Expression of Ghrelin and Its Functional Receptor, the Type 1a Growth Hormone Secretagogue Receptor, in Normal Human Testis and Testicular Tumors F. GAYTAN, M. L. BARREIRO, J. E. CAMINOS, L. K. CHOPIN, A. C. HERINGTON, C. MORALES, L. PINILLA, R. PANIAGUA, M. NISTAL, F. F. CASANUEVA, E. AGUILAR, C. DIE ´ GUEZ, AND M. TENA-SEMPERE Departments of Cell Biology, Physiology, and Immunology (F.G., M.L.B., L.P., E.A., M.T.-S.), and Pathology (C.M.), University of Cordoba, 14004 Cordoba, Spain; Departments of Physiology (J.E.C., C.D.) and Medicine (F.F.C.), University of Santiago de Compostela, 15705 Santiago de Compostela, Spain; Center for Molecular Biotechnology, Queensland University of Technology (L.K.C., A.C.H.), Brisbane, Queensland, Australia; Department of Cell Biology and Genetics, University of Alcala (R.P.), 28871 Madrid, Spain; and Department of Morphology, University Autonoma (M.N.), 28029 Madrid, Spain Ghrelin, the endogenous ligand for the GH secretagogue re- ceptor (GHS-R), has been primarily linked to the central neu- roendocrine regulation of GH secretion and food intake, al- though additional peripheral actions of ghrelin have also been reported. In this context, the expression of ghrelin and its cognate receptor has been recently demonstrated in rat testis, suggesting a role for this molecule in the direct control of male gonadal function. However, whether this signaling system is present in human testis remains largely unexplored. In this study we report the expression and cellular location of ghrelin and its functional receptor, the type 1a GHS-R, in adult human testis. In addition, evaluation of ghrelin and GHS-R1a immunoreactivity in testicular tumors and dysge- netic tissue is presented. The expression of the mRNAs en- coding ghrelin and GHS-R1a was demonstrated in human tes- tis specimens by RT-PCR, followed by direct sequencing. In normal testis, ghrelin immunostaining was demonstrated in interstitial Leydig cells and, at lower intensity, in Sertoli cells within the seminiferous tubules. In contrast, ghrelin was not detected in germ cells at any stage of spermatogenesis. The cognate ghrelin receptor showed a wider pattern of cellular distribution, with detectable GHS-R1a protein in germ cells, mainly in pachytene spermatocytes, as well as in somatic Ser- toli and Leydig cells. Ghrelin immunoreactivity was absent in poorly differentiated Leydig cell tumor, which retained the expression of GHS-R1a peptide. In contrast, highly differen- tiated Leydig cell tumors expressed both the ligand and the receptor. The expression of ghrelin and GHS-R1a was also detected in dysgenetic Sertoli cell-only seminiferous tubules, whereas germ cell tumors (seminoma and embryonal carci- noma) were negative for ghrelin and were weakly positive for GHS-R1a. In conclusion, our results demonstrate that ghrelin and the type 1a GHS-R are expressed in adult human testis and testicular tumors. Overall, the expression of ghrelin and its functional receptor in human and rat testis, with roughly similar patterns of cellular distribution, is highly suggestive of a conserved role for this newly discovered molecule in the regulation of mammalian testicular function. (J Clin Endo- crinol Metab 89: 400–409, 2004) G HRELIN HAS BEEN recently identified as the en- dogenous ligand for the GH secretagogue receptor (GHS-R) (1, 2). Ghrelin is a 28-amino acid peptide in which the serine 3 residue is n-octanoylated, a modification es- sential for its endocrine activity (1). In addition, a second ligand for the GHS-R, termed des-Gln 14 -ghrelin, has been described, whose biological activity and sequence are identical to those of ghrelin, except for one glutamine in position 14 (3). As expected for the endogenous counter- part of GHSs, this molecule has been proven to elicit GH secretion in different species, including humans (1, 2, 4 – 6). In addition, ghrelin is able to induce food intake and adiposity in rodents (6 – 8), and its involvement in the long-term control of body weight in humans has been recently proposed (9). Moreover, emerging evidence in- dicates that ghrelin exhibits an array of additional endo- crine and nonendocrine biological actions, which include neuroendocrine modulation of lactotropic, corticotropic, and gonadotropic axes, as well as metabolic, cardiovas- cular, and antiproliferative effects (2, 5, 10 –12). The biological actions of ghrelin are mostly conducted through interaction with its specific cell surface receptor, namely the GHS-R. The cognate ghrelin receptor belongs to the large family of G protein-coupled, seven-transmembrane domain receptors (13, 14). This receptor is highly expressed at central neuroendocrine tissues such as the pituitary and hypothalamus (15). Two GHS-R subtypes, generated by al- ternative splicing of a single gene, have been described: the full-length type 1a receptor and the truncated GHS-R type 1b (13, 14). The GHS-R1a is the functionally active, signal-trans- ducing form of the receptor. In contrast, the GHS-R1b lacks transmembrane domains 6 and 7, and it is apparently devoid of high affinity ligand binding and signal transduction ca- pacity (13). Thus, its functional role, if any, remains unclear. In addition, evidence for GHS-R-independent biological ac- Abbreviations: AMV-RT, Avian myeloblastosis virus reverse tran- scriptase; GHS-R, GH secretagogue receptor; HPRT, hypoxanthine gua- nine phosphoribosyl transferase; SCF, stem cell factor; SCO, Sertoli cell-only syndrome. JCEM is published monthly by The Endocrine Society (http://www. endo-society.org), the foremost professional society serving the en- docrine community. 0021-972X/04/$15.00/0 The Journal of Clinical Endocrinology & Metabolism 89(1):400 – 409 Printed in U.S.A. Copyright © 2004 by The Endocrine Society doi: 10.1210/jc.2003-031375 400

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Expression of Ghrelin and Its Functional Receptor, theType 1a Growth Hormone Secretagogue Receptor, inNormal Human Testis and Testicular Tumors
F. GAYTAN, M. L. BARREIRO, J. E. CAMINOS, L. K. CHOPIN, A. C. HERINGTON, C. MORALES,L. PINILLA, R. PANIAGUA, M. NISTAL, F. F. CASANUEVA, E. AGUILAR, C. DIEGUEZ, AND
M. TENA-SEMPERE
Departments of Cell Biology, Physiology, and Immunology (F.G., M.L.B., L.P., E.A., M.T.-S.), and Pathology (C.M.),University of Cordoba, 14004 Cordoba, Spain; Departments of Physiology (J.E.C., C.D.) and Medicine (F.F.C.), University ofSantiago de Compostela, 15705 Santiago de Compostela, Spain; Center for Molecular Biotechnology, Queensland Universityof Technology (L.K.C., A.C.H.), Brisbane, Queensland, Australia; Department of Cell Biology and Genetics, University ofAlcala (R.P.), 28871 Madrid, Spain; and Department of Morphology, University Autonoma (M.N.), 28029 Madrid, Spain
Ghrelin, the endogenous ligand for the GH secretagogue re-ceptor (GHS-R), has been primarily linked to the central neu-roendocrine regulation of GH secretion and food intake, al-though additional peripheral actions of ghrelin have alsobeen reported. In this context, the expression of ghrelin andits cognate receptor has been recently demonstrated in rattestis, suggesting a role for this molecule in the direct controlof male gonadal function. However, whether this signalingsystem is present in human testis remains largely unexplored.In this study we report the expression and cellular location ofghrelin and its functional receptor, the type 1a GHS-R, inadult human testis. In addition, evaluation of ghrelin andGHS-R1a immunoreactivity in testicular tumors and dysge-netic tissue is presented. The expression of the mRNAs en-coding ghrelin and GHS-R1a was demonstrated in human tes-tis specimens by RT-PCR, followed by direct sequencing. Innormal testis, ghrelin immunostaining was demonstrated ininterstitial Leydig cells and, at lower intensity, in Sertoli cellswithin the seminiferous tubules. In contrast, ghrelin was notdetected in germ cells at any stage of spermatogenesis. The
cognate ghrelin receptor showed a wider pattern of cellulardistribution, with detectable GHS-R1a protein in germ cells,mainly in pachytene spermatocytes, as well as in somatic Ser-toli and Leydig cells. Ghrelin immunoreactivity was absent inpoorly differentiated Leydig cell tumor, which retained theexpression of GHS-R1a peptide. In contrast, highly differen-tiated Leydig cell tumors expressed both the ligand and thereceptor. The expression of ghrelin and GHS-R1a was alsodetected in dysgenetic Sertoli cell-only seminiferous tubules,whereas germ cell tumors (seminoma and embryonal carci-noma) were negative for ghrelin and were weakly positive forGHS-R1a. In conclusion, our results demonstrate that ghrelinand the type 1a GHS-R are expressed in adult human testisand testicular tumors. Overall, the expression of ghrelin andits functional receptor in human and rat testis, with roughlysimilar patterns of cellular distribution, is highly suggestiveof a conserved role for this newly discovered molecule in theregulation of mammalian testicular function. (J Clin Endo-crinol Metab 89: 400–409, 2004)
GHRELIN HAS BEEN recently identified as the en-dogenous ligand for the GH secretagogue receptor
(GHS-R) (1, 2). Ghrelin is a 28-amino acid peptide in whichthe serine 3 residue is n-octanoylated, a modification es-sential for its endocrine activity (1). In addition, a secondligand for the GHS-R, termed des-Gln14-ghrelin, has beendescribed, whose biological activity and sequence areidentical to those of ghrelin, except for one glutamine inposition 14 (3). As expected for the endogenous counter-part of GHSs, this molecule has been proven to elicit GHsecretion in different species, including humans (1, 2, 4 – 6).In addition, ghrelin is able to induce food intake andadiposity in rodents (6 – 8), and its involvement in thelong-term control of body weight in humans has been
recently proposed (9). Moreover, emerging evidence in-dicates that ghrelin exhibits an array of additional endo-crine and nonendocrine biological actions, which includeneuroendocrine modulation of lactotropic, corticotropic,and gonadotropic axes, as well as metabolic, cardiovas-cular, and antiproliferative effects (2, 5, 10 –12).
The biological actions of ghrelin are mostly conductedthrough interaction with its specific cell surface receptor,namely the GHS-R. The cognate ghrelin receptor belongs tothe large family of G protein-coupled, seven-transmembranedomain receptors (13, 14). This receptor is highly expressedat central neuroendocrine tissues such as the pituitary andhypothalamus (15). Two GHS-R subtypes, generated by al-ternative splicing of a single gene, have been described: thefull-length type 1a receptor and the truncated GHS-R type 1b(13, 14). The GHS-R1a is the functionally active, signal-trans-ducing form of the receptor. In contrast, the GHS-R1b lackstransmembrane domains 6 and 7, and it is apparently devoidof high affinity ligand binding and signal transduction ca-pacity (13). Thus, its functional role, if any, remains unclear.In addition, evidence for GHS-R-independent biological ac-
Abbreviations: AMV-RT, Avian myeloblastosis virus reverse tran-scriptase; GHS-R, GH secretagogue receptor; HPRT, hypoxanthine gua-nine phosphoribosyl transferase; SCF, stem cell factor; SCO, Sertolicell-only syndrome.JCEM is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving the en-docrine community.
0021-972X/04/$15.00/0 The Journal of Clinical Endocrinology & Metabolism 89(1):400–409Printed in U.S.A. Copyright © 2004 by The Endocrine Society
doi: 10.1210/jc.2003-031375
400

tions of ghrelin as well as of synthetic GHSs has been pre-sented recently (16, 17).
Despite the fact that most of the biological actions ofghrelin are carried out centrally, additional peripheral ac-tions of ghrelin have recently emerged. Among those, a rolefor this molecule in the direct control of rat testicular functionhas been suggested. Thus, we have previously reported theexpression of the ghrelin gene and protein in interstitial ratLeydig cells under the control of pituitary LH (18, 19). Sim-ilarly, expression of the cognate ghrelin receptor in rat testishas been demonstrated (18, 20), and evidence of the abilityof ghrelin to modulate stimulated testosterone secretion invitro has been presented (18). To our knowledge, however,detailed evaluation of the pattern of expression of ghrelinand its functional receptor in testes from nonrodent species,including humans, has not been conducted, although detec-tion of ghrelin mRNA in human testicular tissue has beenbriefly reported recently (21). In this context, in the presentstudy we evaluated the expression and pattern of cellulardistribution of ghrelin and its functional receptor, i.e. theGHS-R1a, in adult human testis. In addition, considering itsproposed effects on the proliferative rate of different tumorcell lines (12, 22), ghrelin and GHS-R1a immunoreactivitywas also evaluated in different human testicular tumors anddysgenetic testis tissue.
Materials and MethodsTissue samples
Tissue sections from adult human testes, testicular tumors, and dys-genetic testis tissue were obtained from the archives of Department ofPathology, University of Cordoba (Cordoba, Spain), and Department ofMorphology, University Autonoma (Madrid, Spain), upon approval ofthe respective local ethical committees. In detail, from a larger series,sections of five normal testicular specimens were obtained from adultpatients undergoing therapeutic orchidectomy unrelated to testicularpathology. In these samples, normal spermatogenesis and homogenoushistological appearance were used as indexes to define tissue lacking anypathological condition, following the criteria described previously (23).In addition, representative tissue sections of several testicular disorderswere obtained from archival samples. These were previously diagnosedspecimens that included germ cell aplasia [Sertoli cell-only syndrome
(SCO); two specimens], stromal cell-derived tumors (Leydig cell tumors;three specimens), and germ cell-derived tumors (seminoma and em-bryonal carcinoma; three and two specimens, respectively). A compi-lation of normal and pathological testicular samples used in the presentstudy is included in Table 1.
RNA analysis by RT-PCR
Total RNA was isolated from adult human testes (BD Biosciences,Palo Alto, CA) using the single-step, acid guanidinium thiocyanate-phenol-chloroform extraction method (24). Human testes were obtainedfrom apparently healthy individuals without history of testicular pa-thology and hormonal treatment, who had suffered sudden death. Aspositive controls, total RNA samples were obtained from human stom-ach and brain tissues (BD Biosciences). The expression of ghrelin andtype 1a GHS-R mRNAs was assessed by RT-PCR using the primer pairsindicated in Table 2. These sets of primers were synthesized accordingto the published cDNA sequences of human ghrelin and GHS-R (1, 13).Amplification of human ghrelin cDNA was conducted using a previ-ously reported primer pair (22), and the expression of the mRNA en-coding GHS-R1a was assessed in human samples using a type 1a-specific primer pair spanning the single intron of the GHS-R gene, thusallowing amplification of a 205-bp fragment of GHS-R cDNA unique tothe 1a form (13). In addition, as an internal control, amplification of a285-bp fragment of hypoxanthine guanine phosphoribosyl transferase(HPRT) mRNA or a 232-bp fragment of �-actin mRNA was carried outin parallel in each sample using the primer pair and conditions indicatedin Table 2.
For amplification of the targets, RT and PCR were run in two separatesteps. In addition, PCR amplification of specific targets (ghrelin andGHS-R1a) and internal controls (HPRT and �-actin) was carried out inseparate reactions with different numbers of cycles, but using similaramounts of the corresponding cDNA templates, generated in single RTreactions, as previously described (18–20). Briefly, for analysis of ghrelinmRNA expression, 4 �g total testicular RNA were heat-denaturized andreverse transcribed by incubation at 42 C for 90 min with 12.5 U avianmyeloblastosis virus reverse transcriptase (AMV-RT; Promega, Madi-son, WI), 20 U of the ribonuclease inhibitor RNasin (Promega), 200 nmdeoxy-NTP mixture, and 500 ng random hexamers (Promega) in a finalvolume of 30 �l 1� AMV-RT buffer. For analysis of GHS-R1a transcript,RT reactions were conducted similarly, but using 1 nm specific antisenseprimer GHS-R1a-RT (5�-CCCAGAAGTCTGAACACTGCCACC-3�).PCR reactions were conducted using 10-�l aliquots of the cDNA sam-ples, subjected to amplification in 50 �l 1� PCR buffer in the presenceof 2.5 U Taq-DNA polymerase (Promega), 200 nm deoxy-NTPs mixture,and 1 nm of the appropriate primer pairs. PCR reactions consisted of aninitial denaturing cycle at 97 C for 5 min, followed by a number of cyclesof amplification defined by denaturation at 96 C for 30 sec, annealing for
TABLE 1. Compilation of normal and pathological human testis samples, and semiquantitative expression data of ghrelin andGHS-R1a
Specimens Age (yr) No. ofsamples Ghrelin-IR GHS-R1a-IR Ghrelin mRNA GHS-R1a mRNA
Normal testisAdult testis 43–65 5 Leydig cells: ��� Leydig cells: ���
Sertoli cells:�� Sertoli cells:�� Detectable DetectableGerm cells: �� Germ cells: ��
Testicular disordersLeydig cell tumors 29–47 3
Differentiated Tumor cells: �� Tumor cells: �� NT NTPoorly differentiated Tumor cells: �� Tumor cells: �� NT NT
Germ cell aplasia 33–49 2SCO syndrome Leydig cells: ��� Leydig cells �� NT NT
Sertoli cells: �� Sertoli cells: �Germ cell tumors 31–51 5
Seminoma Tumor cells: �� Tumor cells: �� NT NTEmbryonal carcinoma Tumor cells: �� Tumor cells: �� NT NT
Intensity of ghrelin and GHS-R1a immunostaining was estimated on a semiquantitative basis, using a 0–3 scale: ���, strongly positive;��, clearly positive; �, weakly positive; ��, negative. In tumor samples (Leydig cell and germ cell tumors), remnants of testicular tissue outsidethe tumor area (free of tumor) showed a pattern of cellular immunostaining for ghrelin and GHS-R1a similar to that of control samples, andthis is not indicated in the table. IR, Immunoreactivity; NT, not tested.
Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis J Clin Endocrinol Metab, January 2004, 89(1):400–409 401

30 sec, and extension at 72 C for 1 min. A final extension cycle of 72 Cfor 15 min was included. The annealing temperature was adjusted foreach target: 59 C for ghrelin and GHS-R1a subtype, and 56 C for HPRTand �-actin transcripts. For qualitative analysis, 38 (ghrelin and GHS-R1a) and 30 (HPRT and �-actin) PCR amplification cycles were used.
PCR-generated DNA fragments were resolved in Tris-borate-buff-ered 1.5% agarose gels and were visualized by ethidium bromide stain-ing. The specificity of PCR products was confirmed by direct sequencingusing a fluorescent dye termination reaction and an automated se-quencer (Central Sequencing Service, University of Cordoba). In allassays, reactions without RT were included that yielded negative am-plification, thus ruling out the possibility of spurious amplification of thesignals.
Immunohistochemistry and polyclonal antighrelin and anti-GHS-R1a antibodies
For analysis of ghrelin peptide expression, a rabbit antighrelin poly-clonal antiserum, provided by Drs. Kojima and Kangawa (NationalCardiovascular Center Research Institute, Osaka, Japan), was used asprimary antibody. This antibody was generated as described in detailpreviously (25), using [Cys0]rat ghrelin (13–28) as antigen. This anti-serum is able to recognize both rat and human ghrelin (26), and it isdevoid of significant cross-reactivity with other peptides, as reportedpreviously (26). In addition, immunohistochemical labeling of GHS-R1aprotein was conducted using a rabbit polyclonal antibody generatedagainst a synthetic peptide corresponding to the C-terminal fragment(RAWTESSINTC) of the human GHS-R1a protein conjugated to diph-theria toxin (Mimotopes, Melbourne, Australia), as described in detailpreviously (22). Western analyses using this antibody demonstrated asingle specific band of approximately the predicted size (45 kDa) for theGHS-R1a in the ALVA-41 and DU145 prostate cancer cell lines (Chopin,L. K., and A. C. Herington, unpublished observation), which have beenproven to express the GHS-R1a mRNA isoform and GHS-R1a protein(22).
Immunohistochemistry was performed on routinely neutral-buff-ered, formaldehyde-fixed, paraffin-embedded tissues. In detail, testic-ular sections (5 �m thick) were placed on poly-l-lysine-coated slides,and after dewaxing in xylene and rehydration in ethanol, the sampleswere incubated in 2% hydrogen peroxide in methanol for 30 min toquench endogenous peroxidase, followed by washing in PBS. In addi-tion, sections for GHS-R1a immunolabeling were immersed in 10 mmcitrate buffer and submitted to antigen retrieval in a microwave oven(twice, 5 min each time, 700 watts). According to routine immunohis-tochemical procedure, sections were allowed to cool at room tempera-ture, washed in PBS, blocked with normal serum, and incubatedovernight with the primary antibody, antighrelin (diluted 1:600) oranti-GHS-R1a (diluted 1:10). The sections were then processed accordingto the avidin-biotin-peroxidase complex technique following previouslydescribed methods (19, 27). Negative controls were run routinely inparallel by replacing the primary antibody with preimmune serum orPBS. In addition, positive controls for ghrelin and GHS-R1a immuno-staining were assayed. These included reactions in rat testicular samplesand human ovary and pituitary sections, conducted using antighrelin
and anti-GHS-R1a primary antibodies, respectively. These yieldedstrong immunoreactivity, in line with our previous findings (18–20, 28).As an additional control for the specificity of GHS-R1a antibody, im-munohistochemical reactions were carried out in human pituitary andtesticular tissues after preabsorption of the antiserum overnight at 4 Cwith 1 mg/ml of the synthetic peptide (RAWTESSINTC) against whichit was raised. In keeping with our previous results (28), this procedurecompletely abolished immunolabeling of pituitary and testicular sec-tions (data not shown).
ResultsGhrelin expression in normal human testis
Expression of the ghrelin gene in normal adult humantestis was demonstrated by means of primer-specific RT-PCR(Fig. 1). Thus, RT-PCR reactions from human testicular RNA,using a primer pair designed to amplify a 264-bp region ofpreproghrelin mRNA transcript, resulted in the generation ofa single amplicon of expected size; its identity was confirmedby direct sequencing. In our assays, the possibility of spu-rious amplification of the signal was ruled out by the lack ofamplification in reactions without RT. For comparative pur-poses, RT-PCR amplification of ghrelin mRNA was also con-ducted in RNA samples from human stomach, the majorsource of systemic ghrelin (1). These reactions yielded strongamplification signals.
In addition, the pattern of cellular distribution of ghrelinprotein in normal adult human testis was analyzed by im-
FIG. 1. Expression of the gene encoding ghrelin in adult humantestis. A representative RT-PCR assay of ghrelin mRNA in a normalhuman testis sample (Te) is presented. A single amplicon of theexpected 264-bp size was obtained, whose identity was confirmed bydirect sequencing. In addition, positive [human stomach (St)] andnegative [without AMV-RT (�)] controls are shown. Amplification ofHPRT transcript served as an internal control.
TABLE 2. Oligo-primer pairs used for RT-PCR amplification of ghrelin, isoform-specific GHS-R1a, HPRT, and �-actin transcripts inhuman testis samples
Target Oligo-primers Expected size (bp) PCR cycles
Ghrelin Ghrelin sense (5�-GAG GAT GAA CTG GAA GTC CG-3�) 264 38Ghrelin as (5�-CAT TTA TTC GCC TCC TGA GC-3�)
GHS-R type1a GHS-R1a sense (5�-CTG CGC TCA GGG ACC AGA ACC A-3�) 205 38GHS-R1a as (5�-GTT GAT GGC AGC ACT GAG GTA GAA-3�)
HPRT HPRT sense (5�-CAG CCC TGC CGT CGT GAT TA-3�) 285 30HPRT as (5�-AGC AAG ACG TTC AGT CCT GTC-3�)
�-Actin �-actin sense (5�-CTG TCT GGC GGC ACC ACC AT-3�) 232 30�-actin as (5�-GCA ACT AAG TCA TAG TCC GC-3�)
For each target, the primer pair used for amplification is included. In addition, the expected size of the generated cDNA products and thenumber of cycles selected for RT-PCR analysis is indicated for each signal. as, Antisense.
402 J Clin Endocrinol Metab, January 2004, 89(1):400–409 Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis

munohistochemistry, using a well characterized specific an-tighrelin polyclonal antibody (25, 26). This antiserum hasbeen previously used by our group for the immunodetectionof ghrelin peptide in rat male and female gonads as well asin human ovary (18–20, 28, 29). In keeping with our RNAdata, immunohistochemical assays demonstrated the pres-ence of ghrelin peptide in normal human testicular tissue(Fig. 2, A and B). The pattern of cellular distribution ofghrelin immunoreactivity was highly conserved within thecontrol group, with almost negligible variation among sam-ples. Strong ghrelin immunostaining was detected in steroi-dogenic Leydig cells within the testicular interstitium. Inaddition, ghrelin immunoreactivity was observed, albeit atlower intensity, in seminiferous tubules, where somatic Ser-toli cells showed a specific ghrelin signal. In contrast, nostaining was detected in germ cells at any stage of spermat-ogenesis. Similarly, interstitial cell types other than Leydigcells failed to show specific ghrelin immunoreactivity. Inpositive cells, the cytoplasm was uniformly stained, whereascell nuclei remained negative. As a control for specificity ofdetection, omission of primary antighrelin antibody and itssubstitution by either PBS or preimmune serum resulted innegative staining of Leydig and Sertoli cells (data notshown).
Ghrelin expression in pathological testicular samples
The pattern of cellular expression of ghrelin protein wasalso assessed in representative tissue samples of differenttesticular disorders. Samples of germ cell aplasia (SCO), stro-mal-cell derived tumors (Leydig cell tumors), and germ cell-derived tumors (seminoma and embryonal carcinoma) wereanalyzed. SCO testes were considered as those where germcells were absent from the seminiferous tubules. Amongthese, hypoplastic tubules were defined as those bearingdysgenetic Sertoli cells, following previously described cri-teria (30). As was the case in normal testis tissue, strongghrelin immunolabeling was detected in discernible Leydigcells within the interstitial space of SCO testes. In addition,ghrelin immunoreactivity was observed at lower intensity indysgenetic Sertoli cells of hypoplastic tubules (Fig. 2C).
Ghrelin immunohistochemistry was also conducted inrepresentative Leydig cell tumors with distinct morpholog-ical patterns of cell differentiation. Poorly differentiated Ley-dig tumor cells, characterized by small cell volume andscarce cytoplasm, failed to show discernible ghrelin immu-noreactivity, whereas interstitial Leydig cells in the adjacentnormal tissue presented a strong ghrelin signal (Fig. 2, D andE). In contrast, Leydig cell tumors with a higher degree of celldifferentiation, defined by a large vacuolated cytoplasm, ex-hibited specific ghrelin immunostaining, albeit at lower in-tensity than normal Leydig cells (Fig. 2, F and G). Finally,analysis of ghrelin peptide expression was also carried out ingerm cell-derived tumors. Our analysis failed to demonstratedetectable ghrelin signals in sections from testicular semi-noma and embryonal carcinoma (Fig. 2, H and I) despiteclear-cut ghrelin immunoreactivity detected in remnants ofnormal testicular tissue in these samples.
A comprehensive compilation of ghrelin expression data
in normal and pathological human testis samples is includedin Table 1.
GHS-R expression in normal human testis
In addition to the ligand, expression of the mRNA encod-ing the functional form of the cognate ghrelin receptor, thetype 1a GHS-R, was assessed in normal adult human testisby primer-specific RT-PCR. Our molecular analysis demon-strated that, as was the case for ghrelin, GHS-R1a mRNA isexpressed in adult human testis (Fig. 3). Thus, RT-PCR re-actions from human testicular RNA, using a type 1a-specificprimer pair spanning the single intron of GHS-R gene anddesigned to amplify a 205-bp area unique to the 1a form (13),generated a single amplicon of expected size; its identity wasconfirmed by direct sequencing. The possibility of spuriousamplification of the signal was excluded by the lack of am-plification in reactions without RT. As a positive control,RT-PCR amplification of GHS-R1a mRNA was also con-ducted in human brain samples.
The pattern of cellular expression of GHS-R1a protein wasassessed in normal adult human testis by means of immu-nohistochemistry, using a previously characterized, specificanti-GHS-R1a polyclonal antibody (22). This antiserum hasbeen previously used by our group for the immunodetectionof GHS-R peptide in rat testis and human ovary (20, 28). Aswas the case for ghrelin, our analyses demonstrated thepresence of GHS-R1a protein in normal human testiculartissue (Fig. 4, A–C). Also, as reported for the ligand, thepattern of cellular location of GHS-R1a immunoreactivitywas highly conserved within the control group, with almostnegligible variation among samples. However, comparedwith that of ghrelin, GHS-R1a protein showed a somewhatwider pattern of cellular distribution within the human testis.Thus, clear-cut GHS-R1a immunostaining was detected insteroidogenic Leydig cells within the testicular interstitium,whereas other interstitial cell types were negative. In addi-tion, GHS-R1a immunoreactivity was detected in the semi-niferous tubules, where somatic Sertoli cells showed specificGHS-R1a signal. In addition, GHS-R1a immunolabeling wasobserved in germ cells, mainly in pachytene spermatocytesthat showed strong GHS-R1a-specific staining (Fig. 4C). Asa control for specificity of detection, substitution of the pri-mary anti-GHS-R1a antibody with preimmune serum or itsimmunoabsorption with the synthetic peptide (RAWTESS-INTC) against which it was raised resulted in negative stain-ing of testicular tissue (data not shown).
GHS-R expression in pathological testicular samples
In addition to ghrelin, the pattern of cellular distributionof GHS-R1a protein was evaluated in different testiculardisorders, namely germ cell aplasia (SCO), Leydig cell tu-mors, and germ cell-derived tumors (seminoma and embry-onal carcinoma). As was the case in normal testis tissue,intense GHS-R1a immunostaining was detected in discern-ible Leydig cells within the interstitial space of SCO testes.However, in contrast to normal tubules (see Fig. 4, A and B),GHS-R1a immunoreactivity was observed at negligible lev-els in dysgenetic Sertoli cells of hypoplastic tubules despiteclear GHS-R1a signals in adjacent interstitial Leydig cells
Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis J Clin Endocrinol Metab, January 2004, 89(1):400–409 403
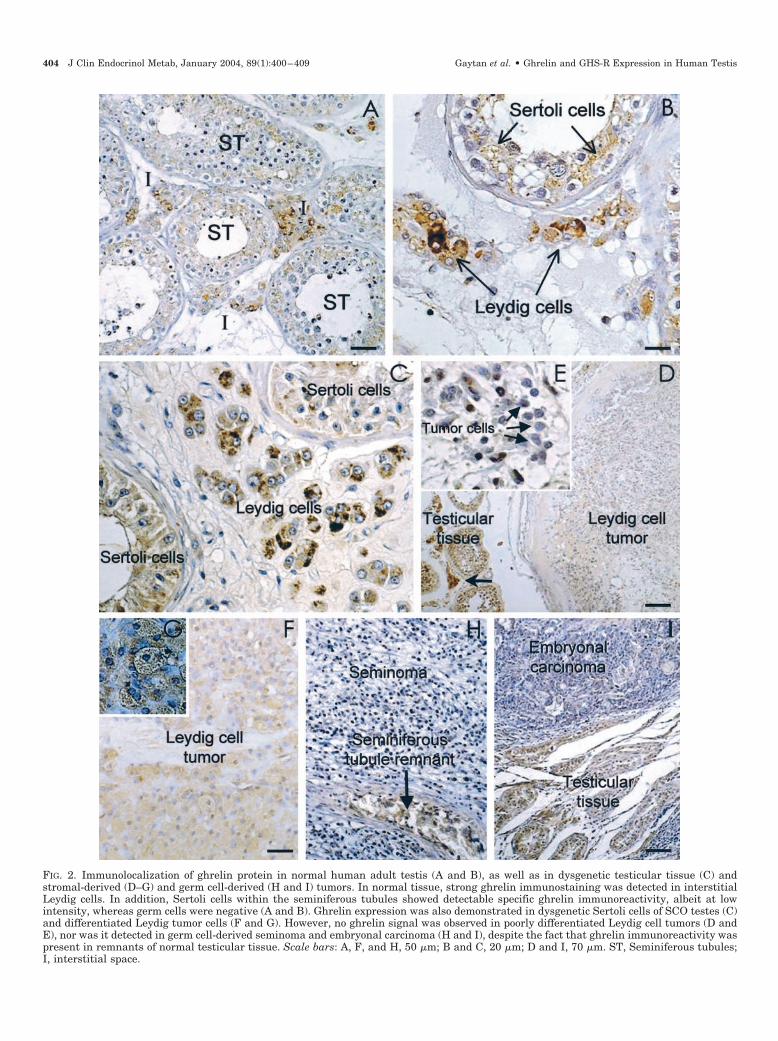
FIG. 2. Immunolocalization of ghrelin protein in normal human adult testis (A and B), as well as in dysgenetic testicular tissue (C) andstromal-derived (D–G) and germ cell-derived (H and I) tumors. In normal tissue, strong ghrelin immunostaining was detected in interstitialLeydig cells. In addition, Sertoli cells within the seminiferous tubules showed detectable specific ghrelin immunoreactivity, albeit at lowintensity, whereas germ cells were negative (A and B). Ghrelin expression was also demonstrated in dysgenetic Sertoli cells of SCO testes (C)and differentiated Leydig tumor cells (F and G). However, no ghrelin signal was observed in poorly differentiated Leydig cell tumors (D andE), nor was it detected in germ cell-derived seminoma and embryonal carcinoma (H and I), despite the fact that ghrelin immunoreactivity waspresent in remnants of normal testicular tissue. Scale bars: A, F, and H, 50 �m; B and C, 20 �m; D and I, 70 �m. ST, Seminiferous tubules;I, interstitial space.
404 J Clin Endocrinol Metab, January 2004, 89(1):400–409 Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis

(Fig. 4D). In addition, strong GHS-R1a immunolabeling wasdetected in Leydig cell tumors regardless of their morpho-logical patterns of cell differentiation. Thus, both poorly dif-ferentiated Leydig tumor cells, characterized by small cellvolume and scarce cytoplasm (Fig. 4E), and highly differen-tiated Leydig cell tumors (Fig. 4, F and G) showed specificGHS-R1a signals. Finally, analysis of GHS-R1a peptide dis-tribution was conducted in germ cell-derived tumors. De-spite the lack of ghrelin expression in these tumors, ourassays demonstrated detectable GHS-R1a immunoreactivity,albeit at low intensity, in sections from testicular seminomaand embryonal carcinoma (Fig. 4, H and I).
A comprehensive compilation of GHS-R1a expressiondata in normal and pathological human testis samples isincluded in Table 1.
Discussion
In the present communication we have used molecularand immunohistochemical approaches to characterize theexpression and pattern of cellular location of ghrelin and thefunctional GHS-R1a subtype in the adult human testis. Im-portantly, a role for ghrelin as a putative regulator of rodentgonadal function has been recently suggested. Thus, a spe-cific ghrelin gene-derived transcript has been identified inmouse testis (31). Moreover, our group has provided evi-dence for the expression of ghrelin and its receptor in rattestis (18–20), and ghrelin expression has been recently dem-onstrated in rat and human ovary (28, 29). Yet, the presenceof ghrelin and its cognate functional receptor in human tes-ticular tissue remained largely unexplored. Our present re-sults indicate that both components (ligand and receptor) ofthe ghrelin signaling system are present in the human testis,with patterns of cellular distribution largely similar to thosein the rat testis (20).
Using real-time RT-PCR, the expression of ghrelin gene inhuman testis has been postulated very recently (21). Ourpresent data further substantiate this initial observation anddemonstrate for the first time the actual presence of ghrelinpeptide in the human male gonad. Immunolocalization ofghrelin within human testis tissue indicated that this proteinis strongly expressed in interstitial mature Leydig cells. No-tably, our previous molecular and immunohistochemical
analyses in the rat indicated that testicular expression ofghrelin is restricted to mature Leydig cells (18, 19). Moreover,in the human ovary, intense ghrelin immunoreactivity hasbeen demonstrated in hilus interstitial cells (28), i.e. a cell typethat shows distinctive morphological characteristics, such asthe presence of crystals of Reinke, identical to those of dif-ferentiated testicular Leydig cells (32). Thus, the expressionof ghrelin is apparently a characteristic feature of both rodentand human Leydig cells that may serve as a useful markerof Leydig cell differentiation in mammalian testis, as previ-ously proposed for other signals, such as relaxin-like factor(33). Interestingly, Leydig cell-specific expression of ghrelinin rat testis is under regulation of pituitary LH (19), and a rolefor ghrelin as a direct modulator of LH-driven testiculartestosterone secretion has been reported (18). Furthermore,ghrelin expression in human ovary has been demonstratedin steroidogenically active luteal and hilus cells (28). In thiscontext, elucidation of the potential direct role of ghrelin inthe regulation of androgen secretion in the human testismerits further investigation.
Despite clear similarities in the pattern of expression ofghrelin in rat and human testicular Leydig cells, a specificfeature of ghrelin expression in the human testis is the pres-ence of this peptide in Sertoli cells. In fact, ghrelin protein inrat testis was solely detected in interstitial Leydig cells, andseminiferous tubules did not show any significant ghrelinimmunoreactivity (18, 19). The basis for such a species di-vergence remains obscure. In this sense, the promoter of thehuman ghrelin gene has been recently cloned, and it wasshown to be activated by cAMP (34). Notably, cAMP is themajor intracellular second messenger in Sertoli cells for aplethora of paracrine and endocrine regulators, includingpituitary FSH (35). This phenomenon might also be the basisfor ghrelin expression in interstitial Leydig cells that usecAMP as intracellular messenger for its major endocrineregulator, namely pituitary LH (35). In any case, given thepresence of the cognate ghrelin receptor in different tubularcell types reported herein, it is possible that local expressionof ghrelin, in both Leydig and Sertoli cells may play a role inthe paracrine control of seminiferous tubule function. In thiscontext, it is noticeable that ghrelin apparently regulatesexpression of the stem cell factor (SCF) gene in seminiferoustubules of the rat testis (our unpublished observation). SCFhas been pointed out as the major paracrine stimulator ofgerm cell development, acting as a survival factor for sper-matogonia, spermatocytes, and spermatids in the adult ratseminiferous epithelium (36, 37). Whether ghrelin is able tomodulate SCF gene expression, or it conducts additionalbiological functions in human seminiferous tubules remainsto be evaluated.
In addition to the ligand, our immunohistochemical anal-yses of the presence and cellular location of GHS-R1a proteinwithin the adult human testis demonstrated a somewhatwider pattern of distribution than that of ghrelin, with de-tectable specific signals in germ cells, mainly in pachytenespermatocytes, as well as in somatic Sertoli and Leydig cells.Partially conflicting results have been reported to date on theactual expression of the functional GHS-R1a in rodent andhuman gonads. In this sense, our recent analyses demon-strated expression of the type 1a GHS-R subtype at the
FIG. 3. Expression of the isoform-specific type 1a GHS-R mRNA inadult human testis. A representative RT-PCR assay of GHS-R1amRNA in a normal human testis sample (Te) is presented. A singleamplicon of the expected 205-bp size was obtained, whose identity wasconfirmed by direct sequencing. In addition, positive [human brain(Br)] and negative [without AMV-RT (�)] controls are shown. Am-plification of �-actin mRNA served as an internal control.
Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis J Clin Endocrinol Metab, January 2004, 89(1):400–409 405

FIG. 4. Immunolocalization of GHS-R type 1a protein in normal human adult testis (A–C) as well as in dysgenetic testicular tissue (D) andstromal-derived (E–G) and germ cell-derived (H and I) tumors. In normal tissue, clear-cut GHS-R1a immunostaining was detected in interstitialLeydig cells (long arrows). In addition, GHS-R1a immunoreactivity was detected in seminiferous tubules, in both Sertoli cells (asterisks) andgerm cells, mainly pachytene spermatocytes (short arrows; B and C). A detail of GHS-R1a labeling in the cytoplasm of pachytene spermatocytesis presented in C. GHS-R1a was almost negligible in dysgenetic Sertoli cells of SCO testes (D) despite the fact that a clear GHS-R1a signal wasdemonstrated in adjacent interstitial Leydig cells. In addition, GHS-R1a peptide was present in Leydig tumor cells regardless of their degreeof cellular differentiation (E–G), whereas germ cell-derived seminoma and embryonal carcinoma showed weak, but detectable, ghrelin im-munostaining (H and I). Scale bars: A, E, and F, 50 �m; B, 20 �m; H and I, 60 �m. TR, Tubule remnant.
406 J Clin Endocrinol Metab, January 2004, 89(1):400–409 Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis

mRNA and peptide levels in rat testis during the adult period(20). In keeping with these data, high levels of GHS/ghrelin-binding sites were demonstrated in human testis (38). Sim-ilarly, ghrelin-binding sites were identified in the humanovary (38), thus suggesting the presence of the functionalreceptor, which was recently confirmed by our immunohis-tochemical analyses (28). In contrast, however, in a recentreport systematic screening of GHS-R1a mRNA expressionin a wide array of human tissues, using real-time RT-PCR,failed to detect the GHS-R1a transcript in the human testis(21). Conversely, positive amplification of the mRNA en-coding the truncated GHS-R1b form was observed (21).These observations are apparently in conflict with ourpresent RT-PCR and immunohistochemical data. The rea-sons for such a discrepancy remain unclear. It has to be noted,however, that changes in the balance of expression between1a and 1b forms of GHS-R may take place in the human testisduring development and under certain endocrine condi-tions. This is certainly the case for the rat testis, wherechanges in the pattern of alternative splicing of GHS-R geneare observed throughout postnatal development; strong ex-pression of GHS-R1a mRNA is detected from puberty on-ward, whereas in earlier stages of testicular development thepredominant receptor form is probably the truncated GHS-R1b type (20). Thus, detailed description of the endocrine anddevelopmental background of the assayed testis samplesmay help to explain the conflicting results indicated above(21). Moreover, pending on the availability of a specific an-tibody, it will be of interest to analyze the cellular expressionof GHS-R1b subtype in human testis. Overall, data fromprevious binding studies (38) and our present immunohis-tochemical analyses indicate that functional GHS-Rs are ex-pressed in the adult human testis. Furthermore, comparisonof the distribution of GHS-R1a protein in rat and humanspecies (Ref. 20 and the present results) is suggestive of ahighly conserved pattern of cellular expression of the func-tional ghrelin receptor in mammalian testis.
Besides analyses in normal testis tissue, immunolocaliza-tion of ghrelin, and GHS-R1a proteins were conducted inrepresentative sections of several types of testicular tumorsand dysgenetic syndrome. It is worthy noting that the ex-pression of ghrelin has been previously demonstrated inseveral endocrine tumors, such as pituitary-, stomach-, andpancreas-derived tumors, i.e. tissues that normally expressghrelin (39–41). In keeping with these observations, ourimmunohistochemical analyses demonstrated that ghrelinexpression is conserved in Leydig cell tumors and dysgeneticSertoli cells. However, the expression of ghrelin in Leydigtumor cells is apparently linked to the degree of cell differ-entiation. Thus, highly differentiated Leydig cell tumorsshowed specific ghrelin immunostaining, albeit at lower in-tensity than normal Leydig cells. In contrast, poorly differ-entiated Leydig tumor cells, characterized by small cell vol-ume and scarce cytoplasm, failed to show any discernibleghrelin immunoreactivity. The mechanisms for such a phe-nomenon have not been directly addressed in our study.However, on the basis of the present findings and our pre-vious data on the expression of ghrelin in fully differentiatedrat Leydig cells (18, 19), it is tempting to propose that ghrelinmay operate as a marker of Leydig cell differentiation, both
during normal development and in tumor transformation.On the latter, loss of ghrelin expression might be an index ofcell dedifferentiation. Moreover, ghrelin and its syntheticcounterparts (GHSs) have been shown to carry out antipro-liferative actions in different tumor cell lines (12, 17). Nota-bly, rat and human mature Leydig cells, which do expressghrelin and its functional receptor, are devoid of significantproliferative activity (42). In contrast, proliferative rat Leydigcell progenitors (19) and poorly differentiated human Leydigtumor cells do not show ghrelin immunoreactivity. On thisbasis, we are presently investigating whether ghrelin mayfunction as an autocrine regulator of Leydig cell proliferationin both normal and tumor conditions.
In addition to ghrelin, analysis of the expression of GHS-R1a peptide in representative testicular tumors was con-ducted. In this sense, the expression of GHS/ghrelin-bindingsites and/or the GHS-R gene has been previously demon-strated in a wide variety of tumors, including pituitary ad-enomas and other neuroendocrine tumors (43), neoplasticthyroid tissue (44), human breast carcinoma (45), prostatecancer cell lines (22), and pancreatic islet cell tumors (40). Ourpresent results indicate that GHS-R1a is expressed in Leydigcell tumors regardless of their degree of cellular differenti-ation as well as in germ cell-derived seminoma and embry-onal carcinoma. Given the proposed regulatory effect of gh-relin and GHSs on cell proliferation in different tumor celllines (12, 17, 46), our present findings provide the basis fora direct action of these compounds on different testiculartumors. It has to be noted, however, that both antiprolifera-tive and proliferative effects of ghrelin and synthetic GHSshave been reported (for a review, see Ref. 46), and that at leastsome of the antiproliferative actions of GHSs are apparentlyconducted through a GHS-R1a-independent pathway (17).
In conclusion, our immunohistochemical analyses providecompelling evidence for the presence of ghrelin and its cog-nate functional receptor, namely the type 1a GHS-R, in theadult human testis. Indeed, the simultaneous expression ofboth components (ligand and receptor) of this signaling sys-tem in different testicular compartments is compatible witha potential action of locally produced ghrelin in the auto-/paracrine regulation of human testis function. Additionally,the wide pattern of GHS-R1a expression in human testismakes it possible that circulating ghrelin may operate uponspecific testicular cell targets, as previously proposed forother peripheral signals with key actions in the regulation ofbody weight and energy expenditure, such as the adipocyte-derived plasma hormone, leptin (47). Moreover, the expres-sion of ghrelin and GHS-R1a was also demonstrated in dif-ferent testicular tumors. The fact that the pattern of cellulardistribution of ghrelin and its receptor is roughly similar inrat and human testes is highly suggestive of a conserved roleof this newly discovered molecule in the regulation of mam-malian testicular function. The relevance of this novel tes-ticular regulatory network in physiological and pathophys-iological conditions merits further investigation.
Acknowledgments
Rabbit antighrelin polyclonal antibody was kindly donated by Drs.M. Kojima and K. Kangawa (Department of Biochemistry, NationalCardiovascular Center Research Institute, Osaka, Japan). We are in-
Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis J Clin Endocrinol Metab, January 2004, 89(1):400–409 407

debted to P. Cano for her excellent technical support in conductingimmunohistochemical analyses. The skillful assistance of E. Tarradaswith the preparation of the photomicrographs is appreciated.
Received August 6, 2003. Accepted October 14, 2003.Address all correspondence and requests for reprints to: Dr. Manuel
Tena-Sempere, Physiology Section, Department of Cell Biology, Phys-iology, and Immunology, Faculty of Medicine, University of Cordoba,Avda. Menendez Pidal s/n, 14004 Cordoba, Spain. E-mail:[email protected].
This work was supported by Grants BFI2000-0419-CO3-03 andBFI2002-00176 from DGESIC (Ministerio de Ciencia y Tecnologıa,Spain), European Union Research Contract EDEN QLK4-CT-2002-00603,and the National Health and Medical Research Council of Australia.
References
1. Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K 1999Ghrelin is a growth-hormone acylated peptide from stomach. Nature 402:656–666
2. Kojima M, Hosoda H, Matsuo H, Kangawa K 2001 Ghrelin: discovery of thenatural endogenous ligand for the growth hormone secretagogue receptor.Trends Endocrinol Metab 12:118–122
3. Hosoda H, Kojima M, Matsuo H, Kangawa K 2000 Purification and charac-terization of rat des-Gln14-ghrelin, a second endogenous ligand for the growthhormone secretagogue receptor. J Biol Chem 275:21995–22000
4. Takaya K, Ariyasu H, Kanamoto N, Iwakura H, Yoshimoto A, Harada M,Mori K, Komatsu Y, Usui T, Shimatsu A, Ogawa Y, Hosoda K, Akamizu T,Kojima M, Kangawa K, Nakao K 2000 Ghrelin strongly stimulates growthhormone release in humans. J Clin Endocrinol Metab 85:4908–4911
5. Arvat E, Maccario M, di Vito L, Broglio F, Benso A, Gottero C, Papotti M,Muccioli G, Dieguez C, Casanueva FF, Deghenghi R, Camanni F, Ghigo E2001 Endocrine activities of ghrelin, a natural growth hormone secretagogue(GHS), in humans: comparison and interactions with hexarelin, a nonnaturalpeptidyl GHS, and GH-releasing hormone. J Clin Endocrinol Metab 86:1169–1174
6. Wren AM, Small CJ, Ward HL, Murphy KG, Dakin CL, Taheri S, KennedyAR, Roberts GH, Morgan DG, Ghatei MA, Bloom SR 2000 The novel hy-pothalamic peptide ghrelin stimulates food intake and growth hormone se-cretion. Endocrinology 141:4325–4328
7. Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H, Wakabayashi I 2000Central effect of ghrelin, an endogenous growth hormone secretagogue, onhypothalamic peptide gene expression. Endocrinology 141:4797–4800
8. Tschop M, Smiley DL, Heiman ML 2000 Ghrelin induces adiposity in rodents.Nature 407:908–913
9. Cummings DE, Weigle DS, Frayo RS, Breen PA, Ma MK, Dellinger EP,Purnell JQ 2002 Plasma ghrelin levels after diet-induced weight loss or gastricbypass surgery. N Engl J Med 346:1623–1630
10. Furuta M, Funabashi T, Kimura F 2001 Intracerebroventricular administrationof ghrelin rapidly suppresses pulsatile luteinizing hormone secretion in ovari-ectomized rats. Biochem Biophys Res Commun 288:780–785
11. Okumura H, Nagaya N, Enomoto M, Nakagawa E, Oya H, Kangawa K 2002Vasodilatory effect of ghrelin, an endogenous peptide from the stomach.J Cardiovasc Pharmacol 39:779–783
12. Broglio F, Arvat E, Benso A, Papotti M, Muccioli G, Deghenghi R, Ghigo E2002 Ghrelin: endocrine and non-endocrine actions. J Pediatr EndocrinolMetab 15:1219–1227
13. Howard AD, Feighner SC, Cully DF, Arena JP, Liberator PA, Rosenblum CI,Hamelin M, Hreniuk DL, Palyha OC, Anderson J, Paress PS, Diaz C, ChouM, Liu KK, McKee KK, Pong SS, Chaung LY, Elbrecht A, Dashkevicz M,Heavens R, Rigby M, Sirinathsinghji DJ, Dean DC, Melillo DG, Patchett AA,Nargund RP, Griffin PR, DeMartino JA, Gupta SK, Schaeffer JM, Smith RG,van der Ploeg LHT 1996 A receptor in pituitary and hypothalamus thatfunctions in growth hormone release. Science 273:974–977
14. McKee KK, Palyha OC, Feighner SD, Hreniuk DL, Tan CP, Phillips MS,Smith RG, van der Ploeg LH, Howard AD 1997 Molecular analysis of ratpituitary and hypothalamic growth hormone secretagogue receptors. MolEndocrinol 11:415–423
15. Guan XH, Yu H, Palyha OC, McKee KK, Feighner SD, Sirinathsinghji DJ,Smith RG, van der Ploeg LH, Howard AD 1997 Distribution of mRNAencoding the growth hormone secretagogue receptor in brain and peripheraltissues. Brain Res Mol Brain Res 48:23–29
16. Baldanzi G, Filigheddu N, Cutrupi S, Catapano F, Bonissoni S, Fubini A,Malan D, Baj G, Granata R, Broglio F, Papotti M, Surico N, Bussolino F,Isgaard J, Deghenghi R, Sinigaglia F, Prat M, Muccioli G, Ghigo E, GrazianiA 2002 Ghrelin and des-acyl ghrelin inhibit cell death in cardiomyocytes andendothelial cells through ERK1/2 and PI 3-kinase/AKT. J Cell Biol 159:1029–1037
17. Ghe C, Cassoni P, Catapano F, Marrocco T, Deghenghi R, Ghigo E, Muccioli
G, Papotti M 2002 The antiproliferative effect of synthetic peptidyl GH secre-tagogues in human CALU-1 lung carcinoma cells. Endocrinology 143:484–491
18. Tena-Sempere M, Barreiro ML, Gonzalez LC, Gaytan F, Zhang FP, CaminosJP, Pinilla L, Casanueva FF, Dieguez C, Aguilar E 2002 Novel expression andfunctional role of ghrelin in rat testis. Endocrinology 143:717–725
19. Barreiro ML, Gaytan F, Caminos JE, Pinilla L, Casanueva FF, Aguilar E,Dieguez C, Tena-Sempere M 2002 Cellular location and hormonal regulationof ghrelin in rat testis. Biol Reprod 67:1768–1776
20. Barreiro ML, Suominen JS, Gaytan F, Pinilla L, Chopin LK, Casanueva FF,Dieguez C, Aguilar E, Toppari J, Tena-Sempere M 2003 Developmental,stage-specific, and hormonally regulated expression of growth hormone secre-tagogue receptor messenger RNA in rat testis. Biol Reprod 68:1631–1640
21. Gnanapavan S, Kola B, Bustin SA, Morris DG, McGee P, Fairclough P,Bhattacharya S, Carpenter R, Grossman AB, Korbonits M 2002 The tissuedistribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R, inhumans. J Clin Endocrinol Metab 87:2988–2991
22. Jeffery PL, Herington AC, Chopin LK 2002 Expression and action of thegrowth hormone releasing peptide ghrelin and its receptor in prostate cancercell lines. J Endocrinol 172:R7–R11
23. Suarez-Quian CA, Martınez-Garcia F, Nistal M, Regadera J 1999 Androgenreceptor distribution in adult human testis. J Clin Endocrinol Metab 84:350–358
24. Chomczynski P, Sacchi N 1987 Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156–159
25. Hosoda H, Kojima M, Matsuo H, Kangawa K 2000 Ghrelin and des-acylghrelin: two major forms of rat ghrelin peptide in gastrointestinal tissue.Biochem Biophys Res Commun 279:909–913
26. Nagaya N, Kojima M, Uematsu M, Yamagishi M, Hosoda H, Oya H, HayashiY, Kangawa K 2001 Hemodynamic and hormonal effects of human ghrelin inhealthy volunteers. Am J Physiol 280:R1483–R1487
27. Garcia-Pardo L, Granados MD, Gaytan F, Padilla CA, Martinez-Galisteo E,Morales C, Sanchez-Criado JE, Barcena JA 1999 Immunolocalization of glu-taredoxin in the human corpus luteum. Mol Hum Reprod 5:914–999
28. Gaytan F, Barreiro ML, Chopin LK, Herington AC, Morales C, Pinilla L,Casanueva FF, Aguilar E, Dieguez C, Tena-Sempere M 2003 Immunolocal-ization of ghrelin and its functional receptor, the type 1a growth hormonesecretagogue receptor, in the cyclic human ovary. J Clin Endocrinol Metab88:879–887
29. Caminos JE, Tena-Sempere M, Gaytan F, Sanchez-Criado JE, Barreiro ML,Nogueiras R, Casanueva FF, Aguilar E, Dieguez C 2003 Expression of ghrelinin the cyclic and pregnant rat ovary. Endocrinology 144:1594–1602
30. Regadera J, Martınez-Garcıa F, Gonzalez-Peramato P, Serrano A, Nistal M,Suarez-Quian C 2001 Androgen receptor expression in Sertoli cells as a func-tion of seminiferous tubule maturation in the human cryptorchid testis. J ClinEndocrinol Metab 86:413–421
31. Tanaka M, Hayashida Y, Nakao N, Nakai N, Nakashima K 2001 Testis-specific and developmentally induced expression of a ghrelin gene-derivedtranscript that encodes a novel polypeptide in the mouse. Biochim BiophysActa 1522:62–65
32. Erickson GF, Magoffin DA, Dyer CA, Hofeditz C 1985 The ovarian androgenproducing cells: a review of structure/function relationships. Endocr Rev6:371–399
33. Balvers M, Spiess A-N, Domagalski R, Hunt N, Kilic E, Mukhopadhyay AK,Hanks E, Charlton HM, Ivell R 1998 Relaxin-like factor expression as markerof differentiation in the mouse testis and ovary. Endocrinology 139:2960–2970
34. Kishimoto M, Okimura Y, Nakata H, Kudo T, Iguchi G, Takahashi Y, KajiH, Chihara K 2003 Cloning and characterization of the 5�-flanking region ofthe human ghrelin gene. Biochem Biophys Res Commun 305:186–192
35. Tena-Sempere M, Huhtaniemi I 2003 Gonadotropins and gonadotropin re-ceptors. In: Fauser BCJM, ed. Reproductive medicine: molecular, cellular andgenetic fundamentals. New York: Parthenon; 225–244
36. Yan W, Linderborg J, Suominen J, Toppari J 1999 Stage-specific regulation ofstem cell factor gene expression in the rat seminiferous epithelium. Endocri-nology 140:1499–1504
37. Yan W, Suominen J, Toppari J 2000 Stem cell factor protects germ cells fromapoptosis in vitro. J Cell Sci 113:161–168
38. Papotti M, Ghe C, Cassoni P, Catapano F, Deghenghi R, Ghigo E, MuccioliG 2000 Growth hormone secretagogue binding sites in peripheral humantissues. J Clin Endocrinol Metab 85:3803–3807
39. Pappoti M, Cassoni P, Volante M, Deghenghi R, Muccioli G, Ghigo E 2001Ghrelin-producing endocrine tumors of the stomach and intestine. J ClinEndocrinol Metab 86:5052–5059
40. Volante M, Allia E, Gugliotta P, Funaro A, Broglio F Deghenghi R, MuccioliG, Ghigo E, Pappoti M 2002 Expression of ghrelin and of the GH secretagoguereceptor by pancreatic islet cells and related endocrine tumors. J Clin Endo-crinol Metab 87:1300–1308
41. Korbonits M, Bustin SA, Kojima M, Jordan S, Adams EF, Lowe DG, Kan-gawa K, Grossman AB 2001 The expression of the growth hormone secret-agogue receptor ligand ghrelin in normal and abnormal human pituitary andother neuroendocrine tumors. J Clin Endocrinol Metab 86:881–887
42. Teerds KJ, de Rooij DG, Rommerts FFG, Wensing CJG 1988 The regulation
408 J Clin Endocrinol Metab, January 2004, 89(1):400–409 Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis

of the proliferation and the differentiation of rat Leydig cell precursors afterEDS administration or daily hCG treatment. J Androl 9:343–351
43. Korbonits M, Jacobs RA, Aylwin SJ, Burrin JM, Dahia PL, Monson JP,Honegger J, Fahlbush R, Trainer PJ, Chew SL, Besser GM, Grossman AB1998 Expression of the growth hormone secretagogue receptor in pituitaryadenomas and other neuroendocrine tumors. J Clin Endocrinol Metab 83:3624–3630
44. Cassoni P, Papotti M, Catapano F, Ghe C, Deghenghi R, Ghigo E, MuccioliG 2000 Specific binding sites for synthetic growth hormone secretagogues innontumoral and neoplastic human thyroid tissue. J Endocrinol 165:139–146
45. Cassoni P, Papotti M, Ghe C, Catapano F, Sapino A, Graziani A, DeghenghiR, Reissmann T, Ghigo E, Muccioli G 2001 Identification, characterization andbiological activity of specific receptors for natural (ghrelin) and syntheticgrowth hormone secretagogues in human breast carcinomas and cell lines.J Clin Endocrinol Metab 86:1738–1745
46. Jeffery PL, Herington AC, Chopin LK 2003 The potential autocrine/paracrineroles of ghrelin and its receptor in hormone-dependent cancer. CytokineGrowth Factor Rev 14:113–122
47. Tena-Sempere M, Barreiro ML 2002 Leptin in male reproduction: the testisparadigm. Mol Cell Endocrinol 188:9–13
JCEM is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
Gaytan et al. • Ghrelin and GHS-R Expression in Human Testis J Clin Endocrinol Metab, January 2004, 89(1):400–409 409
Related Documents











