Expression and modulation of TUB by insulin and thyroid hormone in primary rat and murine 3T3-L1 adipocytes Clare Stretton a,1 , Gary J. Litherland b,1 , Audrey Moynihan a , Eric Hajduch c,d , Harinder S. Hundal a, * a Division of Molecular Physiology, College of Life Sciences, University of Dundee, Dundee DD1 5EH, UK b Institute of Cellular Medicine, The Medical School, Newcastle University, Newcastle NE2 4HH, UK c INSERM U872, Université Pierre et Marie Curie, Paris 6, France d Université Paris Descartes, Paris F-75006, France article info Article history: Received 26 October 2009 Available online 1 November 2009 Keywords: 3T3-L1 Adipogenesis Zucker rat Insulin Tubby abstract tub encodes a protein of poorly understood function, but one implicated strongly in the control of energy balance and insulin sensitivity. Whilst tub expression is particularly prominent in neurones it is also detectable in extraneuronal tissues. We show here, for the first time, expression of TUB protein in rat adi- pocytes and the murine adipocyte model 3T3-L1 and demonstrate that insulin induces its tyrosine phos- phorylation and association with the insulin receptor. TUB expression is regulated developmentally during adipogenic differentiation of 3T3-L1 cells and in response to cell treatment with thyroid hormone or induction of insulin resistance. TUB was upregulated 5- to 10-fold in adipocytes from obese Zucker rats and 3T3-L1 adipocytes that had been rendered insulin resistant, a response that could be antagonised by rosiglitasone, an insulin-sensitising drug. Our data are consistent with a previously unforeseen role for TUB in insulin signalling and fuel homeostasis in adipocytes. Ó 2009 Elsevier Inc. All rights reserved. Introduction TUB and the tubby-like proteins (TULPs) 1–3 comprise a small gene family whose gene sequences are highly conserved across species [5,11,15,21,23]. This suggests that they may serve a basic function within the cell but, as yet, their exact role remains unclear. The four proteins of the family are highly homologous and have over-lapping expression profiles, but gene-inactivation experiments suggest they serve non-redundant roles. tub was originally identified as the gene involved in the tubby mouse syndrome, characterized by late-onset obesity, insulin resistance and sensory impairments [6,22]. This defect is due to expression of a mutant TUB protein in which the C-terminal 44 amino acid residues are replaced by 24 incorrect residues encoded intronical- ly. Gene knockout studies on TUB result in the same phenotype as the tubby mouse suggesting that the phenotype is caused by a loss of function mutation [22]. In neuronal cell lines TUB can be regulated by a range of stimuli including insulin and thyroid hormone and may regulate several functions including vesicle trafficking, gene transcription and en- ergy homeostasis [12,13,28]. In these latter studies TUB was shown to become tyrosine phosphorylated and associate with the insulin receptor following insulin treatment suggesting that it may act as an adaptor protein in insulin signalling [13]. The ‘‘tubby domain” permits interaction of TUB with phosphatidylinositol-4,5-bisphos- phate (PI(4,5)P 2 ) [27] and, as such, this may facilitate its associa- tion/retention with the inner leaflet of the plasma membrane where it may be placed in close proximity to receptor proteins that alter its function through covalent modification. More recent work has indicated that tubby proteins are also secreted from cells raising the suggestion that they may also possess extracellular functions [4]. Although most abundant in neuronal tissue, tub expression has also been noted previously in insulin-sensitive tis- sues of mice and humans using RT-PCR [10,22] suggesting that, in addition to its role within the CNS, TUB may also regulate fuel and energy balance in peripheral tissues targeted by hormones such as insulin and tri-iodothyronine (T3, i.e. thyroid hormone). We show here that TUB is expressed in primary and cultured adipocytes and that expression of TUB in the latter is regulated in a differentiation- dependent manner. Intriguingly, expression of TUB is enhanced in adipocytes from insulin resistant animals and in 3T3-L1 adipocytes rendered insulin resistant by sustained exposure to a high insulin concentration and in cells incubated with T3. These findings indi- cate that TUB expression is under hormonal control in adipocytes and that TUB may participate in insulin signalling and modulate adipocyte physiology during insulin resistance and altered thyroid status. 0006-291X/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2009.10.147 * Corresponding author. Fax: +44 1382 385514. E-mail address: [email protected] (H.S. Hundal). 1 These authors have contributed equally to this work. Biochemical and Biophysical Research Communications 390 (2009) 1328–1333 Contents lists available at ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemical and Biophysical Research Communications 390 (2009) 1328–1333
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Expression and modulation of TUB by insulin and thyroid hormonein primary rat and murine 3T3-L1 adipocytes
Clare Stretton a,1, Gary J. Litherland b,1, Audrey Moynihan a, Eric Hajduch c,d, Harinder S. Hundal a,*
a Division of Molecular Physiology, College of Life Sciences, University of Dundee, Dundee DD1 5EH, UKb Institute of Cellular Medicine, The Medical School, Newcastle University, Newcastle NE2 4HH, UKc INSERM U872, Université Pierre et Marie Curie, Paris 6, Franced Université Paris Descartes, Paris F-75006, France
a r t i c l e i n f o
Article history:Received 26 October 2009Available online 1 November 2009
Keywords:3T3-L1AdipogenesisZucker ratInsulinTubby
0006-291X/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.bbrc.2009.10.147
* Corresponding author. Fax: +44 1382 385514.E-mail address: [email protected] (H.S. Hu
1 These authors have contributed equally to this wo
a b s t r a c t
tub encodes a protein of poorly understood function, but one implicated strongly in the control of energybalance and insulin sensitivity. Whilst tub expression is particularly prominent in neurones it is alsodetectable in extraneuronal tissues. We show here, for the first time, expression of TUB protein in rat adi-pocytes and the murine adipocyte model 3T3-L1 and demonstrate that insulin induces its tyrosine phos-phorylation and association with the insulin receptor. TUB expression is regulated developmentallyduring adipogenic differentiation of 3T3-L1 cells and in response to cell treatment with thyroid hormoneor induction of insulin resistance. TUB was upregulated 5- to 10-fold in adipocytes from obese Zucker ratsand 3T3-L1 adipocytes that had been rendered insulin resistant, a response that could be antagonised byrosiglitasone, an insulin-sensitising drug. Our data are consistent with a previously unforeseen role forTUB in insulin signalling and fuel homeostasis in adipocytes.
� 2009 Elsevier Inc. All rights reserved.
Introduction
TUB and the tubby-like proteins (TULPs) 1–3 comprise a smallgene family whose gene sequences are highly conserved acrossspecies [5,11,15,21,23]. This suggests that they may serve a basicfunction within the cell but, as yet, their exact role remainsunclear. The four proteins of the family are highly homologousand have over-lapping expression profiles, but gene-inactivationexperiments suggest they serve non-redundant roles. tub wasoriginally identified as the gene involved in the tubby mousesyndrome, characterized by late-onset obesity, insulin resistanceand sensory impairments [6,22]. This defect is due to expressionof a mutant TUB protein in which the C-terminal 44 amino acidresidues are replaced by 24 incorrect residues encoded intronical-ly. Gene knockout studies on TUB result in the same phenotype asthe tubby mouse suggesting that the phenotype is caused by a lossof function mutation [22].
In neuronal cell lines TUB can be regulated by a range of stimuliincluding insulin and thyroid hormone and may regulate severalfunctions including vesicle trafficking, gene transcription and en-ergy homeostasis [12,13,28]. In these latter studies TUB was shown
ll rights reserved.
ndal).rk.
to become tyrosine phosphorylated and associate with the insulinreceptor following insulin treatment suggesting that it may act asan adaptor protein in insulin signalling [13]. The ‘‘tubby domain”permits interaction of TUB with phosphatidylinositol-4,5-bisphos-phate (PI(4,5)P2) [27] and, as such, this may facilitate its associa-tion/retention with the inner leaflet of the plasma membranewhere it may be placed in close proximity to receptor proteins thatalter its function through covalent modification. More recent workhas indicated that tubby proteins are also secreted from cellsraising the suggestion that they may also possess extracellularfunctions [4]. Although most abundant in neuronal tissue, tubexpression has also been noted previously in insulin-sensitive tis-sues of mice and humans using RT-PCR [10,22] suggesting that, inaddition to its role within the CNS, TUB may also regulate fuel andenergy balance in peripheral tissues targeted by hormones such asinsulin and tri-iodothyronine (T3, i.e. thyroid hormone). We showhere that TUB is expressed in primary and cultured adipocytes andthat expression of TUB in the latter is regulated in a differentiation-dependent manner. Intriguingly, expression of TUB is enhanced inadipocytes from insulin resistant animals and in 3T3-L1 adipocytesrendered insulin resistant by sustained exposure to a high insulinconcentration and in cells incubated with T3. These findings indi-cate that TUB expression is under hormonal control in adipocytesand that TUB may participate in insulin signalling and modulateadipocyte physiology during insulin resistance and altered thyroidstatus.

C. Stretton et al. / Biochemical and Biophysical Research Communications 390 (2009) 1328–1333 1329
Methods
Materials. Donor bovine serum, foetal bovine serum (FBS) andpenicillin/streptomycin solution were obtained from Invitrogen(Paisley, UK). Dulbecco’s Modified Eagle’s Medium (DMEM) andall other reagents were obtained from Sigma–Aldrich (St. Louis,MO) unless specified otherwise. Anti-TUB antibodies were ob-tained from Chemicon and Santa-Cruz (Santa Cruz, CA), anti-PKBantibodies and anti-phosphotyrosine antibody from Cell SignallingTechnology (Beverly, MA) and anti-insulin receptor b-subunit fromUpstate (Billerica, MA). Rosiglitasone was a gift from Dr. CalumSutherland (University of Dundee).
Animal work. Male Sprague–Dawley rats and male obese Zuckerrats (fa/fa) and their lean littermates were purchased from Harlanand Olac, Bicester, Oxon, UK. Adipocytes were isolated from epidid-ymal fat pads and fractionated to isolate membranes enriched inplasma membranes (PM), low density microsomes (LDM) and highdensity microsomes (HDM) as described previously [9]. The pro-tein content of membrane fractions was determined by the methodof Bradford [3].
Culture of 3T3-L1 adipocytes and hormone treatments. 3T3-L1 adi-pocytes were maintained in DMEM supplemented with 10% (v/v)donor calf bovine serum, 100 U/ml penicillin, 100 lg/ml strepto-mycin and 0.25 lg/ml amphotericin B. Adipocyte differentiationwas induced by culturing confluent cells in DMEM containing10% (v/v) FBS, 100 mM isobutylmethylxanthine, 0.25 mM dexa-methasone and 1 mg/ml insulin for two days followed by DMEMcontaining FBS and insulin only for a further two days. Cells werecultured in DMEM containing 10% FBS (v/v) alone for the remain-der of the differentiation period. Differentiated cells were har-vested at periods up to 10 days after induction of differentiation.Acute insulin treatment was carried out after a period of 3–4 h ser-um starvation. Insulin was added to the cultures at a final concen-tration of 100 nM for 10 min. T3 incubations were carried out for7 h at concentrations indicated in the figure.
Isolated rat adipocytes and 3T3-L1 adipocytes were lysed usinglysis buffer, whereas in some experiments subcellular membranefractions from 3T3-L1 adipocytes were prepared as described pre-viously [8]. Protein content in cell and tissue lysates and mem-brane fractions was quantified using the Bradford method [3].
Immunoblotting and immunoprecipitation. Cell lysates were sep-arated by SDS/PAGE and immunoblotted with primary antibodiesagainst target proteins of interest prior to incubation with appro-priate peroxidise-conjugated IgG antibodies and chemilumines-cence detection involving exposure to autoradiographic film [8].Immunoprecipitation was performed using protein G-sepharosebeads complexed to anti-TUB or anti-phosphotyrosine antibodies.The bead/antibody complex was incubated with 500 lg cell lysateprotein and immunoprecipitates pelleted as described previously[8] and subjected to SDS–PAGE and immunoblotting using anti-bodies against proteins of interest.
RNA extraction and quantitative PCR. RNA extraction was carriedout using TriReagent (Sigma–Aldrich). RNA was treated withDNase I (Promega) and first-strand cDNA synthesis performedfrom 1 lg RNA using M-MLV reverse transcriptase (Promega, Mad-ison, WI) and oligo d(T)s. qPCR was performed using an ABI prism7000 (Applied Biosystems, Foster City, CA) real-time thermocyclerand SYBRGreen kit (Sigma–Aldrich) and primers targeting TUB andthe a1-subunit of the sodium/potassium ATPase as a control. PCRconditions were as follows: initial denaturation 95 �C for 2 min fol-lowed by 40 cycles of denaturation at 95 �C for 15 s, annealing at55 �C for 15 s and extension at 68 �C for 30 s. Primer sequencesused were as follows: tub sense 50-ACAATGGCGTCAACCCTCAG-30,tub anti-sense 50-CTGGGACGATCACACTCATCTTC-30, Na+/K+ ATPasea1 sense 50-CAGCAGGGAATGAAGAACAAGATC-30, Na+/K+ ATPase
a1 anti-sense 50-GTAGGTTTGAGGGGATACATCCTAAG-30. The ratioof TUB mRNA expression versus Na+/K+ ATPase a1 mRNA expres-sion was calculated using an equation developed by Pfaffl [25].
Results
Localisation and regulation of TUB expression during adipocytedifferentiation
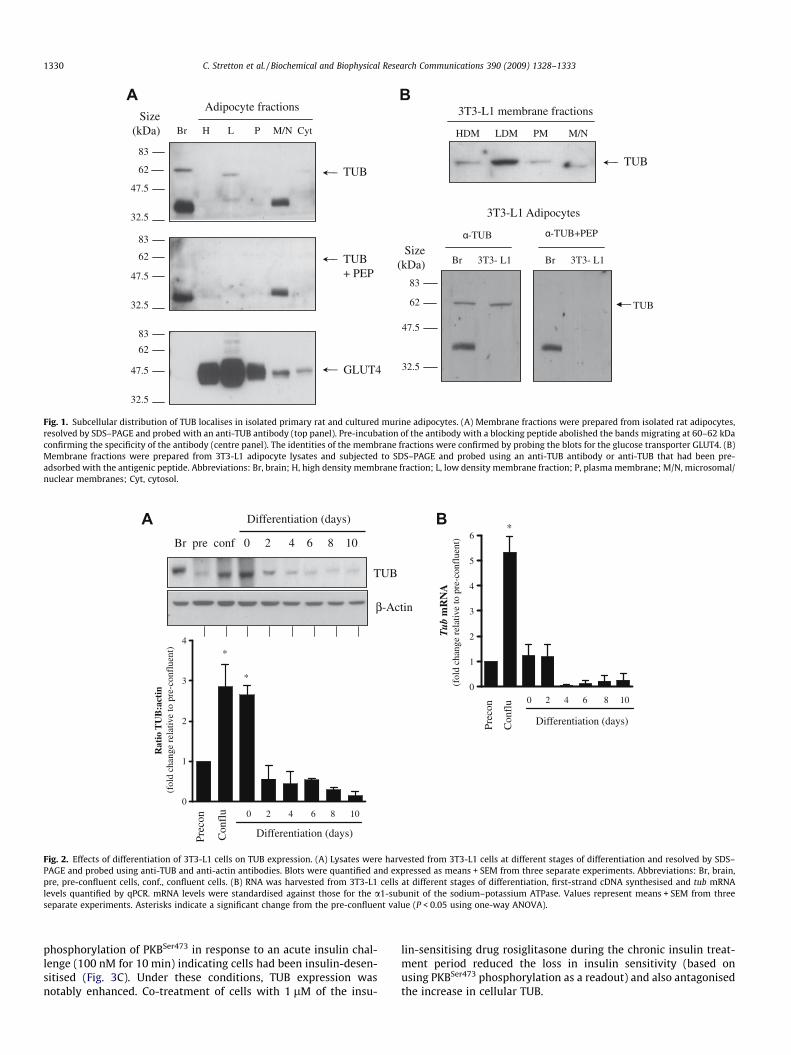
The presence and localisation of TUB in adipocytes was initiallydetermined using cell fractionation of isolated primary rat adipo-cytes. Whilst TUB was detectable in both the low density mem-brane (LDM) and cytoplasmic fractions it migrated as a slightlyfaster band (�60 kDa) than that observed in brain tissue(�62 kDa; Fig. 1A). This band appears to correspond to TUB sinceit was not detectable when using TUB antibody that had beenpre-adsorbed with the TUB antigenic peptide (Fig. 1A, centre pa-nel). A second band was also observed in brain tissue and in themicrosomal/nucleosomal membrane fraction at �35–40 kDa butthis could not be eliminated using the blocking peptide suggestingthat the band is due to non-specific binding of the antibody. Theidentity of the isolated membrane fractions was confirmed by blot-ting for the GLUT4 glucose transporter (Fig. 1A, lower panel),which, in unstimulated fats cells, is known to be present in muchhigher abundance in the LDM fraction compared with that en-riched with plasma membranes.
Similarly, in cultured murine 3T3-L1 adipocytes, TUB was ex-pressed predominantly in the LDM with a weaker signal detectablein the other membrane fractions (Fig. 1B). Again, adipocyte TUBwas seen to migrate slightly ahead of protein band observed in braintissue. Specificity of the immunoreactive TUB band in 3T3-L1 adipo-cytes was confirmed using peptide-blocked serum which failed todetect the 60 kDa band that we propose represents adipocyte TUB.
The effects of adipogenesis on TUB expression were assessed byimmunoblotting lysates from 3T3-L1 cells at various stages of dif-ferentiation (Fig. 2A). TUB was most abundant in confluent pre-adi-pocytes and at the onset of differentiation, but as differentiationprogressed TUB abundance fell progressively and was only weaklydetectable in fully differentiated adipocytes. tub mRNA exhibited asimilar expression profile to that of the protein (Fig. 2B), beinggreatest as cells approached confluence, but falling rapidly upondifferentiation.
Regulation of TUB by insulin and T3
TUB becomes tyrosine phosphorylated following insulin treat-ment of neuronal cells and in vitro assays have demonstrated thatTUB associates with and is a direct substrate for several tyrosineprotein kinases including the insulin receptor, Abl and Jak2 [13].We therefore sought to determine whether this might also be thecase in 3T3-L1 cells. Post-confluent pre-adipocytes were used inorder to maximise endogenous TUB content. Fig. 3A shows thatincubation of 3T3-L1 cells with insulin induced tyrosine phosphor-ylation of TUB. Probing TUB immunoprecipitates with an anti-phosphotyrosine antibody revealed a band at �60 kDa only in pre-cipitates from insulin-treated cells. To confirm the identity of thisprotein band, we performed the converse experiment and probedfor TUB in phosphotyrosine immunoprecipitates from 3T3-L1 celllysates. TUB was found in phosphotyrosine precipitates only insamples that had been treated with insulin (Fig. 3A).
Since the interaction between the two proteins appears to beregulated by insulin we investigated if insulin-sensitivity of adipo-cytes may play a role in regulating TUB expression. Differentiated3T3-L1 adipocytes were rendered insulin resistant by treatmentwith 1 lM insulin for 24 h. Chronic insulin treatment (Ic) reduced
83
62
47.5
32.5
83
62
47.5
32.5
83
62
47.5
32.5
Size(kDa) Br H L P M/N Cyt
Adipocyte fractions
TUB
TUB + PEP
GLUT4
HDM LDM PM M/N
3T3-L1 membrane fractions
TUB
83
62
47.5
32.5
Br 3T3- L1 Br 3T3- L1
α-TUB α-TUB+PEP
TUB
3T3-L1 Adipocytes
Size(kDa)
Fig. 1. Subcellular distribution of TUB localises in isolated primary rat and cultured murine adipocytes. (A) Membrane fractions were prepared from isolated rat adipocytes,resolved by SDS–PAGE and probed with an anti-TUB antibody (top panel). Pre-incubation of the antibody with a blocking peptide abolished the bands migrating at 60–62 kDaconfirming the specificity of the antibody (centre panel). The identities of the membrane fractions were confirmed by probing the blots for the glucose transporter GLUT4. (B)Membrane fractions were prepared from 3T3-L1 adipocyte lysates and subjected to SDS–PAGE and probed using an anti-TUB antibody or anti-TUB that had been pre-adsorbed with the antigenic peptide. Abbreviations: Br, brain; H, high density membrane fraction; L, low density membrane fraction; P, plasma membrane; M/N, microsomal/nuclear membranes; Cyt, cytosol.
β-Actin
TUB
Br pre conf 0 2 4 6 8 10
Differentiation (days)
0
1
2
3
4*
Prec
on
Con
flu 0 2 4 6 8 10
Differentiation (days)
Rat
io T
UB
:act
in(f
old
chan
ge r
elat
ive
to p
re-c
onfl
uent
)
*0
1
2
3
4
5
6*
Prec
on
Con
flu 0 2 4 6 8 10
Differentiation (days)
Tub
mR
NA
(fol
d ch
ange
rel
ativ
e to
pre
-con
flue
nt)
Fig. 2. Effects of differentiation of 3T3-L1 cells on TUB expression. (A) Lysates were harvested from 3T3-L1 cells at different stages of differentiation and resolved by SDS–PAGE and probed using anti-TUB and anti-actin antibodies. Blots were quantified and expressed as means + SEM from three separate experiments. Abbreviations: Br, brain,pre, pre-confluent cells, conf., confluent cells. (B) RNA was harvested from 3T3-L1 cells at different stages of differentiation, first-strand cDNA synthesised and tub mRNAlevels quantified by qPCR. mRNA levels were standardised against those for the a1-subunit of the sodium–potassium ATPase. Values represent means + SEM from threeseparate experiments. Asterisks indicate a significant change from the pre-confluent value (P < 0.05 using one-way ANOVA).
1330 C. Stretton et al. / Biochemical and Biophysical Research Communications 390 (2009) 1328–1333
phosphorylation of PKBSer473 in response to an acute insulin chal-lenge (100 nM for 10 min) indicating cells had been insulin-desen-sitised (Fig. 3C). Under these conditions, TUB expression wasnotably enhanced. Co-treatment of cells with 1 lM of the insu-
lin-sensitising drug rosiglitasone during the chronic insulin treat-ment period reduced the loss in insulin sensitivity (based onusing PKBSer473 phosphorylation as a readout) and also antagonisedthe increase in cellular TUB.

IR βIP: α-TUBIB: α-IRβ
IR βWCLIB: α-IRβ
Con Ins
IP: α-TUBWB: α-PYTUB -
WCLWB: α-TUB
IP: α-PYWB: α-TUB
IP: α-TUBIB: α-PY
Con Ins
75TUB -
TUB -
105
kDa
TUB
3T3-L1 Adipocytes
PKBS473
PKB
- + + + + Ins (100 nM)
Ic
Ic+
RO
SI
RO
SI- -
β-actin-
5 10 15 5 10 15 5 5 Age (wk)
Lean Obese Lean ObeseAdipose Tissue Brain
TUB -
Fig. 3. Regulation of TUB by insulin treatment. (A) Murine 3T3-L1 pre-adipocytes were treated with 100 nM insulin for 30 min at 37 �C and lysed prior toimmunoprecipitation using a-TUB. Immune pellets were immunoblotted using a-phosphotyrosine (PY). The top panel shows an immunoreactive TUB band at �60 kDa.Lysates were also subject to immunoprecipitation using a-PY. Immune pellets were subjected to SDS–PAGE and immunoblotting using a-TUB (middle panel). (B) Cells weretreated with 100 nM insulin for 30 min at 37 �C and lysates were subjected to immunoprecipitation using a-TUB. The top panel shows an immunoreactive band atapproximately 97 kDa. The middle and lower panels portray a-TUB immunoprecipitates and cell lysates, respectively, probed with a-IR-b subunit. Abbreviations: IB,immunoblot; IP, immunoprecipitation; WCL, whole cell lysate. (C) Murine 3T3-L1 adipocytes were rendered insulin resistant by chronic exposure to 1 lM insulin (Ic) for 24 hand/or treated with the insulin sensitiser rosiglitasone (10 nM) prior to an acute insulin challenge (100 nM, 10 min) and cell lysis and SDS–PAGE. Lysates wereimmunoblotted with antibodies against TUB, PKB and phospho-PKBSer473. The blots shown are representative of three separate experiments. (D) Adipocytes and brain tissuefrom lean and obese Zucker rats aged 5, 10 and 15 weeks were analysed. Adipocyte fractions (30 lg) were subjected to SDS–PAGE and immunoblotting using a-TUB antibodyand b-actin (loading control). At least three animals per condition were analysed and a representative blot shown.
β-actin
0 10 100 1000
[T3]/nM
TUB
Fig. 4. TUB expression is positively regulated by thyroid hormone in 3T3-L1adipocytes. Murine 3T3-L1 adipocytes were incubated in media containing 1% (v/v)foetal bovine serum and supplemented with concentrations of T3 indicated for 7 hat 37 �C. Cells were lysed and proteins resolved by SDS–PAGE, prior to immuno-blotting and detection of TUB and b-actin as described. Blots shown are represen-tative of three separate experiments.
C. Stretton et al. / Biochemical and Biophysical Research Communications 390 (2009) 1328–1333 1331
The above findings suggest that cell sensitivity to insulin maybe an important determinant of TUB expression in adipocytes. Toexplore this idea further we examined TUB expression in a rodentmodel for insulin resistance, the obese fa/fa Zucker rat. Adipocytesfrom lean and obese animals at 5, 10 and 15 weeks of age wereanalysed. At 5 weeks of age, fa/fa Zucker rats were not significantlyobese (body weight 162 ± 5 g lean, 165 ± 5 g obese, [17]). Uponreaching maturity, however, they became progressively more ob-ese compared to lean littermates (315 ± 2 g lean, 409 ± 6 g obeseat 10 weeks; 313 ± 9 g lean, 520 ± 12 g obese at 15 weeks, [17]).It is clear that the expression of TUB in adipocytes from young,5-week-old animals was lower than in adipocytes from mature(10 and 15 week) animals, in both lean (7-fold) and obese (3.5-fold) rats (Fig. 3D). At all ages studied, we also observed a strikingincrease in expression of TUB in adipocytes from obese rats, com-pared to their lean littermates (5.1-fold at 5 weeks, 4.3-fold at10 weeks, 5.8-fold at 15 weeks; Fig. 3D). Despite the markedincrease in TUB expression in adipocytes of obese Zucker rats nodifference in brain TUB expression was observed between leanand obese Zucker rats (Fig. 3D).
In addition to insulin, TUB is also regulated by thyroid hormonein neuronal cells [14]. We therefore investigated if a similar mech-anism of regulation exists in adipocytes. Confluent 3T3-L1 pre-adi-pocytes were treated with increasing concentrations of T3 for 7 hand abundance of TUB assessed by immunoblotting (Fig. 4). TUB
expression was elevated in response to T3 treatment in a dose-dependent manner whilst total b-actin levels remained unaffected.This data suggests that T3 regulation of TUB expression is active inadipose tissue as well as in the brain [14].
Discussion
Previous work has shown that tub mRNA is expressed in numer-ous non-neuronal cell types, including adipose tissue [22]. How-

1332 C. Stretton et al. / Biochemical and Biophysical Research Communications 390 (2009) 1328–1333
ever, our understanding of TUB biology in such cell types has beenlargely overlooked given the much greater impetus to establishTUB function within the CNS, where TUB and its related familymembers are most prominently expressed [5,10]. It is surprisingtherefore that whilst data from a number of different groups haveimplicated TUB and the TULPS in various cellular responses [2,12–14,27] we still know very little of their precise functional role atthe molecular level. Moreover, whilst the phenotype of the tubbymouse supports a critical role for TUB in preserving normal sensoryfunction and energy homeostasis there are no convincing links be-tween the loss of neuronal TUB function and causal events in thedevelopment of obesity. Consequently, the exact role of TUB inthe aetiology of the obese tubby phenotype remains unknown,but it is of considerable interest that TUB is expressed in adipo-cytes; a tissue whose primary function is one of storing energy inthe form of neutral triglycerides.
The findings presented here extend the initial observation of tubmRNA in mouse adipose tissue [22] and demonstrate that TUB pro-tein is detectable in both primary rat and cultured murine 3T3-L1adipocytes. Of particular interest was the differentiation-linkedexpression of TUB in 3T3-L1 adipocytes, which was found to peakprior to adipogenesis but then decline rapidly as cells differentiateinto adipocytes. This expression profile is likely to be directed by reg-ulated changes in tub gene transcription based on the observationthat changes in TUB protein not only mirror, but slightly lag thosein tub mRNA expression. We currently do not know the nature ofthe stimulus or the signalling pathways that are responsible for insti-gating these changes in TUB expression, but the transient elevationin TUB abundance at a critical juncture within the differentiationprogramme may indicate that TUB controls early differentiationevents through transcriptional activation of differentially-expressedgenes. Crystal structure analysis of TUB shows that it contains aputative DNA binding groove in its C-terminal domain and potentialtranscriptional activation domains within its N-terminal region,which are highly suggestive of a function for TUB as a transcriptionfactor [2]. Consistent with this idea, studies in neuronal cells havealso shown that activation of the 5HT-2C serotonin receptor inducesTUB translocation to the nucleus where it may act to modulate genetranscription [27]. However, as yet, no direct transcriptional targetsor signalling pathways regulated as a consequence of nuclear local-isation of TUB have been identified.
Differentiation of 3T3-L1 adipocytes is associated with markedacquisition of insulin sensitivity [26] and it is plausible that the dif-ferentiation-linked reduction in TUB may, in part, be due to insu-lin-induced repression of the tub gene. Insulin repressesexpression of numerous genes encoding proteins with importantmetabolic functions (e.g. glucose-6-phosphatase and phosphoenol-pyruvate carboxykinase) [19] and it is plausible that tub may be-long to this gene group. This possibility is supported by ourfinding that 3T3-L1 adipocytes rendered insulin resistant bychronic exposure to high insulin exhibit a marked increased inTUB content, which can be attenuated by treating cells with theinsulin-sensitising drug rosiglitasone. Such regulation is not re-stricted to cultured adipocytes based on the striking increase inTUB abundance in adipocytes isolated from obese (fa/fa) Zuckerrats. The increase in adipocyte TUB was apparent in rats as youngas 5 weeks, which, while normoglyceamic, are already insulinresistant based on a significant increase in serum insulin comparedto that of their lean litter mates [17]. We have previously shownthat as these animals age they become increasingly more hyperins-ulinemic and hyperglycaemic [17] and so it is interesting to notethat with this progressive increase in insulin resistance TUBexpression in fa/fa adipocytes was also further elevated. Intrigu-ingly, similar changes in TUB were not observed in brain consistentwith the view that such regulation may not occur in a tissue that isnot a primary target of insulin action.
Studies in both neuronal cell lines and Caenorhabditis eleganshave suggested that TUB has a role to play in insulin-dependent re-sponses in the cell. In C. elegans TUB is detectable only in neuronsbut has been implicated in the regulation of both fat metabolismand life-span via independent mechanisms [20]. Regulation oflife-span by TUB is mediated through the insulin/IGF1 signallingpathway and requires the forkhead transcription factor daf16, anorthologue of the mammalian transcription factor FOXO3a[1,16,18,24]. Insulin treatment of CHO-IR and PC12 has previouslybeen shown to induce tyrosine phosphorylation of TUB and pro-mote association of TUB with SH2 domain-containing proteins[9]. However, precisely what cellular responses are targeted byphosphorylated TUB remains currently unknown. Nevertheless,our finding that acute insulin treatment induces TUB/insulin recep-tor association and tyrosine phosphorylation of TUB indicates thatthe signalling mechanism involved in the covalent modification ofneuronal TUB is also present in adipocytes.
T3 plays crucial roles in processes such as body growth, develop-ment and energy metabolism [7]. T3 increases the rate of energyexpenditure and consequently a prominent feature of hypothyroid-ism in humans is increased body weight and obesity. This is also afeature of the tubby mouse, and it has been shown previously thattub expression can be stimulated by T3 in rat Purkinje cells [14], mostlikely through the transcriptional activating activity of nuclear thy-roid hormone receptor (TR). However, this does not occur in the‘‘satiety centre” of the hypothalamus, the brain area thought to bemost intimately linked with energy homeostasis. Given this incon-clusive link between thyroid status, tub expression and energyhomeostasis, we assessed whether a similar situation might existin adipocytes, since adipose tissue is also crucial in the regulationof energy metabolism. The finding that T3 positively regulates TUBabundance in 3T3-L1 adipocytes suggests a novel pathway throughwhich thyroid hormone might exert control over energy metabo-lism, and reinforces the idea that adipocyte TUB expression may playa role in this vital physiological process.
In summary our data supports the idea that TUB may be a novelmediator of insulin signalling and energy metabolism in adiposetissue. Such a role for TUB would represent a previously overlookedmechanism by which loss of TUB function in tubby mice might re-sult in obesity and insulin resistance in this rodent model. Sincetubby mice share many of the characteristics of maturity-onsetobesity syndromes found in human populations, the elucidationof TUB functions in adipose tissue represents a novel and impor-tant focus for further study.
Acknowledgments
We are grateful for financial support from the European Com-mission (Contract LSHM-CT-20004-005272), Diabetes Researchand Wellness Foundation and Diabetes, UK.
References
[1] K. Ashrafi, F.Y. Chang, J.L. Watts, A.G. Fraser, R.S. Kamath, J. Ahringer, G.Ruvkun, Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatorygenes, Nature 421 (2003) 268–272.
[2] T.J. Boggon, W.S. Shan, S. Santagata, S.C. Myers, L. Shapiro, Implication of tubbyproteins as transcription factors by structure-based functional analysis,Science 286 (1999) 2119–2125.
[3] M.M. Bradford, A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding, Anal.Biochem. 72 (1976) 248–254.
[4] N.B. Caberoy, W. Li, Unconventional secretion of tubby and tubby-like protein1, FEBS Lett. 583 (2009) 3057–3062.
[5] K. Carroll, C. Gomez, L. Shapiro, Tubby proteins: the plot thickens, Nat. Rev.Mol. Cell Biol. 5 (2004) 55–63.
[6] D.L. Coleman, E.M. Eicher, Fat (fat) and tubby (tub): two autosomal recessivemutations causing obesity syndromes in the mouse, J. Hered. 81 (1990) 424–427.

C. Stretton et al. / Biochemical and Biophysical Research Communications 390 (2009) 1328–1333 1333
[7] H.C. Freake, J.H. Oppenheimer, Thermogenesis and thyroid function, Annu. Rev.Nutr. 15 (1995) 263–291.
[8] E. Hajduch, D.R. Alessi, B.A. Hemmings, H.S. Hundal, Constitutive activation ofProtein Kinase Ba (PKBa) by membrane targeting promotes glucose andsystem A amino acid transport, protein synthesis and GSK3 inactivation in L6muscle cells, Diabetes 47 (1998) 1006–1013.
[9] E. Hajduch, F. Darakhshan, H.S. Hundal, Fructose uptake in rat adipocytes:GLUT5 expression and the effects of streptozotocin-induced diabetes,Diabetologia 41 (1998) 821–828.
[10] W. He, S. Ikeda, R.T. Bronson, G. Yan, P.M. Nishina, M.A. North, J.K. Naggert,GFP-tagged expression and immunohistochemical studies to determine thesubcellular localization of the tubby gene family members, Brain Res. Mol.Brain Res. 81 (2000) 109–117.
[11] A. Ikeda, P.M. Nishina, J.K. Naggert, The tubby-like proteins, a family with rolesin neuronal development and function, J. Cell Sci. 115 (2002) 9–14.
[12] A. Ikeda, Q.Y. Zheng, A.R. Zuberi, K.R. Johnson, J.K. Naggert, P.M. Nishina,Microtubule-associated protein 1A is a modifier of tubby hearing (moth1), Nat.Genet. 30 (2002) 401–405.
[13] R. Kapeller, A. Moriarty, A. Strauss, H. Stubdal, K. Theriault, E. Siebert, T.Chickering, J.P. Morgenstern, L.A. Tartaglia, J. Lillie, Tyrosine phosphorylationof tub and its association with Src homology 2 domain-containing proteinsimplicate tub in intracellular signaling by insulin, J. Biol. Chem. 274 (1999)24980–24986.
[14] N.P. Koritschoner, M. Alvarez-Dolado, S.M. Kurz, M.F. Heikenwalder, C. Hacker,F. Vogel, A. Munoz, M. Zenke, Thyroid hormone regulates the obesity gene tub,EMBO Rep. 2 (2001) 499–504.
[15] C.P. Lai, C.L. Lee, P.H. Chen, S.H. Wu, C.C. Yang, J.F. Shaw, Molecular analyses of theArabidopsis TUBBY-like protein gene family, Plant Physiol. 134 (2004) 1586–1597.
[16] K. Lin, J.B. Dorman, A. Rodan, C. Kenyon, daf-16: an HNF-3/forkhead familymember that can function to double the life-span of Caenorhabditis elegans,Science 278 (1997) 1319–1322.
[17] G.J. Litherland, E. Hajduch, G.W. Gould, H.S. Hundal, Fructose transport andmetabolism in adipose tissue of Zucker rats: diminished GLUT5 activity duringobesity and insulin resistance, Mol. Cell Biochem. 261 (2004) 23–33.
[18] L.X. Liu, J.M. Spoerke, E.L. Mulligan, J. Chen, B. Reardon, B. Westlund, L. Sun, K.Abel, B. Armstrong, G. Hardiman, J. King, L. McCague, M. Basson, R. Clover, C.D.Johnson, High-throughput isolation of Caenorhabditis elegans deletion mutants,Genome Res. 9 (1999) 859–867.
[19] C. Mounier, B.I. Posner, Transcriptional regulation by insulin: from thereceptor to the gene, Can. J. Physiol. Pharmacol. 84 (2006) 713–724.
[20] A. Mukhopadhyay, B. Deplancke, A.J. Walhout, H.A. Tissenbaum, C. eleganstubby regulates life span and fat storage by two independent mechanisms, CellMetab. 2 (2005) 35–42.
[21] P.M. Nishina, M.A. North, A. Ikeda, Y. Yan, J.K. Naggert, Molecularcharacterization of a novel tubby gene family member, TULP3, in mouse andhumans, Genomics 54 (1998) 215–220.
[22] K. Noben-Trauth, J.K. Naggert, M.A. North, P.M. Nishina, A candidate gene forthe mouse mutation tubby, Nature 380 (1996) 534–538.
[23] M.A. North, J.K. Naggert, Y. Yan, K. Noben-Trauth, P.M. Nishina, Molecularcharacterization of TUB, TULP1, and TULP2, members of the novel tubby genefamily and their possible relation to ocular diseases, Proc. Natl. Acad. Sci. USA94 (1997) 3128–3133.
[24] S. Ogg, S. Paradis, S. Gottlieb, G.I. Patterson, L. Lee, H.A. Tissenbaum, G. Ruvkun,The Fork head transcription factor DAF-16 transduces insulin-like metabolicand longevity signals in C. elegans, Nature 389 (1997) 994–999.
[25] M.W. Pfaffl, A new mathematical model for relative quantification in real-timeRT-PCR, Nucleic Acids Res. 29 (2001) e45.
[26] M.D. Resh, Development of insulin responsiveness of the glucose transporterand the (Na+,K+)-adenosine triphosphatase during in vitro adipocytedifferentiation, J. Biol. Chem. 257 (1982) 6978–6986.
[27] S. Santagata, T.J. Boggon, C.L. Baird, C.A. Gomez, J. Zhao, W.S. Shan, D.G.Myszka, L. Shapiro, G-Protein signaling through tubby proteins, Science 292(2001) 2041–2050.
[28] H. Stubdal, C.A. Lynch, A. Moriarty, Q. Fang, T. Chickering, J.D. Deeds, V.Fairchild-Huntress, O. Charlat, J.H. Dunmore, P. Kleyn, D. Huszar, R. Kapeller,Targeted deletion of the tub mouse obesity gene reveals that tubby is a loss-of-function mutation, Mol. Cell Biol. 20 (2000) 878–882.
Related Documents











