Expression of NTRK1/TrkA affects immunogenicity of neuroblastoma cells Kristian W. Pajtler 1 , Vera Rebmann 2 , Monika Lindemann 2 , Johannes H. Schulte 1,3 , Stefanie Schulte 1 , Michael Stauder 1 , Ivo Leuschner 4 , Kurt-Werner Schmid 5 , Ulrike K€ ohl 6 , Alexander Schramm 1 and Angelika Eggert 1 1 Department of Pediatric Oncology and Hematology, University Hospital Essen, Germany 2 Department for Transfusion Medicine, University Hospital Essen, Germany 3 Centre for Medical Biotechnology, University Duisburg-Essen, Essen, Germany 4 Institute of Pediatric Pathology, University Hospital Schleswig-Holstein, Germany 5 Department of Pathology, University Hospital Essen, Germany 6 Institute of Cellular Therapeutics, Medical School Hannover, Germany High levels of the NTRK1/TrkA receptor are expressed in low-stage neuroblastomas, which are characterized by a good patient prognosis and often undergo spontaneous regression. In addition to apoptosis, tumor-immune responses might contribute to this regression. We hypothesized that TrkA expression might enhance the immune response to neuroblastomas. Immunohisto- chemistry on neuroblastoma tissue microarrays confirmed significantly higher lymphocyte infiltration in low-stage compared with high-stage tumors. Flow cytometry of human SH-SY5Y cells stably transfected with NTRK1/TrkA cDNA revealed significant upregulation of major histocompatibility complex (MHC) class I complexes on TrkA-expressing cells. Corresponding to this up- regulation, T cell activity and cytoxicity was enhanced in the presence of SY5Y-TrkA cells or by medium conditioned by them, suggesting the existence of additional soluble factors stimulating the T cell response. Activation of natural killer (NK) cells was only increased in the presence of SY5Y-TrkA conditioned medium (CM) and not in co-culture assays, suggesting a domi- nant inhibitory effect of upregulated MHC class I as the primary NK cell escape mechanism of TrkA-expressing neuroblasto- mas. We reanalyzed gene expression data obtained from the cell culture model to identify additional genes involved in the TrkA-mediated modulation of immune responses. Upregulation of selected target genes in SY5Y-TrkA cells was confirmed on transcript and protein levels. However, none of the proteins were detected in medium conditioned by SY5Y-TrkA cells, arguing against these factors as soluble mediators of the TrkA-induced immune response. We here provide evidence that TrkA expres- sion in neuroblastoma leads to an increased immunogenicity that may contribute to a less malignant phenotype and/or spon- taneous regression of neuroblastoma cells. Neuroblastoma is a common solid tumor of childhood and originates from primitive cells of the sympathetic nervous system. It is characterized by a broad biological heterogeneity comprising oncogene amplification or allelic loss, chromo- somal ploidy and expression of neurotrophin receptors corre- lating to a different degree with clinical outcome. 1 The identification of an IgM natural antibody in sera from neuro- blastoma patients and healthy individuals, which exerts potent antitumor activity, suggests that immune responses play a role in controlling this tumor. 2 Although neuroblas- toma cells express tumor-associated antigens that may be rec- ognized by cytotoxic T cells, they often lack constitutive Key words: neurotrophin receptor, spontaneous regression, cytotoxic lymphocytes, NK cells Abbreviations: BDNF: brain-derived neurotrophic factor; CM: conditioned medium; CPM: counts per minute; DC: dendritic cell; IFN-g: interferon-g; MFI: mean fluorescence intensity; MHC: major histocompatibility complex; MICA: MHC class I polypeptide-related sequences A; MICB: MHC class I polypeptide-related sequences B; NCR: natural cytoxicity receptor; NGF: nerve growth factor; PBMC: Peripheral blood mononuclear cells; PBS: phosphate buffered saline; PHA: phytohemagglutinin; TIL: tumor-infiltrating lymphocyte; TMA: Tissue microarrays; ULBP: UL16 binding protein Additional Supporting Information may be found in the online version of this article. Grant sponsors: Kind-Philipp-Stiftung f€ ur Leuk€ amieforschung and the Faculty of Medicine (IFORES grant), University Duisburg-Essen (to K.W.P.); Grant sponsor: European Union (European Network for Cancer Research in Children and Adolescents/ENCCA: 7th Framework Program (to A.E.); Grant number: NoE 261474; Grant sponsor: European Union (Analyzing and Striking the Sensitivities of Embryonal Tumours/ASSET: 7th Framework Program) (to A.E.); Grant number: CP 259348 DOI: 10.1002/ijc.28096 History: Received 29 Aug 2012; Accepted 24 Jan 2013; Online 7 Feb 2013 Correspondence to: Dr. Kristian W. Pajtler, Department of Pediatric Oncology and Hematology, University Hospital Essen, Hufelandstraße 55, Essen 45147, Germany, Tel.: 149-201-723 85157, Fax: 149-201-723 5386, E-mail: [email protected] Tumor Immunology Int. J. Cancer: 00, 000–000 (2013) V C 2013 UICC International Journal of Cancer IJC

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Expression of NTRK1/TrkA affects immunogenicity ofneuroblastoma cells
Kristian W. Pajtler1, Vera Rebmann2, Monika Lindemann2, Johannes H. Schulte1,3, Stefanie Schulte1, Michael Stauder1,
Ivo Leuschner4, Kurt-Werner Schmid5, Ulrike K€ohl6, Alexander Schramm1 and Angelika Eggert1
1 Department of Pediatric Oncology and Hematology, University Hospital Essen, Germany2 Department for Transfusion Medicine, University Hospital Essen, Germany3 Centre for Medical Biotechnology, University Duisburg-Essen, Essen, Germany4 Institute of Pediatric Pathology, University Hospital Schleswig-Holstein, Germany5 Department of Pathology, University Hospital Essen, Germany6 Institute of Cellular Therapeutics, Medical School Hannover, Germany
High levels of the NTRK1/TrkA receptor are expressed in low-stage neuroblastomas, which are characterized by a good patient
prognosis and often undergo spontaneous regression. In addition to apoptosis, tumor-immune responses might contribute to
this regression. We hypothesized that TrkA expression might enhance the immune response to neuroblastomas. Immunohisto-
chemistry on neuroblastoma tissue microarrays confirmed significantly higher lymphocyte infiltration in low-stage compared
with high-stage tumors. Flow cytometry of human SH-SY5Y cells stably transfected with NTRK1/TrkA cDNA revealed significant
upregulation of major histocompatibility complex (MHC) class I complexes on TrkA-expressing cells. Corresponding to this up-
regulation, T cell activity and cytoxicity was enhanced in the presence of SY5Y-TrkA cells or by medium conditioned by them,
suggesting the existence of additional soluble factors stimulating the T cell response. Activation of natural killer (NK) cells
was only increased in the presence of SY5Y-TrkA conditioned medium (CM) and not in co-culture assays, suggesting a domi-
nant inhibitory effect of upregulated MHC class I as the primary NK cell escape mechanism of TrkA-expressing neuroblasto-
mas. We reanalyzed gene expression data obtained from the cell culture model to identify additional genes involved in the
TrkA-mediated modulation of immune responses. Upregulation of selected target genes in SY5Y-TrkA cells was confirmed on
transcript and protein levels. However, none of the proteins were detected in medium conditioned by SY5Y-TrkA cells, arguing
against these factors as soluble mediators of the TrkA-induced immune response. We here provide evidence that TrkA expres-
sion in neuroblastoma leads to an increased immunogenicity that may contribute to a less malignant phenotype and/or spon-
taneous regression of neuroblastoma cells.
Neuroblastoma is a common solid tumor of childhood andoriginates from primitive cells of the sympathetic nervoussystem. It is characterized by a broad biological heterogeneitycomprising oncogene amplification or allelic loss, chromo-somal ploidy and expression of neurotrophin receptors corre-lating to a different degree with clinical outcome.1 The
identification of an IgM natural antibody in sera from neuro-blastoma patients and healthy individuals, which exertspotent antitumor activity, suggests that immune responsesplay a role in controlling this tumor.2 Although neuroblas-toma cells express tumor-associated antigens that may be rec-ognized by cytotoxic T cells, they often lack constitutive
Key words: neurotrophin receptor, spontaneous regression, cytotoxic lymphocytes, NK cells
Abbreviations: BDNF: brain-derived neurotrophic factor; CM: conditioned medium; CPM: counts per minute; DC: dendritic cell;
IFN-g: interferon-g; MFI: mean fluorescence intensity; MHC: major histocompatibility complex; MICA: MHC class I polypeptide-related
sequences A; MICB: MHC class I polypeptide-related sequences B; NCR: natural cytoxicity receptor; NGF: nerve growth factor; PBMC:
Peripheral blood mononuclear cells; PBS: phosphate buffered saline; PHA: phytohemagglutinin; TIL: tumor-infiltrating lymphocyte;
TMA: Tissue microarrays; ULBP: UL16 binding protein
Additional Supporting Information may be found in the online version of this article.
Grant sponsors: Kind-Philipp-Stiftung f€ur Leuk€amieforschung and the Faculty of Medicine (IFORES grant), University Duisburg-Essen (to
K.W.P.); Grant sponsor: European Union (European Network for Cancer Research in Children and Adolescents/ENCCA: 7th Framework
Program (to A.E.); Grant number: NoE 261474; Grant sponsor: European Union (Analyzing and Striking the Sensitivities of Embryonal
Tumours/ASSET: 7th Framework Program) (to A.E.); Grant number: CP 259348
DOI: 10.1002/ijc.28096
History: Received 29 Aug 2012; Accepted 24 Jan 2013; Online 7 Feb 2013
Correspondence to: Dr. Kristian W. Pajtler, Department of Pediatric Oncology and Hematology, University Hospital Essen, Hufelandstraße
55, Essen 45147, Germany, Tel.: 149-201-723 85157, Fax: 149-201-723 5386, E-mail: [email protected]
Tum
orIm
mun
olog
y
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC
International Journal of Cancer
IJC

expression of co-stimulatory molecules and surface HLA classI and II molecules. This renders them a difficult target for thehost T cell compartment.3 In contrast, it has been shown thatNK cells are able to lyse neuroblastoma cells in vitro andinhibit neuroblastoma growth in mice and humans.4–6 Neuro-blastoma cells display numerous strategies to escape immunecontrol mechanisms. Nonetheless, immunotherapy withmonoclonal antibodies directed against tumor-associated anti-gens, such as the GD2 disialoganglioside, has been tested inadvanced clinical trials with promising results.7 Despite theneed to better understand tumor-immune response mecha-nisms to further improve immunotherapeutic approachesagainst neuroblastoma, few reports have investigated thenature of the tumor-immune effector cell interaction.
High expression of the neurotrophic tyrosine kinase type 1receptor, NTRK1, also named TrkA, is associated with a favor-able prognosis in neuroblastoma.8 TrkA activation by bindingof the specific ligand, nerve growth factor (NGF), inhibitsangiogenesis, induces differentiation and growth arrest andmediates apoptosis.9,10 In contrast, high tumor expression ofNTRK2/TrkB and its specific ligand, brain-derived neurotro-phic factor (BDNF), correlates with progressive disease andunfavorable prognosis in neuroblastoma patients based on anautocrine loop leading to enhanced proliferation, metastaticbehavior and chemoresistance of neuroblastoma cells.11 Trk re-ceptor signalling is a contributing factor to the diverse spec-trum of clinical presentation in neuroblastoma patients, whichranges from widespread metastatic disease that is refractory toaggressive multimodal therapies to complete spontaneousregression or differentiation in low-stage or stage 4s neuroblas-tomas, even without therapeutic intervention.1
Neuroblastoma has the highest rate of spontaneous regres-sion observed among human cancers.12 Based on the biologi-cal characteristics of neuroblastomas, various mechanismsincluding delayed activation of normal apoptotic pathways,13
inhibition of angiogenesis14 and anti-tumor reactions by theimmune system15 have been suggested to contribute to thisphenomenon. Most low-stage neuroblastomas express highTrkA levels, leading to differentiation in the presence of NGFor apoptosis in its absence. Therefore, TrkA is likely involvedin the regulation of spontaneous regression as well as differ-entiation of neuroblastomas with favorable biology.16
Low-stage neuroblastomas are often heavily infiltrated bytumor-infiltrating lymphocytes (TILs), suggesting that theimmune system carries out surveillance for these tumors.17
Furthermore, differentiation of neuroblastoma cells hasrecently been shown to increase antigenicity of these cells,making them better targets for immune effector cells.18
Based on these observations, we hypothesized that TrkAexpression might enhance immunogenicity of neuroblastomacells in addition to its previously described biological effects.The resulting increased immune response might contributeto spontaneous neuroblastoma regression and finally lead toa less malignant tumor phenotype. To further investigate thepotential implications of TrkA expression for the immuneresponse, we examined the interaction between TrkA- orTrkB-expressing neuroblastoma cells with immune effectorcells in more detail.
Material and MethodsCell culture, conditioned medium and cell models
Neuroblastoma cell lines were cultured as described.19 Cell lineidentity was authenticated by DSMZ (Braunschweig, Germany).NTRK1/TrkA or NTRK2/TrkB cDNAs were cloned into the ret-roviral expression vectors, pLNCX or pLNCX2 (Clontech, PaloAlto, CA), respectively.20 NTRK1/TrkA was also stablyexpressed in NB69 human neuroblastoma cells. Single-cellclones of transfected cells were obtained by limiting dilution.The empty pLNCX vector (SY5Yvec, NB68vec) served as nega-tive control. Sequencing confirmed the identity of all transfec-tants. For generation of conditioned medium (CM),neuroblastoma cell cultures were washed with phosphate buf-fered saline (PBS) and incubated 48 hr with serum-free me-dium. CM was centrifuged after collection and completeprotease inhibitor cocktail (Roche Applied Science, Mannheim,Germany) was added before 50-fold concentration using Ami-con Ultra-15 centrifugal filter units (Millipore, Billerica, MA).Human NTRK1/TrkA cDNA was also cloned into the pT-REx-DEST30 vector (Invitrogen, Carlsbad, CA) for Tet-conditionalexpression. The SH-SY5Y cell line was sequentially transfectedwith pT-REx-DEST30, then pcDNA6/TR (Invitrogen), harbor-ing the tetracycline repressor gene. Single-cell clones wereselected by limiting dilution in blasticidine-containing medium.
Tissue microarrays and immunohistochemistry
Tissue microarrays (TMAs) were prepared from paraffin-embedded tissue of 78 primary neuroblastomas from thearchival files of the Institutes of Pathology, Universities ofKiel and Essen. Clinical data and informed patient consentwere obtained within the German Neuroblastoma Trial BFM-
What’s new?
Elevated expression of neurotrophic tyrosine kinase type 1 receptor (NTRK1, or TrkA) previously was found to be associated
with favorable prognosis in neuroblastoma. That effect may be the result of TrkA-induced enhancement of the immune
response toward neuroblastoma cells, as reported here. The findings suggest that TrkA-associated immune activity might
partly explain spontaneous regression of low-stage neuroblastomas. They also have novel translational implications, among
them the possibility for the development of T cell-based immunotherapy approaches.
Tum
orIm
mun
olog
y
2 NTRK1 affects neuroblastoma immunogenicity
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC
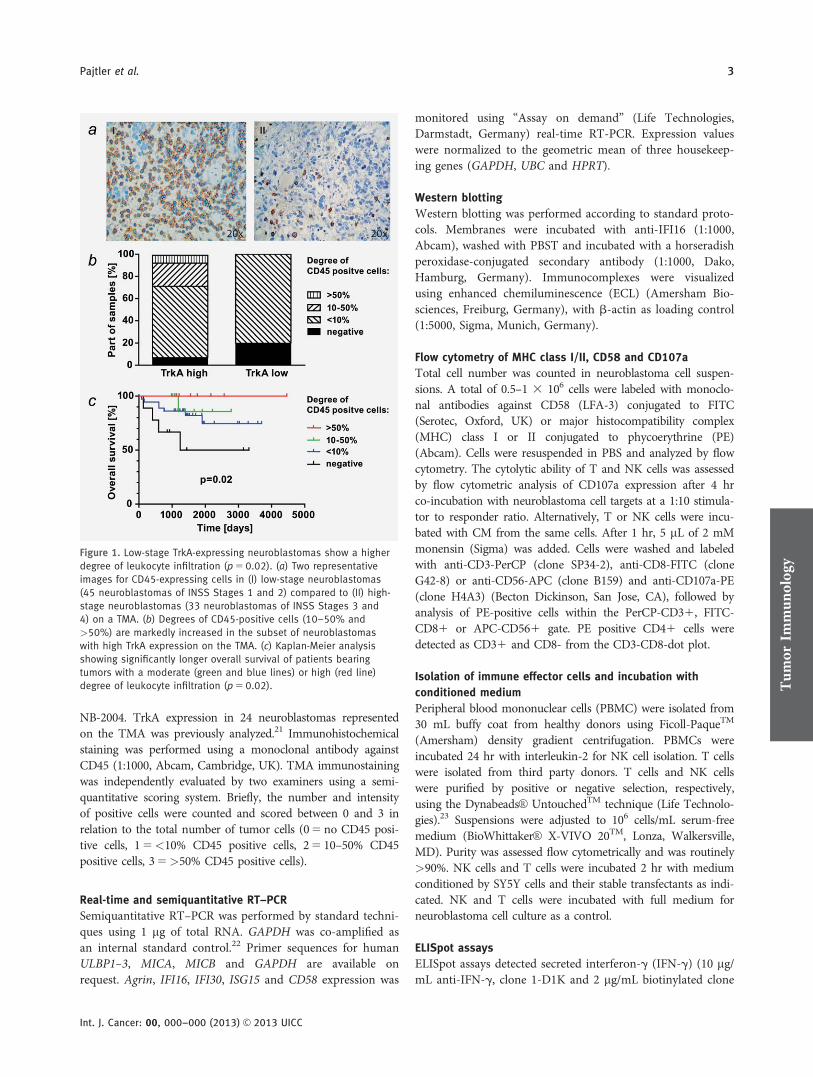
NB-2004. TrkA expression in 24 neuroblastomas representedon the TMA was previously analyzed.21 Immunohistochemicalstaining was performed using a monoclonal antibody againstCD45 (1:1000, Abcam, Cambridge, UK). TMA immunostainingwas independently evaluated by two examiners using a semi-quantitative scoring system. Briefly, the number and intensityof positive cells were counted and scored between 0 and 3 inrelation to the total number of tumor cells (05 no CD45 posi-tive cells, 15<10% CD45 positive cells, 25 10–50% CD45positive cells, 35>50% CD45 positive cells).
Real-time and semiquantitative RT–PCR
Semiquantitative RT–PCR was performed by standard techni-ques using 1 mg of total RNA. GAPDH was co-amplified asan internal standard control.22 Primer sequences for humanULBP1–3, MICA, MICB and GAPDH are available onrequest. Agrin, IFI16, IFI30, ISG15 and CD58 expression was
monitored using “Assay on demand” (Life Technologies,Darmstadt, Germany) real-time RT-PCR. Expression valueswere normalized to the geometric mean of three housekeep-ing genes (GAPDH, UBC and HPRT).
Western blotting
Western blotting was performed according to standard proto-cols. Membranes were incubated with anti-IFI16 (1:1000,Abcam), washed with PBST and incubated with a horseradishperoxidase-conjugated secondary antibody (1:1000, Dako,Hamburg, Germany). Immunocomplexes were visualizedusing enhanced chemiluminescence (ECL) (Amersham Bio-sciences, Freiburg, Germany), with b-actin as loading control(1:5000, Sigma, Munich, Germany).
Flow cytometry of MHC class I/II, CD58 and CD107a
Total cell number was counted in neuroblastoma cell suspen-sions. A total of 0.5–1 3 106 cells were labeled with monoclo-nal antibodies against CD58 (LFA-3) conjugated to FITC(Serotec, Oxford, UK) or major histocompatibility complex(MHC) class I or II conjugated to phycoerythrine (PE)(Abcam). Cells were resuspended in PBS and analyzed by flowcytometry. The cytolytic ability of T and NK cells was assessedby flow cytometric analysis of CD107a expression after 4 hrco-incubation with neuroblastoma cell targets at a 1:10 stimula-tor to responder ratio. Alternatively, T or NK cells were incu-bated with CM from the same cells. After 1 hr, 5 lL of 2 mMmonensin (Sigma) was added. Cells were washed and labeledwith anti-CD3-PerCP (clone SP34-2), anti-CD8-FITC (cloneG42-8) or anti-CD56-APC (clone B159) and anti-CD107a-PE(clone H4A3) (Becton Dickinson, San Jose, CA), followed byanalysis of PE-positive cells within the PerCP-CD31, FITC-CD81 or APC-CD561 gate. PE positive CD41 cells weredetected as CD31 and CD8- from the CD3-CD8-dot plot.
Isolation of immune effector cells and incubation with
conditioned medium
Peripheral blood mononuclear cells (PBMC) were isolated from30 mL buffy coat from healthy donors using Ficoll-PaqueTM
(Amersham) density gradient centrifugation. PBMCs wereincubated 24 hr with interleukin-2 for NK cell isolation. T cellswere isolated from third party donors. T cells and NK cellswere purified by positive or negative selection, respectively,using the DynabeadsVR UntouchedTM technique (Life Technolo-gies).23 Suspensions were adjusted to 106 cells/mL serum-freemedium (BioWhittakerVR X-VIVO 20TM, Lonza, Walkersville,MD). Purity was assessed flow cytometrically and was routinely>90%. NK cells and T cells were incubated 2 hr with mediumconditioned by SY5Y cells and their stable transfectants as indi-cated. NK and T cells were incubated with full medium forneuroblastoma cell culture as a control.
ELISpot assays
ELISpot assays detected secreted interferon-g (IFN-g) (10 mg/mL anti-IFN-g, clone 1-D1K and 2 mg/mL biotinylated clone
Figure 1. Low-stage TrkA-expressing neuroblastomas show a higher
degree of leukocyte infiltration (p 5 0.02). (a) Two representative
images for CD45-expressing cells in (I) low-stage neuroblastomas
(45 neuroblastomas of INSS Stages 1 and 2) compared to (II) high-
stage neuroblastomas (33 neuroblastomas of INSS Stages 3 and
4) on a TMA. (b) Degrees of CD45-positive cells (10–50% and
>50%) are markedly increased in the subset of neuroblastomas
with high TrkA expression on the TMA. (c) Kaplan-Meier analysis
showing significantly longer overall survival of patients bearing
tumors with a moderate (green and blue lines) or high (red line)
degree of leukocyte infiltration (p 5 0.02). Tum
orIm
mun
olog
y
Pajtler et al. 3
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC

7-B6-1) and IL-10 (10 mg/mL anti-IL-10, clone 9D7 and1 mg/mL biotinylated clone 12G8). All antibodies wereobtained from Mabtech (Nacka, Sweden). ELISpot plateswere incubated with 60 mL of primary antibody overnight at4�C before NK and T cell isolation, washed three times andblocked with cell culture medium.24 Three different types ofexperiments were performed: (i) using nitrocellulose ELISpotplates, duplicates of 160,000 NK cells were incubated with10,000 K562 erythroleukemic cells for 48 hr after activationwith neuroblastoma cell CM for 2 hr, (ii) using nitrocelluloseELISpot plates, 400,000 T cells were incubated with neuro-blastoma cell CM for 2 hr, then stimulated by PHA (1 mg/mL) for 24 hr and (iii) using PVDF ELISpot plates, 100,000allogeneic T cells were stimulated by co-culture with 10,000neuroblastoma cells for a total of 72 hr. After co-culture/stimulation at 37�C, plates were washed six times. Capturedcytokines were detected by incubation for 2 hr with 60 mL ofthe second antibody and color reactions.24,25
Cytotoxicity assays
NK cell cytotoxicity was quantified colorimetrically using theLDH-Cytotoxicity Assay kit (Biovision). Effector and target
cells were mixed 20:1 and 10:1, plated in triplicates in a 96-well plate and incubated for 2 hr at 37�C. Cell supernatantswere then transferred to a plate containing the kit substratemix to detect LDH activity. After 60 min at RT, the absorb-ance at 450 nm was determined. Cell-mediated cytotoxicitywas calculated using: cytotoxicity effector (%)5 [(target cellmix2 spontaneous effector LDH release2 spontaneous targetLDH release)/(maximum target LDH release2spontaneoustarget LDH release)]3100. NK cell cytotoxicity toward neu-roblastoma cells in direct contact was assessed flow cytomet-rically. After 2 hr of NK and neuroblastoma cell co-culture,dead cells were determined by propidium iodide staining.MHC depletion of neuroblastoma cells was performed usingantibodies against MHC class I (clone w6/32) and MHC classII (anti-CD74) as a control (Abcam).
Lymphocyte transformation test
Proliferative T cell responses to the mitogen PHA (1 mg/mL)were measured using 200,000 T cells in 200 mL of Bio-WhittakerVR X-VIVO 20 culture medium/well in microtitreplates (37�C, 5% CO2). Tests were conducted in triplicate.Cells were cultured 3 days and 1 mCi 3H-thymidine(TRA.120, specific activity 5 Ci/mmol, Amersham) was addedper well for the last 16 hr of culture. Cells were harvestedonto filter pads, and the incorporated radioactivity was quan-tified by liquid scintillation counting.26
Microarray data and statistical analyses
Correlation of gene expression and Kaplan-Meier analyseswas carried out using microarray data previously obtained forthe same cell culture model on R2 (http://r2.amc.nl).19 Dataanalyses in R2 were performed using “Time Series” tools,including “Correlation analyses” and “Gene Filtering” usingKEGG pathways on all arrays deployed in this study.SPSS18.0 was used for further statistical analysis. GO termanalysis was conducted for genes showing a >3-fold expres-sion difference between SY5Y-TrkA and SY5Y-TrkB cells. GOterms resulting in a p< 0.01 corrected for multiple testingwere considered. GO categories were first ranked according tothe corresponding UniGene entries of differentially expressedgenes, solely considering terms having more than ten entries,then the resulting GO categories were sorted by p-value cor-rected for multiple testing. A Student’s two-sided t-test wasused to compare all interval variables, and a chi-square testwas used to compare all categorical variables. Graph PadPrism 5.0 was used to perform Kaplan-Meier survival analysiswith log-rank statistics. Microarray data have been depositedin the GEO database, accession no. GSE18409.
ResultsLow-stage neuroblastomas expressing NTRK1/TrkA show a
higher degree of leukocyte infiltration
To gain first insights about interactions between neuroblas-toma cells and the immune system, we assessed the degreethat primary neuroblastomas were infiltrated by immune
Figure 2. MHC class I molecules are upregulated on TrkA-express-
ing neuroblastoma cells independently of differentiation and pres-
ence of NGF. Flow cytometric analysis of MHC class I expression on
the surface of the indicated neuroblastoma cell lines. Bars indicate
percentages of MHC class I-positive cells, mean 6 SD of three or
more experiments is presented. (a) **P<0.01 indicates signifi-
cantly higher expression of MHC class I on SY5Y-TrkA cells com-
pared to SY5Y-TrkB and the empty vector control, SY5Yvec, cells.
(b) Significantly higher expression of MHC class I on TrkA-express-
ing cells *p<0.05 was confirmed in a second stably transfected
neuroblastoma cell line (NB69).
Tum
orIm
mun
olog
y
4 NTRK1 affects neuroblastoma immunogenicity
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC
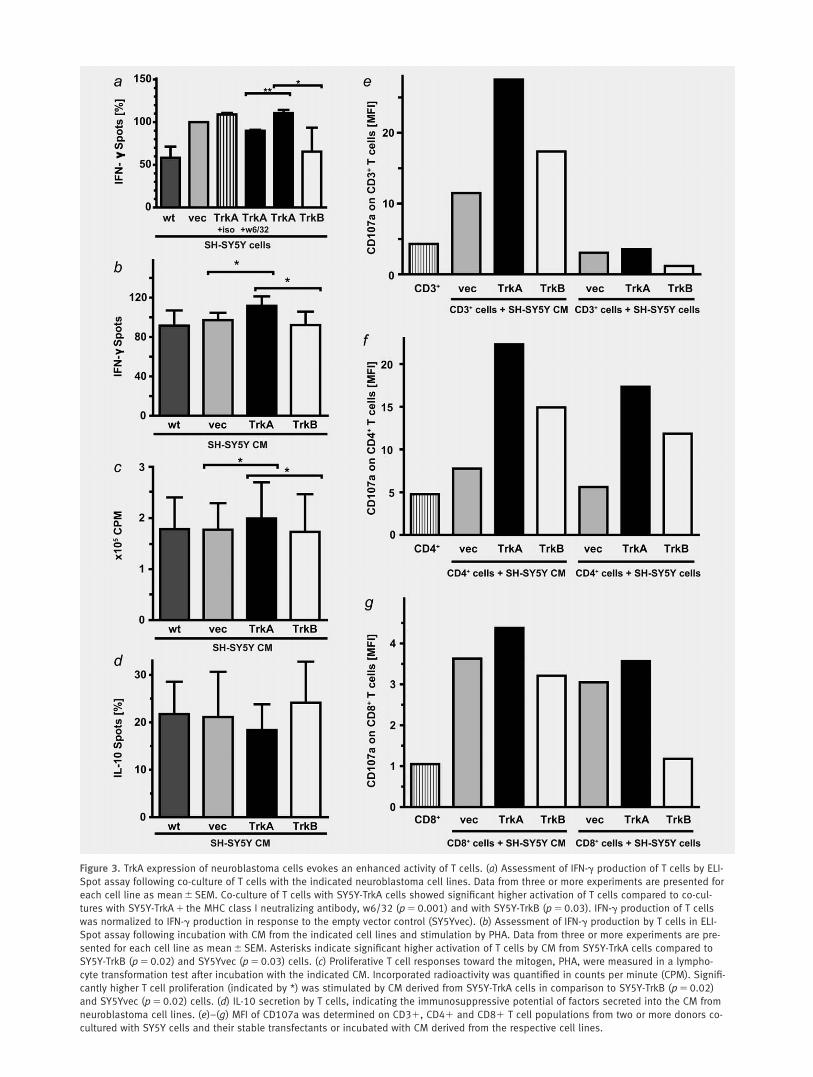
Figure 3. TrkA expression of neuroblastoma cells evokes an enhanced activity of T cells. (a) Assessment of IFN-g production of T cells by ELI-
Spot assay following co-culture of T cells with the indicated neuroblastoma cell lines. Data from three or more experiments are presented for
each cell line as mean 6 SEM. Co-culture of T cells with SY5Y-TrkA cells showed significant higher activation of T cells compared to co-cul-
tures with SY5Y-TrkA 1 the MHC class I neutralizing antibody, w6/32 (p 5 0.001) and with SY5Y-TrkB (p 5 0.03). IFN-g production of T cells
was normalized to IFN-g production in response to the empty vector control (SY5Yvec). (b) Assessment of IFN-g production by T cells in ELI-
Spot assay following incubation with CM from the indicated cell lines and stimulation by PHA. Data from three or more experiments are pre-
sented for each cell line as mean 6 SEM. Asterisks indicate significant higher activation of T cells by CM from SY5Y-TrkA cells compared to
SY5Y-TrkB (p 5 0.02) and SY5Yvec (p 5 0.03) cells. (c) Proliferative T cell responses toward the mitogen, PHA, were measured in a lympho-
cyte transformation test after incubation with the indicated CM. Incorporated radioactivity was quantified in counts per minute (CPM). Signifi-
cantly higher T cell proliferation (indicated by *) was stimulated by CM derived from SY5Y-TrkA cells in comparison to SY5Y-TrkB (p 5 0.02)
and SY5Yvec (p 5 0.02) cells. (d) IL-10 secretion by T cells, indicating the immunosuppressive potential of factors secreted into the CM from
neuroblastoma cell lines. (e)–(g) MFI of CD107a was determined on CD31, CD41 and CD81 T cell populations from two or more donors co-
cultured with SY5Y cells and their stable transfectants or incubated with CM derived from the respective cell lines.

effector cells. TMAs of 78 tumors representing all stages wereimmunohistochemically stained using an unrestrictedantibody against the CD45 leukocyte tyrosine phosphatase.High-stage neuroblastomas (Stages 3 and 4, n5 33) had sig-nificantly less leukocyte infiltration compared to low-stageneuroblastomas (Stages 1 and 2, n5 45) (p5 0.02; Fig. 1a).Neuroblastoma infiltrating leukocytes have previously beenshown to predominantly originate from the T cell lineage,characterized by CD4 or CD8 expression, with some CD56-positive cells being of NK cell origin or representing NK-likeT cells.27 A specific TrkA antibody, not cross-reacting withTrkB and suitable for immunohistochemistry, is currently notcommercially available. Thus, TrkA expression in the arrayedtumors was assessed at the mRNA level from existing expres-sion microarray data. A substantial part of tumors with highTrkA expression contained moderate and high levels ofCD45-positive cells (defined as >10% and >50% infiltration,respectively), whereas all tumors with low TrkA expressionshowed <10% or no CD45-positive cells (Fig. 1b). The pres-ence of TILs in the tumor mass has previously been reportedto correlate with favorable prognosis of neuroblastomapatients.17 Accordingly, the degree of leukocyte infiltration inour tumor cohort was predictive for overall patient survival(log-rank test, p5 0.02) in Kaplan-Meier analysis (Fig. 1c).Multivariate analysis revealed only stage and CD45 positivityas independent prognostic markers (p5 0.0007 and p5 0.04,respectively). However, the important prognostic marker,MYCN amplification, could not be included in the analysisdue to the low number of MYCN-amplified tumors on theTMA, limiting the significance of the multivariate analysis.Taken together, low-stage neuroblastomas with high TrkAexpression were characterized by a higher percentage of leu-kocyte infiltration, suggesting a more active tumor responsefrom the immune system, which is likely to contribute to abetter patient outcome.
MHC class I molecules are upregulated on TrkA-expressing
neuroblastoma cells independently of differentiation and
the presence of NGF
Since the mechanisms underlying recruitment of lymphoidinfiltrates to low-stage primary neuroblastomas are poorlyunderstood to date, we aimed to further assess a potentialrelationship between TrkA receptor expression and neuro-blastoma cell immunogenicity in vitro. We first focussed onMHC class I molecules, as (i) they are major players in regu-lating immune effector cell interactions with target cells and(ii) have previously been implicated in TrkA-mediated differ-entiation of neuroblastoma cells.18 It has been shown that thecells in most primary neuroblastomas express very low MHCclass I levels on their surfaces, thereby, generating a mecha-nism for immune escape.28 However, neuroblastoma cell lineswere previously demonstrated to upregulate expression ofMHC class I molecules during differentiation, regardless ofTrkA expression.18 To discriminate between the effect of dif-ferentiation and TrkA expression on MHC class I molecule
expression, the percentage of cells expressing MHC class Icomplexes was analyzed by flow cytometry in an establishedhuman neuroblastoma cell culture model stably expressingfull-length constructs for NTRK1/TrkA, NTRK2/TrkB or theempty vector in the SH-SY5Y cell line (designated SY5Y-TrkA, SY5Y-TrkB and SY5Yvec, respectively).10,29 Interest-ingly, twice as many SY5Y-TrkA cells expressed MHC class Imolecules even in an undifferentiated state and in the ab-sence of exogenous NGF compared to the parental cell line,and ten-fold more SY5Y-TrkA cells expressed MHC class Imolecules than SY5Y-TrkB cells (Fig. 2a). These findingswere confirmed in a second human neuroblastoma cell cul-ture model, NB69 cells stably transfected with NTRK1/TrkA(NB69-TrkA) or empty vector (NB69vec) (Fig. 2b).30 Theseresults demonstrate that TrkA expression and basal activationin neuroblastoma cells is sufficient to upregulate MHC class Iexpression independently of differentiation or the presence ofexogenous NGF. In addition, cytokine expression profiles ofneuroblastoma cell lysates and their CM did not display sig-nificant overexpression of cytokines known to be involved inMHC class I upregulation (data not shown). Thus, MHCclass I upregulation in TrkA transfectants is (i) not artificiallyinduced by the transfection process itself and (ii) presumablyindependent of the expression of major cellular cytokines.
TrkA expression in neuroblastoma cells enhances T cell
activity and cytotoxicity
Since the expression level of MHC class I complexes on thetarget cells is a crucial parameter for T cell activation, thehigher MHC class I expression by SY5Y-TrkA cells is likelyto affect, for example, the T cell alloresponse to these cells.To test this hypothesis, SY5Y-TrkA cells were co-culturedwith human allogeneic T cells from healthy, MHC class Iand II disparate donors. After 2 days in co-culture on ELI-Spot plates, T cell activation was analyzed by measuringIFN-g production as a surrogate marker for functional ac-tivity in the T cells. As neuroblastoma cell lines onlyexpress MHC class I, but no MHC class II, as determinedby flow cytometric analysis (data not shown), the IFN-gproduction can be attributed to MHC class I disparity. Ourprevious experiments indicate that after 3 days in culture (1day pre-incubation and 2 days in ELISpot plates) MHCclass I differences lead to IFN-g production. Co-culture ofT cells with SY5Y-TrkA cells enhanced IFN-g productionby the T cells in comparison to co-culture with the parentalcell line or TrkB-expressing cells (Fig. 3a). IFN-g produc-tion also was slightly elevated in T cells co-cultured withSY5Yvec cells, suggesting some unspecific activation ofimmune effector cells by the vector system itself or follow-ing stable transfection. Enhanced IFN-g production by Tcells was reduced by approximately 15%, when MHC class Imolecules on SY5Y-TrkA cells were blocked using a specificantibody against MHC class I (w6/32) (Fig. 3a). This sug-gests additional factors to be involved in increased T cellactivation.
Tum
orIm
mun
olog
y
6 NTRK1 affects neuroblastoma immunogenicity
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC

Figure 4. TrkA expression increases NK cell activation, but inhibits direct NK cell cytotoxicity. (a) NK cell-mediated lysis of indicated cells in
co-culture experiments. NK cell cytotoxicity against SY5Y-TrkA cells significantly increased after depletion of MHC class I (p 5 0.01). An anti-
body against MHC class II (anti-CD74) was used as control. (b), (c) NK cells from three or more healthy donors were incubated with CM
from the stably transfected NB cells, SY5Yvec, SY5Y-TrkA or SY5Y-TrkB. NK cell activity was measured by their IFN-g production in an ELI-
Spot assay, NK cell-mediated lysis by colorimetric quantification of LDH-release. Medium without supplements was used as control. (d)
CD107a expression in NK cells as a read out for their cytotoxic capability measured as MFI was comparably low in response to direct cell
contact with SY5Y cells and their stable transfectants. (e) Expression of activating NKG2D ligands was analyzed on mRNA levels by semi-
quantitative RT-PCR in the indicated cell lines.

To assess a potential additional contribution of secretedsoluble factors to the observed effect of SY5Y-TrkA cells onT cell function, we repeated the IFN-g production assayusing CM collected after 48 hr of neuroblastoma cell culture.
T cells were incubated for 2 hr with CM, then stimulated byphytohemagglutinin (PHA). Interestingly, CM from SY5Y-TrkA cells showed a significant higher ability to induceIFN-g production in T cells in comparison to SY5Y-TrkB,SY5Yvec and SY5Ywt cells (Fig. 3b). In addition, lymphocyteproliferation measured by lymphocyte transformation testwas higher in T cells incubated with SY5Y-TrkA CM thanwith SY5Y-TrkB, SY5Yvec or SY5Ywt CM (Fig. 3c). In con-trast, T cells incubated with SY5Y-TrkA CM secreted lessinterleukin-10 (IL-10) than T cells incubated with mediumconditioned by SY5Y-TrkB, SY5Ywt or SY5Yvec cells (Fig.3d), indicating lymphocyte suppression. These results suggestthat SY5Y-TrkA cells harbor a significantly higher potentialto activate T cells by both membrane-adherent and solublesecreted factors. Direct cell contact is likely to contribute tothe tumor-immune cell interaction, but is not solely responsi-ble for the TrkA-mediated effect on T cells.
To further resolve the functional properties of different Tcell subgroups during TrkA-mediated T cell effects, weassessed the degranulation capacity of the subgroups by flowcytometric analysis of CD107a/LAMP-1 surface expression.31
CD107a is a vesicle membrane protein in immune effectorcells that becomes transiently mobilized to the cell surface dur-ing the degranulation process, leading to the release of cyto-toxic mediators such as perforin or granzymes. Its surfaceexpression has previously been shown to correlate withimmune effector cell cytotoxic activity, and can be used as amarker for the capability of T or NK cells to kill tumor tar-gets.31 The mean fluorescence intensity (MFI) of CD107a wasdetermined on CD31, CD41 and CD81 T cell subpopula-tions co-cultured with SY5Y cells or their stable transfectantsor incubated with medium conditioned by the respective celllines. CD107a expression was markedly elevated in all T cellsubpopulations co-cultured with SY5Y-TrkA cells or CMderived from these cells (Figs. 3e–3g), demonstrating a TrkA-mediated stimulation of both cytotoxic CD81 T cells as wellas CD41 T helper cells, by membrane-adherent and solublefactors. CD81 cytotoxic T cells, but not CD31 and CD41 Tcells, also exhibited a limited amount of degranulation activityafter co-culture with SY5Yvec cells or incubation with theirCM. These results corroborate the results from our cytokine
Figure 5.
Figure 5. Immune modulating genes are differentially expressed in
SY5Y-TrkA versus SY5Y-TrkB expressing cells. (a) Reanalysis of
microarray expression profiles obtained in 68 primary neuroblasto-
mas shows a correlation between the mRNA expression levels of
TrkA and agrin in these tumors (p 5 4 3 1028). (b) Real-time PCR
validation of gene expression in SY5Y cells and their stable trans-
fectants confirmed upregulation of all five target genes in SY5Y-
TrkA cells. (c) Target gene expression in a SY5Y cell line with tetra-
cycline-inducible expression of TrkA. Induction of TrkA led to
increased expression of all five selected target genes, with mark-
edly enhanced expression after addition of the TrkA-specific ligand,
NGF. (d), (e) Upregulation of the selected target proteins on the
surface of neuroblastoma cells was confirmed by flow cytometry
for CD58 or western blot analysis for IFI16.
Tum
orIm
mun
olog
y
8 NTRK1 affects neuroblastoma immunogenicity
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC

response assay, and together demonstrate that TrkA expressionon neuroblastoma cells enhances T cell activity and cytoxicity.
TrkA expression increases NK cell activation, but inhibits
direct NK cell cytotoxicity
Potential interactions between TrkA-expressing neuroblas-toma cells and NK cells are also of interest for neuroblastomaimmunotherapy development. We, therefore, also assessedthe effects of TrkA expression on NK cell-mediated lysis oftumor cells in co-culture experiments. Co-culture of SY5Y-TrkA cells with NK cells did not significantly enhance NKcell cytotoxicity in comparison to co-culture with SY5Y,SY5Yvec and SY5Y-TrkB cells (Fig. 4a). The absence of a sig-nificant difference could be explained by MHC class I upreg-ulation in TrkA-expressing neuroblastoma cells counteractingother potentially positive TrkA-mediated effects on the NKcells. To test whether MHC class I expression inhibited NKcell cytotoxicity toward SY5Y-TrkA cells, MHC class I mole-cules were depleted in cell co-cultures by adding an MHCclass I antibody to culture media. MHC class I depletionrestored NK cell cytotoxicity toward SY5Y-TrkA cells, whereaslysis of SY5Yvec and SY5Y-TrkB cells remained low (Fig. 4a).NK cells were significantly less cytotoxic toward SY5Y cells andtheir stable transfectants than toward K562 cells, an erythroleu-kemia cell line lacking MHC class I expression and known tobe an NK cell-sensitive target. These results suggest an inhibi-tion of NK cell cytotoxicity in TrkA-expressing neuroblastomacells via upregulation of MHC class I as the primary immuneescape mechanism, whereas different immune escape mecha-nisms seem to be in place in SY5Yvec and SY5Y-TrkB cells.
To assess the need for direct cell–cell contact for theobserved effects on tumor-immune cell interaction, NK cellactivation and cytoxicity were measured after exposure tomedium conditioned by SY5Y-TrkA, SY5Y-TrkB, SY5Yvec orSY5Ywt cells. NK cell activity measured by IFN-g productionincreased only in response to SY5Y-TrkA CM (Fig. 4b).SY5Y-TrkA CM also enhanced NK cell cytotoxicity towardK562 cells in comparison to medium conditioned by SY5Y,SY5Yvec or SY5Y-TrkB cells (Fig. 4c). In contrast to theresults for T cells, SY5Y-TrkA cells or their CM did notenhance degranulation in NK cells assessed by analysis ofCD107a expression (Fig. 4d). CD107a expression in NK cellswas comparably low in response to SY5Y cells and all theirstable transfectants, whereas K562 cells demonstrated ahigher capability to induce CD107a. These results suggestthat humoral factors secreted by SY5Y-TrkA cells can stimu-late NK cell cytotoxicity toward non-neuroblastoma targetcells, such as K562 cells, to some extent. However, theinduced NK cell cytotoxicity is insufficient to kill neuroblas-toma target cells, which are likely protected by membrane-associated molecules on the tumor cell surface. MHC class Iexpression is only one possible immune escape mechanism,as SY5Yvec and SY5Y-TrkB cells also proved to be pro-tected from NK cell lysis independently of MHC class Iexpression.
Tumor cells may also express ligands of NK cell activatingreceptors, such as the UL16 binding proteins (ULBPs), MHCclass I polypeptide-related sequences A (MICA) or B (MICB)or natural ligands of the NKG2D receptor, to regulate theinteraction with immune effector cells. We analyzed mRNAexpression of these activating ligands in the SY5Y cell culturemodel. ULBP2, ULBP3, MICA and MICB were indeed signif-icantly upregulated on the surface of SY5Y-TrkA cells incomparison to SY5Y, SY5Yvec and SY5Y-TrkB cells (Fig. 4e).ULBP1 expression was upregulated on the surfaces of bothSY5Y-TrkA and SY5Y-TrkB cells in comparison to SY5Yvecand the parental SY5Y cells. These results suggest that thetumor-protecting effects of membrane-associated factors,including MHC class I molecules, cancel out the activatingeffects of expressing NKG2D ligands toward NK cells.
Immune modulating genes are differentially expressed in
TrkA- and TrkB-expressing neuroblastoma cells
To search for additional underlying molecular mechanismsfor TrkA-mediated immune responses, we reanalyzed Affyme-trix U95A expression data previously obtained for the sameSH-SY5Y cell culture model to identify further TrkA-regu-lated genes capable of enhancing tumor immunogenicity,apart from MHC class I.19 Comparison of SY5Y-TrkA andSY5Y-TrkB cell transcriptomes revealed multiple differentiallyexpressed genes, even in the absence of the respective exoge-nous ligands, NGF or BDNF. An automated search in the R2molecular database32 for a distinct subgroup of genes relevantfor tumor immunology based on gene ontology annotationsidentified 17 differentially expressed genes (p< 0.002). Weselected five TrkA-induced candidate genes with the highestpotential relevance for immune modulations for further anal-yses (Supporting Information Table 1). To assess the potentialrelevance of these five candidate genes in vivo, we reanalyzedmicroarray expression profiles obtained from 68 primary neu-roblastomas for correlations with TrkA expression levels inthese tumors. Agrin expression significantly correlated withTrkA expression (Fig. 5a), whereas only a trend toward posi-tive correlation with TrkA expression was observed for theother four candidates. We next investigated this connection inthe SH-SY5Y cell culture model using real-time RT-PCR. Allfive candidate genes were upregulated in SY5Y-TrkA cellscompared with parental cells and SY5Y-TrkB cells (Fig. 5b).However, the five candidate genes were also slightly upregu-lated in SY5Yvec cells compared with the parental cell line.As an inducible model can better control for artificial andunspecific effects induced by stable transfection of the emptyvector, we repeated the experiments using a vector system forconditional TrkA expression operating under a tetracyclinerepressor. Induction of NTRK1/TrkA resulted in reproducibleupregulation of CD58, IFI16 and ISG15 expression, which wasfurther enhanced by addition of NGF to the culture medium(Fig. 5c). Enhanced CD58 expression on the cell surface wasconfirmed by flow cytometry, and enhanced IFI16 expressionwas detected in western blots of whole-cell lysates (Fig. 5d).
Tum
orIm
mun
olog
y
Pajtler et al. 9
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC

However, western blotting of CM conditioned by the stablytransfected SY5Y-TrkA cells or the inducible TrkA modelcells could not detect soluble CD58 and IFI16 (data notshown), arguing against these factors as the underlying molec-ular mechanism of immune effector cell activation mediatedby secreted factors. High-throughput proteomic analyses arewarranted to identify additional molecular players of theTrkA-mediated immune response to neuroblastoma cells.
DiscussionThe molecular mechanisms regulating neuroblastoma cell im-munogenicity are not yet completely understood. We showhere that TrkA overexpression in human neuroblastoma cellsaffects tumor cell antigenicity and markedly enhances activa-tion of immune effector cells. We demonstrate (i) increasedleukocyte infiltration of TrkA-expressing low-stage neuroblas-tomas, (ii) an enhanced ability of TrkA-expressing neuroblas-toma cells to stimulate T cell activation, proliferation andcytotoxicity in vitro, (iii) a corresponding upregulation ofMHC class I molecules as major players in the regulation ofimmune responses by TrkA-expressing neuroblastoma cellsand (iv) an increased ability of TrkA-expressing neuroblas-toma cells to stimulate NK cell activation, but not cytotoxic-ity toward neuroblastoma cells.
We show that the percentage of TILs is significantly higherin low-stage neuroblastomas expressing high levels of TrkA.This observation is in agreement with a previous report dem-onstrating a correlation between TILs and neuroblastomatumor stage.27 TILs infiltrating low-stage neuroblastomas havepreviously been characterized to be predominantly cells of theT cell lineage with a smaller fraction of CD56-positive cellsbeing of NK cell origin or representing NK-like T cells.27 Thepresence of TILs in solid tumors has been described as afavorable prognostic factor in numerous tumor entitiesincluding neuroblastoma33 and our results verify this for afurther cohort of primary neuroblastomas.
A successful T cell-mediated immune response requiresexpression of MHC classes I and II. It has been shown thatneuroblastoma cells express antigens suitable for recognitionby cytotoxic T cells, however, they display little to no surfaceMHC class I.28,34,35 Defects in the antigen-processingmachinery have been described as a potential underlyingmechanism.36 MHC class I expression can be raised by INFg
or retinoid treatment.3,37,38 T cell-based immunotherapy can-not be successful unless MHC class I expression is upregu-lated on the neuroblastoma cell surface. We show here thatTrkA overexpression or induction in a neuroblastoma cellmodel upregulated MHC class I expression. TrkA-expressingneuroblastoma cells induced to differentiate in vitro by NGFtreatment have previously been shown to upregulate MHCclass I expression, however, MHC class I upregulation wasattributed to differentiation rather than a direct effect ofTrkA upregulation.18 We demonstrate here that TrkA expres-sion in the absence of NGF is sufficient to upregulate MHCclass I expression at the neuroblastoma cell surface. Trk
receptors have previously been shown to have a high capacityfor ligand-independent activation particularly in neuroblas-toma, presumably via spontaneous interactions.19
MHC class I upregulation on neuroblastoma cells haspreviously been shown to result in increased CTL cytoxic-ity.18 Indeed, we observed an increase in T cell activation,proliferation and cytotoxicity in response to TrkA-express-ing neuroblastoma cells, with both CD81 and CD41 Tcells demonstrating enhanced responses. Interestingly, theseeffects were not only restricted to cell–cell contacts andincreased MHC class I expression, but were also mediatedby CM, suggesting that at least some effector molecules aresoluble factors. Our first attempts to identify these solublefactors based on differentially expressed mRNAs wereunsuccessful. An unbiased proteomic approach with massspectrometric identification of differentially expressed pro-teins in conditioned media derived from the SY5Y andNB69 cell culture models should be a more promisingapproach.
Although not detected as soluble proteins in CM of theneuroblastoma cell culture models, the TrkA-upregulated andvalidated candidate genes, CD58, IFI16, ISG15 and agrinmight nonetheless play a role in the modulation of neuro-blastoma immunogenicity. CD58 is a ligand of the T lympho-cyte CD2 protein that functions in adhesion andT lymphocyte activation by advancing the contact betweencytolytic effector cells and target cells.39 Monoclonal antibod-ies neutralizing CD58 have been shown to inhibit melanomacell lysis by allogeneic NK cells and autologous TILs.40 IFI16has recently been identified as a double-strand DNA(dsDNA) receptor involved in T cell effector functions, as in-tracellular dsDNA is a potent activator of human dendriticcells (DCs).41 ISG15 is a ubiquitin-like protein that becomesconjugated to many cellular proteins upon activation byIFN-a.42 Interestingly, it is located on chromosome 1p36.3 ina region hypothesized to harbor one or more neuroblastomatumor suppressor genes. ISG15 stimulates IFN-g productionby CD31 T cells, as well as NK cell proliferation and non-MHC-mediated tumor cell cytotoxicity.43 Overexpression ofagrin was very recently shown to occur in activated T cells,leading to further activation of these immune effecor cells bysynergizing with the T cell receptor.44 If and how the hereidentified increased expression of CD58, IFI16, ISG15 andagrin, in TrkA-expressing neuroblastoma cells contributes tothe increased immunogenicity of neuroblastoma cells remainsto be elucidated in further analyses.
The “missing self” recognition theory describes malig-nantly transformed cells lacking MHC class I expression astriggering NK cell-dependent activation.45 Vice versa, upregu-lation of class I MHC expression in TrkA-expressing neuro-blastoma cells inhibited NK cell cytoxicity as expected, andneutralizing MHC class I using an antibody restored NK cellcytotoxicity toward SY5Y-TrkA cells. In contrast, mediumconditioned by SY5Y-TrkA cells activated NK cells andstimulated cytotoxicity against K562 cells, which do not
Tum
orIm
mun
olog
y
10 NTRK1 affects neuroblastoma immunogenicity
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC

express MHC class I. As a balance of activating and inhibi-tory signals mediates NK cell recognition and killing of tar-get cells,46,47 we also assessed the presence of NK cellactivating signals in the SY5Y cell culture model. The natu-ral cytoxicity receptors (NCRs), NKp30, NKp44 andNKp46, are the main activating receptors, for which onlyfew ligands are known and remain poorly characterized onthe molecular level.48 NKG2D, 2B4 and DNAM-1 are alsoactivating receptors, and recognize a variety of well-definedligands.46 MICA, MICB and ULBPs are among the beststudied ligands, and bind NKG2D. Upregulation of theseligands has been detected in solid tumors of multiple ori-gins, including neuroblastomas,49 and might be a cell intrin-sic protective mechanism to render transformed cellssusceptible to killing. NK cells expressing high NKG2D lev-els, especially following IL-2 stimulation, have previouslybeen reported to lyse MHC class I-positive target cells.5,50
Although TrkA-expressing neuroblastoma cells express highNKG2D ligand levels, inhibitory signals mediated by MHCclass I expression appear to override the effect, since weobserved NK cell activation but no significant cytotoxicity
toward SY5Y-TrkA cells unless MHC class I expression wasdepleted.
Taken together, we demonstrate that TrkA-expressionenhances the immune response toward neuroblastoma cellsby regulating immune-modulating surface molecules includ-ing MHC class I molecules as well as secreted factors yet tobe identified. These results point to three potentially impor-tant clinical implications: (i) the TrkA-induced immuneresponse might explain a part of the molecular mechanismsresulting in spontaneous regression of low-stage neuroblasto-mas; (ii) therapeutic induction of TrkA might be a promisingapproach to create a less malignant and more treatable tumorphenotype and (iii) the subgroup of neuroblastomas withhigh TrkA expression might not be a suitable target for NKcell-based immunotherapy, but would rather benefit from Tcell-based immunotherapy approaches.
AcknowledgementsThe authors thank E. Mahlow and A. Rieb for excellent technical assistance,K. Astrahantseff for article proofreading and H. Stephan for figurepreparation.
References
1. Brodeur GM. Neuroblastoma: biological insightsinto a clinical enigma. Nat Rev Cancer2003;3:203–16.
2. Ollert MW, David K, Schmitt C, et al. Normalhuman serum contains a natural IgM antibodycytotoxic for human neuroblastoma cells. ProcNatl Acad Sci U S A 1996;93:4498–503.
3. Prigione I, Corrias MV, Airoldi I, et al.Immunogenicity of human neuroblastoma. AnnN Y Acad Sci 2004;1028:69–80.
4. Huenecke S, Zimmermann SY, Kloess S, et al. IL-2-driven regulation of NK cell receptors withregard to the distribution of CD161 and CD16-subpopulations and in vivo influence afterhaploidentical NK cell infusion. J Immunother2010;33:200–10.
5. Kloess S, Huenecke S, Piechulek D, et al. IL-2-activated haploidentical NK cells restore NKG2D-mediated NK-cell cytotoxicity in neuroblastomapatients by scavenging of plasma MICA. Eur JImmunol 2010;40:3255–67.
6. Cheung NK, Modak S. Oral (1–>3),(1–>4)-beta-D-glucan synergizes with antiganglioside GD2monoclonal antibody 3F8 in the therapy ofneuroblastoma. Clin Cancer Res 2002;8:1217–23.
7. Yu AL, Gilman AL, Ozkaynak MF, et al. Anti-GD2 antibody with GM-CSF, interleukin-2, andisotretinoin for neuroblastoma. N Engl J Med2010;363:1324–34.
8. Nakagawara A, Arima-Nakagawara M, ScavardaNJ, et al. Association between high levels ofexpression of the TRK gene and favorableoutcome in human neuroblastoma. N Engl J Med1993;328:847–54.
9. Eggert A, Grotzer MA, Zuzak TJ, et al. Resistanceto tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis inneuroblastoma cells correlates with a loss ofcaspase-8 expression. Cancer Res 2001;61:1314–9.
10. Eggert A, Ikegaki N, Liu X-G, et al. Moleculardissection of TrkA signal transduction pathways
mediating differentiation in humanneuroblastoma cells. Oncogene 2000;19:2043–51.
11. Nakagawara A, Azar CG, Scavarda NJ, et al.Expression and function of Trk-B and BDNF inhuman neuroblastomas. Mol Cell Biol1994;14:759–67.
12. Yamamoto K, Hanada R, Kikuchi A, et al.Spontaneous regression of localizedneuroblastoma detected by mass screening. J ClinOncol 1998;16:1265–9.
13. Brodeur GM, Castle VP. In: Hickman JA, DiveC, eds. Apoptosis and cancer chemotherapy. NewJersey: Humana, 1999. 305–18.
14. Papac RJ. Spontaneous regression of cancer:possible mechanisms. In Vivo 1998;12:571–8.
15. Bolande RP. A natural immune system inpregnancy serum lethal to human neuroblastomacells: a possible mechanism of spontaneousregression. Perspect Pediatr Pathol 1992;16:120–33.
16. Brodeur GM, Minturn JE, Ho R, et al. Trkreceptor expression and inhibition inneuroblastomas. Clin Cancer Res 2009;15:3244–50.
17. Martin RF, Beckwith JB. Lymphoid infiltrates inneuroblastomas: their occurrence and prognosticsignificance. J Pediatr Surg 1968;3:161–4.
18. Carlson LM, Pahlman S, De Geer A, et al.Differentiation induced by physiological andpharmacological stimuli leads to increasedantigenicity of human neuroblastoma cells. CellRes 2008;18:398–411.
19. Schulte JH, Schramm A, Klein-Hitpass L, et al.Microarray analysis reveals differential geneexpression patterns and regulation of single targetgenes contributing to the opposing phenotype ofTrkA- and TrkB-expressing neuroblastomas.Oncogene 2005;24:165–77.
20. Eggert A, Ho R, Ikegaki N, et al. Different effectsof TrkA expression in neuroblastoma cell lines
with or without MYCN amplification. MedPediatr Oncol 2000;35:623–27.
21. Schramm A, Schulte JH, Klein-Hitpass L, et al.Prediction of clinical outcome and biologicalcharacterization of neuroblastoma by expressionprofiling. Oncogene 2005;24:7902–12.
22. Eggert A, Brodeur GM, Ikegaki N. A relativequantitative RT-PCR protocol for TrkB expressionin neuroblastoma using GAPD as an internalcontrol. Biotechniques 2000;28:681–91.
23. Riley JL, Blair PJ, Musser JT, et al. ICOScostimulation requires IL-2 and can be preventedby CTLA-4 engagement. J Immunol2001;166:4943–8.
24. Lindemann M, Bohmer J, Zabel M, Grosse-WildeH. ELISpot: a new tool for the detection of nickelsensitization. Clin Exp Allergy 2003;33:992–8.
25. Lindemann M, Saure C, Klinkenbusch H, et al.Alloreactivity in recipients prior to and postliving kidney and liver transplantation. Scand JImmunol 2011;73:344–5.
26. Lindemann M, Virchow S, Ramann F, et al. TheG protein beta3 subunit 825T allele is a geneticmarker for enhanced T cell response. FEBS Lett2001;495:82–6.
27. Facchetti P, Prigione I, Ghiotto F, et al.Functional and molecular characterization oftumour-infiltrating lymphocytes and clonesthereof from a major-histocompatibility-complex-negative human tumour: neuroblastoma. CancerImmunol Immunother 1996;42:170–8.
28. Wolfl M, Jungbluth AA, Garrido F, et al.Expression of MHC class I, MHC class II, andcancer germline antigens in neuroblastoma.Cancer Immunol Immunother 2005;54:400–6.
29. Eggert A, Grotzer MA, Ikegaki N, et al.Expression of the neurotrophin receptor TrkAdown-regulates expression and function ofangiogenic stimulators in SH-SY5Yneuroblastoma cells. Cancer Res 2002;62:1802–8.
Tum
orIm
mun
olog
y
Pajtler et al. 11
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC

30. Eggert A, Sieverts H, Ikegaki N, et al. p75mediated apoptosis in neuroblastoma cells isinhibited by expression of trkA. Med PediatrOncol 2000;35:573–76.
31. Betts MR, Brenchley JM, Price DA, et al. Sensitiveand viable identification of antigen-specific CD81 Tcells by a flow cytometric assay for degranulation. JImmunol Methods 2003;281:65–78.
32. Koster J. R2: microarray analysis andvisualization platform Amsterdam, 2008.
33. Okada K, Komuta K, Hashimoto S, et al.Frequency of apoptosis of tumor-infiltratinglymphocytes induced by fas counterattack inhuman colorectal carcinoma and its correlationwith prognosis. Clin Cancer Res 2000;6:3560–4.
34. Lampson LA, Fisher CA, Whelan JP. Strikingpaucity of HLA-A, B, C and beta 2-microglobulinon human neuroblastoma cell lines. J Immunol1983;130:2471–8.
35. Ponzoni M, Casalaro A, Lanciotti M, et al. Thecombination of interferon and tumor necrosis factorcauses a rapid and extensive differentiation ofhuman neuroblastoma cells. Cancer Res1992;52:931–39.
36. Raffaghello L, Prigione I, Bocca P, et al. Multipledefects of the antigen-processing machinerycomponents in human neuroblastoma:immunotherapeutic implications. Oncogene2005;24:4634–44.
37. Drew PD, Lonergan M, Goldstein ME, et al.Regulation of MHC class I and beta 2-microglobulin gene expression in humanneuronal cells. Factor binding to conserved cis-acting regulatory sequences correlates withexpression of the genes. J Immunol1993;150:3300–10.
38. Vertuani S, De Geer A, Levitsky V, et al. Retinoidsact as multistep modulators of the majorhistocompatibility class I presentation pathway andsensitize neuroblastomas to cytotoxic lymphocytes.Cancer Res 2003;63:8006–13.
39. Wang JH, Smolyar A, Tan K, et al. Structure of aheterophilic adhesion complex between thehuman CD2 and CD58 (LFA-3) counterreceptors.Cell 1999;97:791–803.
40. Altomonte M, Gloghini A, Bertola G, et al.Differential expression of cell adhesion moleculesCD54/CD11a and CD58/CD2 by humanmelanoma cells and functional role in theirinteraction with cytotoxic cells. Cancer Res1993;53:3343–8.
41. Kis-Toth K, Szanto A, Thai TH, et al. CytosolicDNA-activated human dendritic cells are potentactivators of the adaptive immune response.J Immunol 187:1222–34.
42. Farrell PJ, Broeze RJ, Lengyel P. Accumulation ofan mRNA and protein in interferon-treatedEhrlich ascites tumour cells. Nature1979;279:523–5.
43. D’Cunha J, Ramanujam S, Wagner RJ, et al. Invitro and in vivo secretion of human ISG15, anIFN-induced immunomodulatory cytokine.J Immunol 1996;157:4100–8.
44. Kabouridis PS, Pimentel TA, Brancaleone V,et al. Distinct localization of T cell Agrin duringantigen presentation—evidence for the expressionof Agrin receptor(s) in antigen-presenting cells.FEBS J 2012;279:2368–80.
45. Ljunggren HG, Karre K. In search of the“missing self”: MHC molecules and NK cellrecognition. Immunol Today 1990;11:237–44.
46. Lanier LL. NK cell recognition. Annu RevImmunol 2005;23:225–74.
47. Vivier E, Tomasello E, Baratin M, et al.Functions of natural killer cells. Nat Immunol2008;9:503–10.
48. Groth A, Kloss S, von Strandmann EP, et al.Mechanisms of tumor and viral immune escapefrom natural killer cell-mediated surveillance.J Innate Immun 2011;3:344–54.
49. Raffaghello L, Prigione I, Airoldi I, et al.Downregulation and/or release of NKG2Dligands as immune evasion strategy of humanneuroblastoma. Neoplasia 2004;6:558–68.
50. Diefenbach A, Jensen ER, Jamieson AM, et al.Rae1 and H60 ligands of the NKG2D receptorstimulate tumour immunity. Nature2001;413:165–71.
Tum
orIm
mun
olog
y
12 NTRK1 affects neuroblastoma immunogenicity
Int. J. Cancer: 00, 000–000 (2013) VC 2013 UICC
Related Documents











