pathway, inAcremonium chrysogenum. J Bacteriol 176: 985±991
Zanca DM, MartõÂ n JF (1983) Carbon catabolite regulation of theconversion of penicillin N into cephalosporin C. J Antibiot(Tokyo) 36: 700±708
614
Related Documents











